Embed Size (px)
Citation preview

Eur. J. Biochem. 194, 693-698 (1990) 0 FEBS 1990
Review
Self recognition by the immune system Harald VON BOEHMER
Basel Institute for Immunology, Basel, Switzerland
(Received July 3, 1990) - EJB 90 0776
In each organism, the immune system must acquire the ability to distinguish self from nonself. Experiments in T cell receptor transgenic mice indicate that this process involves the selection of lymphocytes in the thymus.
The immune system is often regarded as a system that protects us from harmful microorganisms such as bacteria and viruses. Apart from this beneficial effect, the immune system can also cause grief because it can reject transplanted organs such as a kidney or a heart.,While the immune system can attack the kidney from a foreign donor, or nonself, it usually does not attack organs from the same organism, or self. This means that the immune system can distinguish be- tween self and nonself. The aim of the experiments described here was to elucidate the principles and mechanisms of this distinction. In order to understand these experiments, one needs to know a few facts about the components as well as the development of the immune system.
CELLS AND MOLECULES OF THE IMMUNE SYSTEM The immune system is, to a great part, represented by a
portion of white blood cells called lymphocytes. Morphologi- cally, one lymphocyte looks like another, but in reality each lymphocyte differs from all others because it carries a unique receptor on its surface. These receptors allow lymphocytes to recognize specific antigens. The antigen receptors are proteins that consist of constant and variable parts. The variable part binds antigen and makes each lymphocyte unique [I - 41. Anti- gen can be anything, such as toxins from bacteria, proteins from viruses or even synthetic chemicals that do not occur in nature.
There are various classes of lymphocytes which have differ- ent roles in the immune system. The most ‘cruel’ class consists of so-called killer lymphocytes [5]. Their major task is to screen other cells in the body, whether they are infected by virus, develop into cancer cells or are abnormal in any other way. The antigen receptors of these cells recognize other cells con- taining fragments of proteins, called peptides, which should not be there. Proteins are synthesized and degraded in cells, and peptides are part of the proteins. By an astonishing mech- anism, the killer lymphocytes can be informed of what is going on inside other cells in the body. For instance, when a cell is infected with a virus, the virus has entered the cell and multi-
Correspondence to H. von Boehmer, Basel Institute for Immu-
Abbreviations. MHC, major histocompatibility complex; TCR, T nology, Grenzacherstrasse 487, CH-4005 Basel, Switzerland
cell receptor.
plies inside the living cell. Although it is inside the cell, the virus is not hidden from the immune system: peptides from the virus will bind to so-called major histocompatibility complex (MHC) molecules inside the cell, which then transport the peptides to the cell surface and present them to a killer lym- phocyte, or better, a pre-killer cell with an appropriate antigen receptor (Fig. 1) [6-91. This antigen receptor is composed of CI and p receptor chains [lo] and binds to both the peptide as well as the presenting MHC molecule: by receptor-gene transfection experiments it could be shown that, in fact, one c@ receptor has specificity for both the peptide and the presenting MHC molecule [ I l , 121. The binding of the ap receptor to MHC plus peptide is, however, not sufficient to activate the pre-killer cell. The activation requires another nonvariable receptor on the pre-killer cell, the CD8 coreceptor which also binds MHC molecules (Fig. 1) [13, 141. When the variable up receptor and the invariable CD8 coreceptor are brought together by binding to different sites on the same MHC mol- ecule [15, 161, the pre-killer cell receives a signal to divide and to differentiate, such that the initially inactive pre-killer lymphocyte produces many active killer cells which can lyse the virus-infected target cell. This makes the virus accessible to other components of the immune system, normally the antibodies, that will eliminate it. The role of the killer cell in the elimination of a virus is, in fact, twofold: first, it deprives the virus of its support, the living cell, and second, it makes the virus accessible to antibodies. Without the killer cells the virus could multiply enormously inside the cell and finally destroy it. Thousands of virus particles could then infect other cells, and this cycle could be repeated until the organism is destroyed.
When discussing the activation of a pre-killer cell by a foreign peptide, one should realize that the MHC molecules not only present peptides derived from foreign proteins, but also peptides from proteins that are normal constituents of the cell. This is the reason why, when a transplanted kidney comes from a genetically distinct donor whose cell proteins differ from that of the recipient, pre-killer cells will be acti- vated by peptides from nonself proteins, and destroy the kid- ney.
Very briefly, two other classes of lymphocytes are con- sidered here, namely the helper cells and the antibody-produc- ing cells. Killer and helper lymphocytes are called T lympho- cytes because they are formed in the thymus [17]. The anti-

694
,v
killer cell killer cell
4
'perforin 8 coreceptor . .
c 7
Fig. 1 . The activution oj a pre-killer T lymphocyte by peptides and M H C molecule.s,fiorn a virus infected cell. A pre-killer T lymphocyte with an x p TCR (white) and a CD8 coreceptor (grey) binds to both the MHC molecule (black) as well as the peptide (triangle) which is derived from a virus protein made inside the cell. The peptide binds to the MHC molecule inside the cell and the whole complex is trans- ported to the cell surface and 'presented' to the T cell. As a result of the binding of both the ap TCR and the CD8 coreceptor to the same MHC molecule, the pre-killer cells receives a signal, starts to divide and differentiate into an active killer cell. When the receptor of an activated killer cell binds to MHC molecules and peptides, the killer cell will produce a substance called perforin which destroys the virus- infected cell. Note that MHC molecules not only present peptides from foreign proteins as shown here, but also peptides from proteins which occur normally in cells
body-producing cells are formed in the bone marrow and are called B lymphocytes. T helper cells and B cells cooperate in the antibody response to antigens, like bacterial toxins, which again are proteins [18]. Once a toxin is released into the blood stream, it will be taken up by macrophages; cells which ingest a large variety of substances. Inside the macrophage, the toxin will be degraded, and peptides from the toxin will be presented by MHC molecules. There are two types of MHC molecules: the class I MHC molecules that present peptides from proteins which are made inside the cell (like viral proteins), and the class I1 MHC molecules that present peptides from proteins which enter the cell (like bacterial toxins) [8]. The ap T cell receptor (TCR) of a T helper cell binds to the class-11-MHC- presented peptide. and another invariable CD4 coreceptor 1191, characteristic of pre-helper cells, binds to the same class 11 MHC molecule [20] (like the CD8 coreceptor of a killer cell binds to class I MHC molecules). The pre-helper cell divides and differentiates, and in this way produces many active helper cells (Fig. 2). In the meantime, the toxin has also bound to a specific antibody molecule on a B cell and the whole complex, i.e. the antibody and the toxin, has been ingested by the B cell. The complex is degraded and peptides from the toxin are again transported to the cell surface by class I1 MHC mol- ecules [21]. Such B cells are now detected by the active T helper cells whose $? receptors and CD4 coreceptors bind to the peptide MHC complex at the surface of the B cell. As a result, the T helper cells produce factors, called interleukins, that stimulate the B cells to divide, differentiate and secrete large amounts of antibodies which can neutralize the free toxin in the blood stream (Fig. 2). Thus, both T killer cells as well as
bacterial toxins antibodies
Fig. 2. The activation of a pre-helper cell and antibody-producing B cells by a bacterial toxin. A bacterial toxin (square with triangle) is ingested and degraded into peptides by a macrophage. A certain peptide (triangle) binds to a class I1 MHC molecule (black) and is presented to a pre-helper cell. As a result of the binding of both the L@' TCR (white) and the CD4 coreceptor (black) to the same class I1 MHC molecule, the pre-helper cell receives a signal, starts to divide and differentiate into an active helper cell. A B cell can only internalize the toxin when it is bound by an antibody on the surface of a cell. Inside the B cell, the toxin is degraded and peptides are transported to the cell surface by class I1 MHC molecules. When the c$ TCR and the CD4 coreceptor of an activated helper cell bind to a class I1 MHC molecule and peptide, the helper cell will produce interleukins which stimulate the B cell to divide and secrete large amounts of antibodies. These antibodies can then bind to free toxins and neutralize them
T helper cells have a variable ap TCR, while killer cells have an invariable CD8 and helper cells have an invariable CD4 receptor. T cells can only be activated by peptides presented by MHC molecules because T cell activation does not require binding of the aP TCR, but also binding of the CD4 or CD8 to the same class I1 or class I MHC molecule, respectively. T cell activation does not occur when the ap TCR binds to one MHC molecule, and the CD4 or CD8 coreceptor to another MHC molecule on the same cell.
Fig. 3 illustrates how an up TCR might bind to a peptide presented by MHC molecules [12]. One important feature of antigen recognition by the immune system that is not illus- trated in this figure is the great variability on the interacting molecules. Thus, a very large number of different peptides can be recognized which involve an equally large number of TCR and a diverse set of MHC molecules, each of which can present many, but not all, peptides. The variability of the antigen receptor is somatically generated : the genes encoding these receptors do not exist as a continuous stretch of genetic infor- mation but occur in the germline as little pieces or gene seg- ments. These segments recombine randomly in lymphocytes, and by this mechanism form a very large number of different genes encoding in the order of 10' different receptors [22] (Fig. 4). The variability of the MHC molecules is largely an allelic variability; this means that one individual contains a limited set of different MHC molecules and that different individuals have different allelic forms of this set 1231.
Both the randomly generated receptor diversity and the allelic MHC polymorphism preclude a heritable basis for self/ nonself discrimination by the immune system. Thus, in each

69 5
The Molecules of the Immune System Tlieproteiiis that mcngnizr fi>rri& invaders arr the must diverse purr ins known. Tliry are encoded by hundreds ofscarreredgene trngments, which ciln he cumhinrd in rnil/ions or billions of ways
by Surumu TonegPws
Fig. 3. A computer model of how an a/3 T C R couldpossibly bind to a class I MHC molecule andpeptide. Taken from [12]. In the lower part, a class I MHC molecule containing a peptide (circular structure in the middle) is shown. In the upper part the TCR is aligned over the MHC molecule. Contacts between the MHC molecule, peptide and T cell receptor are in white
Fig. 4. The title of and a picture f rom an article written by Susumu Tonegawa [53/ on the origin of antibody diversity. While the picture describes the gene segments encoding antibodies, very similar events are involved in the generation of TCRs
NORMAL MOUSE TCELLRECEPTORTRANS- GENIC SClD MOUSE
-10' DIFFERENT RECEP- NO LYMPHOCYTES I 1 RECEPTOR TORS
Fig. 5 . The construction of T C R transgenic scid mice. A normal mouse may contain as many as lo8 different TCRs (left). In a scid mouse, the rearrangement mechanism, generating productive TCR-antibody and antibody genes, is defective. As a consequence, this mouse does not produce any antigen receptors nor lymphocytes. The introduction of productively rearranged CI and /3 TCR genes cures the defect, at least partially, and T lymphocytes, all expressing the same receptor, are generated
individual, the immune system must 'learn' to distinguish self from nonself. With regard to T cell receptors, this means that initially, the developing immune system also generates receptors specific for self peptides presented by self MHC molecules. Cells with these receptors are potentially harmful to the organism and thus, the immune system must have ways to preclude the activation of such cells. The developing immune system will also generate receptors which cannot bind to self MHC molecules at all. Cells with these receptors are useless to the organism because they cannot detect foreign peptides presented by self MHC molecules.
This scenario of antigen recognition poses two major ques- tions with regard to self/nonself dsicrimination. First, what happens to potentially harmful pre-killer cells which have receptors specific for self peptides presented by self MHC
molecules? Second, what happens to useless T cells which cannot bind to self MHC molecules at all, and therefore can- not recognize nonself peptides presented by self MHC mol- ecules?
SELFiNONSELF DISCRIMINATION IN T CELL RECEPTOR TRANSGENIC MICE
We have learned from experiments during the last decade that one cannot study these questions in any conclusive detail in normal animals, because normal animals have about lo8 different T cell receptors [24-331. For this reason it is very difficult to follow a single cell with one particular receptor throughout lymphocyte development. To overcome this prob-

696
@'
-1 4'8'
000 000 000
aD
--- Fig. 6. T cell development in the thymus. The thymus is represented by the rectangle. Large dividing cells (lower left) enter the thymus and begin to express TCR (symbolized by /) as well as CD4 and CD8 coreceptors. As a consequence many CD4'8' immature thymocytes are produced ( 5 x 10' cells/day). Only a few mature further and be- come CD4'8- pre-helper and CD4-8' pre-killer cells which leave the thymus (1 x 10' cellsiday)
t
0 n
MHCDb+. MALE NORMAL THYMUS
d d 0
CD8 CD8 Fig. I. The cellular composition of the thymus of u TCR transgenic I?IOUSP ( u normul thjniuS for coipar2ron on the right handside) express- ing both the peptide (mule untigrn) und the M H C molecule (class I M H C Dh) recognized by the transgenic TCR. The thymocytes are stained by antibodies against CD4 (red fluorescence on y-axis) and against CD8 (grecn fluroescence on x-axis) coreceptor. CD4'8' thymocytes are eliminated (deletion of harmful cells at an immature stage)
lem, we decided to study self/nonself discrimination in TCR transgenic mice which have only one receptor with one known specificity [34-381. We have constructed such a mouse by introducing productively rearranged CY and p TCR genes from one particular killer lymphocyte into a mouse strain which suffers from a genetic defect, the scid mutation. scid stands for severe combined immune deficiency, which results from the inability of this mouse strain to properly rearrange antigen receptor gene segments. Therefore, mice from this strain can- not produce any antigen receptors [39]. The introduction of one functionally rearranged r and one functionally rearranged B TCR gene into the germ line of this mouse strain should result in a new strain with an immune system expressing only one receptor instead of lo8 (Fig. 5). With the help of various collaborators we produced such a strain and could address questions concerning the selfinonself discrimination of the immune system in a much more conclusive way.
For our purposes we chose a receptor specific for an intra- cellular peptide that is present in male but absent in female mice (HY peptide). and which is presented by Db MHC mol-
MHCD~ +, FEMALE NORMAL THYMUS
d
0 n
d
0 n
CD8 CD8 Fig. 8. The cellular composition of the thymus of a TCR transgenic mouse (a normal thymusfor comparison on the right handside) express- ing the M H C molecule (class I M H C Db) hut not the peptide (male antigen) recognized by the transgenic TCR. Staining was carried out as in Fig. 7 . CD4-8' pre-killer cells are selected (selection of useful cells) from CD4'8 ' precursors. Specificity of receptor determines whether cells become CD4' helpers and CD8+ killers. No CD4'8- pre-helper cells are detectable
ecules. The genes encoding this receptor were isolated from a killer cell with a CD8 coreceptor and the productively re- arranged CI and p TCR genes were introduced into scid mice. In various scid mice we could then analyze the fate of T lymphocytes expressing the transgenic receptor : we could study the fate of potentially harmful cells in transgenic mice that have both the HY peptide as well as H-2Db MHC mol- ecules. We could study the fate of potentially useful cells in female transgenic mice that do not have the HY peptide, but do have Db MHC molecules. Finally, we could analyze the fate of the useless T cells in female transgenic mice, that lack both the HY peptide as well as Db MHC molecules. The various transgenic mice were obtained by introducing the T cell receptor transgenes in one particular mouse strain, and by further crossing this transgenic strain with other nontransgenic strains with MHC molecules which were appro- priate for our purposes.
T LYMPHOCYTE DEVELOPMENT IN THE THYMUS
For a better understanding of the results obtained with the various transgenic mice, a few facts about T cell development need to be introduced here (Fig. 6). T cell development takes place in the thymus. Throughout life the thymus is colonized by hemopoietic cells that contain the precursors of T cells (40, 411. As soon as the precursors enter the thymus they start to rearrange their receptor gene segments and express receptors and coreceptors on the cell surface [42-441. Initially, the immature T cells each express one particular ag TCR and CD4 as well as CD8 coreceptors. These cells are produced in large quantities (in a mouse 5 x lo7 cellsiday) and most of them die within a few days [45]. A few of them, however, become mature pre-killer or pre-helper cells with CD8 and CD4 coreceptors, respectively, and leave the thymus (Fig. 6). These mature cells can be stimulated by antigen to become effector cells 1461, while their immature CD4'8' precursors cannot be induced in this way, even though they express a/3 TCR, as well as both coreceptors on the cell surface [16, 471. The overproduction of immature T cells has been suspected to reflect some cellular selection in the thymus [47]. The analy- sis of the various transgenic mice has confirmed this suspicion
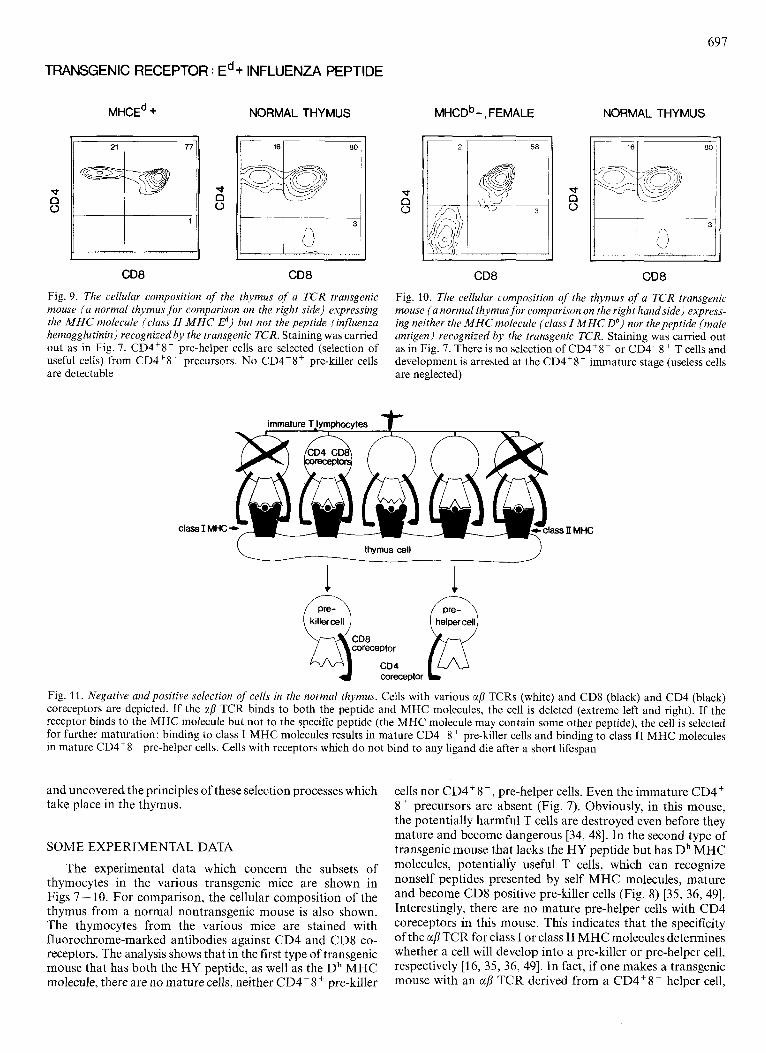
697
TRANSGENIC RECEPTOR : Ed + INFLUENZA PEPTIDE
MHCE~ + NORMAL THYMUS
1 J I I
CD8 CD8
Fig. 9. The cellular composition of the thymus of a T C R transgenic mouse ( a normal thymus for compurison on the right side) expressing the M H C molecule (class II M H C E d ) but not the peptide (influenza hemagglutinin) recognized by the transgenic TCR. Staining was carried out as in Fig. 7. CD4'8- pre-helper cells are selected (selection of useful cells) from CD4'8' precursors. No CD4-8' pre-killer cells are detectable
class I MIX
MHCD~-, FEMALE NORMAL THYMUS
CD8 CD8
Fig. 10. The cellular composition of the thymus of a TCR transgenic mouse ( a normal thymus fo r comparison on the right handside) express- ing neither the M H C molecule (class I M H C Db) nor thepeptide (male antigen) recognized by the transgenic TCR. Staining was carried out as in Fig. 7. There is no selection of CD4'8- or CD4-8' T cells and development is arrested at the CD4'8' immature stage (useless cells are neglected)
,11 MHC
thymus cell
1 1 QcD*cD4 killercell coreceptor @
coreceptor
Fig. 11. Negative and positive selection of cells in the normal thymus. Cells with various afi TCRs (white) and CD8 (black) and CD4 (black) coreceptors are depicted. If the afi TCR binds to both the peptide and MHC molecules, the cell is deleted (extreme left and right). If the receptor binds to the MHC molecule but not to the specific peptide (the MHC molecule may contain some other peptide), the cell is selected for further maturation: binding to class I MHC molecules results in mature CD4-8' pre-killer cells and binding to class I1 MHC molecules in mature CD4'8- pre-helper cells. Cells with receptors which do not bind to any ligand die after a short lifespan
and uncovered the principles of these selection processes which take place in the thymus.
SOME EXPERIMENTAL DATA
The experimental data which concern the subsets of thymocytes in the various transgenic mice are shown in Figs 7 - 10. For comparison, the cellular composition of the thymus from a normal nontransgenic mouse is also shown. The thymocytes from the various mice are stained with fluorochrome-marked antibodies against CD4 and CD8 co- receptors. The analysis shows that in the first type of transgenic mouse that has both the HY peptide, as well as the Db MHC molecule, there are no mature cells, neither CD4-8 + pre-killer
cells nor CD4+8-, pre-helper cells. Even the immature CD4' 8' precursors are absent (Fig. 7). Obviously, in this mouse, the potentially harmful T cells are destroyed even before they mature and become dangerous [34, 481. In the second type of transgenic mouse that lacks the HY peptide but has Db MHC molecules, potentialiy useful T cells, which can recognize nonself peptides presented by self MHC molecules, mature and become CD8 positive pre-killer cells (Fig. 8) [35, 36, 491. Interestingly, there are no mature pre-helper cells with CD4 coreceptors in this mouse. This indicates that the specificity of the ap TCR for class I or class 11 MHC molecules determines whether a cell will develop into a pre-killer or pre-helper cell, respectively [16, 35, 36, 491. In fact, if one makes a transgenic mouse with an aj3 TCR derived from a CD4'8- helper cell,

698
one obtains only CD4+8-, but not CD4'8- mature thymocytes in the trangenic mouse (Fig. 9) [50, 511. Finally, in the third mouse which lacks both the HY peptide as well as the Db MHC molecule, cells expressing the transgenic recep- tor are useless. They die after a short lifespan and mature T cells do not emerge (Fig. 10) [37]. Thus, it is the interaction of the TCR with peptides and MHC molecules in the thymus that determine whether a cell is potentially harmful and, therefore, deleted, potentially useful and selected for maturation, or useless and not selected at all [52].
CONCLUSION
Extrapolating from these data, we arrive at the following picture of self/nonself discrimination in a normal mouse (Fig. 11). Immature CD4'8' thymocytes express a large va- riety of receptors that are generated by random recombination of receptor gene segments and x and f l TCR chain pairing. If the receptor does not bind to any ligand in the thymus, the cell will die after about three days. If the receptor binds to both the peptide and the presenting MHC molecule, the cell will be deleted. If the receptor binds to the MHC molecule only (which may be occupied by some other peptide), the cell will be selected for further maturation. Depending on whether the receptor binds to a class I or class I1 MHC molecule, the selected cell will assume the CD4-8' or CD4'8- phenotype, respectively. Thus. the initial learning by the immune system does involve selection of lymphocytes: the useful are selected, the useless are neglected and the harmful are deleted.
Prof. Pawel Kisielow's contribution to these studies is especially acknowledged. Other important collaborators include Drs Anton Berns, Horst Bliithmann, Peter Borgulya, Hiroyuki Kishi, Bernadette Scott, Michael Steinmetz, Hung-Sia Teh and Yasushi Uematsu. The expert technical support by Katrin Hafen and Verena Stauffer throughout the conduction of these experiments is gratefully acknowl- edged. The help of Nicole Schoepflin in the preparation of this manu- script is also gratefully acknowledged.
REFERENCES 1. 2. 3. 4. 5 .
6.
7.
8.
9.
10.
11.
12. 13.
Jerne, N. K . (1955) Proc. Nut1 Acad. Sci. USA 41, 849-857. Burnet, F. M. ( 1 957) Aust. J . Sci. 20, 67. Edelman, G. M. (1970) Sci. Am. 223, 34-42. Jerne. N . K. (1973) Sci. Am. 229, 52-60. Cerottini, J . C.. Nordin, A. A. & Brunner, K. T. (1970) Nature
Babbitt, B. P., Allen, P. M., Matsueda, G., Haber, E. & Unanue,
Buus, S., Sette. A., Colon, S. M., Jenis, D. M. & Grey, H. M.
Townsend, A. R., Rothbard, J., Gotch. F. M., Bahadur, G. &
Bjorkman. P. S., Saper, M. A,, Samraoni, B., Bennett, W. S. &
Allison, J. P. & Lanier. L. L. (1987) Annu. Rev. Immunol. 5, 503-
Dembic, Z., Haas, W., Wciss, S., McCubrey, J. & Kiefer, H.
Davis, M. M. & Bjorkman, P. J. (1988) Nature 334, 395-402. Dembic, 2.. Haas. W., Zamoyska, R., Parnes, J. & Steinmetz, M.
227, 72 - 73.
E. R. (1985) N a t ~ ~ 3 1 7 , 359-361.
(1986) Cell 47. 1071 -1077.
Wraith, I). (2986) Cell44, 959-968.
Strominger, J. L. (1987) Nature 329, 509-512.
540.
(1986) NCLtur(J 320, 232-238.
(1987) Nature 326, 510-511.
14. Connolly, S. M., Potter, T. A., Wormstall, E. & Hansen, T. H.
IS. Emmerich, F., Strittmatter, V. & Eichmann, K. (1986) Proc. Nut1
16. von Boehmer, H. (1986) Immunol. Today 7, 333-336. 17. Miller, J. F. A. (1961) Lancet 2, 748. 18. Mitchell, G. F. &Miller, S. F. (1968) J . Exp. Med. 128, 821 -837. 19. Dialynas, D. P., Quan, Z. S., Wall, K. A., Pierres, A. & Quintans,
S. (2983) J . Immunol. 131,2445-2451. 20. Doyle, C. & Strominger, J. L. (1987) Nature 330. 256-259. 21. Lanzavecchia, A. (1985) Nature 314, 537-539. 22. Tonegawa, S. (1983) Nature 302, 575 - 581. 23. Klein, J. (1979) Science 203, 516-521. 24. von Boehmer, H. & Sprent, J . (1976) Transplant. Rev. 29, 3-23. 25. Bevan, M. J. (1977) Nature 269,417-418. 26. Zinkernagel, R. M., Callahan, G. N., Althage, A,, Cooper, S. &
27. von Boehmer, H., Haas, W. & Jerne, N. K. (1978) Proc. Nut1
28. Zinkernagel, R. M., Althage, A,, Waterfield, E., Kindred, B. &
29. Wagner, H., Hardt, C., Stockinger, H., Pfizenmaier, K. & Bartlett,
30. Matzinger, P. (1981) Nuture 292, 497- 501. 31. von Boehmer, H., Teh, H. S., Bennick, H. R., Haas, W. (1985) in
Recognition and regulation in cell-mediated immunity (Watson, J. D. & Mabrook, J., eds) pp. 89 - 101, Dekker, New York and Basel.
(1988) J . Exp. Med. 168,325-341.
Acad. Sci. USA 83, 8298 - 8302.
Klein, P. A. (1978) J . E.xp. Med. 147. 882-896.
Acad. Sci. USA 75, 2439 - 2442.
Welsh, R. M. (1980) J. Exp. Med. 151, 376-399.
R. (1981) Immunol. Rev. 58,95-129.
32. Ishii, N., Nagy, Z. A. & Klein, J. (1982) Nature 295, 531 - 533. 33. Goverman, J., Hunkapillar, T. & Hood, L. E. (1986) Cell 45,
34. Kisielow, P., Bliithmann, H., Staerz, U. D., Steinmetz, M. & von
35. Teh, H. S., Kisielow, P., Scott,B., Kishi, H. & Uematsu, Y. (1988)
36. Kisielow, P., Teh, H. S., Bliithmann, H. & von Boehmer, H. (1988)
37. Scott, B., Bliithmann, H.. Teh, H. S. & von Boehmer, H. (2989)
38. von Boehmer, H. (1990) Annu. Rev. Imrnunol. 8, 531 -556. 39. Bosma, G. C., Custer, P. R. & Bosma. M. J . (1983) Nature 301,
40. Moore, M. A. &Owen, J. J. (1967) J . Exp. Med. 126, 715-726. 41. Le Douarin, N. M. & Jotereau, F. V. (2975) J . Bxp. Med. 142,
42. Snodgrass, R. H., Dembic, Z., Steinmetz, M. & von Hoehmer, H.
43. Raulet, D. H., Garman, R. D., Saito, H. & Tonegawa, S. (1985)
44. Crisanti, A,, Colantoni, A,, Snodgrass, R. H. & von Boehmer, H.
45. Shortman, K. &Jackson, H. (1974) Cell. Immunol. 12, 230-246. 46. Kisielow, P., von Boehmer, H. & Haas, W. ( 1 982) ELK J . Immunol.
47. von Boehmer, H. (1988) Annu. Rev. Immunol. 6,309-326. 48. Sha, W. C., Nelson, C. A,, Newberry. R. D., Kranz, D. M. &
49. Sha, W. C., Nelson, C. A., Newberry, R. D., Kranz, D. M. &
50. Berg, L. J., Pullem, A. M., Fazekas de St. Groth, B., Mathis,
51, Kaye, J., Hsu, M. L., Sauron, M. E., Jameson, J. C. & Gascoigne,
52. von Boehmer, H., Teh, H. S. & Kisielow, P. (1989) Immunol.
53. Tonegawa, S. (1985) Sci Am. 253, 122- 131.
475 - 484.
Boehmer, H. (1988) Nature 333, 742- 746.
Nature 335,229 - 233.
Nature 335, 730-733.
Nature 338, 591 - 593.
527 - 529.
17-40.
(1985) Nature 315,232-233.
Nature 314, 103- 107.
(1986) EMBO J . 11,2837-2843.
12,463-467.
Russell, S. H. (1988) Nature 336, 73 - 76.
Russell, J. H. (1988) Nature 335, 271 -274.
D. & Benoist, C. (1989) Cell 58, 1035 - 1046.
R. S. (1989) Nature 341, 746-749.
Today. 10, 57 - 61.





























