Embed Size (px)
Citation preview

Plant Physiol. (1974) 54, 76-81
Seasonal Patterns of Acid Metabolism and Gas Exchange inOpuntia basilaris'
Received for publication January 18, 1974 and in revised form March 20, 1974
STAN R. SZAREK AND IRWIN P. TINGDepartment of Biology anid the Philip L. Boyd Deep Canyoni Deser-t Research Cenite,-, Uniiversity of California,Riverside, Californiiia 92502
ABSTRACT
Acid metabolism and gas exchange studies were conductedin situ on the cactus Opuntia basilaris Engelm. and Bigel. Apattern of significant seasonal variation was evident. The pat-tern was controlled by rainfall, which significantly influencedplant water potentials, total gas transfer resistances, and noc-turnal organic acid synthesis. In winter and early spring, whenplant water stress was mild, stomatal and mesophyll resistancesremained low, permitting enhanced nocturnal assimilation of"CO2. The day/night accumulation of acidity was large duringthese seasons. In summer and fall, plant water stress was mod-erate, although soil water stress was severe. The nocturnal as-similation of "CO2 was very low during these seasons, even instems with open stomata, indicating large mesophyll resistancesrestricting exogenous gas incorporation. The day night ac-cumulation of acidity was reduced, and a low level of acidmetabolism persisted throughout this period. The rapid re-sponse to a midsummer rainfall emphasizes the importance ofplant water potential as a parameter controlling over-all meta-bolic activity. The seasonal variations of acid metabolism andgas exchange significantly influenced the efficiency of wateruse and carbon dioxide assimilation. Periods of maximal effici-ency followed rainfall throughout the course of the year.
Historically, succulents have been considered a group ofplants with morphological and anatomical similarities, and re-cently this classification has been expanded to include physio-logical similarities (7, 16). Some of these plants demonstrate acircadian fluctuation of organic acids typical of CAM,2 andthe gas exchange of CAM plants has been proposed to be anadaptation to dry, arid environments (14). Numerous researchreports have appeared concerning the physiology of theseplants, although only a few studies have investigated CAMplants in their native environment. Daily patterns of acidmetabolism and gas exchange have been investigated in pine-
'This research was supported by National Science FoundationGrant GB 15886, through the United States International Biologi-cal Program (Desert Biome) and the University of CaliforniaChancellor's Patent Fund.
2 Abbreviations: CAM: Crassulacean acid metabolism; r,: sto-matal resistance to water vapor; r,': mesophyll resistance to C02;-: water potential; TR: transpiration ratio; CAE: carbon assimila-tion efficiency; C3: reductive pentose phosphate cycle; C4: dicar-boxylic acid cycle.
apple under agricultural growing conditions (1), and earlierwork with pineapple centered exclusively around diurnal pat-terns of gas exchange (8). Seasonal patterns of gas exchangewere recently investigated in Dlidleya farinosa growing in itsnative coastal environment (2), and some preliminary informa-tion has been reported on seasonal patterns of acid metabolismin Bryophyllutin calycinum7l (4) and Sedumi praealtlwnz (3). Toour knowledge there have not been any reports investigatingseasonal patterns both of acid metabolism and of gas exchangein any CAM plant.The following study was initiated to investigate concurrently
these two parameters in a desert succulent, growing in an aridenvironment where severe soil water stress occurs during theyear. Opuntia basilaris was selected for this study because it isa typical CAM plant (20), and it is a wide ranging species ina large genus important in the arid soLthwestern United States.
MATERIALS AND METHODS
Gas exchange measurements consisted of measuring stoma-tal diffusion resistances and rates of CO2 assimilation. Stomataldiffusion resistances were determined with a diffusion resistancehygrometer, similar in design to those constructed by vanBavel, Nakayama, and Ehrler (24). The instrument was cali-brated regularly, from 0.2 to 0.3 full scale, with evaporativediscs constructed to lose water at a known, constant diffusionalresistance. Direct transpiration measurements on a single stemcould be obtained within a few minutes. The measurementswere conducted during the nighttime and early morning whentemperature differences between the hygrometer cup and theevaporating surface were lowest.
Carbon dioxide-'4C uptake was measured with a portable in-strument constructed following the design by Shimshi (18).Air containing "CO2 was applied to stem surfaces for 20 secthrough a 1-cm diameter leaf chamber. A cylinder of tissuewith the exposed stem surface was immediately removed witha cork borer (8 mm diameter). frozen, and stored under DryIce until returned to the laboratory. The radioactive sampleswere subsequently lyophilized and combusted in oxygen follow-ing a modified Schoniger technique (15). Radioactive gas as-similated by the stem tissue was measured by liquid scintillationcounting. The specific radioactivity of the radioactive gas wasinitially determined by infrared gas analysis of the 12C-fraction,and liquid scintillation counting of the "C-fraction. Ambientconcentrations of C02, i.e., 300 [ul/liter, were utilized through-out the year. In all cases, the same location on the stem surfaceof the previous stomatal diffusion measurement was exposedto radioactive gas. Much of the nocturnal sampling was re-stricted to the predawn hours.
Plant and soil water potentials were determined with a com-mercially available thermocouple psychrometer (Wescor, Inc.).
76 www.plantphysiol.orgon December 26, 2018 - Published by Downloaded from
Copyright © 1974 American Society of Plant Biologists. All rights reserved.
77SEASONAL PATTERNS OF ACID METABOLISM
The soil psychrometer probes were permanently placed at thestudy area at soil depths of 2, 5, 10, 15, and 30 cm, with thesensor axes positioned parallel to the soil surface. Plant 1values were also determined with the ceramic-capped probes.At the beginning of the sampling period sensors were placedin the center of three separate stems, through an incision in theepidermis. This opening was sealed with a viscous grease andallowed to equilibrate overnight. In all cases, the plant i wasdetermined shortly after sunrise.The titratable acidity of the stem tissue was determined uti-
lizing a technique modified from Sideris, Young, and Chun(19). All of the samples were titrated to a pH 6.4 end pointwith 0.01 N NaOH. Experimental results from our lab indicatethat 95% of the organic acids contributing to the circadianacid fluctuation are neutralized at pH 6.4. Boundary andstomatal resistances to CO2 were calculated from the transpira-tion resistance measurements by multiplying the latter valuesby the ratio of the diffusion coefficients of water and CO2 inair, i.e., 1.6 for D.H.o/Dc,.. Total gas diffusion resistances toCO2 transfer and assimilation were calculated from the ob-served rates of carbon assimilation, utilizing a maximal CO2concentration gradient of 300 pl/liter. Mesophyll resistancesto CO2 were calculated by subtracting the boundary and stoma-tal resistances to CO2 from the total resistances to CO2 transferand assimilation. Important microclimatic measurements weredetermined with portable thermocouple sensors during thecourse of the field work.
Only plants of 0. basilaris Engelm. and Bigel. were includedin this study. This species, a low-growing spineless platyopuntia,occupies sandy, gravelly, or rocky soils from sea level to 1500m. Plants of this species range throughout the Colorado andMojave Deserts of California and also occur in Nevada, Utah,Arizona, and northern Sonora in Mexico. The study area wasat the University of California's Philip L. Boyd Deep CanyonDesert Research Center, near Palm Desert, California (116007' W, 330 30' N). The primary climatic characteristics are:(a) scant, erratic precipitation, averaging 9.8 cm annually; (b)high summer daytime temperatures, generally less than 46 C;(c) few freezing nights during the winter, with minimal tem-peratures generally greater than 2 C; and (d) high solar radia-tion throughout most of the year, with occasional cloud cover.The elevation of the study area is 305 m and the vegetationconsists of numerous species of succulent and woody peren-nials.
RESULTS
Environmental Conditions. A circadian fluctuation of acidaccumulation and depletion is typical of all CAM plants, al-though the amplitude of the day/night acidity fluctuation isdependent upon effects of environmental conditions. The re-sults of this study indicate that precipitation is the major en-vironmental condition influencing the seasonal acid metabo-lism of 0. basilaris. The effect of rainfall is upon plant b,stomatal regulation, and CO2 assimilation capacity, such thatatmospheric CO2 is readily available for assimilation and en-hancement of acid accumulation can occur.
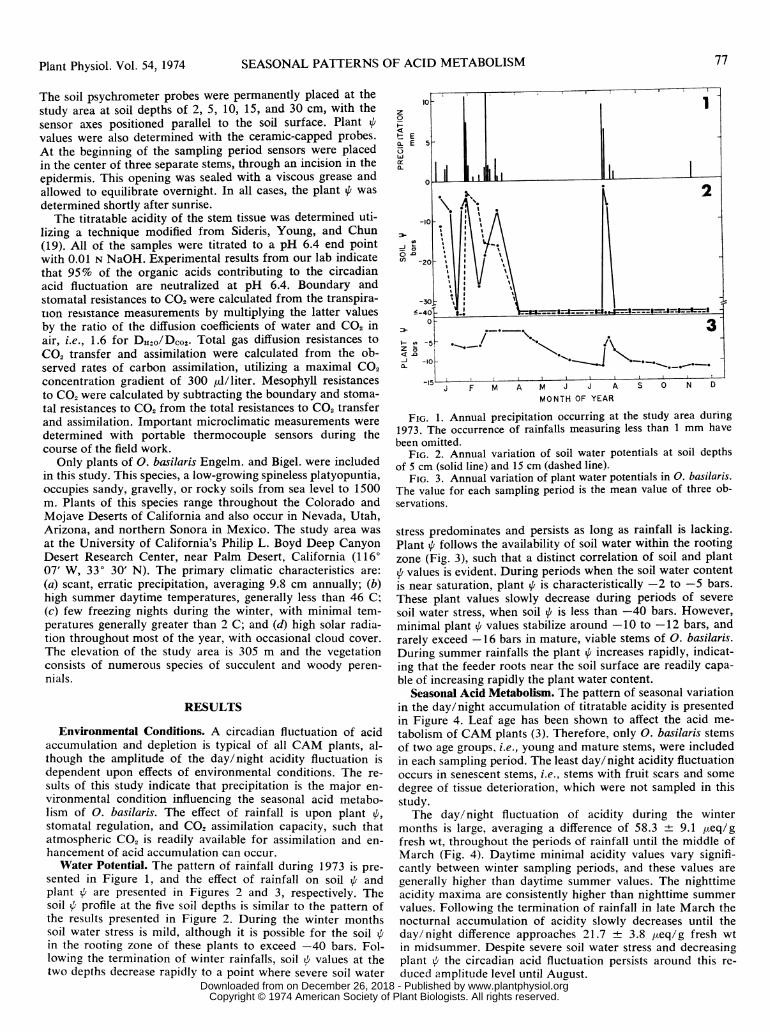
Water Potential. The pattern of rainfall during 1973 is pre-sented in Figure 1, and the effect of rainfall on soil a andplant a are presented in Figures 2 and 3, respectively. Thesoil V profile at the five soil depths is similar to the pattern ofthe results presented in Figure 2. During the winter monthssoil water stress is mild, although it is possible for the soil iin the rooting zone of these plants to exceed -40 bars. Fol-lowing the termination of winter rainfalls, soil / values at thetwo depths decrease rapidly to a point where severe soil water
z0
a--
a:a.
:-ja:U,
F~~~~~~~~-
\,;5L . \ |
5;*
J F M A M J J A S 0 N DMONTH OF YEAR
FIG. 1. Annual precipitation occurring at the study area during1973. The occurrence of rainfalls measuring less than 1 mm havebeen omitted.
FIG. 2. Annual variation of soil water potentials at soil depthsof 5 cm (solid line) and 15 cm (dashed line).
FIG. 3. Annual variation of plant water potentials in 0. basilaris.The value for each sampling period is the mean value of three ob-servations.
stress predominates and persists as long as rainfall is lacking.Plant f follows the availability of soil water within the rootingzone (Fig. 3), such that a distinct correlation of soil and plant
+ values is evident. During periods when the soil water contentis near saturation, plant i is characteristically -2 to -5 bars.These plant values slowly decrease during periods of severesoil water stress, when soil i is less than -40 bars. However,minimal plant i values stabilize around -10 to -12 bars, andrarely exceed -16 bars in mature, viable stems of 0. basilaris.During summer rainfalls the plant i increases rapidly, indicat-ing that the feeder roots near the soil surface are readily capa-ble of increasing rapidly the plant water content.
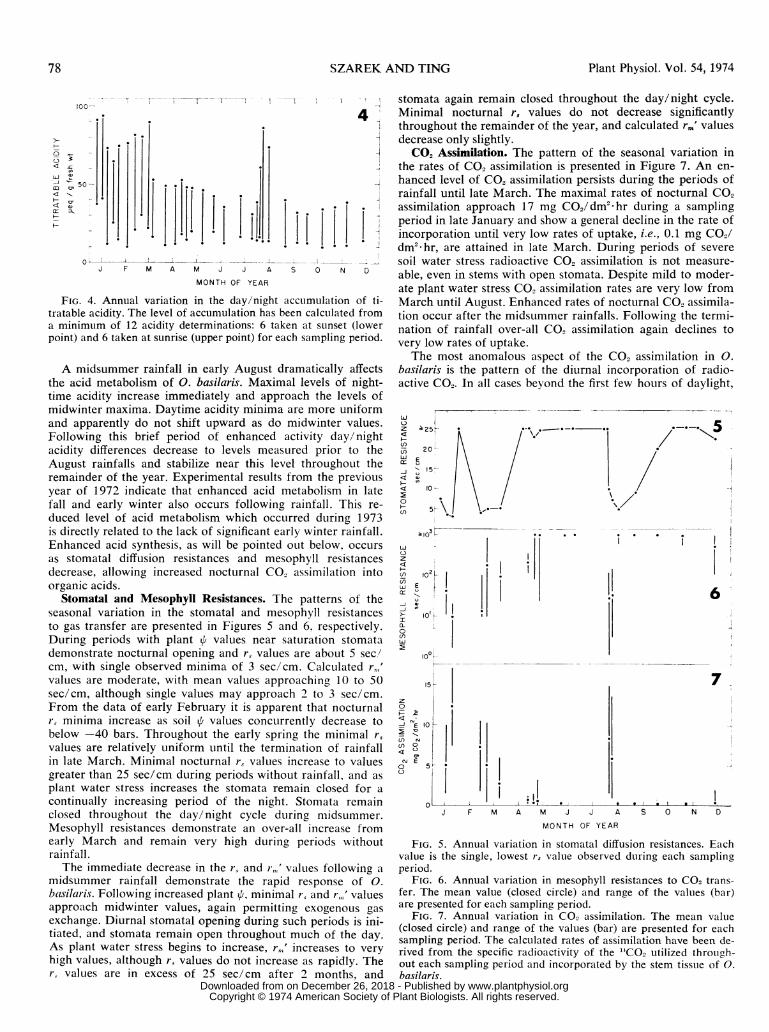
Seasonal Acid Metabolism. The pattern of seasonal variationin the day/night accumulation of titratable acidity is presentedin Figure 4. Leaf age has been shown to affect the acid me-tabolism of CAM plants (3). Therefore, only 0. basilaris stemsof two age groups, i.e., young and mature stems, were includedin each sampling period. The least day/night acidity fluctuationoccurs in senescent stems, i.e., stems with fruit scars and somedegree of tissue deterioration, which were not sampled in thisstudy.
The day/night fluctuation of acidity during the wintermonths is large, averaging a difference of 58.3 ± 9.1 ,ueq/gfresh wt, throughout the periods of rainfall until the middle ofMarch (Fig. 4). Daytime minimal acidity values vary signifi-cantly between winter sampling periods, and these values aregenerally higher than daytime summer values. The nighttimeacidity maxima are consistently higher than nighttime summervalues. Following the termination of rainfall in late March thenocturnal accumulation of acidity slowly decreases until theday/night difference approaches 21.7 ± 3.8 ,ueq/g fresh wtin midsummer. Despite severe soil water stress and decreasingplant e the circadian acid fluctuation persists around this re-duced amplitude level until August.
Plant Physiol. Vol. 54, 1974
www.plantphysiol.orgon December 26, 2018 - Published by Downloaded from Copyright © 1974 American Society of Plant Biologists. All rights reserved.
SZAREK AND TING Plant Physiol. Vol. 54, 1974
r T--- I
100--
'I
J
CD
n~~~~~~~L
J F M A M J J A
MONTH OF YEAR
I - T
'11 111~ 1I_L__1 _ _
SO N D~i
FIG. 4. Annual variation in the day/night accumulation of ti-tratable acidity. The level of accumulation has been calculated froma minimum of 12 acidity determinations: 6 taken at sunset (lowerpoint) and 6 taken at sunrise (upper point) for each sampling period.
A midsummer rainfall in early August dramatically affectsthe acid metabolism of 0. basilaris. Maximal levels of night-time acidity increase immediately and approach the levels ofmidwinter maxima. Daytime acidity minima are more uniformand apparently do not shift upward as do midwinter values.Following this brief period of enhanced activity day/nightacidity differences decrease to levels measured prior to theAugust rainfalls and stabilize near this level throughout theremainder of the year. Experimental results from the previousyear of 1972 indicate that enhanced acid metabolism in latefall and early winter also occurs following rainfall. This re-
duced level of acid metabolism which occurred during 1973is directly related to the lack of significant early winter rainfall.Enhanced acid synthesis, as will be pointed out below, occurs
as stomatal diffusion resistances and mesophyll resistancesdecrease, allowing increased nocturnal CO. assimilation intoorganic acids.
Stomatal and Mesophyll Resistances. The patterns of theseasonal variation in the stomatal and mesophyll resistancesto gas transfer are presented in Figures 5 and 6. respectively.During periods with plant A values near saturation stomatademonstrate nocturnal opening and r, values are about 5 sec!
cm, with single observed minima of 3 sec/cm. Calculated r,,'values are moderate, with mean values approaching 10 to 50sec/cm, although single values may approach 2 to 3 sec/cm.From the data of early February it is apparent that nocturnalrs minima increase as soil values concurrently decrease tobelow -40 bars. Throughout the early spring the minimal r,
values are relatively uniform until the termination of rainfallin late March. Minimal nocturnal r, values increase to valuesgreater than 25 sec/cm during periods without rainfall, and as
plant water stress increases the stomata remain closed for a
continually increasing period of the night. Stomata remainclosed throughout the day/night cycle during midsummer.Mesophyll resistances demonstrate an over-all increase fromearly March and remain very high during periods withoutrainfall.The immediate decrease in the r, and r,,' values following a
midsummer rainfall demonstrate the rapid response of 0.basilari.v. Following increased plant i, minimal rs and r,,,' valuesapproach midwinter values, again permitting exogenous gas
exchange. Diurnal stomatal opening during such periods is ini-tiated, and stomata remain open throughout much of the day.As plant water stress begins to increase, r<,' increases to very
high values, although r, values do not increase as rapidly. The,., values are in excess of 25 sec/cm after 2 months, and
stomata again remain closed throughout the day/night cycle.Minimal nocturnal r, values do not decrease significantlythroughout the remainder of the year, and calculated rm' valuesdecrease only slightly.CO2 Assimilation. The pattern of the seasonal variation in
the rates of CO2 assimilation is presented in Figure 7. An en-hanced level of CO2 assimilation persists during the periods ofrainfall until late March. The maximal rates of nocturnal CO2assimilation approach 17 mg C02/dm2 hr during a samplingperiod in late January and show a general decline in the rate ofincorporation until very low rates of uptake, i.e., 0.1 mg CO2/dm2 hr, are attained in late March. During periods of severe
soil water stress radioactive CO2 assimilation is not measure-able, even in stems with open stomata. Despite mild to moder-ate plant water stress CO2 assimilation rates are very low fromMarch until August. Enhanced rates of nocturnal CO2 assimila-tion occur after the midsummer rainfalls. Following the termi-nation of rainfall over-all CO2 assimilation again declines tovery low rates of uptake.The most anomalous aspect of the CO2 assimilation in 0.
basilaris is the pattern of the diurnal incorporation of radio-active CO2. In all cases beyond the first few hours of daylight,
LUI
z
Uf)U)
-J
2
0
51\ 1 \,-
03 -. _.I1
z
<2
>f) 102
U)
X E
a, 101
oLL0
15
z0_- E
=n E 10 _
V)N
<0°
(U Ea0 5I
J F M A M J J A
MONTH OF YEAR
/-.--Nl5
V/ i
\ ,
I 6 '~~~~~~~~~~~~~~~~~~~~~~~~~
7
. .,
S 0 N D
FIG. 5. Annual variation in stomatal diffusion resistances. Eachvalue is the single, lowest r, value observed during each samplingperiod.
FIG. 6. Annual variation in mesophyll resistances to CO2 trans-fer. The mean value (closed circle) and range of the values (bar)are presented for each sampling period.
FIG. 7. Annual variation in CO2 assimilation. The mean value(closed circle) and range of the values (bar) are presented for eachsampling period. The calculated rates of assimilation have been de-rived from the specific radioactivity of the "CO2 utilized through-out each sampling period and incorporated by the stem tissue of 0.basilaris.
78
www.plantphysiol.orgon December 26, 2018 - Published by Downloaded from Copyright © 1974 American Society of Plant Biologists. All rights reserved.
SEASONAL PATTERNS OF ACID METABOLISM
the rates of photosynthetic uptake of 14CO2 have generallybeen less than 0.5 mg CO./ dM2 hr. We have measured suchrates when stomata were open, i.e., r, values of 3 to 5 sec/cm,and plant f values near -5 bars. This phenomenon has beendemonstrated in both winter and summer during this study.
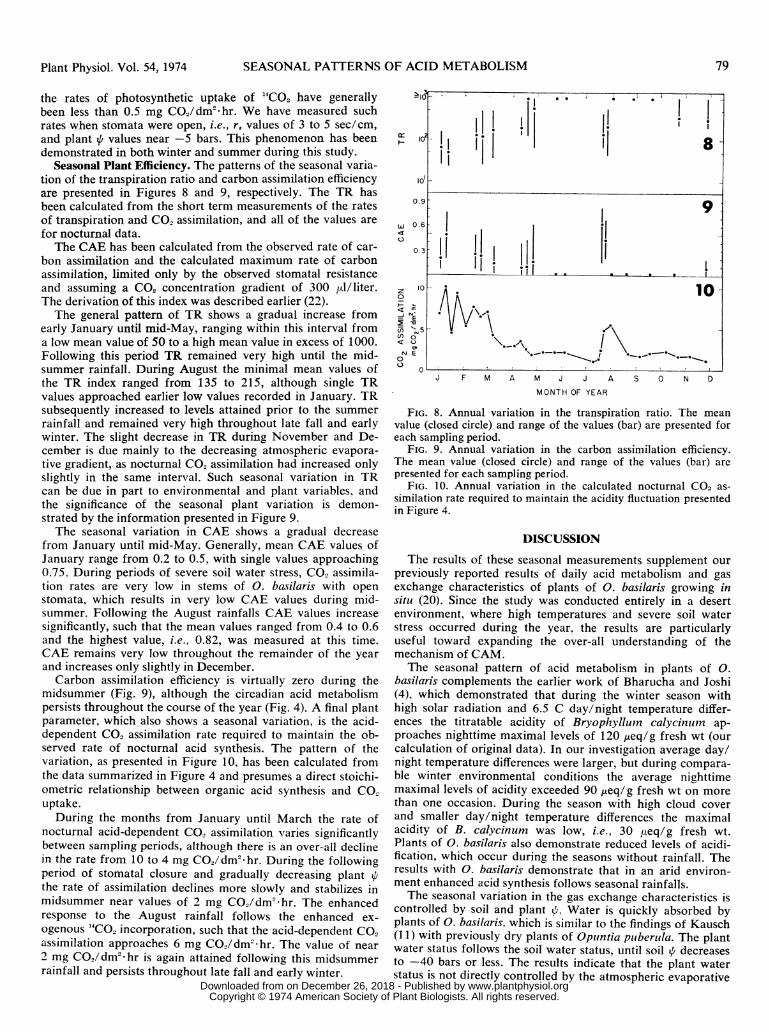
Seasonal Plant Efficiency. The patterns of the seasonal varia-tion of the transpiration ratio and carbon assimilation efficiencyare presented in Figures 8 and 9, respectively. The TR hasbeen calculated from the short term measurements of the ratesof transpiration and CO2 assimilation, and all of the values arefor nocturnal data.
TIhe CAE has been calculated from the observed rate of car-bon assimilation and the calculated maximum rate of carbonassimilation, limited only by the observed stomatal resistanceand assuming a CO, concentration gradient of 300 pl/liter.The derivation of this index was described earlier (22).The general pattern of TR shows a gradual increase from
early January until mid-May, ranging within this interval froma low mean value of 50 to a high mean value in excess of 1000.Following this period TR remained very high until the mid-summer rainfall. During August the minimal mean values ofthe TR index ranged from 135 to 215, although single TRvalues approached earlier low values recorded in January. TRsubsequently increased to levels attained prior to the summerrainfall and remained very high throughout late fall and earlywinter. The slight decrease in TR during November and De-cember is due mainly to the decreasing atmospheric evapora-tive gradient, as nocturnal CO2 assimilation had increased onlyslightly in the same interval. Such seasonal variation in TRcan be due in part to environmental and plant variables, andthe significance of the seasonal plant variation is demon-strated by the information presented in Figure 9.The seasonal variation in CAE shows a gradual decrease
from January until mid-May. Generally, mean CAE values ofJanuary range from 0.2 to 0.5, with single values approaching0.75. During periods of severe soil water stress, CO2 assimila-tion rates are very low in stems of 0. basilaris with openstomata, which results in very low CAE values during mid-summer. Following the August rainfalls CAE values increasesignificantly, such that the mean values ranged from 0.4 to 0.6and the highest value, i.e., 0.82, was measured at this time.CAE remains very low throughout the remainder of the yearand increases only slightly in December.Carbon assimilation efficiency is virtually zero during the
midsummer (Fig. 9), although the circadian acid metabolismpersists throughout the course of the year (Fig. 4). A final plantparameter, which also shows a seasonal variation, is the acid-dependent CO2 assimilation rate required to maintain the ob-served rate of nocturnal acid synthesis. The pattern of thevariation, as presented in Figure 10, has been calculated fromthe data summarized in Figure 4 and presumes a direct stoichi-ometric relationship between organic acid synthesis and CO.uptake.
During the months from January until March the rate ofnocturnal acid-dependent CO2 assimilation varies significantlybetween sampling periods, although there is an over-all declinein the rate from 10 to 4 mg CO2/dm2 hr. During the followingperiod of stomatal closure and gradually decreasing plant Qthe rate of assimilation declines more slowly and stabilizes inmidsummer near values of 2 mg CO2/dm2 hr. The enhancedresponse to the August rainfall follows the enhanced ex-ogenous 14CO2 incorporation, such that the acid-dependent CO2assimilation approaches 6 mg CO2/dm2 hr. The value of near2 mg CO2/dm2-hr is again attained following this midsummerrainfall and persists throughout late fall and early winter.
J F M A M J J A S 0 N D
MONTH OF YEAR
FIG. 8. Annual variation in the transpiration ratio. The meanvalue (closed circle) and range of the values (bar) are presented foreach sampling period.
FIG. 9. Annual variation in the carbon assimilation efficiency.The mean value (closed circle) and range of the values (bar) arepresented for each sampling period.
FIG. 10. Annual variation in the calculated nocturnal CO2 as-
similation rate required to maintain the acidity fluctuation presentedin Figure 4.
DISCUSSION
The results of these seasonal measurements supplement ourpreviously reported results of daily acid metabolism and gasexchange characteristics of plants of 0. basilaris growing insitu (20). Since the study was conducted entirely in a desertenvironment, where high temperatures and severe soil waterstress occurred during the year, the results are particularlyuseful toward expanding the over-all understanding of themechanism of CAM.The seasonal pattern of acid metabolism in plants of 0.
basilaris complements the earlier work of Bharucha and Joshi(4), which demonstrated that during the winter season withhigh solar radiation and 6.5 C day/night temperature differ-ences the titratable acidity of Bryophyllumn calycinumn ap-proaches nighttime maximal levels of 120 ,ueq/g fresh wt (ourcalculation of original data). In our investigation average day/night temperature differences were larger, but during compara-
ble winter environmental conditions the average nighttimemaximal levels of acidity exceeded 90 ,ueq/g fresh wt on more
than one occasion. During the season with high cloud cover
and smaller day/night temperature differences the maximalacidity of B. calycinum was low, i.e., 30 ,ueq/g fresh wt.Plants of 0. basilaris also demonstrate reduced levels of acidi-fication, which occur during the seasons without rainfall. Theresults with 0. basilaris demonstrate that in an arid environ-ment enhanced acid synthesis follows seasonal rainfalls.The seasonal variation in the gas exchange characteristics is
controlled by soil and plant t@. Water is quickly absorbed byplants of 0. basilaris, which is similar to the findings of Kausch(11 ) with previously dry plants of Opuntia puberula. The plantwater status follows the soil water status, until soil q' decreasesto -40 bars or less. The results indicate that the plant waterstatus is not directly controlled by the atmospheric evaporative
I_-/\ ,ff . X X .
79Plant Physiol. Vol. 54, 1974
www.plantphysiol.orgon December 26, 2018 - Published by Downloaded from Copyright © 1974 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 54, 1974
demand during periods of severe soil water stress. Nocturnaland diurnal opening of stomata is common during periods ofmild plant water stress. As plant water stress begins to in-crease, diurnal stomatal opening is first restricted, and as thestress continues nocturnal opening is restricted in a rathercontinual manner until the stomata remain closed throughoutthe day/night cycle in midsummer. During the month of Maythe restriction of nocturnal stomatal opening was initiated andby early June stomata did not show nocturnal opening. Plant tvalues in May ranged from -6.5 to -7.5 bars, and by earlyJune had decreased to values of -10 bars. These results sug-gest the threshold level of plant b required for nocturnal stoma-tal opening would be in the range of -8 to -10 bars. Theseasonal results confirm that nocturnal stomatal opening isrestricted to those periods when plant f is characteristicallygreater than -8 to -10 bars. Diurnal stomatal opening is moresensitive to moderate plant water stress, since stomatal openingis restricted beyond the first few hours of daylight when plantf is less than -5 to -6 bars.The results of the seasonal variation in gas exchange agree
with the daily and seasonal patterns of gas exchange of otherCAM plants. Increasing water stress has been shown to reducediurnal gas exchange initially to a greater extent than nocturnalgas exchange in Agave americana and Aeonium haworthii(14), while the results with Bryophyllum daigreinontianumdemonstrate a continual reduction in nocturnal gas exchangeonce diurnal gas exchange is completely restricted (12). Recentseasonal results with Dudleya farinosa demonstrated a decreasein leaf conductivity to water vapor as drought-induced waterstress increased (2). The seasonal results during the midsummerof our study extend the observations of gas exchange character-istics of CAM plants, since continuous stomatal closurethroughout the day/night cycle occurs during periods of severesoil water stress and moderate plant water stress.
Nocturnal CO2 assimilationin 0. basilaris exhibits a signifi-cant seasonal variation in rates of incorporation. Earlier labo-ratory investigations with CAM plants have emphasized thesignificance of nighttime temperatures on nocturnal CO2 as-similation, with netCO2 assimilation generally at temperaturesless than 25 C (13). However, these experimental results werederived from conditions of ample soil water and mild plantwater stress. The work of Kausch (l1), with a CAM plantwhich was previously exposed to several weeks of dry condi-tions, demonstrated that increased amounts of total nocturnalCO2 uptake followed remoistening. Little information is avail-able from in situi investigations, except the recent results ofBartholomew (2), which suggested that nocturnal CO2 uptakein conditions of moderate soil water stress was not significantlycorrelated with nocturnal temperatures. His results appear tobe more closely correlated with plant water status. Our findingswith 0. basilaris demonstrate that CO. assimilation during pe-riods of stomatal opening correlates with plant k. Enhancedlevels of CO2 assimilation have been recorded during periodswith low, as well as moderate nighttime temperatures. Theprincipal mechanism by which water stress reduces nocturnalCO. assimilation appears to be increased total gas transfer re-sistances, with r,,' being more sensitive than r_.
Water stress has long been known to reduce photosyntheticcarbon assimilation, although numerous conflicting studies con-cerning the effects of water stress on r,,,' values have been re-ported. Some reports from laboratory investigations with agri-cultural plants indicate that the ri value is unaffected by waterstress over a wide range of plant t (23). Other reports indicatethat only a portion of the water stress-induced reduction inphotosynthesis can be accounted for by increased stomatalresistances (17). The seasonal variation in nocturnal CO2 as-
similation rates in plants of 0. basilaris cannot be solely ac-counted for by increased r, values. Other workers have dem-onstrated rhythmic fluctuations in the CO2 compensationconcentration in CAM plants which are influenced by envi-ronmental factors, i.e., photoperiod and thermoperiod (9). Thedependency of CO2 assimilation on nonstomatal transfer re-sistances is consistent throughout our study, although theeffects of expected fluctuations in the CO2 compensation con-centration on the magnitude ofrm' are difficult to estimate atthis time.The seasonal variation in the TR and CAE demonstrate that
the actual values can vary significantly between sampling pe-riods. Moreover, environmental differences can cause discrep-ancies in the efficiency values calculated from a single sam-pling period. The difference in the indices can be pointed outfrom the values recorded during the months of January andAugust. While maximal TR efficiency occurred in the midwin-ter month of January, maximal CAE occurred during August.Since the maximal mean January and August CAE values arecomparable, i.e., 0.5 to 0.6, the greater differences in the re-spective maximal mean TR values, i.e., 50 to 135, are probablydue solely to higher atmospheric evaporative gradients duringthe August nights. Unlike the TR index of efficiency, CAE isdependent only upon plant variables and therefore is less am-biguous in describing the over-all plant response.The TR of plants of 0. basilaris is within the range of the
values reported for the three photosynthetic groups of plants.Typical values are 450 to 950 for C3 plants, 250 to 350 for C4plants, and 50 to 125 for CAM plants (7, 22). The large varia-tion of TR reported in this study indicates that during periodsof mild soil and plant water stress the efficiency of water useis very high, while during periods of severe soil water stress theefficiency is very low. In some instances in our study TR ap-proaches values of 25 to 80, which are similar to previouslyreported minimal nocturnal values of other CAM plants (10,14). However, TR values throughout most of the year in ourstudy are comparable to the typical diurnal values of the othertwo photosynthetic groups of plants.
The CAE of plants of 0. ba*ilaris is also within the range ofthe values of the three photosynthetic groups of plants. Calcu-lating from the data of Bierhuizen and Slatyer (5) with cotton,a C3 plant, the maximal CAE value is 0.22. Calculating fromthe data of Bjorkman et al. (6) with Tidestroinia oblongifolia,a C, plant, the maximal CAE value is 1.00. Calculating fromthe data of Neales, Patterson, and Hartney (14) for pineapple,a CAM plant, the maximal nocturnal and diurnal values are0.72 and 0.93, respectively. Maximal nocturnal values for 0.basilaris are 0.75 during midwinter and 0.82 during midsum-mer. These values are from periods with ample soil water andmild plant water stress and are significantly higher than thevalues from periods with severe soil water stress. During pe-riods of increasing plant water stress CAE values are low andapproach zero when ri,,' values are very large. The CAE indexdemonstrates that during some environmental conditions aCAM plant may operate as efficiently as a C4 plant, while dur-ing other periods CAM plant efficiency is below the level ofC. plants.
The results of this study demonstrate that during arid en-vironmental conditions a reduced level of acid metabolism per-sists, despite the cessation of atmospheric gas exchange. Undersevere soil water stress conditions a rate of nocturnal CO. as-similation into organic acids stabilizes at 2 mg CO2/dm2 hr.Previously, we reported that 80% of the carbon assimilatedinto organic acids was subsequently converted to photosyn-thetic products (21). Our earlier results would suggest thatphotosynthetic rates in 0. basilaris. utilizing CO2 available
80 SZAREK AND TING
www.plantphysiol.orgon December 26, 2018 - Published by Downloaded from Copyright © 1974 American Society of Plant Biologists. All rights reserved.

SEASONAL PATTERNS
solely from endogenous sources, would be on the order of 1.6mg CO2/dm2 hr during periods of severe soil water stress. Thisreduced level of metabolic activity, which is dependent uponthe recycling of endogenous C02, maintains a moderate energystate throughout periods without rainfall.
Acknowledgments-We would especially like to thank Dr. H. B. Johnson forvaluable advice in many phases of this study and Dr. F. C. Vasek for critical re-view of this manuscript. We are grateful for the skilled assistance of F. Grgurevicand T. Yonkers.
LITERATURE CITED
1. ALBERT, B. 1971. Effects de la radiation globale sur la synthese d'acides or-ganiques et la regulation stomatique des plantes succulentes. Exempled'Ananas comosus (L.) Merr. Oecol. Plant. 6: 25-34.
2. BARTHOLOMEW, B. 1973. Drought response in the gas exchange of Dudleyafarinosa (Crassulaceae) grown under natural conditions. Photosynthetica 7:114-120.
3. BENNETT-CLARK, T. A. 1933. The role of organic acids in plant metabolism.Part I. New Phytol. 32: 37-71.
4. BHARU-CHA, F. R. AND G. V. JOSHI. 1958. Studies in crassulacean metabolismin Bryophyllum calycinum under tropical conditions. J. Biol. Sci. 1: 5-12.
5. BIERHUIZEN, J. F. AND R. 0. SLATYER. 1964. Photosynthesis of cotton leavesunder a range of environmental conditions in relation to internal and ex-ternal diffusive resistance. Aust. J. Biol. Sci. 17: 348-359.
6. BJ6RKMAN, O., R. W. PEARCY, A. T. HARRISON, AND H. MOONEY. 1972. Photo-synthetic adaptation to high temperatures: a field study in Death V-alley,California. Science 175: 786-789,
7. BLACK, C. C., JR. 1973. Photosynthetic carbon fixation in relation to net CO2uptake. Annu. Rev. Plant Physiol. 24: 253-286.
8. EKERN, P. C. 1965. Evapotranspiration in pineapple in Hawaii. Plant Physiol.40: 736-739.
9. JONES, Ml. B. AND T. A. MANSFIELD. 1972. A circadian rhythm in the level ofcarbon dioxide compensation in Bryophyllum fedfschenkoi with zero v-aluesduring the transient. Planta 103: 134-146.
10. JOSHI, M. C., J. S. BOYER, AND P. J. KRAMIER. 1965. Growth, carbon dioxideexchange, transpiration, and transpiration ratio of pineapple. Bot. Gaz. 126:174-179.
OF,ACID METABOLISM 81
11. KACSCH, W. 1965. Beziehungen zwischen Wurzelwachstum, Transpiration, undCO2-Gaswechsel bei einigen Kakteen. Planta 66: 229-238.
12. KLrGE, M. AND K. FISCHER. 1967. Uber Zusammenhange zwischen dem CO2-Austausch und der Abgabe von Wasserdampf durch Bryophyllum daigre-montianum Berg. Planta 77: 212-223.
13. KLUGE, M., 0. L. LANGE, M. v. EICHMA'NN, AND R. SCHMID. 1973. DiurnalerSauerhythmus bei Tillandsia usneoides: Untersuchungen uiber den Weg desKohlenstoffs sowie die Abhiingigkeit des CO2-Gaswechsels von Lichtinten-sitat, Temperatur und Wassergehalt der Pflanze. Planta 112: 357-372.
14. NEALES, T. F., A. A. PATTERSON, AND V. J. HARTN-EY. 1968. Physiologicaladaptation to drought in the carbon assimilation and water loss of xero-phytes. Nature 219: 469-472.15. OLIVERIO, V. T., C. DENHAM, AND J. D. DAVIDSON. 1962. Oxygen flask com-
bustion in determination of 14C and 3H in biological materials. Anal. Bio-chem. 4: 188-189.
16. RH-NsoN-, S. L. AND M. THOMsAS. 1960. Crassulacean acid metabolism. Annu.Rev. Plant Physiol. 11: 81-110.
17. REDSHAw, A. J. AN-D H. MEIDN'ER. 1972. Effects of water stress on the re-sistance to uptake of carbon dioxide in tobacco. J. Exp. Bot. 23: 229-240.
18. SHIMSHI, D. 1969. A rapid field method for measuring photosynthesis withlabelled carbon dioxide. J. Exp. Bot. 20: 381-401.19. SIDERIS, C. P., H. Y. YOUNG, AND H. H. Q. CHL'N. 1948. Diurnal changes and
growth rates as associated with ascorbic acid, titratable acidity, carbo-hydrate and nitrogeneous fractions in the leaves of Ananas comosus (L.)Merr. Plant Physiol. 23: 38-69.
20. SZAKEK, S. R., H. B. JOHNSON. AND I. P. TING. 1973. Drotught adlaptat!on inOpuntia basilaris. Significance of recycling carbon through crassulaceanacid metabolism. Plant Physiol. 52: 539-541.
21. TING, I. P. AND W. M. DUGGER, JR. 1968. Non-autotrophic carbon dioxidemetabolism in cacti. Bot. Gaz. 129: 9-15.
22. TING, I. P., H. B. JOHNSON, AND S. R. SZAREK. 1972. Net C02 fixation incrassulacean acid metabolism plants. Izn: C. C. Black, ed., Net CarbonDioxide Assimilation in Higher Plants. Proc. Symp. S. Sect. Am. Soc. PlantPhysiol. and Cotton, Inc., Raleigh. pp. 26-53.
23. TROUGHTON, J. H. AND R. 0. SLATYER. 1969. Plant water status, leaf tempera-ture, and the calculated mesophyll resistance to carbon dioxide of cottonleaves. Aust. J. Biol. Sci. 22: 815-827.24. VAN BAVEL, C. H. MI., F. S. NAKAYAMA, AND WV. L. EHRLER. 1965. Measuring
transpiration resistance of leaves. Plant Physiol. 40: 535-540.
Plant Physiol. Vol. 54, 1974
www.plantphysiol.orgon December 26, 2018 - Published by Downloaded from Copyright © 1974 American Society of Plant Biologists. All rights reserved.





























