Embed Size (px)
Citation preview

Roles of Glutamate Receptors and the Mammalian Target ofRapamycin (mTOR) Signaling Pathway in Activity-dependentDendritic Protein Synthesis in Hippocampal Neurons*□S
Received for publication, November 22, 2005, and in revised form, April 6, 2006 Published, JBC Papers in Press, May 1, 2006, DOI 10.1074/jbc.M512524200
Ruomu Gong, Chang Sin Park, Nima Rezaei Abbassi, and Shao-Jun Tang1
From the Department of Neurobiology and Behavior, Center for Neurobiology of Learning and Memory,University of California, Irvine, California 92697-3800
Local protein synthesis in neuronal dendrites is critical for syn-aptic plasticity. However, the signaling cascades that couple synap-tic activation todendritic protein synthesis remain elusive.Thepur-pose of this study is to determine the role of glutamate receptors andthemammalian target of rapamycin (mTOR) signaling in regulatingdendritic protein synthesis in live neurons. We first characterizedthe involvement of various subtypes of glutamate receptors and themTOR kinase in regulating dendritic synthesis of a green fluores-cent protein (GFP) reporter controlledby�CaMKII5�and3�untrans-lated regions in culturedhippocampal neurons. Specific antagonists ofN-methyl-D-aspartic acid (NMDA), �-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA), and metabotropic glutamate recep-tors abolished glutamate-induced dendritic GFP synthesis, whereasagonists of NMDA and metabotropic but not AMPA glutamatereceptors activated GFP synthesis in dendrites. Inhibitions of themTOR signaling, as well as its upstream activators, phosphatidyli-nositol 3-kinase and AKT, blocked NMDA receptor-dependentdendritic GFP synthesis. Conversely, activation of mTOR signalingstimulated dendritic GFP synthesis. In addition, we also found thatinhibition of the mTOR kinase blocked dendritic synthesis of theendogenous�CaMKII andMAP2 proteins induced by tetanic stim-ulations in hippocampal slices. These results identify critical rolesof NMDA receptors and themTOR signaling pathway for control ofsynaptic activity-induced dendritic protein synthesis in hippocam-pal neurons.
Protein synthesis is essential for the expression of long-term synapticplasticity (1). Recent studies indicate that synaptic proteins can be syn-thesized locally in dendrites and at synaptic regions (2). Local proteinsynthesis is required for BDNF2- and tetanus-induced long-termpotentiation (LTP) (3–5),metabotropicglutamate receptor (mGluR)-
dependent long-term depression (LTD) (6), and serotonin-inducedlong-term facilitation ofAplysia neurons (7, 8). A number of mRNAsthat encode proteins involved in synaptic plasticity, including�CaMKII, MAP2, and Arc/Arg3.1, have been found in dendrites (2).Importantly, dendritic synthesis of the �CaMKII protein is essentialfor late-phase LTP and various forms of memory (9).Synaptic activation rapidly increases �CaMKII protein in hippocampal
dendrites (10, 11), causes the accumulation of newly synthesized Arc pro-tein at the activated synapses (12, 13), and enriches the polyribosome inspines (14). In addition, stimulation of group 1 mGluRs by 3,5-dihydroxy-phenylglycine activates protein synthesis in synaptosomes (15), transecteddendrites (16, 17), andhippocampaldendrites in vivo (18).TheactivationofNMDA receptors induced synthesis of�CaMKII protein in synaptosomes(19) and the activation of dopamine D1/D5 receptor in cultured neuronsstimulates GluR1 synthesis in dendrites (20). Despite these importantobservations, the role of glutamate receptors in regulatingdendritic proteinsynthesis in live neurons has not been systematically investigated. Besidessynaptic activity, other factors such as estrogen and BDNF also regulatedendritic protein synthesis (21–25).Dendritic mRNAs are often translationally repressed before stimula-
tions. Miniature activities may play a critical role in repressing dendriticmRNAs from translation (26). Recent studies indicated that fragile Xmen-tal retardation protein can repress the translation from multiple dendriticmRNAs, including CaMKII and Arc (27). Other translational repressorproteins in dendrites or postsynaptic regions include RNG105 and Pumilio(28–30). Non-translated RNAs such as BC1may also be involved in trans-lational repression of dendritic mRNAs (27, 31).The mechanisms that couple synaptic activities to the activation of
dendritic protein synthesis are poorly understood. TheAurora signalingwas suggested to control activity-dependent translational activation byregulating polyadenylation of cytoplasmic polyadenylation element-containing mRNAs (e.g. �CaMKII) (32–34). Cytoplasmic polyadenyl-ation element-dependent polyadenylation of the �CaMKII mRNA inneurons is also regulated by CaMKII signaling (35). MAPK signalingwas found to be required for NMDA receptor-mediated activation ofMnk1 kinase, which phosphorylates eukaryotic initiation factor 4E (eIF-4E) (36). Down-regulation of MAPK signaling impaired not only L-LTPandmemory consolidation, which are protein synthesis-dependent, butalso activity-regulated phosphorylation of multiple translation initia-tion factors (37). In eukaryotic cells, mTOR signaling regulates transla-tion by modulating the activity of eIF-4E, the cap-binding protein, andother proteins that are involved in translation initiation and elongationsuch as p70S6K, eIF-4GI, eIF-4B, and eEF2 (38). The mTOR signalingpathway is present in the hippocampal neurons at the synaptic region(39), activated by synaptic activity (40, 41), and required for LTP (5, 39,40) and LTD expression (42) and memory consolidation (43). Despitethe suggestive evidence, the role of the mTOR signaling in control of
* The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
□S The on-line version of this article (available at http://www.jbc.org) contains supple-mental Figs. S1–S6.
1 Supported by the American Heart Association, the EJLB Foundation, the WhitehallFoundation, and the United States Army Medical Research Command. To whom cor-respondence should be addressed: 303 Qureshey Research Laboratory, Center forNeurobiology of Learning and Memory, University of California, Irvine, CA 92697-3800. Tel.: 949-824-9580; Fax: 949-824-9762; E-mail: [email protected].
2 The abbreviations used are: BDNF, brain-derived neurotrophic factor; LTP, long-termpotentiation; NMDAR, N-methyl-D-aspartate receptor; APV, 2 amino-5-phosponovalericacid; AMPA, �-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor; mGluR,metabotropic glutamate receptor; mTOR, mammalian target of rapamycin; ROI, region ofinterest; MAPK, mitogen-activated protein kinase; eIF 4E, eukaryotic initiation factor 4E;GFP, green fluorescent protein; UTR, untranslated region; PI3K, phosphatidylinositol 3-ki-nase; MCPG, (�)-�-methyl-4-carboxyphenylglycine; PA, phosphatidic acid; CNQX,6-cyano-7-nitroquinoxaline-2,3-dione; PBS, phosphate-buffered saline; ACPD, trans-(�)-1-aminocyclopentane-1,3-dicarboxylic acid; mTOR, mammalian target of rapamycin.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 27, pp. 18802–18815, July 7, 2006© 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
18802 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

synaptic activity-dependent dendritic proteins remains to be directlydemonstrated.In this study, we specifically tested the role of glutamate receptors
and the mTOR signaling pathway in regulating activity-dependentprotein synthesis in the dendrites of hippocampal neurons. Using aGFP reporter with the �CaMKII 5� and 3� UTRs (21), we first inves-tigated the role of NMDA, AMPA, and mGlu receptors and mTORsignaling in regulation of dendritic protein synthesis in cultured hip-pocampal neurons. We found that NMDARs, AMPARs, and mGluRsare required for glutamate-induced dendritic GFP synthesis, whereasactivation of NMDARs and mGluRs but not AMPARs led to GFP syn-thesis in dendrites. Inhibition of mTOR signaling abolished NMDAR-and mGluR-dependent synthesis of dendritic GFP. Similarly, inhibitionof themTORupstream activators PI3K andAKT also disruptedNMDAreceptor-dependent dendritic GFP synthesis. On the other hand, acti-vation of mTOR signaling was sufficient to induce GFP synthesis indendrites. Furthermore, we also demonstrated that tetanus-induceddendritic synthesis of the �CaMKII protein in hippocampal slices alsorequired the activation of NMDA receptors and the mTOR signaling.
EXPERIMENTAL PROCEDURES
Reagents—All cell culture media and Lipofectamine 2000 were fromInvitrogen; phosphatidic acid (1-palmitoyl 2-oleoyl) from Avanti PolarLipids; Ascomycin and SH-6 from Calbiochem; LY294002 and wort-mannin from BioMol; poly-D-lysine, laminin, glutamate, anisomycin,rapamycin, d-(�)-2-amino-5-phosphonovaleric acid (APV), NMDA,AMPA, 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX), (�)-�-methyl-4-carboxyphenylglycine (MCPG), and trans-ACPD from Sigma; anti-ratCaMkinase IImonoclonal antibodies fromAffinity BioReagents; and fluo-rescein isothiocyanate-conjugated secondary antibody from JacksonImmunoResearch Laboratories. The rapamycin-resistant mTOR mutantcDNAwas kindly provided by Dr. Jie Chen.
Neuron Culture and Transfection—Primary cultures of hippocampalneurons were prepared from E17 mouse embryos as previouslydescribed (61). Neurons were plated at a density of 200–400 cells/mm2
on glass coverslips coated with poly-D-lysine and laminin. Cells weremaintained in 24-well plates with Neurobasal medium supplementedwith B-27 and 0.5mM L-glutamine for 10–14 days before use. Cells weretransfected with pcDNA3.1-5�UTR-dGFP-3�UTR plasmids (1 �g/cov-erslip), using Lipofectamine 2000 according to the manufacturer’sinstructions. In co-transfection experiments, 1�g ofGFP reporterDNAand 2 �g of rapamycin-resistant mTOR expression DNA were used.Experiments were performed 24 h after transfection.
Electrophysiology—For all electrophysiology experiments, hip-pocampal slices (400 �M) were prepared from 6–8-week-old maleSprague-Dawley rats. Area CA1 of the hippocampal slice was dividedinto two separate sections by introducing a lesion extending from stra-tum pyramidal through stratum radiatum up to the hippocampal fis-sure. Prior to recording, slices were allowed to recover first for 2 h atroom temperature in oxygenated (95% O2, 5% CO2) artificial cerebro-spinal fluid (ACSF: 119 mM NaCl, 2.5 mM KCl, 1.3 mM MgSO4, 2.5 mM
CaCl2, 1.0mMNaH2PO4, 26.2mMNaHCO3, 11.0mMglucose), and thenfor 30 min at 27 °C in the submerged recording chamber that was con-tinuously perfusedwith oxygenatedACSF. Field excitatory postsynapticpotentials of Schaffer Collateral-Commissural pathway synapses wereevoked with concentric bipolar tungsten stimulating electrodes andrecorded with low-resistance glass microelectrodes filled with 3 MNaClevery 2 min. Both stimulating and recording electrodes were visuallyplaced in the middle of stratum radiatum in the section of area CA1proximal to CA3. LTP was induced by high-frequency stimulations (4
trains of 1-s 100 Hz stimulations spaced by 30-s intervals). All drugs inoxygenated ACSF were perfused into the recording chamber through-out the entire duration of the recording.
Immunohistochemistry—Hippocampal slices (400 �M) after elec-trophysiology experiments were immediately fixed in ice-cold 4%paraformaldehyde with 0.2% glutaraldehyde (in phosphate-bufferedsaline (PBS)) for 1 h. Slices were sectioned (25 �M) using a Vibratome3000, permeabilized in 0.7% Triton X-100 in PBS for 1 h, washed withPBS (2 � 5 min), rinsed in 0.1 M glycine in PBS for 1 h, treated with1% sodium borohydride in ddH2O at room temperature for 20 min,and incubated in pre-block buffer (0.05% Triton X-100, 5% donkeyserum in PBS) for 90 min and then with primary antibody (1:100) inpre-block buffer overnight (4 °C). After three washes (3 � 30 min)with the pre-block buffer at room temperature, sections were incu-bated with fluorescein isothiocyanate-conjugated donkey anti-mouse IgG antibody (1:100) in the pre-block buffer at room temper-ature for 1 h, rinsed with PBS (3 � 5 min) and mounted with ProlongGold Antifade Reagents (Molecular Probes).
Image Acquisition and Analysis—For culture neurons, 24 h aftertransfection, neurons on coverslips were placed in a 35-mM dish con-taining HEPES-buffered solution (62) without glycine or picrotoxin.Healthy pyramidal neurons that expressed GFP were chosen for exper-imentation. Selected dendrites were photobleached by exciting GFP at494 nM (100-W mercury lamp) for 10–60 s until the GFP signals werebarely seen. Appropriate drugs were added to the medium immediatelyafter photobleaching. Images were acquired (Olympus BX61 coupledwith an FV12 CCD camera) every 15 min immediately after photo-bleaching and drug applications. Same acquisition parameters and set-tings were applied to controls and experiments. Fluorescent imageswere analyzed withMicroSuite software (Olympus). Fluorescent inten-sity in a distal segment (�100 �M from the cell body) of the photo-bleached dendrites was quantified with Image J (NIH). Changes of themean fluorescent intensity in dendrites over time were determined by�F � (Ft � F0)/F0 (F, mean fluorescent intensity in dendrites; �F, thedifference of the mean fluorescent intensity in dendrites between time tand time 0 (immediately after photobleaching and drug applications);Ft, mean fluorescent intensity of photobleached dendrites after treat-ments at time t; F0, mean fluorescent intensity of the dendritic segmentat time 0). Statistical significance of differences between groups wasdetermined by independent Student’s t tests or analysis of variance(SPSS, Chicago, IL).
Hippocampal Sections—Fluorescent images of the whole hippocam-pal section after immunostaining were acquired with a �4 object on anOlympus BX61 microscope. Acquisition parameters were optimized toavoid signal saturation. To quantify fluorescent signals, we used Image Jsoftware (NIH) to select regions of interest (ROIs) in stimulated andcontrol sides of original images. ROI is a square (100� 100 �M) 100 �M
away from the cutting edge and the CA1 cell body layer. Themean pixelintensity of individual ROIs was quantified with Image J. The ratio ofmean intensities between stimulated and control ROIs was determined.Student’s t test was used to determine the statistical significance of ratiodifferences between groups.
RESULTS
Membrane Depolarization Activates NMDA Receptor-dependentSynthesis of the GFP Reporter Protein in Dendrites of Cultured Hip-pocampal Neurons—We adapted the GFP imaging approach developedby Aakalu et al. (21) to monitor dendritic protein synthesis in culturedhippocampal neurons. Briefly, primary mouse hippocampal neurons
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18803
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

were transfectedwith theGFP reporter plasmid that contains the 5� and3� UTRs from �CaMKII; selective dendrites were photobleached toenhance the detection sensitivity of newly synthesized GFP; GFP recov-ery in photobleached dendrites was monitored by time lapse imaging.Because the GFP was tethered to membrane by a myristoylationsequence to limit its movement, GFP recovery in dendritic segmentsdistal to the cell body (�100 �M) was due to new protein synthesisrather than diffusion from other cell compartments (21). To determine
whether neuronal activities can stimulate GFP synthesis in dendrites,we examined the effect of KCl-mediated depolarization on GFP recov-ery in photobleached dendrites. Compared with sham treatments, KCl(35 mM) stimulation dramatically increased the magnitude of GFPrecovery in distal dendrites (Fig. 1). Consistent with previous observa-tions that the BDNF-induced GFP increase in dendrites was sensitive totranslation inhibitors (21), theKCl-stimulated enhancement of dendriteGFP recovery was blocked by anisomycin (Fig. 1). These observations
FIGURE 1. Membrane depolarization by high potassium results in NMDA receptor-dependent GFP synthesis in dendrites. A, representative cultured hippocampal neurons24 h after transfections of the GFP report plasmid. a, sham treatments; b, KCl (35 mM); c, KCl and anisomycin (10 �g/ml); d, KCl and APV (50 �M). In this and all subsequent experiments,vehicles for chemicals were used in sham treatments. Arrows indicate the dendrites shown in B. B, time lapse images of straightened dendrites at a higher magnification. Shown aredendritic processes immediately following the cell body. The first image (0 min) was taken immediately after brief photobleaching and drug applications, and subsequent imageswere recorded every 15 min. In this and subsequent figures, distal dendritic segments (the upstream boundary is 100 �M from the cell body) used for quantification of GFP signals areindicated by boxes; bars in A and B, 10 �M. C, summary of changes of GFP signals in distal dendritic segments. GFP signals in distal dendritic segments were quantified. The change ofGFP signals is indicated by �F/F (mean � S.E.). p values were determined by Student’s t tests.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
18804 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from
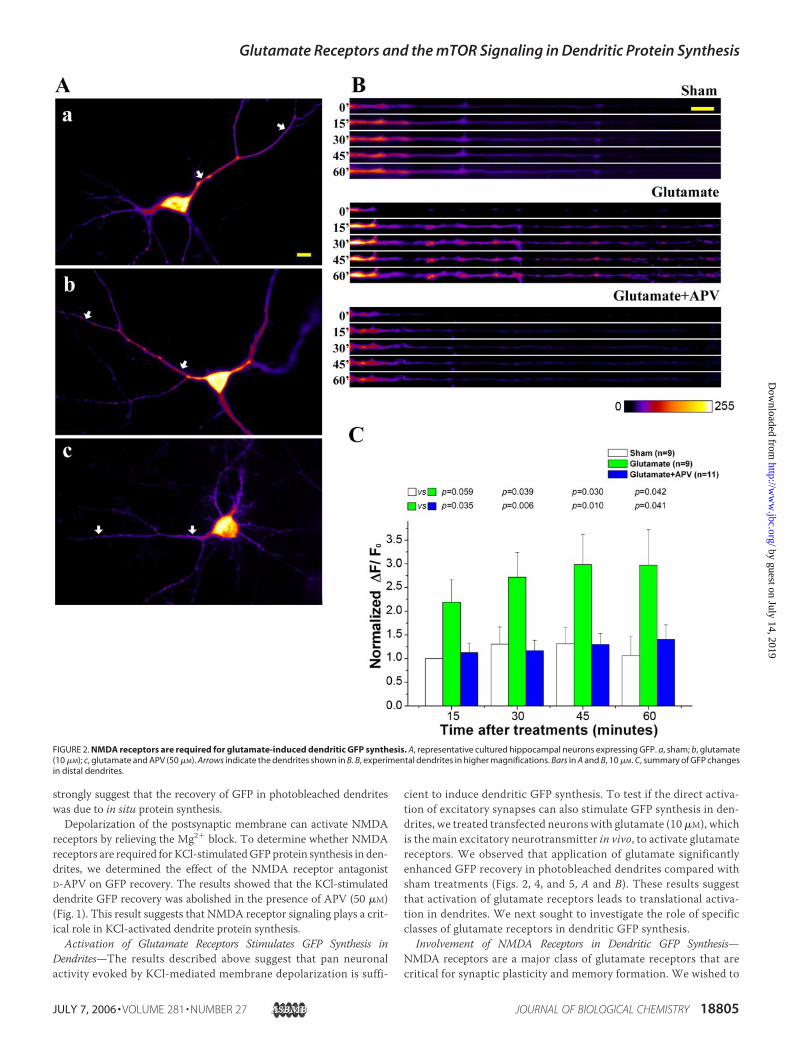
strongly suggest that the recovery of GFP in photobleached dendriteswas due to in situ protein synthesis.Depolarization of the postsynaptic membrane can activate NMDA
receptors by relieving the Mg2� block. To determine whether NMDAreceptors are required for KCl-stimulatedGFP protein synthesis in den-drites, we determined the effect of the NMDA receptor antagonistD-APV on GFP recovery. The results showed that the KCl-stimulateddendrite GFP recovery was abolished in the presence of APV (50 �M)(Fig. 1). This result suggests that NMDA receptor signaling plays a crit-ical role in KCl-activated dendrite protein synthesis.
Activation of Glutamate Receptors Stimulates GFP Synthesis inDendrites—The results described above suggest that pan neuronalactivity evoked by KCl-mediated membrane depolarization is suffi-
cient to induce dendritic GFP synthesis. To test if the direct activa-tion of excitatory synapses can also stimulate GFP synthesis in den-drites, we treated transfected neurons with glutamate (10 �M), whichis the main excitatory neurotransmitter in vivo, to activate glutamatereceptors. We observed that application of glutamate significantlyenhanced GFP recovery in photobleached dendrites compared withsham treatments (Figs. 2, 4, and 5, A and B). These results suggestthat activation of glutamate receptors leads to translational activa-tion in dendrites. We next sought to investigate the role of specificclasses of glutamate receptors in dendritic GFP synthesis.
Involvement of NMDA Receptors in Dendritic GFP Synthesis—NMDA receptors are a major class of glutamate receptors that arecritical for synaptic plasticity and memory formation. We wished to
FIGURE 2. NMDA receptors are required for glutamate-induced dendritic GFP synthesis. A, representative cultured hippocampal neurons expressing GFP. a, sham; b, glutamate(10 �M); c, glutamate and APV (50 �M). Arrows indicate the dendrites shown in B. B, experimental dendrites in higher magnifications. Bars in A and B, 10 �M. C, summary of GFP changesin distal dendrites.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18805
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

investigate if disruption of NMDA receptor signaling can block glu-tamate-stimulated GFP synthesis in dendrites. Toward this end, weused the NMDA receptor antagonist APV. We demonstrated thatAPV (50 �M) was able to completely abolish glutamate-induced den-dritic GFP synthesis (Fig. 2). These observations indicated thatNMDA receptors are required for glutamate-induced GFP synthesisin dendrites.
We next determined if the direct activation of NMDA receptors isable to cause GFP synthesis in dendrites. The results showed thatNMDA treatments enhanced GFP recovery in photobleached den-drites; this GFP recovery was blocked by APV and anisomycin (Fig. 3).The NMDA-induced GFP recovery was also abolished by anothertranslation inhibitor emetine (supplemental Fig. 1). These results indi-cate theGFP recovery induced byNMDAreceptor activationwas due to
FIGURE 3. Activation of NMDA receptors leads to dendritic GFP synthesis. A, representative hippocampal neurons. a, sham; b, NDMA (50 �M); c, NMDA and anisomycin (10 �g/ml);d, NMDA and APV (50 �M). Arrows indicate the dendrites shown in B. B, experimental dendrites in higher magnifications. Bars in A and B, 10 �M. C, summary of GFP changes in distaldendrites.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
18806 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

protein synthesis rather than other posttranslational processes such asprotein transport from cell bodies. To further exclude the contributionof GFP and mRNA transport, we determined the effect of nocodazoleand cytochalasin B, the inhibitors for microtubule- or actin-basedcytoskeleton, respectively. The result showed that NMDA still stimu-lated GFP recovery in the presence of these inhibitors (supplementalFig. 2). Both nocodazole and cytochalasin B induced an appreciable butnot significant decrease of GFP recovery, probably due to their effectson cell healthiness. Together, these observations suggest that NMDARactivation induced GFP synthesis in dendrites of live neurons.
Involvement of AMPA Receptors in Dendritic GFP Synthesis—TheAMPAclass of glutamate receptors plays a critical role in gating theactivity of NMDA receptors (44). We sought to investigate whetherAMPA receptors are involved in regulation of glutamate-stimulatedGFP synthesis in dendrites. For this purpose, we used CNQX, a specific
antagonist for AMPA receptors. We observed that applications ofCNQX (10 �M) abolished glutamate-induced GFP synthesis (Fig. 4).These results suggest a requirement of AMPA receptor dendritic GFPsynthesis activated by glutamate stimulations. We also determined theeffect of AMPA, a specific agonist for AMPA receptors. In contrast toNMDA treatments, applications of AMPA (10 �M) showed an inhibi-tory effect on GFP recovery in photobleached dendrites (supplementalFig. 3).
Involvement of Metabotropic Glutamate Receptors in DendriticGFP Synthesis—mGluRs are a class of G protein-coupled glutamatereceptors that regulate Ca2� release from intracellular calciumstores. We first studied the effect of MCPG, a specific antagonist forgroup I and II mGluRs. The results showed that applications ofMCPG (200 �M) abolished glutamate-induced GFP synthesis in den-drites (Fig. 5, A and B). These observations indicate a requirement of
FIGURE 4. Requirements of AMPA receptors forglutamate-induced GFP synthesis in dendrites.A, experimental dendrites for determining theeffect of CNQX (10 �M). Bar, 10 �M. B, summary ofGFP changes in distal dendrites.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18807
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

group I and II mGluRs in dendritic GFP synthesis induced by gluta-mate stimulations.To investigate if the activation of mGluRs is able to induce dendritic
GFP synthesis, we used trans-(�)-1-aminocyclopentane-1,3-dicar-boxylic acid (ACPD) to stimulate group I and II mGluRs.We found thatapplications of ACPD (50 �M) stimulated GFP synthesis in dendrites(Fig. 5, C and D). This ACPD-stimulated GFP synthesis was blocked byMCPG (Fig. 5, C and D). These results suggest that the activation ofgroup I and IImGluRs is sufficient to induceGFP synthesis in dendrites.However, in contrast to the stimulation effects of glutamate andNMDA(Figs. 2–4), ACPD-induced GFP synthesis did not reach a significantlevel at 15min compared to the shamcontrol (Fig. 5D). This observationsuggests that the kinetics of group I and II mGluR-activated GFP syn-
thesis was slower than that induced by NMDA receptor activation(Fig. 3C).
Requirement of the mTOR Signaling Pathway for NMDA Receptor- andmGluR-regulated GFP Synthesis in Dendrites—Previous studies revealedthe existence of themTOR signaling pathway at the synaptic region and itsactivation in a NMDA receptor-dependent manner (39, 40).We hypothe-sized that mTOR signaling mediates NMDA receptor-activated dendriteGFP synthesis. To test this hypothesis, we used rapamycin, a specific inhib-itor of the mTOR kinase, to disrupt this signaling pathway. We observedthat rapamycin (200nM) completely blockedNMDA-inducedGFPsynthe-sis indendrites (Fig. 6,AandB).Ontheotherhand, ascomycin, a rapamycinanalog that does not inhibitmTOR, did not have an effect (Fig. 6,A andB).To verify that rapamycin inhibited NMDA-induced dendrite GFP synthe-
FIGURE 5. Involvement of group I and II mGluRs in regulation of dendritic protein synthesis. A, experimental dendrites for determining the effect of MCPG (200 �M) onglutamate-induced dendritic protein synthesis. B, summary of GFP changes in distal dendrites of A. C, experimental dendrites for determining the effect of ACPD (50 �M) on dendriticprotein synthesis. D, summary of GFP changes in distal dendrites of C. Bar, 10 �M.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
18808 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from
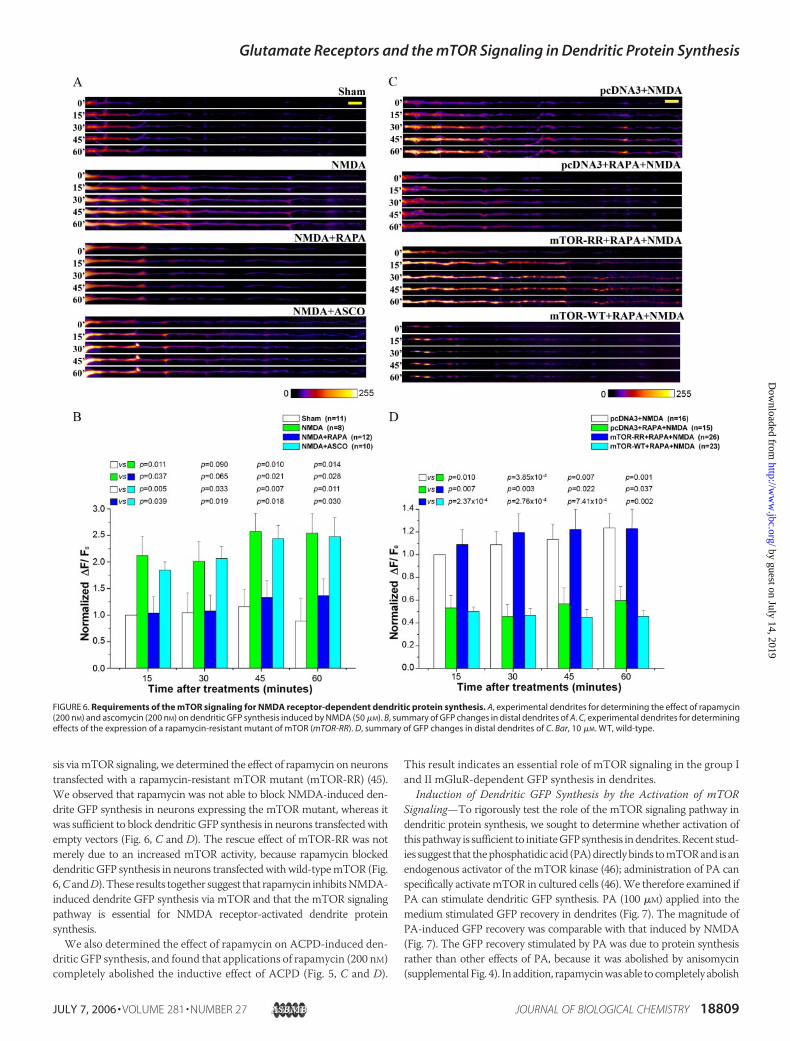
sis viamTOR signaling, we determined the effect of rapamycin on neuronstransfected with a rapamycin-resistant mTOR mutant (mTOR-RR) (45).We observed that rapamycin was not able to block NMDA-induced den-drite GFP synthesis in neurons expressing the mTOR mutant, whereas itwas sufficient to block dendritic GFP synthesis in neurons transfectedwithempty vectors (Fig. 6, C and D). The rescue effect of mTOR-RR was notmerely due to an increased mTOR activity, because rapamycin blockeddendritic GFP synthesis in neurons transfectedwithwild-typemTOR (Fig.6,CandD).These results together suggest that rapamycin inhibitsNMDA-induced dendrite GFP synthesis via mTOR and that the mTOR signalingpathway is essential for NMDA receptor-activated dendrite proteinsynthesis.We also determined the effect of rapamycin on ACPD-induced den-
dritic GFP synthesis, and found that applications of rapamycin (200 nM)completely abolished the inductive effect of ACPD (Fig. 5, C and D).
This result indicates an essential role of mTOR signaling in the group Iand II mGluR-dependent GFP synthesis in dendrites.
Induction of Dendritic GFP Synthesis by the Activation of mTORSignaling—To rigorously test the role of the mTOR signaling pathway indendritic protein synthesis, we sought to determine whether activation ofthispathway is sufficient to initiateGFPsynthesis indendrites.Recent stud-ies suggest that thephosphatidicacid (PA)directlybinds tomTORand isanendogenous activator of the mTOR kinase (46); administration of PA canspecifically activatemTOR in cultured cells (46).We therefore examined ifPA can stimulate dendritic GFP synthesis. PA (100 �M) applied into themedium stimulated GFP recovery in dendrites (Fig. 7). The magnitude ofPA-induced GFP recovery was comparable with that induced by NMDA(Fig. 7). The GFP recovery stimulated by PA was due to protein synthesisrather than other effects of PA, because it was abolished by anisomycin(supplementalFig. 4). Inaddition, rapamycinwasable tocompletely abolish
FIGURE 6. Requirements of the mTOR signaling for NMDA receptor-dependent dendritic protein synthesis. A, experimental dendrites for determining the effect of rapamycin(200 nM) and ascomycin (200 nM) on dendritic GFP synthesis induced by NMDA (50 �M). B, summary of GFP changes in distal dendrites of A. C, experimental dendrites for determiningeffects of the expression of a rapamycin-resistant mutant of mTOR (mTOR-RR). D, summary of GFP changes in distal dendrites of C. Bar, 10 �M. WT, wild-type.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18809
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

PA-induced GFP synthesis (Fig. 7), indicating that PA stimulated dendriteprotein synthesis viamTOR.These results suggest that activationofmTORsignaling is sufficient to induce GFP synthesis in dendrites.To assess if NMDA receptors and mTOR use a common mecha-
nism to activate GFP synthesis in dendrites, we compared the effects
of NMDA, PA, or both together. We reasoned that, if NMDA recep-tors and mTOR use different mechanisms to activate dendrite pro-tein synthesis, treatments with both NMDA and PA may lead to anadditive stimulation effect on dendritic GFP synthesis than that fromeither treatment alone. On the other hand, if NMDA receptors andmTOR employ a commonmechanism to regulate GFP synthesis, onewould expect that treatments with both NMDA and PA may notcause an additive stimulation effect compared with that from indi-vidual treatments. The result indicated that the magnitudes of stim-ulated dendritic GFP synthesis from NMDA, PA, and both togetherwere not significantly different from each other (Fig. 7). These obser-vations are consistent with the idea that NMDA receptors andmTOR signaling are in the same pathway to control dendritic GFPsynthesis. However, we cannot exclude the possibility of the ceilingeffect from single stimulation.
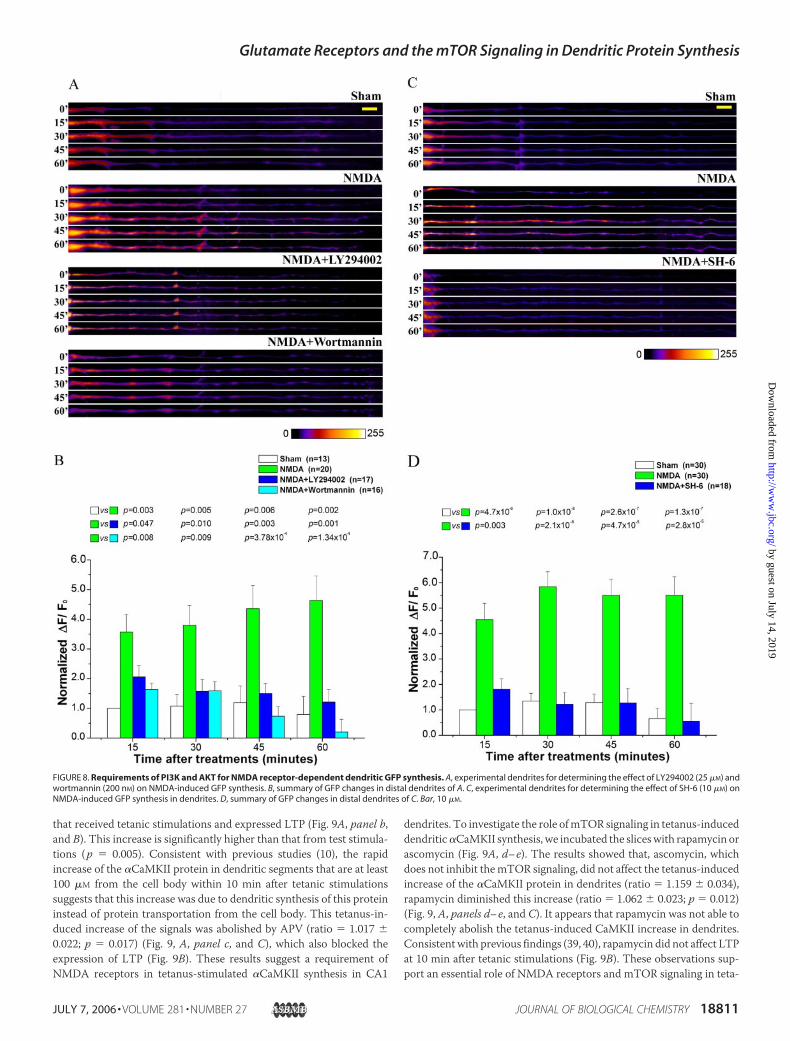
Involvement of the PI3K/AKT Signaling in NMDA Receptor-dependentDendrite GFP Synthesis—The fact that the activation of themTOR signal-ing pathway is not only required for NMDA receptor-dependent den-dritic protein synthesis but also sufficient to initiate this process suggeststhat the activationofNMDAreceptors activates themTORsignalingpath-way that then induces dendritic GFP synthesis. To understand themecha-nism that links NMDA receptors to the activation of mTOR signaling, wetested if PI3K, which is considered an upstream regulator ofmTOR signal-ing (47), is required for NMDA-induced dendritic synthesis of GFP.Toward this end, we used two selective PI3K inhibitors, wortmannin andLY294002. We observed that both inhibitors were able to block NMDA-induced GFP synthesis in dendrites (Fig. 8, A and B). Wortmannin causedan appreciable reduction of GFP recovery in NMDA-stimulated neuronsbelow that of control neurons at late stages (Fig. 8B), indicating a role ofPI3K in basal dendritic protein synthesis. Indeed, wortmannin treatmentled to a consistent but insignificant (p� 0.05) reduction ofGFP recovery inneurons not stimulatedwithNMDA (supplemental Fig. 5). In addition, wealso tested the potential role of AKT, another mTOR upstream activator(47).We found that the AKT selective antagonist SH-6 abolishedNMDA-inducedGFP synthesis in dendrites (Fig. 8,C andD). These results supportan essential role of PI3KandAKT inNMDAreceptor-dependent dendriticGFP synthesis.
Requirement of NMDA Receptors and the mTOR Signaling for Teta-nus-induced Dendrite Synthesis of the �CaMKII Protein in Hippocam-pal CA1—Results from the above experiments suggest an essential roleof the mTOR signaling pathway in NMDA receptor- and mGluR-de-pendent dendritic translation of the GFP mRNA controlled by�CaMKII UTRs. To investigate if this signaling is required for the den-dritic synthesis of endogenous �CaMKII, we sought to characterize theeffect of disruption of mTOR signaling on tetanus-induced �CaMKIIsynthesis in dendrites. Toward this end, we performed fluorescentimmunostaining experiments to reveal the �CaMKII protein on hip-pocampal slices. We introduced a microlesion perpendicular to the cellbody layer to divide the CA1 field of slices (Fig. 9A). No electric stimu-lation was applied on the side that was used as an internal control; theother side received either test stimulations (Fig. 9A, a) or tetanic stim-ulations (Fig. 9A, b–e) to induce LTP (Fig. 9B). The presence ofmicrole-sions should prevent potential spreading of electric stimulation intocontrol sides. Slices were fixed for immunostaining 10 min after stimu-lations. Fluorescent signals in a CA1 dendritic region (100 � 100 �M)100�Maway from the cell body layer and the edge ofmicrolesions in thecontrol and stimulated sides were quantified, and the signal ratiobetween the stimulated side and control side was determined (Fig. 9C).This ratio in slices that received test stimulations (Fig. 9A, a) is 0.978 �0.037. On the other hand, this ratio increased to 1.132 � 0.033 in slices
FIGURE 7. PA-mediated activation of the mTOR signaling results in GFP synthesis indendrites. A, experimental dendrites for determining the effect of PA (100 �M) on den-dritic GFP synthesis. Bar, 10 �M. B, summary of GFP changes in distal dendrites.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
18810 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from
that received tetanic stimulations and expressed LTP (Fig. 9A, panel b,and B). This increase is significantly higher than that from test stimula-tions (p � 0.005). Consistent with previous studies (10), the rapidincrease of the �CaMKII protein in dendritic segments that are at least100 �M from the cell body within 10 min after tetanic stimulationssuggests that this increase was due to dendritic synthesis of this proteininstead of protein transportation from the cell body. This tetanus-in-duced increase of the signals was abolished by APV (ratio � 1.017 �0.022; p � 0.017) (Fig. 9, A, panel c, and C), which also blocked theexpression of LTP (Fig. 9B). These results suggest a requirement ofNMDA receptors in tetanus-stimulated �CaMKII synthesis in CA1
dendrites. To investigate the role ofmTOR signaling in tetanus-induceddendritic�CaMKII synthesis, we incubated the sliceswith rapamycin orascomycin (Fig. 9A, d–e). The results showed that, ascomycin, whichdoes not inhibit themTOR signaling, did not affect the tetanus-inducedincrease of the �CaMKII protein in dendrites (ratio � 1.159 � 0.034),rapamycin diminished this increase (ratio � 1.062 � 0.023; p � 0.012)(Fig. 9, A, panels d–e, and C). It appears that rapamycin was not able tocompletely abolish the tetanus-induced CaMKII increase in dendrites.Consistentwith previous findings (39, 40), rapamycin did not affect LTPat 10 min after tetanic stimulations (Fig. 9B). These observations sup-port an essential role of NMDA receptors and mTOR signaling in teta-
FIGURE 8. Requirements of PI3K and AKT for NMDA receptor-dependent dendritic GFP synthesis. A, experimental dendrites for determining the effect of LY294002 (25 �M) andwortmannin (200 nM) on NMDA-induced GFP synthesis. B, summary of GFP changes in distal dendrites of A. C, experimental dendrites for determining the effect of SH-6 (10 �M) onNMDA-induced GFP synthesis in dendrites. D, summary of GFP changes in distal dendrites of C. Bar, 10 �M.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18811
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

nus-induced dendritic synthesis of the �CaMKII protein. In addition,rapamycin also blocked tetanus-induced dendritic synthesis of MAP2(supplemental Fig. 6).
DISCUSSION
We used the CaMKII-GFP reporter (21) to investigate the regulatorymechanisms underlying activity-dependent protein synthesis in den-drites of cultured neurons. Several lines of evidence suggest that theactivity-induced GFP recovery we observed in photobleached dendriticsegments resulted from local protein synthesis rather than other cellularprocesses such as mRNA and protein transport from cell bodies. First,two different translational inhibitors, anisomycin and emetine, blockedNMDA-induced GFP recovery in photobleached dendrites (Fig. 3 andsupplemental Fig. 1). These results directly show that protein synthesisin response to NMDAR activation accounts for the observed GFPrecovery in photobleached dendrites. Second, disruption of cytoskel-etons by cytochalasin B and nocodazole did not block NMDA-induceddendritic GFP recovery (supplemental Fig. 2), indicating that cytoskel-eton-based active cellular transport is not responsible for the GFPrecovery. Third, GFP recovery in distal dendrites at least 100 �M awayfrom the soma approached plateaus 15 min after KCl, glutamate, orNMDA stimulation (Figs. 1–4). The simplest explanation for this rapidrecovery is dendritic synthesis of GFP, instead of somatic transport. TheCaMKII 3� UTR in this reporter construct contains cis-elements fordendriticmRNA targeting that is regulated by neuronal activity (48, 49).One possibility is that activity-induced dendritic RNA targeting mightcontribute to the enhanced GFP recovery in photobleached dendrites.Although we cannot conclusively exclude this possibility, our resultsoutlined above strongly suggest that the activity-induced GFP recoverymainly resulted from dendritic protein synthesis. In addition, previousstudies have shown that RNA targeting in cultured neurons is mediatedby microtubule but not actin filaments (50–52). However, bothcytochalasin B and nocodazole, which disrupt actin filaments andmicrotubules, respectively, were not able to abolish NMDA-inducedGFP recovery, with no significant differences in their effects (supple-mental Fig. 2). If dendritic RNA targeting accounted for the observedGFP recovery in dendrites, one should expect that nocodazole but notcytochalasin B would impair NMDA-induced GFP recovery.
Roles of Glutamate Receptors in the Regulation of Dendrite ProteinSynthesis—We systematically investigated the role of different classes ofglutamate receptors in regulation of dendrite protein synthesis in cul-tured hippocampal neurons, using their selective agonists and antago-nists. We found that APV abolished the dendrite protein synthesis acti-vated by KCl-mediated depolarization and glutamate applications (Figs.1 and 2), indicating that activation of NMDA receptors is necessary forprotein synthesis in dendrites induced by these stimulations. Theseobservations are consistent with the findings of the requirement ofNMDA receptors for dendrite �CaMKII synthesis in the hippocampusinduced by synaptic stimulations (Fig. 9) (10, 11). We also showed thatstimulations of NMDA receptors resulted in dendritic synthesis of GFP(Fig. 3), suggesting the activation of this class of glutamate receptors issufficient to initiate mRNA translation in dendrites. As NMDA recep-tors are important calcium channels at the synapse, calcium influxthrough NMDA receptor channels is likely critical to activate mRNAtranslation. Given the important role of NMDA receptors and dendritic�CaMKII synthesis in LTP (9, 53), NMDA receptor-regulated dendritic�CaMKII synthesis may be an essential part of molecular processesinvolved in LTP expression.Whereas the AMPA receptor antagonist CNQX blocked glutamate-
induced dendrite GFP synthesis (Fig. 4), the agonist AMPA inhibited
GFP synthesis in dendrites (supplemental Fig. 3). These observationssuggest that the activation of AMPA receptors is necessary for gluta-mate-stimulated protein synthesis in dendrites but not sufficient to acti-vate the translational process by itself. As AMPA receptors play impor-tant roles in gating the activation of NMDA receptors (44), theregulatory role of AMPA receptors in dendrite protein synthesis is likelymediated by NMDA receptors. Why stimulation of AMPA receptorsinhibits dendritic protein synthesis is an interesting problem thatremains to be investigated.We also observed that blocking group I and II mGluRs with MCPG
abolished glutamate-induced GFP synthesis in dendrites and that acti-vating these classes of glutamate receptors with ACPD led to dendriteGFP synthesis (Fig. 5). These findings are consistent with previous stud-ies showing that activation of mGluRs leads to association of multiplepotential translational regulatory kinases with ribosomes (54), proteinsynthesis in synaptoneurosome preparations (15), and translation oftransfected GFP mRNAs in severed hippocampal dendrites (17).Because activation of group I mGluRs is involved in expression of proteinsynthesis-dependent formsof synaptic plasticity, includingLTD (6, 42) andLTP (55), mGluR-dependent dendritic protein synthesis likely plays animportant role for expression of these forms of synaptic plasticity. It wouldbe interesting to investigate the potential functional interactions betweenmGluRs and other regulatory factors such as NMDARs and BDNF duringdendritic protein synthesis.Our results indicate that the excitatory neurotransmitter glutamate
can activate dendritic protein synthesis through distinct subtypes ofglutamate receptors. However, the fact that NMDA, AMPA, and mGlureceptors are all required for glutamate-stimulated dendritic proteinsynthesis suggests a coordinated mechanism. How different classes ofglutamate receptors coordinate translational activation is a significantproblem that remains to be addressed. As numerous previous studiesindicated regulatory roles of the AMPA and mGlu receptors on thefunction of NMDA receptors during synaptic plasticity, one possibilityis that the activities of NMDA receptors in regulation of dendrite pro-tein synthesis are modulated by AMPA and mGlu receptors. On theother hand, both NMDA and mGluRs are sufficient to initiate GFPsynthesis in dendrites (Figs. 3 and 5). Because activation of NMDAreceptors and group I mGluRs leads to a local Ca2� increase by Ca2�
influx and intracellular release, respectively, Ca2� signals likely mediatethe stimulation effects of these two classes of glutamate receptors ondendritic GFP synthesis. Furthermore, as mTOR signaling is requiredfor both NMDA- and mGluR-regulated dendritic GFP synthesis, thetranslational regulatory processes initiated by these glutamate receptorsare at least partially overlapped.A recent study showed that miniature activities mediated by NMDA
and AMPA glutamate receptors play an inhibitory role on protein syn-thesis in dendrites (26). However, we observed that both NMDA andAMPA receptors are required for glutamate-activated dendritic GFPsynthesis and that NMDA receptor activation is sufficient to initiateGFP synthesis in dendrites (Figs. 2–4). It is intriguing that the sametypes of glutamate receptors display opposite effects on dendritic pro-tein synthesis in different experimental paradigms. These oppositeeffects may be due to the difference in the amount of NMDA receptoractivities and the dynamics of NMDA receptor activation. For example,whereas bath applications of glutamate and NMDA are expected toresult in the activation of a large number of NMDA receptors and thusa big Ca2� influx, spontaneous glutamate releases from presynaptic ter-minals can only open a limited number of NMDA receptor channels,which leads to a small Ca2� influx. A large NMDA receptor-mediatedCa2� influxmay activate the positive regulator of translational signaling,
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
18812 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

FIGURE 9. Requirements of NMDA receptors and mTOR signaling for tetanus-induced dendritic synthesis of the �CaMKII protein in hippocampal slices. A, representativemouse hippocampal slices after fluorescent immunostaining. Microlesions (indicated by asterisks) were introduced to divide the CA1 field into two parts. a, test stimulations; b, tetanicstimulations; c, tetanic stimulations and APV applications; d, tetanic stimulations and ascomycin applications; e, tetanic stimulations and rapamycin applications. Circles, position ofstimulation electrodes; boxes, ROI in CA1 dendrites for signal quantifications; arrowheads, CA1 cell body layers. Bar, 100 �M. A color scale for signal intensities is provided under thefluorescent graphs. B, averaged synaptic responses between 8 and 10 min after baseline or tetanic stimulations. LTP was induced by tetanus and tetanus with ascomycin orrapamycin, but not by tetanus with APV. C, summary of signal ratios (mean � S.E.) between the stimulated side and unstimulated side under different conditions. At least 20 slicesfrom 5 mice were used for each experiment. One-way analysis of variance reveals significant differences among the groups (F(4,159) � 5.84: p 0.01); p values for individual pairs ofcomparison determined by a LSD (least-significant difference) post hoc test are indicated.
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18813
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

whereas a small Ca2� influx may activate negative translational regula-tors. Consistent with this notion, it is known that the magnitude anddynamics of postsynaptic Ca2� signals play a crucial role in modulatingactivities of specific kinases and phosphatases during LTP and LTDinduction (56).
mTOR Signaling and Synaptic Activity-regulated Dendritic ProteinSynthesis—Howdoes the activation of glutamate receptors lead to proteinsynthesis in dendrites? It is conceivable that glutamate receptor activationinitiates signaling cascades that control translational initiation.We specifi-cally tested the role of the mTOR signaling pathway. Previous studiesrevealed the localization ofmultiple components in this translational regu-latory pathway at the postsynaptic region (39). The synaptic localization ofthis signaling pathway makes it well suited to couple the activation of glu-tamate receptors with local translation initiation.We observed that inhibi-tion of mTOR activities with rapamycin blocked dendritic GFP synthesisinduced by stimulations of NMDA receptors and mGluRs (Figs. 5 and 6).The inhibitory effect of rapamycin ondendriticGFP synthesiswas depend-ent on themTOR kinase, because a rapamycin-resistantmutant ofmTORwas able to suppress the rapamycin-mediated inhibition (Fig. 6). In addi-tion, inhibitions of the mTOR upstream activators, PI3K and AKT, alsoabolishedNMDAreceptor-regulatedprotein synthesis indendrites (Fig. 8).These observations are consistent with previous findings that synapticstimulations activate a downstream effector, p70S6K (40). Our findingsprovidedirect evidence foranessential roleof themTORsignalingpathwayin control of glutamate receptor-dependent protein synthesis in dendritesof living neurons. These results support the idea that the activation of glu-tamate receptors activates the mTOR signaling pathway that then leads totranslational activation in dendrites. Consistent with this notion, we foundthat applications of PA, an endogenous mTOR activator, activated den-dritic protein synthesis (Fig. 7), and that the PA-mediated activation ofdendrite protein synthesis was abolished by rapamycin (Fig. 7).The mTOR signaling pathway is implicated in synaptic plasticity,
including LTP and LTD (5, 39, 40, 42). Therefore, mTOR signalingregulated-dendritic protein synthesis may directly contribute to theseforms of synaptic plasticity. This notion is consistent with our observa-tions of rapid dendritic protein synthesis in response to glutamate andtetanic stimulations (Figs. 2, 4, 5, and 9); the rapid protein synthesis islikely required for the expression of L-LTP as rapamycin applicationsduring tetanic stimulations but not 5 min after induction impairedL-LTP (40). What are the proteins that are synthesized in dendrites,regulated by synaptic activity and the mTOR signaling pathway, andcontribute to synaptic plasticity? Our results indicate that �CaMKII, aprotein that is enriched at the synaptic region and critical for synapticplasticity (57, 58), is one of them. In support of this idea, �CaMKIIprotein is rapidly induced in CA1 dendrites after LTP-inducing tetanicstimulations (Fig. 9) (10). More importantly, dendritic synthesis of the�CaMKII protein is required for the expression of L-LTP (9). Othersynaptic activity and mTOR signaling-regulated targets include MAP2(supplemental Fig. 6) and eEF1A (59). A comprehensive characteriza-tion of the dendritically synthesized target proteins of mTOR signalingmay significantly improve our understanding of the molecular basis oflong-lasting synaptic plasticity.In summary, our results suggest that NMDA receptors activate pro-
tein synthesis in dendrites viamTOR signaling. However, it is likely thatthis NMDA receptor-mTOR signaling pathway interacts with othertranslational signaling cascades to coordinate dendritic protein synthe-sis. For example, previous studies indicated a role of cytoplasmic poly-adenylation in regulation of activity-dependent �CaMKII protein syn-thesis in neurons (32–34). During translational activation, poly(A) tailsinteract with the translational initiation complex that is regulated by the
mTOR signaling (60). Therefore, mTOR signaling and cytoplasmic pol-yadenylation may cooperatively regulate �CaMKII synthesis in den-drites. In addition, inhibition of extracellular signal-regulated kinase(ERK) activation blocked the phosphorylation of downstream proteinsin the mTOR signaling pathway, including 4E-BP1 and ribosomal pro-tein S6 (37), suggesting a cross-talk between the mTOR and MAPKsignaling pathways. Understanding the cross-talk between the NMDAreceptor-mTOR and other translational signaling pathways during den-dritic protein synthesis is a significant problem to be addressed.
Acknowledgments—We are very grateful to Drs. Erin Schuman and ChangangJiang for theGFP reporter, Dr. Jie Chen for rapamycin-resistantmTORmutantplasmids, and Dr. Schuman for comments on the manuscript.
REFERENCES1. Kandel, E. R. (2001) Science 294, 1030–10382. Steward, O., and Schuman, E. M. (2001) Annu. Rev. Neurosci. 24, 299–3253. Kang, H., and Schuman, E. M. (1996) Science 273, 1402–14064. Bradshaw, K. D., Emptage, N. J., and Bliss, T. V. P. (2003) Eur. J. Neurosci. 18,
3150–31525. Cracco, J. B., Serrano, P., Moskowitz, S. I., Bergold, P. J., and Sacktor, T. C. (2005)
Hippocampus 15, 551–5566. Huber, K. M., Kayser, M. S., and Bear, M. F. (2000) Science 288, 1254–12577. Martin, K. C., Casadio, A., Zhu, H., E, Y., Rose, J. C., Chen, M., Bailey, C. H., and
Kandel, E. R. (1997) Cell 91, 927–9388. Sherff, C. M., and Carew, T. J. (1999) Science 285, 1911–19149. Miller, S., Yasuda, M., Coats, J. K., Jones, Y., Martone, M. E., and Mayford, M. (2002)
Neuron 36, 507–51910. Ouyang, Y., Rosenstein, A., Kreiman, G., Schuman, E. M., and Kennedy, M. B. (1999)
J. Neurosci. 19, 7823–783311. Steward, O., and Halpain, S. (1999) J. Neurosci. 19, 7834–784512. Steward, O., Wallace, C. S., Lyford, G. L., and Worley, P. F. (1998) Neuron 21,
741–75113. Steward, O., and Worley, P. E. (2001) Neuron 30, 227–24014. Ostroff, L. E., Fiala, J. C., Allwardt, B., and Harris, K. M. (2002) Neuron 35, 535–54415. Weiler, I. J., and Greenough, W. T. (1993) Proc. Natl. Acad. Sci. U. S. A. 90,
7168–717116. Kacharmina, J. E., Job, C., Crino, P., and Eberwine, J. (2000) Proc. Natl. Acad. Sci.
U. S. A. 97, 11545–1155017. Job, C., and Eberwine, J. (2001) Proc. Natl. Acad. Sci. U. S. A. 98, 13037–1304218. Huang, F., Chotiner, J. K., and Steward, O. (2005) J. Neurosci. 25, 7199–720919. Scheetz, A. J., Narin, A. C., and Constantine-Paton, M. (2000) Nature Neurosci. 3,
211–21620. Smith, W. B., Starck, S. R., Roberts, R. W., and Schuman, E. M. (2005) Neuron 45,
765–77921. Aakalu, G., Smith,W. B., Nguyen,N., Jiang, C., and Schuman, E.M. (2001)Neuron 30,
489–50222. Yin, Y., Edelman, G. M., and Vanderklish, P. W. (2002) Proc. Natl. Acad. Sci. U. S. A.
99, 2368–237323. Smart, F. M., Edelman, G. M., and Vanderklish, P. W. (2003) Proc. Natl. Acad. Sci.
U. S. A. 100, 14403–1440824. McCarthy, J. B., and Milner, T. A. (2003) Neuroreport 14, 1357–136025. Schratt, G. M., Nigh, E. A., Chen, W. G., Hu, L., and Greenberg, M. E. (2004) J. Neu-
rosci. 24, 7366–737726. Sutton, M. A., Wall, N. R., Aakalu, G. N., and Schuman, E. M. (2004) Science 304,
1979–198327. Zalfa, F., Giorgi, M., Primerano, B., Moro, A., Di Penta, A., Reis, S., Oostra, B., and
Bagni, C. (2003) Cell 112, 317–32728. Shiina, N., Shinkura, K., and Tokunaga, M. (2005) J. Neurosci. 25, 4420–443429. Menon, K. P., Sanyal, S., Habara, Y., Sanchez, R.,Wharton, R. P., Ramaswami,M., and
Zinn, K. (2004) Neuron 44, 663–67630. Ye, B., Petritsch, C., Clark, I. E., Gavis, E. R., Jan, L. Y., and Jan, Y. N. (2004)Curr. Biol.
14, 314–32131. Wang, H., Iacoangeli, A., Popp, S., Muslimov, I. A., Imataka, H., Sonenberg, N., Lo-
makin, I. B., and Tiedge, H. (2002) J. Neurosci. 22, 10232–1024132. Wu, L.,Wells, D., Tay, J., Mendis, D., Abbott, M.-A., Barnitt, A., Quinlan, E., Heynen,
A., Fallon, J. R., and Richter, J. D. (1998) Neuron 21, 1129–113933. Huang, Y. S., Jung, M. Y., Sarkissian, M., and Richter, J. D. (2002) EMBO J. 21,
2139–214834. Wells, D. G., Dong, X., Quinlan, E. M., Huang, Y. S., Bear, M. F., Richter, J. D., and
Fallon, J. R. (2001) J. Neuronsci. 21, 9541–9548
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
18814 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

35. Atkins, C. M., Nozaki, N., Shigeri, Y., and Soderling, T. R. (2004) J. Neurosci. 24,5193–5201
36. Banko, J. L., Hou, L., and Klann, E. (2004) J. Neurochem. 91, 462–47037. Kelleher, R. J., III, Govindarajan, A., Jung, H. Y., Kang, H., and Tonegawa, S. (2004)
Cell 116, 467–47938. Raught, B., Gingras, A.-C., and Sonenberg, N. (2001) Proc. Natl. Acad. Sci. U. S. A. 98,
7037–704439. Tang, S. J., Reis, G., Kang, H., Gingras, A. C., Sonenberg, N., and Schuman, E. M.
(2002) Proc. Natl. Acad. Sci. U. S. A. 99, 467–47240. Cammalleri,M., Lutjens, R., Berton, F., King, A. R., Simpson, C., Francesconi,W., and
Sanna, P. P. (2003) Proc. Natl. Acad. Sci. U. S. A. 100, 14368–1437341. Lenz, G., and Avruch, J. (2005) J. Biol. Chem. 280, 38121–3812442. Hou, L., and Klann, E. (2004) J. Neurosci. 24, 6352–636143. Tischmeyer, W., Schicknick, H., Kraus, M., Seidenbecher, C. I., Staak, S., Scheich, H.,
and Gundelfinger, E. D. (2003) Eur. J. Neurosci. 18, 942–95044. Malenka, R. C., and Bear, M. F. (2004) Neuron 44, 5–2145. Vilella-Bach, M., Nuzzi, P., Fang, Y., and Chen, J. (1999) J. Biol. Chem. 274,
4266–427246. Fang, Y., Vilella-Bach, M., Bachmann, R., Flanigan, A., and Chen, J. (2001) Science
294, 1942–194547. Hay, N., and Sonenberg, N. (2004) Genes Dev. 18, 1926–194548. Mayford, M., Baranes, D., Podsypanina, K., and Kandel, E. R. (1996) Proc. Natl. Acad.
Sci. U. S. A. 93, 13250–1325549. Mori, Y., Imaizumi, K., Katayama, T., Yoneda, T., and Tohyama, M. (2000) Nature
Neurosci. 3, 1079–108450. Bassell, G., Singer, R. H., and Kosik, K. (1994) Neuron 12, 571–58251. Knowles, R. B., Sabry, J. H., Martone, M. E., Deerinck, T. J., Ellisman, M. H., Bassell,
G. J., and Kosik, K. S. (1996) J. Neurosci. 16, 7812–782052. Tang, S. J., Meulemans, D., Vazquez, L., Colaco, N., and Schuman, E. (2001) Neuron
32, 463–47553. Bliss, T. V. P., and Collingridge, G. L. (1993) Nature 361, 31–3954. Angenstein, F., Greenough, W. T., and Weiler, I. J. (1998) Proc. Natl. Acad. Sci.
U. S. A. 95, 15078–1508355. Raymond, C. R., Thompson, V. A., Tate, W. R., and Abraham,W. C. (2000) J. Neuro-
sci. 20, 969–97656. Bear, M. F., and Linden, D. J. (2001) in Synapses (Cowan, W. M., Sudhof, T. C.,
Stevens, C., and Davies, K., eds) pp. 455–517, The Johns Hopkins University Press,Baltimore
57. Silva, A. J., Stevens, C. F., Tonegawa, S., and Wang, Y. (1992) Science 257, 201–20658. Lisman, J., Schulman, H., and Cline, H. (2002) Nat. Rev. Neurosci. 3, 175–19059. Tsokas, P., Grace, E. A., Chan, P.,Ma, T., Sealfon, S. C., Iyengar, R., Landau, E.M., and
Blitzer, R. D. (2005) J. Neurosci. 25, 5833–584360. Sachs, A. (2000) in Translational Control of Gene Expression (Sonenberg, N., Her-
shey, J. W. B., andMathews, M. B., eds) pp. 447–465, Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, NY
61. Goslin, K., Asmussen, H., and Banker, G. (1998) in Culturing Nerve Cells (Banker, G.,Goslin, K., eds) pp. 339–370, MIT Press, Cambridge, MA
62. Malgaroli, A., and Tsien, R. W. (1992) Nature 357, 134–139
Glutamate Receptors and the mTOR Signaling in Dendritic Protein Synthesis
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18815
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from

Ruomu Gong, Chang Sin Park, Nima Rezaei Abbassi and Shao-Jun TangHippocampal Neurons
Signaling Pathway in Activity-dependent Dendritic Protein Synthesis in Roles of Glutamate Receptors and the Mammalian Target of Rapamycin (mTOR)
doi: 10.1074/jbc.M512524200 originally published online May 1, 20062006, 281:18802-18815.J. Biol. Chem.
10.1074/jbc.M512524200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2006/05/02/M512524200.DC1
http://www.jbc.org/content/281/27/18802.full.html#ref-list-1
This article cites 59 references, 32 of which can be accessed free at
by guest on July 14, 2019http://w
ww
.jbc.org/D
ownloaded from










![Targeting of PI3K/AKT/mTOR pathway to inhibit T cell activation … · 2017. 8. 25. · AKT/mammalian target of rapamycin (PI3K/AKT/ mTOR) [1]. This pathway controls numerous cellular](https://img.dokumen.tips/doc/110x75/60af5eaa6ab71f4bc15363aa/targeting-of-pi3kaktmtor-pathway-to-inhibit-t-cell-activation-2017-8-25-aktmammalian.jpg)



![Effects of rapamycin and curcumin on inflammation and ......(mTOR) pathway has been studied as a possible target for anti-epileptogenic strategies [5–7]. The mTOR path-way regulates](https://img.dokumen.tips/doc/110x75/6098f8d4914d1836e3143c4d/effects-of-rapamycin-and-curcumin-on-inflammation-and-mtor-pathway-has.jpg)














