Embed Size (px)
Citation preview

•SEMINARS!!!!Newborn Screening for Enzyme Deficiency Diseases: From Bench to Practice
Michael Gelb, UW
F 3:15 pm. SL 140. BE THERE!

Lyrics
• http://books.google.com/books?id=oq9ENyL_d9YC&lpg=PP1&pg=PA1#v=onepage&q&f=false
http://books.google.com/books?id=oq9ENyL_d9YC&lpg=PP1&pg=PA1#v=onepage&q&f=false



• LEHNINGER • PRINCIPLES OF BIOCHEMISTRY
• Fifth Edition
David L. Nelson and Michael M. Cox
© 2008 W. H. Freeman and Company
CHAPTER 15Principles of Metabolic Regulation


Regulation of Enzyme Activity often occurs by
phosphorylation



Role of AMP-activated protein kinase (AMPK) in carbohydrate and fat metabolism. AMPK is activated by elevated [AMP] or decreased [ATP], by exercise, by the sympathetic nervous system,
or by peptide hormones produced in adipose tissue (leptin and adiponectin). When activated, AMPK phosphorylates target proteins and shifts metabolism away from energy-consuming
processes; shifts metabolism in extrahepatic tissues to the use of fatty acids as a fuel; and triggers gluconeogenesis in the liver to provide glucose for the brain. In the hypothalamus, AMPK stimulates
feeding behavior to provide more dietary fuel.


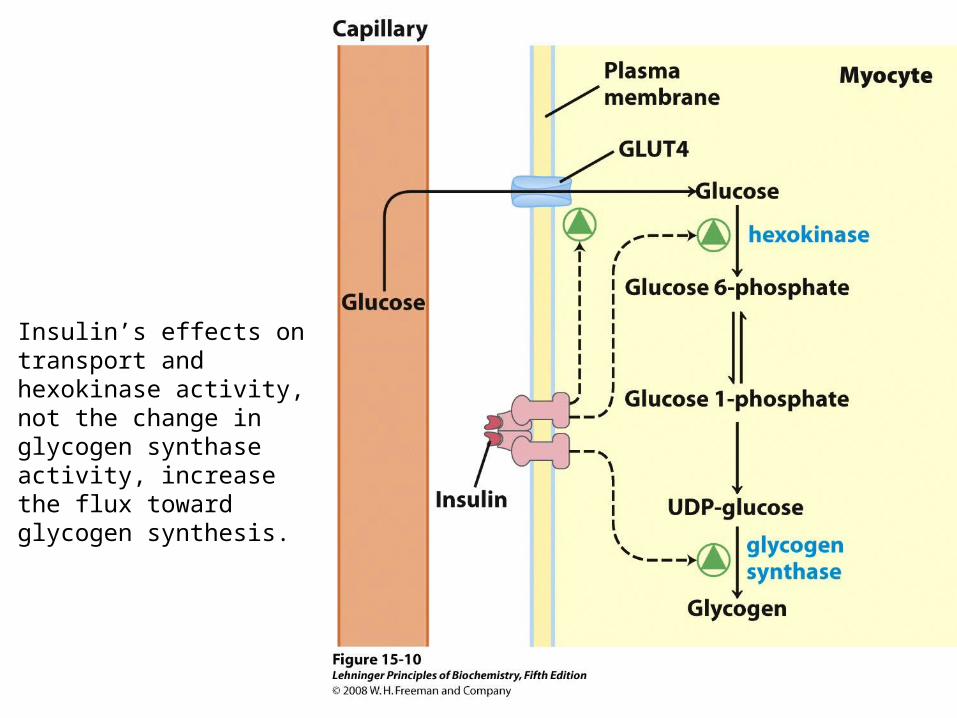
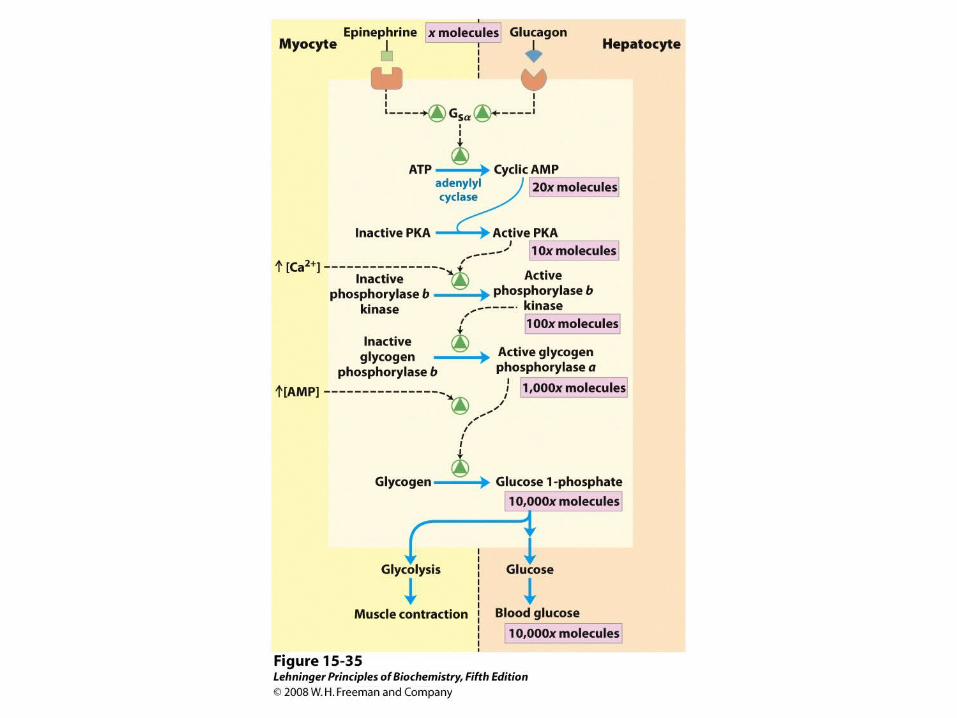
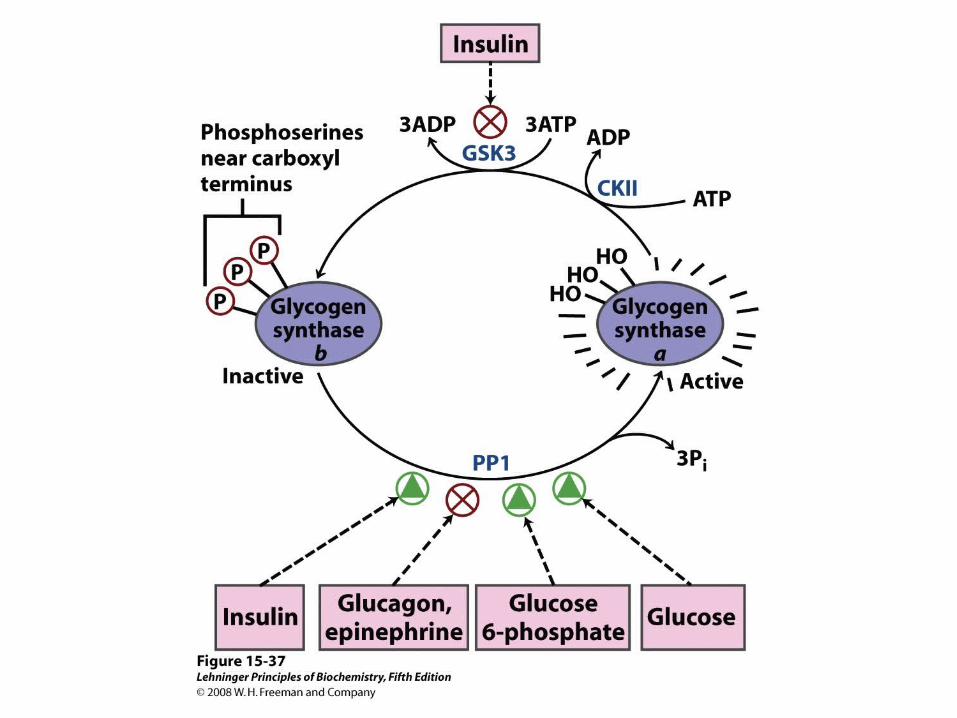
Insulin’s effects on transport and hexokinase activity, not the change in glycogen synthase activity, increase the flux toward glycogen synthesis.

Describe these differences!
When blood glucose rises above 5 mM, hexokinase IV activity increases, but hexokinase I is already operating near Vmax and cannot respond to an increase in glucose concentration.
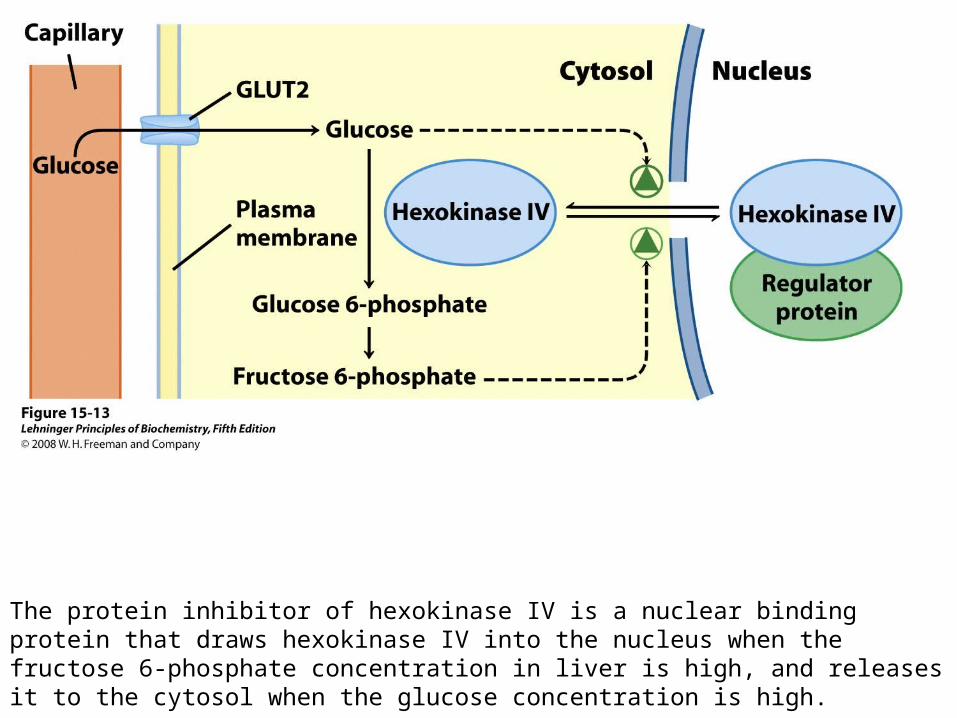
The protein inhibitor of hexokinase IV is a nuclear binding protein that draws hexokinase IV into the nucleus when the fructose 6-phosphate concentration in liver is high, and releases it to the cytosol when the glucose concentration is high.

FIGURE 15-14a Phosphofructokinase-1 (PFK-1) and its regulation. Ribbon diagram of E. coli PFK-1, showing two of its four identical subunits (PDB ID 1PFK). Each subunit has its own catalytic site, where the products ADP and fructose 1,6-bisphosphate are almost in contact, and its own binding sites for the allosteric regulator ADP (blue), located at the interface between subunits.
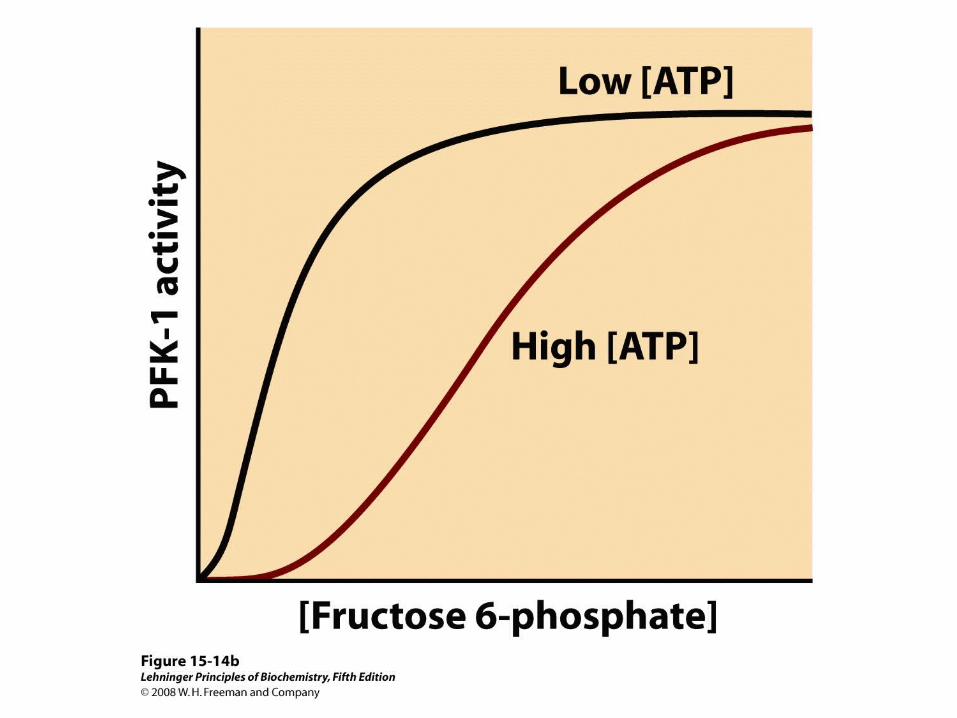
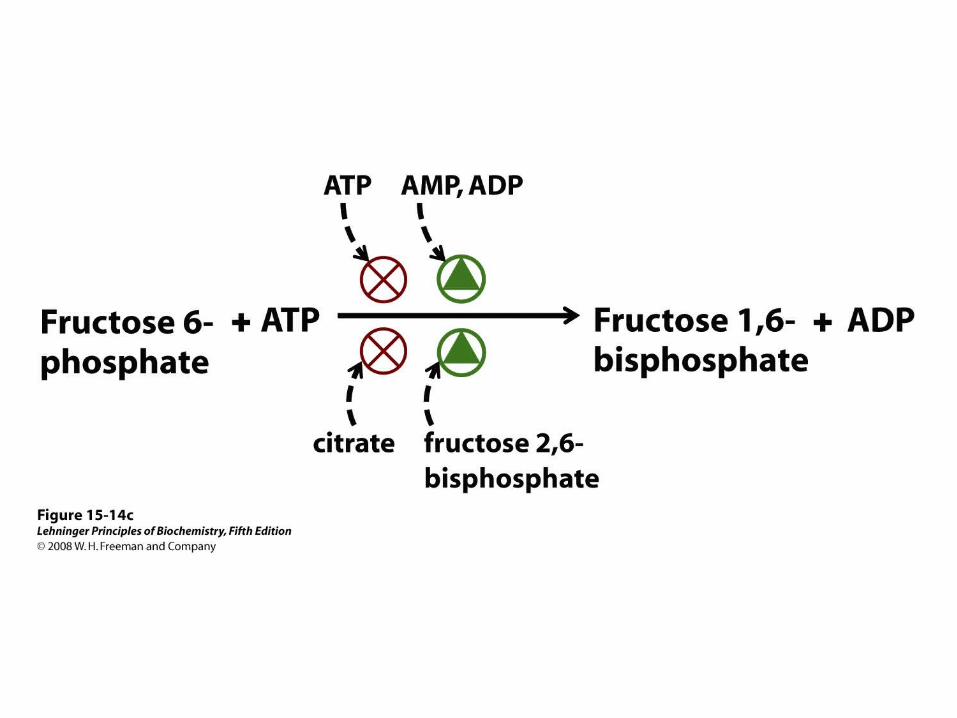



Fructose 2,6-bisphosphate (F26BP) has opposite effects on the enzymatic activities of phosphofructokinase-1 and fructose 1,6-bisphosphatase.
(a) PFK-1 activity in the absence of F26BP is half-maximal when the concentration of fructose 6-phosphate is 2 mM. When 0.13 μM F26BP is present, the K0.5 for fructose 6-phosphate is only 0.08 mM.
(b) FBPase-1 activity is inhibited by as little as 1 μM F26BP and is strongly inhibited by 25 μM. In the absence of this inhibitor the K0.5 for fructose 1,6-bisphosphate is 5 μM, but in the presence of 25 μM F26BP the K0.5 is >70 μM. Fructose 2,6-bisphosphate also makes FBPase-1 more sensitive to inhibition by another allosteric regulator, AMP.


Both enzyme activities are part of the same polypeptide chain, and they are reciprocally regulated by insulin and glucagon.
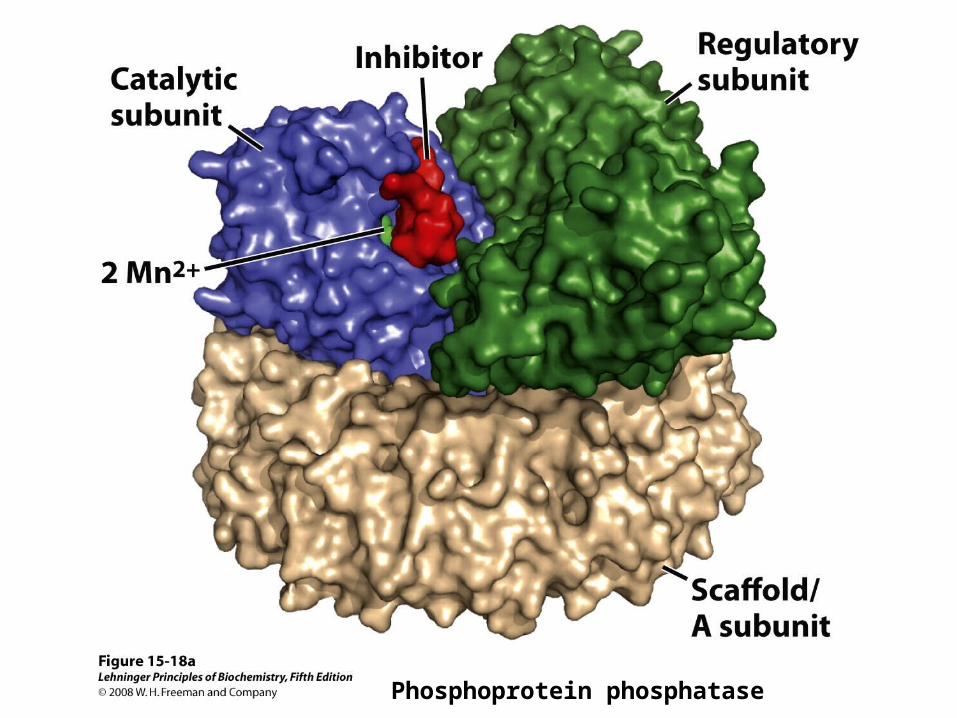
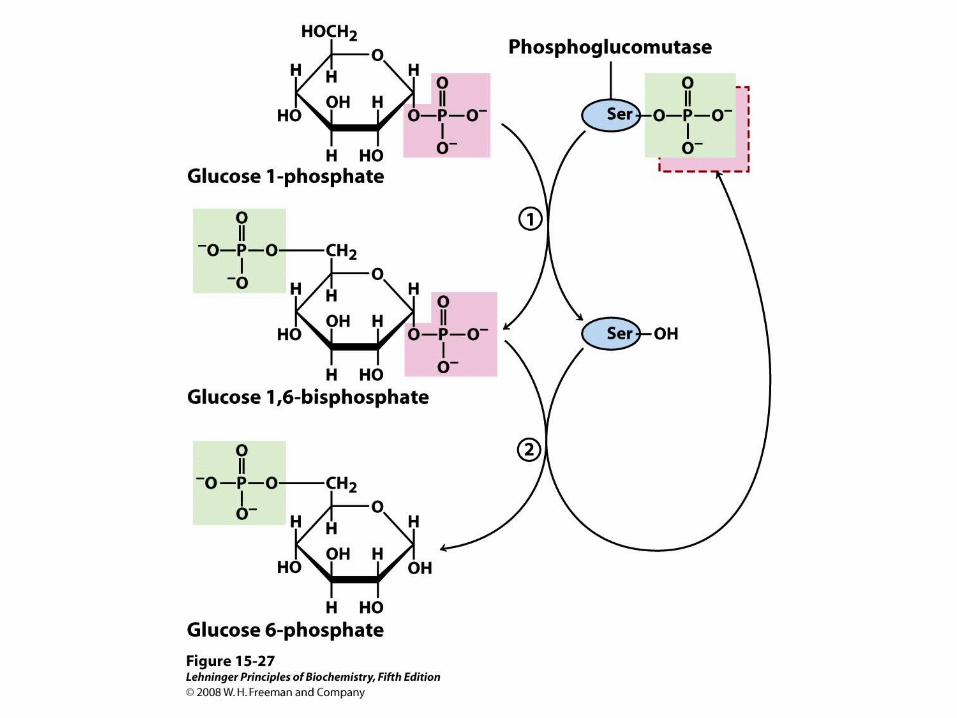
Phosphoprotein phosphatase



The PEP carboxykinase promoter region, showing the complexity of regulatory
input to this gene.
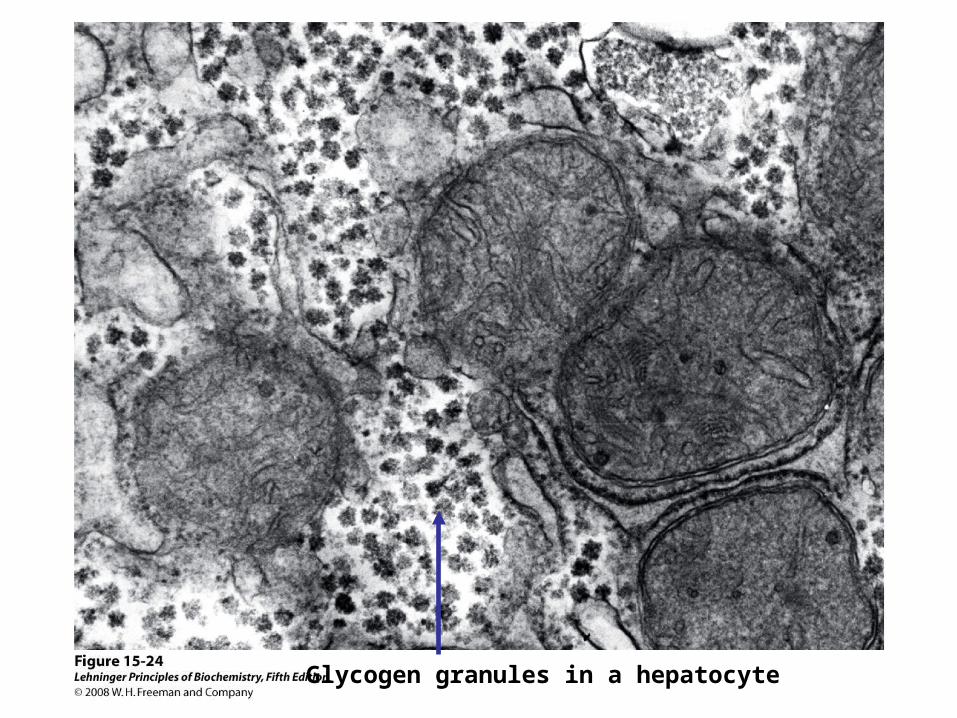
Glycogen granules in a hepatocyte






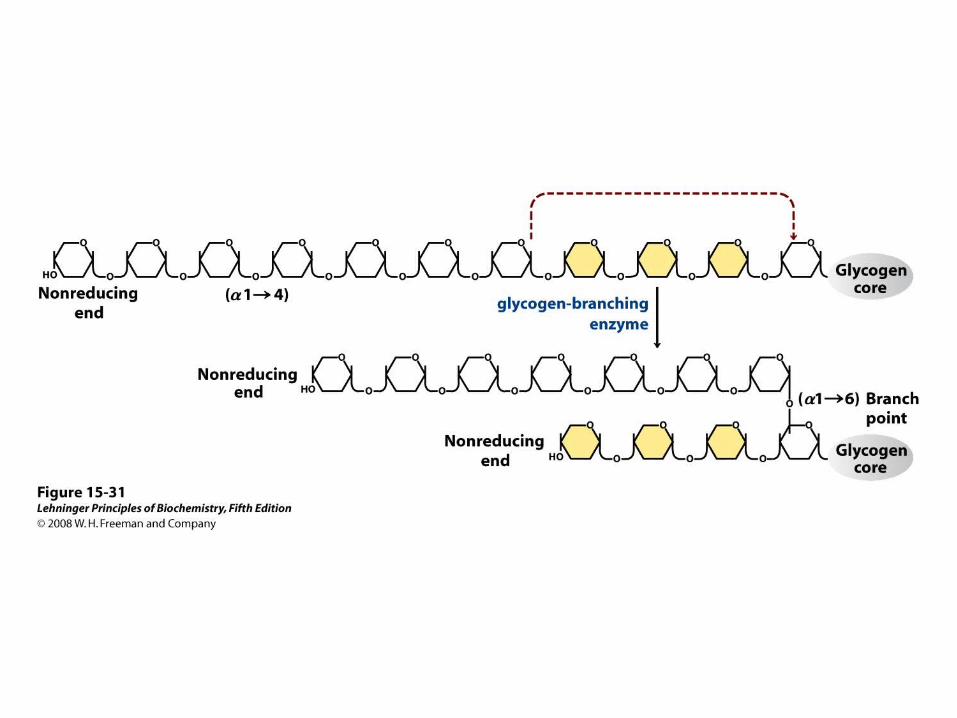

Glycogenin structure. Muscle glycogenin (Mr 37,000) forms dimers in solution. The substrate, UDP-glucose, is bound to a Rossmann fold near the amino terminus and is some distance from the Tyr194 residues—15 Å from the Tyr in the same monomer, 12 Å from the Tyr in the dimeric partner. Each UDP-glucose is bound through its phosphates to a Mn2+ ion that is essential to catalysis. Mn2+ is believed to function as an electron-pair acceptor to stabilize the leaving group, UDP. The glycosidic bond in the product has the same configuration about the C-1 of glucose as the substrate UDP-glucose, suggesting that the transfer of glucose from UDP to Tyr194 occurs in two steps. The first step is probably a nucleophilic attack by Asp162, forming a temporary intermediate with inverted configuration. A second nucleophilic attack by Tyr194 then restores the starting configuration













“Alfonse, Biochemistry makes my head hurt!!”\















![Newborn Screening: Because you touch the future everyday Guidelines.… · Newborn Screening ACT Sheet [Absent/Reduced biotinidase activity] Biotinidase Deficiency Differential Diagnosis:](https://img.dokumen.tips/doc/110x75/5f03bfe47e708231d40a94f1/newborn-screening-because-you-touch-the-future-everyday-guidelines-newborn-screening.jpg)














