Embed Size (px)
Citation preview

Aquatic Botany, 39 ( 1991 ) 57-73 57 Elsevier Science Publ ishers B.V., Ams te rdam
Root adaptation to soil waterlogging
W. Armstrong 1, S.H.F.W. Justin 1, P.M. Becket t 2 and S. Ly the ~ ~ Department of Applied Biology and 2Department of Applied Mathematics, University of Hull,
Hull, HU6 7RX (Gt. Britain)
(Accepted for publication 4 July 1990 )
ABSTRACT
Armstrong, W., Justin, S.H.F.W., Beckett, P.M. and Lythe, S., 1991. Root adaptation to soil waterlog- ging. Aquat. Bot., 39: 57-73.
The structural diversity of roots is emphasised and adaptation to soil waterlogging is reviewed. Results pertinent to the mechanism of aerenchyma development are presented, and a modelling ap- proach is used to explore the implications for aeration and rhizosphere oxidation of the structural and physiological adaptations of adventitious roots and laterals.
I N T R O D U C T I O N
Oxygen (O2) deficiency and phytotoxin accumulation are major character- istics of waterlogged soil because of imposed liquid-phase diffusive aeration. The low diffusion coefficient for 02 in water ensures that even relatively low levels ofO2 demand are sufficient to deplete 02 from all but the surface layers of soil (Table 1 ); anaerobic microbes create the phytotoxins, examples of which are MnlI, Fell and H2S. The characteristics of waterlogged soils have
TABLEI
Surface oxygenation in saturated soils. Predictions of the depth (cm) to which 02 will diffuse in waterlogged soils for different levels of soil 02 demand and 02 diffusivity (a function of soil type). (After Armstrong, 1979)
Soil O: consumption (g cm-3 s - l )
02 diffusivity in the wet soil ( 10 -6 cm -2 s - t )
0.56 3.54 10
5.27× 10 -s 0.013 0.034 0.057 5.27 × 10 -9 0.042 0.107 0.180 5.27)< 10 - l ° 0.135 0.339 0.570 5.27× 10 - t t 0.426 1.073 1.803 5.27× 10 -2 1.349 3.393 5.702
0304 -3770 /91 /$03 .50 © 1991 - - Elsevier Science Publ ishers B.V.

58 W. ARMSTRONG ET AL
RICE
; ' , - , i E i
2 f / 9 8 0 m m at 25h
/ {o} 8.25 mm at 25h
cn
c t (ol g clo.38} ~ 1 . 9 9 m m
[Air] [Air] (o} f r
/ ~ ( 963 )
~ l ~ ~ ~ 0.87ram ot 25h e - - - l.25m ot25h ~ L.45mm at 25h
lo; [Air] (a59)
[N 2] (all treatments) 0
J i i I i i J J I ~ i i , 1
0 5 10 15 Time (hours]
Fig. 1. The effects of (a) 2h, (b) 3h, (c) and (d) 4h, (e) 5h, ( f ) 6 h a n d (g) 8 h anoxia on the elongation of intact rice roots bathed in a de-oxygenated medium devoid of a carbohydrate source. Roots which were anoxic for > 4 h failed to regrow on re-aeration and shrank before or after re-aeration. Figures in parentheses indicate the approximate 02 concentration within the root apex (%) calculated from radial 02 loss to a cylindrical platinum electrode. Figures in square brackets indicate the 02 concentration around the shoot system. (After Webb and Arm- strong, 1983).
been extensively reviewed in recent years (Gambrell and Patrick, 1978; Armstrong, 1982; Ponnamperuma, 1984).
The consequences for the plant of soil 02 deficiency and phytotoxin accu-

ROOT ADAPTATION TO SOIL WATERLOGG1NG 5 9
mulation are far reaching: not only is 02 an essential requirement for root growth, but total internal anoxia in roots readily kills the tips, either directly (Fig. 1 ) or through self-induced post-anoxic injury involving free 02 radicals (Hendry and Brocklebank, 1985; Crawford et al., 1987 ). 02 is essential to the long-term survival of non-dormant root tips and poorly aerated roots will be less tolerant of phytotoxin accumulation.
In this paper, we have chosen to consider the ways in which the growth and development of roots affect their aeration and thereby influence the plant's ability to survive and compete in wetland conditions. The topic is not new and, regarding general principles, we have made extensive reference to earlier studies. We have included, however, some recent and as yet unpublished ob- servations, and have also tried to stress the importance of something which is often overlooked: the enormous diversity of structure and the variations in 02 demand in roots not only at generic or species level, but even within indi- vidual roots (see also Luxmoore et al., 1970). Such diversity must influence performance and survival in wetland conditions, but it is an area for research which is largely overlooked.
STRATAGEMS OF SURVIVAL
Although plants often succumb to soil waterlogging, at least two successful developmental rooting stratagems for survival can be identified and both in- volve anoxia avoidance. They are (1) superficial rooting and (2) enhanced development of internal gas space. Internal gas spaces provide a conduit for the transport of 02 from the shoot system and, provided that there is a gas continuum, even a small fractional volume of gas space will provide at least some transport in submerged roots (Greenwood, 1967; Armstrong et al., 1983). In roots, the transport is chiefly diffusive (see also Armstrong, 1979; Higuchi, 1982; Kon~alovfi et al., 1988; Beckett et al., 1988); convective gas flows of the water lily and Phragmites type (Dacey, 1981; Grosse and Mevi- Schutz, 1987; Armstrong and Armstrong, 1991; Grosse et al., 1991 ) should not operate to any significant degree in roots. It should also be noted that wherever there is internal 02 transport there will be a potential for radial O2 loss from root to soil, and this constitutes an additional factor in survival: the oxidative removal of soil-borne phytotoxins (Armstrong, 1982 - review; Wheeler et al., 1985; Marschner, 1986 ).
Both avoidance stratagems are recognisable in the data shown in Fig. 2. Ninety-one plant species, comprising non-wetland, facultatively wetland (in- termediate) and strictly wetland species, are represented, and their rooting depths in flooded soil are plotted in relation to overall fractional root porosity.
In general, the superficial rooters exhibit low fractional root porosities (Fig. 3a). They survive presumably by exploiting chiefly the aerobic surface horizons (cf. Table 1 ), although in many cases aeration will be supplemented

60 w . A R M S T R O N G E T AL.
O
350
300
250
200
150
100
50 t , ,
o o'.1 o.1 0'.2 FRP [flooded )
45 I
36trz ~o St
ta) A
/ " • A
/ / ,',
,/X
I I i , | 0 3 0.4 0.5
A
A
Fig. 2. The relationship for individual species between fractional root porosity (FRP) and root penetration in flooded soil. ©, non-wetland; T, intermediate; A, wetland types; t , species which died or where root growth was so affected that insufficient tissue was available for the determi- nation of root porosity in flooded soil. For these species, symbol positions indicate porosities in drained soil (Redrawn from Justin and Armstrong, 1987 ).
to some degree by internal transport. Usually, the roots which develop, or perhaps the only ones to survive, are very thin and this accords with the pre- sumption of a dependence on the dissolved 02 in the aerobic layers. A high surface area:volume ratio is just what is required for successful exploitation of the scarce 02 resource. It should be noted, however, that confinement of
Fig. 3. Transverse sections of roots: (a) Luzula campestris (L.) DC, hexagonal packing, low porosity from triquetrous spaces; (b) maize, basal region with non-porous outer cortex of cells in hexagonal packing, inner and middle cortex showing lysigenous aerenchyma developed from cells in cubic packing; (c) maize, apical region of long root, cortical cells in cubic packing only; (d) Cirsium eriophorum (L.) Scop., dicotyledonous cortex with cubic packing and no second- ary growth; (e) Elymus pycnanthus (Godr.) Melderis, hexagonal, radial packing, some lysigen- ous aerenchyma; ( f ) Juncus inflexus L., inner porous cortex with radial hexagonal packing, middle cortex of lysigenous aerenchyma from cells in cubic packing, outer aporous cortex of cells in hexagonal non-radial packing. Bar=0.1 mm. (For details, see Justin and Armstrong, 1987.)

©
Z
Lb~
©
©
Z

62 W. ARMSTRONG ET AL.
the roots is generally associated with poorer top growth; for most of the sur- face rooting species we have examined, shoot dry weights were lower by at least one-half in permanently flooded soils with a stagnant water regime (Justin and Armstrong, 1987). It is interesting to find a significant propor- tion of wetland plants represented in the superficial rooting category. In the field, however, such plants often grow in flushed soils where the detrimental effects of root confinement will be offset, at least to some extent, by the en- hanced nutrient supply. Why roots should develop a superficial rooting habit has not been thoroughly researched. It may be that in some cases roots pene- trating below the aerobic zone simply die. On the other hand, root orientation is frequently altered by flooding; horizontal rooting is common, as is upward growth, and it seems reasonable to suppose that hormonal control might be involved (see also Reid and Bradford, 1984).
The major stratagem for survival and competitiveness in waterlogged soils is, however, the enhanced development of gas space in the primary and/or secondary cortex of the root. A majority of the wetland and intermediate spe- cies shown in Fig. 2 have fractional root porosities exceeding 10% and in most cases this can be traced to aerenchyma development. In some species, this is produced only in response to flooding, in others it is an almost constant fea- ture of the root system, although it may be made more extensive by flooding (Justin and Armstrong, 1987 ). Lysigenous aerenchyma, caused by the sepa- ration and almost concomitant collapse of cortical cells (Fig. 3b) is a very ancient feature which can be seen in stem sections of the Devonian lycopod Asteroxylon mackiei Kidston & Lang.
Aerenchyma development in roots serves two purposes: it can greatly re- duce diffusive resistance to longitudinal (axial) transport of gases from and to the shoot; it also results in a reduction in 02 demand per unit volume and this naturally enhances further the 02 supply to the apical parts of the root. Of the two, however, it is the reduced diffusive resistance which accomplishes most in terms of elevated aeration, a conclusion arrived at in analogue mo- delling studies in which the anatomical and respiratory demand characteris- tics of non-wetland and wetland root systems were intermixed (Armstrong, 1979).
S T R U C T U R A L A N D METABOLIC DIVERSITY, M A T H E M A T I C A L M O D E L L I N G
The anatomical basis of differences in root porosity depends on species, on root type and even on the stage of development of individual roots. On the subject of primary root anatomy, we are often schooled into thinking in terms of monocotyledonous or dicotyledonous stereotypes, e.g. Zea mays L. and Ranunculus. We also learn of anomalous secondary growth, but are inclined to believe it to be of very limited distribution. The anatomical study of the

ROOT ADAPTATION TO SOIL WATERLOGGING 63
roots of the 91 species represented in Fig. 2 (Justin and Armstrong, 1987) very much emphasises the fallacy of this conception.
Pre- or non-aerenchymatous gas space in roots is a function of the type and closeness of cell packing, and because of the potential for varying combina- tions there is considerable scope for diversity. As seen in transection, cell packing is mostly of two types: hexagonal, in which each cell makes contact with six near neighbours (Fig. 3a and b) and cubic, in which each cell con- tacts four near neighbours (Fig. 3c and d).
Packing may be so close that gas space is extremely small to non-existent. This is most characteristic of hexagonal packing in outer cortical tissues where the cells are also often thick walled, and the form and location of this type of tissue indicates a provision for resisting bending and pulling strains (exam- ples may be found in the basal parts of maize (Fig. 3b) and Spartina nodal roots). In terms of aeration, such tissues are detrimental, at least in the young root, and because of the metabolic demands of secondary wall deposition, they may have a high 02 demand; they also make no direct contribution to internal gas space provision. Because of their location chiefly in basal regions, however, their O2 demands will be relatively easily satisfied. In the basal part of the maize root, this type of tissue can occupy up to 50% of the total root cross-sectional area. As roots extend into wetland sites, the proportion of this outer cortical tissue diminishes and may ultimately disappear (Fig. 3c); it is a good example of structural and probably metabolic diversity within individ- ual roots. Elsewhere in the root cortex, hexagonally packed tissues usually contain gas space. Individual spaces are triquetrous in section (Fig. 3a and e ), but the maximum achievable porosity with such an arrangement, cells in point contact only, is just 9.3%. For a whole root section, including the non- porous stele, porosities must therefore be significantly < 9.3%. Hexagonally packed tissues may be small celled with the cells radially aligned as often in the inner cortex of the Juncaceae (Fig. 3f), or larger celled and radially aligned as in the mid-cortex of Elymus pycnanthus (Godr.) Melderis (Fig. 3e) or larger celled and non-radially aligned as in some other grasses and many di- cotyledonous non-wetland species. Another characteristic of cells in hexago- nal packing is that they seem to be less likely to subsequently form aeren- chyma. Where they do (e.g. wheat and Elymus pycnanthus), the aerenchyma is generally less extensive and much less regular than aerenchyma formed from cells in cubic packing (Trought and Drew, 198 l; Erdmann and Weidenroth, 1986 ). From the point of view of root aeration, the development of hexagonal cell packing may be generally regarded as being disadvantageous.
As seen in transection, the cubic packing of cells is a much better arrange- ment for aeration provision by internal gas transport than is hexagonal pack- ing. The maximum achievable porosity is ~ 21.5% and cells in cubic packing generally, although not invariably, more readily form aerenchyma in sub-ap- ical regions. Again, however, smaller cells in cubic packing may form an inner
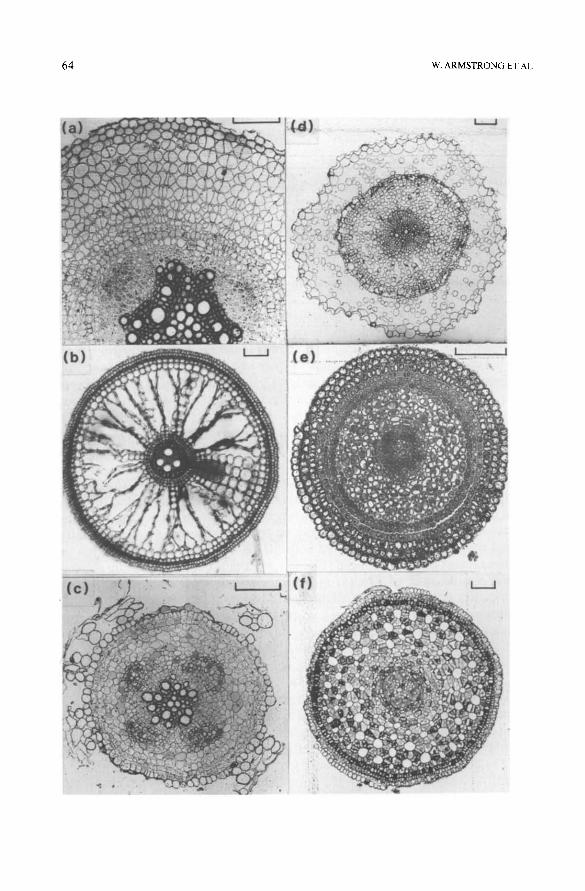

ROOT ADAPTATION TO SOIL WATERLOGGING 6 5
cortex which is resistant to lysigeny and this further adds to the potential for diversity in cortical structure. The mechanism and types of aerenchyma de- velopment are discussed again in the next section.
In some dicotyledonous species from a range of families, cubic packing does not normally lead on to aerenchyma development irrespective of cell size. Instead, there is an anomalous secondary growth: the cells remain potentially meristematic and divide to form the secondary cortex while still retaining a cubic packing arrangement (Fig. 4a, cf. Fig. 3d). Examples are to be found in the Asteraceae and Scrophulariaceae. Although the higher porosity of this cu- bic packing in these cases might be expected to confer some aeration advan- tage in wet sites, the continuing meristematic nature of the cells and the de- position of secondary walls probably create a high 02 demand which negates the porosity benefits. In any event, plants such as Cirsium eriophorum (L.) Scop. are highly sensitive to waterlogging, whilst even Senecio aquaticus Hill, which does eventually form some aerenchyma in the outer cortex, is not par- ticularly successful when grown in stagnant waterlogged compost (Justin and Armstrong, 1987).
Structural change, in the form of aerenchyma development, is invariably accompanied causally and coincidentally by basipetal changes in 02 demand, both in the cortex, wall layers and the stele (Armstrong et al., 1990a). The changes in 02 demand are most frequently a reduction in demand associated with an ageing of tissues or their collapse. On the other hand, this decrease in demand can be offset to some extent by lateral root development sub-apically (Armstrong et al., 1983). Lateral roots may inhibit lysigenous aerenchyma formation (Fig. 4b) and can occlude cortical gas space for axial transport. This may be particularly acute if lateral roots are large in relation to the root from which they develop, e.g. in fibrous branching root systems such as wheat (Thomson et al., 1989).
Other changes which considerably add to the diversity and complexity in terms of root aeration, are changes in permeability to gases such as 02. We find this extremely developed in the hypodermal and surface layers of wet- land grasses such as rice and Phragmites, and sedges such as Eriophorum angustifolium Honckeny, such that 02 may not leak from the root in some
Fig. 4. Transverse sections of roots: (a) Cirsium eriophorum, secondary thickening involves renewed division of the primary cortex, cubic packing retained; (b) rice, lysigenous aeren- chyma from cells in cubic packing, inhibition in region of lateral root development; (c) Rumex acetosella L., "normal" low/non-porous secondary growth in stele and isolated, originally non- aerenchymatous, aerenchymatous primary cortex, cells originally in cubic packing; (d) Plantago maritima L, schizogenous aerenchymatous primary cortex isolated by secondary growth in which secondary cortex is also schizogenously aerenchymatous; (e) Rumex crispus L., cubic packing in cortex 2 mm behind apex, and (f) same root 20/tin further on, showing the division which has taken place to create schizogenous aerenchyma. Bar= 0.1 ram. (For details, see Justin and Armstrong, 1987. )

6 6 W. ARMSTRONG ET AL.
A B
L : I 0 L :10
L ( i n f ) : 2 e , 3 5 ! L :20 L :16
C D
L : IO L : IO
r IIlllt L : 1 9 L :19
I
L
k ( I n f ) : 2 5 . 6 L ( l e f ) : 2 2 . 4 L ( I n f ) : 2 9 , 1 5
I L_J , Fig. 5. Longitudinal half-sectional diagrams of roots in waterlogged soil at various stages of growth (L) and predicting maximum rooting depths (Lmf.: root apex totally anoxic) for roots in which respiratory demand declines and fractional porosity increases with distance from the apex, but in which 02 permeability of the outer stele and wall layers varies. (A) Root wall impermeable from z=0 to z=L (see Fig. 3), no rhizosphere oxygenation; (B) root wall fully permeable to 02 throughout its length; (C) root wall permeability decreasing basipetally to become minimal at 50 mm from apex, rhizosphere oxygenation concentrated near root apex; (D) as (C), but with stelar 02 diffusivity declining by an order of magnitude over the apical 40 ram. Note that the changing permeability characteristics of stele and wall layers can increase the predictable potential rooting depth by ~ 30%. (Redrawn from Armstrong and Beckett, 1987.)
parts (Armstrong, 1971; Gaynard and Armstrong, 1987; Amstrong and Arm- strong, 1988). It can also be detected, however, in a whole range of species. Sub-apical reductions in root wall permeability have now been found in wheat (Thomson et al., 1989) and in several R u m e x species (Laan, et al., 1989b). A reduction in root wall permeability will of course reduce radial 02 losses into the rhizosphere and presumably also the intake of phytotoxic materials, particularly those which move into the apoplast. A reduction in 02 permea- bility also presumably attends the various depositions of suberin and second- ary wall in the endodermis and in the stele, further adding to the complexity

Cor ticaL oxygen COnCenlrolion
r=O rl z2 / r3 r4 "co
z=L
ROOT ADAPTATION TO SOIL WATERLOGGING 67
Anoxic wall
Fig. 6. Longitudinal half-sectional plan of root depicting the multicylindrical framework for the aeration modelling shown in Fig. 5. Symbols: r = 0, central axis; r~, r2 = radii of inner and outer stele; r3=outer radius of porous cortex; r4=outer radius of non-porous wall layers (radius of root); ro~ = radius of oxygenated rhizosphere; rz= oxygenated rhizosphere; r,, radius of internal anoxic region (stele); z= distance from root base; L = root length. The predicted 02 concentra- tion profile is inset within the limits r2-r3 and z=0 to z = L . Areas of dark shading indicate anoxia (stele and soil). (Redrawn from Armstrong and Beckett, 1987.)
of the aeration process whether 0 2 transport is from the shoot or, as in drained conditions, from the soil. Further elements of diversity and complexity arise because of normal or anomalous secondary growth of both stems and roots; "normally" in roots the secondary cortex arises from a secondary meristem in the pericycle of the stele and the primary cortex of more apical parts, in- eluding the gas space system, is soon isolated from the shoot (Fig. 4c). For many plants, secondary growth may be the single most detrimental character- istic to flood tolerance (Justin and Armstrong, 1987). In some cases, how- ever, soil waterlogging appears to suppress or very much restrict secondary growth in roots; in other cases the secondary cortex itself, particularly in tap- roots, may be highly aerenchymatous (Fig. 4d) and particularly well adapted for supporting the growth of lateral roots (Justin and Armstrong, 1987 ).
We have recently made some attempts to discover how some of these de- velopmental patterns might affect root aeration in flooded soil. Our approach has been that of diffusion-based mathematical modelling by numerical itera- rive techniques, techniques made possible by the now much improved com- puter technology. Our findings have been presented in detail elsewhere (Armstrong and Beckett, 1987) and some of the results are summarised in

68 W. ARMSTRONG ET AL.
Fig. 5 (see also Fig. 6). They demonstrate how a lowering of sub-apical root wall 02 permeability will lead (a) to better aeration of the root as a whole and (b) to a potential increase in the aeration-supportable rooting depth in flooded soil. The results also predict that a sub-apical to basal lowering of endodermal and stelar 02 permeability might lead to some degree of sub-ap- ical stelar anoxia, again leading to improved aeration for the remainder of the root and a greater aeration-rooting depth (see also, Smith et al., 1986 ). Some further observations on the results of modelling are made later in the section on "Rhizosphere considerations".
AERENCHYMA DEVELOPMENT
Aerenchyma forms in roots either (a) lysigenously, by concomitant cell separation and collapse (collapse may be somewhat delayed), or (b) schizo- genously, by cell separation (often accompanying cell division) without col- lapse. Aerenchyma can arise in primary or secondary tissues.
Recent years have seen a particularly interesting development in our un- derstanding of the mechanism underlying lysigenous aerenchyma formation. Previously, it had often been assumed that aerenchyma was caused by severe 02 stress, but this failed to explain why aerenchyma once formed, and thus creating improved aeration, should then continue to develop. Research has now shown that the hormone ethene (C2H4) can trigger lysigenous aeren- chyma formation in maize roots and that various inhibitors of ethene action and ethene production in roots can inhibit aerenchyma development (Drew et al., 1979, 1981; Jackson et al., 1981; Konings, 1982). Ethene production is common in roots, but the accumulation to levels which induce aerenchyma development is probably a consequence of soil flooding which helps partially to trap the gas within the roots (Konings and Jackson, 1979 ). Further careful experimentation has shown that the greatest stimulus for endogenous ethene production and aerenchyma formation in maize is a lowering of 02 levels in the cortex to ~3% (Jackson et al., 1985b). The partial anoxia thereby in- duced in the stele stimulates a greater production of the ethene precursor 1- aminocyclopropane-l-carboxylic acid (ACC) which can be transformed into ethene if transported to the aerated outer stele or cortex (Atwell et al., 1988 ). Aerenchyma development occurs, therefore, before the root suffers severe 02 stress and the proposed mechanism explains why it is that aerenchyma con- tinues to form despite the enhanced degree of aeration which it creates.
Although for maize the role of ethene seems established beyond doubt, un- certainty has been expressed over whether ethene is similarly involved in ly- sigenous aerenchyma development in other species. With the International Rice Research Institute rice cultivar RB3 for example, Jackson et al. ( 1985a) could find no evidence for ethene involvement and suggested that the rice had become genetically programmed to develop aerenchyma regardless of the soil

ROOT ADAPTATION TO SOIL WATERLOGGING 69
condition. We are not entirely satisfied with this explanation since it has often been shown that rice roots will develop less aerenchyma if grown in drained soils (Armstrong, 1971; Das and Jat, 1977 ). Contemporaneous with the stud- ies of Jackson et al. (1985a), we investigated root aerenchyma development in a different rice cultivar, previously shown to be responsive to soil flooding (Norin 36) (Armstrong, 1971 ). Our findings (Justin and Armstrong, 1990) were that, as in maize, the ethene inhibitor Ag + could reduce or almost totally prevent aerenchyma development, whilst at the same time it did not inhibit root growth. Also, the effects of the silver could be partially overcome by si- multaneously exposing the roots to exogenous ethene. Subsequently, with cv. RB3, we have found some evidence of responsiveness to ethene and silver, although this variety is apparently very much less responsive than cv. Norin 36. Why should two varieties respond differently? No doubt a whole variety of reasons could be proposed; for example, it may be that RB3 has a greater inherent sensitivity to ethene, it may also produce more endogenous ethene than N36, or it may be that root wall permeability may decline more severely in RB3 so reducing endogenous ethene losses and also helping to prevent the entry of Ag + when it is applied as an inhibitor. Whatever the reason, how- ever, it would seem that at this stage it would be wrong to dismiss the possi- bility that ethene production by roots might be the major trigger for lysigen- ous aerenchyma development.
Of the mechanism underlying schizogenous aerenchyma formation, almost nothing is known. Recently, however, it has been shown that among the Rumex species the degree of schizogenous aerenchyma development appears to re- spond to the degree of external aeration (Laan et al., 1989a,). Close to the root/root cap junction in Rumex the cortical initials are, in transection, ra- dially aligned in cubic packing (Fig. 4e ). Aerenchyma development arises by subsequent anticlinal, periclinal and oblique cell divisions accompanied by cell separation. This creates a honeycomb of tissue bounding lacunae which are longitudinally uninterrupted throughout the root (Fig. 4f). In Rumex thyrsiflorus Fingerh., a relatively flood-intolerant species, the aerenchyma does not develop in response to low O2 or to soil flooding. Similarly, Rumex crispus L. does not develop aerenchyma in forcibly aerated solutions, but in stagnant solution or in flooded soil the development is extensive. The roots of Rumex maritimus L. develop aerenchyma both in aerated and non-aerated media, although the lacunae may be smaller in the aerated medium. It will be inter- esting to discover whether the Rumex type of aerenchyma is also responsive to ethene and ethene inhibitors.
RHIZOSPHERE CONSIDERATIONS
In this final section, we return again to the likely importance of root dimen- sion as an adaptation in flooded soil. It has been emphasised earlier that ra-

70 W. ARMSTRONG ET AL.
d i a l 0 2 loss from root to rhizosphere might be of considerable importance for protecting the plant against the re-oxidisable phytotoxins which accumulate in anoxic soils. Rhizosphere oxygenation may also support an aerobic nitri- fying microflora, a feature which may be important in the "root-zone" treat- ment of sewage effluent in reed beds. However, just as superficial and narrow roots may, by virtue of their high surface to volume ratio, be advantageous for survival in flooded soils (see "Stratagems of survival") it can readily be deduced that narrow but short roots should also be advantageous for soil ox- idation. Some years, ago, it was demonstrated how the zones of oxidation around the lateral roots of Eriophorum angustifolium were, relative to their size, much greater than around the parent adventitious root (Armstrong, 1967 ), an observation which was corroborated by mathematical modelling (Armstrong, 1970). More recently, it has been shown that rhizosphere oxi- dation by Phragmites australis (Trin.) Cav. ex Steudel is most intense within the zones of fine laterals which are borne sub-apically on the adventitious roots (Armstrong and Armstrong, 1988 ). We have recently explored this ob- servation using our more advanced mathematical models and arrived at some interesting conclusions (Armstrong et al., 1990b) namely ( 1 ) that for a plant such as Phragmites australis, the oxidising power of the laterals on just 10 mm of adventitious roots could be 5 times greater than for the much longer parent root (it should be noted that much of the sub-apical surface of the parental roots can be almost impermeable to O2) and (2) that because of their low fractional porosities the lateral roots cannot be other than very short i.e. < 20 mm. The implications of these findings would seem to be that the shortness and narrowness of the fine lateral roots in such wetland species as Phragmites and rice ought to be genetically rather than environmentally determined de- velopmental features. We have yet to ascertain whether this is so.
CONCLUDING REMARKS
This paper has centred on the diversity/variability among and within root systems. This diversity is considerable and embraces root morphology, anat- omy and physiology; it extends even to variation within individual roots. It is thus of considerable consequence for the aeration of root systems and the differential flood tolerance of plants. One must conclude that although the general principles of root aeration may now be understood, there is still much scope for further study into the aeration biology of roots. An important aspect in this should be further investigations into the mechanisms underlying aerenchyma formation, but equally important are topics which have been be- yond the scope of this paper, namely the role of anaerobic metabolism in roots, their tolerance of anoxia and, also, the dynamics of root and soil aeration, which have so far rarely been the subject of investigation.

ROOT ADAPTATION TO SOIL WATERLOGGING
ACKNOWLEDGEMENTS
We thank E.M. Sharpe for typing the manuscript.
71
REFERENCES
Armstrong, J. and Armstrong, W., 1988. Phragmites australis: A preliminary study of soil-oxi- dising sites and internal gas transport pathways. New Phytol., 108: 373-382.
Armstrong, J. and Armstrong, W., 1991. A convective through-flow of gases in Phragmites australis. Aquat. Bot. 39: 73-86.
Armstrong, W., 1967. The oxidising activity of roots in waterlogged soils. Physiol. Plant., 20: 920-926.
Armstrong, W., 1970. Rhizosphere oxidation in rice and other species: a mathematical model based on the oxygen flux component. Physiol. Plant., 23: 623-630.
Armstrong, W., 1971. Radial oxygen losses from intact rice roots as affected by distance from the apex, respiration and waterlogging, Physiol. Plant., 25: 192-197.
Armstrong, W., 1979. Aeration in higher plants. In: H.W. Woolhouse (Editor), Advances in Botanical Research, 7. Academic Press, London, pp. 225-232.
Armstrong, W., 1982. Waterlogged soils. In: J.R. Etherington (Editor), Environment and Plant Ecology. John Wiley and Son, pp. 290-330.
Armstrong, W. and Beckett, P.M., 1987. Internal aeration and the development of stelar anoxia in submerged roots: a multishelled mathematical model combining axial diffusion of oxygen in the cortex with radial losses to the stele, the wall layers and the rhizosphere. New Phytol., 105: 221-245.
Armstrong, W., Healy, M.T. and Lythe, S., 1983. Oxygen diffusion in pea. II. The oxygen status of the primary root as affected by growth, the production of laterals and radial oxygen loss. New Phytol., 94: 549-559.
Armstrong, W., Beckett, P.M., Justin, S.H.F.W. and Lythe, S., 1990a. Modelling and other as- pects of root aeration. In: M.B. Jackson, D.D. Davies and H. Lambers (Editors), Plant Life under Oxygen Deprivation. S.P.B. Academic Publishings, hv The Hague, pp. 283-302.
Armstrong, W., Armstrong, J. and Bechett, P.M., 1990b. Measurement and modelling of oxygen release from roots of Phragmites australis. In: P.F. Cooper and B.C. Findlater (Editors), The Use of Constructed Wetlands in Water Pollution Control. Pergamon, Oxford, pp. 41-52.
Atwell, B.J., Drew, M.C. and Jackson, M.B., 1988. The influence of oxygen deficiency on eth- ylene synthesis, l-aminocyclopropane- l-carboxylic acid levels and aerenchyma formation in Zea mays. Physiol. Plant., 72:15-22.
Beckett, P.M., Armstrong, W., Justin, S.H.F.W. and Armstrong, J., 1988. On the relative im- portance of convective and diffusive gas-flows in plant aeration. New Phytol., 110: 463-468.
Crawford, R.M.M., Monk, L.S. and Zochowski, Z.M., 1987. Enhancement of anoxia tolerance by removal of volatile products of anaerobiosis. In: R.M.M. Crawford (Editor), Plant Life in Aquatic and Amphibious Habitats. Br. Ecol. Soc., Spec. Publ. Ser. No. 5, Blackwell, Oxford, pp. 375-384.
Dacey, J.W.H., 1981. Pressurised ventilation in the yellow water-lily. Ecology, 62:1137-1147. Das, D.K. and Jat, R.L., 1977. Influence of three soil-water regimes on root porosity and growth
of four rice varieties. Agron. J., 64:197-200. Drew, M.C., Jackson, M.B. and Giffard, S., 1979. Ethylene promoted adventitious rooting and
the development of cortical air spaces (aerenchyma) in roots may be adaptive responses to flooding in Zea mays L. Planta, 147: 83-88.
Drew, M.C., Jackson, M.B., Giffard, S. and Campbell, R., 1981. Inhibition by silver ions of gas

72 W. ARMSTRONG ET AL.
space (aerenchyma) formation in adventitious roots of Zea mays L. subjected to exogenous ethylene or to oxygen deficiency. Planta, 153:217-224.
Erdmann, B. and Weidenroth, E.M., 1986. Changes in the root system of wheat seedlings follow- ing root anaerobiosis. II. Morphology and anatomy of evolution forms. Ann. Bot., 58: 607- 616.
Gambrell, R.P. and Patrick, W.H., Jr., 1978. The chemical and microbiological properties of anaerobic soils and sediments. In: D.D. Hook and R.M.M. Crawford (Editors), Plant Life in Anaerobic Environments. Ann Arbor Science, MI, pp. 375-424.
Gaynard, T.J. and Armstrong, W., 1987. Some aspects of internal plant aeration in amphibious habitats, in: R.M.M. Crawford (Editor), Plant Life in Aquatic and Amphibious Habitats. Br. Ecol. Soc. Spec. Pubi. Ser. No. 5, Blackwell, Oxford, pp. 303-320.
Greenwood, D.J., 1967. Studies in oxygen transport through mustard seedlings (Sinapsis alba, L). New Phytol., 66: 597-606.
Grosse, W. and Mevi-Schutz, J., 1987. A beneficial gas transport system in Nymphoidespehata. Am. J. Bot., 74: 941-952.
Grosse, W., Biichel, H.B. and Tiebel, H., 1991. Pressurised ventilation in wetland plants. Aquat. Bot., 39: 87-96.
Hendry, G.A.F. and Brocklebank, K.J., 1985. Iron-induced oxygen radical metabolism in waterlogged plants. New Phytol., 101: 199-206.
Higuchi, T., 1982. Gaseous carbon dioxide transport through the aerenchyma and intercellular spaces in relation to the uptake of carbon dioxide by rice roots. Soil Sci. Plant Nutr., 28: 491-497.
Jackson, M.B., Drew, M.C. and Giffard, S.C., 1981. Effects of applying ethylene to the root system of Zea mays on growth and nutrient concentration in relation to flooding tolerance. Physiol. Plant., 52: 23-28.
Jackson, M.B., Fenning, T.M. and Jenkins, W., 1985a. Aerenchyma (gas-space) formation in adventitious roots of rice (Oryza sativa L.) is not controlled by ethylene or small partial pressures of oxygen. J. Exp. Bot., 36:1566-1572.
Jackson, M.B., Fenning, T.M., Drew, M.C. and Saker, L.R., 1985b. Stimulation of ethylene, production and gas-space (aerenchyma) formation in adventitious roots of Zea mays. L. by small partial pressures of oxygen. Planta, 165: 486-492.
Justin, S.H.F.W. and Armstrong, W., 1987. The anatomical characteristics of roots and plant response to soil flooding. New Phytol., 106: 465-495.
Justin, S.H.F.W. and Armstrong, W., 1990. Evidence for the involvement of ethene in aeren- chyma formation in adventitious roots of rice. New Phytol., in press.
Kon~alov~i, H., Pokorn~, J. and Kv6t, J., 1988. Root ventilation in Carex gracilis Curt.: diffu- sion or mass flow? Aquat. Bot, 30:149-155.
Konings, H., 1982. Ethylene-promoted formation of aerenchyma in seedling roots of Zea mays L. under aerated and non-aerated conditions. Physiol. Plant., 54:119-124.
Konings, H. and Jackson, M.B., 1979. A relationship between rates of ethylene production by roots and the promoting or inhibiting effects of exogenous ethylene and water on root elon- gation. Z. Pflanzenphysiol., 92: 385-397.
Laan, P., Berrevoets, M.J., Lythe, S., Armstrong, W. and Blom, C.W.P.M., 1989a. Root mor- phology and aerenchyma formation as indicators of flood tolerance in Rumex species. J. Ecol., 77: 693-703.
Laan, P., Smolders, A., Blom, C.W.P.M. and Armstrong, W., 1989b. The relative roles of inter- nal aeration, radial oxygen losses, iron exclusion and nutrient balance in flood-tolerance of Rumex species. Acta Bot. Neerl., 38:131 -145.
Luxmoore, R.J., Stolzy, L.H. and Letey, J., 1970. Oxygen diffusion in the soil-plant system. Agron. J., 62:317-322.
Marschner, H., 1986. Mineral Nutrition of Higher Plants. Academic Press, London, 674 pp.

ROOT ADAPTATION TO SOIL WATERLOGGING 7 3
Ponnamperuma, F.N., 1984. Effects of flooding on soils. In: T.T. Kozlowski (Editor), Flooding and Plant Growth. Academic Press, London, pp. 9-45.
Reid, D.M. and Bradford, K.J., 1984. Effects of flooding on hormone relations. In: T.T. Kozlowski (Editor), Flooding and Plant Growth. Academic Press, London, pp. 195- 219.
Smith, A.M., Hylton, C.M., Koch, L. and Woolhouse, H.W., 1986. Alcohol dehydrogenase ac- tivity in the roots of marsh plants in naturally waterlogged soils. Planta, 168:130-138.
Thomson, C.J., Armstrong, W., Waters, I. and Greenway, H., 1989. Aerenchyma formation and associated oxygen movement in seminal and crown roots of wheat grown in hypoxic solu- tions. Plant Cell Environ., 13: 395-403.
Trought, M.C.T. and Drew, M.C., 1981. The development of waterlogging damage in young wheat plants in anaerobic solution culture. J. Exp. Bot., 31: 1573-1585.
Webb, T. and Armstrong, W., 1983. The effects of anoxia and carbohydrates on the growth and viability of rice, pea and pumpkin roots. J. Exp. Bot., 34: 579-603.
Wheeler, B.D., AI-Farraj, M.M. and Cook, R.E.D., 1985. Iron toxicity to plants in base-rich wetlands: comparative effects on the distribution and growth of Epilobium hirsutum L. and Juncus subnodulosus Schrank. New Phytol., 100: 653-669.





























