Embed Size (px)
DESCRIPTION
.
Citation preview

ARTICLE IN PRESS
0147-6513/$ - se
doi:10.1016/j.ec
�CorrespondConservation a
India. Fax: +9
E-mail addr
Ecotoxicology and Environmental Safety 60 (2005) 324–349
www.elsevier.com/locate/ecoenv
Salt tolerance and salinity effects on plants: a review
Asish Kumar Paridaa, Anath Bandhu Dasa,b,�
aNational Institute for Plant Biodiversity Conservation and Research, Nayapalli, Bhubaneswar 751015, Orissa, IndiabRegional Plant Resource Centre, Nayapalli, Bhubaneswar 751015, Orissa, India
Received 18 September 2003; received in revised form 8 March 2004; accepted 8 June 2004
Available online 7 August 2004
Abstract
Plants exposed to salt stress undergo changes in their environment. The ability of plants to tolerate salt is determined by multiple
biochemical pathways that facilitate retention and/or acquisition of water, protect chloroplast functions, and maintain ion
homeostasis. Essential pathways include those that lead to synthesis of osmotically active metabolites, specific proteins, and certain
free radical scavenging enzymes that control ion and water flux and support scavenging of oxygen radicals or chaperones. The
ability of plants to detoxify radicals under conditions of salt stress is probably the most critical requirement. Many salt-tolerant
species accumulate methylated metabolites, which play crucial dual roles as osmoprotectants and as radical scavengers. Their
synthesis is correlated with stress-induced enhancement of photorespiration. In this paper, plant responses to salinity stress are
reviewed with emphasis on physiological, biochemical, and molecular mechanisms of salt tolerance. This review may help in
interdisciplinary studies to assess the ecological significance of salt stress.
r 2004 Elsevier Inc. All rights reserved.
Keywords: Antioxidative enzymes; Compatible solutes; Ion homeostasis; Photosynthesis; Salt stress
1. Introduction
Salinity is the major environmental factor limitingplant growth and productivity (Allakhverdiev et al.,2000b). The detrimental effects of high salinity on plantscan be observed at the whole-plant level as the death ofplants and/or decreases in productivity. Many plantsdevelop mechanisms either to exclude salt from theircells or to tolerate its presence within the cells. Duringthe onset and development of salt stress within a plant,all the major processes such as photosynthesis, proteinsynthesis, and energy and lipid metabolism are affected.The earliest response is a reduction in the rate of leafsurface expansion, followed by a cessation of expansionas the stress intensifies. Growth resumes when the stressis relieved. Carbohydrates, which among other sub-
e front matter r 2004 Elsevier Inc. All rights reserved.
oenv.2004.06.010
ing author. National Institute for Plant Biodiversity
nd Research, Nayapalli, Bhubaneswar 751015, Orissa,
1-674-2550274.
ess: [email protected] (A.B. Das).
strates are needed for cell growth, are supplied mainlythrough the process of photosynthesis, and photosynth-esis rates are usually lower in plants exposed to salinityand especially to NaCl.Salinity stress biology and plant responses to high
salinity have been discussed over two decades (Flowerset al., 1977; Greenway and Munns, 1980; Ehret andPlant, 1999; Hasegawa et al., 2000; Zhu, 2002) and it hasbeen over a decade since salinity tolerance in marinealgae has been covered (Kirst, 1989). These reviewscovered organismal, physiological, and the then-knownbiochemical hallmarks of stress and the bewilderingcomplexity of plant stress responses. We summarize inthis review physiological, biochemical, and molecularmechanisms of salt tolerance with the salient features ofsalinity stress effects on plants. In this review, muchresearch information about cellular, metabolic, molecu-lar, and genetic processes associated with the response tosalt stress, some of which presumably function tomediate salt tolerance, has been gathered.

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 325
2. Salt tolerance of plants
Salt tolerance is the ability of plants to grow andcomplete their life cycle on a substrate that containshigh concentrations of soluble salt. Plants that cansurvive on high concentrations of salt in the rhizosphereand grow well are called halophytes. Depending on theirsalt-tolerating capacity, halophytes are either obligateand characterized by low morphological and taxonomi-cal diversity with relative growth rates increasing up to50% sea water or facultative and found in less salinehabitats along the border between saline and nonsalineupland and characterized by broader physiologicaldiversity which enables them to cope with saline andnonsaline conditions.
2.1. Mechanism of salt tolerance
Plants develop a plethora of biochemical and mole-cular mechanisms to cope with salt stress. Biochemicalpathways leading to products and processes thatimprove salt tolerance are likely to act additively andprobably synergistically (Iyengar and Reddy, 1996).Biochemical strategies include (i) selective accumulationor exclusion of ions, (ii) control of ion uptake by rootsand transport into leaves, (iii) compartmentalization ofions at the cellular and whole-plant levels, (iv) synthesis
Extracellular and cell-wall space• Ion exclusion• Ion export• Cell-wall modification
Cytosol and org• Osmotic adjustment • Radical scavenging • Ion-selectivity changes • Enhancement of proton pumping • Aquaporin-activity control
Vacuolar space
• Ion (sodium, calcium) st• Ion (potassium) export• Osmotic adjustment• Proton-gradient mainten
Fig. 1. Biochemical functions associated with tolerance to plant salt stress. T
that are defined by the plasma membrane and tonoplast (reproduced from B
of compatible solutes, (v) change in photosyntheticpathway, (vi) alteration in membrane structure, (vii)induction of antioxidative enzymes, and (viii) inductionof plant hormones (Fig. 1). These are discussed underseparate headings.Salt tolerance mechanisms are either low-complexity
or high-complexity mechanisms. Low-complexity me-chanisms appear to involve changes in many biochem-ical pathways. High-complexity mechanisms involvechanges that protect major processes such as photo-synthesis and respiration, e.g., water use efficiency, andthose that preserve such important features as cytoske-leton, cell wall, or plasma membrane–cell wall interac-tions (Botella et al., 1994) and chromosome andchromatin structure changes, i.e., DNA methylation,polyploidization, amplification of specific sequences, orDNA elimination (Walbot and Cullis, 1985). It isbelieved that for the protection of higher-order pro-cesses, low-complexity mechanisms are induced coordi-nately (Bohnert et al., 1995).
2.2. Ion regulation and compartmentalization
Ion uptake and compartmentalization are crucial notonly for normal growth but also for growth under salineconditions (Adams et al., 1992b) because the stressdisturbs ion homeostasis. Plants, whether glycophyte or
anelle space• Plant growth regulatorsensitivity adjustment• Osmoprotection• Ion partitioning
orage
ance
he schematic presentation of a plant cell includes three compartments
ohnert and Jensen, 1996).

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349326
halophyte, cannot tolerate large amounts of salt in thecytoplasm and therefore under saline conditions theyeither restrict the excess salts in the vacuole orcompartmentalize the ions in different tissues to facil-itate their metabolic functions (Reddy et al., 1992;Iyengar and Reddy, 1996; Zhu, 2003). Glycophytes limitsodium uptake or partition sodium in older tissues thatserve as storage compartments that are eventuallysacrificed (Cheeseman, 1988).Removal of sodium from the cytoplasm or compart-
mentalization in the vacuoles is done by a salt-inducibleenzyme Na+/H+ antiporter (Apse et al., 1999). Twoelectrogenic H+ pumps, the vacuolar type H+-ATPase(V-ATPase) and the vacuolar pyrophosphatase (V-PPase), coexist at membranes of the secretory pathwayof plants (Dietz et al., 2001). The V-ATPase is thedominant H+ pump at endomembranes of most plantcells with regard to protein amount and, frequently,activity. The V-ATPase is indispensable for plantgrowth under normal conditions due to its roles inenergizing secondary transport, maintaining solutehomeostasis, and, possibly, facilitating vesicle fusion.Under stress conditions such as salinity, drought, cold,acid stress, anoxia, and excess heavy metals in the soil,survival of the cells depends strongly on maintaining oradjusting the activity of the V-ATPase. Regulation ofgene expression and activity are involved in adapting theV-ATPase on long- and short-term bases (Dietz, et al.,2001). Salt modulation of the tonoplast H+-pumping V-ATPase and H+-pyrophosphatase has been evaluated inhypocotyls of Vigna unguiculata seedlings (Otoch et al.,2001). Salt stress induces V-ATPase expression in V.
unguiculata with concomitant enhancement of itsactivity as a homeostatic mechanism to cope with saltstress, whereas under the same conditions V-PPase isinhibited (Otoch et al., 2001). The main strategy of salttolerance in the halophyte Suaeda salsa seems to be anup-regulation of V-ATPase activity, which is required toenergize the tonoplast for ion uptake into the vacuole,while V-PPase plays only a minor role (Wang et al.,2001).When under salt stress, plants maintain high concen-
trations of K+ and low concentrations of Na+ in thecytosol. They do this by regulating the expression andactivity of K+ and Na+ transporters and of H+ pumpsthat generate the driving force for transport (Zhu et al.,1993). Although salt-stress sensors remain elusive, someof the intermediary signaling components have beenidentified. Evidence suggests that a protein kinasecomplex consisting of the myristoylated calcium-bindingprotein SOS3 and the serine/threonine protein kinaseSOS2 is activated by a salt-stress-elicited calciumsignal. The protein kinase complex then phosphorylatesand activates various ion transporters, such as theplasma membrane Na+/H+ antiporter SOS1 (Zhuet al., 1993).
The Arabidopsis thaliana AtNHX1 gene encodes avacuolar Na+/H+ antiporter that is important in salttolerance. The tissue distribution and regulation ofAtNHX1 expression by salt stress and abscisic acid(ABA) have been reported by Shi and Zhu (2002).Experimental evidence shows that the steady state levelof AtNHX1 transcript is up-regulated by treatment withNaCl, KCl, or ABA. AtNHX1 promoter (GUS) analysisin transgenic Arabidopsis shows that AtNHX1 isexpressed in all tissues except the root tip. StrongGUS expression was detected in guard cells, suggestingthat AtNHX1 may play a role in pH regulation and/orK+ homeostasis in the specialized cells. AtNHX1
promoter activity is substantially up-regulated by NaCl,KCl, or ABA, demonstrating that salt and ABAregulation of AtNHX1 expression occurs at the tran-scriptional level. Strong induction of GUS activity inroot hair cells suggests a role of AtNHX1 in storing Na+
in the enlarged vacuoles in root hair cells. The up-regulation of AtNHX1 transcript levels by NaCl isreduced in abil-1, aba2-1, and aba3-1 but not in abi2-1,sos1, sos2, or sos3 mutants of Arabidopsis. ABA-induced AtNHX1 expression is also decreased in abil-1but not in abi2-1. This evidence suggests that salt stressup-regulates AtNHX1 expression transcriptionally andthat the up-regulation is partially dependent on ABAbiosynthesis and ABA signaling through ABI1 (Shi andZhu, 2002).With a homologous gene region a Na+/H+ anti-
porter gene has been isolated from a halophytic plant,Atriplex gmelini, and named AgNHX1 (Hamada et al.,2001). The isolated cDNA is 2607 bp in length andcontains one open reading frame, which comprises 555amino acid residues with a predicted molecular mass of61.9 kDa. The amino acid sequence of the AgNHX1
gene shows more than 75% identity with those of thepreviously isolated NHX1 genes from the glycophytes A.
thaliana and Oryza sativa. The migration pattern ofAgNHX1 is shown to correlate with H+-pyropho-sphatase and not with P-type H+-ATPase, suggestingthe localization of AgNHX1 in a vacuolar membrane.Induction of the AgNHX1 gene has been observed bysalt stress at both mRNA and protein levels. Theexpression of the AgNHX1 gene in the yeast mutant,which lacks the vacuolar-type Na+/H+ antiporter gene(NHX1) and has poor viability under the high-saltconditions, shows partial complementation of theNHX1 functions. It has been suggested that theAgNHX1 products have important roles in salt toler-ance (Hamada et al., 2001).Experimental evidence implicates Ca2+ function in
salt adaptation. Externally supplied Ca2+ reduces thetoxic effects of NaCl, presumably by facilitating higherK+/Na+ selectivity (Liu and Zhu, 1997; Lauchli andSchubert, 1989). High salinity also results in increasedcytosolic Ca2+ that is transported from the apoplast

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 327
and intracellular compartments (Knight et al., 1997).The resultant transient Ca2+ increase potentiates stresssignal transduction and leads to salt adaptation(Mendoza et al., 1994; Knight et al., 1997). A prolongedelevated Ca2+ level may, however, also pose a stress; ifso, reestablishment of Ca2+ homeostasis is a requisite.Other mechanisms of salt regulation are salt secretion
and selective salt accumulation or exclusion. Saltsecretion occurs through development of unique cellularstructures called salt glands. These salt glands secretesalt (especially NaCl) from leaves and maintain internalion concentration at lower level (Hogarth, 1999). Saltexclusion occurs through roots to regulate the saltcontent of their leaves in many halophytes (Levitt,1980). Selective accumulation of ions or solutes enablesthe plants to make osmotic adjustments, which occurthrough mass action, and results in increased waterretention and/or sodium exclusion.
2.3. Induced biosynthesis of compatible solutes
To accommodate the ionic balance in the vacuoles,cytoplasm accumulates low-molecular-mass compoundstermed compatible solutes because they do not interferewith normal biochemical reactions (Yancey et al., 1982;Ford, 1984; Ashihara et al., 1997; Hasegawa et al., 2000;Zhifang and Loescher, 2003); rather, they replace waterin biochemical reaction. With accumulation propor-tional to the change of external osmolarity withinspecies-specific limits, protection of structures andosmotic balance supporting continued water influx (orreduced efflux) are accepted functions of osmolytes(Hasegawa et al., 2000). These compatible solutesinclude mainly proline (Khatkar and Kuhad, 2000;Singh et al., 2000), glycine betaine (GB) (Rhodes andHanson, 1993; Khan et al., 2000a; Wang and Nil, 2000),sugars (Kerepesi and Galiba, 2000; Bohnert and Jensen,1996; Pilon-Smits et al., 1995), and polyols (Ford, 1984;Popp et al., 1985; Orthen et al., 1994; Bohnert et al.,1995).Polyols make up a considerable percentage of all
assimilated CO2 and serve several functions: as compa-tible solutes, as low-molecular-weight chaperones, andas scavengers of stress-induced oxygen radicals (Smirn-off and Cumbes, 1989; Bohnert et al., 1995). Polyols areclassified as acyclic (e.g., manitol) and cyclic (e.g.,pinitol). Mannitol, a sugar alcohol that may serve as acompatible solute to cope with salt stress, is synthesizedvia the action of a mannose-6-phosphate reductase(M6PR) in celery (Zhifang and Loescher, 2003). Incontrast to previous approaches that have used abacterial gene to engineer mannitol biosynthesis inplants and other organisms, A. thaliana, a non-mannitol-producer, has been transformed with thecelery leaf M6PR gene under control of the CaMV
35S promotor. In all independent Arabidopsis M6PR
transformants, mannitol accumulates throughout theplants in amounts ranging from 0.5 to 6 mmol g�1 freshweight. A novel compound, not found in either celery orArabidopsis, 1-O-beta-D-glucopyranosyl-D-mannitol,also accumulates in vegetative tissues of mature plantsin amounts up to 4 mmol g�1 fresh weight but not inflowers and seeds. In the absence of NaCl, alltransformants are phenotypically the same as the wildtype; however, in the presence of NaCl, maturetransgenic plants shows a high level of salt tolerance,i.e., growing, completing normal development, flower-ing, and producing seeds in soil irrigated with 300mMNaCl in the nutrient solution. These results demonstratea major role in developing salt-tolerant plants by meansof introducing mannitol biosynthesis using M6PR
(Zhifang and Loescher, 2003). Pinitol is synthesizedfrom myo-inositol by the sequential catalysis of inositol-o-methyltransferase and ononitol epimerase (Bohnertand Jensen, 1996). Polyols function in two ways that aredifficult to separate mechanistically: osmotic adjustmentand osmoprotection. In osmotic adjustment, they act asosmolytes facilitating the retention of water in thecytoplasm and allowing sodium sequestration to thevacuole or apoplast. These osmolytes protect cellularstructures by interacting with membranes, proteincomplexes, or enzymes. These compounds have hydro-gen-bonding characteristics that allow them to protectmacromolecules from the adverse effects of increasingionic strength in the surrounding media (Crowe et al.,1992). By tight association with protein and membranecomponents, they compensate for water loss duringstress (Yancey et al., 1982). Those polyols that arenonreducing sugars may also store excess carbon underenvironmental stress conditions (Vernon et al., 1993).The cyanobacterium Synechocystis sp. PCC 6803accumulates the compatible solute glucosylglycerol(GG) and sucrose under salt stress (Hagemann andMurata, 2003). The molecular mechanisms for GGsynthesis including regulation of the GG-phosphatesynthase (ggpS) gene, which encodes GgpS, has beenintensively investigated. Experimental evidences showsthat GG is important for salt tolerance and thus for theproper division of cells under salt stress conditions inSynechocystis (Hagemann and Murata, 2003). Accumu-lation of the polyol myo-inositol in leaf tissues ofActidinia takes place under salt stress and accumulationof myo-inositol is negatively correlated to fructose andglucose (Klages et al., 1999). Severe salt stress(4120mM NaCl) led to a preferential accumulationof D-pinitol in gametophytes of Acrostichum aureum,whereas the sporophyte accumulates D-1-O-methyl-muco-inositol (Sun et al., 1999).Carbohydrates such as sugars (glucose, fructose,
sucrose, fructans) and starch accumulate under saltstress (Parida et al., 2002). Their major functions areosmoprotection, osmotic adjustment, carbon storage,

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349328
and radical scavenging. Salt stress increases reducingsugars (glucose, fructose), sucrose, and fructans in anumber of plants (Kerepesi and Galiba, 2000; Khatkarand Kuhad, 2000; Singh et al., 2000). In Vicia faba
salinity decreases soluble and hydrolyzable sugars(Gadallah, 1999). Sugar content increases in somegenotypes of rice but also decreases in some genotypes(Alamgir and Ali, 1999). Under salinity, the starchcontent in roots of rice plants declines and remainsunchanged in shoots. A decrease in starch content andan increase in both reducing and nonreducing sugarhave been reported in leaves of Bruguiera parviflora
(Parida et al., 2002) (Fig. 2). The contents of reducingand nonreducing sugars and the activity of sucrosephosphate synthase increase under salt stress, whereasstarch phosphorylase activity decreases (Dubey andSingh, 1999). In leaves of tomato the contents of solublesugars and total saccharides are significantly increased,but the starch content is not affected significantly byNaCl treatment (Khavarinejad and Mostofi, 1998). Gaoet al. (1998) have reported that in tomato (Lycopersicon
esculentum L.) salinity enhances sucrose concentrationand activity of sucrose phosphate synthase (EC 2.3.1.14)in leaves but decreases the activity of acid invertase (EC2.3.1.14). It has also been reported that polyphenol level
C 1 2 3 4 M
66.0
43.0
20.1
29.0
14.3
Fig. 2. Changes in protein profle in leaf of B. parviflora after 7, 14, and
30 days recovery from salt stress. Lanes C and 1 represent proteins
extracted from control and 400mM NaCl-treated (45 day) leaves.
Lanes 2, 3, and 4 represent proteins extracted from leaves after 7, 14
and 30 days of desalinization, respectively. Lane M represents the
molecular weight marker. The boldface arrow on the left indicates the
23-kDa polypeptide, which reappears after desalinization (from Parida
et al., 2004c).
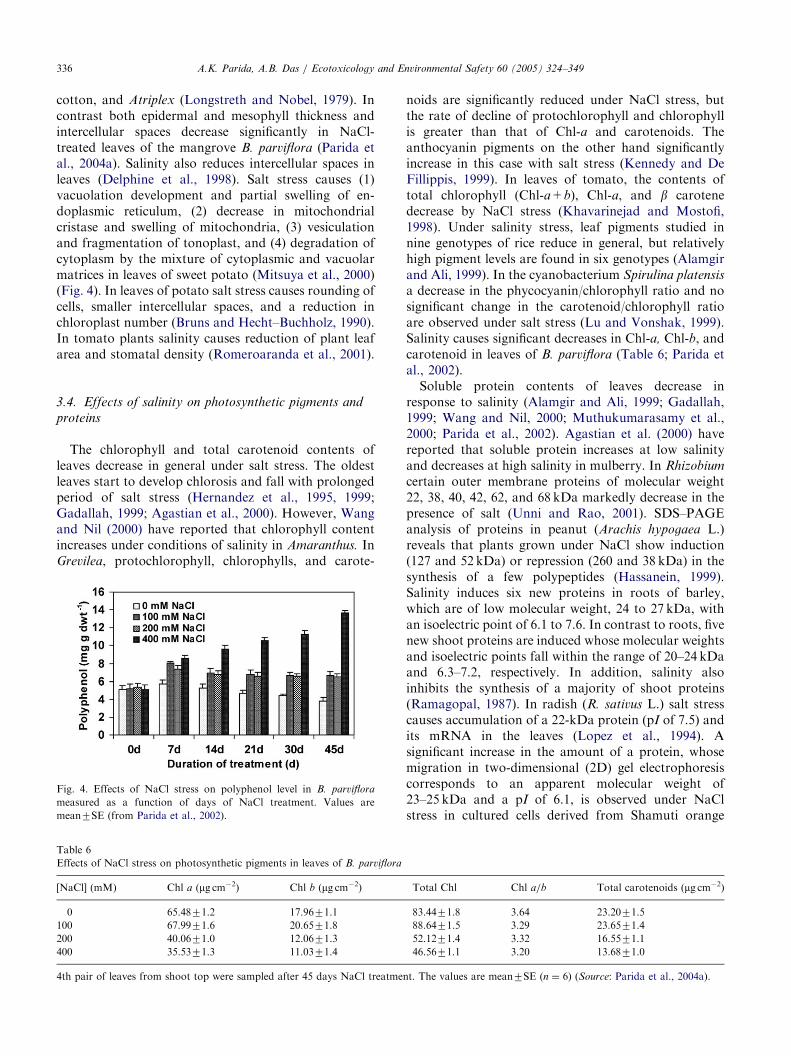
increases in leaves of B. parviflora by salt stress (Fig. 4)(Parida et al., 2002).A number of nitrogen-containing compounds (NCC)
accumulate in plants exposed to saline stress. The mostfrequently accumulating NCC include amino acids,amides, imino acids, proteins, quaternary ammoniumcompounds, and polyamines. The specific NCC thataccumulate in saline environments varies with plantspecies. Osmotic adjustment, protection of cellularmacromolecules, storage of nitrogen, maintenance ofcellular pH, detoxification of the cells, and scavenging offree radicals are proposed functions of these compoundsunder stress conditions. NCC accumulation is usuallycorrelated with plant salt tolerance (Mansour, 2000).There are several reports of accumulation of free aminoacids and other NCC under salt stress. Glycine betainecontent increases by salt stress in a number of plants(Khan et al., 1998, 1999, 2000a; Saneoka et al., 1999;Muthukumarasamy et al., 2000; Wang and Nil, 2000).Khan et al. (2000a) have reported that glycine betainecontent increases in shoots and does not differsignificantly in roots of Haloxylon recurvum under saltstress. Many plants accumulate proline as a nontoxicand protective osmolyte under saline conditions (Leeand Liu, 1999; Khatkar and Kuhad, 2000; Muthuku-marasamy et al., 2000; Singh et al., 2000; Jain et al.,2001). It has been reported that proline levels increasesignificantly in leaves of nonsecretor mangrove B.
parviflora (Fig. 3) (Parida et al., 2002) and in salt-secretor mangrove Aegiceras corniculatum (Parida et al.,unpublished data). The most abundant amino acids(cysteine, arginine, methionine), constituting about 55%of total free amino acid content, are reduced in NaCl-treated plants of wheat, but valine, isoleucine, asparticacid, and proline increase in response to NaCl stress andNaCl-treated wheat seedlings show 1.6-fold increase intotal free amino acids compared to the control(Elshintinawy and Elshourbagy, 2001). In Zea mays,
Fig. 3. Effects of NaCl stress on proline level in B. parviflorameasured
as a function of days of NaCl treatment. Values are mean7SE (fromParida et al., 2002).

ARTIC
LEIN
PRES
S
Table 1
Responses to salt-stress-accumulating products and their function(s) in conferring tolerance
Product group Specific compound Suggested function(s) References
Ions Sodium, chloride Osmotic adjustment Blumwald et al. (2000); Hasegawa et al. (2000); Nilu et al. (1995)
Potassium exclusion/Export Koyro (2000)
Khan et al. (2000a, b)
Proteins Osmotin Pathogenesis-related proteins Singh et al. (1987); King et al. (1988)
SOD/Catalase Osmoprotection Bohnert and Jensen (1996)
Radical detoxification Bohnert and Jensen (1996); Allen et al (1997); Hernandez et al. (2000)
Amino acids Proline Osmotic adjustment Khatkar and Kuhad (2000); Singh et al. (2000); Lin et al. (2002)
Ectoine Osmoprotection Lippert and Galinski (1992)
Sugars Glucose, fructose, sucrose Osmotic adjustment Kerepesi and Galiba (2000)
Fructans Osmoprotection, carbon storage Bohnert and Jensen (1996); Pilon-Smits et al. (1995)
Polyols Acyclic (e.g., manitol) Carbon storage, osmotic adjustment Popp et al. (1985); Bohnert et al. (1995)
Cyclic (e.g., pinitol) Osmoprotection, osmotic adjustment Ford (1984); Bohnert et al. (1995)
Retention of photochemical efficiency of PSII Sun et al (1999)
Radical scavenging Smirnoff and Cumbes (1989); Orthen et al. (1994)
Polyamines Spermine, spermidine Ion balance, chromatin protection Tiburico et al. (1993); SantaCruz et al. (1998)
Quaternary amines Glycine betaine Osmoprotection Khan et al. (2000a); Wang and Nil (2000)
Preservation of thylakoid and plasma membrane integrity Rhodes and Hanson (1993)
b-Alanine betaine, Osmoprotection Rhodes and Hanson (1993)
Dimethyl-sulfonio propionate, Osmoprotection Hanson (1998); Trossat et al. (1998)
Choline-o-sulfate Osmoprotection Nuccio et al. (1999)
Trigonelline Rajasekaran et al. (2001)
Pigments Carotenoids, anthocyanins, betalaines Protection against photoinhibition Foyer et al. (1994); Adams et al. (1992a); Kennedy and De Fillippis (1999)
A.K
.P
arid
a,
A.B
.D
as
/E
coto
xico
log
ya
nd
En
viron
men
tal
Sa
fety6
0(
20
05
)3
24
–3
49
329

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349330
with increasing external salt concentration, amino acidsand quaternary ammonium compounds accumulate inleaves and roots, with concentrations in leaves exceedingthose in roots (AbdElBaki et al., 2000). Prolineaccumulates in leaves, stems, and roots of Pringlea
antiscorbutica (kerguelen cabbage) and this osmolyteaccumulate at a 2–3� higher concentration in thecytoplasm than in the vacuole (Aubert et al., 1999).Mattioni et al. (1997) have reported that most of theamino acids show an increase with the induction ofwater or salt stress in durum wheat (Triticum durum),but proline increases more markedly than other aminoacids and the activity of the enzyme delta (1)-pyrroline-5-carboxylate reductase (EC 1.5.1.2) involved in prolinebiosynthesis is enhanced during both water and saltstress, while the activity of proline dehydrogenase (EC1.5.1.2) involved in proline catabolism is inhibited onlyduring salt stress. These results indicate that synthesis denovo is the predominant mechanism in proline accu-mulation in durum wheat. In mulberry, free amino acidsincrease at low salinity and decrease at high salinity andglycine betaine accumulates more than proline (Agas-tian et al., 2000). Proline, quaternary ammonium, andtertiary sulfonium osmolytes are zwitter ions at physio-logical pH. Although they are ionic, they have no netcharge. Their osmotic function is due to their uniquechemistry.Experimental observations shows that transformation
of plants with the codA gene for choline oxidaseenhances tolerance of the transgenic plants to salt stressat the reproductive stage as a result of the accumulationof GB in the reproductive organs (Sulpice et al., 2003).The compatible solute N-epsilon-acetyl-beta-lysine isunique to methanogenic archaea and is produced undersalt stress only (Pfluger et al., 2003). However, themolecular basis for the salt-dependent regulation of N-epsilon-acetyl-beta-lysine formation is unknown. Genespotentially encoding lysine-2, 3-aminomutase (abl4) andbeta-lysine acetyltransferase (ablB), which are assumedto catalyze N-epsilon-acetyl-beta-lysine formation fromalpha-lysine, have been identified on the chromosomesof the methanogenic archaeaMethanosarcina mazei Gol,Methanosarcina acetivorans, Methanosarcina barkeri,Methanococcus jannaschii, and Methanococcus maripa-
ludis (Pfluger et al., 2003). Table 1 provides examples ofplant products whose action correlates with metabolicfunctions that are known or assumed to enable plants tocope with salt tolerance.A common feature of compatible solutes is that these
compounds can accumulate to high levels withoutdisturbing intracellular biochemistry (Bohnert andJensen, 1996). Compatible solutes have the capacity topreserve the activity of enzymes that are in salinesolutions. These compounds have minimal effect on pHor charge balance of the cytosol or lumenal compart-ments of organelles. The synthesis of compatible
osmolytes is often achieved by diversion of basicintermediary metabolites into unique biochemical reac-tions. Often, stress triggers this metabolic diversion. Forexample, higher plants synthesize glycine betaine fromcholine by two reactions that are catalyzed in sequenceby choline monooxygenase and betaine aldehyde dehy-drogenase (Rhodes and Hanson, 1993).
2.4. Induction of antioxidative enzymes
Salt stress is complex and imposes a water deficitbecause of osmotic effects on a wide variety of metabolicactivities (Greenway and Munns, 1980; Cheeseman,1988). This water deficit leads to the formation ofreactive oxygen species (ROS) such as superoxide (O2
d�),hydrogen peroxide (H2O2), hydroxyl radical (
dOH)(Halliwell and Gutteridge, 1985), and singlet oxygen(1O2) (Elstner, 1987). These cytotoxic activated oxygenspecies can seriously disrupt normal metabolism throughoxidative damage to lipids (Fridovich, 1986; Wise andNaylor, 1987) and to protein and nucleic acids (Frido-vich, 1986; Imlay and Linn, 1988). Since internal O2concentrations are high during photosynthesis (Steiger etal., 1977), chloroplasts are especially prone to generateactivated oxygen species (Asada and Takahashi, 1987).Once produced, O2
d� will rapidly dismutate, eitherenzymatically or nonenzymatically, to yield H2O2 andO2. In addition, H2O2 may interact in the presence ofcertain metal ions or metal chelates to produce highlyreactive dOH (Imlay and Linn, 1988). In varyingdegrees, present day plants possess a number ofantioxidants that protect against the potentially cyto-toxic species of activated oxygen. The metalloenzymesuperoxide dismutase (SOD; EC 1.15.1.1) converts O2
d�
to H2O2. Catalase and a variety of peroxidases (Chang etal., 1984) catalyze the breakdown of H2O2. Althoughcatalase is apparently absent in the chloroplast, H2O2can be detoxified in a reaction catalyzed by an ascorbate-specific peroxidase often present in high levels in thisorganelle (Chen and Asada, 1989) through the ascorba-te–glutathione cycle (Halliwell and Gutteridge, 1986;Asada, 1992). Both ascorbate and glutathione have beenreported in millimolar concentrations within the chlor-oplast (Halliwell, 1982). Ascorbate can also be oxidizedby direct reaction with O2
d� or by serving as a reductantof the a-chromoxyl radical of oxidized a-tocopherol(Foyer et al., 1991). The thylakoid membranes are rich ina-tocopherol which disrupts lipid peroxidation reactionsnot only by reacting with O2
d� but also by scavenginghydroxyl, peroxyl, and alkoxyl radicals (Halliwell, 1987).When plants are subjected to environmental stress
conditions such as high light intensity, temperatureextremes, drought, high salinity, herbicide treatment, ormineral deficiencies, the balance between the productionof reactive oxygen species and the quenching activity ofthe antioxidants is upset, often resulting in oxidative

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 331
damage (Harper and Harvey, 1978; Dhindsa andMatowe, 1981; Wise and Naylor, 1987; Spychalla andDesborough, 1990). Plants with high levels of antiox-idants, either constitutive or induced, have beenreported to have greater resistance to this oxidativedamage (Harper and Harvey, 1978; Dhindsa andMatowe, 1981; Wise and Naylor, 1987; Spychalla andDesborough, 1990). The activities of the antioxidativeenzymes such as catalase (CAT), ascorbate peroxidase(APX), guaicol peroxidase (POD), glutathione reductase(GR), and superoxide dismutase increase under saltstress in plants and a correlation of these enzyme levelsand salt tolerance exists (Gossett et al., 1994; Hernandezet al., 1995, 2000; Sehmer et al., 1995; Kennedy and DeFillippis, 1999; Sreenivasulu et al., 2000; Benavides etal., 2000; Lee et al., 2001; Mittova et al., 2002, 2003). Insoybean root nodules ascorbate peroxidase, catalase,and glutathione reductase activities decrease under saltstress, while superoxide dismutase and reduced glu-tathione increase and malondialdehyde and total proteinremain unchanged (Comba et al., 1998).Transgenic plants have been generated to probe the
effects of ROS scavenging on salinity stress tolerance,based on observations of gene expression changes insalt-stressed plants. A putative phospholipid hydroper-oxide glutathione peroxidase (PHGPX) transcript in-creases during salt stress in Arabidopsis and citrus(Gueta-Dahan et al., 1997; Sugimoto and Sakamoto,1997); also in citrus, transcripts and enzyme activities ofCu/Zn-SOD, glutathione peroxidase, and cytosolic APXincrease (Holland et al., 1993; Gueta-Dahan et al.,1997). Catalase-deficient (antisense) tobacco showsenhanced sensitivity to oxidative stress under conditionsof light and salinity (Willkens et al., 1997). ROSscavenging as an important component of abiotic stressresponses is documented by mutant analysis. Theascorbic-acid-deficient Arabidopsis semidominant soz1
accumulates only 30% of ascorbate compared with wildtype, and plants show significantly higher sensitivity tooxidative stress conditions (Conklin et al., 1996).Further support comes from the study of transgenicmodels, which have been generated to study antioxidantdefenses (Orr and Sohal, 1992; Creissen et al., 1996;Allen et al., 1997; Noctor and Foyer, 1998; Smirnoff,1998). Overexpression of genes leading to increasedamounts and activities of mitochondrial Mn-SOD, Fe-SOD, chloroplastic Cu/Zn-SOD, bacterial catalase, andglutathione-S-transferase (GST)/glutathione peroxidase(GPX) can increase the performance of plants understress (Bowler et al., 1991; Gupta et al., 1993a, b; VanCamp et al., 1996; Shikanai et al., 1998; Roxas et al.,2000). A cDNA clone encoding a cytosolic SOD hasbeen isolated from a cDNA library constructed frompoly(A)+ RNA from epicotyls of 5-day-old Cicer
arietinum, L. etiolated seedlings after a differentialscreening to select clones whose expression decreases
with epicotyl growth (Hernandez et al., 2002). Analysisof its deduced amino acid sequence shows all the typicalstructural motifs of plant cytosolic SODs (EC 1.15.1.1.).The expression of this clone is always higher in youngand growing tissues than in old and storage tissues anddiminishes throughout the development of the seedlings.Cytosolic Cu/Zn-SOD activity is also higher in radiclesand younger internodes. Under stress conditions onlycold increases the gene expression whereas the activity isclearly increased by a saline medium (Hernandez et al.,2002). To analyze the potential of the active oxygen-scavenging system of the cytosol in leaves of salt-stressed mangrove B. gymnorrhiza, Takemura et al.(2002) have isolated a full-length cDNA encoding a 153-amino-acid sequence of cytosolic Cu/Zn-SOD and apartial cDNA encoding catalase. Northern blot analysesshow that the transcript level of cytosolic Cu/Zn-SODincreases after 1 and 5 days of NaCl treatment, but nosignificant change occurs in the expression of thecatalase gene. The transcript of cytosolic Cu/Zn-SODis also induced by mannitol treatment. This suggeststhat the increase in cytosolic Cu/Zn-SOD 1 day afterNaCl treatment is a response to osmotic stress.After 5 days of treatment with NaCl, the transcriptlevel of cytosolic Cu/Zn-SOD increases in young andmature leaves but not in old leaves. Expression of thecytosolic Cu/Zn-SOD gene is induced by exogenousabscisic acid, while the catalase gene is induced byapplication of 2-chloroethylphosphonic acid, which is agenerator of ethylene. The results from this studysuggest that salt stress leads to the generation ofsuperoxide in the cytosol and that the oxygen-scaven-ging system in the cytosol contributes to the salttolerance capacity of B. gymnorrhiza (Takemura et al.,2002).
2.5. Induction of plant hormones
High salt concentration triggers an increase in levelsof plant hormones such as ABA and cytokinins(Thomas et al., 1992; Aldesuquy, 1998; Vaidyanathanet al., 1999). Abscisic acid is responsible for thealteration of salt-stress-induced genes (de Bruxelles etal., 1996). ABA-inducible genes are predicted to play animportant role in the mechanism of salt tolerance in rice(Gupta et al., 1998). Salt stress results in increased levelsof ABA, aminocyclopropane-1-carboxylic acid, andethylene production in Citrus sinensis (GomezCadenaset al., 1998). ABA has been found to alleviate theinhibitory effect of NaCl on photosynthesis, growth andtranslocation of assimilates (Popova et al., 1995). ABAalso promotes switch from C3 to crassulacean acidmetabolism (CAM) in Mesembryanthemum crystallinum
under salt stress (Thomas et al., 1992). ABA promotesstomatal closure by rapidly altering ion fluxes in guardcells under stress conditions. Other ABA actions involve

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349332
modifications of gene expression, and the analysis ofABA-responsive promoters has revealed diversity ofpotential cis-acting regulatory elements. The nature ofthe ABA receptors(s) remains unknown. In contrast,combined biophysical, genetic, and molecular ap-proaches have led to considerable progress in thecharacterization of more downstream signaling ele-ments. In particular, substantial evidence points to theimportance of reversible protein phosphorylation andmodification of cytosolic calcium levels and pH asintermediates in ABA signal transduction (Leung andGiraudat, 1998). Experimental evidence shows that theincrease of Ca2+ uptake is associated with the rise ofABA under salt stress and thus contributes to mem-brane integrity maintainance, which enables plants toregulate uptake and transport under high levels ofexternal salinity in the longer term (Chen et al., 2001). Ithas been reported that ABA reduces ethylene releaseand leaf abscission under salt stress in citrus probably bydecreasing the accumulation of toxic Cl� ions in leaves(GomezCadenas et al., 2002). Salt tolerance of faculta-tively halophytic Lophopyrum elongatum and the closelyrelated but less salt tolerant wheat T. aestivum L. isenhanced when plants are allowed to gradually accli-mate to salt stress in comparison to when they aresuddenly shocked (Noaman et al., 2002). This acclima-tion to salt stress is regulated by ABA; a pretreatmentwith ABA substitutes for the acclimation and increasestolerance of salt shock. The ABA-induced acclimation israpid and coincides with enhanced expression of theearly salt induced (ESI) genes in the roots. L. elongatum
tolerates salt shock better than wheat and its genomeconfers greater tolerance of salt shock in theiramphiploid than wheat. The tolerance of salt shock isprincipally controlled by chromosome 3E in theL. elongatum genome. Wheat homologous chromo-somes 3A and 3D also control salt shock response. Itis speculated that chromosome 3 in both speciesmediates salt shock response via ABA (Noaman et al.,2002).Jasmonates also have important roles in salt toler-
ance. Experimental evidence shows that salt-toleranttomato cultivars have higher levels of jasmonates thansalt-sensitive cultivars (Hilda et al., 2003). Jasmonatesare generally considered to mediate signaling, such asdefense responses, flowering, and senescence. However,factors involved in the jasmonate signal-transductionpathway remain unclear. To clarify the functions andsignaling mechanisms of jasmonates on a genomewidelevel, Sasaki et al. (2000) have adopted a cDNAmacroarray technique. By analyzing the data from thecDNA macroarray, many function-known and -un-known genes have been identified as MeJA-responsivegenes, and the profiles of expression show goodagreement with Northern blot analysis (Sasaki et al.,2000).
2.6. Change in photosynthetic pathway
Salt stress inhibits photosynthesis by reducing waterpotential. So the main aim of salt tolerance is to increasewater use efficiency under salinity. To this end,facultative halophytic plants such as M. crystallinum
shift their C3 mode of photosynthesis to CAM (Cush-man et al., 1989). This change allows the plant to reducewater loss by opening stomata at night, thus decreasingtranspiratory water loss under prolonged salinity con-ditions. There is also a shift from the C3 to the C4pathway in response to salinity in salt-tolerant plantspecies such as Atriplex lentiformis (Zhu and Meinzer,1999).
2.7. Molecular mechanism of salt tolerance
Physiologic or metabolic adaptations to salt stress atthe cellular level are the main responses amenable tomolecular analysis and have led to the identification of alarge number of genes induced by salt (Ingram andBartels, 1996; Bray, 1997; Shinozaki et al., 1998).Selective examples of genes/proteins induced by saltstress are given in Table 2. These genes can be classifiedin functional groups (Table 3) related to their physio-logic or metabolic function predicted from sequencehomology with known proteins. Salt tolerance is amultigenic trait and a number of genes categorized intodifferent functional groups are responsible for encodingsalt-stress proteins: (i) genes for photosynthetic en-zymes, (ii) genes for synthesis of compatible solutes, (iii)genes for vacuolar-sequestering enzymes, and (iv) genesfor radical-scavenging enzymes. Most of the genes in thefunctional groups have been identified as salt inducibleunder stress conditions. Other genes have been detectedby a salt-hypersensitivity assay in Arabidopsis, which ledto the identification of mutants in potassium uptake asbeing critical in salt sensitivity (Wu et al., 1996).However, other physiological systems may be equallylimiting under stress conditions and mutants in thesephysiological pathways could lead to increased salttoxicity and would affect survival in a negative manner.Transcript regulation in response to high salinity has
been investigated for salt-tolerant rice (var Pokkali) withmicroarrays including 1728 cDNAs from libraries ofsalt-stressed roots (Kawasaki et al., 2001). NaCl at150mM reduces photosynthesis to one-tenth of the pre-stress value within minutes. Hybridizations of RNA tomicroarray slides were probed for changes in transcriptsfrom 15min to 1 week after salt shock. Beginning 15minafter the shock, Pokkali shows up-regulation oftranscripts. Approximately 10% of the transcripts inPokkali are significantly up- or down-regulated within1 h of salt stress. The initial differences between controland stressed plants continue for hours but become lesspronounced as the plants adapted over time. The

ARTIC
LEIN
PRES
S
Table 2
Selective examples of genes/proteins induced by salt stress
Plant species Salt responsive genes/proteins Characteristic feature(s) Reference
Arabidopsis thaliana Sal 1 Induced by salt stress, overexpression in Arabidopsis or yeast overcomes
Na+ and Li+ toxicity, homologous to hal 1 of yeast
Quintero et al. (1996)
Brassica napus Bnd 22 22 kDa protein, level increased by progessive or rapid water stress and
salinity
Reviron et al. (1992)
Citrus cinensis Salt associated 23 to 25 kDa protein Induced by salt stress, ABA, and water stress Benhayyim et al. (1993)
Dunaliella salina P 150 150kDa protein, induced by salt stress, de novo synthesized protein Sadaka et al. (1991)
Hordeum ulgare 26 and 27 kDa proteins (salt-induced
polypeptides SIP S1–S4)
Salt stress induced S1–S4 polypeptides but water deficit did not induce S2polypeptides
Hurkman and Tanaka (1988)
hva 1 Induced by ABA, drought, NaCl, cold, and heat treatment Hong et al. (1992)
Lycopersicom esculantum TAS-12 Salt and water stress induced lipid transfer protein Torres-Schumann et al. (1992)
le-16 gene Induced by drought, PEG, salinity, cold, and heat stress Plant et al. (1991)
Mesembryanthemum
crystallinum
ppc-1 and ppc-2 Encodes PEP carboxylase, induced by salt and water stress, exogenous
ABA is a poor substitute for NaCl to induce it
Cushman et al. (1989)
Isogenes
lmt 1 Encodes myo-inositol o-methyl transferase 1; induced by NaCl and
osmotic stress
Vernon and Bohnert (1992)
Inps 1 Encodes myo-inositol 1-phosphate synthase; shows significant
homology to corresponding genes in plants and yeast
Ishitani et al. (1996)
Nicotiana tabacum 26- and 43-kDa polypeptides Levels increase in both NaCl, and PEG-induced water stress adapted
cells but are not detected in unadapted
Singh et al. (1985)
58-, 37-, 35.5-, 34-, 26-, 21-, 19.5-, and
18-kDa polypeptides
Increased levels with increased NaCl tolerance Singh et al. (1985)
30-kDa polypeptide Heat shock at 38 1C induces cross tolerance to salt stress Harrington and Alm (1988)
Vitronectin and fibronectin-like
proteins
Found in membranes and cell wall of NaCl-adapted cells Zhu et al. (1993)
Osmotin 26-kDa protein, protein level enhanced in both NaCl- and PEG-induced
water stress adapted cells but not in unadapted cells
Singh et al. (1987)
O. sativa RAB21 Induced when plants are subjected to water stress, rab21 mRNA and
protein accumulate in rice embryos, leaves, roots, and callus derived
suspension cells upon treatment with NaCl and/or ABA
Mundy et al. (1990)
salT mRNA accumulates rapidly in shoots and roots of mature seedlings
with ABA salts, PEG, NaCl, and KCl; no induction in leaf lamina
Claes et al. (1990)
em Induced by ABA and salt stress, salt interacts synergistically with ABA Bostock and Quatrano (1992)
The names of various genes and proteins have been by and large reproduced here as per the original publications of the authors.
A.K
.P
arid
a,
A.B
.D
as
/E
coto
xico
log
ya
nd
En
viron
men
tal
Sa
fety6
0(
20
05
)3
24
–3
49
333

ARTICLE IN PRESS
Table 3
Functional groups of genes/proteins activated in salt stress with
potential for providing tolerance (reproduced from Winicov, 1998)
1 Carbon metabolism and energy production/photosynthesis
2 Cell wall/membrane structural components
3 Water channel proteins
4 Ion transport
5 Oxidative stress defenses
6 Detoxifying enzymes
7 Proteinases
8 Proteins involved in signaling
9 Transcription factors
Table 4
Differences between halophytes and glycophytes with respect to salt
tolerance mechanism
Halophytes Glycophytes
1. Salinity has a smaller effect
upon the extent to which
stomata limit photosynthesis.
1. Higher.
2. High water use efficiency. 2. Comparatively less.
3. Low internal CO2concentration.
3. Comparatively less.
4. Low light saturated
photosynthetic rate.
4. It is less.
5. Efficient accumulating solutes. 5. Less efficient.
6. Low level of Na+ and Cl� ions
in the cytoplasm and
chloroplast.
6. It is higher.
A.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349334
interpretation of an adaptive process is supported by thesimilar analysis of salinity-sensitive rice (var IR29), inwhich the immediate response exhibited by Pokkali isdelayed and later results in down-regulation of tran-scription and death. The up-regulated functions ob-served with Pokkali at different time points during stressadaptation change over time. Increased protein synth-esis and protein turnover are observed at early timepoints, followed by the induction of known stress-responsive transcripts within hours and the induction oftranscripts for defense-related functions later. After 1week, the nature of up-regulated transcripts (e.g.,aquaporins) indicates recovery (Kawasaki et al., 2001).Acclimation of microorganisms to environmental
stress is closely related to the expression of variousgenes (Kanesaki et al., 2002). It has been reported thatsalt stress and hyperosmotic stress have different effectson the cytoplasmic volume and gene expression inSynechocystis sp. PCC 6803 (Kanesaki et al., 2002).DNA microarray analysis indicates that salt stressstrongly induces the genes for some ribosomal proteins.Hyperosmotic stress strongly induces the genes for 3-ketoacyl-acyl carrier protein reductase and rare lipo-protein A. Genes whose expression is induced both bysalt stress and by hyperosmotic stress include those forheat shock proteins and the enzymes for the synthesis ofglucosylglycerol. It has also been reported by Kanesakiet al. (2002) that each kind of stress induces a number ofgenes for proteins of unknown function. Findings ofKanesaki et al. (2002) suggests that Synechocystis sp.recognizes salt stress and hyperosmotic stress asdifferent stimuli, although mechanisms common to theresponses to each form of stress might also contribute togene expression.
2.8. Salinity effect: halophytes vs. glycophytes
The salt tolerance mechanism is a multigenic trait andtherefore biochemical pathways leading to products orprocesses that improve salt tolerance are likely to actadditively and probably synergistically. So the advan-tage of halophytes over glycophytes can be throughmore efficient performance of a new basic biochemical
tolerance mechanism. Table 4 summarizes differences inhalophytes and glycophytes with respect to salt toler-ance mechanism.
3. Salinity effects on plants
Salinity of soil and water is caused by the presence ofexcessive amounts of salts. Most commonly, high Na+
and Cl� cause the salt stress. Salt stress has threefoldeffects; viz. it reduces water potential and causes ionimbalance or disturbances in ion homeostasis andtoxicity. This altered water status leads to initial growthreduction and limitation of plant productivity. Since saltstress involves both osmotic and ionic stress (Hagemannand Erdmann, 1997; Hayashi and Murata, 1998),growth suppression is directly related to total concen-tration of soluble salts or osmotic potential of soil water(Flowers et al., 1977; Greenway and Munns, 1980). Thedetrimental effect is observed at the whole-plant level asdeath of plants or decrease in productivity. Suppressionof growth occurs in all plants, but their tolerance levelsand rates of growth reduction at lethal concentrations ofsalt vary widely among different plant species. Althoughthe change in water status is the cause of growthsuppression, the contribution of subsequent processes toinhibition of cell division and expansion and accelera-tion of cell death has not been well documented(Hasegawa et al., 2000). Salt stress affects all the majorprocesses such as growth, photosynthesis, proteinsynthesis, and energy and lipid metabolism. These arediscussed under separate headings.
3.1. Effects of salinity on growth
Salinity stress results in a clear stunting of plants(Hernandez et al., 1995; Cherian et al., 1999; Takemuraet al., 2000). The immediate response of salt stress isreduction in the rate of leaf surface expansion leading to

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 335
cessation of expansion as salt concentration increases(Wang and Nil, 2000). Salt stress also results in aconsiderable decrease in the fresh and dry weights ofleaves, stems, and roots (Hernandez et al., 1995;AliDinar et al., 1999; Chartzoulakis and Klapaki,2000). The optimum growth of plants is obtained at50% seawater and declines with further increases insalinity in Rhizophora mucronata (Aziz and Khan, 2001).Fresh and dry weights of plants increase with anincrease in salinity in Salicornia rubra while the optimalgrowth occurrs at 200mM NaCl and the growthdeclines with a further increase in salinity (Khan,2001). In Raphanus sativus (radish) total plant dryweight decreases at higher salinities and about 80% ofthe growth reduction at high salinity can be attributed toreduction of leaf area expansion and hence to reductionof light interception. The remaining 20% of the salinityeffect on growth is most likely explained by a decrease instomatal conductance. The small leaf area at highsalinity is related to a reduced specific leaf area andincreased tuber/shoot weight ratio and the latter can beattributed to tuber formation starting at a smaller plantsize at high salinity (Marcelis and VanHooijdonk, 1999).Kurban et al. (1999) have reported that in Alhagi
pseudoalhagi (a leguminous plant), total plant weightincreases at low salinity (50mM NaCl) but decreases athigh salinity (100 and 200mM NaCl). Khan et al. (1999)have reported that when Halopyrum mucronatum (aperennial grass found on the coastal dunes of Karachi,Pakistan) is treated with 0, 90, 180, and 360mMNaCl insand culture, fresh and dry mass of roots and shootspeaks at 90mM NaCl, a further increase in salinityinhibits plant growth, ultimately resulting in plant deathat 360mM NaCl, and maximum succulence is noted at90mM NaCl. Experimental evidence shows that in a saltnonsecretor mangrove B. parviflora the plant growth isoptimal at 100mM NaCl under hydroponic culture,whereas further increase in NaCl concentration retardsplant growth (Table 5) and 500 mM NaCl is found to belethal in this species (Parida et al., 2004a). On the otherhand a salt secretor mangrove Aegiceras corniculatum
can tolerate upto 250mM NaCl and 300mM is foundlethal in this case (Mishra and Das, 2003). Increasingsalinity is accompanied by significant reductions in
Table 5
Growth of B. parviflora after 45 days in nutrient solution containing 0–400m
[NaCl] (mM) Plant height
(cm/plant)
Leaf area (cm2/
plant)
F
p
0 24.070.62a 54.970.35a 8
100 25.570.82a 58.970.90b 8
200 23.270.55a 46.270.62c 8
400 22.570.91a 44.670.55c 7
Values are mean7SE of three independent experiments. Different letters nParida et al., 2004a).
shoot weight, plant height, number of leaves per plant,root length, and root surface area per plant in tomato(Mohammad et al., 1998). Increased NaCl levels resultsin a significant decrease in root, shoot, and leaf growthbiomass and increase in root/shoot ratio in cotton(Meloni et al., 2001).
3.2. Effects of salinity on water relations
Water potential and osmotic potential of plantsbecome more negative with an increase in salinity,whereas turgor pressure increases with increasingsalinity (Morales et al., 1998; Hernandez et al., 1999;Khan et al., 1999; Meloni et al., 2001; Khan, 2001;Romeroaranda et al., 2001). Leaf water and osmoticpotentials and xylem tension increase with an increase inmedia salinity in R. mucronata (Aziz and Khan, 2001).Leaf osmotic potentials decrease with increases in NaClin Chrysanthemum and sea aster (Matsumura et al.,1998). Relative water content, leaf water potential,water uptake, transpiration rate, water retention, andwater use efficiency decrease under short-term NaClstress in jute (Chaudhuri and Choudhuri, 1997). Waterpotential, osmotic potential, and stomatal conductancebecome more negative with an increase in salinity, whilepressure potential decreases with increasing salinity inthe halophytic perennial grass Urochondra setulosa
(Trip.) (Gulzar et al., 2003). With increasing saltconcentration, leaf water potential and evaporation ratedecrease significantly in the halophyte S. salsa whilethere are no changes in leaf relative water content (Lu etal., 2002). Leaf water potential and osmotic potentialdecline depending on the osmotic potential of therooting medium and the mode of stress imposition. Agreater decline in osmotic potential compared with thetotal water potential led to turgor maintenance in plantsunder progressive or prolonged NaCl stress (Rajasekar-an et al., 2001).
3.3. Effects of salinity on leaf anatomy
Salinity causes increases in epidermal thickness,mesophyll thickness, palisade cell length, palisadediameter, and spongy cell diameter in leaves of bean,
M NaCl
resh matter of
lant (g/plant)
Dry matter of
plant (g/plant)
Water content per unit leaf
area (gm�2)
.3870.51a 1.9470.05a 301.175.51a
.7270.35a 2.0470.04a 302.072.70a
.1170.25a 1.7770.04a 319.372.35b
.0270.20a 1.3470.05a 324.471.85c
ext to values indicate statistically different means at Pp0:05 (Source:
ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349336
cotton, and Atriplex (Longstreth and Nobel, 1979). Incontrast both epidermal and mesophyll thickness andintercellular spaces decrease significantly in NaCl-treated leaves of the mangrove B. parviflora (Parida etal., 2004a). Salinity also reduces intercellular spaces inleaves (Delphine et al., 1998). Salt stress causes (1)vacuolation development and partial swelling of en-doplasmic reticulum, (2) decrease in mitochondrialcristase and swelling of mitochondria, (3) vesiculationand fragmentation of tonoplast, and (4) degradation ofcytoplasm by the mixture of cytoplasmic and vacuolarmatrices in leaves of sweet potato (Mitsuya et al., 2000)(Fig. 4). In leaves of potato salt stress causes rounding ofcells, smaller intercellular spaces, and a reduction inchloroplast number (Bruns and Hecht–Buchholz, 1990).In tomato plants salinity causes reduction of plant leafarea and stomatal density (Romeroaranda et al., 2001).
3.4. Effects of salinity on photosynthetic pigments and
proteins
The chlorophyll and total carotenoid contents ofleaves decrease in general under salt stress. The oldestleaves start to develop chlorosis and fall with prolongedperiod of salt stress (Hernandez et al., 1995, 1999;Gadallah, 1999; Agastian et al., 2000). However, Wangand Nil (2000) have reported that chlorophyll contentincreases under conditions of salinity in Amaranthus. InGrevilea, protochlorophyll, chlorophylls, and carote-
Fig. 4. Effects of NaCl stress on polyphenol level in B. parviflora
measured as a function of days of NaCl treatment. Values are
mean7SE (from Parida et al., 2002).
Table 6
Effects of NaCl stress on photosynthetic pigments in leaves of B. parviflora
[NaCl] (mM) Chl a (mg cm�2) Chl b (mg cm�2)
0 65.4871.2 17.9671.1100 67.9971.6 20.6571.8200 40.0671.0 12.0671.3400 35.5371.3 11.0371.4
4th pair of leaves from shoot top were sampled after 45 days NaCl treatmen
noids are significantly reduced under NaCl stress, butthe rate of decline of protochlorophyll and chlorophyllis greater than that of Chl-a and carotenoids. Theanthocyanin pigments on the other hand significantlyincrease in this case with salt stress (Kennedy and DeFillippis, 1999). In leaves of tomato, the contents oftotal chlorophyll (Chl-a+b), Chl-a, and b carotenedecrease by NaCl stress (Khavarinejad and Mostofi,1998). Under salinity stress, leaf pigments studied innine genotypes of rice reduce in general, but relativelyhigh pigment levels are found in six genotypes (Alamgirand Ali, 1999). In the cyanobacterium Spirulina platensis
a decrease in the phycocyanin/chlorophyll ratio and nosignificant change in the carotenoid/chlorophyll ratioare observed under salt stress (Lu and Vonshak, 1999).Salinity causes significant decreases in Chl-a, Chl-b, andcarotenoid in leaves of B. parviflora (Table 6; Parida etal., 2002).Soluble protein contents of leaves decrease in
response to salinity (Alamgir and Ali, 1999; Gadallah,1999; Wang and Nil, 2000; Muthukumarasamy et al.,2000; Parida et al., 2002). Agastian et al. (2000) havereported that soluble protein increases at low salinityand decreases at high salinity in mulberry. In Rhizobium
certain outer membrane proteins of molecular weight22, 38, 40, 42, 62, and 68 kDa markedly decrease in thepresence of salt (Unni and Rao, 2001). SDS–PAGEanalysis of proteins in peanut (Arachis hypogaea L.)reveals that plants grown under NaCl show induction(127 and 52 kDa) or repression (260 and 38 kDa) in thesynthesis of a few polypeptides (Hassanein, 1999).Salinity induces six new proteins in roots of barley,which are of low molecular weight, 24 to 27 kDa, withan isoelectric point of 6.1 to 7.6. In contrast to roots, fivenew shoot proteins are induced whose molecular weightsand isoelectric points fall within the range of 20–24 kDaand 6.3–7.2, respectively. In addition, salinity alsoinhibits the synthesis of a majority of shoot proteins(Ramagopal, 1987). In radish (R. sativus L.) salt stresscauses accumulation of a 22-kDa protein (pI of 7.5) andits mRNA in the leaves (Lopez et al., 1994). Asignificant increase in the amount of a protein, whosemigration in two-dimensional (2D) gel electrophoresiscorresponds to an apparent molecular weight of23–25 kDa and a pI of 6.1, is observed under NaClstress in cultured cells derived from Shamuti orange
Total Chl Chl a/b Total carotenoids (mg cm�2)
83.4471.8 3.64 23.2071.588.6471.5 3.29 23.6571.452.1271.4 3.32 16.5571.146.5671.1 3.20 13.6871.0
t. The values are mean7SE (n ¼ 6) (Source: Parida et al., 2004a).

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 337
(Citrus sinensis L. Osbeck) ovular callus cells (Ben-hayyim et al., 1993). Yen et al. (1997) have reported theaccumulation of five polypeptides with estimatedmolecular masses of 40, 34, 32, 29, and 14 kDa bySDS and 2D-PAGE in calli of M. crystallinum underNaCl stress and these polypeptides are classified intotwo distinct groups according to their course ofinduction: early responsive (40-, 34-, and 29-kDa) andlate responsive (32- and 14-kDa) proteins. The disap-pearance of a 60-kDa polypeptide in response to salinityhas been observed in Prosopsis (Munoz et al., 1997). Inwheat, the content of a 26-kDa protein increases inNaCl-treated plants, stimulation is more pronounced inroots than in shoots, and, in contrast, the contents of 13-and 20-kDa proteins decrease and the 24-kDa proteindisappears with NaCl treatment (Elshintinawy andElshourbagy, 2001). NaCl induces accumulation of fourpolypeptides with molecular masses of 61, 51, 39, and29 kDa in maize roots (Tamas et al., 2001). Fractiona-tion of cells followed by SDS–PAGE and 2D-PAGEreveals an increase in the levels of membrane proteins of39 and 50 kDa and a decrease in the level of a membraneprotein of 52 kDa with increasing levels of externalNaCl in Rhodobacter sphaeroides. The proteins havebeen isolated and sequenced, the polypeptide of 50 kDain the inner membrane is assigned to an ATP synthasebeta chain and that of 52 kDa in the outer membrane toa flagellar filament protein, and, as the N terminus of the39 kDa protein in the outer membrane is blocked,partial proteolysis was carried out and four peptideswere sequenced. Each sequence exhibits no significanthomology with those available in databases, suggestingthat the polypeptide of 39 kDa (named SspA) is a novelsalt-stress-induced protein (Xu et al., 2001). It has beenreported that salinity causes a decrease in intensity ofseveral protein bands of molecular weight 17, 23, 32, 33,and 34 kDa in B. parviflora (Parida et al., 2004b) and thedegree of decrease of these protein bands seems to beroughly proportional to the external NaCl concentra-tion. The most obvious change concerns a 23-kDapolypeptide, which disappears after 45 days of treatmentin 400mM NaCl (Fig. 2). Moreover, the 23-kDapolypeptide, which disappears in B. parviflora undersalinity stress, reappears when these salinized seedlingsare desalinized (Fig. 2) and these observations suggestthe possible involvement of these polypeptides forosmotic adjustment under salt stress in this species(Parida et al., 2004c).
3.5. Effects of salinity on lipids
Lipids are the most effective sources of storageenergy, they function as insulators of delicate internalorgans and hormones, and they play important roles asthe structural constituents of most of the cellularmembranes (Singh et al., 2002). They also have vital
roles in the tolerance to several physiological stressors ina variety of organisms including cyanobacteria. Themechanism of desiccation tolerance relies on phospho-lipid bilayers, which are stabilized during water stress bysugars, especially by trehalose. Unsaturation of fattyacids also counteracts water or salt stress. Hydrogenatoms adjacent to olefinic bonds are susceptible tooxidative attack. Lipids are rich in these bonds and are aprimary target for oxidative reactions. Lipid oxidation isproblematic as enzymes do not control many oxidativechemical reactions and some of the products of theattack are highly reactive species that modify proteinsand DNA (Singh et al., 2002). The lipid content inpeanut (A. hypogaea L.) increases at low concentrationof NaCl (up to 45mM) and decreases at higherconcentrations (Hassanein, 1999). Wu et al. (1998) haveanalyzed the changes in lipid composition by NaClstress in root plasma membrane of salt marsh grass(Spartina patens) and reported that molar percentages ofsterols (including free sterols) and phospholipid de-creases with increasing salinity, but the sterol/phospho-lipid ratio is unaffected by NaCl. However, glycolipidshows a statistically significant increase in the total lipidas salinity in the medium is increased and the content ofplasma membrane phosphatidylcholine (PC) and phos-phatidylethalomine (PE) decrease by salinity, but thePC/PE ratios are not affected by salinity. Plasmamembrane vesicles isolated from calli of tomato tolerantto 100mM NaCl exhibit higher phospholipid and sterolcontent and lower phospholipid/free sterol ratio andlower double bond index of phospholipid fatty acids(Kerkeb et al., 2001).
3.6. Effects of salinity on ion levels
High salt (NaCl) uptake competes with the uptake ofother nutrient ions, especially K+, leading to K+
deficiency. Increased treatment of NaCl induces increasein Na+ and Cl� and decrease in Ca2+, K+, and Mg2+
levels in a number of plants (Khan et al., 1999, 2000a;Khan, 2001). Salinity enhances the content of Na+,Ca2+, and Cl� in Vicia faba and the ratio of K+/Na+
decreases (Gadallah, 1999). In A. pseudoalhagi (aleguminous plant), the leaf Na+ concentration in200mM NaCl-treated plants increases to 45 times thatof the control and the plants do not die but continue togrow at such a high leaf Na+ concentration (Kurban etal., 1999). In Ulva fasciata, a marine green macro alga,the contents of Na+, K+, and Cl� accumulate linearlywith increasing salinity. An increase in Na+ and Cl�
content induces proline accumulation but decreasesboth the activity of proline dehydrogenase (PDH EC1.4.3.1; a catabolic enzyme of proline) and the totaland water-soluble Ca2+ contents. These results suggestthat in U. fasciata a loss of cellular Ca2+ is associatedwith NaCl induction of proline accumulation via

ARTICLE IN PRESS
Table 7
Effects of salt stress on ion levels in leaves, stems, and root of B. parviflora treated with different concentrations of NaCl for a period of 45 days
Plant parts [NaCl] (mM) Na K Ca Mg Fe Cu Mn Cl
Leaf 0 14.4770.7 11.0270.1 12.3170.3 155.9471.9 325.2171.4 31.5070.3 103.1071.6 13.1170.1100 33.5470.3 12.2670.7 11.4170.2 162.4971.6 309.0771.8 28.5070.2 97.4170.8 38.5370.6200 38.6370.4 10.8370.6 9.9970.5 145.4671.2 254.1171.5 20.4170.5 53.0670.6 40.2170.4400 42.1370.2 11.8170.2 7.8170.6 117.1471.4 295.7571.3 18.3770.64 7.1070.6 43.4970.4
Stem 0 9.4870.5 6.0070.5 2.4970.9 65.4170.7 137.171.8 4.3570.7 17.0570.9 7.2870.1100 15.8170.7 7.8670.7 2.9170.6 61.5470.3 187.070.8 5.6970.3 21.0770.4 21.9170.3200 16.7070.1 3.8170.3 2.2670.7 88.3470.5 190.570.5 2.3970.5 14.0170.7 26.7070.6400 21.6770.2 2.7170.2 3.6370.5 298.4170.3 220.471.5 1.4870.9 12.6370.5 38.5470.8
Root 0 7.2170.5 25.4570.4 2.7470.6 205.6870.8 632.171.8 65.9070.9 28.4070.5 6.8570.7100 14.4770.3 17.7870.1 2.8970.3 159.5570.4 970.971.1 51.1070.5 37.0070.5 18.3770.3200 17.2570.5 15.0370.9 2.4570.8 142.8770.5 383.970.9 28.1071.2 33.0770.3 24.9570.5400 26.9670.8 11.2070.5 2.3870.3 116.7870.6 277.870.6 14.5270.7 11.9670.7 31.4770.3
Na, K, Ca, Mg, and Cl content are expressed as mg g dwt�1, whereas Fe, Cu, and Mn content are expressed as mg g dwt�1. Values are mean7S.E(n ¼ 6) (Source: Parida et al., 2004a).
A.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349338
an inhibition of PDH activity (Lee and Liu, 1999).Salinity stress causes an increase in levels of Na+ andCl� in guava and the highest ion accumulation is foundin the leaves followed by the roots; the Ca2+ levels arestable in the roots but decrease in stems and leaves andthe K+ content is reduced with increased levels ofsalinity, particularly in the leaves. On the other hand,Mg2+ levels are not affected by salinity in stems androots but decrease in the leaves of guava. There is apositive relationship between Na+ and Cl� and anegative relationship between Na+ and K+ concentra-tion in roots and leaves. Mg2+ concentration in leavesand roots does not vary with the concentration of Na+,and the concentration of Ca2+ does not vary with thatof Na2+ in the leaves but shows an inverse relationshipin the roots (Ferreira et al., 2001). We have reported asignificant increase in Na+ and Cl� content in leaves,stem, and root of the mangrove B. parviflora (Table 7)without any significant alteration of the endogenouslevel of K+ and Fe2+ in leaves (Parida et al., 2004a).Decreases of Ca2+ and Mg2+ content of leaves havealso been reported upon salt accumulation in thisspecies, suggesting increasing membrane stability anddecreased chlorophyll content, respectively (Paridaet al., 2004a).
3.7. Effects of salinity on antioxidative enzymes and
antioxidants
Salt stress causes water deficit as a result of osmoticeffects on a wide variety of metabolic activities of plantsand this water deficit results in oxidative stress becauseof the formation of reactive oxygen species such assuperoxides and hydroxy and peroxy radicals. Thereactive oxygen species that are by-products of hyper-osmotic and ionic stresses cause membrane disfunctionand cell death (Bohnert and Jensen, 1996). The plants
defend against these reactive oxygen species by induc-tion of activities of certain antioxidative enzymes suchas catalase, peroxidase, glutathione reductase, andsuperoxide dismutase, which scavenge reactive oxygenspecies. There are several reports of increasing activityof antioxidative enzymes. Activities of antioxidativeenzymes such as ascorbate peroxidase, glutathionereductase, monodehydroascorbate reductase(MDHAR), dehydroascorbate reductase (DHAR), andMn-SOD increase under salt stress in wheat, while Cu/Zn-SOD remains constant and total ascorbate andglutathione content decrease (Hernandez et al., 2000). Inwheat, activities of APX, MDHAR, DHAR, and GRincrease in the shoots and decrease in the roots(Meneguzzo and Navarilzzo, 1999). Gossett et al.(1994) have reported that in cotton (Gossypium hirsutum
L.) NaCl stress increases the activities of SOD, guaicolperoxidase, and glutathione reductase and decreases theactivities of catalase and ascorbate peroxidase. Saltstress also causes decrease in total ascorbate, totalglutathione, and a-tocopherol levels in this case. Inleaves of rice plant, salt stress preferentially enhances thecontent of H2O2 and the activities of SOD, APX, andGPX, whereas it decreases catalase activity (Lee et al.,2001). On the other hand, salt stress has little effect onthe activity levels of glutathione reductase (Lee et al.,2001). Lechno et al. (1997) have reported that NaCltreatment increases the activities of the antioxidativeenzymes catalase and glutathione reductase and thecontent of the antioxidants ascorbic acid and reducedglutathione but does not affect the activity of SOD incucumber plants. In radish NaCl stress decreases prolineoxidase activity and increases protease, gamma-gluta-myl kinase, and ATPase activities (Muthukumarasamyet al., 2000). SOD activity in plant leaves of barley andH+ ATPase activity in plant roots increase by salinity,whereas malondialdehyde (MDA) concentration in
ARTICLE IN PRESS
0
5
10
15
20
25
30
0 7 14 21 30 45
CA
T (
Um
g-1
pro
tein
)
0mM100mM200mM
(A)
(B)
(C)
(D)
400mM
0mM100mM200mM400mM
0mM100mM200mM400mM
0mM100mM200mM400mM
a
c
aab
00.10.20.30.40.50.60.70.8
0 7 14 21 30 45
AP
X (
Um
g-1
pro
tein
)
bb
aa
00.050.1
0.150.2
0.250.3
0.350.4
0.450.5
0 7 14 21 30 45
GR
(U m
g-1
pro
tein
) c
b
aa
0
5
10
15
20
0 7 14 21 30 45Duration of exposure (days)
Duration of exposure (days)
SO
D(U
mg
-1 p
rote
in) cc
b
a
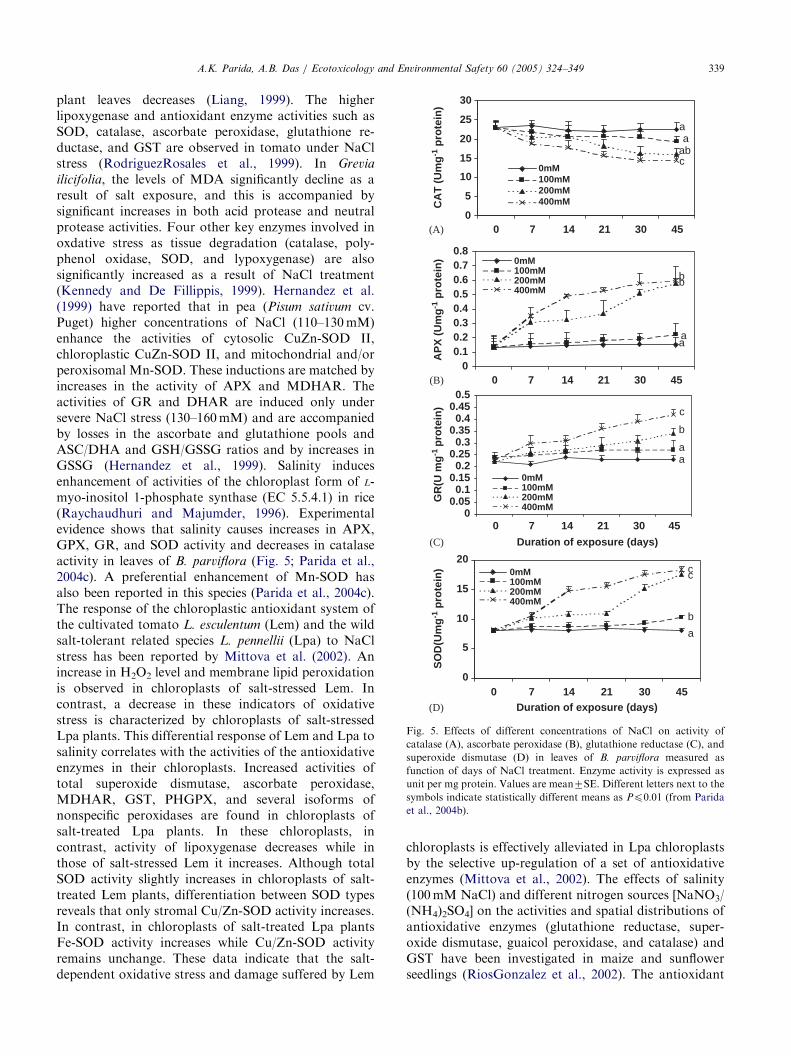
Fig. 5. Effects of different concentrations of NaCl on activity of
catalase (A), ascorbate peroxidase (B), glutathione reductase (C), and
superoxide dismutase (D) in leaves of B. parviflora measured as
function of days of NaCl treatment. Enzyme activity is expressed as
unit per mg protein. Values are mean7SE. Different letters next to thesymbols indicate statistically different means as Pp0:01 (from Parida
et al., 2004b).
A.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 339
plant leaves decreases (Liang, 1999). The higherlipoxygenase and antioxidant enzyme activities such asSOD, catalase, ascorbate peroxidase, glutathione re-ductase, and GST are observed in tomato under NaClstress (RodriguezRosales et al., 1999). In Grevia
ilicifolia, the levels of MDA significantly decline as aresult of salt exposure, and this is accompanied bysignificant increases in both acid protease and neutralprotease activities. Four other key enzymes involved inoxdative stress as tissue degradation (catalase, poly-phenol oxidase, SOD, and lypoxygenase) are alsosignificantly increased as a result of NaCl treatment(Kennedy and De Fillippis, 1999). Hernandez et al.(1999) have reported that in pea (Pisum sativum cv.Puget) higher concentrations of NaCl (110–130mM)enhance the activities of cytosolic CuZn-SOD II,chloroplastic CuZn-SOD II, and mitochondrial and/orperoxisomal Mn-SOD. These inductions are matched byincreases in the activity of APX and MDHAR. Theactivities of GR and DHAR are induced only undersevere NaCl stress (130–160mM) and are accompaniedby losses in the ascorbate and glutathione pools andASC/DHA and GSH/GSSG ratios and by increases inGSSG (Hernandez et al., 1999). Salinity inducesenhancement of activities of the chloroplast form of L-myo-inositol 1-phosphate synthase (EC 5.5.4.1) in rice(Raychaudhuri and Majumder, 1996). Experimentalevidence shows that salinity causes increases in APX,GPX, GR, and SOD activity and decreases in catalaseactivity in leaves of B. parviflora (Fig. 5; Parida et al.,2004c). A preferential enhancement of Mn-SOD hasalso been reported in this species (Parida et al., 2004c).The response of the chloroplastic antioxidant system ofthe cultivated tomato L. esculentum (Lem) and the wildsalt-tolerant related species L. pennellii (Lpa) to NaClstress has been reported by Mittova et al. (2002). Anincrease in H2O2 level and membrane lipid peroxidationis observed in chloroplasts of salt-stressed Lem. Incontrast, a decrease in these indicators of oxidativestress is characterized by chloroplasts of salt-stressedLpa plants. This differential response of Lem and Lpa tosalinity correlates with the activities of the antioxidativeenzymes in their chloroplasts. Increased activities oftotal superoxide dismutase, ascorbate peroxidase,MDHAR, GST, PHGPX, and several isoforms ofnonspecific peroxidases are found in chloroplasts ofsalt-treated Lpa plants. In these chloroplasts, incontrast, activity of lipoxygenase decreases while inthose of salt-stressed Lem it increases. Although totalSOD activity slightly increases in chloroplasts of salt-treated Lem plants, differentiation between SOD typesreveals that only stromal Cu/Zn-SOD activity increases.In contrast, in chloroplasts of salt-treated Lpa plantsFe-SOD activity increases while Cu/Zn-SOD activityremains unchange. These data indicate that the salt-dependent oxidative stress and damage suffered by Lem
chloroplasts is effectively alleviated in Lpa chloroplastsby the selective up-regulation of a set of antioxidativeenzymes (Mittova et al., 2002). The effects of salinity(100mM NaCl) and different nitrogen sources [NaNO3/(NH4)2SO4] on the activities and spatial distributions ofantioxidative enzymes (glutathione reductase, super-oxide dismutase, guaicol peroxidase, and catalase) andGST have been investigated in maize and sunflowerseedlings (RiosGonzalez et al., 2002). The antioxidant

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349340
enzymes exhibit higher activities in salinity-treatedplants. Changes in antioxidant enzyme activities causedby different N sources differ in the two species.Ammonium-fed plants show higher CAT activity inboth plant species and higher GR activity in maize andsunflower leaves, with the highest GST activity in maize.POD and SOD activities are lower in both maize andsunflower seedlings and lower GR activity has beenobserved in maize roots. SOD and POD activities arehigher in the mature sections of the root than in the tips.GR activity is higher in the younger parts of the nitrate-fed plant roots. The antioxidant enzyme activities arehigher in the cortex than in the stele of the nodal roots(RiosGonzalez et al., 2002). The response of theantioxidative systems of leaf cell mitochondria andperoxisomes of the cultivated tomato L. esculentum andthe wild salt-tolerant related species L. pennellii to NaClstress has also been investigated (Mittova et al., 2003).Salt-dependent oxidative stress is evident in Lemmitochondria as indicated by their raised levels of lipidperoxidation and H2O2 content whereas their reducedascorbate and reduced glutathione contents decrease.Concomitantly, SOD activity decreases whereas APXand GPX activities remain at control level. In contrast,the mitochondria of salt-treated Lpa do not exhibit salt-induced oxidative stress. In their case salinity induces anincrease in the activities of superoxide dismutase,ascorbate peroxidase, MDHAR, dehydroascorbate re-ductase, and glutathione-dependent peroxidase. Lpaperoxisomes exhibit increased SOD, APX, MDHAR,and catalase activity and their lipid peroxidation andH2O2 levels are not affected by the salt treatment. Theactivities of all these enzymes remain at control level inperoxisomes of salt-treated Lem plants. The salt-induced increase in the antioxidant enzyme activities inthe Lpa plants confer cross-tolerance toward enhancedmitochondrial and peroxisomal reactive oxygen speciesproduction imposed by salicylhydroxamic acid and 3-amino-1,2,4-triazole, respectively (Mittova et al., 2003).
3.8. Effects of salinity on nitrogen metabolism
Nitrate reductase activity (NRA) of leaves decreasesin many plants under salt stress (AbdElBaki et al., 2000;Flores et al., 2000). The primary cause of a reduction ofNRA in the leaves is a specific effect associated with thepresence of Cl� salts in the external medium. This effectof Cl� seems to be due to a reduction in NO3
� uptakeand consequently a lower NO3
� concentration in theleaves, although a direct effect of Cl�on the activity ofthe enzyme cannot be discarded (Cram, 1973; Smith,1973; Deane-Drummond and Glauss, 1982; Flores et al.,2000). In Zea mays the nitrate content of leavesdecreases, but it increases in roots under NaCl stressand NRA of leaves also decreases under salinity(AbdElBaki et al., 2000). Soussi et al. (1999) have
reported that salinity inhibits nitrogen fixation byreducing nodulation and nitrogenase activity in chick-pea (C. arietinum L.). Considerable inhibition ofnodulation and N2 fixation has also been reported byother workers (Bekki et al., 1987). Severe salt stressreduces the leg hemoglobinn content and nitrogenaseactivity in soybean root nodules (Comba et al., 1998).The exposure of nodulated roots of legumes such assoybean, common bean, and alfalfa to NaCl results in arapid decrease in plant growth associated with a short-term inhibition of both nodule growth and nitrogenaseactivity (Serraz et al., 1998). Khan et al. (1998) havereported that salinity inhibits nitrate content and uptakeand NRA in leaves of maize plants. Nitrogenase activitymeasured with regard to acetylene reduction (C2H2)decreases in common bean by short-term exposure tosalinity (Serraz et al., 2001). Decrease in NRA activityand in total nitrogen and nitrate uptake have beenreported in leaves of B. parviflora (Parida and Das,2004).NADP-specific isocitrate dehydrogenase (ICDH) is a
key cytosolic enzyme that links C and N metabolism bysupplying C skeletons for primary N assimilation inplants. The characterization of the transcript Mc-
ICDH1 encoding an NADP-dependent isocitrate dehy-drogenase (NADP-ICDH; EC 1.1.1.42) has been re-ported from the facultative halophyte M. crystallinum
L., focusing on salt-dependent regulation of the enzyme(Popova et al., 2002). The activity of NADP-ICDH inplants adapted to high salinity increases in leaves anddecreases in roots. By transcript analyses and Western-type hybridizations, expression of Mc-ICDH1 is foundto be stimulated in leaves in salt-adapted M. crystal-
linum. By immunocytological analyses, NADP-ICDHproteins are localized to most cell types with strongestexpression in epidermal cells and in the vascular tissue.In leaves of salt-adapted plants, signal intensitiesincreases in mesophyll cells. In contrast to Mc-ICDH1,the activity and transcript abundance of ferredoxin-dependent glutamate synthase (EC 1.4.7.1), which is thekey enzyme of N assimilation and biosynthesis of aminoacids, decreases in leaves in response to salt stress(Popova et al., 2002).
3.9. Effects of salinity on malate metabolism
NADP-malate dehydrogenase (NADP-MDH; EC1.1.1.82) is responsible for the reduction of oxaloacetateto malate in the chloroplasts of higher plants. In M.
crystallinum, steady state transcript levels for chloro-plast NADP-MDH decrease transiently in the leavesafter salt stress and then increase to levels greater thantwo fold higher than levels in unstressed plants, whereastranscript levels in roots are extremely low and areunaffected by salt-stress treatment and the salt-stress-induced expression pattern of this enzyme suggests that

ARTICLE IN PRESS
0
2
4
6
8
10
12
14
16
18
7 14 30 45
Duration of NaCl treatment (d)
PN (
µmo
lm-2
s-1
)
0 mM
100 mM
200 mM
400 mM
Fig. 6. Effect of NaCl treatment on photosynthetic rate (PN) in B.
parviflora measured as a function of days of NaCl treatment. Values
are mean7SE (from Parida et al., 2004a).
A.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 341
it may participate in the CO2 fixation pathway duringCAM (Cushman, 1993). When Eucalyptus citridora
plants are treated with NaCl, an increased Na+ levelis observed in shoots 3 weeks after treatment, and at thisstage of treatment the growth of plants is not reducedbut the malate metabolism is modified. The malatecontent decreases in leaves while the specific activities ofNAD and NADP-malic enzymes increase. The stimula-tion in enzyme activity is more pronounced for NADP-malic enzyme but, for both enzymes, enzyme activitydiminishes as early as 5 weeks after treatment (DeAr-agao et al., 1997).
3.10. Effects of salinity on chloroplast ultrastructure
Electron microscopy shows that the thylakoidalstructure of the chloroplasts becomes disorganized, thenumber and size of plastoglobuli increases, and theirstarch content decreases in plants treated with NaCl(Hernandez et al., 1995, 1999). In the mesophyll of sweetpotato leaves, thylakoid membranes of chloroplast areswollen and most are lost under severe salt stress(Mitsuya et al., 2000). In potato, salt stress reduces thenumbers and depth of the grana stacks and causes aswelling of the thylakoid and starch grains becomelarger in the chloroplasts (Bruns and Hecht-Buchholz,1990). In leaves of NaCl-treated plants of tomato thetransmission electron microscopy shows that the chlor-oplasts are aggregated, the cell membranes are distortedand wrinkled, and there are no signs of grana orthylakoid structures in chloroplasts (Khavarinejad andMostofi, 1998). Chloroplast ultrastructural changesunder salt stress are apparent in Eucalyptus microcorys;these include the presence of large starch grains, thedilation of the thylakoid membranes, the near absenceof grana, and the presence of enlarged mesophyll cells(Keiper et al., 1998). We have also reported a notabledisorganization of the thylakoid structure of chloro-plasts in leaves of B. parviflora by salt stress (Parida etal., 2003).
3.11. Mechanism of salinity effects on photosynthesis
Plant growth is the result of integrated and regulatedphysiological processes. Physiological processes areaffected by number of environmental factors and theydetermine the response of plants to stress. Limitation ofplant growth by environmental factors cannot beassigned to a single physiological process. The dominantphysiological process is photosynthesis. Plant growth asbiomass production is a measure of net photosynthesisand, therefore, environmental stresses affecting growthalso affect photosynthesis. Salt stress causes either short-or long-term effects on photosynthesis. The short-termeffect occurs after a few hours or within 1 or 2 days ofthe onset of exposure and this response is important, as
there is complete cessation of carbon assimilation withinhours. The long-term effect occurs after several days ofexposure to salt and reduction in carbon assimilation isdue to the salt accumulation in developing leaves(Munns and Termatt, 1986). Although there are reportsof suppression of photosynthesis upon salt stress(Chaudhuri and Choudhuri, 1997; Soussi et al., 1998;AliDinar et al., 1999; Romeroaranda et al., 2001; Kao etal., 2001), there are also reports that photosynthesis isnot slowed down by salinity and is even stimulated bylow salt concentration (Rajesh et al., 1998; Kurban etal., 1999). In A. pseudoalhagi (a leguminous plant), theleaf CO2 assimilation rate increases at low salinity(50mM NaCl) but is not affected significantly by100mM NaCl, while it is reduced to about 60% of thecontrol in 200mM NaCl. Similarly stomatal conduc-tance is consistent with the CO2 assimilation rateregardless of the treatments, and intercellular CO2concentration is lower in the NaCl-treated plants thanin the control (Kurban et al., 1999). In mulberry, netCO2 assimilation rate (PN), stomatal conductance (gs),and transpiration rate (E) decline under salt stress,whereas intercellular CO2 concentration (Ci) increases(Agastian et al., 2000). In B. parviflora net CO2 PNincreases at low salinity (mM) and decreases at highsalinity (Fig. 6), whereas gs remains the same as incontrol at low salinity and decreases at high salinity(Fig. 7) (Parida et al., 2004a). NaCl stress decreases thechlorophyll content and net photosynthetic rate andincreases the rate of respiration and CO2 compensationconcentration and there are no significant changes in thecarotenoid contents in leaves of alfalfa plants (Khavar-inejad and Chaparzadeh, 1998). In A. lentiformis,net CO2 assimilation rate and the ratio of ribulose

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349342
biphosphate carboxylase (Rubisco) activity to that ofphosphoenol pyruvate carboxylase (PEPC) decreaseunder salinity because PEPC activity on a leaf areabasis increases linearly with salinity, while Rubiscoactivity remains relatively constant (Zhu and Meinzer,1999). In the cyanobacterium S. platensis salt stressinhibits the apparent quantum efficiency of photosynth-esis and photosystem II (PSII) activity while stimulatingphotosystem I (PSI) activity and dark respirationsignificantly. Salt stress also results in a decrease inoverall activity of the electron transport chain, whichcannot be restored by diphenyl carbazide, an artificialelectron donor to PSII (Lu and Vonshak, 1999).Experimental evidence shows that PSII mediatedelectron transport activity increases at low salinity(100mM) and decreases at high salinity in B. parviflora
(Table 8; Parida et al., 2003). A significant reduction inCO2 assimilation rate and stomatal conductance by salttreatment is registered in horsegram (Macrotyloma
0
100
200
300
400
500
600
700
800
7 14 30 45
Duration of NaCl treatment (d)
gs
(mm
olm
-2 s
-1)
0 mM
100 mM
200 mM
400 mM
Fig. 7. Effect of NaCl treatment on stomatal conductance (gs) in B.
parviflora measured as a function of days of NaCl treatment. Values
are mean7SE (from Parida et al., 2004a).
Table 8
Effects of NaCl (mM) on PSII-mediated electron transport activity (mmophotoreduction
[NaCl] (mM) 0 days
H2O-DCPIP 0 72.9772.5100 72.977 2.7
200 72.9772.0400 72.9772.3
DPC-DCPIP 0 113.4072.7100 113.8972.0200 112.5772.0400 112.8071.5
The rate was measured at different days after NaCl treatment in a reaction mi
or as DPC-DCPIP. The values are mean7SE (n ¼ 6) (Source: Parida et a
uniflorum) and salinity inhibits the photosynthetic elect-ron transport and the activity of enzymes of the Calvincycle, ribulose-1,5-biphosphate carboxylase (E.C.4.1.1.39),ribulose-5-phosphate kinase (E.C.2.7.1.19), ribulose-5-phosphate isomerase (E.C.5.3.16), and NADP-glyceralde-hyde-3-phosphate dehydrogenase (E.C.1.2.13) (Reddy etal., 1992). In four cultivars of rice (Oryza sativa L.),gradual decreases in the activities of PSI, PSII, andchlorophyll fluorescence transients and emission at 688nmare observed with increase in NaCl concentration and adrastic decrease in net photosynthetic rate is found(Tiwari et al., 1997). Mishra et al. (1991) have reportedthat in wheat (T. aesivum L.) salt stress has no direct effecton electron transport activity and F(v)/F(m) ratio,suggesting that the efficiency of the photochemistry ofPSII is not affected and decrease in F(m) due to salt stressmay influence reduction of Q(A). These results onfluorescence indicate that salt stress predisposes photo-inhibition of plants and reduces their ability to recoverfrom photoinhibition.Light stress and salt stress are major environmental
factors that limit the efficiency of photosynthesis.However, Allakhverdiev et al. (2002) have reported thatthe effects of light and salt stress on PSII in thecyanobacterium Synecchocystis sp. PCC 6803 arecompletely different. Strong light induces photodamageto PS II, whereas salt stress inhibits the repair ofphotodamaged PSII and does not accelearate damage toPSII directly. The combination of light and salt stressappears to inactivate PSII very rapidly as a consequenceof their synergistic effects. Radioactive labeling of cellsreveals that salt stress inhibits the synthesis of proteinsde novo and, in particular, the synthesis of the D1protein of PSII. Northern and Western blotting analysesby Allakhverdiev et al. (2002) demonstrate that saltstress inhibits the transcription and the translation ofpsbA genes, which encode D1 protein. DNA microarrayanalysis indicates that the light-induced expression ofvarious genes is suppressed by salt stress. It has beensuggested that salt stress inhibits the repair of PSII via
l mg chl–1 h�1) in B. parviflora as monitored with regard to DCPIP
15 days 30 days 45 days
71.1271.3 71.8672.0 70.6871.775.1071.5 75.3571.3 75.8171.570.5271.5 65.6371.7 61.2871.068.2772.0 60.8271.0 54.8172.5
111.6271.3 111.5871.0 110.5571.8113.5171.8 115.5472.3 114.6272.5110.7071.5 106.7372.5 93.4772.3104.8272.0 85.8172.4 78.6571.7
xture at pH 7.0 as H2O-DCPIP without any uncoupler and inhibitors
l., 2003).

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 343
suppression of the activities of the transcriptional andtranslational machinery (Allakhverdiev et al., 2002).Photosynthesis involves a long chain of mechanisms,
enzymes, and intermediate products and is regulated byseveral external and internal factors. Photosyntheticefficiency depends on the sequence of metabolic eventssuch as photochemical reactions, on the enzymesinvolved in carbon assimilation, on the structure of thephotosynthetic apparatus, and on the transport ofphotosynthetic intermediates between subcellular com-partments.Photosynthetic rate is lower in salt-treated plants, but
the photosynthetic potential is not greatly affected whenrates are expressed with regard to chlorophyll or leafarea. Decreases in photosynthetic rate are due to severalfactors: (1) dehydration of cell membranes which reducetheir permeability to CO2, (2) salt toxicity, (3) reductionof CO2 supply because of hydroactive closure ofstomata, (4) enhanced senescence induced by salinity,(5) changes of enzyme activity induced by changes incytoplasmic structure, and (6) negative feedback byreduced sink activity (Iyengar and Reddy, 1996).Photosynthetic activity decreases as water potential of
leaves decreases (Iyengar and Reddy, 1996). Thereduction in photosynthetic activity depends on twoaspects of salinization, i.e., the total concentration ofsalt and their ionic composition. High salt concentrationin soil and water create high osmotic potential, whichreduces the availability of water to plants. Decrease inwater potential causes osmotic stress, which reversiblyinactivates photosynthetic electron transport via shrink-age of intercellular space which is due to efflux of waterthrough water channels in the plasma membrane(Allakhverdiev et al., 2000a). Increase in osmoticpotential under high salt conditions causes Na+ ionsto leak into the cytosol (Papageorgiou et al., 1998) andinactivate both photosynthetic and respiratory electrontransport (Allakhverdiev et al., 1999).High salt (NaCl) uptake competes with the uptake of
other nutrient ions, especially K+, leading to K+
deficiency (Ball et al., 1987). Under such conditions ofhigh salinity and K+ deficiency, a reduction in quantumyield of oxygen evolution due to malfunctioning ofphotosystem II occurs (Ball et al., 1987).The reduction in photosynthetic rate is also due to the
reduction in stomatal conductance resulting in restrictedavailability of CO2 for carboxylation reactions (Brugno-li and Bjorkman, 1992). Stomatal closure minimizes lossof water by transpiration and this affects chloroplastlight-harvesting and energy-conversion systems thusleading to alteration in chloroplast activity (Iyengarand Reddy, 1996). The extent to which stomatal closureaffects photosynthetic capacity depends on the magni-tude of partial pressure of CO2 inside the leaf. There arealso reports of nonstomatal inhibition of photosynthesisunder salt stress. This nonstomatal inhibition is due to
increased resistance to CO2 diffusion in the liquid phasefrom the mesophyll wall to the site of CO2 reduction inthe chloroplast and reduced efficiency of RUBPCase(Iyengar and Reddy, 1996).Reduction in photosynthetic capacity is also a
consequence of inhibition of certain carbon metabolismprocesses by feedback from other salt-induced reactions(Greenway and Munns, 1980). Under reduced waterpotential, stromal levels of the substrate fructose-1,6-bisphosphate (FBP) accumulate and the FBPaseproduct fructose-6-phosphate is reduced so thatFBPase becomes rate limiting to photosynthesis (Heuer,1996).To cope with salt stress plants respond with physio-
logical and biochemical changes that aim at theretention of water despite high external osmoticumand the maintenance of photosynthetic activity andthese enable plants to become tolerant. An under-standing of the mechanisms by which salinity affectsphotosynthesis would aid the improvement of growthconditions and crop yield and would provide usefultools for future genetic engineering.
4. Conclusion
Salinity effects and problems with regard to toleranceand ecological performance are discussed briefly in thisreview. This review provides information on physiolo-gical, biochemical, and molecular bases of salt tolerance.Efforts have been made to compare the relativesensitivity of various plant species to salt, and uptakeand transport of NaCl are considered with regard tophytotoxicity and their interactions with nutrients.Present knowledge offers some ways for increasing salttolerance.In conclusion, salinity is the most serious threat to
agriculture and to the environment in many parts of theworld and key molecular factors that can be used forgenetic engineering of salt-tolerant plants include over-expression of specific transcription factors, characteriza-tion of dehydrin proteins, overproduction ofosmoprotectants, expression of water channel proteinsand ion transporters, and expression and characteriza-tion of molecular chaperones.
Acknowledgments
We are grateful to Professor P. Mohanty, VisitingProfessor, Regional Plant Resource Centre, Bhubanes-war for his valuable suggestions during the course ofstudies. The financial assistance from CSIR (Grant No.38(983)/EMR-II), New Delhi is gratefully acknowl-edged.

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349344
References
AbdElBaki, G.K., Siefritz, F., Man, H.M., Welner, H., Kaldenhoff,
R., Kaiser, W.M., 2000. Nitrate reductase in Zea mays L. under
salinity. Plant Cell Environ. 23, 15–521.
Adams, P., Thomas, J.C., Vernon, D.M., Bohnert, H.J., Jensen, R.G.,
1992b. Distinct cellular and organismic responses to salt stress.
Plant Cell Physiol. 33, 1215–1223.
Adams, W.W., Volk, M., Hoehn, A., Demming-Adams, B., 1992a.
Leaf orientation and the response of xanthophylls cycle to incident
light. Oecologia 90, 404–410.
Agastian, P., Kingsley, S.J., Vivekanandan, M., 2000. Effect of salinity
on photosynthesis and biochemical characteristics in mulberry
genotypes. Photosynthetica 38, 287–290.
Alamgir, A.N.M., Ali, M.Y., 1999. Effect of salinity on leaf pigments,
sugar and protein concentrations and chloroplast ATPAase
activity of rice (Oryza sativa L.). Bangladesh J. Bot. 28, 145–149.
Aldesuquy, H.S., 1998. Effect of seawater salinity and gibberllic acid
on abscisic acid, amino acids and water-use efficiency of wheat
plants. Agrochimica 42, 147–157.
AliDinar, H.M., Ebert, G., Ludders, P., 1999. Growth, chlorophyll
content, photosynthesis and water relations in guava (Psidium
guajava L.) under salinity and different nitrogen supply. Garten-
bauwissenschaft 64, 54–59.
Allakhverdiev, S.I., Nishiyama, Y., Suzuki, I., Tasaka, Y., Sakamoto,
A., Murata, N., 1999. Genetic engineering of the unsaturation of
fatty acids in membrane lipids alters the tolerance of Synechocystis
to salt stress. Proc. Natl. Acad. Sci. USA 96, 5862–5867.
Allakhverdiev, S.I., Nishiyama, Y., Miyairi, S., Yamamoto, H.,
Inagaki, N., Kanesaki, Y., Murata, N., 2002. Salt stress inhibits
the repair of photodamaged photosystem II by suppressing the
transcription and translation of psbA genes in Synechocystis. Plant
Physiol. 130, 1443–1453.
Allakhverdiev, S.I., Sakamoto, A., Nishiyama, Y., Inaba, M., Murata,
N., 2000a. Inactivation of photosystems I and II in response to
osmotic stress in Synechococcus, contribution of water channels.
Plant Physiol. 122, 1201–1208.
Allakhverdiev, S.I., Sakamoto, A., Nishiyama, Y., Inaba, M., Murata,
N., 2000b. Ionic and osmotic effects of NaCl-induced inactivation
of photosystems I and II in Synechococcus sp. Plant Physiol. 123,
1047–1056.
Allen, R.D., Webb, R.P., Schake, S.A., 1997. Use of transgenic plants
to study antioxidant defenses. Free Radic. Biol. Med. 23, 473–479.
Apse, M.P., Aharon, G.S., Snedden, W.A., Blumwald, E., 1999. Salt
tolerance conferred by overexpression of a vacuolar Na+/H+
antiport in Arabidopsis. Science 285, 1256–1258.
Asada, K., 1992. Ascorbate peroxidase—a hydrogen peroxide scaven-
ging enzyme in plants. Physiol. Plant. 85, 235–241.
Asada, K., Takahashi, M., 1987. Production and scavenging of active
oxygen radicals in photosynthesis. In: Kyle, D.J., Osmond, C.B.,
Arntzen, C.J. (Eds.), Photoinhibition, vol. 9. Elsivier, Amsterdam,
pp. 227–288.
Ashihara, H., Adachi, K., Otawa, M., Yasumoto, E., Fukushima, Y.,
Kato, M., Sano, H., Sasamoto, H., Baba, S., 1997. Compatible
solutes and inorganic ions in the mangrove plant Avicennia marina
and their effects on the activities of enzymes. Z. Naturforsh. 52c,
433–440.
Aubert, S., Hennion, F., Bouchereau, A., Gout, E., Blingy, R., Dome,
A.J., 1999. Subcellular compartmentation of proline in the leaves
of the subantartic Kerguelen cabbage Pringlea antiscorbutica R-Br.
In vivo C-13 NMR study. Plant Cell Environ. 22, 255–259.
Aziz, I., Khan, M.A., 2001. Effect of seawater on the growth, ion
content and water potential of Rhizophora mucronata Lam. J. Plant
Res. 114, 369–373.
Ball, M.C., Chow, W.S., Anderson, J.M., 1987. Salinity induced
potassium deficiency causes a loss of functional phtosystem II in
leaves of grey mangroves, Avicennia marina, through depletion of
the atrazine-binding polypeptide. Aust. J. Plant Physiol. 14,
351–361.
Bekki, K., Trinchant, J.C., Rigaut, J., 1987. Nitrogen fixation by
Medicago sativa nodules and bacteroids under sodium chloride
stress. Physiol. Plant. 71, 61–67.
Benavides, M.P., Marconi, P.L., Gallego, S.M., Comba, M.E.,
Tomaro, M.L., 2000. Relationship between antioxidant defence
systems and salt tolerance in Solanum tuberosum. Aust. J. Plant
Physiol. 27, 273–278.
Benhayyim, G., Faltin, Z., Gepstein, S., Camoin, L., Strosberg, A.D.,
Eshdat, Y., 1993. Isolation and characterization of salt-associated
protein in Citrus. Plant Sci. 88, 129–140.
Blumwald, E., Aharon, G.S., Apse, M.P., 2000. Sodium transport in
plant cells. Biochim. Biophys. Acta 1465, 140–151.
Bohnert, H.J., Jensen, R.G., 1996. Strategies for engineering water-
stress tolerance in plants. Trends Biotechnol. 14, 89–97.
Bohnert, H.J., Nelson, D.E., Jensen, R.G., 1995. Adaptations to
environmental stresses. Plant Cell 7, 1099–1111.
Bostock, R.M., Quatrano, R.S., 1992. Regulation of Em gene
expression in Rice: interaction between osmotic stress and ABA.
Plant Physiol. 98, 1356–1363.
Botella, M.A., Quesada, M.A., Kononowicz, A.K., Bressan, R.A.,
Pliego, F., Hasegawa, P.M., Valpuesta, V., 1994. Characterization
and in-situ localization of a salt-induced tomato peroxidase
messenger-RNA. Plant Mol. Biol. 25, 105–114.
Bowler, C., Slooten, L., Vandenbranden, S., De Rycke, R., Botterman,
J., Sybesma, C., Van Montague, M., Inze, D., 1991. Manganese
superoxide dismutase can induce cellular damage mediated by
oxygen radicals in transgenic plants. EMBO J. 10, 1723–1732.
Bray, E.A., 1997. Plant responses to water deficit. Trends Plant Sci. 2,
48–54.
Brugnoli, E., Bjorkman, O., 1992. Growth of cotton under continuous
salinity stress: influence on allocation pattern, stomatal and non-
stomatal components of photosynthesis and dissipation of excess
light energy. Planta 187, 335–347.
Bruns, S., Hecht-Buchholz, C., 1990. Light and electron-microscope
studies on the leaves of several potato cultivars after application of
salt at various developmental stages. Potato Res. 33, 33–41.
Chang, H., Siegel, B.Z., Siegel, S.M., 1984. Salinity induced changes in
isoperoxidase in taro, Colocasia esculenta. Phytochemistry 23,
233–235.
Chartzoulakis, K., Klapaki, G., 2000. Response of two green house
pepper hybrids to NaCl salinity during different growth stages. Sci.
Hortic. 86, 247–260.
Chaudhuri, K., Choudhuri, M.A., 1997. Effect of short-term NaCl
stress on water relations and gas exchange of two jute species. Biol.
Plant. 40, 373–380.
Cheeseman, J.M., 1988. Mechanism of salinity tolerance in plants.
Plant Physiol. 87, 547–550.
Chen, G., Asada, K., 1989. Ascorbate peroxidase in tea leaves:
occurrence of two isozymes and the differences in their enzymatic
and molecular properties. Plant Cell Physiol. 30, 987–998.
Chen, S., Li, J., Wang, S., Huttermann, A., Altman, A., 2001. Salt,
nutrient uptake and transport, and ABA of Populus euphratica; a
hybrid in response to increasing soil NaCl. Trees-Struct. Funct. 15,
186–194.
Cherian, S., Reddy, M.P., Pandya, J.B., 1999. Studies on salt tolerance
in Avicennia marina (Forstk.) Vierh.: effect of NaCl salinity on
growth, ion accumulation and enzyme activity. Indian J. Plant
Physiol. 4, 266–270.
Claes, B., Dekeeyser, R., Villarroel, R., Blucke, M.V., Bauw, G.,
Montagu, M.V., Caplan, A., 1990. Characterization of a rice gene
showing organ-specific expression in response to salt stress and
drought. Plant Cell 2, 19–27.

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 345
Comba, M.E., Benavides, M.P., Tomaro, M.L., 1998. Effect of salt
stress on antioxidant defence system in soybean root nodules. Aust.
J. Plant Physiol. 25, 665–671.
Conklin, P.L., Williams, E.H., Last, R.L., 1996. Environmental stress
sensitivity of an ascorbic acid-deficient Arabidopsis mutant. Proc.
Natl. Acad. Sci. USA 93, 9970–9974.
Cram, W.J., 1973. Internal factors regulating nitrate and chloride
influx in plant cells. J. Exp. Bot. 24, 328–341.
Creissen, G., Broadbent, P., Stevens, R., Wellburn, A.R., Mullineaux,
P., 1996. Manipulation of glutathione metabolism in transgenic
plants. Biochem. Soc. Trans. 24, 465–469.
Crowe, J.H., Hoekstra, F.A., Crowe, C.M., 1992. Anhydrobiosis.
Annu. Rev. Plant Physiol. 54, 579–599.
Cushman, J.C., 1993. Molecular-cloning and expression of chloroplast
NADP-malate dehydrogenase during crassulacean acid metabo-
lism induction by salt stress. Photosynth. Res. 35, 15–27.
Cushman, J.C., Meyer, G., Michalowski, C.B., Schmitt, J.M.,
Bohnert, H.J., 1989. Salt stress leads to differential expression of
two isogenes of PEP Case during CAM induction in the common
Ice plant. Plant Cell 1, 715–725.
de Bruxelles, G.L., Peacock, W.J., Dennies, E.S., Dolferus, R., 1996.
Abscisic acid induces the alcohol dehydrogenase gene in Arabi-
dopsis. Plant Physiol. 111, 381–391.
Deane-Drummond, C.E., Glauss, A.D.M., 1982. Studies of nitrate
influx into barley roots by the use of 36ClO3 as a tracer for nitrate.
I. Interactions with chloride and other ions. Can. J. Bot. 60,
2147–2153.
DeAragao, M.E.F., Jolivet, Y., Lima, M.D., deMelo, D.F., Dizen-
gremel, P., 1997. NaCl-induced changes of NAD(P) malic enzyme
activities in Eucalyptus citriodora leaves. Trees-Struct. Funct. 12,
66–72.
Delphine, S., Alvino, A., Zacchini, M., Loreto, F., 1998. Consequences
of salt stress on conductance to CO2 diffusion, Rubisco character-
istics and anatomy of spinach leaves. Aust. J. Plant Physiol. 25,
395–402.
Dhindsa, R.S., Matowe, W., 1981. Drought tolerance in two mosses:
correlated with enzymatic defense against lipid peroxidation. J.
Exp. Bot. 32, 79–91.
Dietz, K.J., Tavakoli, N., Kluge, C., Mimura, T., Sharma, S.S., Harris,
G.C., Chardonnens, A.N., Golldack, D., 2001. Significance of the
V-type ATPase for the adaptation to stressful growth conditions
and its regulation on the molecular and biochemical level. J. Exp.
Bot. 52, 1969–1980.
Dubey, R.S., Singh, A.K., 1999. Salinity induces accumulation of
soluble sugars and alters the activity of sugar metabolizing enzymes
in rice plants. Biol. Plant. 42, 233–239.
Ehret, D.L., Plant, A.L., 1999. Salt tolerance in crop plants. In:
Dhaliwal, G.S., Arora, R. (Eds.), Environmental Stress in Crop
Plants. Commonwealth Publishers, New Delhi, India, pp. 69–120
(Chapter 5).
Elshintinawy, F., Elshourbagy, M.N., 2001. Alleviation of changes in
protein metabolism in NaCl-stressed wheat seedlings by thiamine.
Biol. Plant. 44, 541–545.
Elstner, E.F., 1987. Metabolism of activated oxygen species. In:
Davies, D.D. (Ed.), The Biochemistry of Plants. vol. II, Biochem-
istry of Metabolism. Academic Press, San Diego, CA, pp. 252–315.
Ferreira, R.G., Tavora, F.J.A.F., Hernandez, F.F.F., 2001. Dry
matter partitioning and mineral composition of roots, stems and
leaves of guava grown under salt stress conditions. Pesqui.
Agropecu. Bras. 36, 79–88.
Flores, P., Botella, M.A., Martinez, V., Cedra, A., 2000. Ionic and
osmotic effects on nitrate reductase activity in tomato seedlings.
J. Plant Physiol. 156, 552–557.
Flowers, T.J., Troke, P.F., Yeo, A.R., 1977. The mechanism of salt
tolerance in halophytes. Annu. Rev. Plant Physiol. 28, 89–121.
Ford, C.W., 1984. Accumulation of low molecular solutes in water
stress tropical legumes. Phytochemistry 23, 1007–1015.
Foyer, C.H., Lelandais, M., Edwards, E.A., Mullineaux, P.M., 1991.
The role of ascorbate in plants, interactions with photosynthesis
and regulatory significance. In: Pell, E., Steffen, K. (Eds.), Active
Oxygen/Oxidative Stress and Plant Metabolism. American Society
of Plant Physiology, Rockville, MD, pp. 131–144.
Foyer, C.H., Lelandais, M., Kunert, K.J., 1994. Photooxidative stress
in plants. Physiol. Plant. 92, 696–717.
Fridovich, I., 1986. Biological effects of the superoxide radical. Arch.
Biochem. Biophys. 247, 1–11.
Gadallah, M.A.A., 1999. Effects of proline and glycinebetaine on Vicia
faba response to salt stress. Biol. Plant. 42, 249–257.
Gao, Z.F., Sagi, M., Lips, S.H., 1998. Carbohydrate metabolism in
leaves and assimilate partitioning in fruits of tomato (Lycopersicon
esculentum L.) as affected by salinity. Plant Sci. 135, 149–159.
GomezCadenas, A., Tadeo, F.R., PrimoMillo, E., Talon, M., 1998.
Involvement of abscic acid and ethylene in the responses of citrus
seedlings to salt shock. Plant Physiol. 103, 475–484.
GomezCadenas, A., Arbona, V., Jacas, J., PrimoMillo, E., Talon, M.,
2002. Abscisic acid reduces leaf abscission and increases salt
tolerance in citrus plants. J. Plant Growth Regu. 21, 234–240.
Gossett, D.R., Millhollon, E.P., Lucas, M.C., 1994. Antioxidant
response to NaCl stress in salt tolerant and salt sensitive cultivars
of cotton. Crop Sci. 34, 706–714.
Greenway, H., Munns, R., 1980. Mechanisms of salt tolerance in non-
halophytes. Annu. Rev. Plant Physiol. 31, 149–190.
Gueta-Dahan, Y., Yaniv, Z., Zilinskas, B.A., Ben-Hayyim, G., 1997.
Salt and oxidative stress: similar and specific responses and their
relation to salt tolerance in Citrus. Planta 203, 460–469.
Gulzar, S., Khan, M.A., Ungar, I.A., 2003. Salt tolerance of a coastal
salt marsh grass. Commun. Soil Sci. Plant Anal. 34, 2595–2605.
Gupta, A.S., Heinen, J.L., Holaday, A.S., Burke, J.J., Allen, R.D.,
1993a. Increased resistance to oxidative stress in transgenic plants
that overexpress chloroplastic Cu/Zn-superoxide dismutase. Proc.
Natl. Acad. Sci. USA 90, 1629–1633.
Gupta, A.S., Webb, R.P., Holaday, A.S., Allen, R.D., 1993b.
Overexpression of superoxide dismutase protects plants from
oxidative stress: induction of ascorbate peroxidase in
superoxide dismutase-overexpressing plants. Plant Physiol. 103,
1067–1073.
Gupta, S., Chattopadhyay, M.K., Chatterjee, P., Ghosh, B., SenGup-
ta, D.N., 1998. Expression of abscisic acid-responsive element-
binding protein in salt tolerant indica rice (Oryza sativa L. cv.
Pokkali). Plant Mol. Biol. 137, 629–637.
Hagemann, M., Erdmann, N., 1997. Environmental stresses. In: Rai,
A.K. (Ed.), Cyanobacterial Nitrogen Metabolism and Environ-
mental Biotechnology. Springer, Heidelberg, Narosa Publishing
House, New Delhi, India, pp. 156–221.
Hagemann, M., Murata, N., 2003. Glucosylglycerol, a compatible
solute, sustains cell division under salt stress. Plant Physiol. 131,
1628–1637.
Halliwell, B., 1982. The toxic effects of oxygen on plant tissues. In:
Oberly, L.W. (Ed.), Superoxide Dismutase, vol. I. CRC Press, Boca
Raton, FL, pp. 89–123.
Halliwell, B., 1987. Oxidative damage, lipid peroxidation, and
antioxidant protection in chloroplasts. Chem. Phys. Lipids 44,
327–340.
Halliwell, B., Gutteridge, J.M.C., 1985. Free Radicals in Biology and
Medicine. Clarendon Press, Oxford.
Halliwell, B., Gutteridge, J.M.C., 1986. Free Radicals in Biology and
Medicine. Oxford University Press, London.
Hamada, A., Shono, M., Xia, T., Ohta, M., Hayashi, Y., Tanaka, A.,
Hayakawa, T., 2001. Isolation and characterization of a Na+/H+
antiporter gene from the halophyte Atriplex gmelini. Plant Mol.
Biol. 46, 35–42.

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349346
Hanson, A.D., 1998. Salinity promotes accumulation of 3-dimethyl-
sulfoniopropionate and its precursor S-methylmethionine in
chloroplasts. Plant Physiol. 116, 165–171.
Harper, D.B., Harvey, B.M.R., 1978. Mechanisms of paraquat
tolerance in perennial ryegrass II. Role of superoxide dismutase,
catalase, and peroxidase. Plant Cell Environ. 1, 211–215.
Harrington, H.M., Alm, D.M., 1988. Interaction of heat and salt
shock in cultured tobacco cells. Plant Physiol. 88, 618–625.
Hasegawa, P.M., Bressan, R.A., Zhu, J.K., Bohnert, H.J., 2000. Plant
cellular and molecular responses to high salinity. Annu. Rev. Plant
Physiol. Plant Mol. Biol. 51, 463–499.
Hassanein, A.M., 1999. Alterations in protein and esterase patterns of
peanut in response to salinity stress. Biol. Plant. 42, 241–248.
Hayashi, H., Murata, N., 1998. Genetically engineered enhancement
of salt tolerance in higher plants. In: Sato, K,Murata,N. (Ed.),
Stress Response of Photosynthetic Organisms: Molecular Mechan-
isms and Molecular Regulation. Elsevier, Amsterdam, pp. 133–148.
Hernandez, J.A., Olmos, E., Corpas, F.J., Sevilla, F., del Rio, L.A.,
1995. Salt-induced oxidative stress in chloroplasts of pea plants.
Plant Sci. 105, 151–167.
Hernandez, J.A., Campillo, A., Jimenez, A., Alacon, J.J., Sevilla, F.,
1999. Response of antioxidant systems and leaf water relations to
NaCl stress in pea plants. New Phytol. 141, 241–251.
Hernandez, J., Jimenez, A., Mullineaux, P., Sevilla, F., 2000.
Tolerance of pea plants (Pisum sativum) to long-term salt stress is
associated with induction of antioxidant defences. Plant Cell
Environ. 23, 853–862.
Hernandez, J., Nistal, Dopico, B., Labrador, E., 2002. Cold and salt
stress regulates the expression and activity of a chickpea cytosolic
Cu/Zn superoxide dismutase. Plant Sci. 163, 507–514.
Heuer, B., 1996. Photosynthetic carbon metabolism of crops under salt
stress. In: Pesserkali, M. (Ed.), Hand Book of Photosynthesis.
Marshal Dekar, Baten Rose, USA, pp. 887–896.
Hilda, P., Graciela, R., Sergio, A., Otto, M., Ingrid, R., Hugo, P.C.,
Edith, T., Estela, M.D., Guillermina, A., 2003. Salt tolerant tomato
plants show increased levels of jasmonic acid. Plant Growth Regul.
41, 149–158.
Hogarth, P.J., 1999. The Biology of Mangroves. Oxford University
Press, New York.
Holland, D., Ben-Hayyim, G., Faltin, Z., Camoin, L., Strosberg, A.D.,
Eshdat, Y., 1993. Molecular characterization of salt-stress-asso-
ciated protein in citrus: protein and cDNA sequence homology to
mammalian glutathione peroxidase. Plant Mol. Biol. 21,
923–927.
Hong, B., Barg, R., Ho, T.H.D., 1992. Developmental and organ-
specific expression of an ABA and stress-induced protein in barley.
Plant Mol. Biol. 18, 663–674.
Hurkman, W.J., Tanaka, C.K., 1988. Polypeptide changes induced by
salt stress, water deficit and osmotic stress in barley roots, A
comparison using two-dimensional gel electrophoresis. Electro-
phoresis 9, 781–787.
Imlay, J.A., Linn, S., 1988. DNA damage and oxygen radical toxicity.
Science 240, 1302–1309.
Ingram, J., Bartels, D., 1996. The molecular basis of dehydration
tolerance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47,
377–403.
Iyengar, E.R.R., Reddy, M.P., 1996. Photosynthesis in highly salt-
tolerant plants. In: Pesserkali, M. (Ed.), Handbook of photosynth-
esis. Marshal Dekar, Baten Rose, USA, pp. 897–909.
Ishitani, M., Majumder, A.L., Bornhouser, A., Michealowski, C.B.,
Jensen, R.G., Bohnert, H.J., 1996. Coordinate transcriptional
induction of myo-inositol metabolism during environmental stress.
Plant J. 9, 537–548.
Jain, M., Mathur, G., Koul, S., Sarin, N.B., 2001. Ameliorative effects
of proline on salt stress-induced lipid peroxidation in cell lines of
groundnut (Arachis hypogaea L.). Plant Cell Rep. 20, 463–468.
Kanesaki, Y., Suzuki, I., Allakhverdiev, S.I., Mikami, K., Murata, N.,
2002. Salt stress and hyperosmotic stress regulate the expression of
different sets of genes in Synechocystis sp. PCC 6803. Biochem.
Biophys. Res. Commun. 290, 339–348.
Kao, W.Y., Tsai, H.C., Tsai, T.T., 2001. Effect of NaCl and nitrogen
availability on growth and photosynthesis of seedlings of a
mangrove species, Kandelia candel (L.) Druce. J. Plant Physiol.
158, 841–846.
Kawasaki, S., Borchert, C., Deyholos, M., Wang, H., Brazille, S.,
Kawai, K., Galbraith, D., Bohnert, H.J., 2001. Gene expression
profiles during the initial phase of salt stress in rice. Plant Cell 13,
889–905.
Keiper, F.J., Chen, D.M., DeFilippis, L.F., 1998. Respiratory,
photosynthetic and ultrastructural changes accompanying salt
adaptation in culture of Eucalyptus microcorys. J. Plant Physiol.
152, 564–573.
Kennedy, B.F., De Fillippis, L.F., 1999. Physiological and oxidative
response to NaCl of the salt tolerant Grevillea ilicifolia and the salt
sensitive Grevillea arenaria. J. Plant Physiol. 155, 746–754.
Kerepesi, I., Galiba, G., 2000. Osmotic and salt stress-induced
alteration in soluble carbohydrate content in wheat seedlings.
Crop Sci. 40, 482–487.
Kerkeb, L., Donaire, J.P., RodriguezRosales, M.P., 2001. Plasma
membrane H+-ATPase activity is involved in adaptation of tomato
calli to NaCl. Physiol. Plant. 111, 483–490.
Khan, M.A., 2001. Experimental assessment of salinity tolerance of
Ceriops tagal seedlings and saplings from the Indus delta, Pakistan.
Aquat. Bot. 70, 259–268.
Khan, M.A., Ungar, I.A., Showalter, A.M., Dewald, H.D., 1998.
NaCl-induced accumulation of glycinebetaine in four subtropical
halophytes from Pakistan. Physiol. Plant. 102, 487–492.
Khan, M.A., Ungar, I.A., Showalter, A.M., 1999. Effects of salinity on
growth, ion content, and osmotic relations in Halopyrum mocor-
onatum (L.) Stapf. J. Plant Nutr. 22, 191–204.
Khan, M.A., Ungar, I.A., Showalter, A.M., 2000a. Effects of sodium
chloride treatments on growth and ion accumulation of the
halophyte Haloxylon recurvum. Commun. Soil Sci. Plant Anal.
31, 2763–2774.
Khan, M.A., Ungar, I.A., Showalter, A.M., 2000b. The effect of
salinity on the growth, water status, and ion content of a leaf
succulent perennial halophyte Suadea fruticosa (L.) Forssk. J. Arid
Environ. 45, 73–84.
Khatkar, D., Kuhad, M.S., 2000. Short-term salinity induced changes
in two wheat cultivars at different growth stages. Biol. Plant. 43,
629–632.
Khavarinejad, R.A., Chaparzadeh, N., 1998. The effects of NaCl and
CaCl2 on photosynthesis and growth of alfalfa plants. Photo-
synthetica 35, 461–466.
Khavarinejad, R.A., Mostofi, Y., 1998. Effects of NaCl on photo-
synthetic pigments, saccharides, and chloroplast ultrastructure in
leaves of tomato cultivars. Photosynthetica 35, 151–154.
King, G.H., Turner, V.A., Hussey Jr., C.E., Wurte, Le, E.S., Lee,
S.M., 1988. Isolation and characterization of a tomato cDNA
clone which codes for a salt-induced protein. Plant Mol. Biol. 10,
401–412.
Kirst, G.O., 1989. Salinity tolerance of eukaryotic marine algae. Annu.
Rev. Plant Physiol. Plant Mol. Biol. 40, 21–53.
Klages, K., Boldingh, H., Smith, G.S., 1999. Accumulation of myo-
inositol in Actinidia seedlings subjected to salt stress. Ann. Bot. 84,
521–527.
Knight, H., Trewavas, A.J., Knight, M.R., 1997. Calcium signaling in
Arabidopsis thaliana responding to drought and salinity. Plant J.
12, 1067–1078.
Koyro, H.W., 2000. Effect of high NaCl-salinity on plant growth, leaf
morphology, and ion composition in leaf tissues of Beta vulgaris
ssp maritima. J. Appl. Bot. Angew. Bot. 74, 67–73.

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 347
Kurban, H., Saneoka, H., Nehira, K., Adilla, R., Premachandra, G.S.,
Fujita, K., 1999. Effect of salinity on growth, photosynthesis and
mineral composition in leguminous plant Alhagi pseudoalhagi
(Bieb.). Soil Sci. Plant Nutr. 45, 851–862.
Lauchli, A., Schubert, S., 1989. The role of calcium in the regulation of
membrane and cellular growth processes under salt stress. NATO
ASI Ser. G19, pp. 131–137
Lechno, S., Zamski, E., TelOr, E., 1997. Salt stress-induced responses
in cucumber plants. J. Plant Physiol. 150, 206–211.
Lee, T.M., Liu, C.H., 1999. Correlation of decreases calcium contents
with proline accumulation in the marine green macroalga Ulva
fasciata exposed to elevated NaCI contents in seawater. J. Exp.
Bot. 50, 1855–1862.
Lee, D.H., Kim, Y.S., Lee, C.B., 2001. The inductive responses of the
antioxidant enzymes by salt stress in the rice (Oryza sativa L.). J.
Plant Physiol. 158, 737–745.
Leung, J., Giraudat, J., 1998. Abscisic acid signal transduction. Annu.
Rev. Plant Physiol. Plant Mol. Biol. 49, 199–222.
Levitt, J., 1980. Responses of Plant to Environmental Stress Chilling,
Freezing, and High Temperature Stresses, second ed. Academic
Press, New York.
Liang, Y.C., 1999. Effects of silicon on enzyme activity and sodium,
potassium and calcium concentration in barley under salt stress.
Plant. Soil 209, 217–224.
Lin, C.C., Hsu, Y.T., Kao, C.H., 2002. The effect of NaCl on proline
accumulation in rice leaves. Plant Growth Regul. 36, 275–285.
Lippert, K., Galinski, E.A., 1992. Enzyme stabilization by ecotine-type
compatible solutes: protection against heating, freezing and drying.
Appl. Microbiol. Biotechnol. 37, 61–65.
Liu, J., Zhu, J.K., 1997. An Arabidopsis mutant that requires increased
calcium for potassium nutrition and salt tolerance. Proc. Natl.
Acad. Sci. USA 94, 14960–14964.
Longstreth, D.J., Nobel, P.S., 1979. Salinity effects on leaf anatomy.
Plant Physiol. 63, 700–703.
Lopez, F., Vansuyt, G., Fourcroy, P., Casse-Delbart, F., 1994.
Accumulation of a 22-kDa protein in the leaves of Raphanus
sativus in response to salt stress or water deficit. Physiol. Plant. 91,
605–614.
Lu, C.M., Vonshak, A., 1999. Characterization of PS II photochem-
istry in salt-adapted cells of cyanobacterium Spirulina platensis.
New Phytol. 141, 231–239.
Lu, C.M., Qiu, N.W., Lu, Q.T., Wang, B.S., Kuang, T.Y., 2002. Does
salt stress lead to increased susceptibility of photosystem II to
photoinhibition and changes in photosynthetic pigment composi-
tion in halophyte Suaeda salsa grown outdoors?. Plant Sci. 163,
1063–1068.
Mansour, M.M.F., 2000. Nitrogen containing compounds and
adaptation of plants to salinity stress. Biol. Plant. 43, 491–500.
Marcelis, L.F.M., VanHooijdonk, J., 1999. Effect of salinity on
growth, water use and nutrient use in radish (Raphanus sativus L.).
Plant Soil 215, 57–64.
Matsumura, T., Kanechi, M., Inagaki, N., Maekawa, S., 1998. The
effects of salt stress on ion uptake, accumulation of compatible
solutes, and leaf osmotic potential in safflower, Chrysanthemum
paludosum and sea aster. J. Jpn. Soc. Hortic. Sci. 67, 426–431.
Mattioni, C., Lacerenze, N.G., Troccoli, A., DeLeonardis, A.M.,
DiFonzo, N., 1997. Water and salt stress-induced alterations in
proline metabolism of Triticum durum seedlings. Physiol. Plant.
101, 787–792.
Meloni, D.A., Oliva, M.A., Ruiz, H.A., Martinez, C.A., 2001.
Contribution of proline and inorganic solutes to osmotic adjust-
ment in cotton under salt stress. J. Plant Nutr. 24, 599–612.
Mendoza, I., Rubio, F., Rodriguez-Navarro, A., Prado, J.M., 1994.
The protein phosphatase calcineurin is essential for NaCl tolerance
of Saccharomyces cerevisiae. J. Biol. Chem. 269, 8792–8796.
Meneguzzo, S., Navarilzzo, I., 1999. Antioxidative responses of shoots
and roots of wheat to increasing NaCl concentrations. J. Plant
Physiol. 155, 274–280.
Mishra, S., Das, A.B., 2003. Effect of NaCl on leaf salt secretion and
antioxidative enzyme level in roots of a mangrove, Aegiceras
corniculatum. Indian J. Exp. Biol. 41, 160.
Mishra, S.K., Subrahmanyam, D., Singhal, G.S., 1991. Interrelation-
ship between salt and light stress on primary processes of
photosynthesis. J. Plant Physiol. 138, 92–96.
Mitsuya, S., Takeoka, Y., Miyake, H., 2000. Effects of sodium
chloride on foliar ultrastructure of sweet potato (Ipomoea batatas
Lam.) plantlets grown under light and dark conditions in vitro. J.
Plant Physiol. 157, 661–667.
Mittova, V., Tal, M., Volokita, M., Guy, M., 2002. Salt stress induces
up-regulation of an efficient chloroplast antioxidant system in the
salt-tolerant wild tomato species Lycopersicon pennellii but not in
the cultivated species. Physiol. Plant. 115, 393–400.
Mittova, V., Tal, M., Volokita, M., Guy, M., 2003. Up-regulation of
the leaf mitochondrial and peroxisomal antioxidative systems in
response to salt-induced oxidative stress in the wild salt-tolerant
tomato species Lycopersicon pennellii. Plant Cell Environ. 26,
845–856.
Mohammad, M., Shibli, R., Ajouni, M., Nimri, L., 1998. Tomato root
and shoot responses to salt stress under different levels of
phosphorus nutrition. J. Plant Nutr. 21, 1667–1680.
Morales, M.A., Sancheo-Blanco, M.J., Olmos, E., Torrechlas, A.,
Alarcon, J.J., 1998. Changes in the growth, leaf water relations and
all ultrastructure in Argyranthemum coronopifolium plants under
saline conditions. J. Plant Physiol. 153, 174–180.
Mundy, J., Yamaguchi-Shinozaki, K., Chua, N.H., 1990. Nuclear
proteins bind conserved elements in the abscisic acid responsive
promoter of a rice RAB gene. Proc. Natl. Acad. Sci. USA 87,
1406–1410.
Munns, R., Termatt, A., 1986. Whole plant responses to salinity. Aust.
J. Plant Physiol. 13, 143–160.
Munoz, G.E., Marin, K., Gonzalez, C., 1997. Polypeptide profile in
Prosopsis seedlings growing in saline conditions. Phyton—Int. J.
Exp. Bot. 61, 17–24.
Muthukumarasamy, M., Gupta, S.D., Pannerselvam, R., 2000.
Enhancement of peroxidase, polyphenol oxidase and superoxide
dismutage activities by triadimefon in NaCl stressed Raphanus
sativus L. Biol. Plant. 43, 317–320.
Nilu, X., Bressan, R.A., Hasegawa, P.M., Prado, J.M., 1995. Ion
homeostasis in NaCl stress environments. Plant Physiol. 109,
735–742.
Noaman, M.M., Dvorak, J., Dong, J.M., 2002. Genes inducing Salt
tolerance in wheat, Lophopyrum elongatum and amphiploid and
their responses to ABA under salt stress. Prospects for Saline
Agriculture (Series: Tasks for Vegetation Science) 37, 139–144.
Noctor, G., Foyer, C.H., 1998. Ascorbate and glutathione: keeping
active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol.
Biol. 49, 249–279.
Nuccio, M.L., Rhodes, D., McNeil, S.D., Hason, A.D., 1999.
Metabolic engineering of plants for osmotic stress resistance. Curr.
Opin. Plant Biol. 2, 91–99.
Orr, W.C., Sohal, R.S., 1992. The effects of catalase gene over-
expression on life span and resistance to oxidative stress in
transgenic Drosophila melanogaster. Arch. Biochem. Biophys.
297, 35–41.
Orthen, B., Popp, M., Smirnoff, N., 1994. Hydroxyl radical scavenging
properties of cyclitols. Proc. R. Soc. Edinburg Sect. B 102,
269–272.
Otoch, M.D.L., Sobreira, A.C.M., deAragao, M.E.F., Orellano, E.G.,
Lima, M.D.S., deMelo, D.F., 2001. Salt modulation of vacuolar
H+-ATPase and H+-pyrophosphatase activities in Vigna unguicu-
lata. J. Plant Physiol. 158, 545–551.

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349348
Papageorgiou, G.C., Alygizaki-Zobra, A., Ladas, N., Murata, N.,
1998. A method to probe the cytoplasmic osmolarity and osmotic
water and solute fluxes across the cell membrane of Cyanobacteria
with Chl a florescence: experiments with Synechococcus sp. PCC
7942. Physiol. Plant. 103, 215–224.
Parida, A.K., Das, A.B., 2004. Effects of NaCl stress on nitrogen and
phosphorous metabolism in a true mangrove Bruguiera parviflora
grown under hydroponic culture. J. Plant Physiol. 161, 921–928.
Parida, A., Das, A.B., Das, P., 2002. NaCl stress causes changes in
photosynthetic pigments, proteins and other metabolic components
in the leaves of a true mangrove, Bruguiera parviflora, in
hydroponic cultures. J. Plant Biol. 45, 28–36.
Parida, A.K., Das, A.B., Mittra, B., 2003. Effects of NaCl stress on the
structure, pigment complex compsition and photosynthetic activity
of mangrove Bruguiera parviflora chloroplasts. Photosynthetica 41,
191–200.
Parida, A.K., Das, A.B., Mittra, B., 2004a. Effects of salt on growth,
ion accumulation photosynthesis and leaf anatomy of the
mangrove, Bruguiera parviflora. Trees-Struct. Funct. 18, 167–174.
Parida, A.K., Das, A.B., Mittra, B., Mohanty, P., 2004b. Salt-stress
induced alterations in protein profile and protease activity in the
mangrove, Bruguiera parviflora. Z. Naturforsch. 59c, 408–414.
Parida, A.K., Das, A.B., Mohanty, P., 2004c. Defense potentials to
NaCl in a mangrove, Bruguiera parviflora: differential changes of
isoforms of some antioxidative enzymes.. J. Plant Physiol. 161,
531–542.
Pfluger, K., Baumann, S., Gottschalk, G., Lin, W., Santos, H., Muller,
V., 2003. Lysine-2,3-aminomutase and beta-lysine acetyltransferase
genes of methanogenic archaea are salt induced and are essential
for the biosynthesis of N-epsilon-acetyl-beta-lysine and growth at
high salinity. Appl. Environ. Microbiol. 69, 6047–6055.
Pilon-Smits, E.A.H., Ebskamp, M.J.M., Paul, M.J., Jeuken, M.J.W.,
Weisbeek, P.J., Smeekens, S.C.M., 1995. Improved performance of
transgenic fructan-accumulating tobacco under drought stress.
Plant Physiol. 107, 125–130.
Plant, A.L., Cohen, A., Moses, M.S., Bray, E.A., 1991. Nucleotide
sequence and spatial expression pattern of a drought and ABA-
induced gene of tomato. Plant Physiol. 97, 900–906.
Popova, L.P., Stoinova, Z.G., Maslenkova, L.T., 1995. Involvement of
abscisic acid in photosynthetic process in Hordeum vulgare L
during salinity stress. J. Plant Growth Regul. 14, 211–218.
Popp, M., Larther, F., Weigel, P., 1985. Osmotic adaptation in
Australian mangroves. Vegetatio 61, 247–254.
Popova, O.V., Ismailov, S.F., Popova, T.N., Dietz, K.J., Golldack, D.,
2002. Salt-induced expression of NADP-dependent isocitrate
dehydrogenase and ferredoxin-dependent glutamate synthase in
Mesembryanthemum crystallinum. Planta 215, 906–913.
Quintero, F.J., Garciadeblas, B., Rodriguez-Navarro, A., 1996. The
SAL1 gene of Arabidopsis, encoding an enzyme with 30(20), 50-
Biosphere nucleotidase and inositol polyphosphate 1-phosphate
activities, increases salt tolerance in yeast. Plant Cell 8, 529–537.
Rajasekaran, L.R., Aspinall, D., Jones, G.P., Paleg, L.G., 2001. Stress
metabolism. IX. Effect of salt stress on trigonelline accumulation in
tomato. Can. J. Plant Sci. 81, 487–498.
Rajesh, A., Arumugam, R., Venkatesalu, V., 1998. Growth and
photosynthetic characterics of Ceriops roxburghiana under NaCl
stress. Photosynthetica 35, 285–287.
Ramagopal, S., 1987. Salinity stress induced tissue-specific proteins in
barley seedlings. Plant Physiol. 84, 324–331.
Raychaudhuri, A., Majumder, A.L., 1996. Salinity-induced enhance-
ment of L-myo-inositol 1-phosphate synthase in rice (Oryza sativa
L). Plant Cell Environ. 19, 1437–1442.
Reddy, M.P., Sanish, S., Iyengar, E.R.R., 1992. Photosynthetic studies
and compartmentation of ions in different tissues of Salicornia
brachiata Roxb. under saline conditions. Photosynthetica 26,
173–179.
Reviron, M.P., Vartanian, N., Sallantin, M., Huet, J.C., Pernollet,
J.C., de Vienne, D., 1992. Characerization of a novel protein
induced by progessive or rapid drought and salinity in Brassica
napus leaves. Plant Physiol. 100, 1486–1493.
Rhodes, D., Hanson, A.D., 1993. Quaternary ammonium and tertiary
sulphonium compounds in higher plants. Annu. Rev. Plant
Physiol. Plant Mol. Biol. 44, 357–384.
RiosGonzalez, K., Erdei, L., Lips, S.H., 2002. The activity of
antioxidant enzymes in maize and sunflower seedlings as affected
by salinity and different nitrogen sources. Plant Sci. 162, 923–930.
RodriguezRosales, M.P., Kerkeb, L., Bueno, P., Donaire, J.P., 1999.
Changes induced by NaCl in lipid content and composition,
lipoxygenase, plasma membrane H+ATPase and antioxidant
enzyme activities of tomato (Lycopersicon esculantum, Mill) calli.
Plant Sci. 143, 143–150.
Romeroaranda, R., Soria, T., Cuartero, J., 2001. Tomato plant—
water uptake and plant–water relationships under saline growth
conditions. Plant Sci. 160, 265–272.
Roxas, V.P., Lodhi, S.A., Garrett, D.K., Mahan, J.R., Allen, R.D.,
2000. Stress tolerance in transgenic tobacco seedlings that over-
express glutathione S-transferase/glutathione peroxidase. Plant
Cell Physiol. 41, 1229–1234.
Sadaka, A., Himmelhoch, S., Zamir, A., 1991. A 150 kilodalton cell
surface protein is induced by salt in the haloterant green alga
Dunaliella salina. Plant Physiol. 95, 822–831.
Saneoka, H., Shiota, K., Kurban, H., Chaudhary, M.I., Premachan-
dra, G.S., Fujita, K., 1999. Effect of salinity on growth and solute
accumulation in two wheat lines differing in salt tolerance. Soil Sci.
Plant Nutr. 45, 873–880.
SantaCruz, A., PereAlfocea, F., Caro, M., Acosta, M., 1998.
Polyamines as short-term salt tolerance traits in tomato. Plant
Sci. 138, 9–16.
Sasaki, Y., Asamizu, E., Shibata, D., Nakamura, Y., Kaneko, T.,
Awai, K., Masuda, T., Shimada, H., Takamiya, K., Tabata, S.,
Ohta, H., 2000. Genome-wide expression-monitoring of jasmonate-
responsive genes of Arabidopsis using cDNA arrays. Biochem. Soc.
Trans. 28, 863–864.
Sehmer, L., Alaoui-Sosse, B., Dizengremel, P., 1995. Effect of salt
stress on growth and on the detoxyfying pathway of pedunculate
oak seedlings (Quercus robur L.). J. Plant Physiol. 147, 144–151.
Serraz, R., VasquezDiaz, H., Drevon, J.J., 1998. Effects of salt stress
on nitrogen fixation, oxygen diffusion and ion distribution in
soybean, common bean, and alfalfa. J. Plant Nutr. 21, 475–488.
Serraz, R., VasquezDiaz, H., Hernandez, G., Drevon, J.J., 2001.
Genotypic difference in the short-term response of nitrogenase
activity (C2H2 reduction) to salinity and oxygen in the common
bean. Agronomie 21, 645–651.
Shi, H.Z., Zhu, J.K., 2002. Regulation of expression of the vacuolar
Na+/H+ antiporter gene AtNHX1 by salt stress and abscisic acid.
Plant Mol. Biol. 50, 543–550.
Shikanai, T., Takeda, T., Yamauchi, H., Sano, S., Tomizawa, K.I.,
1998. Inhibition of asorbate peroxidase under oxidative stress in
tobacco having bacterial catalase in chloroplasts. FEBS Lett. 428,
47–51.
Shinozaki, K., Yamaguchi-Shinozaki, K., Mizoguchi, T., Uraro, T.,
Katagiri, T., Nakashima, K., Abe, H., Ichimura, K., Liu, Q.A.,
Nanjyo, T., Uno, Y., Luchi, S., Srki, M., Lto, T., Hirayama, T.,
Mikami, K., 1998. Molecular responses to water stress in
Arabidopsis thaliana. J. Plant Res. 111, 345–351.
Singh, N.K., Handa, A.K., Hasegawa, P.M., Bressan, R.A., 1985.
Proteins associated with adaptation of cultured tobacco cells to
NaCl. Plant Physiol. 79, 126–137.
Singh, N.K., Bracker, C.A., Hasegawa, P.M., Handa, A.K., Buckel,
S., Hermodson, M.A., Pfankock, E., Regnier, F.E., Bressan, R.A.,
1987. Characterization of osmotin. Athumatin-like protein

ARTICLE IN PRESSA.K. Parida, A.B. Das / Ecotoxicology and Environmental Safety 60 (2005) 324–349 349
associated with osmotic adaptation in plant cells. Plant Physiol. 85
(79), 126–137.
Singh, S.K., Sharma, H.C., Goswami, A.M., Datta, S.P., Singh, S.P.,
2000. In vitro growth and leaf composition of grapevine cultivars
as affected by sodium chloride. Biol. Plant. 43, 283–286.
Singh, S.C., Sinha, R.P., Hader, D.P., 2002. Role of lipids and fatty
acids in stress tolerance in cyanobacteria. Acta Protozool. 41,
297–308.
Smirnoff, N., 1998. Plant resistance to environmental stress. Curr.
Opin. Biotechnol. 9, 214–219.
Smirnoff, N., Cumbes, Q.J., 1989. Hydroxyl radical scavenging
activity of compatible solutes. Phyotochemistry 28, 1057–1060.
Smith, F.A., 1973. The internal control of nitrate uptake into excised
barley roots with differing salt contents. New Phytol. 72, 769–782.
Soussi, M., Lluch, C., Ocana, A., 1998. Effects of salt stress on growth,
photosynthesis and nitrogen fixation in chick-pea (Cicer arietinum
L.). J. Exp. Bot. 49, 1329–1337.
Soussi, M., Lluch, C., Ocana, A., 1999. Comparative study of nitrogen
fixation and carbon metabolism in two chick-pea (Ciser arietinum
L.) cultivars under salt stress. J. Exp. Bot. 50, 1701–1708.
Spychalla, J.P., Desborough, S.L., 1990. Superoxide dismutase,
catalase, and alpha-tocopherol content of stored potato tubers.
Plant Physiol. 94, 1214–1218.
Sreenivasulu, N., Grimm, B., Wobus, U., Weschke, W., 2000.
Differential response of antioxidant compounds to salinity stress
in salt-tolerant and salt-sensitive seedlings of fox-tail millet (Setaria
italica). Physiol. Plant. 109, 435–442.
Steiger, H.M., Beck, E., Beck, R., 1977. Oxygen concentration in
isolated chloroplasts during photosynthesis. Plant Physiol. 60,
903–906.
Sugimoto, M., Sakamoto, W., 1997. Putative phospholipid hydroper-
oxide glutathione peroxidase gene from Arabidopsis thaliana
induced by oxidative stress. Genes Genet. Syst. 72, 311–316.
Sulpice, R., Tsukaya, H., Nonaka, H., Mustardy, L., Chen, T.H.H.,
Murata, N., 2003. Enhanced formation of flowers in salt-stressed
Arabidopsis after genetic engineering of the synthesis of glycine
betaine. Plant J. 36, 165–176.
Sun, W.Q., Li, X.P., Ong, B.L., 1999. Preferential accumulation of D-
pinitol in Acrostichum aureum gametophytes in response to salt
stress. Physiol. Plant. 105, 51–57.
Takemura, T., Hanagata, N., Sugihara, K., Baba, S., Karube, I.,
Dubinsky, Z., 2000. Physiological and biochemical responses to
salt stress in the mangrove, Bruguiera gymnorrhiza. Aquat. Bot. 68,
15–28.
Takemura, T., Hanagata, N., Dubinsky, Z., Karube, I., 2002.
Molecular characterization and response to salt stress of mRNAs
encoding cytosolic Cu/Zn superoxide dismutase and catalase from
Bruguiera gymnorrhiza. Trees-Struct. Funct. 16, 94–99.
Tamas, L., Huttova, J., Mistrik, I., 2001. Impact of aluminium, NaCl
and growth retardant tetcyclacis on growth and protein composi-
tion of maize roots. Biologia 56, 441–448.
Thomas, J.C., McElwain, E.F., Bohnert, H.J., 1992. Convergent
induction of osmotic stress—responses. Plant Physiol. 100,
416–423.
Tiburico, A.F., Campos, J.L., Figueras, X., Besford, R.T., 1993.
Recent advances in the understanding of polyamine functions
during plant development. Plant Growth Regul. 12, 331–340.
Tiwari, B.S., Bose, A., Ghosh, B., 1997. Photosynthesis in rice under
salt stress. Photosynthetica 34, 303–306.
Torres-Schumann, S., Godoy, J.A., Pintor-Toro, J.A., 1992. A
probable lipid transfer protein gene is induced by NaCl stress in
tomato plants. Plant Mol. Biol. 18, 749–757.
Trossat, C., Rathinasabapathi, B., Weretilnyk, E.A., Shen, T.L.,
Huang, Z.H., Gage, D.A., Hanson, A.D., 1998. Salinity promotes
accumulation of 3-dimethylsulfoniopropionate and its precursor
S-methylmethionine in chloroplasts. Plant Physiol. 116, 165–171.
Unni, S., Rao, K.K., 2001. Protein and lipopolysaccharide profiles of a
salt-sensitive Rhizobium sp and its exopolysaccharide-deficient
mutant. Soil Biol. Biochem. 33, 111–115.
Vaidyanathan, R., Kuruvilla, S., Thomas, G., 1999. Characterization
and expression pattern of an abscisic acid and osmotic stress
responsive gene from rice. Plant Sci. 140, 21–30.
Van Camp, W., Capiau, K., Van Montagu, M., Inze, D., Slooten, L.,
1996. Enhancement of oxidative stress tolerance in transgenic
tobacco plants overproducing Fe-superoxide dismutase in chlor-
oplasts. Plant Physiol. 112, 1703–1714.
Vernon, D.M., Bohnert, H.J., 1992. A novel methyltransferase induced
by osmotic stress in the facultative halophyte Mesembryanthemum
crystallinum. EMBO J. 11, 2077–2085.
Vernon, D.M., Taraczynski, M.C., Jensen, R.G., Bohnert, H.J., 1993.
Cyclitol production in transgenic tobacco. Plant J. 4, 199–205.
Walbot, V., Cullis, C.A., 1985. Rapid genomic change in higher plants.
Annu. Rev. Plant Physiol. Plant Mol. Biol. 36, 367–396.
Wang, Y., Nil, N., 2000. Changes in chlorophyll, ribulose biphosphate
carboxylase–oxygenase, glycine betaine content, photosynthesis
and transpiration in Amaranthus tricolor leaves during salt stress.
J. Hortic. Sci. Biotechnol. 75, 623–627.
Wang, B.S., Luttge, U., Ratajczak, R., 2001. Effects of salt treatment
and osmotic stress on V-ATPase and V-PPase in leaves of the
halophyte Suaeda salsa. J. Exp. Bot. 52, 2355–2365.
Willkens, H., Chamnogopol, S., Davey, M., Schraudner, M.,
Langebartels, C., 1997. Catalase is a sink for H2O2 and is
indispensable for stress defense in C3 plants. EMBO J. 16,
4806–4816.
Winicov, I., 1998. New molecular approaches to improving salt
tolerance in crop plants. Ann. Bot. 82, 703–710.
Wise, R.R., Naylor, A.W., 1987. Chilling-enhanced photooxidation:
evidence for the role of singlet oxygen and endogenous antiox-
idants. Plant Physiol. 83, 278–282.
Wu, S., Ding, L., Zhu, J., 1996. SOS1, a genetic locus essential for salt
tolerance and potassium acquisition. Plant Cell 8, 617–627.
Wu, J.L., Seliskar, D.M., Gallagher, J.L., 1998. Stress tolerance in the
marsh plane Spartina patens: impact of NaCl on growth and root
plasma membrane lipid composition. Physiol. Plant. 102, 307–317.
Xu, X.Y., Abo, M., Okubo, A., Yamazaki, S., 2001. Salt-stress-
responsive membrane proteins in Rhodobacter sphaeroides f. sp
denitrificans IL106. J. Biosci. Bioeng. 91, 228–230.
Yancey, P., Clark, M.E., Had, S.C., Bowlus, R.D., Somero, G.N.,
1982. Living with water stress: evolution of osmolyte system.
Science 217, 1214–1222.
Yen, H.E., Zhang, D.Z., Lin, J.H., Edwards, G.E., Ku, M.S.B., 1997.
Salt-induced changes in protein composition in light-grown callus
of Mesembryanthemurm crystallinum. Physiol. Plant. 101, 526–532.
Zhifang, G., Loescher, W.H., 2003. Expression of a celery mannose 6-
phosphate reductase in Arabidopsis thaliana enhances salt tolerance
and induces biosynthesis of both mannitol and a glucosyl-mannitol
dimmer. Plant Cell Environ. 26, 275–283.
Zhu, J.K., 2002. Salt and drought stress signal transduction in plants.
Annu. Rev. Plant Biol. 53, 247–273.
Zhu, J.K., 2003. Regulation of ion homeostasis under salt stress. Curr.
Opin. Plant Biol. 6, 441–445.
Zhu, J., Meinzer, F.C., 1999. Efficiency of C-4 photosynthesis in
Atriplex lentiformis under salinity stress. Aust. J. Plant Physiol. 26,
79–86.
Zhu, J.K., Shi, J., Singh, U., Wyatt, S.E., Bressan, R.A., Hasegawa,
P.M., Capita, N.C., 1993. Enrichment of vitronectin and fibro-
nectin like proteins in NaCl-adapted plant cells and evidence for
their involvement in plasma membrane-cell wall adhesion. Plant J.
3, 637–646.





























