Embed Size (px)
Citation preview

��
SECONDARY METABOLITES COMPOSITION AND GEOGRAPHICAL DISTRIBUTION OF MARINE
SPONGES OF THE GENUS DYSIDEA
By
Mayuri CHANDRA
A thesis submitted in partial fulfillment of the requirements for the degree of
Master of Science
School of Biological & Chemical Sciences,
Faculty of Science, Technology & Environment,
The University of the South Pacific
Suva, Fiji Islands
June, 2009


��
�
�
“Man masters nature not by force but by understanding. This is why science has
succeeded where magic failed: because it has looked for no spell to cast on nature.”
- JACOB BRONOWSKI

���
���������� �
This project would not have been completed without the help and support of a
number of people. My heartfelt acknowledgement goes to the following people and
organisations:
This work is part of the C2 component of the Coral Reef Initiative in the South
Pacific (CRISP) project and funded by the Agence Française de Développement. The
research grant for the component completed in Fiji was obtained from the
Government of Taiwan. This project was also partly completed in New Caledonia
and France, and it was funded by the Government of France with the initiative of the
Embassy of France in Fiji and the Centre Régional des Œuvres Universitaires et
Scolaires (CROUS).
I am grateful to my supervisors, Professor Subramaniam Sotheeswaran and Associate
Professor Sadaquat Ali, for giving me an opportunity to pursue my interest in the
field of Natural Products research. This project was a good learning ground for me,
which provided great experiences along the way.
Furthermore, most of this project was completed in New Caledonia and France, and
my sincere gratitude goes to my external supervisors, Dr Sylvain Petek, Dr Isabelle
Bonnard and Dr Bernard Banaigs for their guidance.
Dr Sylvain Petek, my supervisor in New Caledonia, has been a great support and
mentor. The research team at the Institut des Recherche et Développement (IRD –
Noumea, New Caledonia) was really friendly and helped me greatly to adjust to the
new environment and language.
Dr Isabelle Bonnard at the Centre de Phytopharmacie, University of Perpignan via
Domitia (Perpignan, France), was always there for me whether I had any project-
related or personal problems. She is a great mentor and I was able to have a good
learning experience, which broadened my knowledge of research. Thanks to the staff
and students of Centre de Phytopharmacie, I was also able to explore France to some
extent, and the experience was amazing.
The sponge samples used in this study were obtained by diving; hence I would like to
thank the dive team of the IRD Centre of Nouméa, New Caledonia, for harvesting
the sponges. Furthermore, we highly appreciate the governments of New Caledonia,

����
Vanuatu, Solomon and Fiji Islands for permitting us to collect our samples in their
countries, and the Fisheries Departments of these countries for their help and
assistance during the collections.
I would also like to thank Dr Cécile Debitus, Pierre Pério, and Camille Durand at the
Université Paul Sabatier, Toulouse, France, as well as Professor Valeria D’Auria at
the University of Naples, Italy, for providing us with technical support in our sample
analysis.
The anti-microbial testing was carried out partly at the IRD – Noumea, and partly at
the Institute of Applied Sciences (IAS), University of the South Pacific. I would like
to thank the microbiology team at the IAS (especially Pritesh, Kavita, Girish Lakhan,
and Mr. Klaus Feussner) for providing the necessary resources for completing this
part of the project.
I can not forget my family and friends for their never-ending support in the duration
of my project. For all the hours spent in the laboratory, there was always someone to
give company or talk to when things became tensed, confusing, or just tiresome. My
thanks goes to my friends Nirul, Ardarsh, Roselyn, Joslin, Sujlesh, Riteshma, and the
technical staff of the Faculty of Science and Technology, USP, Suva. Kirti and Shital
helped me quite a lot during my stay in France. We had a great time together.
Finally, the support, blessing and sacrifice of my parents, Mr. Vinod Chandra and
Mrs. Kala Chandra, have given me the strength to stay focused on my path to
complete this project.
Thank you, vinaka vakalevu, and merci beaucoup to everyone who has contributed to
this project in one way or another.

�����
��� ��� �
Dysidea species – Lamellodysidea herbacea, Dysidea arenaria, and Dysidea avara –
from New Caledonia were chemically screened to identify the chemical constituents
of these sponges. Each species exhibited a specific profile, thus grouping the three
species chemically together in distinct taxa. The screening also identified the
presence of three different chemical types of L. herbacea present within the same
region, New Caledonia.
The pattern of occurrence of polybrominated diphenyl ethers (PBDE) in L. herbacea
was also mapped for distant geographical sites, which included New Caledonia,
Vanuatu, Fiji Islands, Solomon Islands, Moorea Island in French Polynesia, and
Mayotte. Comparison and analysis of the chemical fingerprints shows that all the
samples belong to the bromo-chemotype of compounds, and similarities and
differences exist within the same species in different geographical locations.
L. herbacea was further explored for compound isolations and characterization to
verify the chemical fingerprints of these sponges. Spectroscopic and chemical
analyses confirmed the presence of PBDEs in the L. herbacea analyzed. Eight
different PBDEs have been identified, where four of these (compounds 2, 6, 7, and 8)
have been successfully isolated and characterized.
Crude extracts of the Dysidea sponges were also tested for activity against the fungi
Cryptococcus neoformans and Candida albicans. Most extracts were active against
Cr. neoformans hence confirming the hypothesis that the action was due to the
inhibition of the proper functioning of the protein farnesyl transferase (PFT), which
is an essential enzyme in the metabolism of this fungus. Ca. albicans is able to
survive even without PFT. Since Plasmodium falciparum (the malarial parasite) is
also dependent on PFT for survival, this anti-fungal test is being developed to be
used as a pre-test for anti-plasmodial studies.

����
��������
L’étude et la comparaison des profils chimiques chez Lamellodysidea herbacea,
Dysidea arenaria, et Dysidea avara ont été menés afin d’identifier les métabolites
secondaires présents dans ces trois espèces d’éponges qui pourraient être utilisés
comme marqueurs taxonomiques. En comparant les profils chimiques, il est apparu
que chaque espèce Lamellodysidea herbacea, Dysidea arenaria, Dysidea avara
pouvaient être groupement, car chacune présente un profil chimique spécifique.
Les profils chimiques des différents échantillons de L. herbacea a ont été étudiés
pour différents sites géographiques. Les pays régions géographiques concernées sont:
la Nouvelle Calédonie, les Vanuatu, les îles Fidji, les îles Salomon, la Polynésie
Française (Moorea) et Mayotte. Les analyses chimiques et spectroscopiques
montrent que tous les échantillons de L. herbacea contiennent des diphénylethers
polybromés (PBDE, bromochémotype).
L. herbacea a été étudiée plus avant pour les composés isolement et la caractérisation
de manière plus approfondie au niveau de l’isolement et de la caractérisation de
composés pour vérifier les ‘empreintes digitales’ chimiques de ces éponges. Les
analysés spectroscopiques et chimiques ont a confirmé la présence de PBDE dans les
extraits de L. herbacea analysées. Huit différents PBDE ont été identifiés; trois types
chimiques de L. herbacea ont été caractérisés; et quatre composés (composés de 2, 6,
7 et 8) ont été isolés et caractérisés.
L’activité antifongique des extrait bruts de Dysidea a ont également été étudiée sur
deux souches de levure testées: pour l'activité contre les champignons Cryptococcus
neoformans et Candida albicans. La plupart des extraits ont été actifs contre sur Cr.
neoformans sans forcément l’être sur Ca. albicans, partant, confirmant l'hypothèse
selon laquelle l'action est le résultat de l'inhibition. Cette différence d’activité
suppose une inhibition du bon fonctionnement de la protéine farnesyl transférase
(PFTase), qui est un enzyme essentielle dans le métabolisme de Cr. neoformans alors
que ce champignon, Ca. albicans est capable de survivre même sans PFTase. Depuis
le Plasmodium falciparum (le parasite du paludisme) est également tributaire des
PFTase pour la survie, cette anti-fongiques test est développé pour être utilisé comme
un pré-test pour les études anti-plasmodiales.

���
���������� � ��
���������� ���������������������������������������������������������������������������������������������������������������������������
��� ��� ���������������������������������������������������������������������������������������������������������������������������������������������
�������������������������������������������������������������������������������������������������������������������������������������������������
��� ���������������������������������������������������������������������������������������������������������������������������������������������
��� ������� ��������������������������������������������������������������������������������������������������������������������������������������
��� ���� �����������������������������������������������������������������������������������������������������������������������������������������
��� ������������������������������������������������������������������������������������������������������������������������������������������
��� ����������� ��������������������������������������������������������������������������������������������������������������������������
��� ����� ��� �������������������������������������������������������������������������������������������������������������������������������
�� �� ���� ������������������������������������������������������������������������������������������������������������������������������
���� ���� ���������������������������������������������������������������������������������������������������������������������������������
���� ����������������������������� ����������������������������������������������������������������������������������
������ ������ ���� ���������������������������������������������������������������������������������������������������
������ ����������� ������ ��������������������������������������������������������������������������������������������������
������ ���� �����!����� ������������������������������������������������������������������������������������������������������������"�
���� �����������������������������������������������������������������������������������������������������������������������
���� ����������������������������������������������������������������������������������������������������
���� �� ������ �������� ������� ������������������������������������������������������������������������������!�
��!� ���� ��������������������������������������������������������������������������������������������������������������������������"�
��"��� �������������# ��������������������������������������������������������������������������������������������������$�
��"��� � ���� � �%�� ����������������������������������������������������������������������������������������������������������������&�
��"��� '�(�� �������)���#��%� ��!���!�*���#�������������������������������������������������������������&�
��#� ��� ��������� ��������������������������������������������������������������������������������������������������������������������
�� ������� ����������������������������������������������������������������������������������������������������������������
���� �����������������������������������������������������������������������������������������������������������������
���� �������������$ �������������������������������������������������������������������������������������������������������������
������ ������+��������������������������������������������������������������������������������������������������������������������������,�
������ ������+��������������������������������������������������������������������������������������������������������������������������-�
���� �����������������������������������������������������������������������������������������������������������������������������������%�
������ �����. �����# �� ����/�.�0������������������������������������������������������������������������������������&�
������ �����1��# ����.�+������# �� ����/�1.�0����������������������������������������������������������&�
������ .�+������# �� ����2�� �������������/.�3��0������������������������������������������������������

����
����,� '���� �� �������4��� ����/'�40���������������������������������������������������������������������
���� �������������� � ���������������������������������������������������������������������������������������
��,��� !����� ���������4��,�������������������������������������������������������������������������������������������������������
��,��� . #��������� ���% �� �������������������������������������������������������������������������������������������������,�
���� ���&� ���������������������������������������������������������������������������������������������������������������������������
��5��� ���� �/� %� ���!�6����)� �2��!)0�1�� ��������������������������������������������������������5�
��5��� )���3������ #�!������������������������������������������������������������������������������������������������������������5�
��5��� 7��� ��8���� ���������������������������������������������������������������������������������������������������������������"�
�� ���� ������������������������������������������������������������������������������������������������������������������������ �
���� ��������������������������������������������������������������������������������������������������������������������������������#�
������ . #��������� ���% �� �������������������������������������������������������������������������������������������������9�
�������� '�(��)*+,��',-.)/-0,)1'*�������������������������������������������������������������������������������������������������#�
�������� ��0'��+,2-,.)(3+���45�6��',-.)/-0,)1'*�7����8���������������������������������������������������������������9�
�������� ��45�6��',-.)/-0,)1'*����)::��1+3/,-:3-1*�7��&��8����������������������������������������������������������
�������� �53;+),��)0(+/�3�+:-()(3+��1+3/,-:3-1*�7��8����������������������������������������������������������������
���������� �-.1-5(6������������������������������������������������������������������������������������������������������������������������
���������� �-.1-5(6����������������������������������������������������������������������������������������������������������������������!�
���������� �-.1-5(6����������������������������������������������������������������������������������������������������������������������9�
������ !����� � �� � �������������������������������������������������������������������������������������������������������������5��
������ !����� � * ������������������������������������������������������������������������������������������������������������������5��
����,� �:'�.;�8:'�������������������������������������������������������������������������������������������������������������������5��
���� ����������������� ���������������������������������������������������������������������������������������!�
������ <�:.: =�:7���<� <';��!=�8!<)����������������������������������������������������������������������������������5"�
�������� �).+;;-6*:�6+)�'+,<)3+)�����������������������������������������������������������������������������������������������������#�
�������� �*:�6+)�),+(),�)�)(6��*:�6+)�)�),)�������������������������������������������������������������������������������������#�
�������� �*:�6+)�:1+3�+:�7�%��=���!�=�)(6�����8������������������������������������������������������������������������"�
������ �8�8.)48�8<��)'!�!877<4<'�<��8'���<�8�).��:�1:�8�8:'�)'!� <'<�8��.8'>) <�
�<�?<<'�.)�<..:!=�8!<)��<4�)�<)��)�<!�:'� <: 4)1�8�).�!8��48�;�8:'�������������������5$�
�������� �-.1),�:-(�-2��).+;;-6*:�6+)�'+,<)3+)�3-;;+3/+6�>�/'�(��+>��);+6-(�)�������������������������9�
���������� �-.1),�:-(�-2�/'+��'+.�3);��,-2�;+:������������������������������������������������������������������������������9�
���������� �-.1),�:-(�-2�/'+��'+.�3);��,-2�;+:�>�/'��+(+��)11�(0���������������������������������������������!��
�������� �-.1),�:-(�-2��).+;;-6*:�6+)�'+,<)3+)��(�-/'+,��+-0,)1'�3);��-3)/�-(:������������������������!��
���������� �-.1),�:-(�-2�/'+��'+.�3);��,-2�;+:�����������������������������������������������������������������������������!��
���������� �-.1),�:-(�-2�/'+��'+.�3);��,-2�;+:�>�/'��+(+��)11�(0���������������������������������������������!#�
������ �:�1)48�:'�:7�!=�8!<)�)4<')48)�)'!�!��)@)4)�74:��'<?��).<!:'8)�)'!�
78A8B�)'!���!=�8!<)��1<�8<��74:����<��:.:�:'�8�.)'!��������������������������������������������������������"-�
����,� �:'�.;�8:'�������������������������������������������������������������������������������������������������������������������9��
���� ���������?�� � ��������������������� ���������������������������������#��
������ !=�8!<)��1<�8<��4��,��������������������������������������������������������������������������������������������������9��

�����
������ .)�<..:!=�8!<)��<4�)�<)�����������������������������������������������������������������������������������������99�
�������� �).1;+�������7�+>��);+6-(�)8�����������������������������������������������������������������������������������������##�
�������� �).1;+����#��7�-;-.-(��:�8����������������������������������������������������������������������������������������������#"�
�������� �).1;+�������7��@���:�8��������������������������������������������������������������������������������������������������������"%�
�������� �).1;+����&%�&�'��7���'+,<)3+)�2,-.��)*-//+8���������������������������������������������������������������"��
������ �:'�.;�8:'�������������������������������������������������������������������������������������������������������������������-��
���� ���&� ������������������������������������������������������������������������������������������������������������������������"��
�� ���������������������������������������������������������������������������������������������������������������������������!"�
#� ������������������������������������������������������������������������������������������������������������������������������������! �
"� ������������������������������������������������������������������������������������������������������������������������������������$��
!��� ����������A������������������ ���������������������������������������������������������������������������������%��
!��� ����������A�� ������������������������������������ �����������������������������������������������%��
!��� ����������A��������������������� �����=��=����������������������������������������������������������%��
"����� )������6�� C���'�4����������.����% �� ����#��������������������������������������������&,�
"����� )������6��%C��3!� ����3!�'�4�1���������#�����������������������������������������������������&"�
"����� )������6���C���'�4�1��������#������������������������������������������������������������������������&$�
!��� ����������A���������������������� ��������������?��������������������������� �
����� �����������������������������������������������������������������������������������������������������������������������������������������������
"�,��� )������6�, C�'�4���������� ����#���������������������������������������������������������������������
"�,��� =������%� �������������� �����#��������������������������������������������������������������������������
!��� ����������A�����������������������������������������������������������������������������������������������������������
!�!� ���������!A�����������A���������� ���������������������������������������������������������������������������������������
!�#� ���������#A������������������������=������������������� ����������������������������������������������������
!�"� ���������"A������������ ��������������������������������������������������������������������������#�
!�9� ���������9A�������������������!������������������������������������������B���������������
!��%� ����������%A���������������������#��������������������������������������������������������������������#�
"��&��� C�)������6��& C�4��,9����������������������������������������������������������������������������������������������������9�
"��&��� C�)������6��&%C�4��,�����������������������������������������������������������������������������������������������������&�
!���� �����������A��������������7���&%�&���8���� � ��������������������������������������������������
!���� �����������A���������������������������������������������������� ���������B�����������
������C��������������������������������������������������������������������������������������������������������������������������������������������������"�
"������ )������6��� C�!����� � �� � �/<.���� �� �����'�40�������������������������������������������������-�
"������ )������6���%C�!����� � * �/<.���� �� �����'�40�����������������������������������������������������$�
!���� �����������A����� �������� � ��������������������������������� ���������������
��##�DDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDDD��%�

������
��� �����������
��� ����&������������������������������������������������������������������������������������������������������������������������������������������
��� ����&�������������������������������������������������������������������������������������������������������������������������������������������
��� ����&��� ��������������������������������������������������������������������������������������������������������������������������������������
��� ����&������ ����������������!=�8!<)�������������������������������������������������������������������������������������������#�
��� ����&���������������� ����������������!=�8!<)���������������������������������������������������������������������"�
��� ��!������������������������������ ������ ����������!=�8!<)����������������������������
.=' �=)��)A;��;.)���������������������������������������������������������������������������������������������������������������������������"�
��� ��#�&������ ����������������������������������� ��������.)�<..:!=�8!<)��<4�)�<)�������������
��� ��"�&���������������� ����������������������������������������������������������������������������������������������#�
��� ��9�&������ ���� ������������������������������������������������������������������������������������������������������"�
��� ���%�&��������������������������������.���<4�)�<)��� ��������B��������������������������������������������%�
��� �����&���������������������� ������B����������������=��=�����!�����������������������������������
��� ��������������������������������� ������� ��������������������������������������������������������������
��� �����&������ ����A���������� �=�����=������ � ����������������������������������������������������������������
��� �����&������������������������������������9�������� &%�&�����������������������������������������������
��� �����&������ �����7�=�&������&�&7�E=�E&������&�E&�������������8������8�������������������������������!�
��� ���!�&������ �����7�=�=�=!&������&�&7�E=�E&������&�E&�������������8������8��������������������"�
��� ���#�&������ �����7�=�=!&�����&�&7�E=�E&������&�E&�������������8������8����������������������������%�
��� ���"�&�������������������.���<4�)�<)����������������������������������������������������������������������������!�
��� ���9�&��������B����������������������.���<4�)�<)������B���������������������������������������������������������!%�
��� ���%�&�������� ������������������������������������������9�������#9����������������������������!��
��� �����&���������������!=�8!<)�����������������������������������������������������������������������������������������!��
��� �����&���������������������������������������� �������.���<4�)�<)������������������������������������������!��
��� �����&�������������������������� ������!=�8!<)�,�99�����������������������������������������������������������������!9�
��� �����&���������������7 �������8���������������������������������������������������������������������������������#��
��� �����&���������������������&��&��"�����������������������������������������������������������������������������������������#��
��� ���!A������������������F���7���������������8����������������������������������������������������������������������#��
��� ���#A�� ������������ �� �����F���������������������������������������������������������������������������#!�
��� ���"�&�� � ����������� ���������������������������������������������������������������������������������������������������##�
��� ���9�&���������� � ���������� �����7�=�=�=!&������&�&7�E=�E&������&�E&
�������������8������8���������������������������������������������������������������������������������������������������������������#"�
��� ���%�&�� � ���������� �����7�=�=!&�����&�&7�E=�E&������&�E&�������������8������8�����#9�
��� �����&������ ����!�����"������������������������������������������������������������������������������������������"%�
��� �����&�� � ���������� ���#�7�=!&������&�&7�E=�E=!E&�����&�E&�������������8������������"��
��� �����&���������������4=1�:�:��;��'<:7:4�)'������B���������������!=�8!<)�������������������������"��
��� �����&���� ������������ ������������������������������������������ �����������������������������������"��

�G��
��� ������� ��
�������&�����������������������������������B�����.���<4�)�<)�������B�����������������������������������������9�
�������&������������������������������B����������������������������������������������������������������������������������������
�������&�!��)@)4)�7�%"��8���������B��������������������������������������������������������������������������������������������
�������&������������&��&��"&�����������������������������������������������������������������������������#��
�������&������������&� ������F�#������������������������������������������������������������������������������������#��
�����!�&���������������4=1�:�:��;��'<:7:4�)'�������������!=�8!<)������������������������������������������������������"�
��� ���� �����
�������&�!=�8!<)��������������������� ��� �������������������������������������������������������������������������������%�
�������&������������������������������������� ������������������������������������������������������������������������������
�������&������������������������������������� �����������������������������������������������������������������������������#�
�������&������������������������������������� �����������������������������������������������������������������������������%�
�������&�������������������������������� ���������������������������������������������������������������������������������������#9�
�����!�&�������������������������������� ����!�����"������������������������������������������������������������������������"%�
�����#�&�������������������������������� ���#�����������������������������������������������������������������������������������"�
��� ����������
���������&�����������������������������H�&��������������������������������������������������������������������������
���������&���$ ��&��$ �����������������������������������������������������������������������������������������������#�
���������&�����&������������� �����������$ �����������������������������������������������������������������������9

G��
��� ����������� �����
� – Chemical Shift
% – Percent
� – Wavelength
4Br-OH-OMe-PBDE – 3,5-dibromo-2-(3’,5’-dibromo-2’-methoxyphenoxy)phenol 79Br – Isomer of bromine79 81Br – Isomer of bromine81 35Cl – Isomer of chlorine35 37Cl – Isomer of chlorine37 13CNMR – Carbon Nuclear Magnetic Resonance Spectroscopy 1HNMR – Proton Nuclear Magnetic Resonance Spectroscopy
ACN – Acetonitrile
AFD – Agence Française pour le Développement (French Agency
for Development)
AIDS – Acquired Immuno-Deficiency Syndrome
APCI – Atmospheric Pressure Chemical Ionisation
Ca. albicans – Candida albicans
CARD-FISH – Catalyzed reporter deposition fluorescence in-situ
hybridization
CD3OCD3 – Deuterated Acetone
CDCl3 – Deuterated chloroform
CI – Conservation International
Cr. neoformans – Cryptococcus neoformans
CRISP – Coral Reef Initiative in the South Pacific
CROUS – Centre Régional des Œuvres Universitaires et Scolaires
D. arenaria – Dysidea arenaria
D. avara – Dysidea avara
D2O – Deuterated water
DCM – Dichloromethane
DMSO-d6 – Deuterated Dimethyl sulphoxide
DNA – Deoxyribonucleic acid
ELS – Evaporative Light Scattering (detector)
ESI – Electrospray Ionisation

G���
EtOAc – Ethyl acetate
EtOH – Ethanol
FACS – Fluorescent Activated Cell Sorter
FDA – Food and Drug Administration
g – Grams
GGTase-1 – Gerany geranyl transferase-1
H2O – Water
HCl – Hydrochloric acid
HIV – Human Immunodeficiency Virus
HMBC – Heteronuclear Multiple Bond Correlation
HPLC – High Performance Liquid Chromatography
HSQC – Heteronuclear Single Quantum Correlation
IRD – Institut de Recherche pour le Développement (Institute of
Research for Development)
IFRECOR – L’Initiative Française sur les Récifs Coralliens (The French
Coral Reef Initiative)
ITS – Internal Transcribed Spacers
J – Coupling constant
L. herbacea – Lamellodysidea (Dysidea) herbacea
LCBE – Laboratoire de Chimie, des Biomolécules et des
l’Environnement
LC-MS – Liquid Chromatography-Mass Spectroscopy
m/z – Mass to charge ratio
MeOD – Deuterated methanol
MeOH – Methanol
mg – Milligrams
MHz – Mega Hertz
mL – Millilitre
mu – Mass units
NMT – N-myristoyl Transferase
NaCl – Sodium chloride
Na2SO4 – Sodium sulphate
NI – Negative Ion
nm – Nanometers

G����
O. spongeliae – Oscillatoria spongeliae
PBDE – Polybrominated Diphenyl Ether
PDA – Photo Diode Array detector
PFT – Protein farnesyl transferase
PI – Positive ion
PLA2 – Phospholipase A2
ppm – Parts per million
RBF – Round bottom flask
Rf – Retention factor
Rt – Retention time
SDA – Sabouraud Dextrose Agar
SPE – Solid Phase Extraction
TLC – Thin Layer Chromatography
μL – Microlitre
UNF – United Nations Foundation
UV – Ultraviolet
VPA – Vanillin Phosphoric Acid
WTCA – Wild Type Candida albicans
WWF – World Wildlife Fund
�
�
�
�
�
�

G�����
��� ����� ��� ����
O
OHOR
R1
Br
R2
Br
Br
Br1
2
34
5
61'2'
3'
4'5'
6'
Compound 1 R = Me; R1 = R2 = H
3,5-dibromo-2-(3’,5’-dibromo-2’-methoxyphenoxy)phenol
Compound 2 R = H; R1 = R2 = Br
3,4,5,6-tetrabromo-2-(3’,5’-dibromo-2’-hydroxyphenoxy)phenol
Compound 3 R = H; R1 = Br; R2 = H
3,5,6-tribromo-2-(3’,5’-dibromo-2’-hydroxyphenoxy)phenol
Compound 4 R = Me; R1 = Br; R2 = H
3,5,6-tribromo-2-(3’,5’-dibromo-2’-methoxyphenoxy)phenol
Compound 5 R = Me; R1 = R2 = Br
3,4,5,6-tetrabromo-2-(3’,5’-dibromo-2’-methoxyphenoxy)phenol
O
OHOR
12
34
5
61'2'
3'
4'5'
6'
Br Br
Br
Br
R3
Compound 6 R = Me; R3 = H
4,6-dibromo-2-(4’,6’-dibromo-2’-methoxyphenoxy)phenol
Compound 7 R = H; R3 = Br
4,6-dibromo-2-(4’,5’,6’-tribromo-2’-hydroxyphenoxy)phenol
Compound 8 R = Me; R3 = Br
4,6-dibromo-2-(4’,5’,6’-tribromo-2’-methoxyphenoxy)phenol

G����
O
OHBr
Br
R4
Br
12
34
5
61'2'
3'
4'5'
6'
Br
Compound 9 R4 = H
3,5-dibromo-2-(2’,4’-dibromophenoxy)phenol
Compound 10 R4 = Br
3,4,5-tribromo-2-(2’,4’-dibromophenoxy)phenol

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
� �����������
��� �����������
Oceans cover more than seventy percent of the surface of the Earth. It is an essential
element for life, and is also home to a vast number of organisms of all kinds, such as
phytoplanktons, fish, marine invertebrates and plants, to name a few. These
organisms have a very challenging environment for survival – it is constantly in
motion, nutrients are difficult to find, and there are risks of predation, competition
and fouling. It is even more challenging for sessile marine organisms to survive in
this environment. However, despite the difficulties of survival, the marine organisms
have developed remarkable means of protection and reproduction, and in many cases
this is achieved by utilizing their own biochemicals.
Biochemical defense mechanisms are subtle and widespread in the plant and animal
kingdom. Most of the marine invertebrates use their secondary metabolites as
chemical defence for protection from predators, to avoid competition and overgrowth
(Debitus, 1998), and to resist malignant microbial infection (Garson, 2001). These
metabolites are produced as a result of transformations and interconversions of the
vast number of organic compounds produced during the metabolic pathways of these
organisms (Voet & Voet, 2004). It is these secondary metabolites that have attracted
researchers from all over the world in the hope to find products that can be useful to
mankind in the form of drugs, as unique templates to serve as a starting point for
synthetic analogues, as interesting tools that can be used to better understand
biological processes (Farnswort, 1984; Sladi� & Gaši�, 2006), and for
chemotaxonomic analysis, which is the classification of organisms based on their
unique chemical components (Erpenbeck & Van Soest, 2006).
The biochemistry of marine invertebrate organisms has been gathering great interest
(Bergquist, 1978) for the past three decades. This has been encouraged mainly by the
recognition that the search for natural products with new molecular structures is
likely to prove rewarding in this little researched area (McConnell, Longley, &
Koehn, 1994).
Considering the identification of these marine organisms however, it is clear that
many specimens had been misclassified previously, and hence need to be re-

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
classified correctly. In the past, classification of organisms was based generally on
morphological characteristics. Recent advancement in technology has led to the use
of electron microscopy to aid in the study of internal sections of organisms, and
genetic mapping for a much closer comparison. However, it is evident with the
various misclassifications of organisms that electron microscopy and genetic
mapping alone can not resolve all the problems of taxonomy. Hence, an additional,
rapid and simple method is required which can be used together with the current
methods to provide the necessary information needed for the proper taxonomy of the
numerous organisms.
�� ����������������������������������������
�� �� ���������� ���!"#��$� ��%��&��#'��
The first marine invertebrate group to be studied in any comprehensive way by those
searching for new compounds was the sponges (sedentary, filter-feeding metazoans
of the phylum Porifera) (Bergquist, 1978; Lévi, 1998). Up to now they have yielded
a great number of novel compounds over a very diverse chemical spectrum (Blunt, et
al., 2006), and there are still possibilities of more novel compounds to be discovered.
Metabolites have been discovered in simple and complex arrangements; many are
also halogenated or contain double bond conjugations and composite configurations,
which are quite unique and difficult to obtain synthetically (Debitus, 1998).
Sponges have exhibited a high incidence of biologically active compounds compared
to other organisms (Belarbi, Gómez, Christi, Camacho, & Grima, 2003; Debitus,
1998) as evidenced either by cytotoxic (Belarbi, Gómez, Christi, Camacho, & Grima,
2003; Haefner, 2003; Debitus, 1998), immunomodulatory (Belarbi, Gómez, Christi,
Camacho, & Grima, 2003; Debitus, 1998) or in some cases, antifouling effects
(Armstrong, McKenzie, & Goldworthy, 1999). It can thus be said that the secondary
metabolites from sponges offer a great potential for application in medicine and
industry (Debitus, 1998).

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
�� � (��&)�*�#��+�*����%�!������
Sponges inhabit a wide variety of marine systems and are found predominantly in
tropical and subtropical oceans but also in polar regions, deep sea and even in fresh
water systems (Hooper & Van Soest, 2002). Marine sponges (Phylum Porifera) are
the most ancient metazoan animal phylum, with a fossil record dating back nearly
600 million years and have existed relatively unchanged in their general structural
organization since the late Cambrian period.
They show a plausible evolutionary jump from unicellular to multicellular
organisms. Because they lack true tissues, these poriferans are also classed as
parazoans, and they exhibit a cellular-level organisation, where each type of cell
specialises to carry out a different task (Mather & Bennett, 1992). Similar cells are
not structured into tissues and bodies, thus forming loose aggregations of similar
types of cells. They also lack muscles, nerves, and internal organs (Brusca & Brusca,
2003).
The taxonomy of sponges depends on the composition of their skeletal materials
(Mather & Bennett, 1992) and their morphological characteristics (Hooper, 1998).
They can either have calcareous spicules (Class Calcarea), siliceous spicules (Class
Hexactinellidae) or spicules made of proteinaceous sponging fibers (Class
Demospongiae).
Furthermore, sponges do not have a definite body shape and are thus divided into
three body types, mainly asconoid, syconoid, and leuconoid (Brusca & Brusca,
2003).
Asconoid sponges are the simplest of all, with a central shaft (spongocoel) and
tubular body. It has several ostia (small pore-like openings) through which water
enters and a central osculum at the top of the sponge to expel water (Brusca &
Brusca, 2003).

Secondary Metabolites Co
�
�
Figu
Syconoid sponges, on
but they contain a
spongocoel (Brusca &
Figu
The most complex or
chambers instead of
canals emptying into
omposition and Geographical Distribution of Marine Sponge
ure 1 - Asconoid sponge (Brusca & Brusca, 2003
n the other hand, also have a tubular body an
thicker body wall with radial canals whi
& Brusca, 2003).
ure 2 - Syconoid sponge (Brusca & Brusca, 2003
rganisation exists in leuconoid sponges consi
a spongocoel, and it also has several incur
a central osculum (Brusca & Brusca, 2003).
es of the Genus Dysidea
� �
3)
nd a single osculum,
ch empty into the
3)
isting of flagellated
rrent and excurrent

Secondary Metabolites Co
�
�
Figu
All the water canals
which beat to create
incoming water, food
in and this sticks to t
and undergo intrace
incoming water is al
metabolic wastes diff
current through the os
Water flow is also a
generally hermaphrod
they do not undergo s
offspring has differen
a blastula forms whic
and floats around unt
into the sessile spo
reproduction is also
budding, whereby the
& Bennett, 1992).
The phylum Porifer
species, with new sp
2003). There are thre
Demospongiae, and
omposition and Geographical Distribution of Marine Sponge
ure 3 - Leuconoid sponge (Brusca & Brusca, 200
s and spongocoel are lined with flagellated
water current to flow through the sponge.
d (mainly bacteria, debris and microscopic alg
the cells in the sponge which are later taken
llular digestion (Mather & Bennett, 1992;
lso rich in oxygen and this is diffused into
fuse out of the cells, flowing out of the spo
sculum (Lévi, 1998).
an important factor for reproduction in spo
dites (male and female gonads are present i
self-fertilization to avoid asexual reproductio
nt characteristics as that of either parent. Ofte
ch is released into the water. The blastula de
til it finds a suitable substrate, after which it s
onge form (Mather & Bennett, 1992; Lév
possible in sponges by the process of e
e parent and offspring have the same geneti
ra encompasses approximately 6000 taxon
pecies still being discovered (Lévi, 1998;
ee main classes of the phylum Porifera as see
d Hexactinellidae. A fourth class of
es of the Genus Dysidea
� �
03)
d choanocyte cells
Together with the
gae) is also brought
into food vacuoles
; Lévi, 1998). The
the cells while the
onge with the water
onges. Sponges are
in one sponge), but
on, and the resulting
en after fertilization,
evelops into a larva
settles and develops
vi, 1998). Asexual
external or internal
c make-up (Mather
nomically validated
Brusca & Brusca,
en earlier: Calcarea,
f sponges (Class

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
Sclerospongiae) has been mentioned in many other texts, but according to the World
Porifera Database, the sclerosponges are polyphyletic, hence accepted as
Demosponges (Van Soest, Boury-Esnault, Janussen, & Hooper, 2005).
Demospongiae are by far the most diverse and ecologically significant group,
whereby 90 % of all species of sponges belong to this class (Lévi, 1998). It includes
approximately 5500 species in more than 10 orders and generally possesses the
leuconoid body type (Brusca & Brusca, 2003). Sponges of this class are asymmetric
and brightly coloured (such as orange, green, yellow, purple, or red) due to the
presence of pigment granules in their amoebocytes.
�� �, �)'��'�$������ �
Sponges of the genus Dysidea belong to the Class Demospongiae, Order
Dictyoceratida, and Family Dysideidae. These sponges lack mineral spicules, and the
main skeleton consists of reticulation of sponging fibers (Bergquist, 1998). By the
year 2001, the genus Dysidea was reported to consist of 17 species (Munro & Blunt,
2001).
Lamellodysidea appears as a new genus in the most recent classification of sponges,
split off from Dysidea, because of the consistent presence of an encrusting basal
plate and the lack of orientation of the skeleton with respect to the surface (Sauleau
& Bourget-Kondracki, 2005). Currently, the genus Lamellodysidea includes the two
species L. herbacea and L. chlorea.
The marine sponge Dysidea is a prolific producer of structurally diverse secondary
metabolites including sesquiterpenes (Sera, Adachi, Nishida, & Shizuri, 1999),
sesterpenes, meroterpenes, and sterols (Cameron, Stapleton, Simonsen, Brecknell, &
Garson, 2000).
The following structures are a few examples of the diversity of compounds found in
sponges of the genus Dysidea:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
O
OMe OH
Br
Br
Br
Br
4,6-dibromo-2-(3',4',6'-tribromo-2'-methoxyphenoxy)phenolN
S
NHO
N
CCl3
O CCl3
Dysidenin
O
OH
OCH3
O
Br
O
OH
H
OHH
OH
OH
H
OH
Herbasterol
H
H
6-hydroxyfurodysinin-O-methyl-lactone
Figure 4 - Compounds isolated from Dysidea species.
Some sesquiterpene derivatives (such as dysidotronic acid; bolaquinone; two
sesquiterpene cyclopentenones [Dysidenones A and B]; and a sesquiterpene
aminoquinone, dysidine) isolated from the Dysidea sp. have shown selective
inhibitor profile against human phospholipase A2 (PLA2) and other leukocyte
functions, such as degranulation process and superoxide production (Giannini, et al.,
2001). PLA2 has also been recognized to be involved in a wide number of
pathophysiological situations, ranging from systemic and acute inflammatory
conditions to cancer (Giannini, et al., 2001; Giannini, et al., 2000).
In addition, certain Dysidea species, exemplified by Lamellodysidea herbacea,
Keller (1889) and L. chlorea, de Laubenfiels (1954), have given rise to
bromophenols (Hanif, et al., 2007) and polychlorinated peptides (Flatt, Gautshci,
Thacker, Musafija-Girt, Crews, & Gerwick, 2005). It is striking that these two
families of halogenated compounds have never been found together in the same
sponge. Hence, marine sponge Lamellodysidea (together with some Dysidea species)
occurs in two chemotypes, one containing sesquiterpenes and polychlorinated
peptides and the other polybrominated diphenyl ethers.
The polychlorinated peptides are an original family of molecules including dysidenin
(Ardá, Jiménez, & Rodríguez, 2004), dysidamides, dysideathiazoles, and chlorinated
diketopiperazines (Goetz, Harrigan, & Likos, 2001). Together with a furodysin
lactone, a well-known Dysidea-derived sesquiterpene, the discovery of the novel

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
polychlorinated dysideaprolines and barbaleucamides was reported by Goetz,
Harrigan, and Likos (2001).
X2HC N
O
R2 O
N
CHY2
R1
S N
1
11 3
5
6
9
12
16
13
19 18
DysideaprolineA - R1 = H, R2 = CH3, X = Y = ClB - R1 = R2 = CH3, X = Y = ClC - R1 = R2 = H, X = Y = ClD - R1 = H, R2 = CH3, X = H, Y = ClE - R1 = H, R2 = CH3, X = Cl, Y = HF - R1 = H, R2 = CH3, X = Cl, Y2 = ClH
Cl3C
O
CH3
N
O
CCl3
R
S N
5
12
14
11
7
1
6
15
10
BarbaleucamideA - R = HB - R = CH3
Figure 5 - Chlorinated compounds isolated from Dysidea sponges
All polychlorinated peptides contain a trichloromethyl functionality on leucine-
derived precursors, and are very similar to pseudodysidenin and barbamide, which
have been isolated from the cyanobacteria Lyngbya majuscula (Jimenez & Scheuer,
2001).
Cl3C
O
CH3
N
OS N
Barbamide
Cl3C N CCl3
O
H
NO CH3
N
SPseudodysidenin
Figure 6 – Examples of chlorinated compounds found in both Dysidea sponges and the
cyanobacteria Lyngbya majuscula

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
Some of these compounds exhibit interesting pharmacological activities such as
inhibition of iodide transport in thyroid cells, neurotoxicity and inhibition of protein
phosphatase (Sauleau, Retailleau, Vacelet, & Bourget-Kondracki, 2005).
About forty different polybrominated diphenyl ethers (PBDEs) have been isolated
from L. herbacea, L. chlorea, D. granulosa, D. dendyi, D. fragilis, and
Phyllospongia foliascens (Table 1). These molecules are reported to exhibit a variety
of bioactivities: antibacterial and antifungal properties, brine shrimp toxicity,
antimicroalgal activity, anti-inflammatory activity, and inhibition of a range of
enzymes implicated in tumor development such as inosine monophosphate
dehydrogenase, guanosine monophosphate synthetase, and 15-lipoxygenase. More
recently, PBDEs have been found to inhibit the assembly of microtubule protein, the
maturation of starfish oocytes, and also Tie2 kinase (Hanif, et al., 2007). This class
of halogenated compounds has been found in unrelated organisms such as the green
alga Cladospora fascicularis (Kuniyoshi, Yamada, & Higa, 1985), red alga
Ceramium tenuicorne, blue mussel Mytilis edilis (Malmvarn, Marsh, Kautsky,
Athanasiadou, Bergman, & Asplund, 2005), and unidentified sponge of the family
Callispongia (Capon, Ghisaberti, Jeffries, Skelton, & White, 1981), gastropterides
Sagaminopteron nigropunctatum and S. psychedelium often found on sponges
(whether they are true sponge feeders or not, is still debated) (Becerro & Paul, 2004),
Baltic salmon and marine mammals (Vetters, Stoll, Garson, Fahey, Gaus, & Muller,
2002). The fact that they are present in a wide variety of marine organisms makes us
think that they are more likely produced by a symbiont than the marine invertebrates
themselves.
Hence, it was suggested that the halogenated compounds from the sponges of this
genus are produced by cyanobacteria living in a symbiotic relationship with the host
sponge (Flowers, Garson, Webb, Dumbei, & Charan, 1998; Ridley, Bergquist,
Harper, Faulkner, Hooper, & Haygood, 2005).
The following table illustrates the different Dysidea species collected at various
geographical locations.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Table 1 - Dysidea species collected in various geographical locations
Dysidea species
Localisation Compound Reference
Lamellodysidea herbacea
Indian Ocean; Zanzibar; Great Barrier Reef, Australia; Indonesia
Polybrominated diphenyl ether
Anjaneyulu, Rao, Radhika, Muralikrishna, & Connolly, 1996; Sionov, et al., 2005; Agrawal & Bowden, 2005; Handayani, et al., 1997; Hanif, et al., 2007
L. herbacea (with symbiotic cyanobacteria Oscillatoria spongeliae)
Papua New Guinea
Polychlorinated peptides; Polychlorinated ketide amino acid (Herbamide A); Polychlorinated tetrapeptide (dysidenin)
Flatt, Gautshci, Thacker, Musafija-Girt, Crews, & Gerwick, 2005; Clark & Crews, 1995
L. herbacea Great Barrier Reef, Australia; Pohnpei, FSM; Red Sea
Chlorinated metabolites; 5,5,5-trichloroleucine metabolite (Herbacic acid); Polychlorinated amino acid derivative; Polychlorinated pyrrolidinones
Dumdei, Simpson, Garson, Byriel, & Kennard, 1997; MacMillan & Molinski, 2000; Unson, Rose, Faulkner, Brinen, Steiner, & Clardy, 1993; Sauleau, Retailleau, Vacelet, & Bourget-Kondracki, 2005
L. herbacea Red Sea; Australia; Ethiopia
Polyhydroxysterols; Ichthyotoxic 9,11-secosterol (Herbasterol)
Sauleau & Bourget-Kondracki, 2005; Capon & Faulkner, 1985; Isaacs, Berman, Kashman, Gebreyesus, & Yosief, 1991
L. herbacea Yap State, Federated States of Micronesia
Betaines Sakai, Suzuki, Shimamoto, & Kamiya, 2004
�,�-disubstituted �-amino acid derivative (Dysibetaine)
Sakai, Oiwa, Takaishi, Kamiya, & Tagawa, 1999
Amino acid (neodysiherbaine A)
Sakai, et al., 2001
L. herbacea Palau; India; Fiji Sesquiterpene; Dysinin-type sesquiterpene
Sera, Adachi, Nishida, & Shizuri, 1999; Venkateswarlu, Biabani, Reddy, Chavakula, & Rao, 1994; Horton, Inman, & Crews, 1990
L. herbacea Ethiopia Dideoxyhexose Isaacs, Berman, Kashman, Gebreyesus, & Yosief, 1991
D. arenaria New Caledonia; FSM; Thailand
Sesquiterpene (9-hydroxyfurodysinin-O-ethyl Lactone); Sesquiterpenoids (arenarol & arenarone); Sesquiterpene ethers (arenarans)
Piggott & Karuso, 2005; Schmitz, Lakshmi, Powell, & van der Helm, 1984; Horton & Crews, 1995
D. arenaria Whitsundays Reef, Australia
Sterol ECTA Jacob, et al., 2003
D. avara Italy; Solomon Is.; Australia
Sesquiterpene hydroquinone (Avarol derivatives); Furanosesquiterpene (Thiofurodysinin)
Crispino, de Giulio, de Rosa, & Strazzullo, 1989; Alvi, Diaz, Crews, Slate, Lee, & Moretti, 1992; Capon & MacLeod, 1987

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
D. fragilis South China Sea Chlorinated diketopiperizine (Dysidamide D)
Fu, Zeng, Su, & Pais, 1997; Su, et al., 1993
D. fragilis Mozambique Brominated diphenyl ether Utkina, Kazantseva, & Denisenko, 1987
D. fragilis Fiji; FSM Azacyclopropene & Derivatives; 2H-Azirines
Molinski & Ireland, 1988; Salomon, Williams, & Faulkner, 1995; Skepper & Molinski, 2008
D. fragilis Spain; Venice; Black Sea; Oahu
Furanosesquiterpenoid; Sesquiterpenes; Steroids; Polyhydroxylated sterols; Sesquiterpenoid bicyclo [3.3.1] nonane aldehyde lactone
Marin, Lopez, Esteban, Meseguer, Munoz, & Fontana, 1998; Aiello, Fattorusso, Menna, & Pansini, 1996; Milkova, Mikhova, Nikolov, Popov, & Andreev, 1992; Schulte, Scheuer, & McConnell, 1980
D. etheria Bermuda �,�-Epoxy �-Lactone Schram & Cardellina, 1985
Polyhydroxylated sterols West & Cardellina, 1988
Indoles Cardellina, Nigh, & VanWagenen, 1986
Sesquiterpene lactone (furodysinin lactone)
Grode & Cardellina, 1984
Furanosesquiterpenes (Nakafurans)
Cardellina & Barnekow, 1988
D. etheria Caribbean Sesterterpene (Dysidiolide) Takahashi, Dodo, Hashimoto, & Shirai, 2000 Gunasekera, McCarthy, Kelly-Borges, Lobkovsky, & Clardy, 1996
D. fusca New Caledonia (East Coast)
Drimane sesquiterpene Montagnac, Martin, Debitus, & Pais, 1996
D. chlorea Yap State, FSM Polychlorinate diketopiperazines (dysidamides I-T)
Fu, Ferreira, Schmitz, & Kelly-Borges, 1998
Sterol
D. chlorea Okinawa Sesquiterpenes (Heterumadysins A-D)
Ueda, Kadekaru, Siwu, Kita, & Uemura, 2006
D. cinerea Dahlak archipelago, Eritrea (Red Sea)
Sesterpenes (Bilosespens A & B)
Rudi, Yosief, Schleyer, & Kashman, 1999
D. frondosa Pohnpei Sesquiterpene hydroquinone deriv.
Patil, et al., 1997
D. cristagalli Spirits Bay, Northland, New Zealand
Sesquiterpene quinones McNamara, et al., 2005
D. amblia CA, USA Diterpenes Walker & Faulkner, 1981; Walker, Rosser, & Faulkner, 1984
D. dendyi Scott reef, North-west Australia
Brominated dibenzo-p-dioxins (spongiadioxin A & B)
Utkina, et al., 2001

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
D. incrustans Coast of Tunisia, Mediterranean Sea
Polyoxygenated steroids (incrustasterol A & B)
Casapullo, Minale, Zollo, Roussakis, & Verbist, 1995
D. reformensis Gulf of California, Pacific Ocean, Mexico
Sesquiterpene hydroquinone
Carballo, Zubía, & Ortega, 2006 D. cachui
D. uriae Furanosesquiterpene (dendrolasin)
Dysidea sp. FSM; Philippines; Fiji
Polybrominated diphenyl ether
Fu & Schmitz, 1996; Zhang, Skildum, Stromquist, Rose-Hellekant, & Chang, 2007; Salva & Faulkner, 1990; Fu, Schmitz, Govindan, & Abbas, 1995
Dysidea sp. Philippines; Indonesia
Polychlorinated compounds (Dysideaprolines A-F & Barbaleucamides A-B); Polychlorinated dipeptide
Harrigan, Goetz, Luesch, Yang, & Likos, 2001; Ardá, et al., 2005
Dysidea sp. Palau; Red Sea; Australia; Philippines; Guam; Vanuatu; USA; PNG; South China Sea
Diterpenes of the dolabellane class; 14-membered macrolide; Diterpenes; Sesquiterpene ester (7-deacetoxy-olepupuane); Sesquiterpenes (Furodysin lactone & Pyrodysinoic acid); Drimane sesquiterpenoid; Sesquiterpene cyclopentenones and aminoquinones; Merosesquiterpenes; Meroterpenoid (Avinosol); Sesquiterpene hydroquinone (Bolinaquinone); Tetracyclic sesquiterpene - substituted quinone (Puupehenone)
Lu & Faulkner, 1998; Carmely, Cojocaru, Loya, & Kashman, 1988; Garson, Dexter, Lambert, & Liokas, 1992; Goetz, Harrigan, & Likos, 2001; Paul, Seo, Cho, Rho, Shin, & Bergquist, 1997; Giannini, et al., 2001; Pérez-García, Zubía, Ortega, & Carballo, 2005; Diaz-Marrero, et al., 2006; de Guzman, et al., 1998; Ciavatta, et al., 2007
Dysidea sp. Okinawa Benzothiazole-2-one analog
Suzuki, et al., 1999
Phyllospongia dendyi
Palau Polybrominated diphenyl ethers
Liu, Namikoshi, Meguro, Nagai, Kobayashi, & Yao, 2004
It was also noticed that most of the specimen collected and studied were obtained
from the tropical regions between the tropics of Cancer and Capricorn, as seen in the
next figure. Furthermore, there were no other distinguishable patterns of distribution
of Dysidea species with regards to the halogenated compounds isolated from them.
The figure below also defines the distribution of brominated and chlorinated
compounds obtained from the Dysidea species.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Figure 7 - Distribution of brominated (green) and chlorinated (pink) compounds from
Lamellodysidea herbacea
��, �������-���(�����������������
Sponges have been the focus of much recent interest due to the following two main
(and often interrelated) factors: they are a rich source of biologically active
secondary metabolites, and they form close association with a wide variety of
microorganisms.
Certain Demosponges are known to be associated with enormous amounts of
microorganisms contributing up to 40 – 60 % of the animal biomass, with density in
excess of 109 microbial cells per mL of sponge tissue, several orders of magnitude
higher than those typical for sea water (Hoffmann, Rapp, & Reitner, 2005). The vast
majority of these microorganisms are located extracellularly within the mesohyl
matrix (Becerro & Paul, 2004).
Investigation of these microorganisms through phylogenetic analysis and microscopy
has shown that there are members of at least eight different bacterial phyla
(Proteobacteria, Acidobacteria, Actinobacteria, Bacteroides, Chloroflexi,
Cyanobacteria, Gemmatimonadetes, Nitrospira), one bacterial candidate phylum
(Poribacteria) and one archaeal phylum (Crenarchaeota) (Grozdanov & Hentschel,
2007).
Although unrelated sponges can harbour similar microbial groups, it is more typical
for the symbiont communities to be specific for a given sponge species, irrespective

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
of the collection site in a geographic region (Thacker & Starnes, 2003). It has been
shown that the microbial communities are passed on to the next sponge generation by
vertical transmission through the reproductive stages such as embryos and larvae
(Grozdanov & Hentschel, 2007).
The symbiont community can serve as a source of nutrition by transferring products
of metabolite processes to the host. For example, symbiotic cyanobacteria have been
shown to transfer organic carbon obtained through photosynthesis to the host
(Ridley, Faulkner, & Haygood, 2005). Other suspected benefits of the symbionts
include contribution to the structural rigidity of sponge and production of
halogenated natural products (Hildebrandt, Waggoner, Lim, Sharp, Ridley, &
Haygood, 2004).
Experimental support for the hypothesis of halogenated metabolites produced by
symbionts was gained when Unson and Faulkner (1993) investigated a specimen of
L. herbacea which contained the chlorinated compound 13-demethylisodysidenin
and were able to separate the auto-fluorescent cyanobacterium from the sponge cells
using a fluorescent activated cell sorter (FACS). Chemical examination revealed that
only cyanobacteria Oscillatoria spongeliae, the major symbiont of the sponge,
contained the chlorinated metabolite. Further experiments on a L. herbacea
containing chlorinated diketopiperazines (Flowers, Garson, Webb, Dumbei, &
Charan, 1998) and on a L. herbacea containing polybrominated diphenyl ethers
(Unson, Holland, & Faulkner, 1991) confirmed this hypothesis.
Additionally, a putative halogenase gene cluster involved in the chlorination of
barbamide has been identified and used to amplify a homolog sequence from L.
herbacea samples. Catalyzed reporter deposition fluorescence in-situ hybridization
(CARD-FISH) analysis showed that halogenase gene homologues are situated in the
cyanobacteria symbiont (Flatt, Gautshci, Thacker, Musafija-Girt, Crews, & Gerwick,
2005). Therefore O. spongeliae is likely the biosynthetic source of the chlorinated
metabolites. But there is still no evidence that this cyanobacterium is also responsible
for the production of brominated metabolites since a marine bacterium (not the
cyanobacterium O. spongeliae) isolated from the tropical Dysidea sp. has been
cultured on medium broth and has biosynthesized polybrominated diphenyl ethers
(Elyakov, Kuznetsova, Mikhailov, Maltsev, Voinov, & Fedoreyev, 1991; Voinov,
Elkin, Kuznetsova, Maltsev, Mikhailov, & Sasunkevich, 1991). There is still no

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
report of O. spongeliae cultured independently from the host sponge producing these
metabolites. Furthermore, the Caribbean sponge Oligoceras violacea (syn. O.
hemorrhages) has been found to be heavily associated with O. spongeliae, but it
contains metabolites consisting of simple terpenes and dendrolasin as a major
constituent (Flatt, Gautshci, Thacker, Musafija-Girt, Crews, & Gerwick, 2005).
��. ���������(�����������������(���/���(��
Comparative biochemistry has been applied to sponge classification at frequent
intervals (Bergquist, 1978), but serious attempts to combine comprehensive
taxonomy with detailed chemistry have been rare. It is known that certain structural
types of sponge metabolites occur in certain sponge families or orders and targeting
of particular sponge groups can be useful in the search for new chemical variants
(Debitus, 1998). Therefore, it is important that taxonomic and chemical programs
proceed together.
Chemotaxonomy is not a new branch of science, and it has been explored previously.
However, interest in the subject with regards to sponges has declined over the past
few years due to certain drawbacks such as ambiguity in the exclusive presence of
chemical markers, availability of specimen, homology of the compounds, their
variability in occurrence, and whether the compound is produced by the organism
being studied or by its symbiont (Erpenbeck & Van Soest, 2006).
In the past, there have been a lot of misclassifications of many organisms, with
sponges being one of them due to their primitive features (Erpenbeck & Van Soest,
2006), and over the accumulated years, many stored specimen would have lost their
very basic morphological characteristics which were initially used to classify them.
By the effective use of chemotaxonomy, these specimens can be reclassified
correctly, providing better knowledge of the organisms and their actual taxonomic
position. Furthermore, the presence or absence of particular chemical markers from
similar groups of organisms can provide a link to the phylogenetic relationships
(Erpenbeck & Van Soest, 2006).
Moreover, it has been seen that only a few stable and suitable chemical markers (for
example sterols, protein banding, steroids, scalarane sesterterpenes, alkylpiperidines,
fatty acids, brominated derivatives, pyrrole-2-carboxylic acid derivatives and other
compounds) have been identified for the higher taxa of sponges (Erpenbeck & Van

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Soest, 2006), but it is still not very clear on the lower species level. Hence, it was
interesting to carry out this study on the sponges of the genus Dysidea, since
considerable information is available on the types of compounds present in these
sponges, but the classification of the genus Dysidea is very difficult to carry out due
to the plasticity of their morphological characteristics. Added to this, any interesting
compounds coming across during this study were subject to isolation and
characterization to further aid in the chemotaxonomical analysis.
��0 ��������������������������1����
Drug discovery, production and sales are a world-wide multi-billion dollar industry
that impacts all levels of modern life. The quality of life is greatly dependent on the
continued access to safe and effective drugs, whether it is in the form of antibiotics
for the treatment of infections; analgesics for pain relief; specific drugs for complex
ailments such as cancer, malaria, or blood pressure, and formulations for pest control
in the homes, agricultural products, and livestock. Most of these drugs mainly have
the active ingredient that is either a bioactive natural product or synthetic derivative
of these natural products.
Although terrestrial plants and organisms have provided great medicines in the past
and still manage to produce effective natural products for pharmacology, researchers
are now slowly moving their focus on marine derived natural products for research
and drug discovery. It has been reported by Walsh (2004) that biotechnology experts
believe that the prospect of finding a useful bioactive natural product from marine
sources is 500 times higher than from terrestrial sources.
Over the past two decades, marine invertebrates have been a rich source of
biologically active and structurally unique secondary metabolites (Faulkner, 2000)
having high potential to be developed for disease treatment (Debitus, 1998). Among
these potential pharmaceuticals include bengamides A and B (isolated from Jaspidae
marine sponges) first reported in 1986 by Quinoa, et al., having antihelminthic
activity; jaspamide (from the genus Jaspis) having herbicidal and fungicidal activities
(Peng, et al., 2003); and cytotoxic activities of arenastatin A, a cyclodepsipeptide
isolated from Dysidea arenaria (Newman & Cragg, 2004).

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
HN
OH OH
OH O
O
N
OR
HH3C(H2C)12CO2Bengamide A: R = HBengamide B: R = CH3
OH
HN O
O
NH
O
N O
O
NH
BrH3C
Jaspamide
O
O
H
OO
O
H
NH
HNO
H HO
H
O
Arenastatin A
Figure 8 - Potential pharmaceuticals from marine sponges
Despite the large number of compounds and the high variety of structurally different
natural products isolated, before the year 2004, no marine derived natural product
had reached the commercial sector (Sladi� & Gaši�, 2006), although quite a number
of interesting marine-derived compounds were in the process of approaching phase II
and phase III clinical trials for cancer, analgesia, allergy and cognitive diseases
(Newman & Cragg, 2004; Haefner, 2003).
However, there were exceptions. After years of research, a medical break-through
was seen when scientists identified the cure for acute myeloid leukemia from a
natural product derived from the Caribbean sponge, Cryptotetheya crypta (Thakur &
Müller, 2004). By the year 2004, 1-beta-D-arabinofuranosylcytosine (also known as
ARA-C or Arabinosyl Cytosine) was a marine derived natural product to provide a
product for clinical use. This drug is marketed under the brand name Cytosar-UR by
Pharmacia & Upjohn Company (Thakur & Müller, 2004; Newman & Cragg, 2004).
There was also another product, adenine arabinoside (Ara-A), which was
commercialized by Glaxo Smith-Kline as an antiviral agent (Newman & Cragg,
2004; Haefner, 2003).
Ziconotide is another natural product of marine origin for clinical use granted
recently by the United States Food and Drug Administration (FDA) (Skov, Beck, de
Kater, & Shopp, 2007). It is the synthetic form of the natural product �-conotoxin
M-VII-A, a peptide derived from the cone snail Conus magus. This compound is an
N-type calcium channel blocker, and is used as an analgesic to improve chronic pain.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Ziconotide is marketed under the brand name Prialt. Trabectedin, also known as
ecteinascidin 743 or ET-743, has been derived from the sea squirt Ecteinascidia
turbinata (Fayette, Coquard, Alberti, Ranchere, Boyle, & Blay, 2005). It is in the late
stages of clinical trials as a chemotherapeutic and anti-neoplastic agent, and is
developed under the brand name Yondelis by Zeltia and Johnson and Johnson as an
anti-tumor drug. ET-743 acts by binding in the minor groove of DNA, blocking
transcription factors activity, and trapping protein from the nucleotide excision repair
system. Thus, it blocks cells in the G2-M phase and causes cell cycle arrest.
��2 �������������������
Fungal infections are increasingly becoming important medical problems as more
strains are now found to be resistant to the classical medications. There are a limited
number of effective antifungal agents in use, even though a lot of progress is being
made in the development of different drugs for the treatment of systemic mycoses
(Sionov, et al., 2005). Human fungal infections are most frequently caused by
Candida and Aspergillus species (Sionov, et al., 2005), and also Cryptococcus
species (Wang & Casadevall, 1996).
Fungi of the genus Cryptococcus and Candida belong to the same family
Cryptococcaceae (Order Cryptococcales) (Mehrotra & Aneja, 1990). They are the
imperfect forms of basidiomycetous and ascomycetous yeasts. Candida albicans and
Cryptococcus neoformans are opportunistic fungal pathogens of this order, and are
greatly found in patients with compromised immune systems (Jacob, et al., 2003;
Wang & Casadevall, 1996; Rhome, et al., 2007). Such patients include human
immunodeficiency virus / acquired immuno deficiency syndrome (HIV/AIDS)
patients, cancer patients undergoing intensive chemotherapy, recipients of bone
marrow and solid organ transplants with acute neutropenia, and also individuals
undergoing prolonged corticosteroid and antibiotic treatments (Sionov, et al., 2005).
Invasive mycoses can be life-threatening for such patients even though some may
have low virulence.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
��2�� ��&��!�!!$���'�%��3����
The Cryptococcus species are encapsulated basidiomycetous yeasts (Rhome, et al.,
2007). They are widely distributed in soil and on decaying plant materials (Mehrotra
& Aneja, 1990). Cryptococcus neoformans and Cryptococcus gattii are two species
pathogenic to humans, and cause life-threatening meningoencephalitis. It is readily
air-borne, and the lung is the primary site of infection, with consequent metastasis to
other parts of the body (Phaff & Macmillan, 1978). It affects the body by the
overgrowth of yeast-like cells that crowd the normal cells so that they do not
function properly. Cr. neoformans, the asexual state of Filobasidiella neoformans
(Mehrotra & Aneja, 1990), causes a vast majority of cryptococcoses and life
threatening infections in 6 – 8 % of AIDS patients (Wang & Casadevall, 1996;
Martinez, Garcia-Rivera, & Casadevall, 2001).
Some of the virulence factors of Cr. neoformans identified in recent studies include
the presence of a capsule, its ability to grow at 37 oC, and the production of melanin
(Wang & Casadevall, 1996). Cr. neoformans can catalyze the formation of melanin
from a variety of phenolic precursors due to the presence of the phenoloxidase
enzyme system. Melanin is said to contribute to virulence after in vitro studies
suggest that it protects fungal cells against oxygen- and nitrogen- derived reactive
intermediates produced by the host effector cells. Melanized cells are hence able to
survive longer since they cannot be efficiently killed in vitro by microglia and
macrophages, and also because they are less susceptible to Amphotericin B, which is
the first line of defence against fungal infections. This suggests that cryptococcal
infections are persistent even with fungicidal therapy.
Another virulence factor under current research is the biosynthetic pathways of the
cryptococcal sphingolipids (Rhome, et al., 2007). It has been identified in this study
that the pathways provide an extremely rich reservoir of sphingolipid molecules and
fungus-specific metabolising enzymes. These enzymes regulate many cellular
functions essential for the viability, virulence and pathogenicity of the fungus.
Further studies are still being carried out on this factor, but it can be said that the
sphingolipid metabolism can provide new and better pharmacological targets.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
��2� �� � ���*4�!����
Fungi of the genus Candida are yeast-like organisms with a strong mycelial or
pseudomycelial growth and multilateral budding (Mehrotra & Aneja, 1990). They
have been isolated from soil, plant and animal material, animal faeces, and water.
Fungal infections caused by Candida species are more common compared to
Cryptococcal infections. Candida species belong to the ascomycetes group of fungi
(Vallim, Fernandes, & Alspaugh, 2004), and Candida albicans is the most common
fungi found in this genus. They are also opportunistic fungi and target mainly
immunocompromised individuals (Sionov, et al., 2005), similar to Cryptococcus
neoformans. Ca. albicans grows as yeast in man (Taber & Taber, 1978), and may
also attack the skin on damp folds on the body, such as the vagina (vaginal thrush) or
the mucous membranes in the mouth (oral thrush) (Stevenson, 1967; Mehrotra &
Aneja, 1990; Moran, Sullivan, & Coleman, 2002), as well as cause cutaneous
candidiasis, bronchocandidiasis and pulmonary candidiasis (Mehrotra & Aneja,
1990; Hunter, Barnett, & Buckelew, 1978, Macmillan & Phaff, 1978).
Over the years, Candida albicans has developed resistance against antifungal agents
such as imidazoles and triazoles (collectively known as “azoles”) because HIV
patients had been kept on low dose prophylactic fluconazole therapy to prevent
opportunistic fungal infections (Jacob, et al., 2003; Moran, Sullivan, & Coleman,
2002). This developing resistance calls for the search of new bioactive compounds
that can help in the cure of these infections.
��2�, �'5����#'���%�������3�!��4��*���$#��'6'*�&3'���
The first anti-fungal agent to be identified was Nystatin, by Hazen and Brown in
1950 (Calderone, 2002). It is a polyene in nature and has since encouraged the
discovery of other polyene-type antifungal agents, including amphotericine B. The
increase in fungal diseases later called for the development of azoles (Calderone,
2002). Azole-based antifungals consist of a range of major clinically important and
useful drugs, such as miconazole, fluconazole, ketoconazole, and itraconazole.
Many of the drugs currently available have undesirable side-effects and may be toxic
to some extent (Sionov, et al., 2005). The steady rise in the occurrence of Candida
infections over the years (Calderone, 2002), and the difficulty in treating

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Cryptococcus and Candida infections calls for the discovery and design of new drug
targets which can be more efficient and effective in treating such life-threatening
fungal infections (Sionov, et al., 2005). The fact that the limited number of antifungal
drugs that are available have restricted sources, and that they have similar modes of
activity makes it important to search for new agents that exhibit antifungal activity
with different modes of activity or that affect different sites of the fungal cells.
Taking this into account, recent studies (Gelb, et al., 2003; Vallim, Fernandes, &
Alspaugh, 2004) have been carried out to explore gene sequences and enzymes of
microbes as new targets for the development of therapeutics. Genetic studies have
shown that the gene coding for protein N-myristoyl transferase (NMT), is essential
for the viability of a range of species including Candida albicans, Cryptococcus
neoformans, Sacharromyces cerevisiae, and Drosophila melanogaster (Gelb, et al.,
2003). Inhibitors of NMT exhibit both selectivity and specificity in the treatment of
such fungal infections. This protein is found to be indispensible for most eukaryotes,
hence making it a candidate as drug target for human pathogens on the condition that
specific reagents can be developed, which are selective against the pathogen rather
than the human enzyme.
Another enzyme, the protein farnesyl transferase (PFT), is well established in anti-
tumor studies. Inhibitors of PFT are able to arrest the growth of transformed cells and
cause tumor xenographs (tissue transplant from one species to an unrelated species)
to shrink (Gelb, et al., 2003). They exhibit known pharmacokinetic properties, and
low cytotoxicity to humans when the therapeutic doses are exceeded. It has been
seen in a study (Vallim, Fernandes, & Alspaugh, 2004) that pharmacological
inhibition of this enzyme can cause dose-dependant cytostasis and prevention of
hyphal differentiation of Cr. neoformans. Hence, the RAM1 gene which codes for
the �-subunit of PFT is essential for the viability of Cryptococcus neoformans.
It has also been seen that in ascomycetous yeasts, such as Candida albicans and
Saccharomyces cerevisiae, PFT works with another enzyme geranylgeranyl
transferase 1 (GGTase-1) to partially compensate the absence of each other, thus
preventing any fatal effects to the fungi due to their absence (Vallim, Fernandes, &
Alspaugh, 2004). On the other hand, in the distantly related basidiomycetous fungus
Cryptococcus neoformans, PFT is not readily complemented by any other enzyme,
therefore its activity is vital for the survival of the fungi. This result suggests that

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
protein prenylation is an interesting target for the further development of anti-
microbial drugs.
In this study, we investigated the activity of crude sponge extracts against Cr.
neoformans and Ca. albicans to see if the growth of both the species of fungi is
inhibited or not. Based on the results obtained, suitable conclusions and hypotheses
about the mode of activity of the products have been suggested for future
investigations.
��7 ����1������������
This study is part of a major project of the Coral Reef Initiative in the South Pacific
(CRISP) program. The CRISP program is funded by major world organizations
including Agence Française pour le Développement (French Agency for
Development - AFD), French Global Environment Facility, United Nations
Foundation (UNF), French Ministry of Foreign Affairs, Conservation International
(CI), World Wild Life (WWF), Institut de Recherche pour le Développement
(Institute of Research for Development - IRD) and L’Initiative Française sur les
Récifs Coralliens (The French Coral Reef Initiative - IFRECOR). Apart from these
organizations, there are several other technical and institutional partners.
The main aim of this program as a whole is to provide initiative for the protection
and sustainable development of coral reefs in the South Pacific, and involves a wide
range of scientific disciplines of study. The CRISP program is divided into three
main components: Marine Protected Areas and Integrated Coastal Management;
Knowledge, Management, Rehabilitation and Development of Coral Ecosystems;
and Institutional and Technical Support, Communication, Coordination and
Extension. This project is a contributing part of the second component of the CRISP
program (Knowledge, Management, Rehabilitation and Development of Coral
Ecosystems).
One of the main objectives of this project is to analyze and compare the chemical
characteristics and constituents of sponges of the Genus Dysidea, which can be
related to the chemical taxonomy of these organisms; and to highlight the external
factors affecting secondary metabolites production in these sponges. The
intraspecific variability often observed in metabolite production may be related to
environmental factors, the physiological state of the organism or its genotype.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Endosymbiotic microorganisms might also influence such a production, particularly
in sponges that contain large amounts of endobiotic bacteria. In the present project,
three sponges of the genus Dysidea, collected in Fiji, New Caledonia, Solomon Is.,
Vanuatu, Moorea and Mayotte are used as biological models.
Other objectives of this study were directed to isolate any interesting compound that
may or may not appear in literature and characterize it to obtain a fair idea of the
class of compound it belongs to. This data would then further aid in the
chemotaxonomic analysis of the Dysidea sponges.
The aim of the study is:
• To obtain baseline levels of the production of secondary metabolites (HPLC);
• To compare the chemical fingerprints and constituents of the three sponges
Lamellodysidea herbacea, Dysidea arenaria, and Dysidea avara (1H-NMR,
HPLC, LC-MS, TLC);
• To characterize the major compounds of the Dysidea sponges (NMR, MS,
UV);
• And to study the geographical variability (HPLC) at different scales within a
single species, Lamellodysidea herbacea.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
��������(�����������
�� ��(�������������������������������
Most of the sponge specimens (Dysidea sp.) for this study were collected in New
Caledonia, with geographically different samples also from Vanuatu, Solomon
Islands and Fiji Islands, during a scientific expedition between 2007 and 2008, by
researchers of Institut de Recherche pour le Développement (IRD), Noumea, New
Caledonia.
The collected sponges were placed in sea water during transportation from the
collection sites to the laboratory. It was then freeze-dried before it could be used. The
identification of the specimens was primarily carried out on-site, and voucher
specimens were sent to the Queensland Museum (Australia) for confirmation by Dr
John Hooper.
Another set of Lamellodysidea herbacea samples was provided at the Laboratoire de
Chimie, des Biomolecules, et des l’Environnement (LCBE), University of Perpignan,
France. These samples were collected from French Polynesia (Moorea Island) and
the Departmental Collectivity of Mayotte, in the year 2002.
� �/�������������8���
Two extraction techniques were used in this project, as outlined by the research
teams worked with. Furthermore, the first technique (Technique 1) was more suitable
for sponge samples of large quantities, while the second technique (Technique 2) was
more convenient for the small quantity of sponge material available. The two
extraction techniques employed are detailed below. In both the techniques employed,
the organic extract obtained was used for this study.
� �� �'!)��9$'���
For the first set of sponge material (Dysidea sp.), collected by researchers of IRD,
Noumea, New Caledonia, approximately 20 g of the freeze-dried sponge material
was weighed out and left to mechanically stir overnight in 100 mL of a mixture of
ethanol and water (80:20). After the initial extraction, the mixture was filtered
(Whatman 103 – 9 cm) using vacuum and the residue was again left to stir in 100 mL
ethanol for 45 minutes. This was then filtered again and the step was repeated a final

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
time. By this step all the ethanol-soluble compounds were expected to have leached
out of the sponge material.
The residue was then freeze-dried and stored separately, while the filtrate was dried
under vacuum using a rotavapor to obtain the organic residue of interest.
The following figure outlines the process of hydro-alcoholic extraction:
Scheme 1 - Hydroalcoholic extraction of freeze-dried sponge material
The residue obtained after drying the filtrate was then dissolved in 100 mL
dichloromethane (DCM), filled in separating funnel 1, and 100 mL of distilled water
was added to it. The content of the separating funnel was mixed carefully and the
layers were let to separate, after which, the organic layer (bottom layer) was drained
out and filled in a clean separating funnel 2. To minimize losses from imperfect layer
separation, 100 mL of water was added to funnel 2 and the two layers were mixed
properly again before obtaining the washed organic layer. Meanwhile, to the first
separating funnel 1, containing the aqueous layer, fresh 100 mL DCM was added,
���������������� ����
����������
�������������������������������������������������������������
������� ����� !��
��������
"#�$��������
��������������%&'��������(�)�
������������������� �����
�����������������������������������*�������� ������������
����+�,���+���
�����������������������������������*�������� ������������
��������
����������
����+�,���+���
����������
�������� ����+�,���+���

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
mixed and organic layer collected in funnel 2. The above steps were repeated twice.
Finally, the aqueous layers from funnels 1 and 2 were combined (ca. 200 mL) and
freeze-dried to remove the water, while the organic layer (ca. 300 mL) was combined
and desiccated by passing it through anhydrous sodium sulphate salt, before filtering
the extract. The filterate obtained thereafter was dried under vacuum using a
rotavapor until a viscous, thick gum texture was reached. It was then transferred to
labeled vials and air dried to completion, after which the vials were stored between 4 oC to –20 oC, until analysed.
The organic extract was labeled as the C extract, while the aqueous extract was
labeled as the B extract. For most of the analyses, the organic extract was used.
These crude extracts obtained were of varying quantities since different sponge
material enabled extraction at varying degrees. Generally, the L. herbacea extracts
were obtained between 3 – 6 % of the freeze-dried sponge material; while the D.
arenaria extracts were obtained between 0.8 – 1.5 %. See Appendix 1 for a list of the
yields and descriptions of the extracts.
The following scheme gives a general outline of the process of liquid-liquid
partitioning:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Scheme 2 - Liquid-liquid partitioning of the Hydroalcoholic extract
�-".���
�-".���
�-".���
������� � �������� � !��"� ��# $ ��#��%���#�� ��
&�����������������' (�)�
��/��+*��/��+�$���������
)� �������������
0��������%&'�0��������(�)�
&�����������������' (���
��/��+*��/��+�$���������
)� ������������1�
0��������(�)�
&�����������������' (�)�2��!��/��������������
��/��+*��/��+�$���������
)� �������������
0��������%&'�
&�����������������' (���2��!��/�������������1�
��/��+*��/��+�$���������
)� ������������1�3��1�
0��������%&'�
&�����������������' (�)�2��!��/��������������� &�����������������' (���
2��!��/�������������1�
��/��+*��/��+�$��������� ��/��+*��/��+�$���������
)� �������������4/��������������
)� ������������15��1�3��1�
4/�������������1�
���6��+���� �
�%���#��*�
%���� ����7���)�� �������������$�$���
"#�$��������
�%���#��+�

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
� � �'!)��9$'� �
The second procedure used was for the Lamellodysidea herbacea samples from
Moorea Island and Mayotte due to the small quantities of freeze-dried sponge
material available.
Two grams of the freeze-dried sponge powder was weighed out and extracted using
DCM/Methanol (1:1) three times with 20 mL of solvent. Each time, the solvent was
added to the sponge material, ultrasonicated for ten minutes, then filtered on cotton
and dried using a rotary evaporator (rotavapor). Before the last drying however, one
gram of reverse-phase (C18) silica was added to the extract to adsorb the sample on
it.
After adsorption on the C18 silica and drying of the solvent, solid phase extraction
(SPE) was carried out on SPE Phenomenex Strata C18 cartridge (vol. = 18 mL; 2 g
C18 silica). The cartridge was first washed and activated using 10 mL of
DCM/Methanol (1:2), followed by conditioning it using 10 mL of Methanol (MeOH)
and then 10 mL of water.
The adsorbed C18 silica was then loaded on the column and washed with 10 mL of
MilliQ water to eliminated salt from the sample. The cartridge was left under
vacuum for 20 to 30 minutes to remove water from the column. Once dry, elution of
the extract was carried out using 9 mL of DCM/Methanol (1:1) under vacuum. The
eluted extracts were collected in 10 mL volumetric flasks and made to the 10 mL
volume with DCM/Methanol (1:1). This was labeled “Extract A”.
From Extract A, 1 mL of the sample was filtered for HPLC analysis (Extract A1) and
another 1 mL for LC-MS analysis (Extract A2). The remaining 8 mL of Extract A,
labeled as Extract A3 was then dried, weighed and stored for later use at -20 oC.
The following scheme briefly outlines this procedure:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
���+��+���8�+�&�������������������
�����'����9�:��������:��!���������������
���$���
%�����+���#�����������$$��;<���������������#��:�����
����%&'='�)(���>�����+���#�����?�
��������+���������#��������������@�
'�@���$�������@�:��!�%&'='�)(���>���A�";������4�
";������4��%���+5�:�� !�+�3�
�����+����*����&�����-�&�3�B���������
";������4�� ������+�����(.�&�
";������4�� ������+������&*'��
� � ���6�*+���+�
����������������3�+��������#�$�������������!�
4++�� �,.*&���������8�����������+���� � �����'�)(?�
�����2�����
�����%&'='�)(�
�."�.!�������;��������&��&�����+ ����;������%&'='�)(���>���:��!�
�������C������������������!�
���� ��������
"�������
&��+������� �
2��!�� �
Scheme 3 - Cold-extraction procedure for small quantities of sample

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
�, ��(�������������
�,�� �)������'��)��3���#��&)��:��;�
The initial TLC of the crude extracts was carried out on normal phase silica gel
(F254) plates with elution using 70 % heptane and 30 % ethyl acetate. Changes were
made to the eluent to 1:1 ratio for fractions obtained during the course of the study in
order to obtain better separation of the products.
The visualization reagents used included ultraviolet Gibb’s reagent for phenols;
vanillin phosphoric acid for steroids; sulfuric acid for organic compounds;
hydrochloric acid/ninhydrin solution for amines, amino acids, and amino sugars; and
dragendorff reagent for alkaloids and other nitrogen-containing compounds. The
procedures for preparation, staining and development of the visualization reagents
was followed closely as stated in “Dyeing reagents for thin-layer and paper
chromatography” (Merck, 1978). The compounds with double bond conjugation
were visualized by ultraviolet (UV) light at 254 nm and 366 nm.
However, in the case of hydrochloric acid/ninhydrin solution, the developed plate
was first sprayed with concentrated hydrochloric acid (HCl) and dried in air for 5
minutes after which it was placed between two glass plates and heated for 10 minutes
at 110 oC, the heating was then continued for another 10 minutes after removing the
top glass plate. This allowed the process for any peptide bond cleavage to take place
and to remove the excess HCl from the plate. After the 20 minutes of heating, the
TLC plate was then sprayed with ninhydrin solution and heated again for 10 minutes
at 110 oC, and this fixed any free nitrogen obtained from the peptide bond cleavage.
After the preliminary results obtained using TLC, the crude samples were analysed
using reverse-phase HPLC.
�,� ��#)��'�%��3��!'���9$� �)��3���#��&)��:���;�
The HPLC system used in this study was the Waters 2695 Separations module with
autosampler, coupled with Waters 996 Photodiode array detector, and the Polymer
Laboratories evaporative light scattering (PL-ELS) detector. The software used to
acquire data was the Waters Corporation Empower Pro (2002).

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
All the crude extracts were put through a primary screening to see the elution of the
compounds of interest and for comparison with other samples. The crude extracts
were prepared at a concentration of 10 mg/mL.
The following conditions (Condition 1) were used for the primary screening:
Column: Phenomenex Gemini 5μ C6-Phenyl 110A (250 x 3.0 mm; 5 micron)
Solvent: Acidified Water (H2O) and Acetonitrile (ACN) (0.1% Formic Acid)
Gradient: 5 minutes at 90 % H2O; 90 to 0 % H2O in 30 minutes; 0 % H2O for 10
minutes
Flow Rate: 0.5 mL/min
Detection: Photo-Diode Array (PDA) detector (254 nm and 280 nm);
Evaporative Light Scattering (ELS) detector
After the primary screening, the HPLC conditions were adjusted to obtain better
separation of the peaks. The following conditions (Condition 2) were adapted for
further analysis:
Column: Phenomenex Gemini 5μ C6-Phenyl 110A (150 x 3.0 mm; 5 micron)
Solvent: Water (H2O) and Acetonitrile (ACN) - pH adjusted to 2.2 with Formic
Acid
Gradient: 50 to 0 % H2O in 15 minutes; hold at 0 % H2O for 15 minutes
Flow Rate: 0.4 mL/min
Detection: PDA detector (240 nm and 280 nm); ELS detector
�,�, ��9$� �)��3���#��&)��-�(�����&'!����!�&��:�<(�;�
Mass analysis was carried out at the Laboratoire de Pharmacochimie des Substances
Naturelles et Pharmacophores Redox, UMR-152-IRD-Université Paul Sabatier,
Toulouse (France).
The liquid chromatography – mass spectroscopy (LC-MS) system used was the
Finnigan LCQ DECA XP Max with positive and negative electrospray (ES) and
atmospheric pressure chemical (APC) ionization techniques, coupled to an ion trap
analyser, with dual pump system.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
For this study, the ionization technique employed was the atmospheric pressure
chemical ionization (APCI), and data was obtained using both positive and negative
modes of analysis. The APCI technique was used because initial analysis with
electrospray ionization (ESI) did not provide appropriate mass profile. As for the
liquid chromatography part, the same column and conditions were used for the
elution as that for the HPLC condition 2. However, instead of the ELS, the column
was connected in series with PDA detector and the Finnigan APCI mass analyser.
�,�. �$!*'���(�#�'��!��'�����!'�:�(�;��&'!����!�&��
1-D and 2-D NMR analyses were carried out at the Laboratoire de Chimie, des
Biomolécules et de l’Environnement (University of Perpignan, France) and
Laboratoire de Pharmacochimie des Substances Naturelles et Pharmacophores Redox
(UMR-152-IRD-Université Paul Sabatier, Toulouse, France).
The NMR system used in Perpignan was the 400 MHz Delta NMR System, while the
system used in Toulouse was the 300 MHz Bruker NMR spectrometer system.
Preliminary proton NMR spectra of the crude extracts were obtained using the 300
MHz system and the isolated compounds were analysed for 1-D and 2-D NMR with
the 400 MHz system.
Based on the solubility of the samples, suitable NMR solvent was chosen for the
analyses. Generally, for most of the samples analyzed in the 300 MHz spectrometer,
deuterated chloroform (CDCl3) was used, but in some cases, deuterated methanol
(MeOD) or deuterated dimethyl sufoxide (DMSO-d6) was required. The analyses of
the isolated compounds on the 400 MHz system were carried out with deuterated
acetone (CD3COCD3 or acetone-d6).

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
�. ��������������������������(��������
�.�� ���� '���&'!�'���,�.��
Fractionation Set 1:
Normal phase flash chromatography of 220 mg of Dysidea species R3141C was
carried out using the Ana-Logix Bottomless Solvent Reservoir (solvent pump 0962-
1). The column used was the SuperFlash SF10-4g Sepra Si-50 (1369600) – 5μm,
60Å, 9.4 x 1.5 cm. It had a mass of packing of 4 grams and a column volume of 4.8
mL, and the flow used during fractionation was 12 mL/min.
A solid deposition of the sample was made by adsorbing the extract onto minimum
amount of normal phase silica gel. The sample was eluted using 100 % heptane,
followed by 90/10 – heptane/ethyl acetate, 50/50 – heptane/ethyl acetate, 100 %
ethyl acetate (EtOAc), 50/50 – EtOAc/methanol, and 100 % methanol (MeOH). The
final washing was carried out using an 84/16 mixture of dichloromethane/MeOH. A
total of 8 fractions were collected in pre-weighed round bottom flasks (RBF), and
dried under vacuum rotavapor at 35 oC.
TLC analysis on the fractions was done using normal phase (NP) silica gel TLC
plates backed with aluminium. Elution was carried out using 70/30 – heptane/EtOAc,
and visualized with 50% sulfuric acid. HPLC analysis was also completed with the
Condition 2 used in the chemical screening.
Following the preliminary analyses, the fraction eluted with 50/50 – heptane/EtOAc
(R3141C-fr2b) was chosen for further fractionation.
Fractionation Set 2:
80 mg of R3141C-fr2b was further fractionated using the same method and column
as in fractionation set 1. However, the solvent composition this time started with
75/25 – heptane/EtOAc, and gradually increased the percentage of EtOAc to 40 %,
50 %, 60 %, 75 %, and 100 %. The final washing was done using 100 % MeOH.
A total of 31 fractions were collected which were reduced to 13 after comparison
using TLC. The 13 fractions were then analysed using HPLC (Condition 2).
Selected fractions from the two sets were sent for proton NMR and LC-MS analysis
in Toulouse. Following data analyses, four fractions (fr. 8, 9, 10, and 11) from the

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
second set of fractionation were selected for compound purification using reverse-
phase semi-preparative HPLC.
Purification of Compound:
The HPLC system used for semi-preparative purification of the sample R3141C-2-
fr8-11 was the Waters 1525 Binary HPLC pump with manual injector, coupled with
Waters 2487 dual wavelength (�) absorbance detector. The software used to acquire
data was the Waters Corporation Breeze version 3.20 (2000).
The following conditions were used for the fractionation:
Column: Phenomenex Gemini C6-Phenyl 5μ
Dimensions: 250 x 10 mm
Flow Rate: 5 mL/min
Solvents: Water and Acetonitrile
Gradient: 40 % water for 30 minutes; 40 – 0 % water in 2 minutes; Hold at 0 %
water for 10 minutes.
The combined fractions (R3141C-2b-fr. 8, 9, 10, and 11) was dissolved in minimum
amount of MeOH and injected. After elution, a total of 7 fractions was collected and
analysed using analytical HPLC (Condition 2) and TLC.
The appropriate products were selected and sent to the University of Naples in Italy
for further analysis.
�.� ��3'**� ��� '��)'�4�!'��
Flash chromatography fractionation similar to the fractionation set 1 of sample
R3141C was carried out with selected L. herbacea samples. Elution solvent was
adjusted according to the samples using heptane and EtOAc. Fractions were air-dried
overnight under ventilator. Analyses were carried out using TLC and HPLC
(Condition 2).
Recrystallization:
For separating the crystals from the mixture, small amounts of hot acetonitrile (ACN)
was added to dissolve the fraction, and let to cool in an ice bath with the vial

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
covered. Over time, the crystals formed and settled to the bottom, after which the
supernatant was decanted and the procedure was repeated.
�0 ����<���������������
�0�� (' ���:��4�$��$ ��'=����'��#���-����;���'&��������
The fungal strains were cultured on Sabouraud dextrose agar prepared at a
concentration of 45 g/L. The medium was prepared as follows:
• Medium powder was suspended in de-ionized water, mixed carefully, and
brought to boil with constant mixing, until the solution turned translucent.
• It was then dispensed into bottles and slants as required while it was still hot
and in liquid state; and autoclaved for 15 minutes at 118 oC.
• After taking the medium out of the autoclave, it was left at room temperature
for 15 seconds and then transferred into a thermostatically controlled water
bath set at approximately 45 – 50 oC. This condition was maintained until the
medium was used.
• For use at a later date, the medium was refrigerated and turned into liquid
state in a thermostatically controlled water bath just before use.
• The tubes containing the media were left to cool in a slanted position.
The sterile petri-dishes to be used were first passed under ultra-violet (UV) light for
15 minutes to ensure that there was no contamination, and then the medium was
poured into the dishes. It was left to cool and settle for about 1 hour, after which the
medium was again passed under UV light for 15 minutes. The petri-dishes were then
closed, inverted, and stored in the refrigerator until used.
�0� ����<%$�#�#��3����!��
Anti-fungigram discs saturated with the sponge extracts were prepared for the
analysis. Each disc had the capacity to hold 10 μL of the sample at a time and the
dosage to be tested was 1 mg of sample per disc. Hence, the samples were prepared
at a concentration of 10 mg/mL, and each sample was loaded 10 times on the disc,
giving enough time for the solvent to evaporate in between loads. The fractionated

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
and purified compounds were loaded at a lower dose (0.1 mg per disc) compared to
the crude extracts. All the tests were done in duplicates.
Anti-fungigram discs of the solvents used for the sample preparation were also
prepared, to act as control for the assay. Furthermore, the antifungal agent Nystatin
was also tested with the samples for reference purposes.
Anti-fungigram discs were prepared in sterile conditions to avoid contamination.
�0�, �$�#�*����!$*������
The culture plates were marked at the bottom with evenly spaced spots each
containing reference numbers to be used for identifying the sample disc used on a
particular plate. Each plate contained seven spots; hence it could be used to test
seven extracts at a time. After marking the plates appropriately, the culture
suspension was prepared.
It was important that all the steps undertaken during the bioassay was done in a
sterile condition so that chances of contamination could be eliminated. The
inoculation loop was first heated until red-hot and introduced into the culture slant
touching at the medium void of fungal growth. This way, the excess heat that could
kill the microorganism, was transferred onto the medium and the loop was left cool
enough to pick out the culture unharmed. A loop full of the fungi from the culture
slant was taken and suspended in test-tube containing sterilized physiological water
(1 % sodium chloride (NaCl) solution). This was then vortexed to give an even
suspension.
The culture plates were inoculated using the spread plate technique. A sterilized
Pasteur pipette was used to transfer approximately 1 mL of the suspension onto the
medium and the plate was turned around gently to evenly spread the suspension. The
excess suspension was drained out using the pastuer pipette and discarded into a
beaker of disinfectant.
The prepared anti-fungigram discs were then placed at the allocated positions on the
culture plates. Fungal cultures incorporated with the anti-fungigram discs were then
incubated for 48 hours, with monitoring at 24 hours.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
, �������+����������
,�� ��(�������������
Chemical analysis is the first approach to identify chemicals/compounds of interest
in any branch of chemistry. It involves a wide range of techniques including use of
chemical identifiers and reactions, and instrumental analysis. It also resolves issues
such as identification, isolation, structure determination, and quantification of
molecules. Chemical screening can be used to identify known compounds and isolate
new ones.
A total of 14 Lamellodysidea herbacea, 9 D. arenaria, and 1 D. avara from New
Caledonia and one L. herbacea from Moorea Island (for comparison) were screened.
Full details of the samples studied are provided in Appendix 1. The techniques used
for chemical screening included thin layer chromatography (TLC), high performance
liquid chromatography (HPLC), 1-D nuclear magnetic resonance (NMR)
spectroscopy, and liquid chromatography - mass spectroscopy (LC-MS).
The aim of this screening was to acquire the chemical fingerprints (TLC and HPLC
chromatograms) of the sponge samples; to set up the chemical identity cards of the
different samples by identifying the compounds that are common within a particular
species; and to see whether the same compound(s) can be found in other species of
the Genus Dysidea. This could indirectly suggest the species that are more closely
related to each other and those that are far away in the phylogeny in terms of
chemical constituents of the sponges.
,���� ��3'**� ��� '��)'�4�!'��
Lamellodysidea herbacea has been one of the most studied species of the genus
Dysidea. It has been found in various locations around the world. The chemical
composition of this species was investigated to identify compounds that can be used
as chemical markers for chemotaxonomy of Dysidea sponges.
3.1.1.1 Thin Layer Chromatography
Thin layer chromatography (TLC) is the primary step in identifying the main groups
of compounds present in the sample of interest, and to obtain a general profile in
order to view the similarities and differences of the chemical composition of the

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
samples. This is achieved by use of different visualization reagents which are
specific for particular functional groups, such as phenols, amines, double bond
conjugations, etc.
The standard compounds used as reference spots for TLC were cholesterol and a
tetrabromo-diphenyl-ether (4Br-OH-OMe-PBDE). The 4Br-OH-OMe-PBDE (3,5-
dibromo-2-(3’,5’-dibromo-2’-methoxyphenoxy)phenol) was chosen as standard
compound for the analysis due to its availability, and also because it was isolated
from L. herbacea collected previously from one of our sampling sites (Moorea
Island).
O
OHO
Br
Br
Br Br
3,5-dibromo-2-(3',5'-dibromo-2'-methoxyphenoxy)phenol(4Br-OH-OMe-PBDE)
HO Cholesterol
12
34
561'
2'3'
4'5'
6'
Figure 9 - Compounds used as standards for TLC analysis
L. herbacea indicated the presence of phenolic compounds due to positive
visualization with Gibb’s reagent (seen as a blue or purple spot, between Rf values of
0.27 and 0.57).
Furthermore, within the L. herbacea extracts from New Caledonia, two out of 14
samples showed differences in TLC profiles and indicated presence of the standard
compound (4Br-OH-OMe-PBDE) through direct correlation (blue spot with Gibb’s
reagent at Rf = 0.55). This visualization was interesting because the other L.
herbacea specimen collected in the same location (South of New Caledonia) did not
contain this compound and they all had similar TLC profiles. The L. herbacea
specimen from Moorea Island contained the 4Br-OH-OMe-PBDE as expected. The
figure below illustrates this point:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Plate 1 - Differences in phenolic composition within L. herbacea from New Caledonia
(TLC eluent: 7/3 - Heptane/EtOAc; Reagent: Gibb's Reagent)
With vanillin phosphoric acid (VPA), all the L. herbacea from New Caledonia had
the same profile, and difference was noted only with the Moorea sample.
Dragendorff reagent and Hydrochloric acid/Ninhydrin solution failed to give
comparable profiles of the samples. It was evident from the TLC analysis of the
samples that two New Caledonian samples (R2259C and R2279C) contained the
standard compound due to the elution of the compound at Rf = 0.55. For
confirmation, the HPLC, NMR and mass profiles of the extracts were obtained.
3.1.1.2 High Performance Liquid Chromatography (HPLC)
The HPLC system was coupled with the photo-diode array (PDA) detector and the
evaporative light scattering (ELS) detector. The PDA detector can scan a wide range
of light absorption from 200 to 800 nm, hence making it suitable for detecting
compounds with chromophores and provide characteristic UV spectra. On the other
hand, the ELS detector is not specific; therefore it can be used to detect all types of
compounds including compounds lacking chromophores. Thus, coupling the HPLC
system with the PDA and ELS detectors enabled efficient total analysis of the
extracts.
The HPLC profiles of the Lamellodysidea herbacea samples were quite simple with
generally two or three main peaks detected by the PDA and ELS detectors. Added to
this, the retention time and UV absorption of the standard 4Br-OH-OMe-PBDE was
comparable to the samples suspected of having this compound. As seen in the TLC

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
profiles, sample R2259C (L. herbacea from New Caledonia) had one extra peak for
4Br-OH-OMe-PBDE in its profile compared to the other samples of the same set,
while R2279C had only one peak corresponding to the standard PBDE.
Figure 10 - Three types of chemically different L. herbacea found in New Caledonia
(Above: Chemical difference seen in TLC visualization using Gibb's reagent. Below:
Chemical difference seen in RP-HPLC and the mass and UV absorption obtained for each
peak [HPLC Condition 2])
Each peak gave a distinctive UV absorption which could be used to identify the
compounds of interest. The standard 4Br-OH-OMe-PBDE absorbed UV light at
232.2 nm and 286.7 nm, and this was also seen in the peaks of compound 1 present
in L. herbacea samples R2259C and R2279C. Thus, LC-MS and NMR analyses were
carried out to determine the mass and orientation of the chemical constituents present

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
in the analytes. The peaks corresponding to compounds 2 and 3 had slightly different
UV spectra with a 2nd absorption maximum at 291.5 nm.
3.1.1.3 Liquid Chromatography – Mass Spectroscopy (LC-MS)
Apart from the mass of a compound, mass analysis can give a fair idea of the type of
halogenation present, if any. The two main types of halogenation patterns seen
generally in L. herbacea are brominated and chlorinated isotopes. These two types of
atoms have a characteristic pattern of isotopic peaks. The chlorine atom has an
abundance ratio of 35Cl : 37Cl = 3:1, while the bromine atom has a ratio of 79Br : 81Br
= 1:1. This can be used as an easy way of identifying the presence of these halogens.
Furthermore, the pattern of the peak obtained can also indicate the number of
halogen atoms in the particular compound. (See Appendix 2 for the guide to identify
the pattern of halogenation and the number of halogen atoms in a compound.)
The LC-MS analysis was carried out using both negative ion (NI) and positive ion
(PI) mode APCI, and it was observed that the NI mode of ionization was more
informative than the PI mode. Therefore, based on the NI-APCI spectra of the
Dysidea samples, L. herbacea exhibited presence of Bromine isotopes corresponding
to the main peaks in the PDA scan. The pattern of spectra obtained for each peak
indicated the presence of 4, 5, or 6 bromo- compound.
Figure 11 - Pattern of halogenated cluster showing presence of 4, 5, and 6 bromine
atoms in the L. herbacea studied [HPLC Condition 2 + Finnigan APCI Mass analyser].

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
This confirmed that the phenolic compounds indicated by the Gibb’s reagent in TLC
analysis of the L. herbacea samples were actually brominated phenols, and apart
from the standard 4Br-OH-OMe-PBDE, the samples also contain other types of
polybrominated diphenyl ethers (PBDE).
However, one complication faced with these results was that for each peak in the
PDA scan, there were two corresponding signals observed in the mass spectra.
Usually, the difference in mass between the two signals matched with the mass of
one bromine atom, and in few cases, it corresponded to a mass of 45 mass units.
Hence, either there were some rearrangements or fragmentations taking place in the
system during mass analysis; or there were two compounds present in the same peak
which could not be separated properly.
The second hypothesis does not quite hold true since this pattern was seen in all the
L. herbacea samples analysed. Furthermore, for two compounds having different
mass and structure, their retention times would differ to some extent, and they cannot
possibly be present in the same peak and retention time.
Therefore, the first hypothesis of rearrangements or fragmentations during the course
of the analysis seemed possible. This pattern has been previously observed in a study
by De Rijke, et al. (2003), where a comparative study was done on flavonoids using
LC-APCI-MS and LC-ESI-MS to determine their analytical performances. In this
study, it was seen that the APCI method, generally produced less fragments
compared to ESI method, thus making it suitable for crude extract analysis. Added to
this observation, it was noticed that with the use of formic acid in the eluent, there
tends to be a formic acid adduct [M+45]- forming with the negative ion mode of
ionization. The adduct formation is highly dependent on the acidity of the compound,
and so this pattern is not seen with all the polybrominated diphenyl ethers (PBDE) in
this analysis. According to their acidity, the formation of the formate ion with neutral
PBDE (monohydroxylated PBDE) successfully competes with proton abstraction
from the weakly acidic PBDE (dihydroxylated PBDE) to form an [M-H]- anion. This
would explain why a mass gain of 45 mass units was observed in some of the
Lamellodysidea herbacea samples only.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Additionally, in another study by De la Fuente, et al. (2003), it was seen that some
PBDEs did lose a bromine atom to form [M-Br]- ion in mass analysis using NI-
APCI. Therefore, this also supports our first hypothesis.
Figure 12 – Pattern of rearrangement of compound observed in mass analysis [HPLC
Condition 2 + Finnigan APCI Mass analyser].
Thus, the L. herbacea samples studied generally contain penta- and hexa-PBDE,
with the exception of a few samples (2 New Caledonian L. herbacea [R2259C and
R2279C], and Moorea samples).
3.1.1.4 Nuclear Magnetic Resonance Spectroscopy (NMR)
NMR analysis of all the samples aided in confirming the observations made with the
other chemical analyses (TLC, HPLC and LC-MS). All the crude extracts of the
Dysidea samples were organic extracts; hence analysis was carried out in deuterated
chloroform (CDCl3). Isolated compounds were studied using acetone-d6.
From the proton NMR profiles of the extracts, it was seen that the L. herbacea
samples had interesting chemical shifts around 7 - 8 ppm. This shift corresponds to
the aromatic protons, and the profile matched with the PBDE possibility.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
The final conclusion was made using the proton NMR data (and in some cases, with
the 13CNMR and 2-D NMR spectra), whereby compounds were assigned to the
different chemical shifts to arrive to a specific structure.
,�����.�� �3&�$� ���
Since compound 1 (ref. Figure 10) gave the major peak in the HPLC profile of
R2259C - type 1 - (and the major spot in the TLC profile), it should show major
peaks in the proton NMR profile. This was confirmed due to the presence of 4 major
chemical shifts in the aromatic region of the profile, which corresponds well to the
presence of 4 protons in the structure with 4 bromine atoms, one hydroxy group and
one methoxy group. The chemical shifts of the hydroxy and methoxy groups were
observed at 5.40 ppm and 4.04 ppm, respectively.
Figure 13 - Compound 1: Mass spectrum [HPLC Condition 2 + Finnigan APCI Mass
analyser], 1HNMR [300 MHz; in CDCl3], and structure
The four protons gave doublets with coupling constant close to 2 Hz, which indicates
that the protons are meta-coupled.
Furthermore, for comparison purposes, the NMR profile of R2259C was compared to
the profile of another L. herbacea from Moorea (MAU-02-Dh1) and New Caledonia

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
(R2279C), which contained only the standard compound, hence having clearer NMR
profiles. It was seen that the three profiles matched well in terms of chemical shift
pattern.
The figure below illustrates the NMR profiles of R2259C and the L. herbacea from
Moorea Island (Full NMR profiles of R2259C, R2279C and L. herbacea from
Moorea are given in Appendix 3a):
Figure 14 - 1HNMR profiles of organic extracts of R2259C and MAU-02-Dh1 (L.
herbacea from New Caledonia and Moorean Is., respectively), in CDCl3, 300 MHz
spectrometer.
The following table (Table 2) shows the comparison between the chemical shifts
obtained in this analysis for R2259C, R2279C, and the L. herbacea from Moorea
Island, with literature values:
Table 2 - 1HNMR chemical shifts obtained for Compound 1 in L. herbacea from
New Caledonia (R2259C and R2279C) and Moorea Is. (in CDCl3, 300 MHz)
Proton � R2259C
(NC) (ppm)
J for R2259C
(Hz)
� R2279C
(NC) (ppm)
J for R2279C
(Hz)
� MAU-02-Dh1
(Moorea) (ppm)
J for MAU-02-Dh1
(Hz)
� Literature
(ppm)*
H4 7.35 1.2 7.36 2.0 7.38 2.2 7.33
H4’ 7.45 1.9 7.46 2.2 7.48 2.2 7.40
H6 7.19 - 7.20 2.0 7.21 2.3 7.09
H6’ 6.78 2.1 6.80 2.2 6.83 2.2 6.54
-OH 5.40 5.08 5.34
-OMe 4.04 3.81 4.06 3.78
* (Fu, Schmitz, Govindan, & Abbas, 1995)

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
A clear correlation could be seen for the chemical shifts obtained for the three
sponge specimens and the reference values available for the compound. Therefore, it
was finalized that Compound 1 in the New Caledonian sponges R2259C and
R2279C is 4Br-OH-OMe-PBDE, and it has the following structure and details:
O
OHO
Br
Br
Br Br
3,5-dibromo-2-(3',5'-dibromo-2'-methoxyphenoxy)phenol (4Br-OH-OMe-PBDE)
Mol. Wt. = 531.82 mu
M/z (APCI-NI mode) = 531.07 mu
TLC Rf = 0.55 - in 7:3 - heptane:EtOAc, Gibb's reagent
HPLC Rt = 31.429 min & 10.804 min (for HPLC condition 1 & 2, respectively)1HNMR shifts (CDCl3, 300 MHz): 7.36 ppm (H4, d, 2.01 Hz); 7.46 ppm (H4', d, 2.19 Hz); 7.20
ppm (H6, d, 2.04 Hz); 6.80 ppm (H6', d, 2.16 Hz); 3.81 ppm (-OMe); 5.08 ppm (-OH)
12
34
56
2'1'
6'5'
4'
3'
Figure 15 - Compound 1 (3,5-dibromo-2-(3’,5’-dibromo-2’-methoxyphenoxy)phenol)
,�����.� �3&�$� � �
Compound 2 was seen to be the most abundant product in all the type 2 L. herbacea
from New Caledonia, and it was the second most abundant compound in the type 3
sample as seen in the HPLC profile in Figure 10. The TLC profile could only be
used as an estimation of the location of the product since it was almost fused with
Compound 3. Therefore, only HPLC, mass spectra and NMR could be used for the
structure determination.
In the HPLC profile, compound 2 eluted at a retention time (Rt) of 32.631 and 10.519
minutes for HPLC conditions 1 and 2, respectively. As for the mass analysis, the
mass data obtained with NI-APCI showed two masses, 593.28 and 674.62 mu, with a
difference of 81.34 mu. This correlates to the loss of a bromine atom, in which case
the mass of 674.62 mass units is the pseudomolecular ion [M-H]- of the product, and
593.28 mu corresponds to [M-Br]- ion, which would have formed due to the
conditions involved in mass analysis.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Furthermore, the mass spectra of compound 2 suggested the presence of 6 bromine
atoms. In the literature cited, only one PBDE could be found which had a molecular
weight of 675.58 mu, and 6 bromine atoms.
However, since the 1HNMR profile of the crude samples did not come out too well,
the L. herbacea sample R2252C was fractionated to see if a better profile could be
obtained. (Details of the fractionation and product isolation are provided in Section
3.3.2.1.)
Following fractionation, the new 1HNMR profile (at 300 MHz in CDCl3) contained 4
proton signals: two were doublets with coupling constants of 3 Hz, while the two
singlets at 6.01 and 6.04 ppm would be due to the hydroxy groups present on the
compound. This has been confirmed by the disappearance of the two peaks when
deuterated water (D2O) was added.
The molecule was also analysed for 13C NMR (at 100 MHz in acetone-d6) and 2-D
NMR (at 400 MHz in acetone-d6).
12 carbon signals and 2 proton signals were visible in the 13C NMR, 1H NMR and
the 2D NMR (HSQC and HMBC). Table 2 indicates the chemical shifts obtained
with the NMR analyses of compound 2.
Table 3 - 1H and 13C NMR shifts obtained for Compound 2
C #
300 MHz (CDCl3)
400 MHz (Acetone-d6)
�H Lit. in CDCl3 (ppm)* 1H (ppm)
13C (ppm)
1H (ppm)
HMBC
1 150.49 2 140.62 3 119.31 4 121.60 5 127.22 6 116.48 1’ 146.89 2’ 145.34 3’ 112.19 4’ 7.45 130.40 7.41 C2’; C3’; C5’; C6’ 7.21 5’ 111.74 6’ 6.68 117.12 6.85 C2’; C4’; C5’ 6.36
-OH 6.01 -OH 6.04
* (Norton, Croft, & Wells, 1981)

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
The positioning of the protons can be justified:
• First, based on the C and H chemical shifts according to the substitution on
the aromatic ring. The low chemical shifts of the C bearing bromines allow us
to conclude that they are on positions 5’ and 3’. Furthermore, the hypothesis
of C bearing bromines on positions 4’ and 6’ is not consistent with the
chemical shifts of the C bearing H according to literature.
• Second, based on the long range H-C coupling data obtained. On the HMBC
spectrum, H6’ proton is coupled with C2’, C4’ and C5’, and the H4’ proton is
coupled with C2’, C3’, C5’ and C6’. The HSQC spectrum allows to correlate
H4’ (7.41 ppm) to the C appearing at 130.4 ppm, and the H6’ (6.85 ppm) to
the C at 117.1 ppm.
This arrangement is obtained for compound 2 illustrated in Figure 16.
Figure 16 illustrates the mass spectra, 1HNMR profile and the structure obtained for
Compound 2 (Full NMR spectra [1HNMR, 13CNMR, HSQC and HMBC] of
compound 2 is attached in Appendix 3b):
Figure 16 - Compound 2 (3,4,5,6-tetrabromo-2-(3’,5’-dibromo-2’-
hydroxyphenoxy)phenol): Mass spectrum [HPLC Condition 2 + Finnigan APCI Mass
analyser], 1HNMR [300 MHz; in CDCl3], and structure

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Hence, with the improved NMR profiles, compound 2 was assigned the structure of
the 3,4,5,6-tetrabromo-2-(3’,5’-dibromo-2’-hydroxyphenoxy)phenol found in
literature (Norton, Croft, & Wells, 1981).
,�����.�, �3&�$� �,�
Compound 3 was the second abundant compound found in type 2 samples. It was
isolated together with compound 2 but could not be purified properly; hence it still
contained traces of compound 2. Thus, the HPLC and NMR profiles of compound 3
contained a mixture of compounds 2 and 3.
From the NI-APCI mass spectra, compound 3 had a pseudomolecular ion mass [M-
H]- of 594.77 mu, and five bromine atoms present in the product.
In the proton NMR profile of the product, there were only three signals observed:
two meta-coupled doublets, and one singlet. Eight possible structures exist for a 5Br-
PBDE with a mass of around 595 mu, and two meta-coupled protons.
These eight possibilities were isomers of each other and three have been isolated
previously in independent research by Hanif, et al. (2007) and Fu, Schmitz,
Govindan, & Abbas (1995) from Dysidea sponges.
Proton and carbon chemical shift data of compound 3 were very similar to compound
2 except for the carbon bearing the 3rd proton at 128.54 ppm and the corresponding
singlet at 7.61 ppm. After correlation of the chemical shifts and comparing them with
the literature, compound 3 was assigned to be 3,5,6-tribromo-2-(3’,5’-dibromo-2’-
hydroxyphenoxy)phenol. This compound has been previously found in literature
(Norton, Croft, & Wells, 1981).
The chemical shifts obtained for compound 3 are provided in the following table:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Table 4 - 1H and 13C NMR shifts obtained for Compound 3
C # 300 MHz (CDCl3) 400 MHz (Acetone-d6) �H Lit. in CDCl3 (ppm)* 1H (ppm) 13C (ppm) 1H (ppm)
1 151.54 2 139.88 3 117.54 4 7.61 128.54 7.64 7.51 5 124.01 6 115.47 1’ 147.12 2’ 145.28 3’ 112.15 4’ 7.47 130.26 7.41 7.41 5’ 111.70 6’ 6.68 117.02 6.74 6.50
*Norton, Croft, & Wells, 1981
The following figure illustrates the mass spectra, NMR profile and structure obtained
for compound 3 (Full NMR profile of compound is provided in Appendix 3c):
Figure 17 - Compound 3 (3,5,6-tribromo-2-(3’,5’-dibromo-2’-hydroxyphenoxy)phenol):
Mass spectrum [HPLC Condition 2 + Finnigan APCI Mass analyser], 1HNMR [300
MHz; in CDCl3], & Structure

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Overall, the three compounds identified in L. herbacea were closely related diphenyl
ethers with varying numbers of bromine, hydroxyl, and methoxy substituents.
Appendix 4 illustrates the 1-D and 2-D NMR shifts and the yields obtained for all the
compounds found in this study.
,��� ���� '����'������
Dysidea arenaria has been noted to produce sterols (Jacob, et al., 2003),
sesquiterpene lactones (Piggott & Karuso, 2005), and sesquiterpene ethers (Horton &
Crews, 1995). In this analysis, however, it was difficult to identify particular
compounds due to the complex analytical profiles obtained.
In the TLC profile of the D. arenaria samples, two to three main groups of
compounds could be identified in the extracts. This was visualized with the help of
vanillin-phosphoric acid reagent and seen as pink or black spots. Furthermore, with
long-wave UV light (366 nm), it was possible to conclude that there were fluorescent
compounds also present in the extract.
Plate 2 - TLC profile of D. arenaria from New Caledonia after visualization using
vanillin phosphoric acid and UV light (360 nm). (Eluent: 7/3 - Heptane/EtOAc)
However, this failed to register clearly in the HPLC profile of these extracts. The
PDA scan gave a lot of peaks hence one peak could not be compared to another, and
in the ELS scan, three clusters of peaks could be observed but there was a lot of
noise registered as well. Overall, the HPLC profiles of all the D. arenaria samples
from New Caledonia were quite similar with differences in their peak intensities. In

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
the mass spectra of these extracts, the masses obtained did not match with the masses
of D. arenaria products reported in literature.
Furthermore, looking at the proton NMR profile of D. arenaria, most of the peaks
were found to have chemical shifts between 0.5 to 3.5 ppm, and there were no peaks
observed in the aromatic region. It can thus be supposed that the main compound(s)
present in the D. arenaria are sterols, aglycones and other hydrocarbons lacking an
aromatic moiety. (See Appendix 5 for PDA + ELS + LC-MS + NMR)
Additional conclusions about the Dysidea arenaria species can not be made without
further study of the samples.
,���, ���� '���6����
Dysidea avara is a rich source of pharmacologically active ingredients such as
avarol, avarone, and their derivative compounds, etc., (Munro & Blunt, 2001; Marin,
et al., 1998; Shen, Lu, Chakraborty, & Kuo, 2003). According to Minale, Riccio, &
Sodano (1974), avarol has been found to be present at about 6 % of the sponge dry
weight, which would in turn make it the most abundant compound present in the
crude extract. However, the different chemical analyses failed to identify this product
in the sample R2082, which has been identified as Dysidea avara.
The TLC profile of the D. avara from New Caledonia had a complex profile similar
to the D. arenaria collected in the same area. This is also true for the HPLC profile
to some extent, because of the presence of similar clusters of peaks observed in the
two sample sets. For mass analysis, the only peaks detected were in the lipidic region
of the chromatogram. This confirmed the proton NMR profile since the main
chemical shifts were observed between 0.5 to 3.0 ppm. (See Appendix 6 for PDA +
ELS + LC-MS + NMR)

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Plate 3 - D. avara (R2082C) compared with reference [Avarol (Left) and D. arenaria
(Right)].
Thus, for D. avara, like the D. arenaria, confirmed conclusions could not be made
due to the complexity of the data obtained, but it is possible that the main compounds
present in this sample are sterols and other hydrocarbons.
When comparing all the species studied (L. herbacea, D. arenaria, and D. avara),
each profile had a distinct characteristic (See Appendix 7). Reverse-phase HPLC
analysis of the crude extracts confirmed the initial conclusions of the TLC results
that the separate species of Dysidea are chemically different also, and that members
of the same species do have similar chemical composition, with a few exceptions. In
comparison to L. herbacea, D. arenaria and D. avara did not exhibit presence of
phenolic or halogenated compounds, and the profiles obtained were not simple also.
A lot of compounds were seen to be UV active and were moderately hydrophobic,
but their quantities were too small to be detected properly.
,���. ��������
Chemical analysis helps to answer quite a lot of questions about the nature of a
compound or a complex mixture of compounds. Chemical identity cards of the
Dysidea sponges have permitted to see that the different species have specific
chemical profiles and that it is possible within a small location (New Caledonia) to
differentiate the sponges at the species level, according to their chemical fingerprints.
Meanwhile, it is not always possible, depending on the complexity of the chemical
fingerprints, to compare and analyze the sponge molecular content. In this study,

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
only the L. herbacea showed simple fingerprints that could be easily analyzed: the
main compounds identified in the L. herbacea samples were polybrominated
diphenyl ethers (PBDE).
The halogenated compounds present in L. herbacea can generally be divided into
two chemotypes: the chloro-chemotype and the bromo-chemotype. This is because,
after observing the several L. herbacea compounds from literature, it was noticed
that when one type of halogenation exists in a sponge metabolite, the other type is
not present, and vice versa. All the New Caledonian L. herbacea samples analyzed in
this study contained polybrominated diphenyl ethers and not chlorinated peptides, so
they belong to the bromo-chemotype of compounds.
The production of the halogenated compounds is suspected to be the result of
symbiotic cyanobacteria living in the sponge tissues, and not the sponge itself. The
difference in the halogenations pattern is correlated to the presence of particular
strains of morphologically similar cyanobacteria, Oscillatoria spongeliae. The
different strains of cyanobacteria have not been found to co-exist on the same
sponge. Therefore, the PBDE’s found in this study can not be used as biomarkers for
sponge chemotaxonomy until it is proven that the compound is actually produced
only in the presence of Lamellodysidea herbacea. More specific compounds will
need to be searched for in order to successfully find a link between the classical
taxonomic technique and chemotaxonomy. Thus, for further study on
chemotaxonomy of this group of sponges, the chemical constituents of both the
sponge and the symbiont will need to be analysed; together and separately. Added to
this, more focus will need to be placed on obtaining sponge-specific compounds
from the sponges, such as sterols, since most are produced by the sponges
themselves.
Overall, the extracts from New Caledonia exhibited three types of profiles: type 1 L.
herbacea contained mainly compound 1 - 4Br-OH-OMe-PBDE (which corresponded
to the standard compound used in the analysis); type 2 was noted to contain two
major compounds, compounds 2 and 3 - 6Br-2OH-PBDE and 5Br-2OH-PBDE,
respectively; while type 3 included the two compounds found in type 2 L. herbacea
as well as the major compound in type 1 sample. Whether these differences in the
sponge molecular content are due to biotic or abiotic factors is an important question
that will be discussed in the next section, including results from a larger geographical

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
location. However, the chemical analysis of different species of the Genus Dysidea
shows that variations in chemical composition of sponges exist which make them
exclusive to their species. This is a very important step towards chemotaxonomic
analyses.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
,� ������������������������������������������
The second aspect of this project was to compare the different ecological niche and
geographical locations the sponges were collected in, and if there were similarities or
differences in the pattern of the constituents present in the samples. Comparisons
were made based on the availability of the samples. The three Dysidea species
focused on were Lamellodysidea herbacea, Dysidea arenaria and Dysidea avara,
and the geographical locations studied were New Caledonia, Vanuatu, Solomon
Islands, Fiji Islands, Moorea Island in French Polynesia, and Mayotte. For
comparison of chemical constituents by geographical location, only Lamellodysidea
herbacea was focused on generally due to its availability, simplicity of composition
for comparison purposes, and due to time constraints.
The following figure outlines the six sampling regions for this study:
�
Figure 18 - Collection sites of L. herbacea for geographical comparison (Source: Google
Maps)
,� �� ���������������������������
Sponges of the genus Dysidea can be found in many colors, shapes and sizes. They
can either be encrusting on hard coral or stone surface, or exist as free-forming
colonies in sediment gaining their stability using a hold-fast. They can also be found
at various depths depending on the substrate, food availability and chances of
survival.
The Dysidea sponges in this study were collected at various depths (from 1 meter in
the lagoon to 20 meters near the reef). Being filter-feeders, they were commonly

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
found in clear waters, but some specimens were also found surrounded by sediment.
Full details of the specimens studied are provided in Appendix 8.
3.2.1.1 Lamellodysidea herbacea
Almost all of the L. herbacea collected in New Caledonia were green or pale green
in color with the base encrusting and spreading horizontally on hard coral surface.
Small leaf-like (lamellate) projections were visible coming out from the base, and
were generally vertical. This was reported by Austin, (2003), to be a form of
protection from sedimentation, since less surface area of the sponge is exposed to the
sediments, while the surface area for water intake would be maximum and free of
clogging.
Some specimens were found to be in competition for space with other sessile marine
organisms while others had dominated their habitats. In certain specimens, where
competition was not too much of a problem, vertical growth could be observed;
hence some samples had larger colonies than others.
Out of the L. herbacea collected in New Caledonia, two specimens were slightly
different in general appearance and environment than the others: samples R2259 and
R2279. R2259 was collected in an area which had some sedimentation, while R2279
contained a slightly different pigmentation and was surrounded by quite a few green
and red algae.
Similarly, small differences could be observed in the appearance of L. herbacea
collected in Fiji and Solomon Islands, especially in the pigmentation of these
sponges. Added to this, the Solomon Island sponge was collected in waters that were
slightly darker than other sites, and as for the Fijian L. herbacea, it was found
growing among other sea-weeds and algae.
For the samples from Moorea and Mayotte, specimens were collected in the lagoon
between the depths of 1 to 1.5 meters. They were found in clear waters close to hard
corals, and were not surrounded by algal or sea-weed growths.
3.2.1.2 Dysidea arenaria and Dysidea avara
The D. arenaria specimens collected in New Caledonia were grey in color and had
lobate finger-like projections coming out from the main stem. The colonies were
quite closely clustered, and they were found growing on sandy substrate, surrounded

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
by sea-weed and algae. Some samples also had sedimentation in the water around
them.
The only D. arenaria obtained from Fiji (R3314), did not quite look like the samples
collected in New Caledonia. This species was pink in color, and had the appearance
of a perforated ball instead of digitate projections from the main stem. Furthermore,
this particular specimen grew out from the middle of a cluster of sea-weed.
For Dysidea avara samples, there was only one specimen each from New Caledonia
(R2082) and Fiji (R3203).
The Fijian sample was red to maroon in color when outside the water, and it had
rigid stems like that of hard corals. The ecological comparison of the two specimens
could not be completed due to lack of data about the ecology of these sponges.
3.2.1.3 Dysidea species (R3033, R3161, and R3141)
Three Dysidea sponges received from the Solomon Islands were unclassified species,
where two of these specimens (R3033 and R3161) were identified to be closer to
each other than the third sponge (R3141). Hence, the two similar species were
classed as Dysidea 4177 while R3141 was labeled Dysidea sp.
Sponge R3033 was purple in color with digitate projections as seen in hard corals.
However, they were not clustered together into one colony. The sponge was actually
scattered on a rocky surface, but still linked to each other. This species was present in
close proximity of algae and sea-ferns.
Sponge R3161 was also purple in color, with digitate projections branching out from
the main stem which was rooted to a rocky surface. This specimen was more colonial
compared to R3033. It was found in an area with less light, and the color of the
sponge out of the water was dark purple.
The last Dysidea sp. (R3141) was found growing on a rocky surface also inhabited
by other algae, sea-weeds and marine invertebrates, and it was dark maroon in color.
It had more of a leaf-like appearance, as if it were a marine plant.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
,� � ��(���������� ���� ����������� ��� ��(���� �(���������
���� ������� ���>���� ���?���� ���������������
������������������������������������������
Geographical distribution of the Dysidea sponges was chosen as a part of this study
due to the fact that the sponges were collected in different geographical locations,
with each specimen providing a distinctive profile. Hence it was interesting to note
the chemical and genetic similarities and differences of these sponges within the
same region, between different regions, and if possible between the South Pacific
Ocean and the Indian Ocean.
Since L. herbacea was the only Dysidea species obtained from all the regions
studied, this analysis was focused only on this particular species.
3.2.2.1 Comparison of Lamellodysidea herbacea collected within New Caledonia
,� � ���� �3&��������%��)'�)'3�!�*����%�*'��
Most of the L. herbacea sponges were obtained from the South of New Caledonia. It
was seen in the chemical screening (Section 3.1) of these samples, that there were
three types of L. herbacea: Type 1 containing mainly the 4Br-OH-OMe-PBDE; Type
2 having 5Br- and 6Br-2OH-PBDE; while the third kind (Type 3) having the two
compounds from type 1 and 4Br-OH-OMe-PBDE.
The following map shows the exact locations where samplings were done and where
the chemically different specimens were collected:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Figure 19 - Map showing collection sites of L. herbacea in New Caledonia. (White: Type
1 sponges; Blue: Type 2 sponges; Yellow: Type 3 sponges)
It can be seen that the Type 3 and Type 1 samples were collected very near to the
mainland, and this could have been a factor in the different chemical patterns seen. A
closer look at the landscape close to the collection sites reveal that these samples
were collected near mountainous terrain. Hence, mineral and sediment wash-off from
the land could have enhanced or suppressed the production of particular compounds
in the two samples. This theory could be feasible since the other collection sites were
either far from land and on the reef, or close to land (not as close as R2259 and
R2279) but near flat terrain (see Figure 20).

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Figure 20 - Map illustrating the terrain near the collection sites of R2259 and R2279
(Source: Google Maps)
The geographical locations of particular sites are therefore also an important factor to
note when sampling in the same region because the slightest difference in conditions
can lead to differences in their chemical constituents (Mather & Bennett, 1992).
These invertebrates are quite sensitive to changing conditions around them, and
constant supplies of minerals and sediment can eventually cause them to change in
abundance and genetic make-up in the long term.
This point was also proven in a study by Maldonado, Giraud, and Carmona (2008),
where survival of asexually produced recruits of the demosponge Scopalina
lophyropoda was observed by exposing them to different levels of sedimentation. It
was seen during this study that the initial rate of mortality of the sponges was due to
sedimentation, but after a certain point, the sponges exhibited reversal of mortality
irrespective of sedimentation and had revived. This pattern was more evident in the
sponges placed in the harbor where the amount of silt was the highest. It was
concluded in this study that an unidentified genetic component was responsible for
the adaptation to sedimentation and thus, in the long term, with constant
sedimentation into their habitat, can eventually alter their genetic composition.
Hence, as perspectives for future work, new sampling of the L. herbacea can be done
in the region of the positions of R2259 and R2279 to see if samples around the

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
vicinity have the same chemical profile or not. Studies can also be done on L.
herbacea exposed to different levels of sedimentation, to see if it is able to survive
the stress or not. If so, then the changes in chemical composition due to stress can
also be studied, thus proving our statement further. This study can be extended by
incorporating changes in genetic composition, if any, due to the exposure to stress.
,� � ��� �3&��������%��)'�)'3�!�*����%�*'��5��)��'�'�(�&&��#�
Selected samples used for obtaining the chemical profiles of the sponges were also
sent to a collaborative laboratory for phylogenetic mapping studies.
The analysis was carried out by sequencing the internal transcribed spacer (ITS)
region, since it is easy to amplify even from small quantities of DNA, and it has a
high degree of variation even between closely related species. The taxon set was
extended with Dysideidae from the collection of the Queensland Museum and
additional sequences from Genbank identified as Dysideidae (Erpenbeck & Van
Soest, 2006; Erpenbeck, Breeuwer, Parra, & Van Soest, 2006). A total of 74 taxa
with 644 characters were used to reconstruct the phylogeny. Character positions
which could not be aligned unambiguously were excluded from the alignment.
Phylogenetic reconstructions were performed with Bayesian Inference and
Maximum Likelihood Methods. The trees were rooted with Carteriospongia
foliascens of the family Thorectidae (Dictyoceratida).
The following taxonomy tree was obtained after analysis:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Figure 21 - Taxonomy tree of Dysidea sp. after gene mapping (Courtesy of Dr. Dirk
Erpenbeck)
It was observed that most of the L. herbacea samples collected from New Caledonia
are genetically very closely related to each other. However, two L. herbacea
specimens from New Caledonia (R2258 and R2279) were seen to be slightly
distantly related to the first set of L. herbacea. Out of these two specimens, the
chemical profile and differences of R2279 correlates well with the phylogenetic tree
obtained of the samples. This however is not true for the samples R2258 and R2259
from New Caledonia. The chemical profiles of these two specimens suggest that
R2279 is more closely related to R2259 rather than R2258. This is quite opposite to
what is observed in the phylogenetic tree.
Since the genetic analysis was carried out using material from sponge tissue
complete with symbionts, phylogenetic study of the symbiotic cyanobacteria present
in these sponge samples is yet to be completed. It will be very interesting to see if the
chemical variations could be related to the genetic variations of the symbiont.
,������������7&�,�������������7&�,��������������7&�,��������������7&�,��������������7&�,�������������7&�,��������������7&�,��������������7&�,���8������������D�������,�������������� �E��
,�������������7&�,�������������7&�,������������ �7&�,����������� �7&�,������������ �7&�
,������������ �7&�,������������ �7&�,������������ � �E��
,�������� �����������������
,����������������������
,�������� �����������������,�������� ����������������
,����������������� �E��

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
However, the hypothesis of the environmental factors influencing the chemical
contents of the sponges is also plausible and needs to be studied.
3.2.2.2 Comparison of Lamellodysidea herbacea in other Geographical
Locations
,� � � �� �3&��������%��)'�)'3�!�*����%�*'��
The total number of locations studied for this comparison was six, and the sites
included New Caledonia (as discussed earlier), Vanuatu, Fiji, Solomon, Moorea and
Mayotte Islands. Appendix 9 illustrates the six sampling sites, and the approximate
distances between each sampling site.
As observed earlier with the ecological comparison of the L. herbacea samples from
all the sites, it was evident that there were minor differences in the samples in terms
of appearance and the area of collection. The extracts were thus put through similar
chemical analyses as seen in Section 3.1, to be able to compare the samples in terms
of their chemical composition.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
The following figure illustrates the comparative HPLC profiles of the geographically
different L. herbacea with their finalized structures:
O
OHOO
OHOH
O
OHOH
O
OHO
O
OHO
O
OHO
O
OHOH
O
OHO
Br
Br
BrBr
Br
Br
Br
Br
Br
Br
Br
Br
Br Br
Br Br
Br
Br
BrBr
Br
Br
Br
Br
Br
Br Br Br
Br
Br
Br
Br
Br
Br
Br
Br
Br
Br
Br
Br
1 2
3 4
5 6
7 8
12
34
56
1'2'3'
4'5'
6'
Figure 22 - Geographical comparison of chemical constituents present in L. herbacea
Following the required chemical analyses, it was clear that the New Caledonian Type
2 L. herbacea had exactly the same chemical profile as that of the Vanuatu sample.
The similarity between these two samples can not be compared directly in terms of
geographical distance, since the two sites are separated by quite a large distance
(approximately 830 km). But, similar phylogenetic profiles of both sponge and
symbiont, or an adaptation to a very similar environment could explain such
chemical similarities. Unfortunately, the ecological environment could not be
compared in this case, since ecological data of the Vanuatu sample was absent.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
On the other hand, looking at the samples from Solomon Islands (R3147), and Fiji
(R3241), variations could be observed when both were compared to the New
Caledonian samples. From the TLC and HPLC analysis, it was concluded that all the
samples contained compound 2 (6Br-2OH-PBDE). The difference in the Solomon
Islands L. herbacea was that it lacked compound 3 (5Br-2OH-PBDE), and instead
showed the presence of compound 4 (5Br-OH-OMe-PBDE) and compound 1 (4Br-
OH-OMe-PBDE); the latter was also seen in the Type 1 and Type 3 L. herbacea
from New Caledonia. The details of structure determination are provided in Section
3.3. These results were confirmed using the LC-MS and 1-D and 2-D NMR spectra
obtained, and the extract was also fractionated to obtain clearer NMR profiles.
After fractionation of the sponge R3147C, the above mentioned compounds 1, 2 and
3 (4Br-OH-OMe-PBDE, 6Br-2OH-PBDE and 5Br-OH-OMe-PBDE) were confirmed
to be present in the sample. Together with these compounds, a fourth brominated
compound was also found in the fractions. The m/z of the compound was 688.89 mu
and it contained 6 bromine atoms. It resembled a 6Br-OH-OMe-PBDE. The 1HNMR
chemical shifts of this 6Br-OH-OMe-PBDE indicate protons that are meta-directed
(discussed in Section 3.3.2.2). However, due to low mass of product and impurities,
further analyses could not be carried forward.
The sponge sample from Fiji (R3241) had been initially classed as Lamellodysidea
herbacea but later it was reclassified to the species Dysidea lizardensis. After
thorough literature search, no further detail could be obtained for this new species. It
was seen that this sample also contained the brominated compounds found in L.
herbacea. Sample R3241 contains the two compounds found in the Type 2 L.
herbacea from New Caledonia – compounds 2 and 3 (6Br-2OH-PBDE and 5Br-
2OH-PBDE, respectively). However, it also contained another peak in the HPLC
profile which did not give a mass and the 1HNMR data at hand was not sufficient to
make any conclusions. Hence, the sample R3241C was also fractionated to see if
better profiles could be obtained for comparison and structure determination.
Presence of compounds 2 and 3 was confirmed in the sample after fractionation. A
third product, compound 8 (5Br-OH-OMe-PBDE) was also found to be present in the
sample and its structure was determined using spectral analyses (LC-MS and 1H and 13C NMR). It can be seen from this result that the D. lizardensis (R3241) studied is
quite closely related to L. herbacea.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
The results of the two fractionations are further discussed in Section 3.3 and spectral
details are provided in Appendix 10.
The five L. herbacea from Moorea Island had exactly the same chemical profiles and
contained mainly compound 1 (4Br-OH-OMe-PBDE), as seen with the Type 1
sample from New Caledonia. This compound was actually predicted since the
standard 4Br-OH-OMe-PBDE used in this study was also isolated from a Moorean L.
herbacea. The similarity between the Type 1 sample and the sponges from Moorea
can not be compared directly in terms of geographical distance since the two sites are
separated by quite a large distance.
The samples from Mayotte Island were exactly the same in their own profiles, but
when compared to all other samples, there was a clear difference in the pattern of
chemical profiles of these Indian Ocean sponges to that of the Pacific Ocean sponges
(from New Caledonia, Solomon Is., Fiji, Vanuatu, and French Polynesia). The L.
herbacea from Mayotte Island consisted of three main compounds, and upon further
investigation using LC-MS and NMR, it was found to consist of compound 6 – an
isomer of Compound 1 (4Br-OH-OMe-PBDE), compound 7 – an isomer of
Compound 3 (5Br-2OH-PBDE), and compound 8 – an isomer of compound 4 (5Br-
OH-OMe-PBDE). The details for structure determination of the compounds in the L.
herbacea from Mayotte are provided in Appendix 11.
,� � � � �3&��������%��)'�)'3�!�*����%�*'��5��)��'�'�(�&&��#�
Several contradictory observations can be made from the phylogenetic tree:
• The L. herbacea from Vanuatu (R2271bis) is closely related to most of the L.
herbacea from New Caledonia when comparing both the chemical
composition of the sponge as well as its phylogenic characteristics. So the
results of the phylogenetic tree and the chemical analysis match perfectly.
• The L. herbacea from Fiji (R3241), which has been later identified as
Dysidea lizardensis, is also genetically and chemically not very different
from the other L. herbacea from New Caledonia.
• On the other hand, the L. herbacea from Solomon Is. (R3147), where the
same type of PBDEs have been found compared to the other L. herbacea,
seems genetically very different from the other L. herbacea. This feature does

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
not correlate with the previous observations. Considering that PBDE are
suspected to be produced by the cyanobacterial symbionts, a close
relationship between PBDE and genetic mapping of the sponges can not be
confirmed without further analyses.
Another very interesting aspect noticed with the phylogenic study is that Dysidea
sponges from different regions form different clades of the taxonomy tree. This
feature is especially visible with the samples collected from the Solomon Islands.
The four Dysidea species of sponges from the Solomon Islands is clustered together
and distinct from the L. herbacea and D. arenaria collected from New Caledonia,
Fiji and Vanuatu.
Although a direct correlation has not been observed between the chemical profile and
the phylogenetic analysis of the Dysidea sponges in this study, chemotaxonomic
studies do not finish here. Further investigation will need to be carried out with focus
on compounds that are specific to certain groups of sponges, and to phylogenetic
analysis of the cyanobacterial symbionts associated with each sponge.
,� �, �(�������� ��� �������� ��������� ���� ��� ������ ���(�
��?����������������@�A�����,��������� ����������(�����
����(�����������
The HPLC analyses of Dysidea arenaria samples from New Caledonia and Fiji
showed considerable differences in their profiles. The New Caledonian specimens
seemed to contain various compounds with slight UV absorption, none of them being
the dominant constituent; and with the ELS detector, three groups of compounds
were detected to be dominant, whereby none of the compounds were UV active.
However, in case of the Fijian specimen, three main groups of compounds were
detected by the ELS detector, and all the signals had corresponding UV absorption as
well.
Similarly, looking at the chemical analyses of the D. avara samples from New
Caledonia and Fiji, it was evident that, like in D. arenaria, the two samples had quite
different profiles in HPLC. The New Caledonian sponge gave a lot of small peaks for
PDA scan, and one main cluster of compounds was observed with many other peaks
equivalent to noise. However, with the Fijian D. avara, two main compounds exist as

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
seen by the correlation between the PDA and the ELS scans of the extracts. (Details
of these correlations can be found in Appendix 12)
With TLC analysis, the Fijian samples of D. arenaria and D. avara also contained
phenolic compounds as visualized by Gibb’s reagent by a brownish-blue spot. These
were seen at Rf 0.11 and 0.21 for D. arenaria, and Rf 0.11 and 0.39 for D. avara.
However, due to the small quantities of the Fiji samples, and also because of time
constraints, further analyses could not be carried forward in order to determine the
exact point of chemical difference between the New Caledonian and Fijian D.
arenaria and D. avara sponges. As observed in the phylogenetic tree (Figure 21),
most of the D. arenaria samples collected from New Caledonia are more closely
related to each other than the L. herbacea obtained from the same region. The
chemically different D. arenaria from Fiji is also genetically different.
Chemical analyses (TLC, HPLC, LC-MS, and 1HNMR) of Dysidea 4177 samples
(R3033C and R3161C) indicated presence of polybrominated diphenyl ethers
(PBDE), basically 4Br-OH-PBDE (3,5-dibromo-2-(2’,4’-dibromophenoxy)phenol -
in both R3033C and R3161C) and 5Br-OH-PBDE (3,4,5-tribromo-2-(2’,4’-
dibromophenoxy)phenol – only in R3161C).
O
Br
Br
OH
Br Br
O
Br
Br
OH
Br
Br
Br
Compound 93,5-dibromo-2-(2',4'-dibromophenoxy)phenol Compound 10
3,4,5-tribromo-2-(2',4'-dibromophenoxy)phenol
12
34
5
61'2'
3'
4'5'
6'
Figure 23 - Brominated diphenyl ethers found in Dysidea 4177
The complete chemical screening details and structure determination is provided in
Appendix 13.
Three species of the genus Dysidea investigated previously have shown the presence
of brominated phenols, such as those described in Figure 23. These include L.
herbacea from Zanzibar (Sionov, et al., 2005), D. fragillis from Mozambique
(Utkina, Kazantseva, & Denisenko, 1987), and D. dendyi from Australia (Utkina,

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Denisenko, Virovaya, Scholokova, & Prokof'eva, 2002). It can be thus hypothesized
that due to the presence of brominated phenols in these Dysidea 4177 samples, they
may be linked more closely to L. herbacea, Dysidea fragillis, and Dysidea dendyi
than the other species. However, this hypothesis can not be proven until
classifications of these specimens are completed using morphological comparison
and genetic mapping.
As far as we know, the hypothesis of close links between Dysidea 4177 and L.
herbacea is not consistent with the genetic mapping of our extracts. As seen in the
phylogenetic tree (Figure 21), Dysidea 4177 from Solomon Is. are genetically
similar, but a little different from other Dysidea sp. from the Solomons, particularly
L. herbacea (R3147). On the other hand, they are very different from the L. herbacea
from New Caledonia. A large range study on the samples of L. herbacea, D. fragilis,
and D. dendyi from the Solomon Is. would be necessary to evaluate the chemical and
genetic proximity of these sponges.
From the different brominated diphenyl ether compounds observed in Dysidea
species, it was seen that there was a variety of the hydroxylated (-OH) or
methoxylated (-OMe) groups present in the molecules; apart from the number of
bromine atoms. Some compounds contained either 1 or 2 –OH’s or –OMe’s, while
others contained both –OH and –OMe. This pattern was not very particular of the
species of Dysidea or the location it was collected in. However, it was observed that
when molecules with 1-OH group were found, compounds with 2-OH groups were
not seen to be present (Handayani, et al., 1997; Zhang, Skildum, Stromquist, Rose-
Hellekant, & Chang, 2007; Fu & Schmitz, 1996). Nonetheless, this observation was
rejected in one study (Fu, Schmitz, Govindan, & Abbas, 1995), where these
compounds have been isolated together.
The chemical screening of Dysidea sp. specimen R3141C, indicated profiles that
were totally different from the other Dysidea species analysed in this study, and it did
not reveal any phenolic compounds by TLC either. This observation is surprising
since this sample is genetically close to the other Dysidea species from Solomon Is.
that contain PBDEs. Due to the lack of information about this species, it was chosen
for fractionation and product isolation, details of which are provided in Section 3.3.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
,� �. ��������
The ecological and geographical environments are major influences on the type and
amount of chemical compounds produced by a particular sponge. Due to their sessile
nature, these invertebrates tend to adapt to their constantly changing environments,
and eventually exhibit properties that are different when compared to the members of
the same species from other regions of the world. If these changes are not constant
throughout one particular region, variations seem to arise within the same region, as
seen with the New Caledonian sponges. This is another reason why chemotaxonomy
is difficult to apply to these invertebrates.
In order to determine the exact conditions due to which variations between the same
species occur in different geographical locations, more extensive studies need to be
carried out, where the sponge biology, ecology, and symbiosis can be analysed in
detail. Added to this, the geography, weather pattern and currents of the surrounding
environment should also be taken into account. This will allow better comparisons to
be made between organisms.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
,�, ����������+�������������(������������(������
���(����������������
Apart from the chemical analysis of the Dysidea sponges from New Caledonia, other
sponges of the genus Dysidea were also analyzed and certain constituents were
successfully isolated and characterized.
,�,�� ����������������,�.��
The shape and appearance of the Dysidea species R3141 was intriguing since it was
not at all like the other sponges. Thus, it was interesting to determine the chemical
constituents of this particular sample. It was also chosen for fractionation since it
contained totally different chromatographic and spectral profiles compared to L.
herbacea, D. arenaria, D. avara and the two Dysidea 4177 species analysed in this
investigation. The general 1HNMR was interesting since most peaks were seen
scattered throughout the profile with major peaks seen in the region of 0.5 to 2.0
ppm. The other peaks were quite small but it could be seen that they were basically
of the same height, which would indicate presence of peptides or related compounds.
Furthermore, no other details were known about this sample from the chemical
analyses since it did not even register a mass in the LC-MS profile of the crude
extract.
Figure 24 - 1HNMR [300 MHz; in CDCl3] and HPLC [Condition 2; UV 254 nm]
profiles of R3141C

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
A series of normal phase column chromatography and reverse phase semi-
preparative HPLC were required for the isolation of three compounds from this
species. (Details of the general isolation procedure are provided under
Methodology.)
The fraction (R3141C-fr2b) eluted using a 50/50 mixture of heptane and ethyl
acetate (EtOAc), was seen to contain most of the compounds found in the crude
extract. Hence it was fractionated a second time.
The interesting fractions in this set were fractions 8 to 11 (R3141C-2b-fr. 8, 9, 10,
and 11). These samples had similar HPLC profiles, and contained a compound which
was detectable by the ELS detector at 8.920 minutes, and which weakly absorbed
UV light at 214 nm. It also gave a mass-to-charge ratio (m/z) of 395.28 mass units.
The NMR profile looked very much like that of a peptide since signals were seen all
over the spectrum and most of the signals were present in the 0.5 to 3.5 ppm region,
with a few signals in the aromatic and -NH region also. Signals which could
correspond to the alpha carbon (C�) were also visible in the region between 4 and 5
ppm.
Figure 25 - 1HNMR profile of R3141C-2b-fr.8 [300 MHz; in CDCl3]
TLC profiling was carried out on the sample, and visualization was done using
Gibb’s reagent, HCl/ninhydrin and ninhydrin only. The Gibb’s visualization
confirmed that the compound did not have a phenolic group. Furthermore, an orange

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
spot appeared when using HCl/ninhydrin reagent and it was absent when only
ninhydrin was used. Plate 4 illustrates the TLC profiles obtained.
From this visualization, it was proposed that there was a peptide without free
nitrogen (free nitrogen can be detected by ninhydrin reagent only). Since the mass
obtained from LC-MS was quite low (395.28 mass units), there was a possibility that
the compound was a small cyclic peptide.
Plate 4 - TLC of R3141C-2b-fr.8-11 for the determination of peptides and phenols.
Eluent – 1/1 - heptane/EtOAc; Reagent - HCl/Ninhydrin (Left); Ninhydrin (Middle); and
Gibb’s reagent (Right)
This fraction was then purified using reverse phase semi-preparative HPLC. Out of
the 7 fractions collected, fraction 5 (R3141C-purif peak 5) was identified as the peak
of interest. However, the mass of the product obtained was very low. After verifying
all the fractions obtained, fraction 7 (R3141C-purif peak 7) was seen to contain 2
new products: which was not detectable in all the HPLC analyses carried out before
the purification using semi-preparative HPLC, and which could be two products of
degradation.
One hypothesis of such a behavior was that degradation of the compound of interest
occurred in the column during semi-preparative fractionation.
Hence, TLC of peak 7 (the supposed degradation products of peak 5) was carried out
with visualization using HCl/Ninhydrin and Ninhydrin only: 2 spots were seen with
different Rf than the original spot of peak 5, one spot being only visible with
HCl/Ninhydrin.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Plate 5 - TLC of R3141C-purif peak 7 for determination of peptides. Eluent – 1/1 -
heptane/EtOAc; Reagent - HCl/Ninhydrin (Left) and Ninhydrin (Right)
The original compound (peak 5) and the products of degradation were sent to the
University of Naples in Italy to be analyzed further.
Conclusions of the products obtained, as determined at the University of Naples,
suggest that the fractions contained a mixture of several compounds related to
Puupehenone. The 1HNMR (Figure 26) was composed of clusters of peaks generally
in the 0.6 – 2.6 ppm region. It is possible to say that these Puupehenone derivatives
are not methanol or ethyl acetate adducts (-OMe or O-CO-Me), since there are no
major peaks seen around 3.5 ppm in the 1HNMR spectrum.
�
Figure 26 - 1HNMR of R3141C peak 5 (fraction of R3141C) in CDCl3
Peak of degradation B
Peak of degradation A

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
These compounds are not peptides as predicted from the HCl/Ninhydrin reagent on
TLC. However, it is possible that when the TLC was sprayed with HCl, it formed a
hemiacetal which reacted with the ninhydrin giving an orange spot. So far, all
reactions with ninhydrin have been correlated to nitrogen-containing compounds.
Due to the low mass of the compounds obtained, it was not possible to confirm the
above hypothesis and the exact structure of the derivatives.
The following figure illustrates the possible Puupehenone derivatives present in the
sample R3141C:
O
O
OO
H
H
H
H
HOO
OOH
C21H28O3Exact Mass: 328.20Mol. Wt.: 328.45
C24H32O5Exact Mass: 400.22
Mol. Wt.: 400.51
Puupehenone
1112
13
14
1518
20
O
H
H
OH
O
O
C21H28O4Exact Mass: 344.20Mol. Wt.: 344.44
Figure 27: Puupehenone derivatives likely to be present in Dysidea species R3141C
Puupehenone and its derivatives belong to the sesquiterpene-substituted quinone
group of compounds (Ciavatta, et al., 2007; Bourguet-Kondracki, Lacombe, &
Guyot, 1999), constituting an important group of marine natural products in terms of
their bioactivities. They have been found to occur in sponges of the order Verongida,
genus Hyortis (Hamann, Scheuer, & Kelly-Borges, 1993; Sladi� & Gaši�, 2006;
Piña, Sanders, & Crews, 2003) and the order Dictyoceratida, genus Dysidea
(Ciavatta, et al., 2007). Puupehenone-related metabolites have been found to exhibit
biological activities such as cytotoxicity, anti-viral, anti-fungal, immunomodulatory
(Hamann, Scheuer, & Kelly-Borges, 1993; Ciavatta, et al., 2007), antibacterial, anti-
plasmodial (Bourguet-Kondracki, Lacombe, & Guyot, 1999), as well as anti-
tuberculosis and antitumor activities (Sladi� & Gaši�, 2006).

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
,�,� ��(��������������������
The L. herbacea samples from New Caledonia (R2252C), Solomon Islands
(R3147C) and Fiji Islands (R3241C) were fractionated in order to get a confirmation
of the exact structure of the PBDE present in them. All three samples were only put
through the first set of fractionation.
3.3.2.1 Sample R2252C (New Caledonia)
It was seen in the sample R2252C after drying of the fractions that crystals had
formed in one of the fractions. After recrystallization using acetonitrile, the product
was obtained as an almost pure product with slight traces of the other compound.
From the HPLC, LC-MS, and 1H and 13C NMR analyses, it was evident that the
crystals obtained was compound 2 (3,4,5,6-tetrabromo-2-(3’,5’-dibromo-2’-
hydroxyphenoxy)phenol), while the supernatant product contained a mixture of
compound 2 and compound 3 (3,5,6-tribromo-2-(3’,5’-dibromo-2’-
hydroxyphenoxy)phenol). The chemical shifts obtained have been presented in
Tables 3 and 4 in Section 3.1.1.4.2 and 3.1.1.4.3, respectively.
The NMR signals of compounds 2 and 3 matched very closely to that found in
literature (Norton, Croft, & Wells, 1981); hence the structure of the compounds were
assigned as follows:
O
OH OH
Br
Br
Br
Br
Br
Br
Compound 23,4,5,6-tetrabromo-2-(3',5'-dibromo-2'-hydroxyphenoxy)phenol
6
5
1
43
22'
1'6'
5'4'
3'
O
OH OH
Br
Br
Br Br
Br
Compound 33,5,6-tribromo-2-(3',5'-dibromo-2'-hydroxyphenoxy)phenol
6
5
1
43
22'
1'6'
5'4'
3'
Figure 28 - Structures for Compounds 2 and 3
Furthermore, the melting point of the main product (Compound 2 – 3,4,5,6-
tetrabromo-2-(3’,5’-dibromo-2’-hydroxyphenoxy)phenol) was also determined and
was found to be in the range of 191.0 oC to 191.6 oC. This is very close to the value
obtained in literature, which is between 194 oC to 195 oC (Norton, Croft, & Wells,
1981). Thus, the conclusions of compounds 2 and 3 made in Section 3.1.1 have been
confirmed.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
3.3.2.2 Sample R3147C (Solomon Is.)
In case of the fractions of sample R3147C, crystals were also observed in the
samples, but recrystallization using hot acetonitrile failed, and crystals of the
products could not be obtained.
From the fractions of R3147C, the first fraction yielded two compounds as observed
in the HPLC chromatogram, where the first compound did not have UV absorption,
but exhibited a mass-to-charge ratio (m/z) of 609.06 mu. This compound did not
show presence of halogenation in the mass spectrum, and was more in the non-polar
region of the spectra. However, there was an aromatic moiety in this compound due
to the chemical shifts observed in the 1HNMR profile. The second (major) compound
(compound 5) in the first fraction had an m/z of 688.89 mu and the isotopic spectra
confirmed the presence of 6 bromine atoms. The closest possible structure with 6
bromine and mass of 689 mu found in literature was a 6Br-OH-OMe-PBDE.
O
O OH
Br
Br
Br
Br
Br
Br
Compound 53,4,5,6-tetrabromo-2-(3',5'-dibromo-2'methoxyphenoxy)phenol
12
34
5
61'2'
3'
4'5'
6'
Figure 29 - Proposed structure for compound 5 (3,4,5,6-tetrabromo-2-(3’,5’-dibromo-
2’-methoxyphenoxy)phenol)
NMR data could not be obtained to confirm this hypothesis due to the small quantity
of product available.
In the second fraction, a mixture of 3 compounds was obtained, out of which the
major compound was determined to be compound 4 (5Br-OH-OMe-PBDE). The m/z
of this compound was obtained to be 608.98 mu with the isotopic spectra indicating 5
bromine atoms. A fragment peak of the compound was also obtained at m/z of
346.47 mu, which corresponds to the loss of the ring containing the methoxy group
and 2 bromine atoms, hence leaving the second phenolic ring with three bromine
atoms. The 13C and 1H-NMR (Table 5) correlate well with the following structure:

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
O
OHO
BrBr
Br
BrBr
165
43
23'
5'
1'
6'4'
2'
Compound 43,5,6-tribromo-2-(3',5'-dibromo-2'-methoxyphenoxy)phenol
Figure 30 - Structure of Compound 4 (3,5,6-tribromo-2-(3’,5’-dibromo-2’-
methoxyphenoxy)phenol)
Table 5 - Chemical shifts obtained for Compound 4 (at 400 MHz, in acetone-d6)
C # 13C (ppm)
1H (ppm)
�H Lit. in Acetone-d6 (ppm)*
13C 1H 1 151.50 150.8 2 139.78 139.0 3 117.65 117.0 4 128.71 7.65 (s) 127.9 7.65 (s) 5 124.05 123.3 6 115.80 115.1 1’ 152.64 151.9 2’ 147.20 146.5 3’ 120.13 119.5 4’ 130.38 7.48 (d, 2.2 Hz) 129.7 7.48 (d, 2.2 Hz) 5’ 117.62 116.9 6’ 118.18 6.82 (d, 2.2 Hz) 117.5 6.81 (d, 2.5 Hz)
-OMe 61.76 4.00 61.1 4.01 *Hanif, et al., 2007
The third fraction yielded compound 2, and thus confirmed the presence of this
compound in the sample. The 1HNMR chemical shifts obtained were also
comparable to the chemical shifts of this compound isolated from the other L.
herbacea from New Caledonia. (Spectral data of the fractions are present in
Appendix 10)

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
3.3.2.3 Sample R3241C (Fiji Is.)
Just like in sample R3147, the fractions obtained for R3241C also showed presence
of crystals, but isolation of the pure compound was not successful using acetonitrile.
From the spectral data of the fractions, two of the compounds were easily identified
to be compound 2 (6Br-2OH-PBDE) and compound 3 (5Br-2OH-PBDE). Two other
compounds present in this specimen were compound 6 (4Br-OH-OMe-PBDE -
isomer of Compound 1) and compound 8 (5Br-OH-OMe-PBDE - isomer of the
Compound 4 obtained in R3147 extracts - Figure 30).
O
Br
R
Br
O
OH
Br
Br
R = H --> Compound 6 : 4,6-dibromo-2-(4',6'-dibromo-2'-methoxyphenoxy)phenolR = Br --> Compound 8 : 4,6-dibromo-2-(4',5',6'-tribromo-2'-methoxyphenoxy)phenol
12
34
5
62'
1'
6'5'
4'
3'
Figure 31 - Compounds 6 and 8 obtained from sample R3241C
Table 6 - Chemical shifts obtained for Compounds 6 and 8 (in Acetone-d6, at
400 MHz)
C # Compound 6 Compound 8 Literature 13C
(ppm)
1H (ppm)
13C (ppm)
1H (ppm)
*Compd. 6 (CDCl3)
**Compd. 8 (CDCl3)
1 145.17 145.13 142.8 142.7 2 147.55 147.24 145.2 144.7 3 117.15 6.62 117.19 6.71 115.7 (6.53) 116.3 (6.51) 4 111.52 111.62 110.0 111.5 5 129.91 7.377 130.11 7.39 129.2 (7.33) 129.3 (7.34) 6 111.90 112.00 111.5 110.1 1’ 141.14 142.16 139.9 140.8 2’ 155.29 154.00 153.4 152.0 3’ 117.76 7.425 119.02 7.64 116.5 (7.09) 116.6 (7.28) 4’ 120.71 119.52 119.8 119.1 5’ 128.31 7.49 123.52 127.5 (7.40) 122.8 6’ 123.02 118.6 122.4
-OMe 3.87 3.87 56.6 (3.78) 56.7 (3.78) *Fu, Schmitz, Govindan, & Abbas, 1995 **Liu, Namikoshi, Meguro, Nagai, Kobayashi, & Yao, 2004
(Spectral data of the fractions are present in Appendix 10)

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
3.3.2.4 Sample MAY-02-Dh2 (L. herbacea from Mayotte)
L. herbacea from Mayotte contained similar brominated compounds as that obtained
from L. herbacea from the other sampling sites. Three main compounds could be
isolated from this sample: compound 6 (4Br-OH-OMe-PBDE - isomer of Compound
1), compound 7 (5Br-2OH-PBDE - isomer of Compound 3), and compound 8 (5Br-
OH-OMe-PBDE - isomer of Compound 4).
The chemical shifts obtained for compounds 6 and 8 were almost the same as that
obtained for these compounds in sample R3241C (the structures were confirmed by
HSQC and HMBC spectra). The structure of compound 7 (isomer of compound 3)
was confirmed as follows using 1-D and 2-D NMR:
OH
Br
Br
Br
O
OH
Br
Br
Compound 7 - 4,6-dibromo-2-(4',5',6'-tribromo-2'-hydroxyphenoxy)phenol
12
34
5
62'
1'
6'5'
4'
3'
Figure 32 - Structure of Compound 7 (4,6-dibromo-2-(4’,5’,6’-tribromo-2’-
hydroxyphenoxy)phenol)
Table 7 - Chemical shifts obtained for Compound 7 (in acetone-d6, 400 MHz)
Carbon #
Compound 7 Literature* in CDCl3 (ppm) 13C (ppm) 1H (ppm) HMBC
1 144.88 2 146.75 3 116.71 6.73 144.88 / 129.76 / 111.34 6.46 4 111.34 5 129.76 7.38 144.88 / 116.71 / 111.69 7.23 6 111.69 1’ 151.63 2’ 140.70 3’ 122.35 7.49 140.70 / 123.05 / 118.23 7.26 4’ 118.23 5’ 123.05 6’ 122.64
*Fu, Schmitz, Govindan, & Abbas, 1995

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
,�,�, ��������
The compounds obtained from sample R3141C seems to be quite sensitive in terms
of stability, and the exact reason for the degradation behavior is not known yet.
The fractionations of L. herbacea sample R2252C from New Caledonia and the L.
herbacea from Mayotte were successful in isolating the major compounds of interest,
and to confirm their structures. Pure compounds of L. herbacea R3147C and D.
lizardensis R3241C could not be obtained but the fractions were clear enough to
provide some data to confirm the compounds present in the samples.
Thus, the compounds obtained from this analysis include tetra-, penta-, and hexa-
brominated diphenyl ethers, with either two hydroxy groups or one hydroxy and one
methoxy groups.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
,�. ����<���������������
Polybrominated diphenyl ethers are a class of phytopathogenic fungicides (Peng, et
al., 2003). The antifungal bioassay in this study was carried out using Cryptococcus
neoformans (ATCC 32045) and wild type Candida albicans (WTCA).
It was evident from the results obtained that although the Dysidea species showed
growth inhibition of Cryptococcus neoformans (and a few samples showed inhibition
of WTCA), the inhibition was not as good as the control (Nystatin) it was tested
with. Nystatin is one of the first commercial drugs used for the treatment of
opportunistic fungal infections. This however, was a preliminary test to see if the
crude samples were active or not, and if purified, it could show interesting activity at
the appropriate dosage.
Figure 33 - Inhibition of Cryptococcus neoformans and WTCA by crude and
fractionated extracts of Dysidea sponges (Dosage of crude extracts = 1 mg per disc;
Dosage of fractionated extracts = 0.1 mg per disc)
It was also interesting to see that the two compounds 1 and 2 had a vast difference in
their activity patterns. Compound 1 (4Br-OH-OMe-PBDE) was more active than
Compound 2 (6Br-2OH-PBDE), which was barely active. Similarly, when
comparing Compound 1 with the brominated metabolites of Dysidea sp. 4177
(Compounds 9 and 10 from samples R3033C and R3161C), it is clear that the penta-
bromo-diphenyl ether (Compound 10) is more active than Compound 1, followed by
the tetra-bromo-diphenyl ether (Compound 9). The following figure shows the
����
�����������
,���
�&,�
���&
,���
�&,�
���&
,���
�&,�
���&
,���
&,�
���&
,��
�&,�
��&
,��
�8��&
,���
&,�
���&
,��
�&,�
�&
,��
�&,�
��&
'��
���%
!�'������%!�
,���
8��&
,���
�&,�
��&
,���
�&,�
���8
��&
,���
�&,�
���&
,���
&,�
��&
,���
�&,�
���&
,��
�&,�
���&
,���
�&,�
���&
,���
�&�B
�*)(*)
'�*.B
%"
�B�*�)
(*.B%
",�
���&
*$����$
��@��
,���
�&*��<�
,���
�&*��<�
,���
�&*��<�
,���
&*��<�
,���
&*��<�
,���
&*��<�
,���
�&*��<�
,���
�&*��<�
,���
�&*��<�
%&'
����
����
7�������
�,��<
,�$�-��� �� ����������� .�/0+1�-"�������#��%���#������ �� �� ����
&��$�� 2-&4
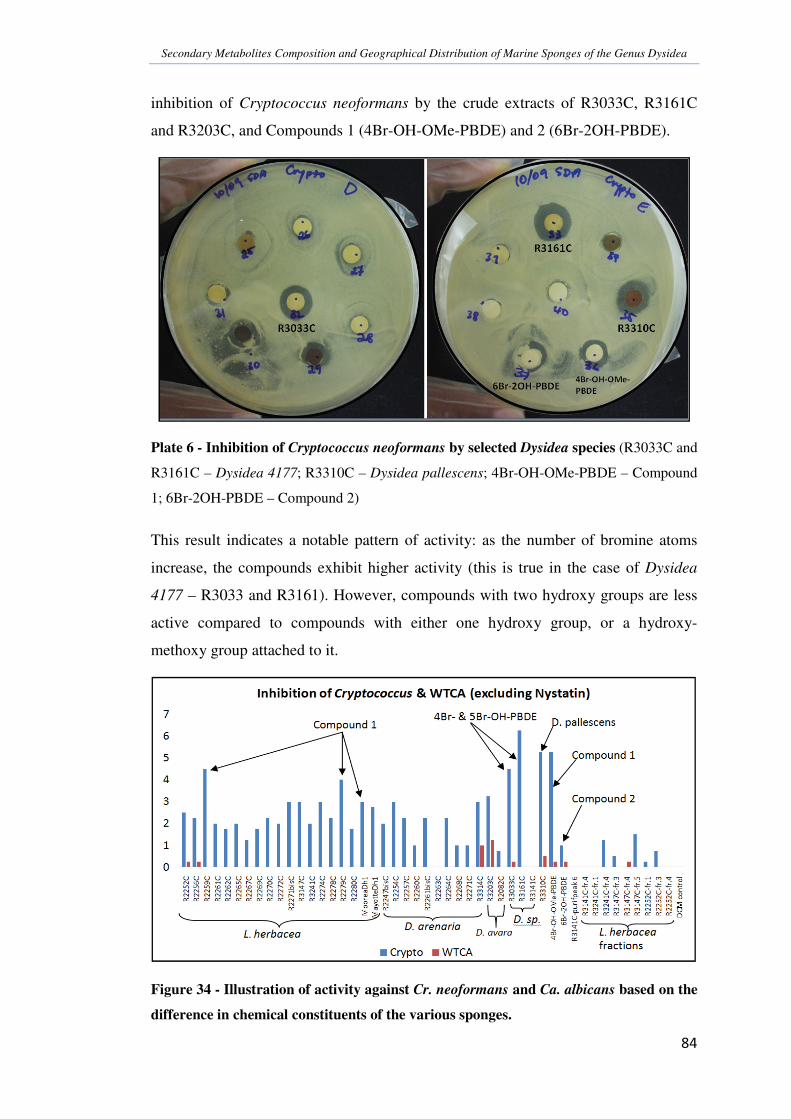
Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
inhibition of Cryptococcus neoformans by the crude extracts of R3033C, R3161C
and R3203C, and Compounds 1 (4Br-OH-OMe-PBDE) and 2 (6Br-2OH-PBDE).
Plate 6 - Inhibition of Cryptococcus neoformans by selected Dysidea species (R3033C and
R3161C – Dysidea 4177; R3310C – Dysidea pallescens; 4Br-OH-OMe-PBDE – Compound
1; 6Br-2OH-PBDE – Compound 2)
This result indicates a notable pattern of activity: as the number of bromine atoms
increase, the compounds exhibit higher activity (this is true in the case of Dysidea
4177 – R3033 and R3161). However, compounds with two hydroxy groups are less
active compared to compounds with either one hydroxy group, or a hydroxy-
methoxy group attached to it.
Figure 34 - Illustration of activity against Cr. neoformans and Ca. albicans based on the
difference in chemical constituents of the various sponges.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Added to this, slight distinction in activity could also be seen within L. herbacea,
especially with the specimens containing Compound 1. These samples (R2259C and
R2279C) have been discussed previously regarding their unique chemical profiles
compared to other L. herbacea samples collected from New Caledonia (Section
3.2.2.1). Furthermore, even though brominated compounds have been found to be
responsible for the biological activity against the fungi, it is important to note that
there are other active compounds present in the Dysidea arenaria and Dysidea avara
specimens that have also showed slight activity.
It is also clear from the results that most of the Dysidea extracts are more active
against Cryptococcus neoformans than Candida albicans. This was the expected
result for the preliminary anti-malarial studies, and it can now be further investigated
for anti-plasmodium activities. As discussed before, one of the differences between
Cr. neoformans and Ca. albicans is that the former species requires the protein
farnesyl transferase (PFT) for viability, whereas the latter fungus can survive without
it. The result obtained in this assay proves that these compounds inhibit the PFT from
functioning normally. Consequently, it suggests a plausible mode of action for any
activity shown against the Plasmodium species, which is also dependent on the
proper functioning of the PFT for viability.
Although the specimens tested for anti-fungal activities did not prove to be very
challenging, it can still be a candidate for various other assays. It can also be tested
directly on Plasmodium species in order to determine another mode of action,
separate from the PFT inhibition.
A study carried out by Peng, et al., (2003) indicated that comparing activity of
sponge-derived PBDEs and manzamine alkaloids against weeds, fungi, and malarial
parasites, the latter group of metabolites showed more promising activity against
Plasmodium berghei. Therefore, this group of metabolites can also be explored in
order to find a drug candidate for Plasmodium infections.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
. ����������������
Sponge metabolites have been the targets of research for the past three decades. Due
to the large biodiversity of these organisms, there are still quite a lot of species that
have not been discovered and studied. Many analyses of sponge metabolites are
directed towards drug discovery. Apart from this, chemotaxonomy is also a very
important research field, whereby chemical constituents of these organisms can be
used together with biological taxonomy to classify the various organisms accurately.
The aspect of chemotaxonomy was partly studied in this research, and it was found
that one of the biggest problems is that the main extractable chemicals are produced
by symbionts and not by the host itself. The main group of compounds found in this
study was PBDEs. These compounds are suspected to be produced by the symbiont
cyanobacteria, Oscillatoria spongeliae, present in L. herbacea sponges. Due to this
reason, a sponge-specific metabolite could not be identified as yet, which could have
been a candidate for a chemical marker in the Dysidea sponges.
An intra- and inter-geographical comparison was also carried out with L. herbacea
from New Caledonia, Vanuatu, Fiji, Solomon Is., Moorea, and Mayotte islands. It
was observed that the sponges are quite sensitive to variations, and due to this,
certain specimen of the same species gave slightly different profiles, while the others
were quite the same. It would be interesting to know if all the L. herbacea studied
contain the same strain of cyanobacteria since all extracts contained brominated
diphenyl ethers (bromo-chemotype).
Also in this study, eight compounds have been identified with the help of spectral
analysis. Out of these eight compounds, four compounds (Compound 2, 6, 7 and 8)
have been isolated successfully, while the others were obtained as mixtures or
impure products. Finally, the anti-fungal assay against Cryptococcus neoformans and
Candida albicans mainly showed moderate activity against Cr. neoformans,
suggesting that the mode of action was via the inhibition of the enzyme protein
farnesyl transferase. This result indicates that the activity of these compounds against
the Plasmodium species is also possible through the same mode.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
0 �������������
Agrawal, M. S., & Bowden, B. F. (2005). Marine sponge Dysidea herbacea
revisited: Another brominated diphenyl ether. Marine Drugs , 3, 9-14.
Aiello, A., Fattorusso, E., Menna, M., & Pansini, M. (1996). The Chemistry of the
demosponge Dysidea fragilis from the Lagoon of Venice. Biochemical
Systematics and Ecology , 24 (1), 37-42.
Alvi, K. A., Diaz, M. C., Crews, P., Slate, D. L., Lee, R. H., & Moretti, R. (1992).
Evaluation of new sesquiterpene quinones from two Dysidea sponge species as
inhibitors of protein tyrosine kinase. J. Org. Chem. , 57, 6604-6607.
Anjaneyulu, V., Rao, K. N., Radhika, P., Muralikrishna, M., & Connolly, J. D.
(1996). A new tetrabromodiphenyl etherfrom the sponge Dysidea herbacea of the
Indian Ocean. Indian Journal of Chemistry , 35B, 89-90.
Ardá, A., Jiménez, C., & Rodríguez, J. (2004). A study of polychlorinated leucine
derivatives: Synthesis of (2S,4S)-5,5-dichloroleucine. Tetrahedron Letters , 45,
3241-3243.
Ardá, A., Rodríguez, J., Nieto, R. M., Bassarello, C., Gomez-Paloma, L., Bifulco, G.,
et al. (2005). NMR J-based analysis of nitrogen-containing moieties and
application to dysithiazolamide, a new polychlorinated dipeptide from Dysidea sp.
Tetrahedron , 61, 10093-10098.
Armstrong, E., McKenzie, J. D., & Goldworthy, G. T. (1999). Aquaculture of
sponges on scallops for natural product research and antifouling. Journal of
Biotechnology , 70, 163-174.
Austin, W. C. (2003). Natural History. Retrieved May 10, 2008, from Sponge
gardens: A hidden treasure in British Columbia:
http://www.mareco.org/khoyatan/spongegardens
Becerro, M. A., & Paul, V. J. (2004). Effects of depth and light on the secondary
metabolites and cyanobacterial symbionts of the sponge Dysidea granulosa.
Marine Ecology progress series , 280, 115-128.
Belarbi, E. H., Gómez, A. C., Christi, Y., Camacho, F. G., & Grima, E. M. (2003).
Producing drugs from marine sponges. Biotechnology Advances , 21, 585-598.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
�
Bergquist, P. R. (1998). Order Dictyoceratida. In C. Lévy (Ed.), Sponges of the New
Caledonian Lagoon (p. 157). Paris: Orstrom Editions.
Bergquist, P. R. (1978). Sponges. London: The Anchor Press Ltd.
Blunt, J. W., Copp, B. R., Munro, M. H., Northcote, P. T., & Prinsep, M. R. (2006).
Marine Natural Products. Nat. Prod. Rep. , 23, 26-78.
Bourguet-Kondracki, M.-L., Lacombe, F., & Guyot, M. (1999). Methanol adduct of
Puupehenone, A biologically active derivative from the marine sponge Hyrtios
species. J. Nat. Prod. , 62 (9), 1304-1305.
Brusca, R. C., & Brusca, G. J. (2003). Phylum Porifera: The Sponges. In
Invertebrates (2nd Edition ed., pp. 179-208). USA: Sinauer Associates, Inc.
Calderone, R. A. (2002). Intorduction and Historical Perspectives. In R. A.
Calderone (Ed.), Candida and Candidiasis (pp. 7-10). Washington D.C., USA:
ASM Press.
Cameron, G. M., Stapleton, B. L., Simonsen, S. M., Brecknell, D. J., & Garson, M. J.
(2000). New sesquiterpene and brominated metabolites from the tropical marine
sponge Dysidea sp. Tetrahedron , 56, 5247-5252.
Capon, R. J., & Faulkner, D. J. (1985). Herbasterol, an Ichthyotoxic 9,11-Secosterol
from the sponge Dysidea herbacea. J. Org. Chem. , 50, 4771-4773.
Capon, R. J., & MacLeod, J. K. (1987). Thiofurodysinin, a sulfur-containing
furanosesquiterpene from the marine sponge Dysidea avara. J. Nat. Prod. , 50 (6),
1136-1137.
Capon, R., Ghisaberti, E. L., Jeffries, P. R., Skelton, B. W., & White, A. H. (1981).
Structural studies of halogenated diphenyl ethers from a marine sponge. J. Chem.
Soc. Perkin , 1, 2464-2467.
Carballo, J. L., Zubía, E., & Ortega, M. J. (2006). Biological and chemical
characterizations of three new species of Dysidea (Porifera: Demospongiae) from
the Pacific Mexican coast. Biochemical Systematics and Ecology , 34, 498-508.
Cardellina, J. H., & Barnekow, D. E. (1988). Oxidized nakafuran 8 sesquiterpenes
from the sponge Dysidea etheria. Structure, stereochemistry, and biological
activity. J. Org. Chem. , 53, 882-884.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Cardellina, J. H., Nigh, D., & VanWagenen, B. C. (1986). Plant growth regulatory
indoles from the sponge Dysidea etheria and Ulosa ruetzleri. J. Nat. Prod. , 49
(6), 1065-1067.
Carmely, S., Cojocaru, M., Loya, Y., & Kashman, Y. (1988). Ten new rearranged
spongian diterpenes from two Dysidea species. J. Org. Chem. , 53 (20), 4801-
4807.
Casapullo, A., Minale, L., Zollo, F., Roussakis, C., & Verbist, J. F. (1995). New
cytotoxic polyoxygenated steroids from the sponge Dysidea incrustans.
Tetrahedron Letters , 36 (15), 2669-2672.
Ciavatta, M. L., Gresa, M. P., Gavagnin, M., Romero, V., Melck, D., Manzo, E., et
al. (2007). Studies on Puupehenone-metabolites of a Dysidea sp.: structure and
biological activity. Tetrahedron , 63, 1380-1384.
Clark, W. D., & Crews, P. (1995). A novel chlorinated ketide amino acid, Herbamide
A, from the marine sponge, Dysidea herbacea. Tetrahedron Letters , 36 (8), 1185-
1188.
Crispino, A., de Giulio, A., de Rosa, S., & Strazzullo, G. (1989). New bioactive
derivative of avarol from the marine sponge Dysidea avara. J. Nat. Prod. , 52 (3),
646-648.
de Guzman, F. S., Copp, B. R., Mayne, C. L., Concepcion, G. P., Mangalindan, G.
C., Barrows, L. R., et al. (1998). Bolinaquinone: A novel cytotoxic sesquiterpene
hydroxyquinone from a Philippine Dysidea sponge. J. Org. Chem. , 63 (22), 8042-
8044.
De la Fuente, J. A., Manzanaro, S., Martin, M. J., De Quesada, T. G., Reymundo, I.,
Luengo, S. M., et al. (2003). Synthesis, activity, and molecular modeling studies
of novel human aldose reductase inhibitors based on a marine natural product. J.
Med. Chem. , 46, 5208-5221.
De Rijke, E., Zappey, H., Ariese, F., Gooijer, C., & Brinkman, U. A. (2003). Liquid
chromatography with atmospheric pressure chemical ionization and electrospray
ionization mass spectrometry of flavonoids with triple-quadrupole and ion-trap
instruments. Journal of Chromatography A , 984, 45-58.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Debitus, C. (1998). The Fascinating world of sponge chemistry & chemical ecology.
In C. Lévy (Ed.), Sponges of the New Caledonian Lagoon (pp. 61-64). Paris:
Orstrom editions.
Diaz-Marrero, A. R., Austin, P., van Soest, R., Matainaho, T., Roskelley, C. D.,
Roberge, M., et al. (2006). Avinosol, A meroterpenoid-nucleoside conjugate with
anti-invasion activity isolated from the marine sponge Dysidea sp. Organic
Letters , 8 (17), 3749-3752.
Dumdei, E. J., Simpson, J. S., Garson, M. J., Byriel, K. A., & Kennard, C. H. (1997).
New chlorinated metabolites from the tropical marine sponge Dysidea herbacea.
Aust. J. Chem. , 50, 139-144.
Elyakov, G. B., Kuznetsova, T., Mikhailov, V. V., Maltsev, I. I., Voinov, V. G., &
Fedoreyev, S. A. (1991). Brominated diphenyl ethers from a marine bacterium
associated with the sponge Dysidea sp. Experientia , 47, 632-633.
Erpenbeck, D., & Van Soest, R. W. (2006). Status and perspective of sponge
chemosystematics. Marine Biotechnology , 9, 2-19.
Erpenbeck, D., Breeuwer, J. A., Parra, F. J., & van Soest, R. W. (2006). Molecular
Phylogenetics and Evolution. 38, 293-305.
Farnswort, R. N. (1984). In P. Krogsgaard-Larsen, S. B. Christensen, & H. Kofod
(Eds.), Natural products and drug discovery (p. 17). Copenhagen: Munksgaard.
Faulkner, J. D. (2000). Marine Natural Products. Nat. Prod. Rep. , 17 (1), 7-55.
Fayette, J., Coquard, I. R., Alberti, L., Ranchere, D., Boyle, H., & Blay, J.-Y. (2005).
ET-743: A novel agent with activity in soft tissue sarcomas. The Oncologist , 10,
827-832.
Flatt, P. M., Gautshci, J. T., Thacker, R. W., Musafija-Girt, M., Crews, P., &
Gerwick, W. H. (2005). Identification of the cellular site of polychlorinated
peptide biosynthesisin the marine sponge Dysidea (Lamellodysidea) herbacea and
symbiotic cyanobacterium Oscillatoria spongeliae by CARD-FISH analysis.
Marine Biology , 147, 761-774.
Flowers, A. E., Garson, M. J., Webb, R. I., Dumbei, E. J., & Charan, R. D. (1998).
Cellular origin of chlorinated diketopiperazines in the dictyoceratid sponge
Dysidea herbacea (Keller). Cell Tissue Res. , 292, 597-607.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Fu, X., & Schmitz, F. J. (1996). New brominateddiphenyl ether from an unidentified
species of Dysidea sponge. 13C NMR data for some brominated diphenyl ethers.
J. Nat. Prod. , 59 (11), 1102-1103.
Fu, X., Ferreira, M. L., Schmitz, F. J., & Kelly-Borges, M. (1998). New
diketopiperazines from the sponge Dysidea chlorea. J. Nat. Prod. , 61 (10), 1226-
1231.
Fu, X., Schmitz, F. J., Govindan, M., & Abbas, S. A. (1995). Enzyme inhibitors:
New and known polybrominated phenols and diphenyl ethers from four Indo-
Pacific Dysidea sponges. J. Nat. Prod. , 58 (9), 1384-1391.
Fu, X., Zeng, L.-M., Su, J.-Y., & Pais, M. (1997). A new diketopiperazine derivative
from the South China Sea sponge Dysidea fragilis. J. Nat. Prod. , 60 (7), 695-696.
Garson, M. J. (2001). Ecological perspectives on marine natural product
biosynthesis. In M. J. B, & B. B. T (Eds.), Marine Chemical Ecology (pp. 71-
114). Boca Raton, Florida: CRC Press.
Garson, M. J., Dexter, A. F., Lambert, L. K., & Liokas, V. (1992). Isolation of the
bioactive terpene 7-deacetoxy-olepupuane from the temperate marine sponge
Dysidea sp. J. Nat. Prod. , 55 (3), 364-367.
Gelb, M. H., Van Voorhis, W. C., Buckner, F. S., Yokoyama, K., R, E., Carpenter, E.
P., et al. (2003). Protein farnesyl and N-myristoyl transferases: Piggy-back
medicinal chemistry targets for the development of anti-trypanosomatid and
antimalarial therapeutics. Molecular and Biochemical Parasitology , 126, 155-
163.
Giannini, C., Debitus, C., Lucas, R., Ubeda, A., Payá, M., Hooper, J. N., et al.
(2001). New sesquiterpene derivatives from the sponge Dysidea species with a
selective inhibitor profile against Human phospholipase A2 and other leukocyte
functions. J. Nat. Prod. , 64 (5), 612-615.
Giannini, C., Debitus, C., Posadas, I., Payá, M., & D'Auria, M. V. (2000).
Dysidotronic acid, a new and selective human phospholipase A2 inhibitor from
the sponge Dysidea sp. Tetrahedron Letters , 41, 3257-3260.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Goetz, G. H., Harrigan, G. G., & Likos, J. (2001). Furodysin lactone and
pyrodysinoic acid, new sesquiterpenes from a Philippines Dysidea species. J. Nat.
Prod. , 64 (11), 1486-1488.
Grode, S. H., & Cardellina, J. H. (1984). Sesquiterpenes from the sponge Dysidea
etheria and the nudibranch Hypselodoris zebra. J. Nat. Prod. , 47 (1), 76-83.
Grozdanov, L., & Hentschel, U. (2007). An environmental genomics perspective on
the diversity and function of marine sponge-associated microbiota. Current
Opinion in Microbiology , 10, 215-220.
Gunasekera, S. P., McCarthy, P. J., Kelly-Borges, M., Lobkovsky, E., & Clardy, J.
(1996). Dysidiolide: A novel protein phosphatase inhibitor from the Caribbean
sponge Dysidea etheria de Laubenfels. J. Am. Chem. Soc. , 118, 8759-8760.
Haefner, B. (2003). Drugs from the deep: Marine natural products as drug
candidates. Drug Discovery Today , 8 (12), 536-544.
Hamann, M. T., Scheuer, P. J., & Kelly-Borges, M. (1993). Biogenetically diverse,
Bioactive constituents of a sponge, Order Verongida: Bromotyramines and
Sesquiterpene-shikimate derived metabolites. J. Org. Chem. , 58 (24), 6565-6569.
Handayani, D., Edrada, R. A., Proksch, P., Wray, V., Witte, L., Van Soest, R. W., et
al. (1997). Four new bioactive polybrominated diphenyl ethers of the sponge
Dysidea herbacea from West Sumatra, Indonesia. J. Nat. Prod. , 60 (12), 1313-
1316.
Hanif, N., Tanaka, J., Setiawan, A., Trianto, A., De Voogd, N. J., Murni, A., et al.
(2007). Polybrominated diphenyl ethers from the Indonesian sponge
Lamellodysidea herbacea. J. Nat. Prod. , 70 (3), 432-435.
Harrigan, G. G., Goetz, G. H., Luesch, H., Yang, S., & Likos, J. (2001).
Dysideaprolines A-F and Barbaleucamides A-B, Novel polychlorinated
compounds from a Dysidea species. J. Nat. Prod. , 64 (9), 1133-1138.
Hildebrandt, M., Waggoner, L. E., Lim, G. E., Sharp, K. H., Ridley, C. P., &
Haygood, M. G. (2004). Approaches to identify, clone, and express symbiont
bioactive metabolite genes. Nat. Prod. Rep. , 21, 122-142.
Hoffmann, F., Rapp, H. T., & Reitner, J. (2005). Anaerobic world in sponges.
Geomicrobiol. J. , 22, 1-10.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Hooper, J. (1998). How can you identify a sponge? In C. Lévy (Ed.), Sponges of the
New Caledonian Lagoon (p. 39). Paris: Orstrom editions.
Hooper, J. N., & Van Soest, R. W. (2002). Introduction. In J. N. Hooper, & R. W.
Soest (Eds.), Systema Porifera: a guide to the classification of sponges (Vol. 1,
pp. 1-3). New York: Kluwer Academic / Plenum Publishers.
Horton, P. A., & Crews, P. (1995). The Arenarans, sesquiterpene ethers from the
marine sponge Dysidea arenaria. J. Nat. Prod. , 58 (1), 44-50.
Horton, P., Inman, W. D., & Crews, P. (1990). Enantiomeric relationships and
antihelmintic activity of Dysinin derivatives from Dysidea marine sponges. J. Nat.
Prod. , 53 (1), 143-151.
Hunter, B. B., Barnett, H. L., & Buckelew, T. P. (1978). Deuteromycetes (Fungi
Imperfecti). In A. I. Laskin, & H. A. Lechevalier (Eds.), CRC Handbook of
Microbiology (2nd Edition ed., Vol. II, p. 283). USA: CRC Press, Inc.
Isaacs, S., Berman, R., Kashman, Y., Gebreyesus, T., & Yosief, T. (1991). New
polyhydroxy sterols, Dysidamides, and a dideoxyhexose from the sponge Dysidea
herbacea. J. Nat. Prod. , 54 (1), 83-91.
Jacob, M. R., Hossain, C. F., Mohammed, K. A., Smillie, T. J., Clark, A. M., Walker,
L. A., et al. (2003). Reversal of fluconazole resistance in multi-drug efflux-
resistant fungi by the Dysidea arenaria sponge sterol 9�,11�-epoxycholest-7-ene-
3�,5�,6�,19-tetrol 6-acetate. J. Nat. Prod. , 66 (12), 1618-1622.
Jimenez, J. I., & Scheuer, P. J. (2001). New lipopeptides from the Caribbean
cyanobacterium Lyngbya majuscula. J. Nat. Prod. , 64, 200-203.
Kuniyoshi, M., Yamada, K., & Higa, T. (1985). A biologically active diphenyl ether
from the green alga Cladosphora fascicularis. Experientia , 41, 523-524.
Lévi, C. (1998). What is a sponge? In C. Lévy (Ed.), Sponges of the New Caledonian
lagoon (pp. 27-34). Paris: Orstrom editions.
Liu, H., Namikoshi, M., Meguro, S., Nagai, H., Kobayashi, H., & Yao, X. (2004).
Isolation and characterization of polybrominated diphenyl ethers as inhibitors of
microtubule assembly from the marine sponge Phyllospongia dendyi collected at
Palau. J. Nat. Prod. , 67 (3), 472-474.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Lu, Q., & Faulkner, D. J. (1998). Three dolabellanes and a macrolide from the
sponge Dysidea sp. from Palau. J. Nat. Prod. , 61 (9), 1096-1100.
MacMillan, J. B., & Molinski, T. F. (2000). Herbacic acid, a simple prototype of
5,5,5-trichloroleucine metabolites from the sponge Dysidea herbacea. J. Nat.
Prod. , 63 (1), 155-157.
Macmillan, J. D., & Phaff, H. J. (1978). Yeasts: General Survey. In A. I. Laskin, &
H. A. Lechevalier (Eds.), CRC Handbook of Microbiology (2nd Edition ed., Vol.
II, p. 226). USA: CRC Press, Inc.
Maldonado, M., Giraud, K., & Carmona, C. (2008). Effects of sediment on the
survival of asexually produced sponge recruits. Marine Biology , 154 (4), 631-
641.
Malmvarn, A., Marsh, G., Kautsky, L., Athanasiadou, M., Bergman, A., & Asplund,
L. (2005). Hydroxylated and methoxylated brominated diphenyl ethers in the red
algae Ceramium tenuicorne and blue mussels from the Baltic Sea. Environ. Sci.
Technol. , 39, 2990-2997.
Marin, A., Lopez, M. D., Esteban, M. A., Meseguer, J., Munoz, J., & Fontana, A.
(1998). Anatomical and ultrastructural studies of chemical defence in the sponge
Dysidea fragilis. Marine Biology , 131, 639-645.
Martinez, L. R., Garcia-Rivera, J., & Casadevall, A. (2001). Cryptococcus
neoformans var. neoformans (Serotype D) strains are more susceptible to heat
than C. neoformans var. grubii (Serotype A) strains. Journal of Clinical
Microbiology , 39 (9), 3365-3367.
Mather, P., & Bennett, I. (1992). Phylum Porifera. In P. Mather, & I. Bennett (Eds.),
A Coral Reef Handbook (pp. 35-46). NSW, Australia: Surrey Beatly & Sons PTY
Limited.
McConnell, O. J., Longley, R. E., & Koehn, F. E. (1994). In V. P. Gullo (Ed.), In the
Discovery of Natural Products with Therapeutic Potential (pp. 109-174). Boston:
Butterworth-Heinemann.
McNamara, C. E., Larsen, L., Perry, N. B., Harper, J. L., Berridge, M. V., Chia, E.
W., et al. (2005). Anti-inflammatory sesquiterpene-quinones from the New
Zealand sponge Dysidea cf. cristagalli. J. Nat. Prod. , 68 (9), 1431-1433.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Mehrotra, R. S., & Aneja, K. R. (1990). An Introduction to Mycology. Kurukshetra,
India: Wiley Eastern Limited.
Merck, E. (1978). Dyeing reagents for thin-layer and paper chromatography.
Darmstadt, Germany.
Milkova, T. S., Mikhova, B. P., Nikolov, N. M., Popov, S. S., & Andreev, S. N.
(1992). Two new polyhydroxylated sterols from the sponge Dysidea fragilis. J.
Nat. Prod. , 55 (7), 974-978.
Minale, L., Riccio, R., & Sodano, G. (1974). Avarol, a novel sesquiterpenoid
hydroquinone with a rearranged drimane skeleton from the sponge Dysidea avara.
Tetrahedron Letters , 38, 3401-3404.
Molinski, T. F., & Ireland, C. M. (1988). Dysidazirine, a cytotoxic azacyclopropene
from the marine sponge Dysidea fragilis. J. Org. Chem. , 53, 2103-2105.
Montagnac, A., Martin, M.-T., Debitus, C., & Pais, M. (1996). Drimane
sesquiterpenes from the sponge Dysidea fusca. J. Nat. Prod. , 59, 866-868.
Moran, G. P., Sullivan, D. J., & Coleman, D. C. (2002). Emergence of Non-Candida
albicans Candida species as pathogens. In R. A. Calderone (Ed.), Candida and
Candidiasis (p. 41). Washington D.C., USA: ASM Press.
Munro, M. H., & Blunt, J. W. (2001). Marine Literature Database (MarinLit).
University of Canterbury, New Zealand.
Newman, D. J., & Cragg, G. M. (2004). Marine natural products and related
compounds in clinical and advanced preclinical trials. J. Nat. Prod. , 67 (8), 1216-
1238.
Norton, R. S., Croft, K. D., & Wells, R. J. (1981). Polybrominated oxydiphenol
derivatives from the sponge Dysidea herbacea. Tetrahedron , 37, 2341-2349.
Patil, A. D., Freyer, A. J., Killmer, L., Offen, P., Carte, B., Jurewicz, A. J., et al.
(1997). Frondosins, five new sesquiterpene hydroquinone derivatives with novel
skeletons from the sponge Dysidea frondosa: Inhibitors of interleukin-8 receptors.
Tetrahedron , 53 (14), 5047-5060.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Paul, V. J., Seo, Y., Cho, K. W., Rho, J.-R., Shin, J., & Bergquist, P. R. (1997).
Sesquiterpenoids of the drimane class from a sponge of the genus Dysidea. J. Nat.
Prod. , 60 (11), 1115-1120.
Peng, J., Shen, X., El Sayed, K. A., Dunbar, D. C., Perry, T. L., Wilkins, S. P., et al.
(2003). Marine natural products as prototype agrochemical agents. J. Agric. Food
Chem. , 51 (8), 2246-2252.
Pérez-García, E., Zubía, E., Ortega, M. J., & Carballo, J. L. (2005).
Merosesquiterpenes from two sponges of the genus Dysidea. J. Nat. Prod. , 68
(5), 653-658.
Phaff, H. J., & Macmillan, J. D. (1978). Yeasts: Description of various genera and
species of special interest. In A. I. Laskin, & H. A. Lechevalier (Eds.), CRC
Handbook of Microbiology (2nd Edition ed., Vol. II, p. 295). USA: CRC Press,
Inc.
Piggott, A. M., & Karuso, P. (2005). 9-Hydroxyfurodysinin-O-ethyl Lactone: A new
sesquiterpene isolated from the tropical marine sponge Dysidea arenaria.
Molecules , 10, 1292-1297.
Piña, I. C., Sanders, M. L., & Crews, P. (2003). Puupehenone congeners from an
Indo-Pacific Hyortis sponge. J. Nat. Prod. , 66 (1), 2-6.
Quinoa, E., Adamczeski, M., Crews, P., & Bakus, G. J. (1986). Bengamides, a
heterocyclic antihelmintics from Jaspidae marine sponge. J. Org. Chem. , 51 (23),
4494-4497.
Rhome, R., McQuiston, T., Kechichian, T., Bielawska, A., Hennig, M., Drago, M., et
al. (2007). Biosynthesis and Immunogenicity of glucosylceramide in
Cryptococcus neoformans and other human pathogens. Eukaryotic Cell , 6 (10),
1715-1726.
Ridley, C. P., Bergquist, P. R., Harper, M. K., Faulkner, D. J., Hooper, J. N., &
Haygood, M. G. (2005). Speciation and biosynthetic variation in four
dictyoceratid sponges and their cyanobacterial symbiont, Oscillatoria spongeliae.
Chemistry & Biology , 12, 397-406.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Ridley, C. P., Faulkner, D. J., & Haygood, M. G. (2005). Investigation of
Oscillatoria spongeliae-dominated bacterial communities in four dictyoceratid
sponges. Applied and Environmental Microbiology , 71 (11), 7366-7375.
Rudi, A., Yosief, T., Schleyer, M., & Kashman, Y. (1999). Bilosespens A and B:
Two novel cytotoxic sesterpenes from the marine sponge Dysidea cinerea.
Organic Letters , 1 (3), 471-472.
Sakai, R., Koike, T., Sasaki, M., Shimamoto, K., Oiwa, C., Yano, A., et al. (2001).
Isolation, structure determination, and synthesis of Neodysiherbaine A, a new
excitatory amino acid from a marine sponge. Organic Letters , 3 (10), 1479-1482.
Sakai, R., Oiwa, C., Takaishi, K., Kamiya, H., & Tagawa, M. (1999). Dysibetaine: A
new �,�-disubstituted �-amino acid derivative from the marine sponge Dysidea
herbacea. Tetrahedron Letters , 40, 6941-6944.
Sakai, R., Suzuki, K., Shimamoto, K., & Kamiya, H. (2004). Novel betaines from a
Micronesian sponge Dysidea herbacea. J. Org. Chem. , 69 (4), 1180-1185.
Salomon, C. E., Williams, D. H., & Faulkner, D. J. (1995). New azacyclopropene
derivatives from Dysidea fragilis collected in Pohnpei. J. Nat. Prod. , 58 (9),
1463-1466.
Salva, J., & Faulkner, D. J. (1990). A new brominated diphenyl ether from a
Philippine Dysidea species. J. Nat. Prod. , 53 (3), 757-760.
Sauleau, P., & Bourget-Kondracki, M.-L. (2005). Novel polyhydoxysterols from the
Red Sea marine sponge, Lamellodysidea herbacea. Steroids , 70 (14), 954-959.
Sauleau, P., Retailleau, P., Vacelet, J., & Bourget-Kondracki, M.-L. (2005). New
polychlorinated pyrrolidinones from the Red Sea marine sponge Lamellodysidea
herbacea. Tetrahedron , 61, 955-963.
Schmitz, F. J., Lakshmi, V., Powell, D. R., & van der Helm, D. (1984). Arenarol and
Arenarone: Sesquiterpenoids with rearranged drimane skeletons from the marine
sponge Dysidea arenaria. J. Org. Chem. , 49, 241-244.
Schram, T. J., & Cardellina, J. H. (1985). An unusual �,�-epoxy �-lactone from the
sponge Dysidea etheria. J. Org. Chem. , 50, 4155-4157.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
Schulte, G., Scheuer, P. J., & McConnell, O. J. (1980). Upial, a sesquiterpenoid
bicyclo[3.3.1]nonane aldehyde lactone from the marine sponge Dysidea fragilis.
J. Org. Chem. , 45 (3), 522-524.
Sera, Y., Adachi, K., Nishida, F., & Shizuri, Y. (1999). A new sesquiterpene as an
anti-fouling substance from a Palauan marine sponge Dysidea herbacea. J. Nat.
Prod. , 62 (2), 395-396.
Shen, Y. C., Lu, C. H., Chakraborty, R., & Kuo, Y. H. (2003). Isolation of
sesquiterpenoids from sponge Dysidea avara and chemical modification of avarol
as potential anti-tumor agents. Nat. Prod. Res. , 17 (2), 83-89.
Sionov, E., Roth, D., Sandovsky-Losica, H., Kashman, Y., Rudi, A., Chill, L., et al.
(2005). Antifungal effect and possible mode of activity of a comppound from the
marine sponge Dysidea herbacea. Journal of Infection , 50, 453-460.
Skepper, C. K., & Molinski, T. F. (2008). Long-chain 2H-azirines with
heterogeneous terminal halogenation from the marine sponge Dysidea fragilis. J.
Org. Chem. , 73 (7), 2592-2597.
Skov, M. J., Beck, J. C., de Kater, A. W., & Shopp, G. M. (2007). Nonclinical safety
of Ziconotide: An intrathecal analgesic of a new pharmaceutical class. Journal of
Toxicology , 26 (5), 411-421.
Sladi�, D., & Gaši�, M. J. (2006). Reactivity and biological activity of the marine
sesquiterpene hydroquinone Avarol and related compounds from sponges of the
Dictyoceratida. Molecules , 11, 1-33.
Stevenson, G. (1967). The Biology of Fungi, Bacteria, and Viruses. London, Great
Britain: Edward Arnold (Publishers) Ltd.
Su, J.-Y., Zhong, Y.-L., Zeng, L.-M., Wei, S., Wang, Q.-W., Mak, T. C., et al.
(1993). Three new diketopiperazines from a marine sponge Dysidea fragilis. J.
Nat. Prod. , 56 (4), 637-642.
Suzuki, H., Shindo, K., Ueno, A., Miura, T., Takei, M., Sakakibara, M., et al. (1999).
S1319: A novel �2-Adrenoceptor agonist from a marine sponge Dysidea sp.
Bioorganic & Medicinal Chemistry Letters , 9, 1361-1364.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Taber, W. A., & Taber, R. A. (1978). Ascomycetes (Ascomycotina). In A. I. Laskin,
& H. A. Lechevalier (Eds.), CRC Handbook of Microbiology (2nd Edition ed.,
Vol. II, p. 175). USA: CRC Press, Inc.
Takahashi, M., Dodo, K., Hashimoto, Y., & Shirai, R. (2000). Concise asymmetric
synthesis of dysidiolide. Tetrahedron Letters , 41, 2111-2114.
Thacker, R. W., & Starnes, S. (2003). Host specificity of the symbiotic
cyanobacterium Oscillatoria spongialide in marine sponges Dysidea spp. Mar.
Biol. , 142, 643-648.
Thakur, N. L., & Müller, W. E. (2004). Biotechnological potential of marine
sponges. Current Science , 86 (11), 1506-1512.
Ueda, K., Kadekaru, T., Siwu, E. R., Kita, M., & Uemura, D. (2006).
Haterumadysins A-D, Sesquiterpenes from the Okinawan marine sponge Dysidea
chlorea. J. Nat. Prod. , 69 (7), 1077-1079.
Unson, M. D., & Faulkner, D. J. (1993). Cyanobacterial symbiont biosynthesis of
chlorinated metabolites from Dysidea herbacea (Porifera. Experientia , 49, 349-
353.
Unson, M. D., Holland, N. D., & Faulkner, D. J. (1991). A brominated secondary
metabolite synthesized by the cyanobacterial symbiont of a marine sponge and
accumulation of the crystalline metabolites in the sponge tissue. Mar. Biol. , 19, 1-
11.
Unson, M. D., Rose, C. B., Faulkner, D. J., Brinen, L. S., Steiner, J. R., & Clardy, J.
(1993). New polychlorinated amino acid derivatives from the marine sponge
Dysidea herbacea. J. Org. Chem. , 58 (23), 6336-6343.
Utkina, N. K., Denisenko, V. A., Scholokova, O. V., Virovaya, M. V., Gerasimenko,
A. V., Popov, D. Y., et al. (2001). Spongiadioxins A and B, Two new
polybrominated dibenzo-p-dioxins from an Australian marine sponge Dysidea
dendyi. J. Nat. Prod. , 64 (2), 151-153.
Utkina, N. K., Denisenko, V. A., Virovaya, M. V., Scholokova, O. V., & Prokof'eva,
N. G. (2002). Two New Minor Polybrominated dibenzo-p-dioxins from the
Marine Sponge Dysidea dendyi. J. Nat. Prod. , 65 (8), 1213-1215.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Utkina, N. K., Kazantseva, M. V., & Denisenko, V. A. (1987). Brominated Diphenyl
Ethers from the Marine Sponge Dysidea fragilis. Khimiya Prirodnykh Soedinenii ,
4, 603-605.
Vallim, M. A., Fernandes, L., & Alspaugh, J. A. (2004). The RAM1 gene encoding a
protein-farnesyltransferase �-subunit homologue is essential in Cryptococcus
neoformans. Microbiology , 150, 1925-1935.
Van Soest, R., Boury-Esnault, N., Janussen, D., & Hooper, J. (2005). Retrieved May
17, 2008, from World Porifera Database: http://www.marinespecies.org/porifera
Venkateswarlu, Y., Biabani, M. A., Reddy, M. V., Chavakula, R., & Rao, J. V.
(1994). A new sesquiterpene from the Andaman sponge, Dysidea herbacea. J.
Nat. Prod. , 57 (6), 827-828.
Vetters, W., Stoll, E., Garson, M. J., Fahey, S. J., Gaus, C., & Muller, J. F. (2002).
Sponge halogenated natural products found at parts-per-million levels in marine
animals. Environmental Toxicology and Chemistry , 10 (21), 2014-2019.
Voet, D., & Voet, J. G. (2004). Biochemistry (3rd ed.). USA: John Wiley and Sons,
Inc.
Voinov, V. G., Elkin, Y. N., Kuznetsova, T. A., Maltsev, I. I., Mikhailov, V. V., &
Sasunkevich, V. A. (1991). Use of mass spectrometry for the detection and
identification of bromine-containing diphenyl ethers. J. Chromatogr. , 586, 360-
362.
Walker, R. P., & Faulkner, D. J. (1981). Diterpenes from the sponge Dysidea amblia.
J. Org. Chem. , 46 (6), 1098-1102.
Walker, R. P., Rosser, R. M., & Faulkner, D. J. (1984). Two new metabolites of the
sponge Dysidea amblia and revision of the structure of Ambliol B. J. Org. Chem.
, 49 (26), 5160-5163.
Walsh, D. (2004). Take two clams and call me in the morning. Proceedings of the
United States Naval Institute, 130(11), p. 90.
Wang, Y., & Casadevall, A. (1996). Susceptibility of melanized and non-melanized
Cryptococcus neoformans to the melanin-binding compounds Trifluoperazine and
Chloroquine. Antimicrobial Agents and Chemotherapy , 40 (3), 541-545.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
West, R. R., & Cardellina, J. H. (1988). Three new polyhydroxylated sterols with the
5�-configuration from the sponge Dysidea etheria. J. Org. Chem. , 54, 3234-3236.
Xu, Y.-M., Johnson, R. K., & Hecht, S. M. (2005). Polybrominated diphenyl ethers
from a sponge of the Dysidea genus that inhibit Tie2 kinase. Bioorganic &
Medicinal Chemistry , 13, 657-659.
Zhang, H., Skildum, A., Stromquist, E., Rose-Hellekant, T., & Chang, L. C. (2007).
Bioactive polybrominated diphenyl ethers from the marine sponge Dysidea sp. J.
Nat. Prod. , (Article in Press).

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2 �������/�
2�� �&&'� �=��B����� ��&'!�'����$ �' �%���)'3�!�*��!�''���#�
# Ext. Code Species Location
Freeze-dried
Sponge (g)
Mass of
Ext. C (g)
% Yield
of Ext. C
Mass of
Ext. B (g)
Description of Ext. C
1 R2252C L. herbacea New
Caledonia (NC)
16.174 0.6989 4.32 1.4202 Green;
Gummy; Semi-solid;
Sticky substance
2 R2256C L. herbacea NC 20.943 0.9612 4.59 1.9702
3 R2259C L. herbacea NC 20.326 1.1974 5.89 2.8710
4 R2261C L. herbacea NC 20.649 0.7715 3.74 1.7336
5 R2262C L.
herbacea? NC
20.320 0.8606 4.23 1.1297
6 R2265C L. herbacea NC 20.536 1.1865 5.78 1.7566
7 R2267C L. herbacea NC 21.186 0.6942 3.28 0.6137
8 R2269C L. herbacea NC 20.664 1.0117 4.90 1.7546
9* R2270C L. herbacea NC
10 R2272C L. herbacea NC 20.421 0.7392 3.62 0.9323
11* R2274C L. herbacea NC
12* R2278C L. herbacea NC
13* R2279C L. herbacea NC
14* R2280C L. herbacea NC
15 MAU-02-
Dh1 L. herbacea Moorea
2.0260 0.1237 6.11 -
16* R2247bisC D. arenaria NC Yellow-brown;
Gummy; Semi-solid;
Sticky substance.
17 R2254C D. arenaria NC 20.824 0.1850 0.89 0.9106
18 R2257C D. arenaria NC 21.664 0.2633 1.22 1.1141
19* R2260C D. arenaria NC
20 R2261bisC D. arenaria NC 21.844 0.3226 1.48 2.0333
21* R2263C D. arenaria NC
22* R2264C D. arenaria NC
23 R2268C D. arenaria NC 21.321 0.2012 0.94 1.2007
24* R2271C D. arenaria NC
25 R2082C D. avara NC 1.0006 0.0316 3.16
Gummy; Yellow-brown;
sticky; semi-solid extract.
*Extracts obtained from IRD Nouméa.

Secondary Metabolites Co
�
�
2� �&&'� �=�
�$34'��
omposition and Geographical Distribution of Marine Sponge
B� �$� '� ��� '�'�3��'� )�*�#'�����
es of the Genus Dysidea
����
��� &���'��� �� �

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2�, �&&'� �=�,B����(�� '���*���%��3&�$� ���A� A��� �,�%��3����
���� �� �
2�,�� �&&'� �=� ,�B� ���(�� &��%�*'�� �%� ��� ���� �� � %��� �3&�$� � ��
������� !�"����#�$�

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2�,� �&&'� �=�,4B��<���� � <���(�����%�*'���%��3&�$� � ��
&��$���+���*��(*7',��������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
&��$���+���*���&*7',��������'(6������������*����
&��$���+���F�(�9&��������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
O
OHOH
Br
Br
Br
Br
Br
Br1
2
34
5
61'2'
3'
4'5'
6'
HH
+ !� ����2� �5�5�5�*�����8����*�*��15�1*+�8����*�1*!�+��;�$!���;��$!�����
&��$���+���F�('B&��������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2�,�, �&&'� �=�,!B����(�����%�*'��%��3&�$� �,�
&��$���+���F��(*7',��������'(6������������*����
&��$���+�����<���$$���
&��$���+������<��$$���
&��$���+�����<���$$���
&��$���+�����<���$$���
&��$���+������<��$$���

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
&��$���+���F���&*7',��������'(6������������*����

Seco
ndar
y M
etab
olit
es C
ompo
sitio
n an
d G
eogr
aphi
cal D
istr
ibut
ion
of M
arin
e Sp
onge
s of
the
Gen
us D
ysid
ea
�
��
���
2�.
�&&'� �=�.B��(���)�%���%����**�!�3
&�$�
��� '���%�' ���
��)'���'* ���4����' �%����)'�%�$��!�3
&�$�
��
2�.��
�&&
'� �=�.�
B����+��, ��(���)�%���%����**�!�3
&�$�
��� '���%�' �
Com
poun
d 1
2 3
4 6
7 8
C#
H
C
H
C
H
C
H
C
H
C
H
C
H
C
1
150.
49
15
1.54
151.
50
14
5.17
144.
88
14
5.13
2
140.
62
13
9.88
139.
78
14
7.55
146.
75
14
7.24
3 -
-
119.
31
11
7.54
117.
65
6.62
11
7.15
6.
73
116.
71
6.71
11
7.19
4 7.
36
-
121.
60
7.64
12
8.54
7.
65
128.
71
11
1.52
111.
34
11
1.62
5 -
-
127.
22
12
4.01
124.
05
7.37
7 12
9.91
7.
38
129.
76
7.39
13
0.11
6 7.
20
-
116.
48
11
5.47
115.
80
11
1.90
111.
69
11
2.00
1’
14
5.34
145.
28
14
7.20
155.
29
15
1.63
154.
00
2’
14
6.89
147.
12
15
2.64
141.
14
14
0.70
142.
16
3’
6.80
6.85
11
7.12
6.
74
117.
02
6.82
11
8.18
118.
23
12
3.02
4’
-
- 11
1.74
111.
70
11
7.62
7.
49
128.
31
12
3.05
123.
52
5’
7.46
7.41
13
0.40
7.
40
130.
26
7.48
13
0.38
120.
71
12
2.64
119.
52
6’
-
- 11
2.19
112.
15
12
0.13
7.
425
117.
76
7.49
12
2.35
7.
64
119.
02
-OH
5.
08
6.
01
-OH
-
6.
04
-OM
e 3.
81
-
4.
00
61.7
6 3.
87
3.
87

Seco
ndar
y M
etab
olit
es C
ompo
sitio
n an
d G
eogr
aphi
cal D
istr
ibut
ion
of M
arin
e Sp
onge
s of
the
Gen
us D
ysid
ea
�
��
���
2�.�
��'* ���4����' �%����)'����*��' �!�3
&�$�
��
+ !
� ��
�'
�!��
1!
���
�
��%�
��#�
���
���!
���
3��
��
�,� �
����
�!��
�4�
���
5��
#���
�� �
� �
+ !
� ��
�
���5�5�5�*�����8
����*�*
��15�
1*+�8����*�1*
!�+��;
�$!�
��;��$!�
����
���<��
�<��
�<��G�
2!�������������
'$�<�A
����
�F����
<��� &
�
���5�*+�8���
�*�*��15�
1*+�8���
�*�1*
���!�
;�$!
���;
��$!
�����
����
<��
�<�
�<�
�G�
'�;���������:���!����
���$�
��+�?�
&����������
������:*
:!�������
$���
+��
��5�*+�8���
�*�*��15�
15�1*
���8����*�1*
���!�
;�$!
���;
��$!
�����
��5�*+�8���
�*�*��15�
15�1*
���8����*�1*
!�+��;
�$!�
��;��$!�
����
����
<��
���
�<��
�G�
&����������
������:*
:!�������
$���
+��

Secondary Metabolites Co
�
�
%�& �&&'� �=�0B
[HPLC Condition 2 +
RT: 0.00 - 34.99
0 50
10
20
30
40
50
60
70
80
90
100
Relative Abun
danc
e
0
10
20
30
40
50
60
70
80
90
100
Relative Abso
rban
ce
2.45
2.68
1.48
5.07
6.308.5
2.52 2.75 96.67
%<���������-�����.%4�
%����������-�����'����
%<����������"��������
%<����������*��(7'
omposition and Geographical Distribution of Marine Sponge
B��&'!���*������-����� � ��" � �
+ Finnigan APCI-MS; 1HNMR at 300 MHz in
10 15 20 25Time (min)
24.32
15.90
20.8315.4712.059.9318.88
24.7017.9557
25.85
13.18
13.52
21.7420.52
22.2318.0215.15
22.7115.3624.68.15 12.71
�������
�����
',��������'(6�
es of the Genus Dysidea
����
n CDCl3]
�
�
�
30
5 29.35
29.60 30.19

Secondary Metabolites Co
�
�
%�% �&&'� �=�2B
���������� � ��������
RT: 0.00 - 34.98
0 50
10
20
30
40
50
60
70
80
90
100
Relativ
e Abun
danc
e
0
10
20
30
40
50
60
70
80
90
100
Relativ
e Abso
rban
ce
2.62
3.42
7.355.83
2.71 3.51
5.952.07
%<��#-����
%<��#��-�����'
%<��#��"������
%<��#����*��(7'
omposition and Geographical Distribution of Marine Sponge
B��&'!���*�����B����� � ' � �
� �� ��!��"#$%&���'$(���)**�$�+� �����
10 15 20 25Time (min)
15.85
18.82 24.3223.0519.92
14.43 17.5724.72
13.3810.18
9.485
21.75
22.26
20.54 22.7622.91
19.8918.00 23.35
14.9213.48 24.90
11.36
#������.%4������
��'���������
����
',��������'(6�
es of the Genus Dysidea
����
��),�
�
�
30
30.23
26.1429.10 30.94

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2�7 �&&'� �=�7B� �3&��������%�������� �� A���� ��" � ��� ����
' � ��
TLC profile of L. herbacea, D. arenaria and D. avara after visualization using
Vanillin Phosphoric Acid:
HPLC profile comparison of the 3 species of Dysidea studied (HPLC Condition 2;
UV absorption at � = 254 nm):
AU
0.00
0.20
0.40
0.60
0.80
1.00
Minutes2.00 4.00 6.00 8.00 10.00 12.00 14.00 16.00 18.00 20.00 22.00 24.00 26.00 28.00 30.00
�<�������
������� �
%<�-��

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
1HNMR of the 3 species investigated (at 300 MHz; in CDCl3):

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
2�C �&&'� �=� CB� �&'!�3'��� ��$ �' � %��� �!�*�#�!�*� �� �
�'�#��&)�!�*�!�3&�������
Station Sample Area of Collection
Photograph of Sample Description
ST720 R2252
L. herbacea
New Caledonia
(22°32.736S 166°26.588E)
Colour - pale
green; Encrusting on hard surface;
Finger-like projections in
some regions, no out-growth on the
other.
ST1048 R2256
L. herbacea
New Caledonia
(22°41.566S 166°38.704E)
Green, lamellate projections from
an encrusting base; in competition for space with other sessile marine
organisms.
ST1052 R2261
L. herbacea
New Caledonia
(22°21.249S 166°14.783E)
Green, lamellate to lobate projections from an encrusting
base; in competition for space with other sessile marine
organisms.
ST1053 R2262
L. herbacea
New Caledonia
(22°16.456S 166°14.311E)
Green, lamellate projections from
an encrusting base; little competition, therefore upward
growth of the leaf-like projections.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
ST1055 R2265
L. herbacea
New Caledonia
(22°01.865S 165°57.579E)
Green sponge with
encrusting base attached to rocky
surface, small projections; found in the middle of
many sessile invertebrates.
ST956 R2267
L. herbacea
New Caledonia
(21°51.695S 165°43.429E)
Green sponge with encrusting base
attached to rocky surface, small
projections; found in the middle of
many sessile invertebrates.
ST1059 R2269
L. herbacea
New Caledonia
(22°10.980S 166°06.066E)
Green sponge with encrusting base
attached to rocky surface, small
projections; found in the middle of
many sessile invertebrates;
small colony seen.
ST254 R2272
L. herbacea
New Caledonia
(22°17.013S 166°11.035E)
Green sponge with encrusting base
attached to rocky surface, small
projections; found in the middle of
many sessile invertebrates;
small colony seen.
ST1060 R2270
L. herbacea
New Caledonia
(22°23.201S 166°17.065E)
Green sponge with encrusting base
attached to rocky surface, small
projections; found in the crevice between two rocks; small colony seen.
Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
ST1132 R2274
L. herbacea
New Caledonia
(22°19.350S 167°02.027E)
Green sponge with
encrusting base attached to flat
rocky surface, not so many
projections seen; small colony.
ST1135 R2278
L. herbacea
New Caledonia
(22°06.832S 167°00.321E)
Blue-green sponge
with encrusting base attached to
flat rocky surface, not so many
projections visible; small colony.
ST1127 R2280
L. herbacea
New Caledonia
(21°32.777S 165°12.996E)
No Photo Available
ST1050 R2259
L. herbacea
New Caledonia
(22°23.295S 166°47.301E)
Green sponge with lamellate/lobate
projections; surrounded by
sediment.
ST1136 R2279
L. herbacea
New Caledonia
(22°08.287S 166°56.955E)
Blue-green, encrusting sponge; large but generally flat colony, very
few lamellate projections visible;
occupying large surface area of a rock containing other organisms.
ST613 R2271bis
L. herbacea
Vanuatu (13°30.910S
167°21.100E)
167°21.100E)
No Photo Available

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
ST840 R3147
L. herbacea
Solomon Is. (09°11.055S
160°11.982E)
Blue-grey sponge with lamellate
projections; small colony encrusted
on a rocky surface; habitat a little
darker than that for New Caledonian
samples.
FJ4 R3241
L. herbacea
Fiji Is. (16°18.847S
178°54.595E)
Blue-grey sponge with lamellate
projections (purple when out of the water); colony encrusted on a
rocky surface with coral and algae growing on it.
MAU-02-Dh1
L. herbacea
Moorea Is.
Found in lagoon
with clear water at a depth of 1 - 1.5 m; Encrusting on hard corals with a
big lamellate colony; in close proximity of two cyanobacteria,
Anabaena torulosa and an unidentified
species.
MAU-02-Dh2
L. herbacea
Moorea Is. No Photo Available
Found in lagoon with clear water at a depth of 1 - 1.5 m; Growing close to hard corals; No
algae seen growing around the samples.
MAU-02-Dh3
L. herbacea
Moorea Is.
Found in lagoon with clear water at a depth of 1 - 1.5 m; Encrusting on hard corals with a
big lamellate colony; in close proximity of two cyanobacteria,
Anabaena torulosa.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
MAY-02-Dh1
L. herbacea
Mayotte Is. No Photo Available
MAY-02-Dh2
L. herbacea
Mayotte Is. No Photo Available
ST124 R2247bis
D. arenaria
New Caledonia
(22°20.611S 166°25.970E)
No Photo Available
ST534 R2254
D. arenaria
New Caledonia
(22°20.700S 166°20.000E)
Grey lobate colony clustered together (only one colony seen); growing on sandy substrate;
surrounded by sea-weed and algae.
ST1049 R2257
D. arenaria
New Caledonia
(22°31.008S 166°36.212E)
Grey lobate colony clustered together (only one colony seen); growing on sandy substrate;
surrounded by sea-weed and algae.
ST1051 R2260
D. arenaria
New Caledonia
(22°19.699S 166°23.463E)
Grey lobate colony clustered together (only one colony seen); growing on sandy substrate;
surrounded by sea-weed and algae.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
ST1054 R2263
D. arenaria
New Caledonia
(22°18.936S 166°21.176E)
Grey lobate colony clustered together (only one colony seen); growing on sandy substrate;
surrounded by sea-weed and algae; sedimentation
seen.
ST1053 R2261bis
D. arenaria
New Caledonia
(22°16.456S 166°14.311E)
Grey lobate colony clustered together (only one colony seen); growing on sandy substrate;
surrounded by sea-weed and algae; sedimentation
seen.
ST1057 R2264
D. arenaria
New Caledonia
(21°51.971S 165°45.628E)
Grey lobate colony clustered together (only one colony seen); growing on sandy substrate;
surrounded by sea-weed and algae; sedimentation
seen.
ST1058 R2268
D. arenaria
New Caledonia
(22°07.065S 166°06.556E)
No Photo Available
ST1061 R2271
D. arenaria
New Caledonia
(22°15.529S 166°20.155E)
Grey lobate colony clustered together (only one colony seen - small in
size); growing on sandy substrate;
surrounded by sea-weed and algae;
sedimentation seen slightly.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
FJ18 R3314
D. arenaria
Fiji Is. (16°06.762S
179°11.264W)
Pink in color; growing in the
middle of a group of sea-weeds; has
the appearance of a perforated ball (not
lobate like the other New Caledonian samples).
ST612 R2082
D. avara
New Caledonia
(20°32.930S 167°10.790E)
No Photo Available
FJ1 R3203 D. cf. avara
Fiji Is. (16°46.600S 178.24.344E)
Red in color when outside water; rigid-looking
stems like that of hard corals.
ST823 R3033
Dysidea 4177
Solomon Is. (08°47.266S
158°14.228E)
Purple in color; growth pattern as
seen in hard corals but scattered;
found on a rocky outgrowth;
surrounged by algae and sea-
ferns.
ST846 R3161
Dysidea 4177
Solomon Is. (08°58.067S
160°44.682E)
Purple in color; growth pattern as
seen in hard corals with finger-like
projections arising from a common
stem; dark purple in color when out
of the water.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
ST840 R3141
Dysidea sp.
Solomon Is. (09°11.055S
160°11.982E)
Maroon in color, and leaf-like appearance;
growing with other sea-weeds and
algae.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2�D �&&'� �=� DB� �'���*' � 3�&� �%� �)'� 2� ��3&*��#� ���'�� �� � �)'�
�&&��=�3��'� �����!'�4'�5''���)'3�

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
Site 1 Site 2 Distance
(km)
NC
Vanuatu 836.89 Solomon 1427.4 Fiji 1420.71 Moorea 4471.05 Mayotte 12115.62
Vanuatu
Solomon 871.29 Fiji 1226.96 Moorea 4488.15 Mayotte 12655.56
Solomon Fiji 2091.66 Moorea 5277.34 Mayotte 12164.15
Fiji Moorea 3237.39 Mayotte 13584.88
Moorea Mayotte 15787.65

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
2��E �&&'� �=� �EB� �&'!���*� '���*�� �%� �,�.7� �� � �, .��
%��!������
2��E�� B��&&'� �=��E�B��,�.7�������������� ������� ������������
���������������������������� ����!"��"�#������������ ���
���$�%�
,���&�F� �(*7',���� ����'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
,���&�F���&*7',��������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2��E� B��&&'� �=��E4B��, .��������������� ������� ������������
������������������� ������$�%�
*Note: The fractions obtained contained a mixture of compounds; hence pure
compounds could not be isolated. However, fractionation did help to confirm the
major products present in the sample.

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2��� �&&'� �=� ��B� (�����'� ��3&*'� :(��<E <�)�;� -� ���$!�$�'�
'�'�3��������
������������� ������� ������������ ���������������� ����!"��"�
#%�
'4H�*�� F� �(*7',� ���� &��$���+�� ����+���������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
'4H�*��F� ��&*7',�����&��$���+������+���������'(6������������*����
'4H�*��F(�9&�����&��$���+������+���������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
O
OHO
12
34
5
61'2'
3'
4'5'
6'
BrBr
Br
Br
H
H
HH
+ !� ���62� �5�*+�8����*�*��15�1*+�8����*�1*���!�;�$!���;��$!�����
�
O
OHO
12
34
5
61'2'
3'
4'5'
6'
BrBr
Br
Br
Br
H
HH
+ !� ����2� �5�*+�8����*�*��15�15�1*���8����*�1*���!�;�$!���;��$!�����
�
'4H�*��F�('B&�����&��$���+������+���������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
'4H�*��F��(*7',�����&��$���+���������'(6������������*����
'4H�*��F���&*7',�����&��$���+���������'(6������������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
'4H�*�� F� (�9&� ����&��$���+� � ��� ����'(6������������*����
'4H�*�� F� ('B&� ���� ����&��$���+� � ��� ���� '(6� �����������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
O
OHOH
12
34
5
61'2'
3'
4'5'
6'
BrBr
Br
Br
Br
HH
H
+ !� ���72� �5�*+�8����*�*��15�15�1*���8����*�1*!�+��;�$!���;��$!����
'4H�*�� F� ('B&� ���� ����&��$���+� � ��� ���� '(6� �����������*����

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
��
2�� �&&'� �=�� B��3&��������%�)'3�!�*����%�*'���%���� ��" � �
�� ���� ' � �%��3��'5��*' ������� ���F��
2�� �� �&&'� �=� � �B����� � ��" � � ������ ������� � �� &��� '!���
��������������� ������$�%�

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
2�� � �&&'� �=� � 4B� ���� '�� �6���� ������ ������� � �� &��� '!���
��������������� ������$�%�

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
%�(� �&&'� �=� �,B� ����� $�' � %��� ���$!�$�'� '�'�3�������� �%� �)'�
4��3����' �!�3&�$� ��������� �)(**�
������������� ������� ������������������������������� ������$�%�

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
�

Secondary Metabolites Composition and Geographical Distribution of Marine Sponges of the Genus Dysidea
�
��
���
1HNMR chemical shifts obtained for Compounds 9 and 10:
Proton � Compd. 9 (ppm)
J for Compd. 9 (Hz)
� Lit. (ppm)*
� Compd. 10 (ppm)
J for Compd. 10 (Hz)
� Lit. (ppm)**
H4 7.36 2.2 7.37 - - -
H6 7.23 2.1 7.12 7.46 - 7.43
H3’ 7.80 2.3 7.85 7.79 2.9 7.84
H5’ 7.32 2.4, 8.8 7.36 7.30 7.35
H6’ 6.47 8.8 6.42 6.44 8.3 6.51
* (Xu, Johnson, & Hecht, 2005)
** (Fu & Schmitz, 1996)


















![Sec. Metabolites [Compatibility Mode]](https://img.dokumen.tips/doc/110x75/549daca0b4795956208b458f/sec-metabolites-compatibility-mode.jpg)










