Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY B 1985 by The American Society of Biological Chemists, Inc.
Vol. 260, No. 9, Issue of May 10, pp. 5641-5647,1985 Printed in U. S.A.
Role of the Nuclear Envelope in Synthesis, Processing, and Transport of Membrane Glycoproteins*
(Received for publication, August 13, 1984)
Lynn Puddington$, Mark 0. Lively, and Douglas S . Lyles From the Deoartment of Microbiology and Immunology and the Department of Biochemistry, Wake Forest Uniuersity Medical ”
Center, Win$ton-Salem; North Carolina 27103
The outer nuclear membrane is morphologically sim- ilar to rough endoplasmic reticulum. The presence of ribosomes bound to its cytoplasmic surface suggests that it could be a site of synthesis of membrane glyco- proteins. We have examined the biogenesis of the ve- sicular stomatitis virus G protein in the nuclear enve- lope as a model for the biogenesis of membrane glyco- proteins. G protein was present in nuclear membranes of infected Friend erythroleukemia cells immediately following synthesis and was transported out of nuclear membranes to cytoplasmic membranes with a time course similar to transport from rough endoplasmic reticulum (&, = 5-7 min). Temperature-sensitive mu- tations in viral membrane proteins which block trans- port of G protein from endoplasmic reticulum also blocked transport of G protein from the nuclear enve- lope. Friend erythroleukemia cells and NIH 3T3 cells differed in the fraction of newly synthesized G protein found in nuclear membranes, apparently reflecting the relative amount of nuclear membrane compared to en- doplasmic reticulum available for glycoprotein synthe- sis. Nuclear membranes from erythroleukemia cells appeared to have the enzymatic activities necessary for cleavage of the signal sequence and core glyco- sylation of newly synthesized G protein. Signal pepti- dase activity was detected by the ability of detergent- solubilized membranes of isolated nuclei to correctly remove the signal sequence of human preplacental lac- togen. RNA isolated from the nuclear envelope was highly enriched for G protein mRNA, suggesting that G protein was synthesized on the outer nuclear mem- brane rather than redistributing to nuclear membranes from endoplasmic reticulum before or during cell frac- tionation. These results suggest a mechanism for in- corporation of membrane glycoproteins into the nu- clear envelope and suggest that in some cell types the nuclear envelope is a major source of newly synthe- sized membrane glycoproteins.
The nuclear envelope consists of an inner and outer mem- brane enclosing a lumen known as the perinuclear space.
* This work was supported in part by National Institutes of Health Grants A115892 and GM 32861 and by pilot grants from the Oncology Research Center, Bowman Gray School of Medicine, and from the North Carolina United Way. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked ‘‘aduertkemnt” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Predoctoral trainee supported by National Institutes of Health Training Grant T32 CA09422. To whom correspondence should be addressed Molecular Biology and Virology Laboratory, The Salk Institute, P. 0. Box 85800, San Diego, CA 92138.
Morphological observations characterizing the structure of the nuclear envelope have led investigators to postulate that the outer membrane could be a site of membrane-bound synthesis of glycoproteins (reviewed in Franke et al., 1981). The presence of ribosomes on the cytoplasmic surface of the outer nuclear membrane and its continuity with the endo- plasmic reticulum (ER’) (Watson, 1955) suggest structural similarity between the two organelles and have led to the hypothesis that the nuclear envelope is functionally a spec- ialized form of ER. The enzymatic activities of nuclear mem- branes and ER are similar (Franke et al., 1981). However, there are some cases of functional differentiation between nuclear membranes and ER as well as some activities that appear to be equivalent in the two membrane systems. An example of the former is the cytochrome P-450-dependent electron transport system, which appears to differ between nuclear membranes and ER with respect to isozyme pattern and drug inducibility (Fahl et al., 1978; Matsuura et al., 1981). In contrast, the glucose-6-phosphatase activity of the two membranes appears to be due to equivalent enzymes (Arion et al., 1983). These results suggest that each activity must be examined independently to determine whether the two organ- elles provide similar functions.
Recent studies have shown that viral and host glycoproteins normally synthesized on ER and transported to plasma mem- branes are also present in the nuclear envelope (Albert and Davies, 1973; Bergmann et al., 1981; Lyles and McConnell, 1981; Rose and Bergmann, 1982). However, neither a mech- anism for incorporation into the nuclear envelope nor the biochemical characteristics of these proteins in the nuclear envelope have been defined. The unexpected result of our previous experiments was that significant proportions (30- 60%) of murine leukemia virus glycoproteins are present in a nuclear fraction obtained from Friend erythroleukemia cells (Lyles and McConnell, 1981). An explanation for this distri- bution could be that some glycoproteins, such as leukemia virus envelope proteins, are preferentially localized in the nuclear envelope of erythroleukemia cells. Alternatively, a more generalized transient association with the nuclear en- velope could be common to newly synthesized membrane glycoproteins in many cell types. These hypotheses were addressed in this study through characterization of the vesic- ular stomatitis virus (VSV) glycoprotein, G, in the nuclear envelope of infected cells.
Biosynthesis of the VSV G protein has been well-charac- terized and serves as a model for many viral and cellular integral membrane proteins destined to be incorporated into the plasma membrane. G protein is synthesized on polyribo-
’The abbreviations used are: ER, endoplasmic reticulum; VSV, vesicular stomatitis virus; endo H, endo-P-N-acetylglucosaminidase H.
5641

5642 Membrane Glycoprotein Biogenesis in Nuclear Membranes
somes bound to the ER (Morrison and Lodish, 1975; Knipe et al., 1977b) and is inserted into the rough ER during syn- thesis (Katz et al., 1977). During translation, two high man- nose oligosaccharides are added to the nascent chain (Robbins et al., 1977; Tabas et al., 1978). G protein remains anchored in the membrane via a hydrophobic region at the carboxyl terminus of the protein (Rose et al., 1980), and a hydrophobic signal sequence at the amino terminus is cleaved by a pepti- dase present in ER membranes (Lingappa et al., 1978). Soon after synthesis in the rough ER, G protein is transported to the Golgi complex (Bergmann et al., 1981) where its oligosac- charides are modified (Tabas and Kornfeld, 1979; Bretz et al., 1980; Griffiths et al., 1982; Roth and Berger, 1982). It is then transported to the plasma membrane where it is incorporated into budding virions (Knipe et al., 197713).
We have examined the possible role of the nuclear envelope in the early stages of glycoprotein maturation. The nuclear envelope appears to contain the necessary functions for the biogenesis of G protein, including G protein mRNA, signal peptidase, and ability to glycosylate G with mannose-rich oligosaccharides. We have determined the distribution and lifetime of G protein in the nuclear envelope of cells infected with wild-type and temperature-sensitive mutants of VSV. The results obtained are consistent with the hypothesis that the nuclear envelope and rough ER are functionally equiva- lent in glycoprotein synthesis and processing and that in some cell types the nuclear membrane is a major source of newly synthesized membrane glycoprotein.
EXPERIMENTAL PROCEDURES AND RESULTS~
Time Course of G Protein Transport from the Nuclear Envelope of Infected Erythroleukemia Cells-The subcellular distribution and rate of transport of the VSV G protein were determined in pulse-chase experiments followed by subcellu- lar fractionation. VSV-infected Friend erythroleukemia cells were labeled with [35S]methionine for 5 min and then incu- bated in medium containing unlabeled methionine for 0,5, or 15 min. Nuclei and total cytoplasmic membranes were isolated from cells at each time point. The protein composition of each fraction was determined by electrophoresis and fluorography. Fig. 2 shows that approximately half of the G protein was present in nuclear membranes initially and was rapidly chased into cytoplasmic membranes. In addition to G protein, the nonglycosylated viral membrane protein, M, was present in the nuclear fraction. However, the higher molecular weight form of G protein, visible after modification by Golgi enzymes, and nucleocapsid proteins (N, NS, and L) were virtually absent from the nuclear fraction, thus providing further evi- dence that cytoplasmic contamination was negligible. Quan- titation of G protein by densitometry demonstrated that 40 & 5% of newly synthesized ( t = 0 ) G protein was present in the nuclear membrane. The same distribution of labeled G was obtained with a 1-min incubation (data not shown) as with a 5-min incubation of cells in [35S]methionine. As dis- cussed below, less than 10-15% of G protein observed in the nuclear fraction could have been contributed by contaminat- ing ER. The half-time of G protein in the nuclear envelope
* Portions of this paper (including “Experimental Procedures,” part of “Results,” and Figs. 1 and 3-9) are presented in miniprint a t the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 84M-2537, cite the authors, and include a check or money order for $7.20 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
L -
G -
N -
”
0 5 1 5 0 5 1 5 0 5 15 wc N C
FIG. 2. Distribution and time course of transport of G pro- tein in the nuclear envelope of erythroleukemia cells. VSV- infected Friend erythroleukemia cells were labeled with [35S]methio- nine for 5 min at 4 h postinfection and then incubated in medium containing unlabeled methionine for the indicated times (minutes). Nuclei and cytoplasmic membranes were obtained from cells at each time point and analyzed on a 10% sodium dodecyl sulfate-polyacryl- amide gel. A fluorograph of the dried gel is shown. Proteins were extracted from unfractionated whole cells (one-third of total a t each time point) (WC), purified nuclei ( N ) , and cytoplasmic membranes (C). The VSV nucleocapsid proteins N and NS co-migrate in this electrophoresis system.
was calculated to be 5.1 f 2.5 min. As the quantity of labeled G protein decreased in the nuclear fraction during the chase, it increased in cytoplasmic membranes, demonstrating quan- titative transport from nuclear to cytoplasmic membranes. Therefore, selective degradation of the nuclear G protein could not account for the half-time observed.
DISCUSSION
The presence of specific glycoproteins in the nuclear enve- lope has been demonstrated by several laboratories. Albert and Davies (1973) showed by 51Cr cytotoxicity inhibition and complement fixation tests that H-2 antigens are present in nuclear membranes obtained from mouse liver or thymus. Viral glycoproteins have been visualized in the nuclear enve- lope by indirect immunofluorescence microscopy of fixed, permeabilized cells, in which labeling of the nuclear envelope is indicated by ring-like staining around the nucleus. Wild- type VSV G protein was detected in the nuclear envelope of

Membrane Glycoprotein Biogenesis i n Nuclear Membranes 5643
cells microinjected with cDNA encoding G protein, but other perinuclear structures, apparently Golgi membranes, were also heavily labeled (Rose and Bergmann, 1982). Labeling of the nuclear envelope by antibody against viral glycoproteins was much clearer in cells expressing mutant glycoproteins of VSV or vaccinia virus, blocked in transport from rough ER to the cell surface (Bergmann et al., 1981; Rose and Bergmann, 1983; Shida and Matsumoto, 1983). A mutant G protein was also detected in the nuclear envelope by ferritin immunoelec- tron microscopy of cells infected with the ts045 mutant of VSV, which is similar to the tsL513 mutant used in this study (Bergmann et al., 1981; Bergmann and Singer, 1983). In the present study, we demonstrated the presence of wild-type G protein in the nuclear envelope using independent techniques, including immunofluorescence microscopy of isolated nuclei without fixation or permeabilization (Fig. 1) and gel electro- phoresis of proteins of isolated nuclei (Fig. 2).
The present study differs from previous morphological stud- ies because it shows that the nuclear envelope is functionally involved in glycoprotein biogenesis. Labeled G protein in the nuclear envelope was shown to be a precursor to the mature glycoprotein, rather than a mutant glycoprotein defective in post-translational modifications and/or intracellular trans- port. The biochemical resemblance of VSV G protein in the nuclear envelope to that in rough ER suggests a similarity of the two organelles in glycoprotein biogenesis. Several lines of evidence support this hypothesis, including presence of G in the nuclear envelope immediately after synthesis, transport of G to membranes in the cytoplasm, glycosylation of G in the nuclear envelope, and presence of signal peptidase activity and G mRNA in the nuclear envelope.
G protein was present in nuclear membranes immediately after synthesis and was transported to cytoplasmic mem- branes with a time course similar to transport of G protein from ER. While it is difficult to rule out the possibility that synthesis of G protein occurred on cytoplasmic rough ER followed by immediate redistribution to the nuclear envelope, this seems unlikely since nuclei purified from infected eryth- roleukemia cells pulse-labeled with [35S]methionine for 1 min also contained approximately half the total labeled G protein. In addition, RNA isolated from nuclei is highly enriched in mRNA for G protein, suggesting that mRNA for this protein is actively translated on the nuclear membrane in infected cells. The G protein of a temperature-sensitive mutant of VSV that is not transported from ER at the nonpermissive temperature was also not transported from nuclear mem- branes, in agreement with morphological observations (Berg- mann et al., 1981, 1983). Mutations in other viral envelope proteins, such as the hemagglutinin of vaccinia virus, result in accumulation of the glycoproteins in both organelles (Shida and Matsumoto, 1983).
Activities required for proteolysis of signal peptides and glycosylation of G protein were demonstrated in the nuclear envelope. Detergent-solubilized extracts of nuclei contained a proteolytic activity that correctly processed preplacental lac- togen to its mature form in a post-translational assay. Signal peptidases studied to date have been shown to act on a variety of substrates from many species. Preplacental lactogen has been shown to be a substrate for solubilized signal peptidases from hen oviduct (Lively and Walsh, 1983) and dog pancreas (Strauss et al., 1979) and purified leader peptidase (Wolfe et al., 1982) from Escherichia coli3 Therefore, the protease ac- tivity observed in Friend erythroleukemia cell nuclei appears to be a true signal peptidase.
M. 0. Lively, unpublished results.
Since glycosylation of G is a cotranslational process (Roth- man and Lodish, 1977), the synthesis of G on the nuclear envelope requires that the appropriate glycosyltransferase activity also be present. Like G protein in ER, oligosaccha- rides on G protein in the nuclear envelope are sensitive to digestion by endo H. In addition, the endo H-sensitive G protein in the nuclear envelope was a precursor to the endo H-resistant G protein in the cytoplasm, since quantitative transport and conversion were observed. Our experiments also showed that transport from the nuclear envelope pre- ceded acquisition of endo H resistance because nuclear G protein was always susceptible to endo H digestion. The half- time of G protein in the nuclear envelope was shorter than the half-time of conversion to the endo H-resistant, higher molecular weight form in cytoplasmic membranes (see Figs. 2 and 3).
The distribution of newly synthesized G protein between nuclear and cytoplasmic membranes was dependent on the cell type examined. Forty per cent of newly synthesized G was present in nuclei from erythroleukemia cells while only 15% was present in nuclei from NIH 3T3 cells. This difference probably reflects the relative amount of ER compared to nuclear membrane in these cells. Electron micrographs of thin sections of these cells supported this hyp~thesis.~ Friend erythroleukemia cells contained very little cytoplasmic rough ER and a large nucleus, while NTH 3T3 cells contained relatively more cytoplasmic rough ER. This difference in cell types probably accounts for the unexpectedly high percentage of murine leukemia virus glycoproteins in nuclear membranes of Friend erythroleukemia cells (Lyles and McConnell, 1981). Determinations made from pulse-chase experiments of the capacity of the nuclear envelope to synthesize glycoproteins are most likely underestimates in both cell types, because portions of the outer nuclear membrane are frequently re- moved during isolation of nuclei (Maggio et al., 1963; Karten- beck et al., 1973; Wishart and Fry, 1980). Consequently, a fraction of labeled glycoprotein synthesized on the nuclear envelope is found in the cytoplasmic membrane fraction. Nonetheless, these observations suggest that the relative amount of rough ER is an important factor determining distribution of glycoprotein biosynthetic capacity within cells.
The fact that Friend erythroleukemia cells contain such a large proportion of newly synthesized G protein in the nuclear membrane has proven advantageous for ruling out the contri- butions of other organelles. In these experiments, the most troublesome contaminant of nuclei would be rough ER. This is the most difficult contaminant to rule out by analysis of marker enzymes, since the enzyme profiles of rough ER and nuclear membranes are so similar. NADPH-cytochrome c reductase is typically used as a marker for ER, but this enzyme is also present in the nuclear envelope (Franke, 1974). How- ever, if NADPH-cytochrome c reductase activity is taken to be an upper estimate of contamination of nuclei by ER, a limit can be placed on the extent to which contaminating rough ER could account for viral glycoproteins in the nuclear fraction of pulse-chase experiments. Nuclei purified from Friend erythroleukemia cells contain 6-10% of the total cel- lular NADPH-cytochrome c reductase activity (Lyles and McConnell, 1981). Since approximately 60% of total newly synthesized (labeled) glycoprotein was in cytoplasmic mem- branes with a distribution similar to NADPH-cytochrome c reductase, 4-6% of total labeled glycoprotein could represent cytoplasmic contamination in the nuclear fraction. Thus, 10- 15% of the glycoprotein in the nuclear fraction obtained from
L. Puddington and D. S. Lyles, unpublished results.

5644 Membrane Glycoprotein Biogenesis in Nuclear Membranes
erythroleukemia cells in radiolabeling experiments would be due to ER contamination (4-6% divided by the 40% of total labeled glycoprotein in the nuclear fraction). As indicated above, this is probably an upper limit, since some of the NADPH-cytochrome c reductase in the nuclear fraction is due to the presence of the enzyme in the nuclear membrane. The absence of significant cytoplasmic contamination in the nuclear fraction was confirmed by electron microscopy. Mi- crosomes derived from rough ER are readily identifiable with the electron microscope. Contamination of nuclei by other organelles is less than 5% (Lyles and McConnell, 1981), and most contaminants present appear to be debris from dead cells or lysed nuclei and not recognizable ER. Contamination of nuclei with smooth cytoplasmic membranes is also negli- gible, since the fully glycosylated form of G protein was never detected in the nuclear fraction (see Fig. 2). Furthermore, our basic conclusion is substantiated by a technique which incor- porates more stringent morphological criteria in the analysis, namely flow cytometry. The cytofluorograph was gated to measure the fluorescence intensity of those particles with light-scattering properties similar to nuclei, therefore exclud- ing fluorescent-labeled whole cells and cytoplasmic vesicles from the analysis. The half-time of G protein in the nuclear envelope determined by this technique was indistinguishable from the half-time determined in pulse-chase experiments.
The results described here suggest a general mechanism by which membrane glycoproteins are incorporated into the outer nuclear membrane. Our recent immunoelectron micro- scopic results have shown that viral glycoproteins, including G protein, incorporated biosynthetically into the outer nuclear membrane, are also present on the lumenal surface of the inner nuclear membrane.5 Viral glycoproteins in the nuclear envelope may have roles other than as transient intermediates in glycoprotein synthesis, particularly if they are also incor- porated into the inner nuclear membrane. An example might be the gp55 glycoprotein encoded by spleen focus-forming virus, the highly oncogenic component of the Friend murine leukemia virus complex. Little, if any, gp55 is expressed on the cell surface. Transport of gp55 from rough ER (Kabat et al., 1980; Ruscetti et ai., 1981) and nuclear membranes (Lyles and McConnell, 1981) appears to be blocked. I t is possible that oncogenesis by gp55 involves interactions specific for the nuclear envelope, such as transmembrane communication with nucleoplasmic elements. Likewise, it is possible that cell surface glycoproteins are normally incorporated into the nu- clear envelope by mechanisms similar to those described here for viral glycoproteins. Additional functions for these glyco- proteins, such as hormone receptors, could also be expressed in the nuclear envelope.
Acknowledgments-We are grateful to Dr. David A. Bass and Pamela Szeda for helpful advice during flow cytometry experiments.
REFERENCES Albert, W. H. W., and Davies, D. A. L. (1973) Immunology 24 , 841-
Arion, W. J., Schulz, L. 0.. Lange, A. J., Telford, J. N., and Walls, 850
H. E. (1983) J. Biol. Chem. 258,12661-12669
L. Puddington, D. S. Lyles, and J. C. Lewis, manuscript in preparation.
Bergmann, J. E., and Singer, S. J. (1983) J. Cell Biol. 9 7 , 1777-1787 Bergmann, J. E., Tokuyasu, K. T., and Singer, S. J. (1981) Proc. Natl.
Bretz, R., Bretz, H., and Palade, G. E. (1980) J. Cell Biol. 84, 87-101 Etchison, J. R., Robertson, J. S., and Summers, D. F. (1977) Virology
Fahl, W. E., Jefcoate, C. R., and Kasper, C . B. (1978) J. Biol. Chem.
Franke, W. W. (1974) Inte. Reu. Cytol. 4 , (suppl.) 71-236 Franke, W. W., Scheer, U., Krohne, G., and Jarasch, E. D. (1981) J.
Friend, C. (1957) J. Exp. Med. 105.307-318 Green, J., Griffiths, G., Louvard, D., Quinn, P., and Warren, G. (1981)
Griffiths, G., Brands, R., Burke, B., Louvard, D., and Warren, G.
Jamieson, J. D., and Palade, G . E. (1967) J. Cell Biol. 34, 577-596 Johnson, G. D., Davidson, R. S., McNamee, K. C., Russell, G.,
Goodwin, D., and Holborow, E. J. (1982) J. Immunol. Methods 5 5 ,
Kabat, D., Ruta, M., Murray, M. J., and Polonoff, E. (1980) Proc.
Kartenbeck, J., Jarasch, E. D., and Franke, W. W. (1973) Exp. Cell
Katz, F. N., Rothman, J. E., Lingappa, V. R., Blobel, G., and Lodish,
Knipe, D. M., Baltimore, D., and Lodish, H. F. (1977a). J. Virol. 21 ,
Knipe, D. M., Baltimore, D., and Lodish, H. F. (1977b). J. Virol. 21,
Laemmli, U. K. (1970) Nature 227 , 680-685 Laskey, R. A., and Mills, A. D. (1975) Eur. J. Biochem. 56, 335-341 Lefrancois, L., and Lyles, D. S. (1982) Virology 121, 157-167 Lingappa, V. R., Katz, F. N., Lodish, H. F., and Blobel, G. (1978) J.
Lively, M. O., and Walsh, K. A. (1983) J . Biol. Chem. 2 5 8 , 9488-
Lyles, D. S., and McConnell, K. A. (1981) J. Virol. 39, 263-272 Maggio, R., Siekevitz, P., and Palade, G. E. (1963) J. Cell Biol. 18,
Matsuura, S., Masuda, R., Omori, K., Negishi, M., and Tashiro, Y.
McMaster, G. K., and Carmichael, G. G . (1977) Proc. Natl. Acad. Sci.
Morrison, T . G., and Lodish, H. F. (1975) J. Biol. Chem. 250,6955-
Rinderknecht, H. (1962) Nature 193 , 167-168 Robbins, P. W., Hubbard, S. C., Turco, S. J., and Wirth, D. F. (1977)
Rose, J. K., and Bergmann, J. E. (1982) Cell 30, 753-762 Rose, J. K., and Bergmann, J. E. (1983) Cell 34, 513-524 Rose, J. K., and Knipe, D. (1975) J. Virol. 15, 994-1003 Rose, J. K., Welch, W. J., Sefton, B. M., Esch, F. S., and Ling, N. C.
(1980) Proc. Natl. Acad. Sci. U. S. A . 77, 3884-3888 Roth, J., and Berger, E. G. (1982) J. Cell Biol. 9 3 , 223-229 Rothman, J. E., and Lodish, H. F. (1977) Nature 269, 775-780 Ruscetti, S., Field, J. A,, and Scolnick, E. M. (1981) Nature 294,
Shida, H., and Matsumoto, S. (1983) Cell 33, 423-434 Straws, A. W., Zimmerman, M., Boime, I., Ashe, B., Mumford, R.
A,, and Alberts, A. W. (1979) Proc. Natl. Acad. Sci. U. S. A . 7 6 ,
Acad. Sci. U. S. A. 7 8 , 1746-1750
78,375-392
253,3106-3113
Cell Biol. 9 1 , 399-50s
J. Mol. Biol. 152, 663-698
(1982) J. Cell Biol. 95. 781-792
231-242
Natl. Acad. Sci. U. S. A. 77 , 57-61
Res. 8 1 , 175-194
H. F. (1977) Proc. Natl. Acad. Sci. U. S. A. 74 , 3278-3282
1149-1158
1128-1139
Biol. Chem. 253,8667-8670
9495
267-291
(1981) J. Cell Biol. 91 , 212-220
U. S. A. 74,4835-4838
6962
Cell 12,893-900
663-665
4225-4229 Tabas, I., and Kornfeld, S. (1979) J. Biol. Chem. 254, 11655-11663 Tabas, I., Schlesinger, S., and Kornfeld, S. (1978) J. Biol. Chem. 253,
Watson, M. L. (1955) J. Biophys. Biochem. Cytol. 1, 257-270 Wishart, G. J., and Fry, D. J. (1980) Biochern. J. 186,687-691 Wolfe, P. B., Silver, P., and Wickner, W. (1982) J . Biol. Chem. 257,
Zilberstein, A., Snider, M. D., Porter, M., and Lodish, H. F. (1980)
716-722
7898-7902
Cell 21,417-427

Membrane Glycoprotein Biogenesis in Nuclear Membranes 5645 S u p p l e m e n t a l M a t e r i a l s t o
PRIlCESSlNG AN0 TRANSPORT UF MEM8HANE BLYCOPROTEINS ROLE UF T H E NUCLEAR ENVELOPE I N SYNTHESIS.
QY
t ynn Pudd ing ton . naru 0. L i v e l y , ana o o u g ~ a s s. ~ y l e r
EXPERIMENTAL PROCEDURES
i n f e c t e d c e l l s were d e p l e t e d o f m e t h i o n i n e b y i n c u b a t & o n w i t h m e t h i o n i n e - f r e e UMEM f o r 10 n i n . C e l l s were p u l s e d f o r 5 m i n w i t h C s l m e t h i o n i n e l 2 5 ~ C i l m l .
serum, t h e n c h a s e d i n m e d i u m c o n t a i n i n g u n l a b e l e d m e t h i o n i n e f o r Y l P y i n g ,6uO C i l m m o l e ) i n m e t h i o n i n e - f r e e medium c o n t a i n i n g 2.5; d i a l y z e d f e t a l C a l f
l e n g t h s o f t i m e . n u c l e i were i s o l a t e d a s d e s c r i b e d above. T o t a l c y t o p l a s m i c membranes p r e s e n t i n t h e l o w s p e e d s u p e r n a t a n t were p e l l e t e a a t 100.000 x g f o r 30 .in. E a c h f r a c t i o n was 1 0 I u b 1 1 i z e d w i t h R l P A b u f f e r ( I O m M T r i s . p H 7.4. 0.15M N a C l . I1 T r i t o n x-100. I; s o d i u m d e o x y c h o l a t e . 0.11 S O S . ImM p h e n y l n e t h y l r u l f o n y ~ f l u o r i d e ) ana n u c l e i c a c i d s removed f r o m u n f r a c t i o n a t e d c e l l s a n a n u c l e i b y c e n t r i f u g a t i o n a t 15.000 x 9 f o r 15 .in. The p r o t e i n c o m p o s i t i o n o f e a c h f r a c t i o n ;as t h e n a e t e r n i n e a ~ b y s o s - p o 1 y a c r y l a m i d c g e l e l e c t r o p h o r e s i s ( L a e r n m l i . 1 9 7 0 ) and f l u o r o g r a p h y u s i n g p r e - e x p o s e d XadaU S B - 5 film ( L a r k e y a n d 1 1 1 1 s . 1 9 7 5 ) . p u a n t i t a t i o n w a s b y s o f t l a s e r s c a n n i n g d e n s i t o m e t r y ( B i o m e d I n s t r u m e n t s . I n c . . F u l l e r t o n . C a l i f o r n i a ) .
t h a t i n f e c t i o n w i t h t s v i r v r e r was a t 3l'C f o r 3.5 h and t h e n s h i f t e d t o 40-C P u l s e - C h a s e a n a l y s i s a f m u t a n t p r o t e i n s was p e r f o r m e d s i m i l a r l y e x c e p t
f o r 0.5 h. T h i s p r o t o c o l f a c i l i t a t e s e f f i c i e n t s y n t h e s i s o f v i r a l p r o t e i n s w i t h o u t p r o d u c t 7 . m o f i n f e c t i o u s v i r i o n s ( K n i p e e t fi.. 1977a). Tr a u t a n t - I n f e c t e d c e l l s were p u l s e - l a b e l e d and chased a t e T t h e r 31° o r 40-c. p r i o r t o f r a c t i o n a t i o n ana a n a l y s i s o f G r o t e i n r a s d e s c r i b e d .
5646 Membrane Glycoprotein Biogenesis in Nuclear Membranes
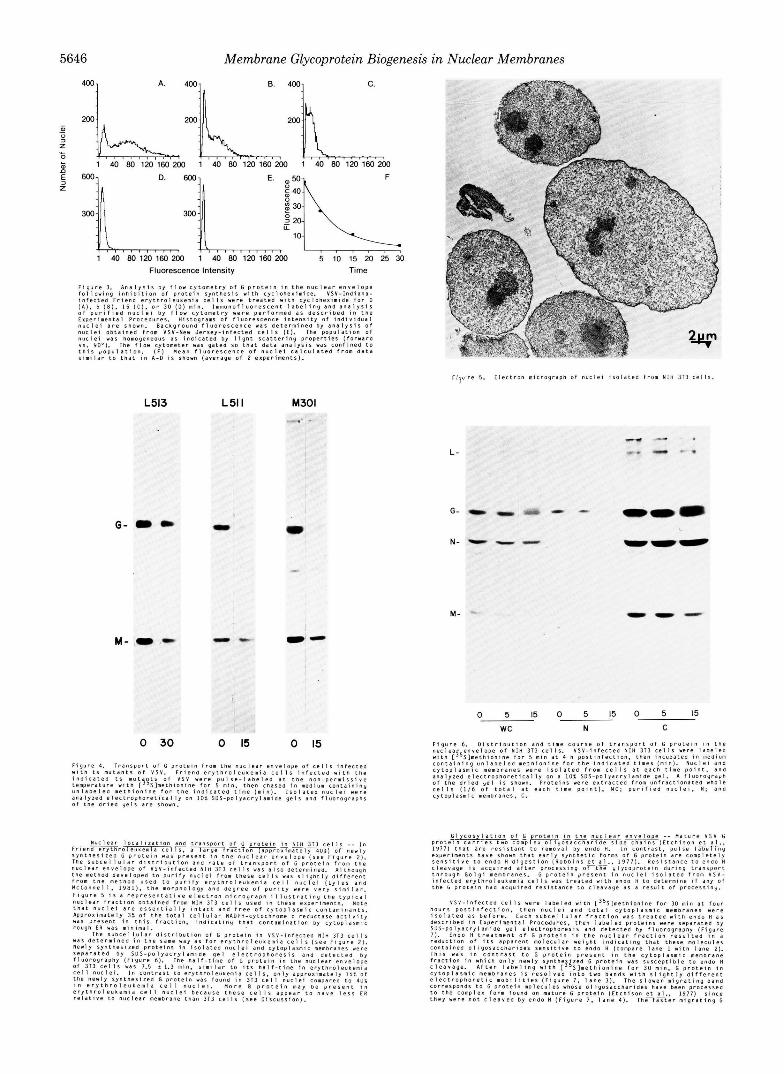
400 1 A. 4001 8. 4001 C.
200L200L 1 40 80 120 160 200 1 40 80 120 160 200 200L 1 40 80 120 160 200
JI 1 40 80 120 160200 1 40 80 I20160200 5 10 15 20 25 30
Fluorescence Intensity Time
F i g u r e 3. A n a l y r l s by f l o w C y t o m e t r y o f G p r o t e i n i n t h e n u c l e a r e n v e l o p e f o l l o w i n g inhibition o f p r o t e i n S y n t h e s i s w i t h C y c l o h e x i m i d e . V S V - I n d i a n a - I n f e c t e d F r i e n d e r y t n P o I e u k e m i a c e l l s were t r e a t e d w i t h C y C l o h e x i r n i d e f o r D
o f p u r l f i e d n u c l e i b y f l o w c y t o m e t r y w e r e p e r f o r m e d a s d e s c r l b e d i n t h e (A) . 5 (n), 15 ( c ) . o r 30 ( 0 ) " i n . ~ r n m u n o f l u o r e ~ c e n t l a b e l i n y a n d a n a l y s i s
E x p e r i m e n t a l Pcocedures. H i s t o g r a m o f f l u o r e s c e n c e i n t e n s i t y o f i n d i v i d u a l
n u c l e i o b t a i n e d f r o m VSV-New J e r s e y - i n f e c t e d c e l l s ( E ) . The p o p u l a t i o n o f n u c l e i a r e s h o w n . B a c k g r o u n d f l u o r e s c e n c e v a s d e t e r m i n e d b y a n a l y s i s O f
n u c l e i w a s hOmogeneOuI a s indicated by I l g h t s c a t t e r i n g p r o p e r t i e s ( f o r w a r d
t h i s p o p u l a t i o n . I F ) M e a n f l u o r e s c e n c e o f n u c l e i c a l c u l a t e d f p o m d a t a VI. Y O ' ) . The f l o w c y t o m e t e r was g a t e d IO t h a t d a t a a n a l y s i s was c o n f i n e d t o
I imi la l . t o t h a t i n A-0 i s shown ( a v e r a g e o f 2 e x p e r i m e n t s ) .
,- , , ? ~ r e 5. E l e c t r o n m i c r o g r a p h o f n u c l e i I s o l a t e d f r o m NIH 313 cells.
L513 L51 I M301
0-
0 30 0 15 0 15
L-
G-
Membrane Glycoprotein Biogenesis in Nuclear Membranes 5647
L- .m.m I . I .
G- +
N-
M-
- + - "
+ N C
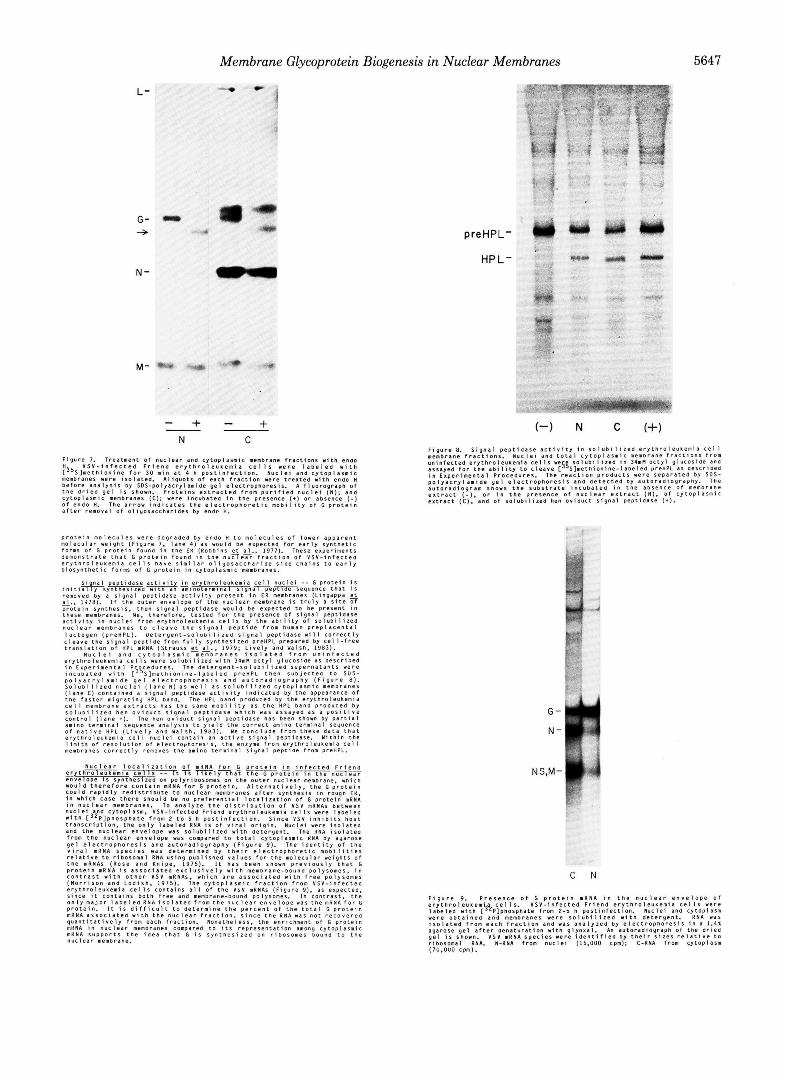
F i g u r e 7. T r e a t m e n t O f n u c l e a r and c.ytOplasmiC membrane f r a c t i o n s w t h enao V S V - i n f e c t e d F r i e n d e r y t h r o l e u t e m i a c e l l s w e r e l a b e l e d w i t h
membranes were i s o l a t e d . A l i q u o t s o f e a c h f r a c t i o n were t r e a t e d w i t h endo H ! 3 5 S l m e t n i o n i n e f o r 3D . in a t 4 h p o s t l n f e c t i o n . N u c l e i a n d c y t o p l a r m i c
b e f o r e a n a l y s i s b y S D S - p o l y a c r y l a m i d e g e l e l e c t r o p h o r e s i s . A f l u o r o g r a p n o f t h e d r i e d g e l i s shown. P r o t e i n s e x t r a c t e d f F o m p u r i f i e d n u c l e i ( N ) ; a n d c y t o p l a s m i c membranes IC): w e r e i n c u b a t e d i n t h e p p e s e n c e (+) o r absence ( - 1 o f e n d o H. The a r r o w i n d i c a t e s t h e e l e c t r O D h o r e t i c m o b i l i t v O f G o r o t e i n
preHPL-
HP L-
a f t e r r e m o v a l o f o l i y o l a c c h d r i d e s by endc H.
p r o t e i n r n o l e ~ u ~ e r were degraded by e n d a H t o m o l e c u l e s o f l o w e r a p p a r e n t m o l e c u l a r w e i g h t ( F i g u r e 7. l a n e 4 ) a s w o u l d be e x p e c t e d f o r e a r l y s y n t h e t i c f o r m O f G p r o t e i n f o u n o i n t h e E N ( H o b b i n s e t a1.. 1977). These exper iments d e m o n s t r a t e t h a t G p r o t e i n f o u n d i n t h e nuTle;Fi f r a c t i o n o f v s v - i n f e c t e a e r y t h r o l e u ~ e m i a c e l l s n a v e s i m i l a r o l i y o s a c c h a r i d e s i d e c h a i n s t o e a r l y b i o s y n t h e t i c f o 7 m o f G p r o t e i n i n C y t o p l a s m i c membranes.
G-
N-
C N
F i g u r e 9. P r e s e n c e o f G p r o t e i n mRNA i n t h e n u c l e a r e n v e l o p e o f e r y t h r o l e u u e n i a c e l l s V S V - i n f e c t e d F r i e n d e r y t h r o l e u u e m i a c e l l s were
were o b t a i n e d a n d membranes were 1 0 1 U Q i l i z e d w i t h d e t e r y e n t . R N A w a s l a b e l e a w i t h [ 3 L P ] p h o r d h a t e f r o m 2 - 5 n p e s t i n r e c t a o n . N u c l e i ana c y t o p l a s m
i s o l a t e d f v o m each f r a c t i o n a n a WII a n a l y z e d b y e ~ e c t r o p h o r e s i s i n a 1.41 agarose g e l a f t e r d e n a t u r a t i o n w i t h g l y o x a l . A" a u t o r a d i o g r a p h o f t h e d r i e d g e l i s shewn. VSV mRNA s p e c i e s were i d e n t i f i e d b y t h e i r s i z e s r e l a t i v e t o r i b o s o m a l RNA. N-RNA f rom nuc le i (15 .000 cpm): C-RNA f rom Cytoplasm (7u.OuD cpm).









![Neotectonics of Tabas Area, Central Iran by Index of ...Active, Tectonics, Index, Tabas, Central Iran 1. Introduction The study area is the north part of Tabas area [1] in the east-central](https://img.dokumen.tips/doc/110x75/60c7cd682ac9086ca8179434/neotectonics-of-tabas-area-central-iran-by-index-of-active-tectonics-index.jpg)



















