Embed Size (px)
Citation preview

Accepted Manuscript
Role of Intracellular Events in the Pathogenesis of Dengue; an Overview
Bhawana Jain, Umesh. C. Chaturvedi, Amita Jain
PII: S0882-4010(14)00037-0
DOI: 10.1016/j.micpath.2014.03.004
Reference: YMPAT 1473
To appear in: Microbial Pathogenesis
Received Date: 27 December 2013
Revised Date: 17 March 2014
Accepted Date: 20 March 2014
Please cite this article as: Jain B, Chaturvedi UC, Jain A, Role of Intracellular Events inthe Pathogenesis of Dengue; an Overview, Microbial Pathogenesis (2014), doi: 10.1016/j.micpath.2014.03.004.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Title: Role of Intracellular Events in the Pathogenesis of Dengue; an Overview 1
AUTHORS: Bhawana Jain*, Umesh. C. Chaturvedi*# and Amita Jain* 2
*Department of Microbiology, K.G. Medical University, Lucknow, India 3
# Indian Council of Medical Research, DHR, New Delhi, India 4
5
Address for correspondence: Dr Amita Jain, Professor, Department of Microbiology, K.G. 6
Medical University, Lucknow, India 7
e-mail: 8
Bhawana Jain – [email protected] (91 9450391235) 9
Umesh. C. Chaturvedi - [email protected] (91 9450913506) 10
Amita Jain – [email protected] (91 9415023928) 11
12
Word Count of Main text (including legend to figure, excluding tables, figure and 13
references): 4,305 14
Word Count of Abstract: 155 15
No. of Tables: 3 16
No. of figures: 1 17
No. of references: 99 18
-----------------------------------------------------------------------------------------------------. 19
20
21
22
#UCC is a Scientific Consultant of Indian Council of Medical Research, New Delhi 23
24

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Abstract 25
Dengue is one of the most important mosquito-borne viral diseases that are relentlessly 26
spreading in newer areas in the tropical and subtropical regions of the World. In last fifty 27
years, in spite of intensive and extensive investigations, pathogenesis of dengue is still not 28
clearly understood. Recently, the research focus is on studying the role of intracellular events 29
in pathogenesis of viral infections. Entry of virion in the host cell is followed by quick 30
succession of events, unfolded protein response, lipid bodies and lipophagy, endoplasmic 31
reticulum stress and recent demonstration of autophagy. The turbulence caused by these 32
events may result in clearance of the virus/ enhanced replication and survival of the host cell/ 33
apoptosis. Both, increased virus load and apoptosis of host cell may have pathological effects 34
on the host. In the present review, we have summed up the role of various intracellular events 35
in viral infections with special emphasis on Dengue virus infection. 36
37
38
Key Words: Dengue/ Autophagy/ Stress Granules/ Processing Bodies/ Lipid droplets 39
40

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
1. Introduction 41
Dengue virus (DENV) affects human of any age group, worldwide, including India1. 42
According to a recent estimate, there are around 390 million (95% credible interval 284–528) 43
dengue infections per year, of which 96 million (95% credible interval 67–136) manifest 44
apparently2. Dengue viruses occur as four antigenically related but distinct serotypes, 45
transmitted to humans by Aedes aegypti mosquitoes. These viruses generally cause either a 46
benign syndrome; dengue fever (DF), or a severe capillary leakage syndrome; dengue 47
hemorrhagic fever/ dengue shock syndrome (DHF/DSS)3. According to WHO 2009 48
classification severe Dengue has been differentiated into three subcategories; severe vascular 49
leakage, severe bleeding, and severe organ dysfunction4. The cardinal feature of DHF/DSS is 50
increased vascular permeability without morphological damage to capillary endothelium. 51
This results in extensive plasma leakage in various serous cavities of the host including 52
pleural, pericardial and peritoneal cavities and tissue spaces in patients with DHF, who may 53
go into profound shock (DSS)3,5. A number of mechanisms of pathogenesis of DHF/ DSS 54
have been discussed including, enhancing antibodies6,7, T cell-mediated response8 including 55
regulatory T cells9, various soluble mediators including a unique Cytotoxic Factor10,11 and 56
cytokines8,12,13, immune complex disease, antibodies cross-reacting with vascular 57
endothelium3, complement and its products, selection of virulent strains, viral virulence and 58
role of host genetics14. Among these the most plausible ones are the enhancing antibodies, 59
shift from Th1 to Th2 cell response and the memory T cells in a secondary infection, 60
resulting in a cytokine tsunami5. Extensive research has been done for more than fifty years 61
in area of dengue pathogenesis; still, the precise mechanism of DHF/DSS is not well 62
understood. In the last ten years or so, the research focus has shifted to intracellular events 63
during viral infections including DENV that may be translated to understand the pathogenesis 64
of severe disease. This review presents an overview of the role of intracellular events during 65
viral infection with special reference to dengue infection. 66
67
2. DENV replication and intracellular events: 68
Biologically diverse cell types, starting from insect cells to highly evolved mammalian cells 69
like endothelium and hepatocytes can be infected by DENV. The first interaction of DENV 70
with its host cell occurs via several putative receptors. They play an important role in 71
capturing, concentrating and transmitting infectious virions, leading to a cascade of events 72
that trigger the virus-cell membrane fusion15,16. A low pH-triggered conformational change of 73
Envelope (E) protein in endosomes leads to virus entry into the host cell through endocytosis 74
and uncoating. Capsid is released into the cell cytoplasm, where it dissociates and release 75
viral genome. Genome is a single RNA molecule of positive polarity and contains single 76
open reading frame (ORF), which is translated into a single large polyprotein. Polyprotein is 77
targeted to the endoplasmic reticulum (ER), where it is processed by virus and host encoded 78
proteases to form three structural proteins (Capsid protein C, a precursor for the membrane 79
protein PreM and envelope protein E) and seven non-structural proteins (NS1, NS2a, NS2b, 80
NS3, NS4a, NS4b, NS5), which help in replication, polyprotein processing and virion 81
assembly17. The replication takes place in virus induced vesicular membrane structures 82
associated with ER. Capsid protein covers the copies of genomes. Immature virions having 83
preM and E proteins on the surface bud into ER lumen18 and then transported through the 84
trans-Golgi network where cleavage of pre/M occurs making the virions infectious and 85
mature and is transported out of the cell by exocytosis19. 86

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
DENV exists as four heterologous serotypes termed serotypes 1, 2, 3 and 4 (DENV 1-4). 87
People once infected with one serotype of dengue virus are usually protected lifelong from 88
subsequent infection with the same serotype (homotypic infection)6. DHF occurs mostly in 89
persons infected with a second DENV serotype after an initial “primary” DENV infection 90
with a different serotype. The antibodies from the previous infection bind to the virus and 91
enhance its uptake by certain Fc receptor bearing monocytes/macrophages cells resulting in 92
high levels of viraemia. This phenomenon is known as antibody dependent enhancement 93
(ADE)6,7,20. Secondary infections cause forty times more DHF cases than primary infections. 94
There are many mechanisms suggested for DHF/DSS but nothing elaborates the pathogenesis 95
precisely. Recent studies have reported important findings of intimate interaction between 96
DENV and the host cell, which can be helpful in elucidating pathogenesis, like; a) autophagy; 97
b) lipid droplets and Lipophagy; c) unfolded protein response; d) Stress Granules and 98
Processing bodies. Further, strategies used by the virus to resist innate antiviral responses 99
have been discovered. Various mechanisms with reference to DENV infection will be 100
reviewed here. 101
2.A Autophagy 102
Autophagy is an intracellular catabolic system which degrades cytoplasmic components 103
within lysosomes. It has a specific role in elimination of micro-organisms, clearance of 104
intracellular proteins and organelles, cell death, and antigen presentation etc.21. The process 105
of autophagy is initiated when cell is subjected to pathogen infection. The signaling pathway; 106
Phosphatidyl Inositol 3-kinase (PI3K) and Beclin 1, gets activated. Beclin 1 first gets 107
separated from Beclin 2 which is a known anti apoptotic as well as anti autophagic protein, 108
and then induces autophagy. Kovacs et al have shown that inhibition of autophagy via Beclin 109
1 gene deletion in T cells results into extensive apoptosis of these cells upon T cell Receptor 110
(TCR) stimulation22. Beclin 1-deficient animals do not mount autoreactive T-cell responses. 111
This pathway leads to the formation of autophagosome, which is a key organelle of 112
autophagy. Certain autophagy related proteins (Atg) help in autophagosome formation. 113
Autophagosomes are formed in response to a number of stimuli, including host- and 114
pathogen-derived molecules, including toll-like receptor ligands and cytokines. Autophagy 115
can itself regulate the production and secretion of cytokines23. Autophagosome fuses with 116
lysosomes where lysosomal hydrolase degrades the inner membrane of autophagosome and 117
contain cytoplasm derived materials, known as autolysosomes or autophagolysosomes24. 118
Bhattacharya and Eissa have reviewed the role of autophagy and autoimmunity and have 119
concluded that autophagy, could modulate the induction or exacerbation of autoimmune 120
processes25. 121
2.A.(1) Role of autophagy in viral infection 122
Autophagy has been studied in a large number of viruses26 (Table 1 and 2). Viruses adopt 123
different mechanisms to induce or inhibit autophagy. Autophagy induction takes place by 124
following mechanisms i) by increase in autophagic flux, ii) by binding to the surface of host 125
cells, iii) by other intracellular events e.g. Endoplasmic Reticulum (ER) stress and Unfolded 126
Protein Response (UPR) (Table 1). Autophagy is inhibited (Table 1) by interfering with 127
either autophagosome formation, maturation and degradation or by degrading autolysosomes. 128
129
130

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Table 1: Impact of Viral infection on Autophagy 131
Mechanisms Viruses Viral molecules responsible for autophagy
Host molecules affected
References
A. Autophagy induction
by increase in autophagic flux Sindbis virus
Hepatitis C virus
-
-
Increases LC3 II
Increases LC3 II
27
28
by binding to the surface of target cells
Human Herpesvirus 6
Adenovirus B and D
Measles Virus
HIV 1
Vesicular Stomatitis Virus
Gp H-L-Q complex
trimeric fiber knob domain
gp41 subunit of viral envelope protein
-
Gp VSV-G
Binds to CD46
Binds to CD46
Binds to CD46
Binds to CXCR4 (CD4+) T cells
binds to TLR 7
-
-
29
30
31
by Endoplasmic Reticulum (ER) stress and Unfolded Protein Response (UPR)
Hepatitis C virus
Dengue Virus
-
NS4A
Induces PERK, ATF6, IRE1
-
32
33
B. Autophagy inhibition
by interfering with autophagosome formation
Kaposi Sarcoma Herpes Virus Murine Gamma Herpes Virus 68
Herpes Simplex Virus 1
vFLIP
M11
ICP34.5
target Beclin-1
encode viral Bcl 2 homologue
encode viral Bcl 2 homologue
34
35
36
by interfering with autophagosome maturation or degradation
HIV 1
Influenza A virus
Nef
Matrix (M2)
interact with Beclin-1
interact with Beclin-1
37
38
by autolysosomal degradation
Coxsackie Virus B3
Polio Virus
-
-
-
-
39
40
LC3 II= microtubule-associated protein 1 light chain 3; TLR= Toll like receptor; PERK= Protein Kinase R like 132 eIF2α kinase; ATF6= Activating Transcription Factor 6; IRE 1= Inositol Requiring Enzyme 1; NS= 133 Nonstructural; NEMO= NF κ B essential modulator; IRGM= Immunity associated GTPase family M; CD= 134 cluster of differentiation 135
136
137

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPTTable 2: Impact of Autophagy on viral pathogenesis 138
Mechanisms Viruses Viral molecules responsible for autophagy
Host molecules affected References
A. Degradation of viruses
by stimulating Innate Immunity
Vesicular Stomatitis Virus
Coxsackie Virus B3
-
-
production of IFN α by TLR7
TLR3 stimulation
-
-
by SLRs (Sequestosome 1/p62-like receptors)
Sindbis Virus
Herpes Simplex Virus 1
Capsid
-
degraded by p62
inhibited by SMURF1, novel adaptor
27
41
by stimulating Adaptive Immune response
Herpes Simplex Virus 1
EBV
Influenza A virus
-
EBNA1 protein
Matrix (M2)
presentation to MHC-I molecule
presentation to MHC-II mol
activates MHC II and CD4+ T cells
42
43
44
B. Promotion of viral replication
by formation of membranous replication factories
Polio Virus
Hepatitis C virus
Rotavirus
2BC and 3A proteins
NS5A, NS5B, nascent viral RNA
NS4P protein
-
-
-
45
46
47
by production of energy Dengue Virus - ATP production by Lipophagy
48
by promoting viral assembly and release
Hepatitis B Virus
HIV 1
Picorna virus
HBsAg
Gag, matrix (MA) protein p17
-
effects envelopment of viral particles
-
promotes nonlytic release
49
37
40
by suppressing antiviral innate immunity
Measles Virus
Hepatitis C virus
HIV 1
Capsid protein
NS3
Nef
target IRGM
target IRGM
target IRGM
50
50
50
LC3 II= microtubule-associated protein 1 light chain 3; TLR= Toll like receptor; PERK= Protein Kinase R like 139 eIF2α kinase; ATF6= Activating Transcription Factor 6; IRE 1= Inositol Requiring Enzyme 1; NS= 140 Nonstructural; NEMO= NF κ B essential modulator; IRGM= Immunity associated GTPase family M; CD= 141 cluster of differentiation 142

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
143
Consequently autophagic induction/inhibition leads to either virus degradation or promotion 144
of viral replication. Autophagy degrades viruses by following mechanisms (Table 2); i) by 145
stimulating innate immunity through Toll Like Receptors (TLRs), ii) by Sequestosome 1/p62-146
like receptors (SLRs) which are autophagic adaptors defined as pattern recognition receptors 147
(PRRs) that can selectively target a variety of pathogens for autophagic degradation, iii) by 148
stimulating adaptive immune response through MHC-I and MHC-II molecules. 149
Viral replication can also be promoted by following mechanisms (Table 2); i) by forming 150
Double Membrane Vesicles (DMVs) which help in replication, ii) by producing energy 151
through the process of lipophagy (discussed in section 2.B) iii) by enhancing viral assembly 152
and release, iv) by suppressing antiviral innate immunity. Innate immunity can be suppressed 153
by delivering cytosolic pathogen-associated molecular patterns (PAMPs) into the proximity 154
of endosomal pattern recognition receptors (PRRs) and MHC loading compartments and also 155
by directly trapping and degrading virions. 156
2.A.(2) Autophagy during dengue virus (DENV) infection 157
Some of the members of family Flaviviridae including DENV use autophagy for enhancing 158
their replication. Apoptosis is induced in macrophages by DENV, which is the primary site 159
for DENV replication. However, infected hepatocytes and epithelial cells which are the 160
secondary target of DENV infection do not die51-53. DENV evades innate immunity by 161
promoting autophagy in liver and kidney cells54. DENV2 induces autophagy in vitro in 162
MDCK renal epithelial cells, hepatocyte cell lines and other cells as well. These findings 163
were confirmed by inhibiting autophagy by the drug, 3-methyladenine as well as by siRNA, 164
targeting autophagy gene expression, which led to inhibition of viral replication. DENV 165
induced autophagosomes co-localize with LAMP 1, a marker of lysosomal fusion54. Other 166
studies also reported co-localization of DENV NS1 protein with autophagosome, LAMP 1 167
and ribosomal protein L2855. M6P-R, one of the endosomal markers, co-localizes with 168
autophagosomes, indicating that autophagosome also fuse with endosomes forming 169
amphisome. Both, autophagosome and amphisome are double membrane vesicles (DMVs) 170
and DENV is known to replicate on virally induced DMVs. DENVs replication on 171
amphisomes possibly influences virus entry and replication. Inhibition of lysosomal fusion 172
with autophagosomes increases replication of DENV 255. NS 4A (Nonstructural protein 4A) 173
of DENV2 is the viral mediator of autophagy upregulation and protection against apoptosis33. 174
DENV utilizes PI3K dependent mechanism to upregulate autophagy. Proteins Beclin 1 and 175
Atg 5 are required for protection of virus infected cells55. Another study suggested that 176
autophagy stimulates DV replication by suppressing innate immune response of the host 177
through Unfolded Protein Response (UPR)/ autophagy pathway28. 178
Role of autophagy is also studied in DENV 3 infection. LAMP 1 co-localizes with 179
autophagosomes like DENV2 but lysosomal fusion inhibitor decreases DENV3 replication, 180
suggesting role of mature autophagosomes in DENV3 infection. Inhibition of formation of 181
autolysosomes by L-arginine, induces the accumulation of autophagosomes and amphisomes. 182
This mechanism stimulates DENV2 replication and inhibits DENV3 replication, suggesting 183
DENV2 uses amphisomes whereas DENV3 uses both amphisomes and autolysosomes for 184
viral RNA replication55,56. Welsch et al showed 3D structures of DENV2 replication 185
complex, which was actually the contiguous invagination of Endoplasmic reticulum (ER)57. 186
He further suggested indirect role of functional components of autophagy in DENV 187
replication, maturation and infectivity by inhibiting autophagy using spautin-1 (specific 188

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
and potent autophagy inhibitor-1), a novel inhibitor and stimulator of autophagy in DENV 189
infection58. Spautin-1 treatment generated heat sensitive and non infectious DENV particles 190
denoting major effect of autophagy on DENV cell cycle. 191
Monocytes are the critical cell in the dengue disease. They are rarely infected directly, but are 192
highly susceptible to antibody enhanced infection59. Panyasrivanit et al have reported that in 193
monocytic cells induction of autophagy reduced viral yield while inhibition of autophagy 194
only slightly increased levels of viruses and there are distinct cell type specific differences in 195
the DENV-autophagy interaction60. 196
There are several studies elucidating the role of autophagy in DENV replication, however, 197
the precise mechanism by which DENV induces autophagy is still unclear. More work is 198
required to further characterize the role(s) and contribution of autophagy to the various 199
aspects of DENV replication, in different cell types. 200
2.B Lipid Droplets and Lipophagy 201
Lipids are stored in a cell in the form of lipid droplets (LDs) and are metabolized by 202
cytoplasmic neutral hydrolases. Lipid droplets (LDs) are dynamic organelles containing 203
neutral lipid core. They arise from Endoplasmic Reticulum (ER) as a result of various stress 204
signals. Lipid metabolism occurring through the lysosomal degradative pathway of 205
autophagy, known as lipophagy, regulates intracellular lipid stores and cellular levels of free 206
lipids such as fatty acids and energy homeostasis. An important role of lipophagy is in the 207
maintenance of adequate levels of β-oxidation to supply ATP which may be responsible for 208
the regulation of cell resistance to death stimuli and in cellular trans-differentiation. Recently 209
LDs have received special attention as they have been found to be associated with viral 210
diseases (eg. HCV, HBV and Rotavirus). Studies on HCV, have shown the positive effect of 211
LDs and associated ER system in promoting viral replication61. Core protein of HCV has 212
been shown to be involved in LDs distribution in perinuclear region and helping viral 213
replication and assembly62-64. Study on DENV demonstrated that LDs sequestrated unfolded 214
proteins and prevented the accumulation of lipotoxic non-esterified fatty acid by diverting 215
them to cytosol for degradation65. Later on Capsid protein of DENV was observed to be 216
present over LDs. In early stages of infection, capsid protein sequestration takes place while 217
later on; LDs serve as platform for assembly of virus. Therefore LDs have indirect role on 218
DENV replication by regulating the capsid protein availability66. Another study also 219
demonstrated the role of ER rearrangement and expansion in early stages of DENV infection 220
which involved reabsorption of LDs67. 221
Lipophagy functions in a number of different cell types, and appears to be a common 222
pathway of cellular lipid metabolism68. Lipophagy eliminates intracellular pathogens, but 223
viruses may subvert the lipophagic pathway to promote their own survival. For example, 224
hepatitis C virus (HCV)69 and hepatitis B virus70 upregulate autophagy which in turn 225
promotes viral replication. The core protein of HCV is known to attach to LD for promoting 226
viral assembly through an unknown mechanism71. Studies with DENV have demonstrated 227
that autophagy increases viral replication through lipophagy. Further, upregulation of 228
autophagy promotes lipid breakdown for supply of energy for viral replication72. DENV 229
infection of hepatoma cells promote the formation of autophagosomes which associate with 230
LDs resulting in stimulation of lipophagy in infected cells. Reduction of lipid droplets as well 231
as cellular triglycerides upon DENV infection indicates that regulation of lipid metabolism by 232

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
autophagy is a major component facilitating DENV replication. Cellular triglycerides that are 233
stored in lipid droplets are depleted by DENV-induced autophagosomes, resulting into 234
increased β-oxidation and energy production. This showed that a virus can trigger autophagy 235
to modulate cellular physiology48,72. Heaton et al have identified three cellular pathways 236
needed for DENV replication: autophagy, actin polymerization, and fatty acid biosynthesis48. 237
Further, a key enzyme in this pathway, fatty acid synthase (FASN), is relocalized to sites of 238
DENV replication by DENV nonstructural protein 3 (NS3). Purified recombinant NS3 also 239
stimulates the activity of FASN in vitro. Thus, it is suggested that DENV co-opts the fatty 240
acid biosynthetic pathway to establish its replication complexes48. In conclusion, this novel 241
mechanism of lipophagy helps DENV not only to promote replication but also to evade 242
immune system of the host by channelizing autophagosomes towards lipid metabolism73. 243
However, despite our improved understanding of lipid metabolism and ER rearrangement, 244
more research is required to precisely delineate the steps leading to DENV replication 245
promotion by LDs or Lipophagy. This will help in development of new therapeutic 246
modalities against DENV. Inhibition of FASN or any of the enzymes/ proteins of autophagy 247
pathway can inhibit DENV replication73. 248
2.C Unfolded Protein Response (UPR) 249
The unfolded protein response (UPR) is cellular response to ER stress which causes 250
accumulation of unfolded or misfolded proteins in the lumen of the endoplasmic reticulum 251
(ER) in a stressed cell. In virus-infected cells, the cellular translation machinery is 252
orchestrated by the infecting virus to produce large amounts of viral proteins, which 253
ultimately perturbs ER homeostasis and causes ER stress74. UPR restores normal function of 254
the cell by attenuating protein translation and activates the signaling pathways to increase the 255
production of molecular chaperones required in protein folding. If these aims are not 256
achieved, the UPR leads to apoptosis. Several viruses are shown to induce UPR. 257
Herpesviruses induce ER stress and activate UPR signaling pathways which help in assembly 258
of infectious particles75. HCV induces the UPR, which activates autophagic pathway by 259
initiating autophagosome formation76. Recently, it has been discovered that autophagy plays 260
a regulatory role in controlling the innate immune response against intracellular pathogens77. 261
HCV induces autophagy through UPR activation. Autophagy is required for promoting HCV 262
RNA replication in human hepatoma cells. This is achieved by suppressing innate antiviral 263
immunity28 through upgradation of HCV-derived pathogen-associated molecular pattern 264
(PAMP), which mediates anti viral response. DENV, like HCV, also utilizes the ER or ER-265
derived membrane structure as the primary site of RNA replication and assembly. Thus, it is 266
conceivable that DENV infected cells experience ER stress and the UPR. Ke and Chen 267
(2011) in the same study showed suppression of DENV-PAMP RNA-induced IFN β 268
promoter activation by UPR inducers, which causes exploitation of UPR-autophagy pathway 269
to escape innate immune response in both DENV and HCV infections28. DENV infection of 270
A549 cells elicits an UPR at the level of translation attenuation and activation of specific 271
pathways. Specific serotypes of DENV can modulate the UPR with different selectivity. The 272
modulators of UPR such as Salubrinal inhibit DENV replication78. Pena and Harris have 273
reported a time-dependent activation of the UPR by DENV-2 which can override inhibition 274
of translation, prevent apoptosis, and prolong the viral life cycle79. Other studies have shown 275
that modulation of intracellular membrane architecture of the cell early after DENV-2 276
infection is driven by viral protein expression and does not require the induction of the UPR 277
and sterol-regulatory-element-binding-protein-2 (SREBP-2) pathways67. Studies on the 278
impact on UPR following direct or ADE infection of THP-1 cells with all the four serotypes 279

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
of DENV showed that the ADE infection with epidemic DENV-2 (high-UPR-gene-280
expressing strains) correlated with severe disease80. 281
2.D Stress Granules and Processing Bodies 282
Viral infection, along with other types of stresses, alters the cells’ protein expression system. 283
In order to adapt to the stress, the supressed expression of constitutive proteins lead to change 284
in translation pathway and diversion of mRNA from polysomes to cytoplasmic granules. 285
These granules are of various types in eukaryotic cells but Stress granules (SG) and 286
Processing bodies (PB) are of special interest in virus infection. 287
During cell stress, untranslated mRNA accumulates in discrete cytoplasmic foci called stress 288
granules (SGs) that have roles in inhibition and degradation of host mRNAs. SGs appear to 289
protect RNAs from harmful conditions, and may safe-guard the information coded in their 290
RNA sequence. These molecules can be stored, degraded, or re-initiate translation81. 291
Processing bodies (P-bodies, PB) are another type of granules packed with translationally 292
silenced mRNPs and enzymes of RNA decay machine. PBs may promote SG assembly82,83. 293
SGs and PBs are actively linked and are believed to transiently bind and exchange mRNP 294
along with remodeling of the mRNP protein constituents, playing roles in decay of unwanted 295
mRNA, nonsense mRNA, AU-rich element mediated mRNA and silencing of microRNA 296
induced mRNA. 297
2.D.(1) Role of SG and PB in viral replication 298
Virus infection interferes with a number of cell processes which directly induce stress 299
responses such as transient global inhibition of protein synthesis to enhance cell survival by 300
restricted consumption of nutrients and energy. Although SG formation is frequently induced 301
by virus infection, many differences exist in the dynamics and outcome of the stress 302
responses induced by various viruses84. Viruses have evolved measures to prevent 303
sequestration of viral mRNA into translationally silenced mRNPs that aggregate in SG and 304
get decayed in PBs. Virus families can be organized into groups on the basis of mechanisms 305
adopting to facilitate their replication by modulating SGs and PBs (Table 3). The gene 306
expression and resources to damage repair pathways may be redirected. The mechanism of 307
SG formation is not understood clearly but may involve remodeling of mRNP which 308
incorporates new proteins. These proteins nucleate SGs and affect mRNP transport on 309
microtubules. Quick reactivation of translation on recovery from stress may be facilitated by 310
SGs. SGs sequester components of apoptotic signal transduction pathways, therefore, may 311
help in cell survival during stress85. Intracellular stress occurs during viral infection which 312
reduces translation through activation of enzymes like eIF2 kinases and changes active 313
polysome mRNPs into stalled translation initiation complex mRNPs. Nucleation of several 314
stress granule marker proteins (viz. G3BP, Tia-1/TIAR, and HDAC6 plus) transport on 315
microtubules (MT) resulting into aggregation of translation initiation complex mRNPs in 316
stress granules, through a series of complex steps. Decapping and decay can occur within or 317
outside PBs. 318
319
320
321

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Table 3: Stress Granules (SGs) and Processing Bodies (PBs) studied in different viral 322
infections 323
Viruses Involved molecules and processes References 1. Inhibition of Stress Granules (SGs)
A. Activation of eIF2α kinase
Polio Virus - 86
Alpha Virus - - Orthoreo Virus - - CMV eIF4G mediated 87
Rotavirus eIF4E, S6, eIF2α phosphorylation 88
Influenza A Virus blocks eIF2α phosphorylation 89
Other Reovirideae eIF4G, eIF4E, eIF3
90,91
Chikungunya Virus - 92
B. Blocking loss of cellular SG formation in response to Oxidative stress
Polio virus Cleavage of SG nucleating protein G3BP, PABP
86,93
West Nile Virus Sequestration of SG nucleating protein TIA-1/TIAR
94
Dengue Virus Sequestration of SG nucleating protein TIA-1/TIAR, G3BP1, Caprin 1, USP 10
94,95
Herpes Simplex Virus 1
Sequestration of SG nucleating protein TIA-1/TIAR
96
HTLV 1 Sequestration of SG nucleation and maintenance protein HDAC 6
97
HIV 1 Sequestration of PABP 1,Staufen 1 98
Rotavirus Sequestration of TIA-1 88
Other Reovirideae Sequestration of TIA-1, TIAR, G3BP 90,91
2. Disruption of Processing Bodies (PBs) Hepatitis C virus Binds with DDX 3, RCK/p 54, Lsm
1, Pat L1
99
Polio Virus - 86,93
Coxsackie Virus - - West Nile Virus - -
3. Alteration of Stress Granule (SG) response Respiratory Syncytial Virus
eIF2α phosphorylation by PKR 100
Vaccinia Virus subvert SG-nucleating proteins 101
324
G3BP= Ras-Gap-SH3 domainbinding protein; TIA-1= T cell intracellular antigen- ; TIAR= T cell 325
intracellular antigen-1 related protein; eIF2α = eukaryotic translation initiation factor 2α; PKR= 326
protein kinase R 327
328
2.D.(2) SGs and PBs in dengue virus (DENV) infection 329

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
DENV is a single stranded positive sense RNA of ≈ 11 kb containing single ORF and is the 330
only viral mRNA produced during viral replication. DENV RNA replication complexes are 331
shown to locate in the perinuclear region of infected cells. Researchers have observed co-332
localization of SG nucleating protein i.e. T cell intracellular antigen-1 (TIA-1) and related 333
protein (TIAR) with DV NS3 and dsRNA in perinuclear region only94. The kinetics of TIAR 334
accumulation was found to be similar to genomic RNA synthesis. DENV components 335
interact with TIAR, facilitate viral RNA synthesis and inhibit SG formation. This prevents 336
stallation of host translation machinery. Co localization of many proteins related to SGs 337
(G3BP 1, G3BP 2, Caprin 1 and USP 10) and PBs (DDX 6 also known as RCK/p 54) with 338
DENV serotype 2 viral components is known95. These proteins bind to DENV at 3’UTR 339
region at different sites, but 3’UTR region of DENV 2 is an important site for PB and SG 340
assembly and DDX 6 had been demonstrated as a pro-viral protein helping in assembly or 341
release of infectious particles. 342
Such studies demarcate the definitive role of SG and PB in DV infection and it is not anti 343
viral, since it is enhancing DV survival. However, further studies may delineate the exact 344
mechanism and help the investigators to find some robust tool to control dengue by 345
modulating these bodies. 346
3. Summary of Intracellular Events and DHF/DSS pathogenesis 347
A number of mechanisms have been considered to explain the pathogenesis of DHF/DSS. 348
The most favored ones are increased virus load by replication via enhancing antibodies, virus 349
virulence, shift from Th1 to Th2 cell response and the memory T cells in a secondary 350
infection, resulting in cytokine tsunami producing severe dengue disease. However, 351
everything is not explained by these mechanisms, therefore a fascinating hypothesis emerged, 352
which stated that intracellular events and pathways may have significant role in DENV 353
infection; mild or severe. The intracellular event that follows entry of DENV in the host cells 354
may indicate the outcome in the form of recovery or severe disease. Viruses are obligate 355
intracellular parasites. Their survival and expansion depends on the health of the host cells 356
providing necessary supporting factors to the viruses. In virus-infected cells, the cellular 357
translation machinery is hijacked by the infecting virus to produce large amounts of viral 358
proteins. The accumulation of unfolded or misfolded proteins, leads to highly specific 359
signaling pathways i.e. the UPR to restore normal ER functions. If this overload is not 360
resolved, the UPR induces ER stress-associated programmed cell death, apoptosis. 361
Autophagy, LD formation and Lipophagy are also among these defensive mechanisms of 362
host against viral infection. To combat such defense system of host, certain viruses like 363
DENV, modulate such events in its favor and use such system to enhance its replication or 364
survival, as an adaptation trick. NS4A protein of DENV induces autophagy by UPR. The 365
UPR activates autophagic pathway which promotes virus replication in cells. This increased 366
viral replication is favored by formation of membranous replication factories also known as 367
double membrane vesicles (DMV) or by production of energy. DMVs provide platform for 368
viral replication. In DENV infection, lipid metabolism leading to the production of ATP by β 369
oxidation (Lipophagy) promotes viral replication. Innate immunity of the host is also 370
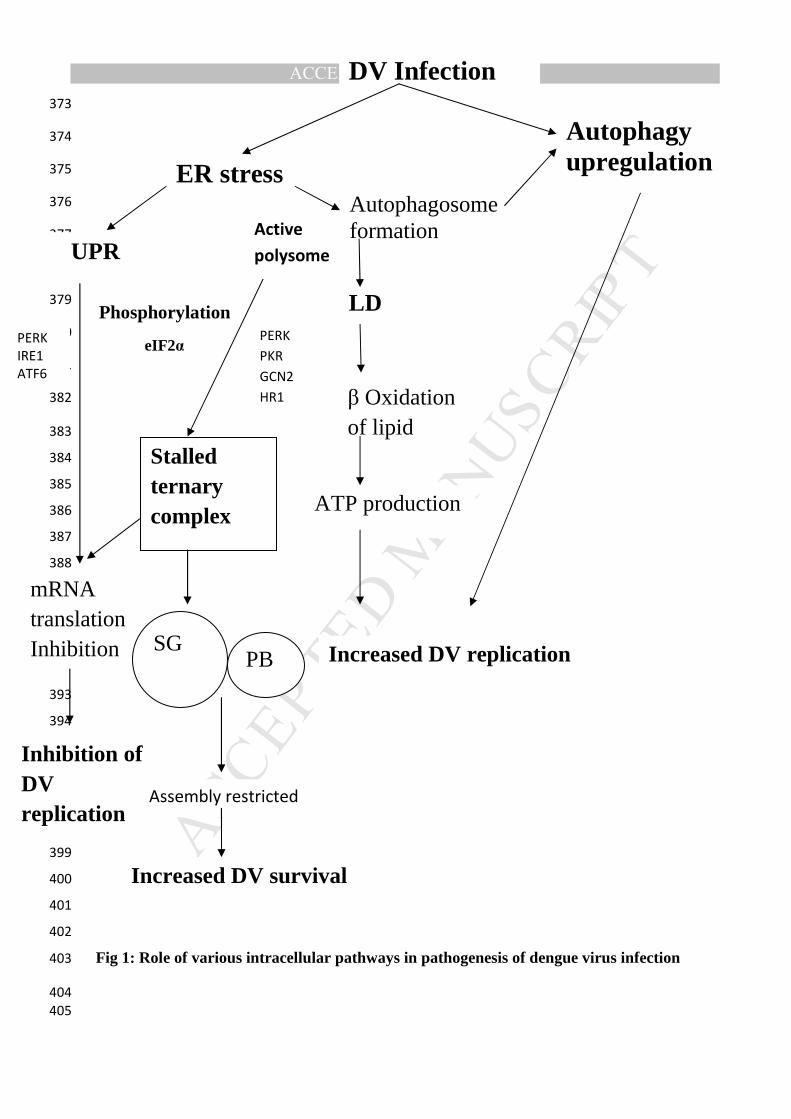
tempered by these events to help DENV infection (Figure 1). 371
372
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT 373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
Fig 1: Role of various intracellular pathways in pathogenesis of dengue virus infection 403
404
405
DV Infection
Autophagy upregulation ER stress
Autophagosome formation
UPR
Phosphorylation
eIF2α
Stalled ternary complex
mRNA translation Inhibition
LD
β Oxidation of lipid
ATP production
Increased DV replication SG
PB
Active
polysome
PERK
IRE1
ATF6
PERK
PKR
GCN2
HR1
Inhibition of DV replication
Assembly restricted
Increased DV survival

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Legend to Figure 1: 406
407
In response to stress caused by DV infection, Endoplasmic reticulum (ER) of host cells, 408
which is otherwise responsible for the synthesis, maturation and proper folding of a wide 409
range of proteins, leads to the induction of inter-organelle signaling pathways. These 410
pathways regulate translation and gene expression through an elaborate adaptive response 411
known as the unfolded protein response (UPR). The UPR functions through the PKR-like ER 412
kinase (PERK), kinase/ endoribonuclease inositol requiring 1 (IRE1)/X-box binding protein 1 413
(XBP1), and/or activation transcription factor 6 (ATF6) pathways which together inhibit 414
translation. PERK along with PKR (Protein Kinase R), GCN2 and HRI lead to 415
phosphorylation of eukaryotic translation initiation factor (eIF) 2α (eIF2α) and arrest of 416
translation due to stalled ternary complex (2(eIF2)+GTPNmet-tRNAi met). The 417
nontranslated mRNA along with proteins is organized into specialized RNA granules, Stress 418
Granules (SGs) and Processing Bodies (PBs). DV sequesters SG proteins thus restricting SG 419
assembly and therefore enhancement in DV survival into host cells occur. Another pathway 420
used by DV is Autophagy which involves sequestration of intact organelles (e.g., 421
mitochondria) and portions of the cytosol via a membrane referred to as the phagophore 422
(derived from ER). This phagophore expands to form double-membrane vesicles, termed 423
autophagosomes. Subsequently, autophagosomes mature by fusing with endosomes and/or 424
lysosomes to form autolysosomes, where degradation of the internal contents occurs by 425
resident lysosomal hydrolases. DV induces degradation of LDs via autophagy in order 426
tomobilize triglycerides that are used for energy (ATP) production via β-oxidation. 427
428
DV: Dengue virus; LD: Lipid Droplets; UPR: Unfolded protein response; ER: Endoplasmic 429
reticulum; SG: Stress Granules; PB: Processing Bodies 430
431
432
4. Conclusions 433
The pathogenesis of dengue is very complex. Impact of intracellular events including 434
autophagy, stress granules and processing bodies, unfolded protein response etc. is not yet 435
fully elucidated in the pathogenesis of dengue. Impact of Dengue virus infection on various 436
intracellular processes is highly complex too and needs to be explained in detail. Delineation 437
of interaction of DENV with each mechanism upto the molecular level is highly needed to 438
find the suitable targets for novel and highly promising anti viral therapeutic modalities, 439
being introduced each day in the scientific field. 440
Acknowledgements 441
Authors acknowledge financial support from ICMR Grant no. 5/8/7/14/2009-ECD-I phase-II 442
and ICMR Grant no. 5/8/7/17/2010-ECD-I. UCC is Scientific Consultant of ICMR/DHR, 443
New Delhi. 444
Conflict of interest: None 445
446

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
References: 447
448
1. Gupta N, Srivastava S, Jain A, Chaturvedi UC (2012) Dengue in India. Indian J Med 449
Res136(3):373-90 450
2. Bhatt S, Gething PW, Brady OJ et al (2013) The global distribution and burden of 451
dengue. Nature, DOI:10.1038/nature12060 452
3. Basu A, Chaturvedi UC (2008) Vascular endothelium: the battlefield of dengue 453
viruses. FEMS Immunol. Med. Microbiol53:287–299. 454
4. World Health Organization, 2009. Dengue: Guideline for Diagnosis, Treatment, 455
Prevention and Control. Geneva: World Health Organization 456
5. Jain A, Chaturvedi UC (2010) Dengue in infants: an overview. FEMS Immunol Med 457
Microbiol59(2):119-30. 458
6. Halstead SB (2003) Neutralization and antibody-dependent enhancement of dengue 459
viruses. Adv Virus Res60:421-467 460
7. Halstead SB (2009) Antibodies determine virulence in dengue. Ann N Y Acad Sci 461
1171 Suppl 1:E48-56. 462
8. Chaturvedi UC, Raghupathy R, Pacsa AS et al (1999) Shift from a Th1-Type 463
Response to Th2-Type in Dengue Haemorrhagic Fever Curr Sci76; 63-69. 464
9. Chaturvedi UC, Shrivastava S, Tripathi RK, Nagar R (2007) Dengue virus-specific 465
suppressor T cell: Current perspectives. FEMS Immunol Med50: 285-299. 466
10. Mukerjee R, Chaturvedi UC, Dhawan R (1995) Dengue Virus-induced human 467
cytotoxic factor: production by peripheral blood leucocytes in vitro. Clin Exp 468
Immunol101:261-267. 469
11. Mukerjee R, Chaturvedi UC, Vaughn DW, Kalayanarooj S (1997) Purification and 470
pathogenicity of the cytotoxic factor from the cases of dengue haemorrhagic fever. 471
Curr Sci72:494-501. 472
12. Chaturvedi UC, Agarwal R, Elbishbishi EA, Mustafa AS (2000) Cytokine Cascade 473
in Dengue Haemorrhagic Fever: Implications for Pathogenesis. FEMS Immunol Med 474
Microbiol28:183-188 475
13. Rathakrishnan A, Wang SM, Hu Y, Khan AM, Ponnampalavanar S, Lum 476
LC, Manikam R, Sekaran SD (2012) Cytokine expression profile of dengue patients at 477
different phases of illness. PLoS One7(12):e52215. doi: 478
10.1371/journal.pone.0052215. 479
14. Chaturvedi UC, Nagar R, Shrivastava S (2006a) Dengue and dengue haemorrhagic 480
fever: implications of host genetics. FEMS Immunol Med Microbiol47:155-166. 481
15. Basu A, Chaturvedi UC (2010) Recent advances in understanding the intracellular 482
responses to dengue virus infection (Invited editorial) Future Virol5:255–257. 483
16. Chaturvedi UC, Nagar R, Shrivastava S (2006b) Macrophage and dengue virus: friend 484
or foe? Indian J Med Res124:23-40 485
17. Bartenachlager R, Miller S. Molecular aspects of Dengue virus replication. Future 486
Microbiol. 2008 Apr;3:155-165 487
18. Mukhopadhyay S, Kuhn RJ, Rossmann MG. A structural perspective of the flavivirus 488
life cycle. Nat. Rev. Microbiol. 2005Jan;3(1):13-22 489

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
19. Mackenzie JM, Jones MK, Westaway EG. Markers for trans Golgi membranes and 490
the intermediate compartment localize to induced membranes with distinct replication 491
functions in flavivirus-infected cells. J. Virol. 1999 Nov;73(11):9555-9567 492
20. Chaturvedi UC, Shrivastava, R, Nagar, R.(2005) Dengue Vaccines: Prospects and 493
Problems. Indian J Med. Res121:639-52 494
21. Mizushima N, Yamamoto A, Matsui M, Yoshimori T, Ohsumi Y (2004) In vivo 495
analysis of autophagy in responose to nutrient starvation using transgenic mice 496
expressing a fluorescent autophagosome marker. Mol Biol Cell15:1101–1111. 497
22. Kovacs JR, Li C, Yang Q, Li G, Garcia IG, Ju S, Roodman DG, Windle JJ, Zhang. X, 498
Lu B. (2012) Autophagy promotes T-cell survival through degradation of proteins of 499
the cell death machinery. Cell Death Differ. 19:144-152. 500
23. Harris J. (2011). Autophagy and cytokines. Cytokine. ;56:140-144. 501
24. Mizushima, N. Autophagy: Process and function. Genes. Dev. 2007, 21, 2861-2873. 502
25. Bhattacharya A, Eissa NT. (2013). Autophagy and autoimmunity crosstalks. Front. 503
Immunol. 15;4:88. doi: 10.3389/fimmu.2013.00088. 504
26. Chiramel AI, Brady NR, Bartenschlager R (2013) Divergent Roles of Autophagy in 505
Virus Infection. Cells2:83-104; doi:10.3390/cells2010083 506
27. Orvedahl A, MacPherson S, Sumpter R Jr, Talloczy Z, Zou Z, Levine B (2010) 507
Autophagy protects against Sindbis virus infection of the central nervous system. Cell 508
Host Microbe7:115-127. 509
28. Ke PY, Chen SS (2011) Activation of the unfolded protein response and autophagy 510
after hepatitis C virus infection suppresses innate antiviral immunity in vitro. J Clin 511
Invest121:37-56. 512
29. Joubert PE, Meiffren G, Gregoire IP et al (2009) Autophagy induction by the 513
pathogen receptor CD46. Cell Host Microbe6: 354-366. 514
30. Denizot M, Varbanov M, Espert L, Robert-Hebmann V, Sagnier S, Garcia E, Curriu 515
M, Mamoun R.and Blanco J, Biard-Piechaczyk, M (2008) HIV-1 gp41 fusogenic 516
function triggers autophagy in uninfected cells. Autophagy 008, 4, 998-1008. 517
31. Nakamoto M, Moy RH, Xu J, Bambina S, Yasunaga A, Shelly SS, Gold B, Cherry S 518
(2012) Virus recognition by Toll-7 activates antiviral autophagy in Drosophila. 519
Immunity36:658-667. 520
32. Kroemer G, Marino G, Levine, B (2010) Autophagy and the integrated stress 521
response. Mol Cell40: 280-293. 522
33. McLean JE, Wudzinska A, Datan E, Quaglino D, Zakeri Z (2011) Flavivirus NS4A-523
induced autophagy protects cells against death and enhances virus replication. J Biol 524
Chem286:22147-22159. 525
34. Liang CEX, Jung JU (2008) Downregulation of autophagy by herpesvirus Bcl-2 526
homologs. Autophagy4: 268-272. 527
35. Ku B, Woo JS, Liang C, Lee KH, Hong HS, E X, Kim KS, Jung JU, Oh BH (2008) 528
Structural and biochemical bases for the inhibition of autophagy and apoptosis by 529
viral BCL-2 of murine gamma-herpesvirus 68. PLoS Pathog 4, e25. 530
36. Orvedahl A, Alexander D, Talloczy Z, Sun Q, Wei Y, Zhang W, Burns D, Leib DA, 531
Levine B (2007) HSV-1 ICP34.5 confers neurovirulence by targeting the Beclin 1 532
autophagy protein. Cell Host Microbe1: 23-35. 533

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
37. Kyei GB, Dinkins C, Davis AS et al (2009) Autophagy pathway intersects with HIV-534
1 biosynthesis and regulates viral yields in macrophages. J Cell Biol186: 255-268. 535
38. Gannage M, Dormann D, Albrecht R et al (2009) Matrix protein 2 of influenza A 536
virus blocks autophagosome fusion with lysosomes. Cell HostMicrobe6: 367-380. 537
39. Kemball CC, Alirezaei M, Flynn CT, Wood MR, Harkins S, Kiosses WB, Whitton JL 538
(2010) Coxsackievirus infection induces autophagy-like vesicles and 539
megaphagosomes in pancreatic acinar cells in vivo. JVI84: 12110-12124. 540
40. Jackson WT, Giddings TH, Jr, Taylor MP, Mulinyawe S, Rabinovitch M, Kopito RR, 541
Kirkegaard K (2005) Subversion of cellular autophagosomal machinery by RNA 542
viruses. Plos Biol3: e156. 543
41. Orvedahl A, Sumpter R Jr, Xiao G et al (2011) Image-based genome-wide siRNA 544
screen identifies selective autophagy factors. Nature480: 113-117. 545
42. English L, Chemali M, Duron J, Rondeau C, Laplante A, Gingras D, Alexander D, 546
Leib D, Norbury C, Lippe R, Desjardins M (2009) Autophagy enhances the 547
presentation of endogenous viral antigens on MHC class I molecules during HSV-1 548
infection. Nat Immunol10: 480-487. 549
43. Paludan C, Schmid D, Landthaler M, Vockerodt M, Kube D, Tuschl T, Munz C 550
(2005) Endogenous MHC class II processing of a viral nuclear antigen after 551
autophagy. Science307: 593-596. 552
44. Schmid D, Pypaert M, Munz C (2007) Antigen-loading compartments for major 553
histocompatibility complex class II molecules continuously receive input from 554
autophagosomes. Immunity26:79-92. 555
45. Schlegel A, Giddings TH Jr, Ladinsky MS, Kirkegaard K (1996). Cellular origin and 556
ultrastructure of membranes induced during poliovirus infection. JVI70: 6576-6588. 557
46. Sir D, Kuo CF, Tian Y, Liu HM, Huang EJ, Jung JU, Machida K, Ou JH (2012) 558
Replication of hepatitis C virus RNA on autophagosomal membranes. J Biol 559
Chem287: 18036-18043. 560
47. Berkova Z, Crawford SE, Trugnan G, Yoshimori T, Morris, AP, Estes MK (2006). 561
Rotavirus NSP4 induces a novelvesicular compartment regulated by calcium and 562
associated with viroplasms. JVI, 80, 6061-6071. 563
48. Heaton NS, Randall G (2011) Dengue virus and autophagy. Viruses3(8):1332-341. 564
49. Li J, Liu Y, Wang Z, Liu K, Wang Y, Liu J, Ding H, Yuan Z (2011) Subversion of 565
cellular autophagy machinery by hepatitis B virus for viral envelopment. JVI 85: 566
6319-6333. 567
50. Gregoire IP, Richetta C, Meyniel-Schicklin L et al (2011) IRGM is a common target 568
of RNA viruses that subvert the autophagy network. PLoS Pathog7: e1002422. 569
51. Tonry J. H., Xiao S. Y., Siirin M., Chen H., da Rosa A. P., Tesh R. B. (2005) Am. J. 570
Trop. Med. Hyg. 72, 320–324. [PubMed: 15772329] 571
52. Siirin M. T., Duan T., Lei H., Guzman H., da Rosa A. P., Watts D. M., Xiao S. Y., 572
Tesh R. B. (2007) Am. J. Trop. Med. Hyg. 76, 299–306. [PubMed: 17297039] 573
53. Murray K., Walker C., Herrington E., Lewis J. A., McCormick J., Beasley D. W., 574
Tesh R. B., Fisher-Hoch S. (2010) J. Infect. Dis. 201, 2–4.[PMCID: 575
PMC2791189] [PubMed: 19961306] 576

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
54. Lee YR, Lei HY, Liu MT, Wang JR, Chen SH, Jiang-Shieh YF, Lin YS, Yeh TM, Liu 577
CC, Liu HS (2008) Autophagic machinery activated by dengue virus enhances virus 578
replication. Virology374:240–248 579
55. Panyasrivanit M, Khakpoor A, Wikan N, Smith DR (2009) Co-localization of 580
constituents of the dengue virus translation and replication machinery with 581
amphisomes. J Gen Virol90:448–456 582
56. Khakpoor A, Panyasrivanit M, Wikan N, Smith DR (2009) A role for 583
autophagolysosomes in dengue virus 3 production in HepG2 cells. J GenVirol90: 584
1093–1103 585
57. Welsch S, Miller S, Romero-Brey I, Merz A, Bleck CK, Walther P, Fuller SD, 586
Antony C, Krijnse-Locker J, Bartenschlager R (2009) Composition and three-587
dimensional architecture of the dengue virus replication and assembly sites. Cell Host 588
Microbe5:365-375. 589
58. Mateo R, Nagamine CM, Spagnolo J, Méndez E, Rahe M, Gale M Jr, Yuan 590
J, Kirkegaard K (2013) Inhibition of cellular autophagy deranges dengue virion 591
maturation. J Virol87(3):1312-21 592
59. Halstead SB, Mahalingam S, Marovich MA, Ubol S, Mosser DM (2010) Intrinsic 593
antibody-dependent enhancement of microbial infection in macrophages: disease 594
regulation by immune complexes. Lancet Infect Dis10(10):712-22 595
60. Panyasrivanit M, Greenwood MP, Murphy D, Isidoro C, Auewarakul P, Smith DR 596
(2011) Induced autophagy reduces virus output in dengue infected monocytic 597
cells. Virology418:74–84. 598
61. Roingeard P, Depla M (2011) The birth and life of lipid droplets: learning from the 599
hepatitis C virus. Biol Cell 103: 223–231. 600
62. Roingeard P, Hourioux C, Blanchard E, Prensier G (2008) Hepatitis C virus budding 601
at lipid droplet-associated ER membrane visualized by 3D electron microscopy. 602
Histochem Cell Biol 130: 561–566. 603
63. Boulant S, Douglas MW, Moody L, Budkowska A, Targett-Adams P, et al. (2008) 604
Hepatitis C virus core protein induces lipid droplet redistribution in a microtubule- 605
and dynein-dependent manner. Traffic 9: 1268–1282. 606
64. Miyanari Y, Atsuzawa K, Usuda N, Watashi K, Hishiki T, et al. (2007) The lipid 607
droplet is an important organelle for hepatitis C virus production. Nat Cell Biol 9: 608
1089–1097. 609
65. Hapala I, Marza E, Ferreira T (2011) Is fat so bad? Modulation of endoplasmic 610
reticulum stress by lipid droplet formation. Biol Cell 103: 271–285. 611
66. Samsa MM, Mondotte JA, Iglesias NG, Assuncao-Miranda I, Barbosa-Lima G, Da 612
Poian AT et al (2009) Dengue virus capsid protein usurps lipid droplets for viral 613
particle formation. PLoS Pathog5: e1000632. 614
67. Peña J, Harris E (2012) Early dengue virus protein synthesis induces extensive 615
rearrangement of the endoplasmic reticulum independent of the UPR and SREBP-2 616
pathway. PLoS One(6):e38202. doi: 10.1371/journal.pone.0038202. 617
68. Liu K, Czaja MJ (2013) Regulation of lipid stores and metabolism by lipophagy. Cell 618
Death and Differentiation20: 3–11. 619

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
69. Ait-Goughoulte M, Kanda T, Meyer K, Ryerse JS, Ray RB, Ray R (2008) Hepatitis C 620
virus genotype 1a growth and induction of autophagy. J Virol82: 2241–2249. 621
70. Sir D, Tian Y, Chen WL, Ann DK, Yen TS, Ou JH (2010) The early autophagic 622
pathway is activated by hepatitis B virus and required for viral DNA replication. Proc 623
Natl Acad Sci USA107:4383–4388. 624
71. McLauchlan J (2009) Hepatitis C virus: viral proteins on the move. Biochem Soc 625
Trans 37: 986–990. 626
72. Heaton NS, Randall G (2010) Dengue virus-induced autophagy regulates lipid 627
metabolism. Cell Host Microbe8: 422–432. 628
73. Heaton NS, Perera R, Berger KL, Khadka S, Lacount DJ, Kuhn RJ, Randall G (2010) 629
Dengue virus nonstructural protein 3 redistributes fatty acid synthase to sites of viral 630
replication and increases cellular fatty acid synthesis. Proc Natl Acad Sci 631
USA107:17345–17350 632
74. Zhang L, Wang A (2012) Virus-induced ER stress and the unfolded protein 633
response. Front Plant Sci3:293. doi: 10.3389/fpls.2012.00293. 634
75. Lee DY, Lee J, Sugden B. The unfolded protein response and autophagy: 635
herpesviruses rule! J Virol. 2009;83(3):1168–1172. 636
76. Schroder M. Endoplasmic reticulum stress responses. Cell Mol Life Sci. 637
2008;65(6):862–894. 638
77. Munz C. Enhancing immunity through autophagy. Annu Rev Immunol. 2009;27:423–639
449. 640
78. Umareddy I, Pluquet O, Wang QY, Vasudevan SG, Chevet E, Gu F (2007) Dengue 641
virus serotype infection specifies the activation of the unfolded protein response. 642
Virol J4:91. 643
79. Peña J, Harris E (2011) Dengue virus modulates the unfolded protein response in a 644
time-dependent manner. J Biol Chem286(16):14226-36. 645
80. Paradkar PN, Ooi EE, Hanson BJ, Gubler DJ, Vasudevan SG (2011) Unfolded protein 646
response (UPR) gene expression during antibody-dependent enhanced infection of 647
cultured monocytes correlates with dengue disease severity. Biosci Rep31(3):221-30. 648
81. Anderson P, Kedersha N (2009) RNA granules: post-transcriptional and epigenetic 649
modulators of gene expression. Nat Rev Mol Cell Biol10: 430–436 650
82. Eulalio A, Behm-Ansmant I, Izaurralde E (2007) P bodies: at the crossroads of post-651
transcriptional pathways. Nat Rev Mol Cell Biol8:9–22 652
83. Buchan JR, Muhlrad D, Parker R (2008) P bodies promote stress granule assembly in 653
Saccharomyces cerevisiae. J Cell Biol183: 441–455 654
84. Lloyd RE (2012) How Do Viruses Interact with Stress-Associated RNA Granules? 655
PLoS Pathog8(6): e1002741. doi:10.1371/journal.ppat.1002741 656
85. White JP, Lloyd RE (2012) Regulation of stress granules in virus systems. Trends 657
Microbiol20:175–183 658
86. Piotrowska J, Hansen SJ, Park N, Jamka K, Sarnow P, et al. (2010) Stable Formation 659
of compositionally unique stress granules in virus-infected cells. J Virol 84: 3654–660
3665 661
87. Isler JA, Skalet AH, Alwine JC (2005) Human Cytomegalovirus Infection Activates 662
and Regulates the Unfolded Protein Response. J Virol79:6890-6899. 663

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
88. Montero H et al. (2008) Rotavirus infection induces the phosphorylation of eIF2alpha 664
but prevents the formation of stress granules. J Virol82:1496–1504 665
89. Khaperskyy DA, Hatchette TF, McCormick C (2011) Influenza A virus inhibits 666
cytoplasmic stress granule formation. Faseb J DOI: 10.1096/fj.11-196915 667
90. Smith JA, Schmechel SC, Raghavan A et al. (2006) Reovirus induces and benefits 668
from an integrated cellular stress response. J Virol80: 2019–2033 669
91. Qin Q, Hastings C, Miller CL (2009) Mammalian orthoreovirus particles induce and 670
are recruited into stress granules at early times postinfection. J Virol 83:11090–11101. 671
92. Rathore AP, Ng ML, Vasudevan SG (2013) Differential unfolded protein response 672
during Chikungunya and Sindbis virus infection: CHIKV nsP4 suppresses eIF2α 673
phosphorylation. Virol J10:36 674
93. White JP, Cardenas AM, Marissen WE, Lloyd RE (2007) Inhibition of cytoplasmic 675
mRNA stress granule formation by a viral proteinase. Cell Host Microbe 2: 295–305. 676
94. Emara MM, Brinton MA (2007) Interaction of TIA-1/TIAR with West Nile and 677
dengue virus products in infected cells interferes with stress granule formation and 678
processing body assembly. Proc Natl Acad Sci USA104(21):9041–9046. 679
95. Ward A M, Bidet K, Yinglin A, Ler S G, Hogue K, Blackstock W, Gunaratne J,. 680
Garcia-Blanco M A (2011) Quantitative mass spectrometry of DENV-2 RNA-681
interacting proteins reveals that the DEAD-box RNA helicase DDX6 binds the DB1 682
and DB2 3' UTR structures. RNA Biology8:1173-1186. 683
96. Esclatine A et al. (2004) The UL41 protein of herpes simplex virus mediates selective 684
stabilization or degradation of cellular mRNAs. Proc Natl Acad Sci U.S.A. 101: 685
18165–18170 686
97. Legros S et al. (2011) The HTLV-1 Tax protein inhibits formation of stress granules 687
by interacting with histone deacetylase 6. Oncogene 30:4050–4062 688
98. Abrahamyan LG, Chatel-Chaix L, Ajamian L et al. (2010) Novel Staufen1 689
ribonucleoproteins prevent formation of stress granules but favour encapsidation of 690
HIV-1 genomic RNA. J Cell Sci123:369–383 691
99. Ariumi Y, Kuroki M, Kushima Y, Osugi K, Hijikata M, et al. (2011) Hepatitis C virus 692
hijacks P-body and stress granule components around lipid droplets. J Virol 85: 6882–693
6892. 694
100. Lindquist ME et al. (2011) Activation of protein kinase R is required for 695
induction of stress granules by respiratory syncytial virus but dispensable for viral 696
replication. Virology413: 103–110 697
101. Simpson-Holley M, Kedersha N, Dower K, Rubins KH, Anderson P, et al. 698
(2011) Formation of antiviral cytoplasmic granules during orthopoxvirus infection. J 699
Virol 85: 1581–1593 700
701
702
703

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
• Role of intracellular events is very important in dengue virus (DENV) infection
• Endoplasmic reticulum (ER) is the key point to provide double membrane structures
• DENV exploits autophagy to increase its replication
• Assembly of Stress granules is decreased by DENV to increase its survival in host cells
• Unfolded protein response (UPR) also plays a critical role





























