Embed Size (px)
Citation preview

SUBJECT: Non-polychaete B’98 problem animals
GUEST SPEAKER: none
DATE: 18 October 1999
TIME: 9:30 a.m. to 3:30 p. m.
LOCATION: City of San Diego Marine Biology Lab4918 N. Harbor Dr. #201San Diego, CA 92106
FUNDS FOR THIS PUBLICATION PROVIDED, IN PART BYTHE ARCO FOUNDATION, CHEVRON, USA, AND TEXACO INC.
SCAMIT Newsletter in not deemed to be valid publication for formal taxonomic purposes.
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
Two October Meetings will be held, bothaddressing Bight’98 problems and specimens.The first, on non-polychaete topics, in SanDiego on October 18; and the second onpolychaete topics at the Worm Lab at the LosAngeles County Museum of Natural History onOctober 25. Be prepared to roll up your sleevesand dig deeply into our problem taxa. We willbe done shortly with sample identification, andQC re-identification will begin in earnest.
OPISTHOBRANCH WORKSHOP
In June of this year the InternationalOpisthobranch Workshop was held in Menfi,Italy. Commentary on the meetings fromparticipants and photographs taken there areavailable on the web as a thread in the Sea SlugForum (run by Dr. Bill Rudman) at
http://www.austmus.gov.au/science/division/invert/mal/forum/menfwkshp.htm .
Arcidae, B’98 station 2425 (Mission Bay),24 July 98, 3.7 m Image by K. Barwick

2
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
The abstracts of the presentations have alsobeen made available on-line at
http://www.futuralink.it/vannarotolo .
NEW LITERATURE
Nemerteans have proven to have a difficulttaxonomy in many ways. Not the least of theseis a basic disagreement between nemerteanspecialists on which characters have meaningin separating taxa. This provides a field ripe fora molecular based approach which can be usedto either validate or refute morphologicallybased taxonomic hypotheses. Sundberg & Saur(1998) provide just such an analysis, dealingwith lineids in the genera Riseriellus, Lineus,and Micrura. The outcome of theirexamination is directly applicable to localnemertean taxonomy as they found both“genera” Lineus and Micrura to bepolyphyletic. Our local Lineus bilineatus is oneof several species ascribed to the genus whichare not in the same clade as the type speciesLineus longissimus. If this analysis isconfirmed by others, lineus will require a newgenus or genera. We assume that the local formidentified as L. bilineatus is the same as theEuropean form of that name included in thepresent analysis. The authors similarly foundMicrura species to be scattered betweenseveral clades, but as no local species wereincluded in the analysis it is unclear how localmembers of the genus would be aligned.Further analysis of a broader spectrum of themany species described in each of these generawill be required to more completely resolve theissue.
Larsen & Wilson (1998) shine a similar, ifmorphologically based, light on some of thecharacters currently considered as significant indefining families within the tanaids. Theyfound, for instance that the number of uropodalarticles in their new species was ambiguousbecause of partial fusion. The authors mentionthat tanaid family definitions are also difficultto apply because of problems in use of seta/
spine differentiations. Sieg, who is largelyresponsible for the current familialarrangement and definitions, did not use theseta/spine distinctions proposed by Watling inhis family definitions, and efforts to do so leadto problems. The senior author is attempting todevelop a better and more consistent separationbetween tanaidomorph families. In the presentpaper the family Paratanaidae has beenredefined, and the status of the others declaredunsatisfactory and in urgent need of re-evaluation.
As in a paper mentioned in the last issue themethodology of cladistics is under continualreview. Jenner (1999) and Jenner & Schram(1999) continue this trend and critically reviewthe assumptions underlying several cladisticmethodologies. The different results presentedby workers using different sets of assumptionsand different methods should hardly besurprising. It is however a bit disappointing tosee in black and white just how methoddependant the “objective and testable”approach of cladistics can be. Instead of a finalgreat truth we are presented with a series ofsuccessively better approximations of theundisputed truth as methodological problemsare discovered and counteracted. As anunregenerate “traditional” morphologist I mustadmit some satisfaction in having the clay feetof the cladistic god highlighted in this fashion.I have no doubt that as the methods mature andthe weak or inappropriate ones are weeded out,the hope of a less subjective systematictechnique will finally be realized. The authorspoint out quite effectively that we ain’t thereyet, especially for phylogenetic reconstruction.
The introduced seaweed Sargassum muticumhit our shores in the 70’s, and rapidly spreadalong the coast. It provided a new substrate fora host of small associates, mostly peracaridsbut also mollusks, bryozoans, urochordates,nemerteans, etc. Viejo (1999) examines thesame algal species in northern Spain, where itis also introduced. She compares the biota ofthe invasive species with that of two local

3
September, 1999 Vol. 18, No.5SCAMIT Newsletter
seaweeds, one with similar and the other withdissimilar morphologies. The local associatedmicrofauna proved well able to colonize anduse the invading alga, and in areas wheremacroalgae had been in low abundance, theSargassum provided additional habitat. AsSargassum also competes with native algae forspace and light, this is not an unmixedblessing. In the spanish case the associates ofthe native algae proved fickle, switching to theinvader without compunction. Anecdotalobservations suggest that the local situation issimilar, with indigenous grazers readilyswitching to the introduced alga.
Invasions do not always end so happily for theecosystem involved. Ruiz et al. (1999) examinethe more stressful interactions between theinvader and the invaded. Using the invaders ofChesapeake Bay as a database, they found that1 in 5 invading species had apparently had asignificant impact on the Bay ecosystem, orone or more of it’s components. They alsodoubt that the behavior of an invader in theBay can be used to predict it’s effect elsewhere.The potential differences in communitycomposition and function are too great betweenlocations to allow easy application ofexperience in one area to another. They alsoclearly perceived non-indigenous species asadditional stressors to alreadyanthropogenically stressed near-shorecommunities. As such their impacts, ifnegative, could be magnified by preexistingstress on the community from these othersources.
Whether or not a species uses larval forms fordispersal is a part of the “life-history strategy”of that species; what it does to insure itspersistence and spread. This is not always asingle path which a species follows, as ourrecently mentioned examples of poecilogonyhave demonstrated. The consequences, bothpositive and negative, of producing larvae havenot often been examined from a practical pointof view; in essence a cost/benefit analysis ofthe larval method. Pechenik (1999) reviews the
question and provides a nice summation of thecosts and benefits of larval production. Thisgreatly simplifies the decisions you out thereneed to make in your search for alternativemodes of reproduction.
30 AUGUST MEETING
President Ron Velarde (CSDMWWD) openedthe business meeting promptly at 9:30a.m. atthe San Diego Marine Biology Lab. The firstorder of business was scheduling meetings forthe next two months. We will be continuingalong the same theme with problematicspecimens from the Bight’98 project.Meetings for non-polychaete topics will beheld in San Diego on September 13 andOctober 18. Meetings for polychaete topicswill be held at the Worm Lab at the LosAngeles County Museum of Natural History onSeptember 27 and October 25.
Sonya Foree from the City and County of SanFrancisco joined us for the day. During theafternoon session, Arleen Navarret (CCSF) andVictoria Diaz and Maricarmen Necoechea,(both from CICESE, Ensenada, BajaCalifornia) were also in attendance.
Larry Lovell reported that the Aphroditaproject is a “go”, and he is putting out a requestfor specimens. Larry, in conjunction withCheryl Brantley and Ron Velarde, will beinvestigating and clarifying the taxonomy ofAphrodita. He would like to get as muchmaterial as possible. It would be especiallyuseful to include wide size ranges. If you cansupply specimens of Aphrodita to Larry, pleaseeither ship them to him at Scripps or give him acall (858) 822-2818 to arrange transportation.
Larry also gave an update on the status of hiswork on the marine invertebrate collection atScripps. There is a lot of re-organization andcleaning up that is going on and still much tobe done. If anyone is interested in seeing the

4
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
collection and putting in a day to help organizea portion of the collection, please call Larry.Larry hopes to put out a projected schedule forthe organization of various taxa.
We then started our parade of problematicpolychaete specimens. Tony Phillips(CLAEMD) handed out a voucher sheet forMarphysa sp B which is synonymous with hisMarphysa sp HYP 1 Phillips 1999. Thisspecies was found at station 2151, Dana PointHarbor, with oyster shell hash, at a depth of 6meters. In this species, the five occipitaltentacles were short, reaching the anterior edgeof the prostomium. The branchiae started onsetiger 10 as a single filament and reached amaximum of three filaments on setigers 45through setiger 102.
Larry Lovell announced that he has contactedAndy Mackie regarding Paradoneis sp HYP 1and Paradoneis sp SD 1, and Andy has agreedto look at our specimens. Larry graciouslyvolunteered to collect specimens, send them toAndy, and also to prepare a voucher sheet onthis species.
Rick Rowe (CSDMWWD) then reported onfour curious specimens from Catalina Island,station 2081, collected at a depth of 50 meters.After some investigation, it was determinedthat they were Pilargidae genus A of Williams.
Next Kelvin Barwick (CSDMWWD) showedus a paraonid specimen, Aricidea (Acmira) spSD1 from Santa Cruz Island. He had foundthis species at three stations. This specimenhad a bluntly rounded prostomium, 12 pairs ofbranchiae starting on setiger 4, and 2 pygidialcirri were evident. There was reddish-brownpigment throughout the worm, and Kelvincommented that usually this pigment wasfaded. There were bristles on the modifiedsetae that could be mistaken for hoods. Kelvinproduced a voucher sheet for this species andposted it on the SCAMIT website.
Next Rick Rowe distributed a voucher sheet onMacrochaeta sp A. He had found this speciesat San Miguel Island (station 2469, 33 mdepth) and also at two ITP stations (I-34, 63 ftdepth and I-7, 171 ft depth). Cheryl Brantleyhad also found this species at San MiguelIsland (station 2490 at 75 m depth and station2491 at 95 m depth). This was a small animalwhich had unusually large setae. To viewmany of the characters for this species, such asthe segmentation, palp and branchial scars, andpapillation, stain the worm with alcian blue.
Rick also reported on a specimen he calledNephtys sp SD 3 that was collected at station2523, Santa Barbara Island. The dorsallamellae were enlarged similar to N. squamosaEhlers 1887.
Kelvin next showed us a specimen of Pista.After examining the specimen, we concludedthat it was Pista disjuncta, following thepresent convention for Pista identification. Adiscussion ensued about the difficulty offinding the long handled setae in specimens ofPista (present on setigers 1 and 2) which is adistinguishing characteristic for this genus. Itwas suggested that one may want to dissect atissue sample from the animal, clean the tissuewith a few drops of bleach, and then look forthe long handled setae. One must observe thetissue frequently before the setae also dissolvein the bleach. Kelvin suggested an alternatemethod; after removing the tissue, he preparesa wet mount using methyl salicylate as themounting medium. This clears the tissueenough to see the handles. This is the samemethod used for clearing flatworms.
Next we examined a specimen of Aricidea(Acmira) cf cerruti brought in by Kelvin. Hefound two specimens at the Channel Islandsstation 2523 at 106 m depth. The specimenhad a rounded prostomium; however, themedian antenna was missing. There were 16pairs of branchiae, and the modified setae werethe same as illustrated in Laubier 1967. We

5
September, 1999 Vol. 18, No.5SCAMIT Newsletter
concluded that this specimen was probably asmall Aricidea (Acmira) cerruti, but withoutthe median antenna for confirmation, Kelvinwill leave the identification as A(A.) cf cerruti.
Larry and Tony both brought in specimens ofChone sp SD 1 from El Segundo and Bightstation 2453. Kathy Langan confirmed theidentification; this species had also been foundat some Tijuana River stations as well asmonitoring stations offshore of San Francisco.See San Diego voucher sheet from February13, 1998.
We then examined a flabelligerid from SanDiego Bay brought in by Rick Rowe. It wascollected at station 2231 at 13 meters depth. Itwas sand encrusted, and anteriorly, it had a rowof 4 small papillae across the dorsum. Thesetae were very similar to Piromis sp A fideHarris 1985. It was decided to call thesePiromis sp SD 1.
Continuing with another species from SanDiego Bay, Kathy Langan showed us somespecimens of Scolelepis. These keyed out to S.texana in Blake 1996 and keyed out to S. sp SD1 in Lovell’s and Pasko’s spionid key ofNovember 1995. This species had shown up atseveral stations in San Diego Bay. There werea few character differences between thesespecimens and the description of S. texana, soit was decided to call these S. sp SD 1 sincethey more closely matched that description.
Kathy also showed us some Dipolydora fromstations 2472 (Santa Cruz Island, 25 m depth),2211 (Orange County Sanitation District, 41 mdepth), and station 2493 (Santa Cruz Island, 44m depth). These specimens were similar to D.bidentata but differed in some significantcharacters. The branchiae started on setiger 9instead of setiger 8 for D. bidentata. Theposterior hooded hooks were bidentate in thesespecimens in contrast to the posteriorunidentate hooded hooks in D. bidentata.Also, no needle packets were seen in thesespecimens. However, with alcian blue stain,we observed some structures in the posterior of
2 worms that at first glance resembled needlepackets. The structures were lobes containinga few to several pointed spines. Some of thepoints extended beyond the ends of the lobes.A voucher sheet will be prepared for this newspecies, Dipolydora sp SD 1.
Tony brought a specimen of Nereiphylla fromstation 2404 which turned out to beNereiphylla sp 3 fide Harris (=Nereiphylla spSD 1).
Ron brought a mystery phyllodocid from SantaCruz Island station 2518, 112 m depth. Heidentified it as Eulalia?. It had pigment circlesaround each segmental line on the dorsum.There was also pigment on the bases of theparapodia. No one had seen anything similarto this specimen before, so it remained asEulalia? for the time being.
We could not put off addressing the cirratulidsany longer, so Rick took the floor and startedoff with 2 specimens of Protocirrineris fromSan Diego Bay station 2226. Rick compared 2different specimens. They were similar in thatthey had no methyl green staining pattern, nospines, and did have multiple cirri on setigers 3and 4. One specimen was similar to P. sp A inthat it had compressed segments, and the otherspecimen had longer segments that were notcrowded. However, Protocirrineris sp A isusually found at deeper stations. Rick islooking for more specimens and for the timebeing will refer to these specimens as P. sp SD1.
There were several new species ofMonticellina that were discovered in theBight’98 study. Rick reviewed severalprovisional species that he had encountered.Most of these had distinguishing methyl greenstaining patterns. The first was Monticellina spSD 2 which is similar to M. elongata. Thisspecies was found in 80 m depth at the ChannelIslands. Rick next described M. sp SD 4 fromSan Diego Bay and noted the differencesbetween it and Blake’s M. serratiseta. It had:1) a light dorsal methyl green stain on the

6
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
prostomium and peristomium and 2) a smallamount of light banding on the anteriordorsum. Tony noted that he finds M.serratiseta at stations that are 80 m and deeper.Finally, M. sp SD 6 from San Diego Bay (15.6m depth) and offshore San Diego (73 m depth)was described by Rick and had dark stainingstripes ventrally on only a few posteriorthoracic setigers and an inflated posterior end.
The next cirratulid genus up for discussion wasAphelochaeta. Cheryl Brantley (CSDLAC)passed out two provisional voucher sheets.The first voucher sheet was for Aphelochaetasp LA 1, found off Santa Cruz Island and SantaRosa Island. A question was posed as towhether these were A. petersenae, and Rickoffered to examine some specimens and see ifhe could answer this question. The secondprovisional species was Aphelochaeta sp LA 2.Five specimens were found at station 2521(Santa Cruz Island) at 75 m depth. TonyPhillips had also reported Aphelochaetaspecimens matching the description of A. spLA 2 in samples from Santa Cruz Island station2515 at a depth of 102 m. There were 45individuals and he recorded them asAphelochaeta sp HYP1. In this species themethyl green staining pattern revealed anunstained “ring” between the prostomium andperistomium. This ring extended around theentire animal at the anterior end.
The presentation of novel Aphelochaetacontinued as Rick outlined the description ofAphelochaeta sp SD 3. There were 11specimens collected from Santa Cruz Island at25 m depth.
A discussion then ensued regarding A.petersenae, A. sp SD 4, and A. sp HYP 1. Aswe have been collecting and viewing more andmore of these animals, we have been noticingthat there are intermediate patterns in themethyl green staining. Rick will be examiningmore specimens with the possibility of lumpingthese provisional species.
13 SEPTEMBER MEETING
The meeting was called to order by PresidentRon Velarde at approximately 9:45 a.m.. Thefirst order of business Ron attended to wasdistributing City of San Diego B’98 sampleswhich had been selected for re-identificationfor the QA/QC aspect of the project.
Ron then passed around a flyer fromSCCWRP which listed upcoming seminars inmarine related topics. The flyer is included asan attachment in the paper version of thisnewsletter, or to those of you who have gone“electronic”, please go to SCCWRP’s website(www.sccwrp.org ) for information on theseseminars.
The current issue of the Festivus, theNewsletter/Journal of the San Diego ShellClub, was passed around as it contained anarticle on the Panamic pearl oyster, Pteriasterna in Carlsbad lagoon.
Megan Lilly (CSDMWWD) then brought up tothose present the idea of having more SCAMITt-shirts and hats created. It was pointed outthat a new silkscreen would need to be made.We decided to see what kind of response AnnDalkey receives for the few remaining itemsshe has in stock. If those sell-out, thenpotentially we will look into re-newing oursupply.
We briefly discussed the SCAMIT website.Even though it’s been said previously, Jay hascreated a beautiful and functional on-linepresence for SCAMIT, and we all owe him ourthanks. We have recently received a series ofnew memberships on-line, most from othercountries. An increasing percentage of websitevisits are also from other countries and itappears the site is now acting, in manyinstances, as an international hub for peoplesearching for marine related links. If that isindeed the case, then we have achieved a gooddeal of our objective of engaging a broaderaudience. [ Now, if they would just write andsubmit some items to the NL...Ed.].

7
September, 1999 Vol. 18, No.5SCAMIT Newsletter
We decided to finally turn our attention toanimals and the first ones that caught our“eyes” were two strange looking crabs broughtby Carol Paquette. The first animal wasCardisoma crassum which is normally found inriver banks in mangrove/estuarine typehabitats in southern Mexico and CentralAmerica; however, this animal was collectedon a heat treatment screen at the ScattergoodGenerating Station in El Segundo. The coolingwater intake, where the animal was collected,is at a depth of 7 m. The animal appeared tohave been alive just prior to being taken basedon its color and condition of its carapace,appendages, etc.. This posed an interestingquestion since this was obviously not a marinecrab. When identifed by Todd Zimmerman(NHMLAC), he was surprised to hear of thelocality at which it was collected. Somesuggestions were that it was a pet storepurchase that was mistakenly “returned to thesea” or perhaps it was dropped from the beakof a hungry gull (said with less seriousness).Whatever the answer, it remains a mystery atthis time. Despite the presence of the animal ina live state at time of capture, it does not, andcould never range into this area. Appropriatehabitat is completely lacking in SouthernCalifornia although an ersatz habitat in aterrarium could be constructed. Much like theoccasional Maine lobsters which escape fromexperimental aquaculture rearing in SouthernCalifornia, and find their way into benthic ortrawl samples, presence of the species in ourlocal waters is completely accidental.
The second crab was not such a mystery, moreof a rarity. It was Euphylax dovii taken againon a heat treatment screen but this time at theSan Onofre power plant. This animal is notusually seen in nearshore collections, being anoceanic species, but has been previously seenand reported from our area.
After much “oohing and ahhing” over the bigcrabs we turned our rapt attention to themollusks. Kelvin Barwick (CSDMWWD) hadbeen working on a sample from Mission Bay
(Station 2425) that was “full” of interesting andunusual (for those of us accustomed to off-shore work) mollusks. The following animalsdiscussed are all from this station.
First Kelvin brought forth a small gastropodwhich turned out to be Barleeia subtenuis. Theanimal was present in high densities (748individuals in the rep). This tiny rissoid is verycommon in Mission Bay, as is its congener B.californicus, grazing on the diatom filmcovering the sandy bay bottom, or on thediatom growth on attached algae and/orseagrass in the bay.
There was a slightly different lookingTurbonilla from this station as well, but it wasdecided to leave it at Turbonilla sp. due to thenomenclatural problems still clinging to thisgenus in local waters although Kelvin willcompare it with the species described andillustrated in the pyramidellid monograph ofDall and Bartsch. Dr. Jim McLean says hethinks our pyramidellid problems have beenresolved in his draft monograph on thegastropods of California, but it is not yetavailable.
Next was a Lirularia parcipictum which theSan Diego lab had not previously seen. Thespecies ID was based on the presence of a basalcarina, and the nature of the spiralornamentation of the whorls. The examinedspecimens were also rather low spired for thespecies.
A juvenile arcid clam was brought forth andnot immediately recognized by those present.It was considered a probable Anadara, but wasunfamiliar to all present. A digital image hasbeen sent to Paul Scott and further ID is stillpending at this time (see cover photo).
Next, a juvenile Leporimetis obesa. Again, ananimal not normally seen by the San Diego labin their standard ocean monitoring program,this, combined with its small size, threw themfor a loop. As with many other bivalves, thejuvenile does not look much like the adult.

8
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
Uneroded adults do, however, carry aroundtheir developmental history in their shells, andprovide the necessary evidence to connect thejuveniles and adults.
Two more juvenile clams were brought forth,one turned out to be a very small Semelevenusta and the other, a young Cumingiacalifornica. All of these animals can, onoccasion, be taken offshore if the sedimentparticle mix and organic load are appropriate,but are much more commonly found in bays,harbors, and estuaries.
Tony Phillips (CLAEMD) brought threevariant forms of Ophiodermella to beexamined by Ron Velarde (CSDMWWD).After much examination and discussion it wasdecided that the three animals were tentativelyall Ophiodermella inermis with slightvariations in color and sculpture. Theseanimals will be checked by Dr. James McLeanat LACMNH.
Megan Lilly (CSDMWWD) then brought forththe ugly question of Lirobittium. She hadexamples of various forms of the genus fromdifferent B’98 stations as well as the standardanimal that the City of San Diego sees in itsregular monitoring stations and callsLirobittium larum. CSDLAC as well asHyperion both call their common formLirobittium rugatum There was somediscussion as to standardizing the various labapproaches to this animal and no conclusionwas reached. The animals brought by Meganwere left at Lirobittium sp. for the time being.The question may be addressed at a latermeeting on gastropods with Hank Chaney ofthe Santa Barbara Museum of Natural History.We have already considered species ofLirobittium at a previous meeting with PaulScott at Santa Barbara, but without muchconsensus on the boundaries of the taxa we see.
The afternoon started (and ended) withCrustacea. Dean Pasko (CSDMWWD)brought forth a strange little animal found inone of the Channel Islands samples. The
animal was recognized by Don Cadien as beinga harpactacoid copepod of the genusScutellidium. The examined specimen had ametallic sheen to its carapace.
Although we had intended to consider anumber of taxa within the amphipod familyOedicerotidae, including several provisionalforms erected by Dean Pasko from San Diegosamples, we got stuck on the Synchelidium.The question of proper generic usage wasraised again. Don Cadien referred to his earlierarguments for not adopting the genusAmerichelidium as proposed by Bousfield andChevrier (1996), but suggested that thecorrection indicated by Bousfield (1997) mightbe adequate to address the problems posed bythe defective generotype selection. Heinformed us that the draft Catalogue of NorthAmerican Aquatic Invertebrates was usingAmerichelidium, and that by the publication ofthe SCAMIT Ed. 4 listing, we might need tochange our current position on the subject(rejection of Americhelidium).
This genus (whether we use Americhelidium orSynchelidium) has always proved to betroublesome. Shoemaker grappled with it, andpassed it off to J. L. Barnard prior to his death.Barnard struggled with it for quite awhile,parsing out the intertidal micropleon, andleaving behind manuscript names for threeother species. None of us were willing to tacklethe problem which would involve criticalexamination of a large body of material fromthe Allan Hancock collections identified as hismanuscript species by J. L. Barnard. This isaccessible, now being on the shelves of theNatural History Museum of Los AngelesCounty. Some desultory attempts were madebut no real progress. Then Amphipacifica Vol.2 No. 2 arrived in May of 1996 and we had theBousfield and Chevrier attempt to make senseof these animals. They retained the two Millsspecies - rectipalmum and shoemakeri, andadded another four from the Eastern Pacific -millsi, pectinatum, setosum, and variabilum.The key they included did not deal well with

9
September, 1999 Vol. 18, No.5SCAMIT Newsletter
the separation of the animals, and was critiquedby Don Cadien in NL Vol 15(6) [October1996]. A replacement key was “inprogress” but has never been sent out.
Dissatisfaction was expressed over the natureof the characters used in both the Bousfield andChevrier key, and in the pictorial key whichDon Cadien had constructed in development ofa replacement key. Sex linked characters suchas the length of the third article of themandibular palp were considered poor forgeneral use, and were to be avoided if at allpossible. Likewise characters which weredifficult to distinguish, such as the shape andrelative posterior extension of the coxae andpleonal epimera, the relative shape and size ofthe articles of the third leg, and the proportionsof the maxilliped outer plate were deemed toodifficult in application to provide a viable key.
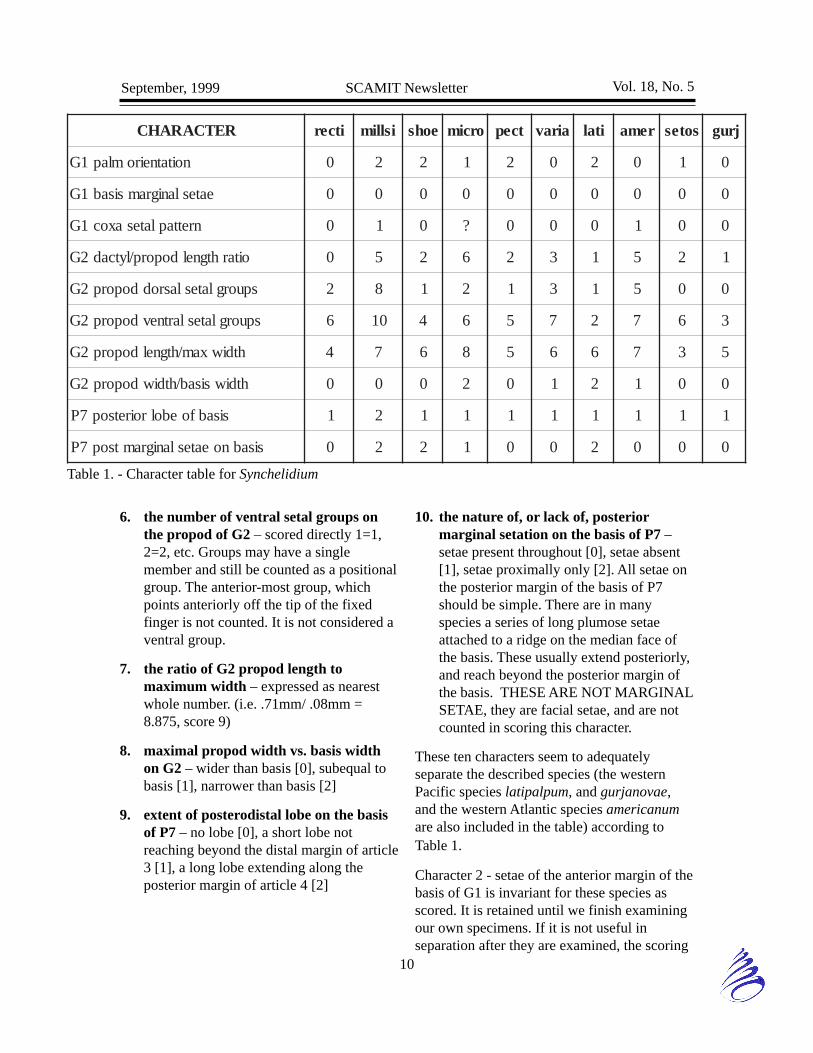
In essence, we stepped back a pace and startedover. Between the members present at themeeting we agreed to try using a series ofcharacters which we felt could be repeatablyevaluated. These were: 1) the orientation of theG1 palm; 2) the setation of the anterodistalmargin of the basis of G1; 3) the pattern ofsetae on the ventral margin of coxa 1; 4) theratio of dactyl to propod length of G2; 5) thenumber of dorsal setal groups on the propod ofG2; 6) the number of ventral setal groups onthe propod of G2; 7) the ratio of G2 propodlength to maximum width; 8) maximal propodwidth vs. basis width on G2; 9) extent ofposterodistal lobe on the basis of P7; and 10)the nature of, or lack of, posterior marginalsetation on the basis of P7. Don Cadien alsosuggested characters based on the spine andserration pattern of the uropods had value, butwere not well enough known as regardsvariability between individuals, sexualdimorphism, and ontogenic change for currentapplication.
We scored each of these characters as follows:
1. G1 palmar orientation – transverse [0],oblique [1], intermediate “can’t decide”[2]. This seemingly simple decision as tothe orientation of the palm has proven to bequite difficult in practice. There isconsiderable perceptual difference betweenindividuals in how a particular palm shouldbe scored. In some cases there is anunequivocally transverse palm, but innumerous other cases a palm that istransverse at the hinge may taper off intoobliquity before it joins the hind margin ofthe propod. Depending on just where thistakes place such a palm could be scored asany of the above states. We must constrainourselves to only the most clear cut casesfor scores of 0 or 1, and place all othermore problematic structures in 2.
2. the setation of the anterodistal margin ofthe basis of G1 – strongly setose, with 4-10+ setae, usually long [0]; weakly setose,with 1-3 setae, usually very short [1]; setaelacking [2]
3. the pattern of setae on the ventralmargin of coxa 1 – setae markedly longerat posteroventral edge of coxa [0], or setaeof posteroventral edge the same length aselsewhere on the ventral margin of coxa [1](this includes cases where long and shortsetae are interspersed along the entireventral margin of the coxa)
4. the ratio of dactyl to propod length inG2 – 1/3 [0], 1/4 [1], 1/5 [2], 1/6 [3], 1/7[4], 1/8 [5], 1/9 [6]. These are all roundedto the nearest choice based on opticalmicrometer measurements of the lengths ofdactyl and propod.
5. the number of dorsal setal groups on thepropod of G2 – scored directly 1=1, 2=2,etc. Groups may have a single member andstill be counted as a positional group. Thegroup at the base of the dactyl is notcounted, as it is present in all species.
10
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
6. the number of ventral setal groups onthe propod of G2 – scored directly 1=1,2=2, etc. Groups may have a singlemember and still be counted as a positionalgroup. The anterior-most group, whichpoints anteriorly off the tip of the fixedfinger is not counted. It is not considered aventral group.
7. the ratio of G2 propod length tomaximum width – expressed as nearestwhole number. (i.e. .71mm/ .08mm =8.875, score 9)
8. maximal propod width vs. basis widthon G2 – wider than basis [0], subequal tobasis [1], narrower than basis [2]
9. extent of posterodistal lobe on the basisof P7 – no lobe [0], a short lobe notreaching beyond the distal margin of article3 [1], a long lobe extending along theposterior margin of article 4 [2]
10. the nature of, or lack of, posteriormarginal setation on the basis of P7 –setae present throughout [0], setae absent[1], setae proximally only [2]. All setae onthe posterior margin of the basis of P7should be simple. There are in manyspecies a series of long plumose setaeattached to a ridge on the median face ofthe basis. These usually extend posteriorly,and reach beyond the posterior margin ofthe basis. THESE ARE NOT MARGINALSETAE, they are facial setae, and are notcounted in scoring this character.
These ten characters seem to adequatelyseparate the described species (the westernPacific species latipalpum, and gurjanovae,and the western Atlantic species americanumare also included in the table) according toTable 1.
Character 2 - setae of the anterior margin of thebasis of G1 is invariant for these species asscored. It is retained until we finish examiningour own specimens. If it is not useful inseparation after they are examined, the scoring
RETCARAHC itcer isllim eohs orcim tcep airav ital rema sotes jrug
noitatneiromlap1G 0 2 2 1 2 0 2 0 1 0
eateslanigramsisab1G 0 0 0 0 0 0 0 0 0 0
nrettaplatesaxoc1G 0 1 0 ? 0 0 0 1 0 0
oitarhtgneldoporp/lytcad2G 0 5 2 6 2 3 1 5 2 1
spuorglateslasroddoporp2G 2 8 1 2 1 3 1 5 0 0
spuorglateslartnevdoporp2G 6 01 4 6 5 7 2 7 6 3
htdiwxam/htgneldoporp2G 4 7 6 8 5 6 6 7 3 5
htdiwsisab/htdiwdoporp2G 0 0 0 2 0 1 2 1 0 0
sisabfoebolroiretsop7P 1 2 1 1 1 1 1 1 1 1
sisabnoeateslanigramtsop7P 0 2 2 1 0 0 2 0 0 0
Table 1. - Character table for Synchelidium

11
September, 1999 Vol. 18, No.5SCAMIT Newsletter
will be redefined to better discriminatebetween species. Please note the illustration ofBousfield and Chevrier of shoemakeri shows acondition which would have been scored as 1in this character. In Mills original description,however, the illustrated condition was scoredas 0 and this was used in the table.
Comparison of specimens with the charactertable should yield presumptive identifications.These should be checked against thedescriptions and illustrations of the species inBousfield and Chevrier (1996) and, in the caseof rectipalmum and shoemakeri, against theoriginal descriptions of Mills (1962), which donot agree in all particulars with the laterreports. The SCAMIT voucher sheet forSynchelidium rectipalmum was found todisagree with the original description in theconfiguration of the lobe on the basis of P7,and should be used with caution.
Our discussion of oedicerotids, as well as othernon-polychaete groups will continue at themeeting scheduled for 18 October at the SanDiego lab. By then we should all have beenable to apply the above character table to ourspecimens, and have comments to make abouttheir applicability and validity as separatorytools. If you find them wanting, try and comeup with suggested alternatives by the meeting.
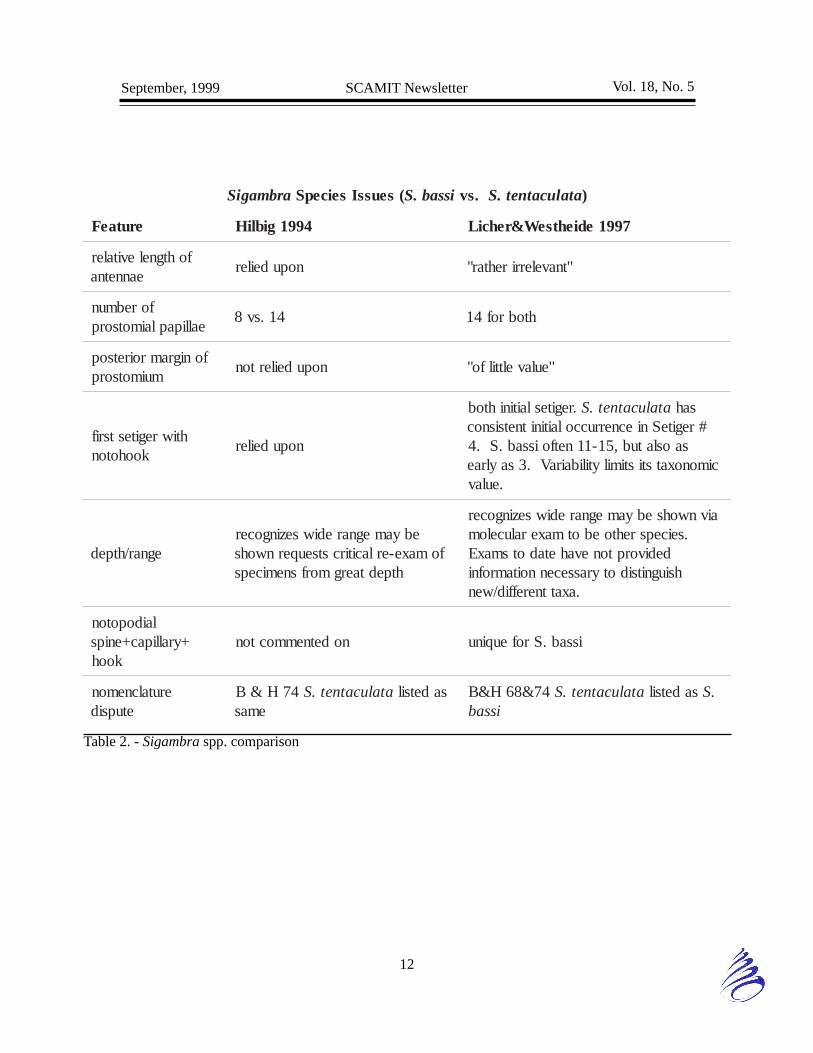
Sigambra ALERT
- Tom Parker (CSDLAC)
Licher & Westheide (1997) review thedescriptions and taxonomy of Sigambra bassiand S. tentaculata. Previously local workershave relied upon various features to identifySigambra species. The use of soft tissuefeatures such as median antennal lengthrelative to lateral antennal length, prostomialmargin shape, and papillae on the proboscishave been utilized. Hard features such as thefirst occurrence of setigers with hooked setaehave also been counted upon as finaldeterminations of whether S. tentaculata, bassi,or setosa specimens were present.
The following brief table (see Table 2., pg 12)is a synopsis comparing the relevantdetermining factors used in the MMS Atlasversus those reviewed by Licher & Westheide.It seems likely that a local review of practicesand specimens is needed to standardize ouridentifications.
FOLLOW UP
The specimen of the crab genus Palicus seen atthe meeting on 16 August has been furtherexamined and is an example of Palicus lucasiifrom California. The first reported specimenfrom local waters was taken in 1994 byCSDLAC off Palos Verdes. Reexamination ofthat specimen with the comparative material ofthe second specimen, and additional materialfrom the Galapagos, Panama, and the Gulf ofCalifornia has shown the original ID as P.lucasii to be incorrect. This second specimen,like the first, is a male. Examination of themale pleopods by Dr. Todd Zimmerman(NHMLAC) confirmed the identity of thesecond specimen, which was taken in 124 ft. ofwater at ITP station 2101 off Imperial Beach.The identify of the first specimen from offPalos Verdes was later established by Todd asP. cortezi (Crane 1937), originally describedfrom the Gulf of California. This is a newrecord for California, and will be added to theemendations to be made to the 3rd Edition ofthe SCAMIT list. Many thanks to Todd for hisefforts in clarifying the identity of thesedifficult animals.
MORE CHANGE
Our coast has two representatives of thesynopiid amphipod genus Tiron, or it used to.It has finally been recognized that the presenceof a mandibular palp in T. biocellata, and it’sabsence in T. tropakis, is of no smallsignificance. Reference to Barnard & Karaman(1991) shows both species still listed underTiron. The same authors, however, include as avalid generic level taxon MetatironRabindranath 1972. As pointed out by Thomas(1993), Barnard & Karaman failed to reallocate
12
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
arbmagiS (seussIseicepS issab.S .sv atalucatnet.S )
erutaeF 4991gibliH 7991ediehtseW&rehciL
fohtgnelevitalereannetna
nopudeiler "tnavelerrirehtar"
forebmuneallipaplaimotsorp
41.sv8 htobrof41
fonigramroiretsopmuimotsorp
nopudeilerton "eulavelttilfo"
htiwregitestsrifkoohoton
nopudeiler
.regiteslaitinihtob atalucatnet.S sah#regiteSniecnerruccolaitinitnetsisnoc
saoslatub,51-11netfoissab.S.4cimonoxatstistimilytilibairaV.3saylrae
.eulav
egnar/htpedebyamegnarediwsezingocer
fomaxe-erlacitircstseuqernwohshtpedtaergmorfsnemiceps
aivnwohsebyamegnarediwsezingocer.seicepsrehtoebotmaxeralucelom
dedivorptonevahetadotsmaxEhsiugnitsidotyrassecennoitamrofni
.axattnereffid/wen
laidopoton+yrallipac+enips
koohnodetnemmocton issab.Srofeuqinu
erutalcnemonetupsid
47H&B atalucatnet.S sadetsilemas
47&86H&B atalucatnet.S sadetsil S.issab
Table 2. - Sigambra spp. comparison

13
September, 1999 Vol. 18, No.5SCAMIT Newsletter
species originally described as Tiron intoMetatiron. One of the salient differencesbetween the two genera is the mandibular palp,which is absent in members of Metatiron. Inconsequence we must recognize that “T.”tropakis belongs in Metatiron, while T.biocellata is properly allocated to Tiron.Barnard explicitly mentioned the absence of amandibular palp in his original description ofTiron tropakis in 1972. In his originaldescription of T. biocellata he indicated that themouthparts were the same as T. spiniferum, thetype of Tiron, with but two exceptions. Themandibular palp was neither illustrated nordescribed, but was not listed as an exception(Barnard 1962). Examination of locallycollected T. biocellata and M. tropakis haveconfirmed the presence of a palp in the former,and its absence in the latter. Jim Roney (HYP)had mentioned this difference in a TaxonomicList Server communication earlier this year, buteither did not appreciate (as I had not), ordidn’t mention the consequent separation at thegeneric level. Edition 3 of the SCAMITTaxonomic Listing is in error as regardsMetatiron tropakis (Barnard 1972); acorrection will be made in Ed. 4. - Don Cadien(CSDLAC)
NEW ISOPOD
Hi all,Just a quick “heads up” or “look out” regardingSouthern California Bight Edotia. Thereappears to be a third species of Edotia in theSCB that we are tentatively calling EDOTIA SPSD 1 in our database. The specimens werecollected in 60 m of water from one of theBight’98 Channel Island stations. The speciesresembles E. sublittoralis very closely,especially in terms of pleonal morphology —i.e., they have an inflated pleon (or pleotelson)with large dorsal swellings rather then the non-inflated pleon with a distinct transverse ridge(carina) characteristic of Edotia sp B. In otherwords, they would key to E. sublittoralis in thekey I distributed some time ago. However, thespecimens looked a little “different” and were
from relatively deep water more characteristicof the habitat of E. sp B. Consequently, Iexamined the pereopods of the “new” speciesand they appear to be distinct from either E.sublittoralis or E. sp B (the pereopods of thesetwo species are quite distinct from each other).
I will try and put out a sheet soon describingthese differences and any others that maybecome apparent. In the meantime, I wouldappreciate it if you could pull any “suspect”specimens and send them to me for additionalexamination. I would consider suspect any“sublittoralis” occurring at depths > about 45m — actually perhaps any Channel Islandcritters in general.
Finally, I am trying to complete my paperdescribing E. sp B and redescribing E.sublittoralis, although I guess I’ll now add the3rd species as well. However, I still haven’thad the opportunity to examine any Edotia spB specimens from waters north of the SanDiego area. Consequently, any specimens oreven location info (i.e., I need a Northern rangelimit) would be appreciated.
Thanks,Tim
Timothy D. StebbinsCity of San Diego Marine Biology Laboratory4918 North Harbor Drive, Suite 101San Diego, CA 92106 USATel: (619) 692-4900; Fax: (619) 692-4902E-mail: [email protected]
JOB OPPORTUNITY
I recently received the following e-mail fromMaggie Dutch and am posting this listing forany of you who may be interested (I for one,would love to live in Washington - M. Lilly,Secretary)
Hi Megan,I work with the Washington State Departmentof Ecology’s Marine Monitoring

14
September, 1999 Vol. 18, No. 5SCAMIT Newsletter
Unit. Our group conducts both sediment andwater column monitoring throughout PugetSound, as part of the Puget Sound AmbientMonitoring Program. The state Department ofPersonnel has recently opened theEnvironmental Specialist 2, 3, 4, and 5registers from which we may hire newemployees. Although we currently have nopositions for which we are hiring, we may inthe near future. I was hoping that you might beable to place a notice in the SCAMITnewsletter indicating that we are interested inhaving folks get on these registers who have 1)strong sediment chemistry, bioassay, andinfaunal monitoring experience, 2) strongstatistical/data analysis and report writingskills, and 3) an interest in moving to the stateof Washington (rain and all :-) !!!).
Anyone with the interest and qualifications cancontact me and send a resume (address, phone,and e-mail below). To get on theEnvironmental Specialist (ES 2 through 5)registers (you must be on the registers to behired), an application should be submitted toour state personnel office:
BIBLIOGRAPHY
Banse, Karl & Katharine D. Hobson. 1968. Benthic Polychaetes from Puget Sound Washington,with Remarks on Four Other Species. Proceedings of the United States NationalMuseum 125:1-52.
—. 1974. Benthic Errantiate Polychaetes of British Columbia and Washington. Bulletin of theFisheries Research Board of Canada 185:1-111.
Barnard, J. Laurens. 1962. Benthic marine Amphipoda of southern California: families Tironidaeto Gammaridae. Pacific Naturalist 3(2):73-115.
—. 1972. A Review of the Family Synopiidae (= Tironidae), Mainly Distributed in the Deep Sea(Crustacea: Amphipoda). Smithsonian Contributions to Zoology 124:1
—, & Gordan S. Karaman. 1991. The Families and Genera of Marine Gammaridean Amphipoda(Except Marine Gammaroids). Records of the Australian Museum Supplement 13:1-866.
Blake, James A. 1996. Chapter 4. Family Spionidae Grube, 1850. Pp. 81-223 IN: Blake, JamesA., Brigitte Hilbig, and Paul H. Scott (eds.) Taxonomic Atlas of the Benthic Fauna of theSanta Maria Basin and Western Santa Barbara Channel. Vol. 6, The Annelida Part 3.Polychaeta: Orbiniidae to Cossuridae. Santa Barbara Museum of Natural History, SantaBarbara, Ca. 418pp.
Washington State Department of PersonnelApplications Unit600 South FranklinPO Box 47561Olympia, WA 98504-7561
Information regarding these open registers, andan electronic version of the state applicationform can be found on Ecology’s EmploymentOpportunity web site:
http://www.wa.gov/ecology/es/jobs.html
Maggie DutchSupervisor, Marine Monitoring UnitWashington State Department of Ecology300 Desmond Drive PO Box 47710Olympia, WA [email protected]

15
September, 1999 Vol. 18, No.5SCAMIT Newsletter
Bousfield, Edward L. 1997. Text Errata. Vol. II(2). (Melitidae and Oedicerotidae). Amphipacifica2(3):140.
Bousfield, Edward L., and Andrée Chevrier. 1996. The amphipod family Oedicerotidae on thePacific coast of North America. Part 1. The Monoculodes and Synchelidium genericcomplexes: systematics and distributional ecology. Amphipacifica 2(2):75-148.
Ehlers, E. 1887. Report on the annelids of the dredging expedition of the U.S. coast surveysteamer BLAKE. Museum of Comparative Zoology, Harvard. Memoir 15:1-335
Hilbig, Brigitte. 1994. Chapter 9. Family Hesionidae. Pp. 243-270 IN: Blake, James A., &Brigitte Hilbig (eds.). Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basinand Western Santa Barbara Channel. Volume 4, The Annelida Part 1. Oligochaeta andPolychaeta: Phyllodocida (Phyllodocidae to Paralacydoniidae). Santa Barbara Museumof Natural History, Santa Barbara, Ca. 377pp.
Jenner, Ronald A. 1999. Metazoan phylogeny as a tool in evolutionary biology: Currentproblems and discrepancies in application. Belgian Journal of Zoology 129(1):245-261.
—, & Frederick R. Schram. 1999. The grand game of metazoan phylogeny: rules and strategies.Biological Reviews of the Cambridge Philosophical Society 74(2):121-142.
Larsen, K. & George D. F. Wilson. 1998. Tanaidomorphan systematics - Is it obsolete? Journalof Crustacean Biology 18(2):346-362.
Licher, Frank & Wilfried Westheide. 1997. Review of the genus Sigambra (Polychaeta:Hesionidae), redescription of S. bassi (Hartman, 1947), and descriptions of two newspecies from Thailand and China. Steenstrupia 23:1-20.
Mills, Eric L. 1962. Amphipod crustaceans of the Pacific coast of Canada: II. FamilyOedicerotidae. Natural History Papers, National Museum of Canada 15:1-21.
Pechenik, Jan A. 1999. On the advantages and disadvantages of larval stages in benthic marineinvertebrate life cycles. Marine Ecology Progress Series 177:269-297.
Rabindranath, P. 1972. Three species of gammaridean Amphipoda (Crustacea) from theTrivandrum Coast, India. Zoologischer Anzeiger 188:84-97.
Ruiz, Gregory M., Paul Fofonoff, Anson H. Hines, & Edwin D. Grosholz. 1999. Non-indigenous species as stressors in estuarine and marine communities: Assessing invasionimpacts and interactions. Limnology and Oceanography 44(3):950-972.
Sundberg, Per & Marianne Saur. 1998. Molecular phylogeny of some Europeanheteronemertean (Nemertea) species and the monophyletic status of Riseriellus, Lineus,and Micrura. Molecular Phylogenetics and Evolution 10(3):271-280.
Thomas, James Darwin. 1993. Identification Manual for the Marine Amphipoda: (Gammaridea)I. Common Coral Reef and Rocky Bottom Amphipods of South Florida. Final Report tothe Florida Department of Environmental Protection, Contract SP290. 80pp.
Viejo, Rosa M. 1999. Mobile epifauna inhabiting the invasive Sargassum muticum and two localseaweeds in northern Spain. Aquatic Botany 64(2):131-149.

September, 1999 Vol. 18, No.5SCAMIT Newsletter
SCAMIT OFFICERS:
If you need any other information concerning SCAMIT please feel free to contact any of theofficers e-mail addressPresident Ron Velarde (619)692-4903 [email protected] Leslie Harris (213)763-3234 [email protected] Megan Lilly (619)692-4901 [email protected] Ann Dalkey (310)648-5544 [email protected] issues of the newsletter are available. Prices are as follows:
Volumes 1 - 4 (compilation)................................. $ 30.00Volumes 5 - 7 (compilation)................................. $ 15.00Volumes 8 - 15 ................................................ $ 20.00/vol.
Single back issues are also available at cost.
Please visit the SCAMIT Website at: http://www.scamit.org





























