Embed Size (px)
Citation preview

J. Zool., Lond. (1984) 202, 535-547
Scent marking strategies in hyaenas (Mammalia)
M. L. G O R M A N Department of Zoology, University ofAberdeen, Scotland, U.K.
AND
M. G. L. MILLS* National Parks Board, Kalahari Gemsbok National Park, Upington, South Africa
(Accepted 21 July 1983)
(With 1 plate and 7 figures in the text)
Brown hyaenas scent mark by making latrines and depositing onto grass stalks two distinct secretions produced in their anal pouch. The latter behaviour is called pasting. Pastings are found spaced throughout a territory but mainly in the interior where the hyaenas spend most of their time. Latrines are clumped near to, but not along, the border. Computer analyses show that intruders encounter scent marks very soon after entering a territory. Comparison with other species indicates that the length of border to be marked by each individual in a group is the important factor in determining whether they mark throughout the temtory or just along the border.
Contents Introduction . . . . . . . . . . . . . . . . . . . . Methods . . . . . . . . . . . . . . . . . . . . . . Results . . . . . . . . . . . . . . . . . . . . . .
Scent marking behaviour . . . . . . . . . . . . . . . .
The spatial distribution of pastings within the temtory . . . . . . . . The spatial distribution of latrines . . . . . . . . . . . . . . The efficiency of paste marking . . . . . . . . . . . . . .
Discussion . . . . . . . . . . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . . . . . . .
The standing crop of pastings . . . . . . . . . . . . . .
Page 535 537 538 538 538 540 54 1 542 543 547
Introduction In the Southern Kalahari, Brown hyaenas (Hyaena brunnea) live in small groups the
members of which share a large territory of 235480 km2 (Mills, 1976). The territories of neighbouring groups overlap to a small extent (Fig. 1) but are defended when neighbours of the same sex meet (Mills, 1983). The members of a group forage alone and at night (Mills, 1978).
Brown hyaenas scent mark throughout their territory using their anal pouches to leave a lipid-rich white secretion and a watery black smear on tall grass stalks (Mills, Gorman & Mills, 1980) (Plate I). We have called this behaviour “pasting”. Paste marking by brown *Present address: National Parks Board, Kruger National Park, Skukuza, South Africa.
0022-5460/84/040535 + 13 $03.00.’0 535
0 1984 The Zoological Society of London

53c M. L. GORMAN A N D M. G. L. MILLS
PLATE 1. (a) Brown hyaena scent marking a grass stalk. (b) A ritual fight between a territory owner and an intruder. (c) A grass stalk marked by a Brown hyaena.
hyaenas involves a significant investment in terms of energy; the white secretion is 97% lipid and during the course of a year one hyaena will deposit some 29,000 paste marks (Mills, Gorman & Mills, 1980). It is assumed, therefore, that the behaviour is adaptive and that the information conveyed by the scent mark increases the inclusive fitness of its author.
Brown hyaenas will defaecate anywhere in their territories but on occasion they will do so at specific places where there are accumulations of 5-50 faeces. These latrines may also serve a function in scent marking as they do in other carnivores (MacDonald, 1980).

SCENT MARKING IN HYAENAS 537
Two functions have been ascribed to scent marking behaviour in the Brown hyaena (Mills, Gorman & Mills, 1980) and these ideas are further developed here.
First, it is postulated that paste marks convey information to other members of the social group. Brown hyaenas forage alone and usually feed on small items with a low renewal rate (Mills, 1978). It is important, therefore, that each knows where other hyaenas have foraged in the recent past so that they can avoid unproductive areas. The black, watery component of the scent mark loses its odour relatively quickly and may contain information on the length of time that has elapsed since it was deposited.
In addition, scent marking may function in communication between social groups. Fights between territorial carnivores are rare because individuals usually avoid violating occupied space (e.g. Erlinge, 1977; Kruuk, 1972). If a conflict develops between resident and intruder it is usually settled in a conventional manner, without escalation to fighting, on the basis of property tenure with the intruder withdrawing. This is true for Brown hyaenas (Mills, 1983).
If territorial conflicts are to be settled by such a convention it is important that the resident animal and the area it occupies can be unambiguously recognized, with no possibility of cheating or bluff. One way to achieve this is for residents to mark their territory with scent; only a long term resident can have had the opportunity to pepper an area with its own scent marks. An individual encountering a scent marked area will quickly perceive that it is already occupied and that there is little to be gained from advancing further, because to do so would mean encountering a resident who would, if necessary, fight to kill. Should an intruder meet an individual with a smell similar to some, or all, of the scent marks in the territory then it can be sure that this is a resident animal and the conflict can be settled con- ventionally. In essence the scent marks act as a cue to establish an asymmetry of contest, allowing a conventional settlement and preventing escalation to real fighting (Maynard Smith & Parker, 1976; Gosling, 1982). There is behavioural and chemical evidence that Brown hyaena pastings contain the necessary information on individual identity (Mills, Gorman & Mills, 1980).
If scent marking behaviour in the Brown hyaena is adaptive then one would predict that scent marks should be distributed in a way that maximizes their chance of being discovered by the individuals for whom they are intended. If the two functions proposed above are correct then these addressees can be outsiders or other members of the social group. The aim of this paper is to describe the dispersion pattern of scent marks in a Brown hyaena territory in order to test the above prediction.
Methods
The data on scent marking were collected in the Kalahan Gemsbok National Park, Republic of South Africa and in the neighbouring Gemsbok National Park, Republic of Botswana. The area is semi-desert and covered with sand-dunes broken by pans and the dry bed of the river Nossob. The dunes and river bed form distinct habitats with the latter providing a much higher density of potential hyaena food during the study (Mills, 1982).
Between 1976 and 1978 we followed 2 females and 3 males who formed the Kwang group (Fig. 1). Each hyaena was fitted with a radio-collar and 2 beta-lights. Hyaenas were located at dusk or as soon after dark as possible and followed in a vehicle, using the beta-lights or a hand held spotlight, for as long as possible before they rested up for the day. When following a hyaena we noted its direction of movement, and whenever it stopped or changed its direction we noted the distance it had moved

538
FIG. 1. Map of part of the Nossob valley showing the territories ofthree Brown hyaena social groups. The territor- ies are convex polygons drawn around the movements of known animals (see Mills, 1976 for details). 0 Nossob Camp; = Nossob River.
by using the vehicle odometer. Each time a hyaena stopped to paste or visit a latrine the odometer reading was again noted. All movements and scent marking locations were later transferred to a large scale map (e.g. Fig. 2).
In this way we gradually built up a representative sample of the places in which hyaenas deposited scent marks. In total we followed the hyaenas for 1947 km and recorded the positions of 5 144 pastings and 169 latrines.
Results Scent marking behaviour
The behaviour associated with pasting has been described in detail elsewhere (Mills, Gorman & Mills, 1980).
Defaecation at a latrine was often accompanied by other scent marking behaviour. Typi- cally a hyaena came to a latrine, spent several minutes sniffing and pawing at the faeces and at nearby grass stalks. It then defaecated, scratched the ground with its forefeet and pasted before continuing on its way.
The standing crop of pastings The number of active scent marks in a territory at any given time is a function of the
rate at which new marks are laid down and of the rate at which old ones decay. In the Kwang territory there were five to seven adult animals each travelling on average 3040 kmhight (Mills, 1978) and pasting at a rate of 2.64 markslkm. We do not have an accurate measure- ment of how long a paste mark remains active but the white component can be detected by the human nose for at least 30 days and probably for much longer by a hyaena. Taking 30 days as a conservative estimate of the average length of life of a scent mark means that the Kwang territory will contain, at any given time, upwards of 20,000 active paste marks.

SCENT MARKING IN HYAENAS 539
Nossob Camp 0 m t
10 km
FIG. 2. Map of the locations at which members of the Kwang group were seen to paste mark in 1977-78.

540
r 7 12- 0 x - 0 m U c
g U " 0 -
D
- &
5 z
M. L. G O R M A N A N D M. G. L. MILLS
1 6 1 L
-
8-
4 -
-
0-
Nearest scent mark (m)
FIG. 3. The distribution of nearest neighbour distances for the scent mark locations in the Kwang territory, together with the distribution that would be expected if the marks were randomly dispersed. 0 Expected; :::observed.
The spatial distribution of pastings within the territory The vertical distribution
The mean distance from the bases of marked grasses to the white secretion was 687.2 k2 1-9 mm ( f standard error, n= 50) and to the black secretion 724 +20 mm. For comparison the mean adult shoulder height, which corresponds to the normal height of the nose, was 787k81 mm (n=20).
The horizontal distribution Figure 2 shows the distribution of the pastings we saw deposited in 1977-78 and serves
to show that they were scattered throughout the whole territory. Combining these data with those collected in 1976 gives 5144 locations, a representative sample of the places within the territory that hyaenas paste. This is probably a much lower number of marks than is actually present at any given time.
This representative pattern of paste mark locations within the territory was subjected to a nearest neighbour analysis and compared to the distribution to be expected if the locations were randomly distributed (Patterson, 1965). The observed distribution of nearest neighbour distances differed significantly from the random one (x2 = 4 19, P< 0-00 1). The paste marks were more regularly spaced than random through the territory (Fig. 3).
The spatial distribution of the sample of paste marking sites was further analysed by means of the computer programs SYMAP and SYMVU (Laboratory for computer graphics, Harvard University, U.S.A.). These programs display, graphically, spatially distributed quantitative data as a three dimensional map in which the vertical dimension reflects the value of the quantitative data, in this case the density of pasting points. To facilitate such an analysis we superimposed on the map of the Kwang area a matrix of 2.5 x 2.5 km squares and then counted the number of pasting sites in each. The density values were analysed using

SCENT MARKING IN HYAENAS 54 1
45 Marks/km* A
FIG. 4. A three dimensional map, generated by S Y M W of the density of scent marks in the Kwang group territory.
10 class intervals of equal size covering the range 0-280 pastings per square. The resultant map is plotted as if seen from the south-east at an altitude of 35" above the horizontal (Fig. 4). The analysis shows clearly that although paste marks were found throughout the territory, the highest densities were near to the centre with a progressive decrease towards the borders. It must be stressed that while the map gives a clear representation of relative differences in scent mark density throughout the territory, in reality the absolute values are probably several times higher.
The spatial distribution of latrines The majority of latrines were situated next to a landmark such as a tree or bush (75%),
the side of a road (15.2%) or at a dried out pool (2.1%). At 34 randomly chosen latrines, associated with Boscia albitrunca trees, 7 1 O/o were situated on the south side of the tree, 2 1 % on the west side and the rest on the east side. The tendency to be clumped on the south side was highly significant (Rayleigh's Test: Z= 17.14, P<O.OOI).
The specific site of the latrines appeared to be important to the hyaenas. When all 50 faeces were removed from a latrine to an equally prominent tree 40 m away, and the original

542 M. L. GORMAN AND M. G. L. MILLS
0 O\\
0
0 0
0
Den ** : 0”
0 0 0 0
0
.. ..
FIG. 5 . Map of the locations of latrines seen to be used by members of the Kwang group during the study. 0 1976: 1977-78.
area covered with a thick layer of sand, the hyaenas continued to use the original site for at least four months.
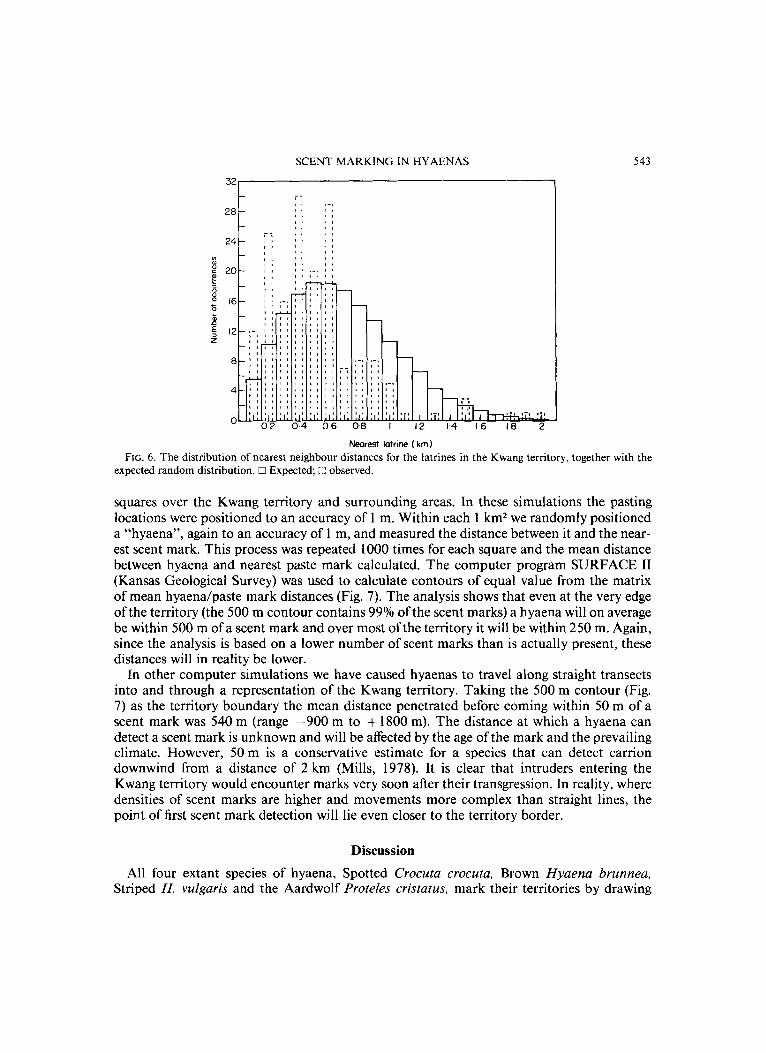
Figure 5 shows the positions of all the latrines we saw used by Brown hyaenas. It is clear that the latrines are scattered throughout the territory, but again not uniformly so. A nearest neighbour analysis of all the latrine locations within the territory revealed that their distri- bution was more clumped than random (Fig. 6, x2=78.36, P<O.OOl) . The groupings of latrines occurred mainly near to the border of the territory, particularly to the north and south where it crossed the dry bed of the river Nossob (Fig. 5).
The frequency at which hyaenas visited latrines, expressed as latrines visited/km travelled, appeared to be higher when they were moving near to the border of the territory than when they were in the hinterland. We analysed this by placing a matrix of 2.5 x 2-5 km squares over the whole area and then calculating the total distance moved within each square by the hyaenas and the number of latrines they visited. We then designated those squares on the periphery of the territory as border squares and the others as internal squares (Mills, Gorman & Mills, 1980). The mean frequency of latrine visitation was significantly higher in border squares than in internal squares in both years of the study (Table I).
We have few quantitative data on the lifespan of latrines. Some are probably relatively shortlived but one, first found in 1972, was still in regular use in 1980.
The eficiency of paste marking We turn now to the problem of how likely it is that hyaenas will encounter paste marks
as they move into, or through, an occupied territory given the density and dispersion of marking sites described above. In computer simulations we superimposed a matrix of 1 km
SCENT MARKING IN HYAENAS
32 _ _ , I
543
I I .-. I , L , t I . I ,
Nearest latrine ( km) FIG. 6. The distribution of nearest neighbour distances for the latrines in the Kwang territory, together with the
expected random distribution. 0 Expected; ;:! observed.
squares over the Kwang territory and surrounding areas. In these simulations the pasting locations were positioned to an accuracy of 1 m. Within each 1 km2 we randomly positioned a “hyaena”, again to an accuracy of 1 m, and measured the distance between it and the near- est scent mark. This process was repeated 1000 times for each square and the mean distance between hyaena and nearest paste mark calculated. The computer program SURFACE I1 (Kansas Geological Survey) was used to calculate contours of equal value from the matrix of mean hyaenalpaste mark distances (Fig. 7). The analysis shows that even at the very edge of the territory (the 500 m contour contains 99% of the scent marks) a hyaena will on average be within 500 m of a scent mark and over most of the territory it will be within 250 m. Again, since the analysis is based on a lower number of scent marks than is actually present, these distances will in reality be lower.
In other computer simulations we have caused hyaenas to travel along straight transects into and through a representation of the Kwang territory. Taking the 500 m contour (Fig. 7) as the territory boundary the mean distance penetrated before coming within 50 m of a scent mark was 540 m (range -900 m to + 1800 m). The distance at which a hyaena can detect a scent mark is unknown and will be affected by the age of the mark and the prevailing climate. However, 50 m is a conservative estimate for a species that can detect carrion downwind from a distance of 2 km (Mills, 1978). It is clear that intruders entering the Kwang territory would encounter marks very soon after their transgression. In reality, where densities of scent marks are higher and movements more complex than straight lines, the point of first scent mark detection will lie even closer to the territory border.
Discussion
All four extant species of hyaena, Spotted Crocuta crocuta, Brown Hyaena brunnea, Striped H. vulgaris and the Aardwolf Proteles cristatus, mark their territories by drawing

544 M. L. GORMAN A N D M. G. L. MILLS
I 10 krn
FIG. 7. A contour map of the average distance (m) between a randomly positioned hyaena and the nearest scent mark belonging to the Kwang group. The 500 m contour approximates to the territory border and contains 99% of the Kwang scent mark locations. (See text for further details).
TABLE I The overallfrequency of latrine visits by Brown hyaenas ofthe Kwanggroup
Latrines visited per km Mann-Whitney followed V-test
Year Border squares Internal squares U Z P
1976 0.14 0.04 131.5 -2 .38 0.009 1911-78 0 .33 0.09 503.0 3 .44 <0 .003
grass stems through their anal pouches and by defaecation at latrines. Each species produces a form of the longlasting, lipid-rich secretion described above, and in each it is thought to be involved in signalling territory ownership (Kruuk, 1972; Kruuk & Sands, 1972; Kruuk, 1976). The Brown hyaena is unique in producing an additional short-lived component.

SCENT MARKING IN HYAENAS 545
Brown hyaenas in the Southern Kalahari pasted throughout their territories but mostly in the central heavily utilized areas (Fig. 4). Apart from pastings, numerous latrines were scattered throughout the territory. These, however, tended to be concentrated in clumps (Fig. 6 ) near to, but not at, the territorial border (Fig. 5). This was particularly so where the border crossed the dry bed of the river Nossob, the richest feeding area for most of the year. When Brown hyaenas were moving near the borders of their territory they visited latrines much more frequently than they did when in the interior (Table I).
The analyses presented in this paper reveal that scent marking behaviour is highly efficient in ensuring the discovery of scent marks by other hyaenas. Pastings had a tight vertical distribution, at a height similar to the normal nose position, which must have maximized their chance of discovery. Latrines were almost always associated with prominent landmarks, such as trees, and there was a very strong tendency for them to be to the south of the tree. The horizontal distribution of pastings, spread throughout the territory but with the highest densities near to the centre where the group members did most of their foraging, would have facilitated the transfer of information among the group as to which areas had recently been harvested. Despite the vast area over which the pastings were dispersed, our computer simulations indicated that potential intruders would have rapidly encountered marks as they moved into a territory (Fig. 7). The latrines, concentrated near to the borders in the most productive parts of the territory, would again have provided an early warning of transgression (Fig. 5).
Hyaena species adopt one of two distinct strategies in terms of how they distribute their scent marks within their territories. The first approach is that seen in the Brown hyaenas of the Kalahari, the second is exemplified by the Spotted hyaenas of the Ngorongoro which place their paste scent marks exclusively along the territorial border (Kruuk, 1972). If scent marking is indeed adaptive then one would expect such differences to have a functional significance.
Spotted hyaenas communicate within their group by posture and vocalization; scent marks are used for inter-group communication. It might be argued that they concentrate their pastings and latrines at the very border of the territory so as to give the earliest possible warning to transgressors. By such an argument the need to signal to both neighbours and fellow group members would be seen as the selective force leading to hinterland marking in Brown hyaenas.
However, the data can be interpreted in a quite different way. Border marking does give the earliest warning of trespass but it involves but a single line of defence which must be maintained intact and renewed regularly if overt conflicts are to be avoided. To maintain a dense, fresh line. of scent marks along the whole border requires a large supply of scent relative to the length of perimeter to be marked. This is possible in the case of the Ngoron- goro Spotted hyaenas where large groups share small territories and the length of border to be marked by each individual is relatively small (Table 11).
Brown hyaenas living in the Kalahari have to maintain very much larger territories in order to maintain an adequate food supply. Given a limited time budget and a finite supply of scent secretion, as territory size increases it becomes progressively more difficult to visit and mark the border with the frequency and regularity of routine necessary to maintain the continuity of the line of scent marks (see also Gosling, 198 1) . The average length of perimeter to be marked by each Kalahari Brown hyaena is an order of magnitude greater than that for a Ngorongoro Spotted hyaena, and their adopted strategy is hinterland marking (Table

546 M. L. GORMAN AND M. G. L. MILLS
TABLE I1 Scent marking patterns in hyaenas
Territory size Circumference*/individual Species and locality Group size (km2) Marking strategy (km)
AardwolP , Serengeti 1-2 1.5 Border 1.7-3,4 Spottedz, Ngorongoro 30-80 30 Border 0 , 2 4 4 6 3 Striped', Serengeti 1 40-70 Hinterland 22-30 Brown4, Kalahari 1 -9 235480 Hinterland 11-16
(Based on a group of live) Spotteds, Kalahari 3-1 5 c. 1000 Hinterland 7.5-3 7.0
Data from Kruuk & Sands (1972)*, Kruuk (1972)z, Kruuk (1976)3, Mills (1982)4, Mills, In prep5. *Based on a circular territory.
11). For populations with a long perimeter to patrol and scent mark, hinterland marking must be a safer strategy; an intruder may penetrate some distance into the territory but sooner or later will encounter scent marks. To attempt to mark only the border in such cases would inevitably mean leaving gaps in the line of marks, with no second line of warning.
The important parameter in determining which strategy to follow when proclaiming territory ownership, border or hinterland marking, appears to be the length of perimeter to be marked by each group member.
Evidence in support of this hypothesis comes from studies of other populations and species of hyaenas.
Spotted hyaenas living in the Kalahari live in much smaller groups and occupy much larger territories than they do in the Ngorongoro (Table 11). The length of border per group member is similar to that of Brown hyaenas and here Spotted hyaenas mark throughout their territory.
Comparison with other species is more difficult since little is known about the detailed biology of the Aardwolf or the Striped hyaena. What is known is in accord with the ideas put forward here. The Aardwolf lives alone, or in pairs, in a very small territory and marks the border (Table 11). The Striped hyaena is a solitary species which occupies a large territory and which adopts the hinterland strategy (Table 11).
It may be, therefore, that the prime factor leading to hinterland marking in the Brown hyaena has been the difficulty of maintaining a continuous line of active scent marks along the border of a very large territory. If this is correct, then signalling to fellow group members with the unique short-lived paste component would appear to be a secondary adaptation.
We would like to thank the Directors of the National Parks Board of Trustees, Republic of South Africa and the Department of Wildlife and National Parks, Republic of Botswana for sanctioning the work in their respective parks.
We are grateful to the National Parks Board for providing facilities, to the Endangered Wildlife Trust for radio tracking equipment, and to the Carnegie Trust and the University of Aberdeen for travel grants.
Finally, we must extend our thanks to Margaret Mills for much help in the field and in the analysis of data, and to Ian Patterson and Hans Kruuk for their constructive criticism of our manuscript.

SCENT MARKING IN HYAENAS 547
REFERENCES Erlinge, S. (1977). Spacing strategy in the stoat Mustela erminea. Oikos 28: 32-42. Gosling, L. M. (1981). Demarcation in a Gerenuk territory: an economic approach. Z. Tierpsychol. 56: 305-322. Gosling, L. M. (1982). A reassessment of the function of scent marking in territories. Z. Tierpsychol. 60: 89-1 18. Kruuk, H. (1972). The spotted hyaena. A study of predation and social behaviour. Chicago: The University of
Kruuk, H. (1976). Feeding and social behaviour of the striped hyaena (Hyaena vulgaris Desmarest). E. A+. Wildl.
Kruuk, H. & Sands, W. A. (1972). The aardwolf (Proteles cristatus Sparrman 1783) as a predator of termites. E.
MacDonald, D. W. (1980). Patterns of scent marking with urine and faeces amongst carnivore communities. Symp.
Maynard Smith, J. & Parker, G. A. (1976). The logic of asymmetric contests. Anim. Behav. 2 4 159-175. Mills, M. G. L. (1976). Ecology and behaviour of the brown hyaena in the Kalahari with some suggestions for
management. Proc. Symp. Endangered Wildl. 1976: 36-42 (Endangered Wildlife Trust, Pretoria). Mills, M. G. L. (1978). Foraging behaviour ofthe brown hyaena (Hyaena brunnea Thunberg, 1820) in the southern
Kalahari. 2. Tierpsychol. 48: 113-141. Mills, M. G. L. (1982). Factors affecting group size and territory size of the brown hyaena Hyaena brunnea in
the southern Kalahari. Z. Tierpsychol. 48: 1 13-14 I . Mills, M. G. L. (1983). Behavioural mechanisms in territory and group maintenance of the brown hyaena Hyaena
brunnea in the southern Kalahari. Anim. Behav. 31: 503-510. Mills, M. G. L., Gorman, M. L. &Mills, M. E. J. (1980). The scent marking behaviour ofthe brown hyaena Hyaena
brunnea. S. A+. J. Zool. 15: 240-248. Patterson, I. J. (1965). Timing and spacing of broods in the black-headed gull. Ibis 107: 433-459.
Chicago Press.
J. 14:91-111.
A@. Wildl. J. 10: 21 1-227.
zool. SOC. Lond. No. 45: 106-12 1 .





























