Embed Size (px)
Citation preview

Although the lungs provide the means for gas exchange
between the external and internal environment, it is thehemoglobin in the red blood cells that transports oxygento the tissues. The red blood cells also function as carriers ofcarbon dioxide and participate in acid-base balance. The functionof the red blood cells, in terms of oxygen transport, is discussedin Chapter 19, and acid-base balance is covered inChapter 6. This chapter focuses on the red blood cell, anemia,and polycythemia.
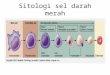
THE RED BLOOD CELLThe mature red blood cell, the erythrocyte, is a non-nucleated,biconcave disk (Fig. 13-1). This shape increases the surface areaavailable for diffusion of oxygen and allows the cell to changein volume and shape without rupturing its membrane. A cytoskeletonof proteins attached to the lipid bilayer provides thisunique shape and flexibility. The biconcave form presents theplasma with a surface 20 to 30 times greater than if the redblood cell were an absolute sphere. The erythrocytes, 500 to1000 times more numerous than other blood cells, are the mostcommon type of blood cell.The function of the red blood cell, facilitated by the hemoglobinmolecule, is to transport oxygen to the tissues. Hemoglobinalso binds some carbon dioxide and carries it from thetissues to the lungs. The hemoglobin molecule is composed oftwo pairs of structurally different polypeptide chains determinedby genes (Fig. 13-2). Alterations in these genes can resultin abnormal hemoglobins. Each of the four polypeptidechains is attached to a heme unit, which surrounds an atom ofiron that binds oxygen. Thus, one molecule of hemoglobin cancarry four molecules of oxygen.The two major types of normal hemoglobin are adulthemoglobin (HbA) and fetal hemoglobin (HbF). HbA consistsof a pair of chains and a pair of chains. HbF is thepredominant hemoglobin in the fetus from the third throughthe ninth month of gestation. It has a pair of chains substitutedfor the chains. Because of this chain substitution, HbFhas a high affinity for oxygen. This facilitates the transfer ofoxygen across the placenta. HbF is replaced within 6 monthsof birth with HbA.The rate at which hemoglobin is synthesized depends onthe availability of iron for heme synthesis. A lack of iron resultsin relatively small amounts of hemoglobin in the red bloodcells. The amount of iron in the body is approximately 35 to50 mg/kg of body weight for males and less for females. Bodyiron is found in several compartments. Most of the functionaliron (80%) is found in hemoglobin, with small amounts beingfound in the myoglobin of muscle, the cytochromes, and ironcontainingenzymes such as catalase. Approximately 15% to20% is stored in the bone marrow, liver, spleen, and other organs.Iron in the hemoglobin compartment is recycled. As redblood cells age and are destroyed in the spleen, iron from their

hemoglobin is released into the circulation and transported tothe bone marrow for use in production of new red cells or tothe liver and other tissues for storage.Dietary iron helps to maintain body stores. Iron, principallyderived from meat, is absorbed in the small intestine,especially the duodenum (Fig. 13-3). When body stores ofiron are diminished or red cell production is stimulated, absorptionis increased. In iron overload, excretion of iron isaccelerated. Normally, some iron is sequestered in the intestinalepithelial cells and is lost in the feces as these cellsslough. The iron that is absorbed enters the circulation, whereit immediately combines with a -globulin, called apotransferrin,to form transferrin, which is then transported in theplasma. From the plasma, iron can be deposited in tissuessuch as the liver. There it is stored as ferritin, a protein–ironcomplex, which can easily return to the circulation. Serumferritin levels, which can be measured in the laboratory, providean index of body iron stores. Clinically, decreased serumferritin levels indicate the need for prescription of iron supplements.Transferrin can also deliver iron to the developingred cell in bone marrow by binding to membrane receptors.The iron is taken up by the developing red cell, where it isused in heme synthesis.Red Cell ProductionErythropoiesis is the production of red blood cells. Afterbirth, red cells are produced in the red bone marrow. Untilage 5 years, almost all bones produce red cells to meet growthneeds. After this period, bone marrow activity gradually declines.After 20 years of age, red cell production takes placemainly in the membranous bones of the vertebrae, sternum,ribs, and pelvis. With this reduction in activity, the red bonemarrow is replaced with fatty yellow bone marrow.The red cells are derived from the erythroblasts or red cellprecursors, which are continuously being formed from thepluripotent stem cells in the bone marrow (Fig. 13-4). As theydevelop into mature red cells, the red cell precursors movethrough a series of divisions, each producing a smaller cell.Hemoglobin synthesis begins at an early stage and continuesuntil the cell becomes a mature erythrocyte. During the maturationtime, the red blood cell accumulates hemoglobin as thenucleus condenses and is finally lost. The period from stem cellto emergence of the reticulocyte in the circulation normallytakes approximately 1 week. Maturation of reticulocyte toerythrocyte takes approximately 24 to 48 hours. During thisprocess, the red cell loses its mitochondria and ribosomes,along with its ability to produce hemoglobin and engage inoxidative metabolism. Most maturing red cells enter the bloodas reticulocytes. Approximately 1% of the body’s total complementof red blood cells is generated from bone marrow eachday, so the reticulocyte count serves as an index of the erythropoieticactivity of the bone marrow.Erythropoiesis is governed for the most part by tissue oxygenneeds. Any condition that causes a decrease in the amountof oxygen that is transported in the blood produces an increasein red cell production. The oxygen content of the blood does

not act directly on the bone marrow to stimulate red blood cellproduction. Instead, the decreased oxygen content is sensed bythe kidneys, which then produce a hormone called erythropoietin.Normally, the kidneys produce approximately 90% oferythropoietin, with the remaining 10% being released by theliver. In the absence of erythropoietin, as in kidney failure,hypoxia has little or no effect on red blood cell production.Erythropoietin takes several days to effect a release of red bloodcells from the bone marrow, and only after 5 days or more doesred blood cell production reach a maximum.Erythropoietin acts in the bone marrow by binding toreceptors on committed stem cells. It functions on many levelsto promote hemoglobin synthesis, increase production ofmembrane proteins, and cause differentiation of erythroblasts.Human erythropoietin can be produced by recombinant deoxyribonucleicacid (DNA) technology. It is used for managementof anemia in cases of chronic renal failure, for treatmentof chemotherapy-induced anemia in persons with malignancies,and treatment of anemia in persons with human immunodeficiencyvirus (HIV) infection who are being treated withzidovudine.Because red blood cells are released into the blood as reticulocytes,the percentage of these cells is higher when there is amarked increase in red blood cell production. For example, insome severe anemias, the reticulocytes may account for asmuch as 30% of the total red cell count. In some situations, redcell production is so accelerated that numerous erythroblastsappear in the blood.
Red Cell DestructionMature red blood cells have a life span of approximately4 months or 120 days. As the red blood cell ages, a number ofchanges occur. Metabolic activity in the cell decreases, andenzyme activity decreases; adenosine triphosphate (ATP) decreases,and the cell membrane becomes more fragile. Once thered cell membrane becomes fragile, the cell ruptures duringpassage through narrowed places in the circulation. Many ofthe red cells self-destruct in the small trabecular spaces in thered pulp of the spleen. The rate of red cell destruction (1% perday) normally is equal to red cell production, but in conditionssuch as hemolytic anemia, the cell’s life span may be shorter.The destruction of red blood cells is facilitated by a group oflarge phagocytic cells found in the spleen, liver, bone marrow,and lymph nodes. These phagocytic cells ingest the hemoglobinfrom the ruptured cells and break it down in a series ofenzymatic reactions. During these reactions, the amino acidsfrom the globulin chains and iron from the heme units are salvagedand reused (Fig. 13-5). The bulk of the heme unit is convertedto bilirubin, the pigment of bile, which is insoluble inplasma and attaches to the plasma proteins for transport.Bilirubin is removed from the blood by the liver and conjugatedwith glucuronide to render it water soluble so that it canbe excreted in the bile. The plasma-insoluble form of bilirubinis referred to as unconjugated bilirubin; the water-soluble form isreferred to as conjugated bilirubin. Serum levels of conjugated

and unconjugated bilirubin can be measured in the laboratoryand are reported as direct and indirect, respectively. If red celldestruction and consequent bilirubin production are excessive,unconjugated bilirubin accumulates in the blood. This resultsin a yellow discoloration of the skin, called jaundice.When red blood cell destruction takes place in the circulation,as in hemolytic anemia, the hemoglobin remains in theplasma. The plasma contains a hemoglobin-binding proteincalled haptoglobin. Other plasma proteins, such as albumin, canalso bind hemoglobin. With extensive intravascular destructionof red blood cells, hemoglobin levels may exceed thehemoglobin-binding capacity of haptoglobin. When this happens,free hemoglobin appears in the blood (i.e., hemoglobinemia)and is excreted in the urine (i.e., hemoglobinuria).Because excessive red blood cell destruction can occur inhemolytic transfusion reactions, urine samples are tested forfree hemoglobin after a transfusion reaction.Red Cell Metabolism andHemoglobin OxidationThe red blood cell, which lacks mitochondria, relies on glucoseand the glycolytic pathway for its metabolic needs. Theenzyme-mediated anaerobic metabolism of glucose generatesthe ATP needed for normal membrane function and ion transport.The depletion of glucose or the functional deficiency ofone of the glycolytic enzymes leads to the premature death ofthe red blood cell. An offshoot of the glycolytic pathway is theproduction of 2,3-diphosphoglycerate (2,3-DPG), which bindsto the hemoglobin molecule and reduces the affinity of hemoglobinfor oxygen. This facilitates the release of oxygen at thetissue level. An increase in the concentration of 2,3-DPG occursin conditions of chronic hypoxia such as chronic lung disease,anemia, and high altitudes.The oxidation of hemoglobin—the combining of hemoglobinwith oxygen—can be interrupted by certain chemicals(e.g., nitrates and sulfates) and drugs that oxidize hemoglobinto an inactive form. For example, the nitrite ion reacts withhemoglobin to produce methemoglobin, which has a low affinityfor oxygen. Large doses of nitrites can result in high levels ofmethemoglobin, causing pseudocyanosis and tissue hypoxia.For example, sodium nitrate, which is used in curing meats,can produce methemoglobin when taken in large amounts.In nursing infants, the intestinal flora is capable of convertingsignificant amounts of inorganic nitrate (e.g., from well water)to nitrite. A hereditary deficiency of glucose 6-phosphate dehydrogenase(G6PD; to be discussed) predisposes to oxidativedenaturation of hemoglobin, with resultant red cell injury andlysis. Hemolysis occurs as the result of oxidative stress generatedby either an infection or exposure to certain drugs.Laboratory TestsRed blood cells can be studied by means of a sample of blood(Table 13-1). In the laboratory, automated blood cell countersrapidly provide accurate measurements of red cell content andcell indices. The red blood cell count (RBC) measures the totalnumber of red blood cells in 1 mm3 of blood. The percentageof reticulocytes (normally approximately 1%) provides an index

of the rate of red cell production. The hemoglobin (grams per100 mL of blood) measures the hemoglobin content of theblood. The major components of blood are the red cell massand plasma volume. The hematocrit measures the volume of redcell mass in 100 mL of plasma volume. To determine the hematocrit,a sample of blood is placed in a glass tube, which isthen centrifuged to separate the cells and the plasma. Thehematocrit may be deceptive because it varies with the quantityof extracellular fluid, rising with dehydration and fallingwith overexpansion of extracellular fluid volume.Red cell indices are used to differentiate types of anemias bysize or color of red cells. The mean corpuscular volume (MCV)reflects the volume or size of the red cells. The MCV falls inmicrocytic (small cell) anemia and rises in macrocytic (largecell) anemia. Some anemias are normocytic (i.e., cells are ofnormal size or MCV). The mean corpuscular hemoglobin concentration(MCHC) is the concentration of hemoglobin in eachcell. Hemoglobin accounts for the color of red blood cells.Anemias are described as normochromic (normal color or MCHC)or hypochromic (decreased color or MCHC). Mean cell hemoglobin(MCH) refers to the mass of the red cell and is less useful inclassifying anemias.A stained blood smear provides information about the size,color, and shape of red cells and the presence of immature orabnormal cells. If blood smear results are abnormal, examinationof the bone marrow may be important. Marrow commonlyis aspirated with a special needle from the posterior iliaccrest or the sternum. The aspirate is stained and observed fornumber and maturity of cells and abnormal types.
Meskipun paru-paru menyediakan sarana untuk pertukaran gas antara lingkungan eksternal dan internal, namun hemoglobin dalam sel darah merah yang mengangkut oksigen ke jaringan. Sel darah merah juga berfungsi sebagai pembawa karbon dioksida dan berpartisipasi dalam keseimbangan asam-basa. Fungsi sel darah merah, dalam hal transportasi oksigen, dibahas di Bab 19, dan asam-basa keseimbangan ditutupi Bab 6. Bab ini berfokus pada sel darah merah, anemia, dan polycythemia.
SEL DARAH MERAH Sel darah merah matang, eritrosit, adalah non-bernukleus, cekung ganda disk (Gambar 13-1). Bentuk ini meningkatkan luas permukaan tersedia untuk difusi oksigen dan memungkinkan sel untuk mengubah volume dan bentuk tanpa pecah membran. sitoskeleton A protein melekat pada lapisan ganda lipid memberikan ini unik bentuk dan fleksibilitas. Bentuk cekung ganda menyajikan plasma dengan permukaan 20 sampai 30 kali lebih besar dibandingkan jika merah sel darah sebuah bola mutlak. Eritrosit, 500 untuk 1000 kali lebih banyak dari sel darah lainnya, adalah yang paling umum jenis sel darah. Fungsi dari sel darah merah, difasilitasi oleh hemoglobin molekul, adalah untuk mengangkut oksigen ke jaringan. Hemoglobin juga mengikat beberapa karbon dioksida dan membawanya dari jaringan ke paru-paru. Molekul hemoglobin terdiri dari dua pasang rantai polipeptida ditentukan secara struktural berbeda oleh gen.

Perubahan pada gen ini dapat mengakibatkan dalam hemoglobin abnormal. Masing-masing dari empat polipeptida rantai terpasang ke unit heme, yang mengelilingi sebuah atom besi yang mengikat oksigen. Jadi, satu molekul hemoglobin dapat membawa empat molekul oksigen. Dua jenis utama dari hemoglobin normal orang dewasa hemoglobin (HbA) dan hemoglobin janin (HbF). HbA terdiri . rantai dan sepasang rantai dari sepasang HbF adalah dominan hemoglobin pada janin dari ketiga melalui bulan kesembilan kehamilan. Ia memiliki sepasang rantai diganti . untuk rantai Karena itu substitusi rantai, HbF memiliki afinitas tinggi untuk oksigen. Hal ini memfasilitasi transfer oksigen di plasenta. HbF diganti dalam 6 bulan lahir dengan HbA. Tingkat di mana hemoglobin disintesis tergantung pada ketersediaan besi untuk sintesis heme. Kurangnya hasil besi dalam jumlah relatif kecil hemoglobin dalam darah merah sel. Jumlah zat besi dalam tubuh adalah sekitar 35 untuk 50 mg / kg berat badan untuk pria dan kurang untuk wanita. Tubuh besi ditemukan dalam beberapa kompartemen. Sebagian besar fungsional besi (80%) ditemukan dalam hemoglobin, dengan jumlah kecil yang ditemukan dalam mioglobin dari otot, sitokrom, dan ironcontaining enzim seperti katalase. Sekitar 15% sampai 20% disimpan di sumsum tulang, hati, limpa, dan organ lainnya.
Besi dalam kompartemen hemoglobin didaur ulang. Sebagai sel redblood usia dan dihancurkan di limpa, besi dari mereka hemoglobin dilepaskan ke sirkulasi dan diangkut ke sumsum tulang untuk digunakan dalam produksi sel darah merah baru atau hati dan jaringan lain untuk penyimpanan. besi Diet membantu untuk mengelola toko tubuh. Besi, terutama berasal dari daging, diserap di usus kecil, terutama duodenum (Gbr. 13-3). Ketika tubuh gudang besi produksi sel berkurang atau merah dirangsang, penyerapan meningkat. Dalam kelebihan zat besi, ekskresi besi dipercepat. Biasanya, besi beberapa adalah diasingkan di usus sel epitel dan hilang dalam tinja sebagai sel-sel ini rawa. Besi yang diserap masuk sirkulasi, di mana -globulin, disebut apotransferrin,segera menggabungkan dengan untuk membentuk transferin, yang kemudian diangkut dalam plasma. Dari plasma, besi dapat disimpan dalam jaringan seperti hati. Ada disimpan sebagai ferritin, protein besi kompleks, yang dengan mudah dapat kembali ke sirkulasi. Serum feritin tingkat, yang dapat diukur di laboratorium, memberikan indeks toko besi tubuh. Klinis, penurunan serum kadar feritin menunjukkan perlunya resep suplemen zat besi. Transferrin juga dapat memberikan besi dengan peningkatan merah sel dalam sumsum tulang dengan cara mengikat reseptor membran. Setrika diambil oleh sel merah berkembang, di mana ia
digunakan dalam sintesis heme. Red Cell Produksi Eritropoiesis adalah produksi sel darah merah. Setelah lahir, sel darah merah diproduksi di sumsum tulang merah. Sampai usia 5 tahun, hampir semua tulang menghasilkan sel darah merah untuk memenuhi pertumbuhan kebutuhan. Setelah periode ini, aktivitas sumsum tulang secara bertahap menurun. Setelah 20 tahun, produksi sel darah merah terjadi terutama dalam tulang membran dari sternum, vertebra, rusuk, dan panggul. Dengan penurunan aktivitas, tulang merah sumsum diganti dengan sumsum tulang lemak kuning. Sel-sel merah berasal dari erythroblasts atau sel darah merah prekursor, yang terus menerus terbentuk dari pluripotent sel induk sumsum tulang (Gbr. 13-4). Saat mereka berkembang menjadi sel darah merah matang, prekursor sel darah merah bergerak melalui serangkaian divisi, masing-masing memproduksi sebuah sel kecil. sintesis Hemoglobin dimulai pada tahap awal dan terus sampai sel menjadi eritrosit matang. Selama pendewasaan waktu, sel darah merah hemoglobin terakumulasi sebagai thenucleus mengembun dan akhirnya

hilang. Periode dari stem cell untuk munculnya retikulosit dalam sirkulasi normal berlangsung sekitar 1 minggu. Pematangan retikulosit untuk eritrosit berlangsung sekitar 24 hingga 48 jam. Selama ini proses, sel merah kehilangan mitokondria dan ribosom, bersama dengan kemampuannya untuk memproduksi hemoglobin dan terlibat dalam metabolisme oksidatif. Sebagian besar sel darah merah jatuh tempo memasuki darah sebagai reticulocytes. Sekitar 1% dari total tubuh komplemen sel darah merah dihasilkan dari sumsum tulang masing-masing hari, sehingga jumlah retikulosit berfungsi sebagai indeks dari erythropoietic aktivitas sumsum tulang. Eritropoiesis diatur untuk sebagian besar oleh oksigen jaringan kebutuhan. Setiap kondisi yang menyebabkan penurunan jumlah oksigen yang diangkut dalam darah menghasilkan peningkatan dalam produksi sel darah merah. Kandungan oksigen dalam darah tidak tidak bertindak langsung pada sumsum tulang untuk merangsang sel darah merah produksi. Sebaliknya, kandungan oksigen menurun dirasakan oleh ginjal, yang kemudian menghasilkan hormon yang disebut eritropoietin. Biasanya, ginjal memproduksi sekitar 90% dari
erythropoietin, dengan 10% sisanya yang dirilis oleh hati. Dengan tidak adanya eritropoietin, seperti gagal ginjal, hipoksia memiliki pengaruh yang kecil atau tidak ada pada produksi sel darah merah. Erythropoietin membutuhkan waktu beberapa hari untuk efek pelepasan darah merah sel-sel dari sumsum tulang, dan hanya setelah 5 hari atau lebih tidak produksi sel darah merah mencapai maksimal. Eritropoietin bertindak di sumsum tulang dengan cara mengikat reseptor pada sel batang berkomitmen. Ini fungsi pada berbagai tingkatan untuk mempromosikan sintesis hemoglobin, meningkatkan produksi membran protein, dan diferensiasi penyebab erythroblasts. Manusia erythropoietin dapat diproduksi dengan rekombinan deoxyribonucleic asam (DNA) teknologi. Hal ini digunakan untuk manajemen anemia pada kasus gagal ginjal kronis, untuk pengobatan kemoterapi-induced anemia pada orang dengan keganasan, dan pengobatan anemia pada orang dengan human immunodeficiency Virus (HIV) yang sedang dirawat dengan AZT.
Karena sel-sel darah merah yang dilepaskan ke dalam darah sebagai reticulocytes, persentase sel-sel ini lebih tinggi bila ada ditandai peningkatan produksi sel darah merah. Misalnya, dalam beberapa anemi berat, mungkin reticulocytes account sebagai sebanyak 30% dari total jumlah sel merah. Dalam beberapa situasi, merah produksi sel begitu dipercepat bahwa banyak erythroblasts muncul dalam darah. Red Cell Destruction Menghasilkan sel darah merah memiliki jangka hidup sekitar 4 bulan atau 120 hari. Karena usia sel darah merah, sejumlah terjadi perubahan. Aktivitas metabolik dalam sel berkurang, dan aktivitas enzim menurun; adenosin trifosfat (ATP) menurun, dan membran sel menjadi lebih rapuh. Setelah membran sel darah merah menjadi rapuh, pecah sel selama melintasi tempat menyempit di sirkulasi. Banyak sel-sel merah merusak diri sendiri dalam ruang trabekular kecil di pulp merah limpa. Laju kerusakan sel darah merah (1% per hari) biasanya adalah sama dengan produksi sel darah merah, namun dalam kondisi seperti anemia hemolitik, rentang hidup sel mungkin lebih pendek.

Penghancuran sel darah merah yang difasilitasi oleh sekelompok sel fagosit besar ditemukan di limpa, hati, sumsum tulang, dan kelenjar getah bening. Sel-sel fagosit menelan hemoglobin dari sel pecah dan memecahnya dalam serangkaian reaksi enzimatik. Selama reaksi, asam amino dari rantai globulin dan besi dari unit heme yang diselamatkan dan digunakan kembali (Gbr. 13-5). Sebagian besar unit heme diubah untuk bilirubin, pigmen empedu, yang tidak larut dalam plasma dan menempel pada protein plasma untuk transportasi. Bilirubin dikeluarkan dari darah oleh hati dan terkonjugasi dengan glukuronat untuk membuat itu larut dalam air sehingga dapat bisa dikeluarkan dalam empedu. Bentuk plasma-larut bilirubin disebut bilirubin sebagai unconjugated; bentuk yang larut dalam air adalah disebut sebagai konjugasi bilirubin. Serum tingkat konjugasi dan bilirubin unconjugated dapat diukur di laboratorium dan dilaporkan sebagai langsung dan tidak langsung, masing-masing. Jika sel darah merah perusakan dan produksi bilirubin akibat yang berlebihan, unconjugated bilirubin terakumulasi dalam darah. Hasil ini dalam warna kuning pada kulit, yang disebut penyakit kuning. Ketika penghancuran sel darah merah terjadi di sirkulasi, seperti pada anemia hemolitik, hemoglobin tetap berada dalam plasma. plasma mengandung protein hemoglobin-mengikat disebut haptoglobin. Lain plasma protein, seperti albumin, dapat juga mengikat hemoglobin. Dengan kehancuran intravaskular yang luas sel darah merah, kadar hemoglobin dapat melebihi mengikat hemoglobin kapasitas haptoglobin. Ketika ini terjadi, hemoglobin bebas muncul dalam darah (yaitu, hemoglobinemia) dan diekskresikan di dalam urin (yaitu, hemoglobinuria). Karena berlebihan penghancuran sel darah merah dapat terjadi pada reaksi transfusi hemolitik, sampel urin diuji untuk free hemoglobin setelah reaksi transfusi. Merah dan sel Metabolisme Hemoglobin Oksidasi Sel darah merah, yang tidak memiliki mitokondria, bergantung pada glukosa dan jalur glikolitik untuk kebutuhan metabolik. The metabolisme anaerobik enzim-dimediasi glukosa menghasilkan ATP dibutuhkan untuk fungsi membran normal dan transpor ion. Penipisan glukosa atau defisiensi fungsional salah satu enzim glikolitik menyebabkan kematian dini sel darah merah. Sebuah cabang dari jalur glikolitik adalah produksi 2,3-diphosphoglycerate (2,3-DPG), yang mengikat ke molekul hemoglobin dan mengurangi afinitas hemoglobin untuk oksigen. Hal ini memudahkan pelepasan oksigen pada tingkat jaringan. Peningkatan konsentrasi 2,3-DPG terjadi dalam kondisi hipoksia kronis seperti penyakit paru-paru kronis, anemia, dan ketinggian tinggi.

Oksidasi hemoglobin-kombinasi dari hemoglobin dengan oksigen dapat terganggu oleh bahan kimia tertentu (Misalnya, nitrat dan sulfat) dan obat-obatan yang mengoksidasi hemoglobin untuk bentuk tidak aktif. Sebagai contoh, ion nitrit bereaksi dengan hemoglobin untuk menghasilkan methemoglobin, yang memiliki afinitas rendah untuk oksigen. Dosis besar nitrit dapat mengakibatkan tingkat tinggi methemoglobin pseudocyanosis, menyebabkan dan hipoksia jaringan. Sebagai contoh, natrium nitrat, yang digunakan dalam menyembuhkan daging, dapat menghasilkan methemoglobin ketika diambil dalam jumlah besar. Pada bayi menyusui, flora usus yang mampu mengkonversi signifikan jumlah nitrat anorganik (misalnya, dari air sumur) menjadi nitrit. Kekurangan turun temurun dari dehidrogenase glukosa 6-fosfat (G6PD, akan dibahas) predisposes untuk oksidatif denaturasi hemoglobin, dengan cedera sel resultan merah dan lisis. Hemolisis terjadi sebagai akibat dari stres oksidatif yang dihasilkan baik oleh infeksi atau paparan obat-obatan tertentu. Laboratorium Pengujian Sel darah merah dapat dipelajari melalui sampel darah (Tabel 13-1). Di laboratorium, counter otomatis sel darah cepat memberikan pengukuran akurat kadar sel darah merah dan sel indeks. Jumlah sel darah merah (RBC) mengukur jumlah jumlah sel darah merah dalam 1 mm3 darah. Persentase dari reticulocytes (biasanya sekitar 1%) memberikan indeks dari tingkat produksi sel darah merah. Hemoglobin (gram per 100 mL darah) mengukur kadar hemoglobin dari darah. Komponen utama dari darah massa sel darah merah dan plasma volume. hematokrit mengukur volume massa redcell dalam 100 mL volume plasma. Untuk menentukan hematokrit, sampel darah ditempatkan dalam tabung kaca, yang kemudian disentrifugasi untuk memisahkan sel dan plasma. The hematokrit mungkin menipu karena bervariasi dengan kuantitas cairan ekstraselular, meningkat dengan dehidrasi dan jatuh dengan overexpansion volume cairan ekstraselular. indeks sel Merah digunakan untuk membedakan jenis anemi oleh ukuran atau warna sel darah merah. Corpuscular volume rata-rata (MCV) mencerminkan volume atau ukuran sel darah merah. The MCV jatuh mikrositik (sel kecil) anemia dan meningkat di makrositik (besar sel) anemia. Beberapa anemi adalah normositik (yaitu, sel-sel dari normal ukuran atau MCV). Konsentrasi hemoglobin rata-rata corpuscular (MCHC) adalah konsentrasi hemoglobin di masing-masing sel. Hemoglobin account untuk warna sel darah merah. Anemi digambarkan sebagai normokromik (warna normal atau MCHC) atau hipokrom (menurun warna atau MCHC). Rata-rata sel hemoglobin (KIA) mengacu pada massa sel darah merah dan kurang berguna dalam mengelompokkan anemi. Pap bernoda darah memberikan informasi tentang ukuran,

warna, dan bentuk sel darah merah dan keberadaan belum dewasa atau sel abnormal. Jika hasil tes darah tidak normal, pemeriksaan dari sumsum tulang mungkin penting. Marrow umum adalah disedot dengan jarum khusus dari iliaka posterior puncak atau sternum. aspirate ini diwarnai dan diamati untuk jumlah dan kematangan sel-sel dan jenis abnormal.