Embed Size (px)
Citation preview

Journal of Pined Research 7:13-22 (1989)
Role of Postsynaptic a-Adrenergic Receptors in the p-Adrenergic Stimulation of
Melatonin Production in the Syrian Hamster Pineal Gland in Organ Culture
Celsa Santana, Juan M. Guerreto, Russel J. Reiter, and Armando Menendez-Pelaez
Department of Cellular and Structural Biology, University of Texas Health Science Center at San Antonio, San Antonio, Texas
The role played by postsynaptic a-adrenergic receptors in the stimulation of pineal melatonin production was investigated in the Syrian hamster. The studies were conducted using organ cultured pineal glands collected from both anatomically intact and superior cervical ganglionectomized hamsters. Results obtained indicate that phenylephrine, an a-adrenergic agonist, by itself has no effect in promoting melatonin production; however, it potentiates the stimulatory effects of isoproterenol, a P-adrenergic agonist, on pineal melatonin production in nonoperated hamsters. Similar observations were obtained with pineal glands whose presynaptic terminals were removed by prior superior cervical ganglionectomy. However, a longer incubation time was required (4-6 hours vs. 2 hours) with pineal glands taken from ganglionectomized animals. Apparently, P-adrenergic activation is an absolute re- quirement to stimulate pineal melatonin production, and an a-adrenergic receptor mechanism potentiates P-adrenergic activation. In addition, the findings obtained with denervated pineal glands suggest that the regulation of pineal melatonin production by both a- and P-adrenergic mechanisms is through receptors located on postsynaptic structures.
Key words: Syrian hamster, isoproterenol, phenylephrine, superior cervical ganglionectomy
INTRODUCTION
Extensive studies by several groups of investigators have demonstrated that in the rat the biochemical events involved in mediating the induction of N-acetyltransferase (NAT) activity are under P-adrenergic control [ Klein and
Received July 15, 1988; accepted October 12, 1988.
Address reprint requests to Russel J. Reiter, Department of Cellular and Structural Biology, The University of Texas Health Science Center at San Antonio, 7703 Floyd Curl Drive, San Antonio, TX 782847762,
0 1989 Alan R. Liss, Inc.

14 Santanaetal.
Meller, 1970; Deguchi and Axelrod, 1972b; Brownstein et al., 19731. Further- more, the administration of either norepinephrine (NE) or isoproterenol (ISO), a (3-adrenergic agonist, readily stimulates production of melatonin within the rat pineal gland [Zatz, 1981; Ebadi, 19841. Furthermore, there is increasing evi. dence to suggest that the induction of rat pineal NAT activity is also modulated by a-adrenergic receptors. In both in vivo and in vitro experiments the a-agonist phenylephrine (PE) both potentiates and prolongs the effects of IS0 on NAT induction [Klein et al., 1983; Alphs and Lovenberg, 19841. These results suggest that rat pineal NAT activity and melatonin production are regulated by both a- and P-adrenergic receptors located on postsynaptic structures.
For a number of years it was thought that P-adrenergic receptors were not involved in mediating the nighttime rise in hamster pineal melatonin production [Binkley, 19761. However, in 1981 Lipton and colleagues found that the nighttime rise in pineal melatonin in the Syrian hamster is inhibited by the administration of propranolol, a P-adrenergic receptor blocker, just as in the rat. It was subsequently shown that the hamster pineal gland is capable of responding to either NE or I S 0 only during the night in both in vivo [Reiter et al., 1987a,c; Vaughan and Reiter, 19871 and in vitro [Vaughan et al., 1986; Santana et al., 19881 studies. This exclusive nighttime sensitivity of the Syrian hamster pineal to P-agonists disappears if animals are exposed to light during the normal dark time period [Reiter et al., 1987b,c].
No data are available concerning the modulatory effects of a-adrenergic agonists on the induction of Syrian hamster pineal melatonin production. For this reason, the role played by the a-adrenergic agonist, PE, on the in vitro stimulation of pineal melatonin production was investigated in the Syrian hamster. In order to determine whether the effects of PHE and IS0 on pineal melatonin production were due to interactions with either pre- or postsynaptic structures, experiments were done with pineals cultured immediately after excision from the hamster, thereby maintaining presynaptic elements associated with the pineal gland, and with cultured pineals whose presynaptic terminals were removed by prior superior cervical ganglionectomy.
MATERIALS AND METHODS Chemicals
The following drugs were used in these studies: ( - )-isoproterenol (ISO) and L-phenylephrine (PE). They were purchased from Sigma Chemical Co. (St. Louis). Other chemicals were obtained from other commercial sources.
Animals
Male Syrian hamsters, weighing between 80-100 gm at the time of the study, were purchased from Sasco, Omaha. Upon arrival in the laboratory they were caged (4-5 per cage) and given food and water ad libitum. They were exposed to an automatically regulated light-dark (LD) cycle of 1410 for a minimum of 1 week before being used for the experiments. Lights were turned off daily from 20.00 through 06.00 hr. Both anatomically intact and ganglion. ectomized animals were used in the experiments. Bilateral superior cervical

Hamster Pineal Gland in Vitro 15
ganglionectomies were done with the animals under ether anesthesia. Operated animals were used in drug studies 2-10 days after surgery to allow both for recovery from the surgery and for degeneration of sympathetic nerves. On the night of the experiments, intact or ganglionectomized animals were exposed to 50-70 pW/cm2 white light after 6 hr of darkness; 30 min later the animals were killed and their pineals were collected for organ culture studies. This irradiance and duration of light was used to suppress high nighttime melatonin levels to basal daytime values [Reiter, 1985I.
Culture Techniques
Hamster pineal glands were cultured using a method described previously by Brammer et al. [ 198 1 ] with some modifications. Two pineals were placed on a filter (HAWP 02500, Millipore Corp., Bedford, MA), which rested on top of a 1-ml well of pH 7.4 culture medium (Falcon organ culture dish 3037, Falcon Plastics, Los Angeles). The glands were incubated at 37°C under 95% O2-5% COz. The culture medium (BGJ,, Fitton-Jackson modification, Grand Island Biological Co., Grand Island, NY) was supplemented with ascorbic acid (0.1 rnglml), glutamine (0.2 mM), BSA fraction V (1 mg/ml), penicillin (100 units/ml), and streptomycin (100 pg/ml). In all in vitro studies, pineal glands were incubated for 30 min prior to treatment with the drugs I S 0 and PHE at the concentrations and times indicated. The drugs were dissolved in the culture medium. Both pineal glands and culture media were collected and quickly frozen on solid COz and stored at - 70°C until assayed for melatonin.
Assays and Statistical Analyses
Melatonin levels were estimated using the radioimmunoassay outlined by Rollag and Niswender [ 19761 and referred to either pg/gland or pg/ml/gland for gland and culture medium, respectively. The purpose of measuring melatonin levels in both pineal gland and culture medium was to study which of these best reflect changes in the response of the pineal gland to the adrenergic stimulation. Results are expressed as means -t standard errors (SEM). Data were statistically analyzed using an ANOVA followed by a Student-Newman-Keuls multiple range test.
RESULTS Effects of wand P-Adrenergic Agonists on Induction of Melatonin Production
The induction of pineal melatonin production by a- and P-adrenergic agonists was studied in pineals from anatomically intact hamsters. Pineals were incubated with either I S 0 (1 pM), PE (1 pM), or I S 0 plus PE at the same concentrations. The incubation time (2 hr) was chosen based on the previous results by Santana et al. [1988] who obtained a maximal response for ISO- induced melatonin production after 2 hr of incubation. As shown in Figure 1 (left panel), IS0 markedly increased the melatonin content in both pineal gland and culture media. PE, which alone did not affect melatonin levels, clearly potentiated the effect of IS0 in increasing the melatonin content in the pineal
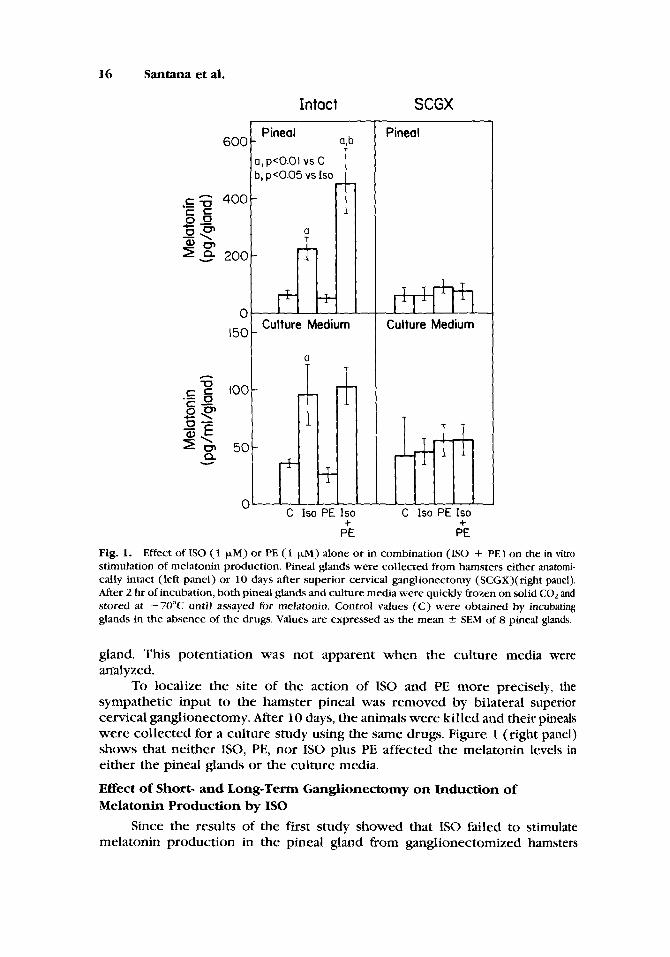
16
600
- 400- .c u c c 0 0 ,on s\, Z G 200-
0 - I50
Santana et al.
Pineal - a,b
a,p<O.Ol vs C b, p<0.05 vs Is0 --
- a
4-
Culture Medium -
Intact
100
50
SCGX
-
-
r' 0-
C 9
so PE Is0
PE +
Pineal
Culture Medium
C Is0 PE Is0
PE +
Fig. 1. Effect of I S 0 (1 pM) or PE (1 p,M) alone or in combination ( I S 0 + PE) on the in vitro stimulation of melatonin production. Pineal glands were collected from hamsters either anatomi- cally intact (left panel) or 10 days after superior cervical ganglionectomy (SCGX)(right panel). After 2 hr of incubation, both pineal glands and culture media were quickly frozen on solid COz and stored at -7OOC until assayed for melatonin. Control values (C) were obtained by incubating glands in the absence of the drugs. Values are expressed as the mean 2 SEM of 8 pineal glands.
gland. This potentiation was not apparent when the culture media were analyzed.
To localize the site of the action of I S 0 and PE more precisely, the sympathetic input to the hamster pineal was removed by bilateral superior cervical ganglionectomy. After 10 days, the animals were killed and their pineals were collected for a culture study using the same drugs. Figure 1 (right panel) shows that neither ISO, PE, nor I S 0 plus PE affected the melatonin levels in either the pineal glands or the culture media.
Effect of Short- and Long-Term Ganglionectomy on Induction of Melatonin Production by I S 0
Since the results of the first study showed that I S 0 failed to stimulate melatonin production in the pineal gland from ganglionectomized hamsters

Hamster Pineal Gland in Vitro 17
Pineal
a, p<O.OOI vs C 400
1 Cultuy Medium
200
50
n " c Is0 c Is0 c Is0 c Is0 - - - ~ 2 6 10
Intact Days After SCGX
Fig. 2. Effect of short- and long-term ganglionectomy on pineal melatonin production by I S 0 (1 kM). Pineal glands were collected from anatomically intact hamsters (INTACT) or at 2,6 , and 10 days after superior cervical ganglionectomy (SCGX). Pineal glands were incubated for 2 hr, and then both glands and culture media quickly frozen on solid C 0 2 and stored at - 70°C until assayed for melatonin. Control values (C) were obtained by incubating glands in the absence of the drug. Values are expressed as the mean k SEM of 8 pineal glands.
killed 10 days after surgery, the study was repeated at different times ( 2 , 6 , and 10 days) after superior cervical ganglionectomy. Again, as shown in Figure 2, IS0 was incapable of stimulating pineal melatonin content at the specified times. These findings suggested that, at least for the incubation time used (2 hr), IS0 was unable to stimulate melatonin production in the pineal gland of ganglion- ectomized Syrian hamsters.
Time-Course Study
To determine if the failure of I S 0 to stimulate pineal melatonin content in superior cervical ganglionectomized hamsters was due to a lag in the response to the P-adrenergic agonist, pineal glands, collected from ganglionectomized hamsters killed 10 days after surgery, were incubated with I S 0 (1 pM) and

18 Santanaeta l .
0 . I I I
200- Culture Medium b T
I I I I 1
0 2 4 6
Hours of Incubation Fig. 3. Time-course study of the in vitro stimulation of pineal and culture medium melatonin levels by I S 0 (1 pM)(O). All hamsters were superior cervical ganglionectomized 10 days before pineal collection, and then their pineals were incubated for the indicated times. Control values (0) were obtained by incubating glands in the absence of the drug. Values are expressed as the mean 5 SEM of 8 pineal glands.
assayed at different times ( 0 , 2 , 4 , or 6 hr) after the beginning of the incubation. The results again indicated that at 2 hr I S 0 did not promote pineal melatonin production; however, when pineal glands were incubated for either 4 or 6 hr, IS0 significantly increased melatonin levels in both pineal gland and culture media (Fig. 3); the increases were similar to those seen in pineal glands from anatomically intact hamsters that were incubated with I S 0 for 2 hr (Fig. 1, left panel).
Effect of Pineal Denemtion on Modulat ion of Melatonin Production by PE
Since the results reported above (Fig. 3) showed that I S 0 was able to stimulate pineal melatonin production when the incubation time was increased to 4-6 hr, in the final experiment the effect of PE on IS0 stimulation of

Hamster Pineal Gland in Vitro 19
- 150- .c u c c 2s - a q 100-
S E v
50 -
a, p<O.OI vs C
Medium Culture 200
a, p<O.OI vs C b, p<0.05 vs Is0
150
50M C Is0 PE Is0
+ PE
,h C Is0 PE Is0 PE +
2- 6-
Hours of Incubation Fig. 4. Effect of I S 0 ( 1 pM) or PE (1 pM) alone or in combination (IS0 + PE) on the in vitro stimulation of melatonin production. All hamsters were superior cervical ganglionectomized 10 days before decapitation and then their pineals were incubated for the indicated times; both pineal glands and culture media were quickly frozen on solid C 0 2 and stored at - 70'C until assayed for melatonin. Control values (C) were obtained by incubating glands in the absence of the drugs. Values are expressed as the mean 2 SEM of 8 pineal glands.
melatonin production was studied in pineal glands collected from superior cervical ganglionectomized hamsters and incubated for 4-6 hr. This experi- ment was performed because, when pineals are cultured immediately after excision from anatomically intact hamsters, presynaptic elements associated with the pineal probably remain viable for the duration of the experiment. Consequently, the potentiation of p-adrenergic-stimulated melatonin produc- tion by PE added to the culture medium (Fig. 1, left panel) in these cases may be via an action on either pre- or postsynaptic mechanisms. As seen in Figure 4, PE alone had no effect on the denervated pineal melatonin content, but significantly potentiated I S 0 stimulation of melatonin production in culture medium. These findings suggest that both I S 0 and PE act through postsynaptic a- and P-adrenergic receptors.

20 Santanaetal.
DISCUSSION
In the Syrian hamster, but only in pineal glands collected at night, @-adrenergic mechanisms have been shown to be involved in pineal melatonin production in both in vivo [Reiter e t al., 1987a,b; Vaughan and Reiter, 19871 and in vitro [Vaughan et al., 1986; Santana et al., 19881 experiments. No dataare available concerning the effects of a-adrenergic agonists on (3-adrenergic stimulation of Syrian hamster melatonin production. In this report we investi- gated this possibility and the results provide evidence that a-adrenergic agonists potentiate in vitro the stimulatory effect of P-adrenergic agonists on the Syrian hamster pineal melatonin production.
In experiments designed to study the response of the hamster pineal gland to a-agonists, PE potentiated the stimulatory effect of IS0 on melatonin production in pineal glands from anatomically intact hamsters. However, PE alone was unable to stimulate pineal melatonin production. These observations, that pineals cultured with PE in the absence of I S 0 do not exhibit a rise in melatonin production, suggest that this a-agonist modulates, rather than directly activates, pineal melatonin production. Conversely, short-term cultured pineals whose presynaptic terminals were removed by prior superior cervical ganglion- ectomy were not responsive to the stimulatory effect of ISO. This apparent lack of responsiveness of IS0 was obvious when pineals were cultured for 2 hr and was unrelated to duration of superior cervical ganglionectomy. When denerva. ted pineals were cultured for a longer time (4-6 hr), I S 0 stimulated pineal melatonin production. In the rat, a similar lag period in the induction of pineal NAT activity by IS0 has been described in both in vivo and in vitro studies (Deguchi and Axelrod, 1972a; Romero and Axelrod, 19753. This lag increasedas rats were exposed to longer periods of light, which reduces release of norepinephrine from nerve endings. These data suggest that the synthesis and accumulation of an intermediate, presumably messenger RNA , is necessary for the induction of pineal NAT activity [Zatz et al., 19751. Our results in denervated hamster pineal glands are likely explained by this mechanism as well.
In experiments designed to localize the site of the action of IS0 and PE more precisely, PE was shown to potentiate the stimulatory effect of IS0 on melatonin production in pineal glands collected from superior cervical ganglion- ectomized hamsters. Since presynaptic elements degenerate after ganglionec- tomy, our results suggest that, as in the rat, melatonin production is regulated by both a- and @-adrenergic receptors located on postsynaptic structures. In order to explain the a-adrenergiclp-adrenergic interactions, several possible mecha- nisms of action have been suggested for the rat [Alphs and Lovenberg, 19841. Hence, a-adrenergic receptors might have an intimate relationship with the postsynaptic @-receptors such that stimulation of the a-receptor induces conformational changes in the (3-receptors, which increase the affinity of the @-receptor for its ligand. Alternatively, a-receptor stimulation may secondarily change @-receptor affinity for P-agonists by changing a physical parameter such as membrane fluidity. A third possibility is that pineal a-receptors operate semi-independently of the (3-receptor and that a-receptor stimulation potenti- ates elements of the induction pathway distal to the @-receptor; however, the @-receptor must obviously be stimulated to initiate enzyme induction and

Hamster Pineal Gland in Vitro 21
melatonin production. Any one of these hypotheses may also explain the modulatory effect of PE on the P-adrenergic stimulation of hamster pineal melatonin production. Further experiments with other agonists and antagonists are required to characterize the nature of both a- and P-adrenergic receptors implicated in the regulation of hamster pineal melatonin production.
ACKNOWLEDGMENTS
Supported by a NSF Grant No. DCB 8410592; C.S. was supported by a postdoctoral fellowship from the Canarias Autonomous Government; J.M.G. was supported by a postdoctoral fellowship from the Fulbright Commission.
LITERATURE CITED
Alphs, L., W. Lovenberg ( 1984) Modulation of rat pineal acetyl-CoA: arylamine N-acetyltransferase
Binkley, S. (1976) Comparative biochemistry of the pineal glands of birds and mammals. Amer.
Bramrner, M., S. Binkley, J. Enrico, M. Duffy (1981) The rise and fall of N-acetyltransferase in vitro: The influence of age. Dev. Biol. 86:256-258.
Brownstein, M., R. Holz, J. Axelrod (1973) The regulation of pineal serotonin by a beta-adrenergic receptor. J. Pharmacol. Exp. Ther. 186:109-113.
Deguchi, T., J. Axelrod ( 1972a) Induction and superinduction of serotonin N-acetyltransferase by adrenergic drugs and denervation in rat pineal organ. Proc. Natl. Acad. Sci. U.S.A.
Deguchi, T., J. Axelrod ( 1972b) Control of circadian changes of serotonin N-act yltransferase activity in the rat pineal organ by beta-adrenergic receptor. Proc. Natl. Acad. Sci. U.S.A.
Ebadi, M. ( 1984) Regulation of the synthesis of melatonin and its significance to neuroendocri- nology. In: The Pineal Gland. R.J. Reiter, ed. Raven, New York, pp. 1-37.
Klein, D.C., J.L. Weller (1970) Indole metabolism in the pineal gland: A circadian rhythm in N-acet yltransferase. Science 169: 1 09 3 - 1 09 5.
Klein, D.C., D. Sudgen, J.L. Weller (1983) Postsynaptic a-adrenergic receptors potentiate the P-adrenergic stimulation of pineal serotonin N-acetyltransferase. Proc. Natl. Acad. Sci. U.S.A.
Lipton, J.S., L.J. Petterborg, RJ. Reiter ( 1981) Influence of propranolol, phenoxybenxamine, or phentolamine on the in vivo nocturnal rise of pineal melatonin levels in the Syrian hamster. Life Sci 28:2377-2382.
Reiter, RJ. ( 1985) Action spectra, dose-response relationships and temporal aspects of light effects on the pineal gland. Annu. N.Y. Acad. Sci. 453215-230.
Reiter, RJ., S. Oaknin, M.E. Troiani, K. Li (1987a) Factors controlling melatonin rhythmicity in the pineal gland of the Syrian hamster. Annu. Rev. Chronopharmacol. 541-44.
Reiter, RJ., M. Puig-Domingo, J.M. Guerrero, A. Gonzalez-Brito (1987b) Nocturnal increase in the sensitivity of the Syrian hamster pineal gland to isoproterenol is darkness dependent Proc. SOC. Exp. Biol. Med. 185:219-222.
Reiter, RJ., G.M. Vaughan, S. Oaknin, M.E. Troiani, B. Cozzi, K. Li ( 1987c) Norepinephrine or isoproterenol stimulation of pineal N-acetyltransferase activity and melatonin content in the Syrian hamster is restricted to the second half of the daily dark phase. Neuroendocrinology
Rollag, M.D., G.D. Niswender ( 1976) Radioimmunoassay of serum concentrations of melatonin in
Romero, J.A., J. Axelrod (1975) Regulation of sensitivity of P-adrenergic stimulation in induction
Santana, C., J.M. Guerrero, R.J. Reiter, M. Puig-Domingo, A. Gonzalez-Brito (1988) Stimulatory
induction by alpha adrenergic drugs. J. Pharmacol. Exp. Ther. 230:431-437.
2001. 16:57-65.
69:2208-2211.
69:2547-2550.
80599-603.
45:249-256.
sheep exposed to different lighting regimes. Endocrinology 98:482-489.
of pineal N-acetyltransferase, Proc. Natl. Acad. Sci. U.S.A. 72:1661-1665.

22 Santanaetal.
effect of isoproterenol but not of dibutyryl cyclic AMP on N-acetyltransferase activity and melatonin content of Syrian hamster pineal gland in organ culture. Neuroendocrinology
Vaughan, G.M., R.J. Reiter (1987) The Syrian hamster pineal gland responses to isoproterenol in vivo at night. Endocrinology 120:1682-1684.
Vaughan, G.M., J. Lasko, S.H. Coggins, B.A. Pruitt, A.D. Mason (1986) Rhythmic melatonin response of the Syrian hamster pineal gland to norepinephrine in vitro and in vivo. J. Pineal Res.
Zatz, M. (1981) Pharmacology of the rat pineal gland. In: The Pineal Gland, Vol. 1. RJ. Reiter, ed.
Zatz, M., J.A. Romero, J. Axelrod ( 1975) Diurnal variations in the requirement for RNA synthesisin
48:229-234.
5235-249.
CRC, Boca Raton, FL, pp. 229-242.
the induction of pineal N-acetyltransferase. Biochem. Pharmacol. 25903-906.





























