Embed Size (px)
Citation preview

Proc. Nat. Acad. Sc. USAVol. 70, No. 5, pp. 1359-1363, May 1973
Independent Control of Ribosomal Gene Replication in PolyteneChromosomes of Drosophila melanogaster
(RNA-DNA hybridization/euchromatin/heterochromatin)
BRIAN B. SPEAR AND JOSEPH G. GALL
Department of Biology, Yale University, New Haven, Connecticut 06520
Contributed by Joseph G. Gall, February £6, 1973
ABSTRACT The saturation values for ribosomal RNAhybridization have been determined for DNA from diploidand polytene tissues of Drosophila melanogaster. Thesevalues have been measured in XO and XX larvae whichhave, respectively, one and two nucleolus organizers inthe diploid chromosome set. The results show that (1)in diploid cells the ribosomal (r)DNA is present in amountsproportional to the number of nucleolus organizers, (2)in polytene cells the rDNA is under-replicated with respectto the euchromatic DNA, and (3) in polytene cells theamount of rDNA is independent of the diploid number ofnucleolus organizers. These observations suggest thatsomatic variations in rDNA content may involve indepen-dent polytenization of the nucleolus organizer withoutchange in the number of ribosomal cistrons per organizer.The independent polytenization ofrl)NA is proposed as thechromosomal explanation for the relative increase inrDNA in flies of the XO constitution.
The chromosomes of Drosophila melanogaster, like those ofmost eukaryotes, contain two distinct types of chromatin, theeuchromatin and the heterochromatin. The euchromatin isdiffuse during interphase, is lightly staining in mitotic pro-phase, and contains almost all of the known genes (1). Theheterochromatin, contrastingly, remains condensed duringinterphase, is consequently darkly staining in mitotic pro-phase, and contains very few genes. In addition to thesegenetic and cytological distinctions, the euchromatin consistsmainly of unique DNA sequences, while most of the hetero-chromatin in diploid cells is highly repeated satellite DNA(2-4).During the formation of the giant polytene chromosomes of
the Drosophila salivary gland, the differences between theeuchromatin and heterochromatin become even more pro-nounced. The euchromatic arms proceed through a series ofDNA replications resulting in as many as 1024 DNA fibers(5). The heterochromatic regions of the chromosomes, on theother hand, clump together forming the chromocenter andreplicate little or not at all (2, 6). The resulting polytenenucleus has a large increase in unique DNA, but little or noincrease in the heterochromatic satellite DNA (2, 7).
In D. melanogaster the DNA that codes for ribosomalRNA is located at or near the nucleolus organizer, a regionwithin the heterochromatin of the X and Y chromosomes (8).The ribosomal cistrons are repeated about 250 times in thenucleolus organizer of wild-type stocks (9). The properties ofthe ribosomal (r)DNA are quite different from those of the
heterochromatic DNA that surrounds it. For instance, therDNA is genetically active, transcribing rRNA and forminga prominent nucleolus in both diploid and polytene tissues.Unlike the satellite DNA, the rDNA replicates during theformation of polytene tissue. This has been demonstrated bythymidine incorporation in D. melanogaster (10, 28), and byrRNA hybridization to DNA from salivary glands of D. hydei(11, 12), Sciara (13), and Rhynchosciara (14).Because the rDNA in polytene nuclei is near the non-
replicating heterochromatin, we felt that it might show un-usual replicative properties in these cells. We have examinedthe rDNA content of diploid and polytene cells of XO andXX larvae. In diploid DNA we found that, as expected, theamount of rDNA was in proportion to the number of nucleolusorganizers. However, the rDNA content of polytene cells isnot only lower than expected, as previously reported byHennig and Meer (12), but it is also independent of thenumber of nucleolus organizers in the zygote. We have con-cluded that the rDNA in polytene chromosomes is underdifferent replicative control than either the euchromaticDNA or the heterochromatic DNA. In addition, this controlis at the individual cell level. Some of these results have beenreported in abstract form (15).
METHODS AND MATERUILS
Drosophila Stocks. Since different wild-type strains ofD. melanogaster may have different amounts of rDNA (9),two strains were used that were expected to have differentrDNA content. These were the Oregon R and Urbana Swild-type strains. Males with a wild-type X as the only sexchromosome (XO) were produced by mating females ofeither wild-type stock to males with theX andY chromosomesattached [Y8X.YLIn(1)EN, Ys B f v y.yL y+]. The XOmales were morphologically normal, though sterile, and hadonly one nucleolus organizer. Every bottle of flies producingXO males was tested by brain squash karyotyping of randomlarvae and by fertility tests.
Tissue Isolatwon and DNA Extraction. The larval brains andimaginal discs were chosen as diploid tissues. Although thereare some polytene and low-level polyploid nuclei observable ina brain squash preparation, these cells are a small fraction ofthe total tissue. The polytene tissue was the larval salivarygland. Late third-instar larvae were dissected in Robb's (16)Drosophila Ringer solution, and brains and imaginal discs orsalivary glands were removed. Some of the tissue was incubatedin the same solution containing [14C]thymidine (10 ,ACi/ml,
1359
Abbreviations: SSC, 0.15 M NaCl-0.015 M Na citrate, pH 7.0;rDNA, DNA coding for ribosomal RNA.
Proc. Nat. Acad. Sci. USA 70 (1973)
007, 0.54z0
C-*t- 0.4z5r
*- 0.3z0
Na 0.2
m
I
_ 0.1zwu
wa.
1 2 1 2XO XX Xo XX
NUMBER OF NUCLEOLUS ORGANIZERS
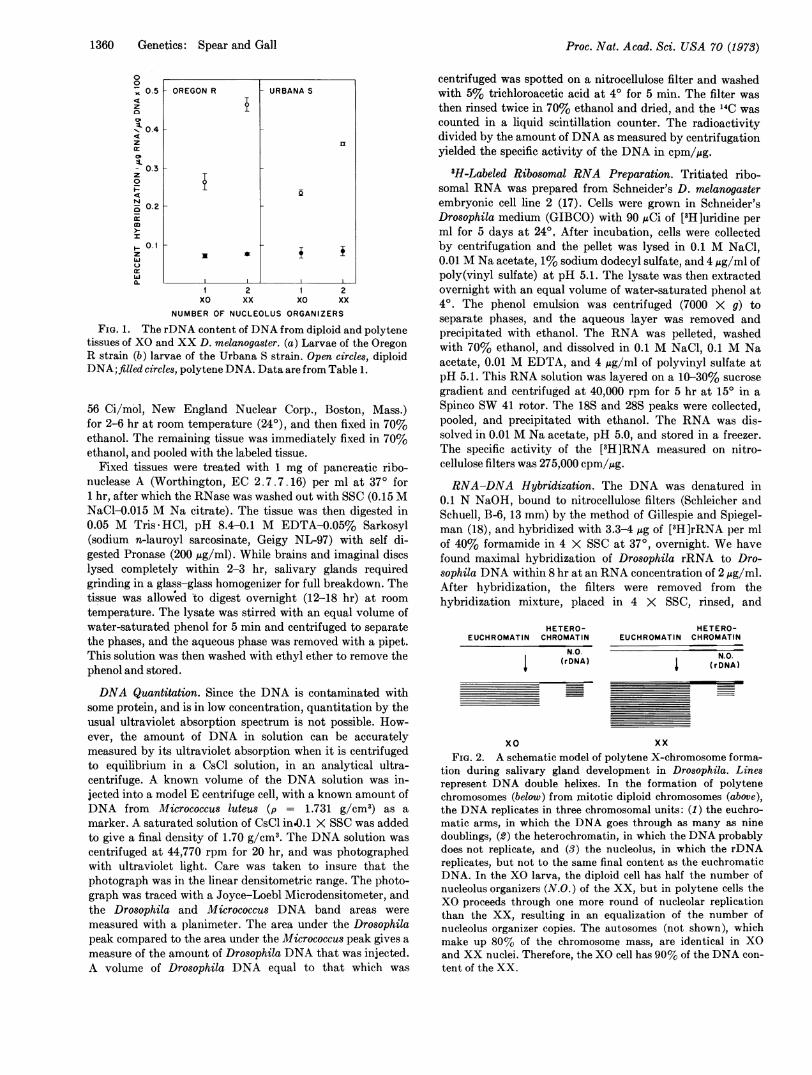
FIG. 1. The rDNA content of DNA from diploid and polytenetissues of XO and XX D. melanogaster. (a) Larvae of the OregonR strain (b) larvae of the Urbana S strain. Open circles, diploidDNA;filled circles, polytene DNA. Data are from Table 1.
56 Ci/mol, New England Nuclear Corp., Boston, Mass.)for 2-6 hr at room temperature (240), and then fixed in 70%ethanol. The remaining tissue was immediately fixed in 70%ethanol, and pooled with the labeled tissue.
Fixed tissues were treated with 1 mg of pancreatic ribo-nuclease A (Worthington, EC 2.7.7.16) per ml at 370 for1 hr, after which the RNase was washed out with SSC (0.15 MNaCl-0.015 M Na citrate). The tissue was then digested in0.05 M Tris * HCl, pH 8.4-0.1 M EDTA-0.05% Sarkosyl(sodium n-lauroyl sarcosinate, Geigy NL-97) with self di-gested Pronase (200 Mug/ml). While brains and imaginal discslysed completely within 2-3 hr, salivary glands requiredgrinding in a glass-glass homogenizer for full breakdown. Thetissue was allowed to digest overnight (12-18 hr) at room
temperature. The lysate was stirred with an equal volume ofwater-saturated phenol for 5 min and centrifuged to separatethe phases, and the aqueous phase was removed with a pipet.This solution was then washed with ethyl ether to remove thephenol and stored.
DNA Quantitation. Since the DNA is contaminated withsome protein, and is in low concentration, quantitation by theusual ultraviolet absorption spectrum is not possible. How-ever, the amount of DNA in solution can be accuratelymeasured by its ultraviolet absorption when it is centrifugedto equilibrium in a CsCl solution, in an analytical ultra-centrifuge. A known volume of the DNA solution was in-jected into a model E centrifuge cell, with a known amount ofDNA from Micrococcus luteus (p = 1.731 g/cm3) as a
marker. A saturated solution of CsCl ine0.1 X SSC was addedto give a final density of 1.70 g/cm3. The DNA solution was
centrifuged at 44,770 rpm for 20 hr, and was photographedwith ultraviolet light. Care was taken to insure that thephotograph was in the linear densitometric range. The photo-graph was traced with a Joyce-Loebl Microdensitometer, andthe Drosophila and Micrococcus DNA band areas were
measured with a planimeter. The area under the Drosophilapeak compared to the area under the Micrococcus peak gives a
measure of the amount of Drosophila DNA that was injected.A volume of Drosophila DNA equal to that which was
centrifuged was spotted on a nitrocellulose filter and washedwith 5% trichloroacetic acid at 40 for 5 min. The filter wasthen rinsed twice in 70% ethanol and dried, and the '4C wascounted in a liquid scintillation counter. The radioactivitydivided by the amount of DNA as measured by centrifugationyielded the specific activity of the DNA in cpm/,gg.
3H-Labeled Ribosomal RNA Preparation. Tritiated ribo-somal RNA was prepared from Schneider's D. melanogasterembryonic cell line 2 (17). Cells were grown in Schneider'sDrosophila medium (GIBCO) with 90 MCi of [3H]uridine perml for 5 days at 240, After incubation, cells were collectedby centrifugation and the pellet was lysed in 0.1 M NaCl,0.01 M Na acetate, 1% sodium dodecyl sulfate, and 4 ,ug/ml ofpoly(vinyl sulfate) at pH 5.1. The lysate was then extractedovernight with an equal volume of water-saturated phenol at4°. The phenol emulsion was centrifuged (7000 X g) toseparate phases, and the aqueous layer was removed andprecipitated with ethanol. The RNA was pelleted, washedwith 70% ethanol, and dissolved in 0.1 M NaCl, 0.1 M Naacetate, 0.01 M EDTA, and 4 Mg/ml of polyvinyl sulfate atpH 5.1. This RNA solution was layered on a 10-30% sucrosegradient and centrifuged at 40,000 rpm for 5 hr at 150 in aSpinco SW 41 rotor. The 18S and 28S peaks were collected,pooled, and precipitated with ethanol. The RNA was dis-solved in 0.01 M Na acetate, pH 5.0, and stored in a freezer.The specific activity of the [3H]RNA measured on nitro-cellulose filters was 275,000 cpmlg.RNA-DNA Hybridization. The DNA was denatured in
0.1 N NaOH, bound to nitrocellulose filters (Schleicher andSchuell, B-6, 13 mm) by the method of Gillespie and Spiegel-man (18), and hybridized with 3.3-4 ,lg of [3H]rRNA per mlof 40% formamide in 4 X SSC at 370, overnight. We havefound maximal hybridization of Drosophila rRNA to Dro-sophila DNA within 8 hr at an RNA concentration of 2 Mg/ml.After hybridization, the filters were removed from thehybridization mixture, placed in 4 X SSC, rinsed, and
HETERO-EUCHROMATIN CHROMATIN
I
HETERO-EUCHROMATIN CHROMATIN
N.O.(rDNA)
N.O.C(rDNA)
XO xx
FIG. 2. A schematic model of polytene X-chromosome forma-tion during salivary gland development in Drosophila. Linesrepresent DNA double helixes. In the formation of polytenechromosomes (below) from mitotic diploid chromosomes (above),the DNA replicates in three chromosomal units: (1) the euchro-matic arms, in which the DNA goes through as many as ninedoublings, (2) the heterochromatin, in which the DNA probablydoes not replicate, and (3) the nucleolus, in which the rDNAreplicates, but not to the same final content as the euchromaticDNA. In the XO larva, the diploid cell has half the number ofnucleolus organizers (N.O.) of the XX, but in polytene cells theXO proceeds through one more round of nucleolar replicationthan the XX, resulting in an equalization of the number ofnucleolus organizer copies. The autosomes (not shown), whichmake up 80% of the chromosome mass, are identical in XOand XX nuclei. Therefore, the XO cell has 90% of the DNA con-
tent of the XX.
- OREGON R - URBANA S
II
i *
xIt + +
I Il
1360 Genetics: Spear and Gall
F

rDNA in Polytene Chromosomes 1361
treated with pancreatic RNase (20 /ug/ml in 2 X SSC) for1 hr. They were then washed with 2 X SSC, then 70% ethanol,dried, and counted. The use of radioactive label in bothnucleic acids allowed for an accurate measurement of theamount of RNA bound per ,ug of DNA.
14C-Labeled Drosophila DNA extracted from cultured cellswas included with every set of filters as a control. This DNAhybridized to a level of 0.30% with rRNA. As a check for non-specific binding of RNA, hybridization was also performedwith blank filters, with Micrococcus DNA, and with theDrosophila DNA solution that had been digested with100 ,ug of DNase per ml at 25° for 1 hr. In no case was therebinding of RNA to the filters.
RESULTS
Since the XO diploid cells have one nucleolus organizer, whileXX diploid cells have two, the percent of rDNA in XO andXX diploid DNA should also be in a ratio of one to two. Theresults (Fig. 1 and Table 1) approach this predicted ratio.In the Oregon R strain the rDNA content is 0.26% for XOand 0.47% for XX, and in the Urbana S strain the amountsare 0.24% for XO and 0.37% for XX.To test for independent replication of rDNA, we have also
measured the rRNA hybridization levels of DNA frompolytene tissues. This has been done for both XO and XXlarvae in the two D. melanogaster strains. Hybridization ofrRNA to these DNAs allows two separate tests: a comparisonof polytene rDNA replication with polytene euchromatic DNAreplication, and a comparison of polytene rDNA replication incells starting with one nucleolus organizer (XO) with polytenerDNA replication in cells starting with two nucleolus orga-nizers (XX). The results in Table 1 and Fig. 1 indicate thatthe polytene rDNA is under-replicated with respect to theeuchromatic DNA, and that the final level of replication isindependent of the number of sex chromosomes.
In wild-type female larvae, the amount of rDNA in totalDNA for diploid cells is about 0.47% for Oregon R and 0.37%for Urbana S. The rDNA content of DNA from the polytenesalivary glands is reduced considerably, to about 0.08% forboth strains. A reduced level of rDNA in the salivary glandcompared to other tissues has been demonstrated previouslyin Drosophila hydei (12) and in Rhynchosciara (14). There isalso a reduction of rDNA in D. melanogaster larvae as com-pared to adults (19).
In polytene chromosome formation, the degree of replica-tion of rDNA is intermediate between those of the euchro-matic DNA and the heterochromatic DNA. In a fully de-veloped polytene nucleus with 1024 strands in the euchromaticarms (5), the polytene rDNA content of 0.08% constitutes150-200 times the amount of rDNA in a diploid nucleolusorganizer. This is equivalent to six or seven doublings of therDNA. It is likely, though, that the level of under-replicationof rDNA will vary with the DNA content of the individualnuclei. Cells with greater total DNA content will probablyhave proportionately lower content of rDNA.The rDNA in polytene cells is not only under-replicated but
its concentration is also disproportional to the diploid numberof nucleolus organizers. The XO polytene cells, arising fromcells with only one nucleolus organizer, have increased theirrDNA replication to have as much rDNA as the XX polytenecells. In both the Oregon R and Urbana S strains, the rDNA
TABLE 1. The hybridization of rRNA to Drosophiladiploid and polytene DNA
No. ofdeter- Ratio ofmina- diploid to
DNA % rDNA i SEM tions polytene
Oregon R XX diploid 0.469 i 0.016 14 6.0Oregon R XX polytene 0.078 + 0.002 5Oregon R XO diploid 0.264 :1= 0.021 12 3.7Oregon R XO polytene 0.074 i 0.007 6Urbana S XX diploid 0.365 i 0.005 12 4.3Urbana S XX polytene 0.085 ± 0.011 4Urbana S XO diploid 0.236 4 0.006 11 2.9Urbana S XO polytene 0.080 4± 0.013 4
1-2 ug of [14C]DNA on a nitrocellulose filter was incubatedwith [3H]rRNA (3.3-4.0,g/ml) in 40%0 formamide in 4 XSSC at 370, overnight. XO values have been multiplied by 0.9 toaccount for a 10%o reduction in the total nuclear DNA content(see legend to Fig. 2.). Radioactivity on the filters generallyranged from 200-1200 cpm for 14C and from 400-1500 cpm for3H.
The XO polytene DNA content is exactly twice the amountexpected from the original number of nucleolus organizers.The DNA from diploid tissues, on the other hand, has per-centages directly proportional to the number of nucleolusorganizer regions: the XO DNA has half as much rDNA asXX DNA. The relationship of XO to XX rDNA hybridizationdata for both diploid and polytene DNA is shown in Fig. 1.Because of the disproportional rDNA synthesis, the degree ofrDNA under-replication in polytene cells is less extreme inXO cells than in XX.For both stocks, the XO diploid rDNA content was slightly
above 0.5 times that of the XX. This may be due to a smallnumber of polytene and polyploid cells in the brain. It is alsointeresting to note that the amounts of rDNA in Oregon Rand Urbana S polytene cells are the same, despite the differ-ences in their diploid values.
DISCUSSION
These results demonstrate three features of ribosomal DNAin D. melanogaster: (1) in diploid tissue the amount of rDNAis in proportion to the number of nucleolus organizers, (2) inpolytene cells rDNA replicates, but not to the same extent asthe euchromatic DNA, and (3) in polytene cells the extent ofrDNA replication is not dependent on the starting number ofnucleolus organizers. These facts bear upon our under-standing of both polytene chromosome organization and theanalysis of rDNA in Drosophila.The DNA in polytene chromosomes is divided into three
classes according to the extent of replication, and each class isassociated with a different type of chromosome structure.First, the euchromatic DNA, which forms the chromosomearms, replicates in a geometric series. This replication maycontinue for up to nine doublings of the DNA, resultingin 1024 DNA strands (5). The second type of DNA is thatfound in the heterochromatin. It consists largely of repetitivesatellite DNA (2-4, 7). This DNA replicates very little or
possibly not at all (2, 6). The third type of DNA is thecontent of XO and XX salivary gland DNA is about 0.08%. rDNA, located in the nucleolus. This DNA does replicate,
Proc. Nat. Acad. Sci. USA 70 (1973)

Proc. Nat. Acad. Sci. USA 70 (1973)
but not to the same extent as the euchromatic DNA. Thesethree types of DNA are under separate replicative control,and are apparently independent of each other to a certainextent.Of particular interest is the independence of polytene rDNA
synthesis, not only from the other DNA fractions in the nu-cleus, but also from the genotype of the fly. In the experi-ments reported here, the XO polytene cells, which havearisen from cells with only one nucleolus organizer, haveundergone additional rDNA replication until, in the late third-larval instar, they have as many ribosomal genes as the XXcells. A comparable case occurs in oocytes of mutant Xenopusthat have only one nucleolus organizer instead of the usualtwo. Despite the lack of one of the nucleolus organizers, theseoocytes amplify rDNA to the same level as the wild type(20, 21).Although the rDNA in XO individuals compensates during
the polytenization process, it should be noted that theX-chromosome as a whole does not (22). Thus, in the malethere is only one X chromosome, and when this polytenizes itpossesses only half the number of strands present in each ofthe synapsed autosome pairs. In the female the two X chromo-somes are paired in the polytene nuclei and together theyhave a number of strands equal to that of an autosomepair.The mechanism of rDNA regulation in XO polytene cells
is not clear at this time. We suggest that the disproportionalrDNA synthesis is best viewed as a differential polytenizationof the entire nucleolus organizer region, as shown in Fig. 2.In such a scheme rDNA increase occurs by successive dou-blings of a nucleolus organizer with a fixed number of cistrons.It is not necessary to postulate variations in the number oftandemly repeated ribosomal cistrons within each organizer.Whether one regards such rDNA replication in polytenenuclei as extrachromosomal or not would seem a matter ofdefinition, depending upon how one regards the tenuous con-nections between the rDNA and the rest of the chromosome.The demonstration of independent rDNA replication in
polytene cells influences the interpretation of earlier data onthe amount of rDNA in whole flies. This is especially true forthe increase in rDNA multiplicity observed by Tartof (23, 24).He found that whole adult XO flies had 0.8 times the rDNApercentage of XX flies. This was interpreted as an increasein the number of ribosomal genes per nucleolus organizer.Our data indicate, however, that diploid tissues do not showthis differential gene multiplicity, but have more nearly theexpected 1: 2 ratio of rDNA between XO and XX individuals.We suggest that the relative rDNA increase in XO flies doesnot result from a change in the number of genes per nucleolusorganizer, but rather from an increase in the number ofnucleolus organizer copies in polytene or polyploid cells.There is evidence for the presence of polytene chromosomesin Drosophila adults (25). The rDNA increase in XO fliesobserved by Tartof could result from independent poly-tenization of rDNA in those adult tissues that have polytenecells.Although this explanation for rDNA variability is not
unequivocal, our results from diploid DNA limit the possiblealternatives. Since in diploid tissues the XO:XX ratio ofrDNA content is 1: 2, directly proportional to the number ofnucleolus organizers, the XO rDNA increase is not a feature
genes is therefore not fixed in the gametes or in early embryo-genesis, but must occur in differentiated cells later in de-velopment. The simplest explanation is that the cells thatcompensate are those whose chromosome structure allowsindependent polyteny of the rDNA.At this point it is important to make a distinction between
the purely somatic phenomenon reported by Tartof and theinherited rDNA increase described by Ritossa under thename "magnification" (26, 27). Magnification involves thebobbed mutations, which are partial deficiencies for the rDNA.Under certain well-defined conditions bobbed stocks revert to
the wild type over a period of 3-4 generations, and thephenotypic reversion is accompanied by a return to wild-typerDNA content. In this case there does seem to be a real changein the number of ribosomal cistrons in the germ-line chromo-somes. Independent polytenization of rDNA that occurs insomatic cells may be used as a reasonable explanation fornoninherited rDNA increase. However, since polytenizationdoes not occur in the germ line, independent polytenization ofrDNA cannot explain magnification. In the whole fly, with itsvariety of cell types, both somatic rDNA variation andmagnification may be present simultaneously. Nucleic acidhybridization on DNA from whole flies measures only thetotal amount of rDNA present. However, measurements on
selected tissues with defined chromosome types, as we havepresented here, provide a means for differentiating thoserDNA variations due to somatic regulation and those due tomagnification.
This research was supported by Grant VC 85 from the Amer-ican Cancer Society and USPHS Grant GM 12427 from theNational Institute of General Medical Sciences. B.B.S. was
supported by USPHS Training Grant 5 TO1 HD 00032.
1. Lindsley, D. L. & Grell, E. H. (1967) "Genetic variations ofDrosophila melanogaster," Carnegie Inst. Wash. Publ. No.627.
2. Gall, J. G., Cohen, E. H. & Polan, M. L. (1971) "RepetitiveDNA sequences in Drosophila," Chromosoma 33, 319-344.
3. Rae, P. M. M. (1970) "Chromosomal distribution of rapidlyreannealing DNA in Drosophila melanogaster," Proc. Nat.Acad. Sci. USA 67, 1018-1025.
4. Botchan, M., Kram, R., Schmid, C. W. & Hearst, J. E.(1971) "Isolation and chromosomal localization of highlyrepeated DNA sequences in Drosophila melanogaster,"Proc. Nat. Acad. Sci. USA 68, 1125-1129.
5. Swift, H. & Rasch, E. (1954) "Composition and structureof giant chromosomes," ed. Alfert, M., in Int. Rev. Cytol. 3,131-175.
6. Rudkin, G. (1969) "Non-replicating DNA in Drosophila,"Genetics Suppl. 61, 227-238.
7. Dickson, E., Boyd, J. B. & Laird, C. D. (1971) "Sequencediversity of polytene chromosome DNA from Drosophilahydei," J. Mol. Biol. 61, 615-627.
8. Ritossa, F. & Spiegelman, S. (1965) "Localization of DNAcomplementary to ribosomal RNA in the nucleolus organizerregion of Drosophila melanogaster," Proc. Nat. Acad. Sci.USA 53, 737-745.
9. Ritossa, F. M., Atwood, K. C. & Spiegelman, S. (1966)"A molecular explanation of the bobbed mutants of Dro-sophila as partial deficiencies of 'Ribosomal' DNA," Genetics54, 819-834.
10. Rodman, T. C. (1969) "Morphology and replication ofintranucleolar DNA in polytene nuclei," J. Cell Biol. 42,575-582.
11. Pardue, M. L., Gerbi, S. A., Eckhardt, R. A. & Gall, J. G.(1970) "Cytological localization of DNA complementary toribosomal RNA in polytene chromosomes of Diptera,"
common to all cells. The change in multiplicity of rRNA
1362 Genetics: Spear and Gall
Chromosoma 29, 268-290.

rDNA in Polytene Chromosomes 1363
12. Hennig, W. & Meer, B. (1971) "Reduced polyteny ofribosomal RNA cistrons in giant chromosomes of Drosophilahydei," Nature New Biol. 233, 70-72.
13. Gerbi, S. A. (1971) "Localization and characterization of theribosomal cistrons in Sciara coprophila," J. Mol. Biol. 58,499-511.
14. Gambarini, A. G. & Meneghini, R. (1972) "RibosomalRNA genes in salivary gland and ovary of Rhynchosciaraangelae," J. Cell Biol. 54, 421-425.
15. Spear, B. B. (1972) "Tissue specific variations in themultiplicity of genes for ribosomal RNA in Drosophilamelanogaster," J. Cell. Biol. 55, 246 abstr.
16. Robb, J. A. (1969) "Maintenance of imaginal discs ofDrosophila melanogaster in chemically defined media,"J. Cell Biol. 41, 876-885.
17. Schneider, I. (1972) "Cell lines derived from late embryonicstages of Drosophila melanogaster," J. Embryol. Exp.Morphol. 27, 353-365.
18. Gillespie, D. & Spiegelman, S. (1965) "A quantitative assayfor DNA-RNA hybrids with DNA immobilized on a mem-brane," J. Mol. Biol. 12, 829-842.
19. Sibatani, A. (1971) "Difference in the proportion of DNAspecific to ribosomal RNA between adults and larvae ofDrosophila melanogaster," Mol. Gen. Genet. 114, 177-180.
20. Brown, D. D. & Dawid, I. (1968) "Specific gene amplifica-tion in oocytes," Science 160, 272-280.
21. Perkowska, E., Macgregor, H. C. & Birnstiel, M. L. (1968)"Gene amplification in the oocyte nucleus of mutant andwild type Xenopus laevis," Nature 217, 649-650.
22. Aronson, J. F., Rudkin, G. T. & Schultz, J. (1954) "Acomparison of giant X chromosomes in male and femaleDrosophila melanogaster by cytophotometry in the ultra-violet," J. Histochem. Cytochem. 2, 458-459.
23. Tartof, K. D. (1971) "Increasing the multiplicity of riboso-mal RNA genes in Drosophila melanogaster," Science 171,294-297.
24. Tartof, K. D. (1973) "Regulation of ribosomal RNA genemultiplicity in Drosophila melanogaster," Genetics 73,57-71.
25. Ashburner, M. (1970) "Function and structure of polytenechromosomes during insect development," Advan. InsectPhysiol. 7, 1-196.
26. Ritossa, F. M. (1968) "Unstable redundancy of genes forribosomal RNA," Proc. Nat. Acad. Sci. USA 60, 509-516.
27. Ritossa, F. M. & Scala, G. (1969) "Equilibrium variationsin the redundancy of rDNA in Drosophila melanogaster,"Genetics, Suppl. 61, 305-317.
28. Nash, D. & Plaut, W. (1965) "On the presence of DNA inlarval salivary gland nucleoli in Drosophila melanogaster,"J. CeU Biol. 27, 682-686.
Proc. Nat. Acad. Sci. USA 70 (1973)





























