-
&p.1:Abstract The sensory innervation of the hard palate
ofthe rhesus monkey was studied by light and electron mi-croscopy.
The mucosa of the hard palate is subject to aparticularly heavy
mechanical load requiring functionalspecialisation of the horny
epithelium in the form ofthickenings – the papilla incisiva and
eight pairs of rugaepalatinae. A thin layer of firm connective
tissue (laminapropria) attaches the mucosa to the periost of the
hardpalate. Sensory nerve fibres were found most abundantlyin the
papilla incisiva and first rugae palatinae. Theirnumber decreases
in an aboral direction. Five types ofsensory nerve endings were
found. Free nerve endingswere ubiquitous in the epithelium and
lamina propria.Merkel nerve endings were found in the basal layer
ofthe epithelium of the papilla incisiva and rugae
palatinae.Meissner corpuscles were located in the connective
tis-sue between epithelial pegs, while lamellated corpuscleswere
seen below the epithelial pegs. Ruffini corpuscleswere found in the
deeper layer of the lamina propria.Thus, a variety of sensory nerve
endings were found inthe hard palate, especially in those areas
that are in closecontact with the tongue during chewing of food.
Thisrich innervation suggests an important role in monitoringthe
mechanical properties of food and the position of thetongue.
&kwd:Key words Mechano-receptors · Palate · Merkel cell
·Ruffini corpuscle · Free nerve ending · Lamellatedcorpuscle ·
Meissner corpuscle&bdy:
Introduction
Parts of the oral mucosa are exposed to a particularlyheavy
mechanical load. In those areas the mucosa has a
horny epithelium with a firm underlying connective tis-sue
(lamina propria). This type is termed masticatorymucosa and can be
found in the gingiva and hard palate.In the palate, the mucosa
forms macroscopically visiblethickenings: a papilla incisiva and
several pairs of rugaepalatinae. Food is pressed with the tongue
against theseregions to check its mechanical properties and
monitorwhether it is ready for swallowing. For this
complexfunction, the mucosa of the hard palate and papilla
incis-iva needs to be well equipped with mechanoreceptors.
Following early light microscopic studies of the hu-man palate
by Gairns (1955), various investigators havecarried out electron
microscopic studies on the innerva-tion of the papilla incisiva in
smaller mammals: rat (Yehand Byers 1983; Byers and Yeh 1984; Chan
and Byers1985; Watanabe and Konig 1986), squirrel monkey(Garant et
al. 1980), mouse (Watanabe and Yamada1988; Tachibana et al. 1990).
However, this is the firstsuch investigation carried out in a
larger primate (Rhesusmonkey) with intact connection to the
underlying bonein order to study the topographic arrangement of
thesemechanoreceptors in situ. This study provides evidencefor a
rich supply with free nerve and Merkel nerve end-ings, and Ruffini,
Meissner and lamellated corpuscles inthe primate hard palate that
are likely to play an impor-tant role in monitoring mechanical
properties of food.
Materials and methods
Two adult Rhesus monkeys were killed with an overdose of
thio-pental and perfused with 6% glutaraldehyde in 0.05 M
phosphatebuffer via the left cardiac ventricle. The upper jaw and
hard palatewere removed from the animals and postfixed for several
days inthe same solution. The tissue was then decalcified by
storing in5% EDTA solution with 0.1 M sodium phosphate buffer (pH
7.4,room temperature) for 5 years. The solution was changed
everymonth. The state of decalcification was checked regularly by
me-chanical probing and by X-ray just before embedding.
Following decalcification the hard palate was cut into
1.5-mm-thick strips, and each strip was divided into 20 sections
from theoral to the aboral end, resulting in several thousand
blocks of tis-sue. These blocks were postfixed in 1% OsO4 in 0.1 M
sodium
Z. Halata (✉) · K.I. BaumannDepartment of Functional Anatomy,
Institute of Anatomy,University Hospital Hamburg- Eppendorf,
Martinistrasse 52,D-20246 Hamburg, Germanye-mail:
[email protected],Fax:
+49-40-4717-2845&/fn-block:
Anat Embryol (1999) 199:427–437 © Springer-Verlag 1999
O R I G I N A L A RT I C L E
&roles:Z. Halata · K.I. Baumann
Sensory nerve endings in the hard palateand papilla incisiva of
the rhesus monkey
&misc:Accepted: 30 September 1998
-
phosphate buffer with 1% sucrose at pH 7.4 for 2 h before
embed-ding in gycidether 100.
Serial semithin sections obtained from these blocks were
stainedaccording to Laczko and Levai (1975) and examined by light
mi-croscopy. Selected semithin sections were re-embedded, cut into
ul-trathin sections and contrasted with 1% uranyl acetate and 1%
leadcitrate (Reynolds 1963) for electron microscopy (Philips
300).
Results
In the Rhesus monkey, the mucosa of the hard palate iscovered by
a multilayered, horny, flat epithelium firmlyconnected with the
periost through the lamina propria.The whole mucosa has a thickness
of 2.5 to 3 mm. Inthis location, the epithelium has no stratum
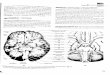
granulosum.The front part behind the processus alveolaris is
thick-ened to form the papilla incisiva, behind which eight ru-gae
palatinae are located (Fig. 1).
Epithelial thickenings (pegs) reach into the laminapropria to
different degrees. In the papilla incisiva and
rugae palatinae these pegs are especially thick and
well-developed. The lamina propria consists of a superficiallayer
facing the epithelium, forming a negative of theepithelial form –
thinner below the epithelial pegs andprotruding between them. By
analogy to the nomencla-ture of the skin this could be called the
papillary layer.The deeper layer consists of dense connective
tissue withsome seromucous salivary glands. This layer is firmly
at-tached to the periost of the hard palate, allowing
minimalmovement against each other.
Five types of sensory nerve endings were found:
1. Free nerve endings within the epithelium and the su-perficial
layer of the lamina propria (Figs. 2, 3, 6–8)
2. Merkel nerve endings in the bases of epithelial thick-enings
(Figs. 4, 5)
3. Ruffini corpuscles in the deeper layer of connectivetissue
about 250–400 µm below the basement mem-brane of the epithelium
(Figs. 9–11).
4. Meissner corpuscles in the lamina propria close tothe
epithelium between the epithelial thickenings(Figs. 12–14)
5. Small lamellated corpuscles below the epithelial thick-enings
(Figs. 15, 16)
Sensory receptors in the epithelium
Free nerve endings
Intraepithelial free nerve endings are supplied by
thinlymyelinated (A) afferent axons with diameters of 1–2 µmand
characterised by accumulation of mitochondria.Their number is
highest at the papilla incisiva and de-creases in the aboral
direction. They can be found in thestratum spinosum and in the
stratum basale. After losingtheir myelin sheath in the lamina
propria the nerve fibrespenetrate the basal lamina. The first type
branches toform enlarged nerve terminals between keratinocytes
ofthe stratum spinosum (Figs. 2, 3).
A second type of intraepithelial free nerve ending isseen just
above the basal lamina in direct contact withcells of the stratum
basale (Fig. 6).
428
Fig. 1 Schematic drawing of the monkey palate with papilla
incis-iva (IP) and eight pairs of rugae palatinae. The surrounding
teethare marked as incisors (I1 and I2), caninus (C), premolars (P1
andP2) and molars (M1 to M3). ×1&/fig.c:
Fig. 2 Free nerve ending in the papilla incisiva. The
enlargednerve terminal (★) is located in the statum spinosum of the
palateepithelium, showing the characteristic accumulation of
mitochon-dria. ×7600&/fig.c:
Fig. 3 Detail view of a free nerve ending (★) located between
ke-ratinocytes of the stratum spinosum. Multiple desmosomal
junc-tions between keratinocytes are indicated by arrows.
×17000&/fig.c:
Fig. 4 Epithelium of the papilla incisiva. Merkel cells (M)
withcytoplasmic protrusions (★) are seen in the stratum basale.
Theyare connected with the surrounding keratinocytes by
desmosomes(arrows). Nerve terminals (T) are associated with the
Merkel cellson their labial side. In the lamina propria an afferent
axon (N) withsheath of peripheral glial cells can be seen.
×7600&/fig.c:
▲
-
429
-
430
-
Fig. 5 Merkel cells (M) with cytoplasmic protrusions (arrows)
inthe papilla incisiva. Nerve terminals (★) are on the labial side
ofthe Merkel cells. ×5400&/fig.c:
Fig. 6 Two types of free nerve endings in the basal layer of
theepithelium and the lamina propria of the papilla incisiva,
respec-tively. The first one (arrow) is located just above the
basal laminamaking direct contact with the keratinocytes of the
stratum basale.The second is formed by a bundle of nerve terminals
(★) andmostly covered by terminal glial cells. Both types contain
accumu-lations of mitochondria. ×5400&/fig.c:
Fig. 7 Free nerve ending (★) surrounded by a simple sheath
ofterminal glial cells (S) and amorphous electron dense
material.Two or three layers of perineural cells (P) encapsulate
the com-plex. ×3200&/fig.c:
Fig. 8 Bundle of free nerve endings in the lamina propria
belowthe epithelium of the first ruga palatina. Terminals (arrows)
can berecognized by their accummulation of mitochondria. They
arecovered by terminal glial cells with their basal lamina.
×5400&/fig.c:
Fig. 9 Ruffini corpuscle in the lamina propria of the second
rugapalatina in cross section. The corpuscle consists of nerve
terminals(arrows) sheathed by terminal glial cells anchored between
bun-dles of collagen fibres. Thin lamellae of perineural cells (C)
formcylinders separating the nerve terminals from the surrounding
con-nective tissue. The nerve terminals are supplied by myelinated
ax-ons (A) with a diameter of about 3 to 5 µm.
×3250&/fig.c:
Fig. 10 Oblique section of a cylinder from a Ruffini corpuscle
inthe lamina propria of the second ruga palatina. Nerve terminals
areindicated by arrows. ×3250&/fig.c:
Fig. 11 Detail of nerve terminals from a Ruffini corpuscle.
Nerveterminals are partly sandwiched between thin lamellae of
terminalglial cells (★) leaving parts of the axolemma (arrows) in
directcontact with the basal lamina and collagen fibrils.
×14500&/fig.c:
431
Merkel nerve endings
The basal layer of the epithelial thickenings containsMerkel
nerve endings. Their number depends on thethickness of the
epithelial pegs. In larger pegs up to 12Merkel nerve endings can be
found. In smaller onesthere may be only individual Merkel nerve
endings. EachMerkel nerve ending consists of a Merkel cell and a
dis-coid nerve terminal. The Merkel cells are oval in shapewith the
long axis oriented perpendicularly to the mucos-al surface. Their
nuclei are lobulated and contain typicaldense core granules in that
part of the cytoplasm facingthe nerve terminal. Desmosomal contacts
with the sur-rounding keratinocytes are often seen. Between
thesecontact sites protoplasmic protrusions extend either be-tween
or are invaginated into neighbouring keratinocytes(Figs. 4, 5). The
discoid nerve terminals are oriented par-allel to the longitudinal
axis of the Merkel cells facingtheir labial side and are
characterised by accumulation ofmitochondria. Synapse-like contacts
can be seen be-tween the axolemma of the nerve terminal and the
cyto-plasmic membrane of the Merkel cell (Fig. 4). Afferentaxons
are myelinated with diameters of 4–5 µm. Afterlosing their myelin
sheath the branches form terminalsinnervating up to 6 Merkel
cells.
Sensory receptors in the lamina propria
Free nerve endings
Two types of free nerve endings can be found innervatedby thinly
myelinated (Aδ-) and unmyelinated (C-) fibres.The endings of
Aδ-fibres show typical thickenings andare only partly covered by
processes of the terminal glialcell (Fig. 6). Very rarely they are
embedded by masses ofbasal lamina-like material and wrapped within
a perineu-ral capsule, resembling a very simple corpuscle (Fig.
7).The endings of unmyelinated fibres form groups of ter-minals
(Fig. 8). The terminals are thin and covered byprocesses of the
terminal glial cells. Only in serial sec-
tions can they be clearly recognised as terminals ratherthan
passing axons.
Ruffini corpuscles
Ruffini corpuscles are found in the lamina propria of thepapilla
incisiva and rugae palatinae (Figs. 9, 10). Theyare cylindrical in
shape. The capsules of the cylinders areformed by flat perineural
cells with both ends open. Col-lagen fibres of the lamina propria
enter via these endsand run through the length of the cylinder to
leave at theother end. These fibres are compartmentalised by thin
la-mellar cells. The afferent axons are thick myelinated fi-bres
(Aβ) with diameters of about 5 µm (Fig. 9) penetrat-ing the capsule
at the long side of the cylinder. The peri-neurium of the nerve
blends with the capsule of the cor-puscle. Within the cylinder the
axon loses the myelinsheath and branches into several terminals.
Each termi-nal is partly sandwiched between thin lamellae of
termi-nal glial cells. Parts of the axolemma and sometimesslim
processes are in direct contact with the basal laminaof the glial
cell and collagen fibrils (Fig. 11). Sometimesseveral Ruffini
corpuscles could be seen close together,oriented in different
directions following the direction ofthe collagen fibre
bundles.
Meissner corpuscles
Meissner corpuscles are located in the connective tissuebetween
the epithelial pegs in the papilla incisiva andfirst two rugae
palatinae and are supplied by one or twomyelinated afferent axons
with diameters of 4–5 µm.Their size increases with the thickness of
the papillarylayer of the lamina propria. They consist of several
flat-tened nerve terminals surrounded by 1–5 flat cytoplas-mic
lamellae of terminal glial cells. They lack a perineu-ral capsule
and are generally located so close below theepithelium that the
basal lamina of the terminal glial cellblends with the basal lamina
of the epithelium (Fig. 12).The nerve terminals have a discoid
shape oriented in par-allel to the epithelial surface. They
resemble a stack ofcoins with lamellae of the terminal glial cell
and materialof connective tissue between them (Figs. 13, 14). The
cy-
▲
-
432
-
433
-
toplasm and nuclei of the glial cells are usually at theside of
this stack.
Lamellated corpuscles
Small lamellated corpuscles are found individually orgrouped
mostly in the connective tissue of the papilla in-cisiva.
Occasionally they can be seen below the epitheli-um of the first
two rugae palatinae. In contrast toMeissner corpuscles they are
located well below the epi-thelial pegs, without direct contact to
the basal lamina,and possess a thin capsule of two or three layers
of peri-neural cells. Their longitudinal axis runs parallel to
themucosal surface.
The lamellated corpuscles consist of a nerve terminal,an “inner
core” and a capsule (Fig. 15). In the centre of
the two symmetric halves of the inner core, formed bythin
cytoplasmic processes of a pair of terminal glialcells covered by
basal laminae, is an oval shaped nerveterminal. Longitudinal clefts
filled with basal membranematerial can be seen between the halves
(Fig. 16). Theslim ends of the oval nerve terminal are oriented
towardsthe clefts. The corpuscle is innervated by a
myelinatedafferent axon of about 5 µm diameter. After losing
themyelin sheath the unmyelinated terminal axon maybranch before
forming the nerve terminals.
Discussion
The present study confirms that in primates a great vari-ety of
sensory nerve endings can be found in the masti-catory mucosa that
are likely to play an important role inchecking the food during the
chewing process. This is inline with previous findings in smaller
mammals. Wefound two morphologically distinct types of free
nerveendings in the epithelium and two in the underlying
con-nective tissue, Merkel cell nerve endings at the bases
ofepithelial thickenings, Ruffini corpuscles in the deeperlayer of
the lamina propria, Meissner corpuscles in thelamina propria with
close contact to the basal lamina ofthe epithelium, and small
lamellated corpuscles in theconnective tissue of the lamina
propria. This suggests a
434
Fig. 12 Small Meissner corpuscle in the lamina propria just
be-low the epithelium. The basal lamina of the glial cell blends
withthe basal lamina of the epithelium (arrow). Discoid nerve
termi-nals (★) are surrounded by thin cytoplasmic processes of the
ter-minal glial cell (S). ×5400&/fig.c:
Figs. 13, 14 Larger Meissner corpuscles in the lamina propria
ofthe papilla incisiva. They are located so close to the
epitheliumthat the basal laminae blend. Each corpuscle consists of
flat nerveterminals (★) and terminal glial cells (S) with its thin
cytoplasmicprocesses both running parallel to the epithelial
surface. ×5400&/fig.c:
Fig. 15 Group of small lamel-lated corpuscles from the lami-na
propria of the papilla incis-iva. In contrast to the
Meissnercorpuscles shown in Figs. 11 to13 they are about 20 µm
belowthe basal lamina and are encap-sulated by thin perineural
cells(C). In the centre of the innercore formed by thin lamellae(L)
of terminal glial cells is thenerve terminal (★). The sup-plying
myelinated axon has adiameter of about 4 µm. ×2500&/fig.c:
▲
-
high degree of specialisation of the various types
ofmechanoreceptors for different parts of the sensory pro-cess
involved in masticatory control.
Free nerve endings
Free nerve endings were found in large numbers in thepapilla
incisiva, with decreasing numbers in the aboraldirection. They are
particularly concentrated in the ele-vated parts of the mucosa
(papilla incisiva and rugaepalatinae). In the mucosa between the
elevated parts onlya few free nerve endings can be seen. Two types
withdifferent characteristics and location were seen in
theepithelium. Those in the stratum spinosum were
simpleenlargements of the peripheral end of the thinly myelin-ated
afferent nerve fibre. In contrast, the second type waslocated
exclusively in the stratum basale, making directcontact with
keratinocytes containing accumulations ofmitochondria. In the skin,
this type is believed to havemainly thermoreceptive functions
(Hensel 1973), andthis is likely to be the case in the oral mucosa
as well.However, we are not aware of studies employing electro-
physiological recordings from such receptors. Thosenerve endings
in the stratum spinosum were described asprobable mechanoreceptors
(Munger 1965) but may alsoact as polymodal nociceptors. In addition
to those in theepithelium, in the lamina propria also two types of
freenerve endings could be distinguished. Nerve endings ofafferent
C-fibres showed minimal terminal enlargement,appearing in groups
wrapped by processes of the termi-nal glial cell. Thus, in order to
be sure that these wereendings rather than passing nerve fibres,
serial sectionshad to be examined. In contrast, nerve endings of
thinlymyelinated fibres had clear terminal enlargements andwere
only partially covered by processes of the terminalglial cells.
Functionally, the free nerve endings in theconnective tissue are
most likely polymodal nociceptors(Kruger et al. 1981; Kruger and
Halata 1996). However,some of the free nerve endings in the
connective tissuemay be efferent autonomic fibres involved in
vasomotorcontrol.
Merkel nerve endings
Merkel nerve endings were found in large numbers inepithelial
pegs throughout the hard palate. All Merkelcells found in this
study were in contact with nerve ter-minals. It is striking that
the location of Merkel cells inrelation to the nerve terminals
seems to depend on the di-rection of the mechanical stimulus to be
perceived. In theskin, the long axis of Merkel cells runs parallel
to thesurface of the epidermis with the nerve terminal directed
435
Fig. 16 Detail of an inner core from a small lamellated
corpusclein the lamina propria of the papilla incisiva. In the
centre of the in-ner core is the nerve terminal (★) with
mitochondria. Pairs of ter-minal glial cell (S) with thin
cytoplasmic processes (L) and basallamina form one half of the
inner core, each separated by a longi-tudinal cleft (arrow) with
collagen fibrils. A thin capsule (C) sur-rounds the inner core of
the corpuscle. ×7360&/fig.c:
-
towards the basal lamina (Halata 1975; Halata 1993). Insinus
hairs their long axis is parallel to the hair shaft,with the nerve
terminal away from the basal lamina(Halata and Munger 1980). In the
monkey hard palate,the alignment is perpendicular to the epithelial
surfacewith the nerve terminal on the labial side of the
Merkelcell, suggesting an optimal arrangement for the percep-tion
of shear forces applied by the tongue. In contrast, inthe large
papilla incisiva of the goat (lacking incisorteeth) Merkel cells
and nerve endings are arranged paral-lel to the mucosal surface as
in the skin (Schwegmann etal. 1993).
Merkel cell receptors are known to have slowly adapt-ing
responses monitoring punctuate pressure (Iggo andMuir 1969).
Characteristic features of Merkel cells haveled to the hypothesis
that the Merkel cell acts as a mech-anoelectric transducer
employing synaptic transmissionto the nerve terminal (Iggo and
Findlater 1984). Thesefeatures could also be seen in the present
study: proto-plasmic protrusions extending between or
invaginatinginto neighbouring keratinocytes, dense core granules
andsynapse-like contacts. However, there is still controversyabout
the precise role of Merkel cells within the mech-ano-electric
transduction process (Mills and Diamond1995; Ogawa 1996; Senok et
al. 1996; Senok andBaumann 1997). In contrast to findings by
Tachibanaand her group (Tachibana et al. 1991; Tachibana et
al.1997; Tachibana et al. 1998), we could not find evidencefor the
presence of so called “transitional” Merkel cellswithout contact to
nerve terminals. All Merkel cellsfound in our study had contact
with nerve terminals.
Ruffini corpuscles
Ruffini corpuscles were found in the deeper layer of thelamina
propria of the papilla incisiva and rugae palati-nae. They are
specially designed to monitor stretch andcan be found ubiquitously
in the connective tissue under-lying the mucosa and skin or in
joint capsules. The nerveterminals are always adjusted to the
structure of the sur-rounding tissue. Larger accumulations of nerve
terminalsare often wrapped by a capsule consisting of either
fibro-blasts or perineural cells (Halata 1988). In contrast,
Ruf-fini corpuscles in the periodontium of teeth (Itoh et al.1981;
Byers 1985; Millar et al. 1989) and those associat-ed with hairs
(Halata and Munger 1980) mainly lacksuch a capsule, which appears
to have no effect on theirfunction. They respond with a slowly
adapting nerve dis-charge (Chambers et al. 1972). The connection
with bun-dles of collagen fibres of the lamina propria makes
themideally suited for this task. In contrast to Ruffini
corpus-cles found in joint capsules (Halata 1988), those in theoral
mucosa and in the periodontium have a thin perineu-ral capsule that
is lacking completely in some places.The usual spindle shape of
Ruffini corpuscles is morediscoid in the palate. In view of the
rather thin layer offirm connective tissue between the mucosa and
bone ofthe hard palate (2.5 to 3 mm) any movement within this
layer is likely to be fairly limited. The presence ofRuffini
corpuscles in this location suggests that shearforces do result in
lateral movement of the mucosaagainst the hard palate of such
dimensions that it needsto be monitored. The ultrastructural
differences betweenRuffini corpusles in the connective tissue of
the hard pal-ate and those in the locomotion apparatus may be an
ad-aptation to the kind of shear forces occurring in this
lo-cation.
Meissner corpuscles
Meissner corpuscles were found only in the papilla incis-iva and
the first two rugae palatinae. They are located inthe lamina
propria just below the basal lamina betweenthe epithelial pegs and
do not possess a perineural cap-sule. Their size depends on the
height of the neighbour-ing epithelial pegs. They are always
supplied by one sin-gle axon, in contrast to those in skin of the
monkey fin-ger tip, which can have up to three supplying nerve
fi-bres and an egg-cup shaped perineural capsule in thelower part
(Halata 1993). In the skin they function asrapidly adapting
mechanoreceptors (Cauna 1956; Iggoand Andres 1982) monitoring
changes in pressure. It islikely that the response characteristics
are the same inthis location. The positioning just below the
epitheliumand the orientation of lamellae parallel to the
surfacemakes them ideally suited for the task of supplementingthe
responses of slowly adapting receptors (i.e. Merkelnerve endings)
to the level of pressure on the mucosawith additional and
independent information about rapidchanges in pressure occurring
during the chewing pro-cess.
Lamellated corpuscles
Lamellated or Pacini-like corpuscles were found in rath-er small
numbers in the lamina propria, mainly in the pa-pilla incisiva.
Often they were found in groups suppliedby one axon. Their
longitudinal axis runs parallel to themucosal surface and the
perineural capsule is thinnerthan in other locations (Zelena 1994).
They are known tobe very rapidly adapting, especially suited for
the per-ception of vibration stimuli (Loewenstein 1971).
Closesimilarities exist to those described in the mucosa of thecat
tongue (Spassova 1974), also called “Krause’s end-bulbs”.
The density of receptors in the mucosa of the hardpalate and in
the papilla incisiva suggests that these dif-ferent receptors play
an important role in sensing the me-chanical properties of food
before swallowing as well asthe position of the tongue within the
mouth. In man thelatter information is likely to be important for
the articu-lation of speech. Usually such a rich supply of
mechano-receptors is only found in the skin of the fingertips.
Herethe active movement for the monitoring of surface prop-erties
plays an important functional role. The hard palate
436
-
does not move itself, but the movement of the tongue orfood
along these receptors is recorded.
&p.2:Acknowledgements This study was supported in part by a
grantfrom Deutsche Forschungsgemeinschaft (Ha 477/633/93). Wewish
to thank Dr. Brian Cooper, Department of Oromaxillar Sur-gery,
State University of Florida, Gainesville Fla., USA, for pro-viding
the monkey palate. The excellent technical skills in prepar-ing the
sections and photographs by Mrs. B. Knuts, Mrs. B. Lücht,Mr. S.
Schillemeit and Dr. A. Spaethe is very much appreciated.We also
wish to thank Mrs. M. Lück for the schematic drawing ofFig. 1.
References
Byers MR (1985) Sensory innervation of periodontal ligament
ofrat molars consists of unencapsulated Ruffini-like
mechanore-ceptors and free nerve endings. J Comp Neurol
231:500–518
Byers MR, Yeh Y (1984) Fine structure of subepithelial “free”
andcorpuscular trigeminal nerve endings in anterior hard palate
ofthe rat. Somatosens Res 1:265–279
Cauna N (1956) Nerve supply and nerve endings of
Meissner’scorpuscles. Am J Anat 99:315–350
Chambers MR, Andres KH, Düring M von, Iggo A (1972) Thestructure
and function of the slowly adapting type II mechano-receptor in
hairy skin. Q J Exp Physiol 57:417–445
Chan KY, Byers MR (1985) Sensory nerve endings of the
incisivepapilla of rat hard palate studied by peroxidase
cytochemicalmethods. J Comp Neurol 234:192–200
Gairns FW (1955) The sensory nerve endings of the human palate.Q
J Exp Physiol 40:40–48
Garant PR, Feldman J, Cho MI, Cullen MR (1980) Ultrastructureof
Merkel cells in the hard palate of the squirrel monkey(Saimiri
sciureus). Am J Anat 157:155–167
Halata Z (1975) The mechanoreceptors of the mammalian
skin.Ultrastructure and morphological classification. Adv
AnatEmbryol Cell Biol 50:1–77
Halata Z (1988) Ruffini corpuscle – a stretch receptor in the
con-nective tissue of the skin and locomotion apparatus. In:Hamann
W, Iggo A (eds) Progress in brain research, vol 74.Transduction and
cellular mechanisms in sensory receptors.Elsevier, Amsterdam New
York, pp 221–229
Halata Z (1993) Die Sinnesorgane der Haut und der
Tiefensensibi-lität. In: Niethammer J, Schliemann H, Starck D,
Wermuth H(eds) Handbook of zoology-Handbuch der Zoologie –
EineNaturgeschichte der Stämme des Tierreiches, vol 8 Part 57.
DeGruyter, Berlin New York, pp 1–81
Halata Z, Munger BL (1980) Sensory nerve endings in Rhesusmonkey
sinus hairs. J Comp Neurol 192:645–663
Hensel H (1973) Cutaneous thermoreceptors. In: Iggo A
(ed)Handbook of sensory physiology. Somatosensory system.Springer,
Berlin Heidelberg New York, pp 79–110
Iggo A, Andres KH (1982) Morphology of cutaneous receptors.Ann
Rev Neurosci 5:1–31
Iggo A, Findlater GS (1984) A review of Merkel cell
mechanisms.In: Hamann W, Iggo A (eds) Sensory receptor
mechanisms.World Scientific Publishing, Singapore, pp 117–131
Iggo A, Muir AR (1969) The structure and function of a
slowlyadapting touch corpuscle in hairy skin. J Physiol (Lond)
200:763–796
Itoh K, Wakita M, Kobayashi S (1981) Innervation of the
peri-odontium in the monkey. Arch Histol Jpn 44:453–466
Kruger L, Halata Z (1996) Structure of nociceptor ‘endings’.
In:Cervero F, Belmonte C (eds) Neurobiology of nociceptors.Oxford
University Press, Oxford New York Tokyo, pp 37–71
Kruger L, Perl ER, Sedivec MJ (1981) Fine structure of
myelinat-ed mechanical nociceptor endings in cat hairy skin. J
CompNeurol 198:137–154
Laczko J, Levai G (1975) A simple differential staining
methodfor semi-thin sections of ossyfying cartilage and bone
tissueembedded in epoxy resin. Mikroskopie 31:1–4
Loewenstein WR (1971) Mechano-electric transduction in the
Pa-cinian corpuscle. Initiation of sensory impulses in
mechanore-ceptors. In: Loewenstein WR (ed) Principles of sensory
physi-ology. Springer, Berlin Heidelberg New York, pp 269–290
Millar BJ, Halata Z, Linden RW (1989) The structure of
physio-logically located periodontal ligament mechanoreceptors
ofthe cat canine tooth. J Anat (Lond) 167:117–127
Mills LR, Diamond J (1995) Merkel cells are not the
mechanosen-sory transducers in the touch dome of the rat. J
Neurocytol24:117–134
Munger BL (1965) The intraepidermal innervation of the snoutskin
of the opossum. J Cell Biol 26:79–97
Ogawa H (1996) The Merkel cell as a possible
mechanoreceptorcell. Prog Neurobiol 49:317–334
Reynolds ES (1963) The use of lead citrate at high pH as an
elec-tron-opaque staining in electron microscopy. J Cell
Biol17:208–212
Schwegmann C, Halata Z, Cooper B (1993) Sensory innervationof
the papilla incisiva in domestic goat. Anat Rec [Suppl]1:102
Senok SS, Baumann KI (1997) Functional evidence for
calcium-induced calcium release in isolated rat vibrissal Merkel
cellmechanoreceptors. J Physiol (Lond) 500:29–37
Senok SS, Baumann KI, Halata Z (1996) Selective phototoxic
de-struction of quinacrine-loaded Merkel cells is neither
selectivenor complete. Exp Brain Res 110:325–334
Spassova I (1974) Ultrastructure of the simple encapsulated
nerveendings (simple end-bulbs of Krause) in the tongue of the
cat.J Anat (Lond) 118:1–9
Tachibana T, Fujiwara N, Sato H, Nawa T (1990) A
comparativeelectron microscopic analysis of mechanoreceptors in the
hardpalate of the mouse (Mus musculus; Rodentia) and the muskshrew
(Suncus murinus; Insectivora). Arch Oral Biol 35:949–956
Tachibana T, Fujiwara N, Nawa T (1991) Mechanoreceptors of
thehard palate of the Mongolian gerbil include special
junctionsbetween epithelia and Meissner lamellar cells: a
comparisonwith other rodents. Anat Rec 231:396–403
Tachibana T, Yamamoto H, Takahashi N, Kamegai T, Shibanai
S,Iseki H, Nawa T (1997) Polymorphism of Merkel cells in therodent
palatine mucosa: immunohistochemical and ultrastruc-tural studies.
Arch Histol Cytol 60:379–389
Tachibana T, Kamegai T, Takahashi N, Nawa T (1998) Evidencefor
polymorphism of Merkel cells in the adult human oral mu-cosa. Arch
Histol Cytol 61:115–124
Watanabe IS, Konig JB (1986) The ultrastructure of Merkel
cor-puscles in the mucosa of the hard palate of rats. Z MikroskAnat
Forsch 100:905–912
Watanabe IS, Yamada E (1988) Fine structure of the
lamellatedcorpuscles in the mouse hard palatine mucosa. Z
MikroskAnat Forsch 102:811–816
Yeh Y, Byers MR (1983) Fine structure and axonal transport
label-ing of intraepithelial sensory nerve endings in anterior
hardpalate of the rat. Somatosens Res 1:1–19
Zelena J (1994) Nerves and mechanoreceptors. Chapmann &
Hall,London New York Tokyo
437