Embed Size (px)
Citation preview

RGD-Recognizing Integrins Mediate Interactionsof Human Prostate Carcinoma Cells With
Endothelial Cells In Vitro
Victor I. Romanov* and Michael S. Goligorsky
Department of Medicine, State University of New York, Stony Brook, New York
BACKGROUND. Interactions of cancer cells with endothelium are a crucial step in metastaticinvasion. RGD-recognizing integrins play a definitive role in these interactions.METHODS. Fluorescence-activated cell sorting (FACS) analysis of RGD-sensitive integrins inprostate epithelial cells was performed. Attachment inhibition assay was used to characterizefunctionality of particular integrins. Potential partners for RGD-binding integrins in humanumbilical vein endothelial cells (HUVEC) were identified by Western blotting and attachmentinhibition assay. To determine the RGD-flanking amino acids optimal for interactions withprostate cell integrins, these cells were biopanned with a phage library.RESULTS. Different expressions of RGD-recognizing integrins and distinctions in RGD-dependent adhesion of nonmalignant and cancer cells were observed. Cancer but not controlcells were detached from culture plastic by incubation with RGD peptide. Adhesion of car-cinoma cells to HUVEC was RGD-sensitive, in contrast to nonmalignant cells. Antibodiesagainst a3, a5, b1, and avb3 inhibited interactions of carcinoma cells with HUVEC. Potentialligands for a5b1, a3b1, and aVb3 integrins, fibronectin, and vitronectin, were detected on theHUVEC surface. Several phages which preferentially bound to the surface of particular pros-tate cells were selected.CONCLUSIONS. Interactions of prostate carcinoma with endothelium are mediated in partvia a5b1, a3b1, and avb3 integrins. Because these interactions are RGD-sensitive, syntheticRGD peptides with optimized flanking amino acids can potentially be used as antimetastaticagents. Prostate 39:108–118, 1999. © 1999 Wiley-Liss, Inc.
KEY WORDS: prostate cancer; integrins; peptide phage display
INTRODUCTION
Every third cancer diagnosed in American males isof prostatic origin [1]. Many histologically detectedasymptomatic prostate cancers never become clini-cally manifest and life-threatening. A small percentageof these tumors traverses all the steps of tumorigenesisrequired to complete malignant progression and pro-duce clinically aggressive metastatic tumors. This typeof prostate cancer becomes a fatal disease for whichcurative treatment is presently limited [1,2]. It is,therefore, important to study the mechanisms of tu-morigenicity and metastasis formation to predict theclinical course of the individual case of prostate ab-normality and to discover ways to prevent cancer pro-gression.
One of the crucial steps in metastasis formation is
an adhesive interaction of tumor cells with other cellsor extracellular matrix (ECM) proteins. Vascular en-dothelium represents the first barrier between tumorcells and extravascular tissues. Such interactions occuras tumor cells migrate locally from primary lesionsand when circulating cells adhere to vascular endo-thelial cells or to basement membrane during the seed-ing of metastasis [3]. The integrin family of cell adhe-sion molecules mediates many of these interactions.
The integrin family is composed of 15 a and 8 b
Grant sponsor: NIH; Grant number: DK41573; Grant sponsor: CaP-CURE 1997 Year Research Award.*Correspondence to: Victor I. Romanov, Ph.D., Division of Nephrol-ogy and Hypertension, Department of Medicine, State University ofNew York, HSC, T15, Rm 20, Stony Brook, NY 11794-8152.Received 4 August 1998; Accepted 22 October 1998
The Prostate 39:108–118 (1999)
© 1999 Wiley-Liss, Inc.

subunits that may form over twenty different ab di-meric combinations on the cell surface. Most of theintegrins can bind two or more components of extra-cellular matrix. Several integrins bind to the canonicaltripeptide sequence (RGD), whereas others recognizedifferent sequences [4,5]. Peptides containing the RGDsequence can specifically inhibit binding of cells, ex-pressing RGD-recognizing integrins, to other cells orECM proteins [4,5]. Differential expression of exracel-lular matrix proteins and integrins in prostate tissueand prostate-derived cell lines was previously inves-tigated [6–12]. The composition of basal lamina sur-rounding normal glands contained laminin, type IVcollagen, entacin, and varying amounts of tenanscinand of collagen VII. The composition of basal laminaof neoplastic acini was similar, except for the loss typeVII collagen [8]. Production of several integrin sub-units has been shown to be decreased in neoplasticprostate tissue and in carcinoma cell lines. Completeloss of b4 expression in prostate carcinoma has beenreported [8]. Blocking of a4-subunit expression in tu-morigenic cells was shown in quantitative studywhere expression of integrin subunits was comparedin nontumorigenic and tumorigenic cell lines [11]. Thepossible role of a6-containing integrins in humanprostate cancer progression was studied [9,10]. How-ever, the special role of RGD-sensitive integrins inprostate cancer progression has not been specificallydiscussed.
The purpose of this study was to investigate therole of individual RGD-recognizing integrins in theinteractions of prostate carcinoma cells with endothe-lium and to define RGD-flanking amino acids optimalfor such interactions.
MATERIALS AND METHODS
Cell Lines and Antibodies
Primary prostate epithelial cells were either pur-chased from Clonetics (San Diego, CA) or derivedfrom histologically normal prostate obtained from pa-tients undergoing cystoprostatectomy. Prostate speci-mens were minced and incubated overnight at 37°Cwith 200 U/ml of collagenase in RPMI 1640. Cellswere washed once in RPMI 1640 and centrifuged at800g for 5 min. The pellet was resuspended in 0.1%(w/v) trypsin and incubated for a further 30 min at37°C on a shaker. The final digest was washed threetimes and centrifuged at 360g for 1 min. A supernatantenriched for stromal cells and a pellet enriched forepithelia were obtained, according to the previouslypublished protocol [13]. Epithelial-enriched pellet wasresuspended in prostate epithelial growth mediaPrEGM (Clonetics). Cells were characterized by im-munostaining for cytokeratins to determine their epi-
thelial origin and were used between 3–8 passages.PZ-HPV-7 cells (ATCC 2231 CRL) were originally
isolated from the peripheral zone of normal humanprostate and were immortalized with human papil-loma virus [14]. These cells were grown in Keratino-cyte Serum-Free Medium (Gibco BRL, Rockville, MD).BPH-1-prostate epithelial cells (derived from a pros-tate with benign prostatic hyperplasia) immortalizedwith SV-40 (a kind gift of Dr. Simon Hayward, Uni-versity of California at San Francisco) were grown inRPMI 1640 medium supplemented with 5% fetal bo-vine serum (FBS) and 100 U/ml penicillin-100 mg/mlstreptomycin (Gibco BRL). DU145 (ATCC HTB 81)was originally isolated from lesions in the brain of apatient with metastatic carcinoma of the prostate. Thecells were maintained in DMEM media supplementedwith 10% FBS and antibiotics. PC3 (CRL-1435 humanmetastasis to bone) was routinely maintained in RPMI1640 medium supplemented with 10% FBS and anti-biotics. HUVEC human endothelial cells, a primaryculture obtained from Clonetics, were maintained inendothelial cell basal medium (EBM-2) medium upto the tenth passage. Antibodies to integrin subunitsand ECM molecules used in this study are listed inTable I. All other chemicals were of the highest puritycommercially available.
Phage Peptide Library
The bacteriophage vector used for library construc-tion was Fuse5 (a generous gift from Dr. G. Smith,University of Missouri); K91 and DH10B (electrocom-petent) Escherichia coli strains were obtained fromGibco BRL. Peptides were expressed as N-terminal fu-sion to the mature protein III of bacteriophage M13. Inthe displayed peptide, the conformation of theCxxxRGDxxxC motif is restricted within a hairpinloop formed by the disulfide bridge between the twocysteine residues. Two synthetic oligonucleotides(58tcactcggccgacggggcttgtnnknnknnkaggggggatnnk-nnknnktgtgggccgctggcccga 38 and 58tcgggcctgcggc38,where k = g + t and n = g + a + t + c) were annealedand filled with T7 DNA polymerase (New EnglandBiolabs, Beverly, MA). The resulting DNA fragmentwas digested with BglII (200 ng, 8 pmol) and ligatedwith gene III of Fuse5 overnight at 15°C with 4,000units of T4 DNA ligase (New England Biolabs). Theligation buffer was substituted by water, and the finaldigest was prepared in 40 ml of water. The ligatedDNA (40 ml) was electroporated into 80 ml of DH10Belectrocompetent cells using a Gene Pulser (BioRad,Medford, MA) at 2.0 kV/cm, 200 ohms, 25 mF. Eachtransfection (total, 16) was resuspended in 2 ml SOCmedium containing 0.2 mg/ml tetracycline and al-lowed to recover and express the tetracycline-resistance gene, at 37°C for 1 hr. Aliquots (20 ml) from
Integrins in Carcinoma-Endothelium Interactions 109

each transfection were plated on LB plates containing20 mg/ml of tetracycline to determine the number oftransformations. The diversity of the library wasevaluated by sequencing phage DNA from 50 ran-domly picked colonies according to the manufactur-er’s protocol for Ph.D Phage Display Peptide library(New England Biolabs). Combined transfections werepropagated in 1 l of LB containing 20 mg/ml tetracy-cline and allowed to grow overnight. Phages were pu-rified twice by PEG 20% polyethylene glycol-8000,2.5 M NaCl precipitation of culture supernatant. ThePEG precipitate was resuspended in the final volumeof 2.5 ml in TBS 50 mM Tris-HCl (pH 7.5), 150 mMNaCl [15,16].
Biopanning of Prostate Epithelial Cells
Cells were grown in 60-mm culture dishes to sub-confluence. Medium was aspirated and monolayerwas incubated for 20 min in phosphate-buffered saline(PBS) with 1% bovine serum albumin (BSA) in a coldroom. Phage (109 colony-forming units/ml) wasadded to blocking solution, and monolayers were in-cubated for 1 hr on a rocker plate in a cold room. Afterfive extensive washes with blocking buffer containing0.1% Tween 20, bound phage was eluted with glycine-HCL, pH 5.0. Tris-HCL, pH 9.0, was immediatelyadded to the eluate, and competent bacteria were in-fected by eluted phages for propagation. The eluatefrom the last round of biopanning was plated on thedishes, and DNA from randomly picked phages wassequenced using an automated sequencer.
Attachment Inhibition of Prostate Cells toDefined Substrates by cRGD Peptide or Selected
xxxRGDxxx Express Phage
Cyclic RGD pentapeptide RGDfV (described previ-ously [17]), commercially available cyclic RGD pep-tide (Gibco BRL), and inactive control peptide
GRGESP (Bachem California, Torrance, CA) in con-centrations of 5–30 mg/ml, or phages with the RGD-containing sequence (1010 virions/well) selected bybiopanning of prostate epithelial cells, were used inthis study. To examine the specificity of the cyclicRGDfV peptide in inhibition of DU145 cell attachmentto defined substrates, cells were detached by 0.05%Trypsin/0.01% EDTA solution, and Trypsin was im-mediately inactivated by addition of the 2-fold excessof Trypsin inhibitor in PBS. Twenty thousand cells perwell were added to 96-well plates. Wells were pre-coated with 10 mg/ml of vitronectin, laminin, fibro-nectin, or poly-L-lysine at 4°C overnight. After wash-ing with PBS, unspecific binding sites were blockedwith 1% BSA in PBS for 1 hr at 37°C. Wells coated with1% BSA were used to determine background adhe-sion. Peptides or phages were added to cell suspen-sion before seeding into wells. After incubation for 1hr at 37°C, unattached cells were washed out and at-tached cells were fixed with 2% glutaraldehyde in PBSfor 20 min. Fixed cells were stained with 0.1% crystalviolet in 0.2 M boric acid, pH 9.0. After solubilizationof crystal violet in 10% acetic acid, the absorbance at590 nm was measured on an ELISA plate reader. Allexperiments were performed in quadruplicate. Thesame experimental protocol was used when the block-ing ability of RGD-containing selected phages in inhi-bition of DU145, PC3, or BPH-1 attachment to fibro-nectin, vitronectin, laminin, or poly-L-lysine wastested.
To determine the ability of cRGDfV to promote celldetachment from culture plastic, peptide (final con-centration, 20 mg/ml) was added to the attached PZ-HPV-7, BPH, PrE PC-3, or DU145 cells growing for16–24 hr since the previous subcloning. Cell morphol-ogy was regularly observed, number of detached cellswas estimated by counting them in a hemocytometer,
TABLE I. Integrin and ECM Antibodies
AntigenClone orsera type Source
Functionallyblocking
Dilution foradhesion assays
Dilution forFACS and
immunocytochemistry
a2 P1E6 Telios Yes 1:100a3 P1B5 Telios Yes 1:500 1:100a4 P4G9 Telios Yes 1:500 1:100a5 P1D6 Gibco BRL Yes 1:500 1:100aVb3 LM609 Chemicon Yes 1:500aV NKI-M9 Chemicon Yes 1:500 1:100a6 NKI-GoH3 Chemicon Yes 1:500 1:100b1 4B4 Coulter Yes 1:1,000 1:500b3 BB10 Chemicon No 1:100 permeabilizedb3 3E1 Gibco BRL Unknown 1:100b5 AB1926 Chemicon Unknown 1:100 permeabilized
110 Romanov and Goligorsky

and attached cells were counted after fixation andcrystal violet staining, as described before [17].
Fluorescence-Activated Cell Sorting (FACS)
Cells were detached from tissue culture plastic withTrypsin-EDTA (0.05%/0.01%) or with Cell Dissocia-tion Solution (Sigma Chemical Co., St. Louis, MO) andwashed twice in cold PBS. Cell suspensions (5 × 105
cells) were incubated with primary antibodies for 30min on ice. Cell pellets were washed three times with5% casein in PBS and incubated in the dark with ap-propriate secondary fluorescent conjugates (MolecularProbes, Eugene, OR) diluted 1:100 for another 30 min.Cells were fixed in 1% paraformaldehyde after an ad-ditional three washes. Labeled cells were analyzed us-ing a Becton Dickinson FACScan system (Medford,MA).
Attachment of Prostate Cells toHUVEC Monolayers
HUVEC cells were grown in 24-well clusters andused for experiments after they formed confluentmonolayers. Experiments were performed in restingcells or in HUVEC activated for 4 hr with 100 U/ml ofrecombinant interleukin-1 (Sigma Chemical Co.), asdescribed before [18,19]. After this pretreatment, su-pernatants were removed and monolayers werewashed with EGM-2 media. PZ-HPV-7, BPH-1, PrE,PC-3, and DU145 cells detached from plastic with CellDissociation Solution (Sigma Chemical Co.) werestained with Cell Tracker intravital fluorescent dye,following the manufacturer’s instructions (MolecularProbes). The stained cells were added to HUVECmonolayers (2.5 × 104 cells per well), and after 60 minof incubation, wells were extensively rinsed with PBSto remove unattached cells. RGD-containing peptideand functionally blocking antibodies were added tosuspended prostate epithelial cells or to HUVEC, asindicated. After fixation for 20 min with 4% paraform-aldehyde in PBS, prostate cells were counted by fluo-rescent microscopy and intensity of fluorescence wasmeasured on Fluorescence Multi-Well Plate ReaderCyto Fluor2 (PerSeptive Biosystems, Framingham,MA). Every experimental condition was repeated atleast three times and cells were used at different pas-sages.
Western Blotting
Equal amounts of total proteins from the lysates ofHUVEC cells were subjected to 10% SDS/PAGE geland, after transfer to the Immobilon membrane (Mil-lipore, Bedford, MA), were blotted with anti-
fibronectin, anti-vitronectin, or anti-laminin antibod-ies. Secondary antibodies (anti-rabbit IgG conjugatedwith peroxidase) were used at a dilution of 1:2,000(Amersham, Arlington Heights, IL). The immunoreac-tive bands were visualized by the Super Signal Sub-strate (Pierce, Rockford, IL).
RESULTS
Effect of cRGD Peptide on Prostate Cell-Matrix Interactions
To examine the functional capacity of expressed in-tegrins, the interactions of BPH-1, PZ-HPV-7, DU145,and PC-3 cells with ECM components and the abilityof cyclic peptide cRGDfK to inhibit these interactionswere studied. Cell-matrix adhesion assays showedthat both types of immortalized cells attached withhighest affinity to fibronectin and with slightly loweraffinity to vitronectin, and these interactions wereRGD-dependent (Fig. 1). Attachment of these cells tolaminin was weaker, and the utilized concentrationsof RGD peptides failed to inhibit these interactions.DU145 cells attached with highest affinity to fibronec-tin, followed by laminin and vitronectin. The attach-ment of DU145 cells to fibronectin and vitronectin wasinhibited by 5–20 mg/ml of cRGDfK peptide. Thistreatment had no effect on the attachment to laminin(Fig. 1). PC-3 cells attached to fibronectin with highestaffinity, followed by laminin and vitronectin. Interac-tions of these cells with fibronectin and vitronectin
Fig. 1. Inhibition of prostate epithelial cell attachment to extra-cellular matrix components by RGD peptide. RGE, control non-inhibitory peptide; RGD, cyclic RGD peptide described in Mate-rials and Methods; FN, fibronectin; LN, laminin; VN, vitronectin.Attachment of DU145 cells to fibronectin with inactive RGE pep-tide was considered 100% standard for each experimental condi-tion.
Integrins in Carcinoma-Endothelium Interactions 111
were RGD-dependent. RGD at a concentration of 20mg/ml did not significantly inhibit adhesion to lami-nin (Fig. 1).
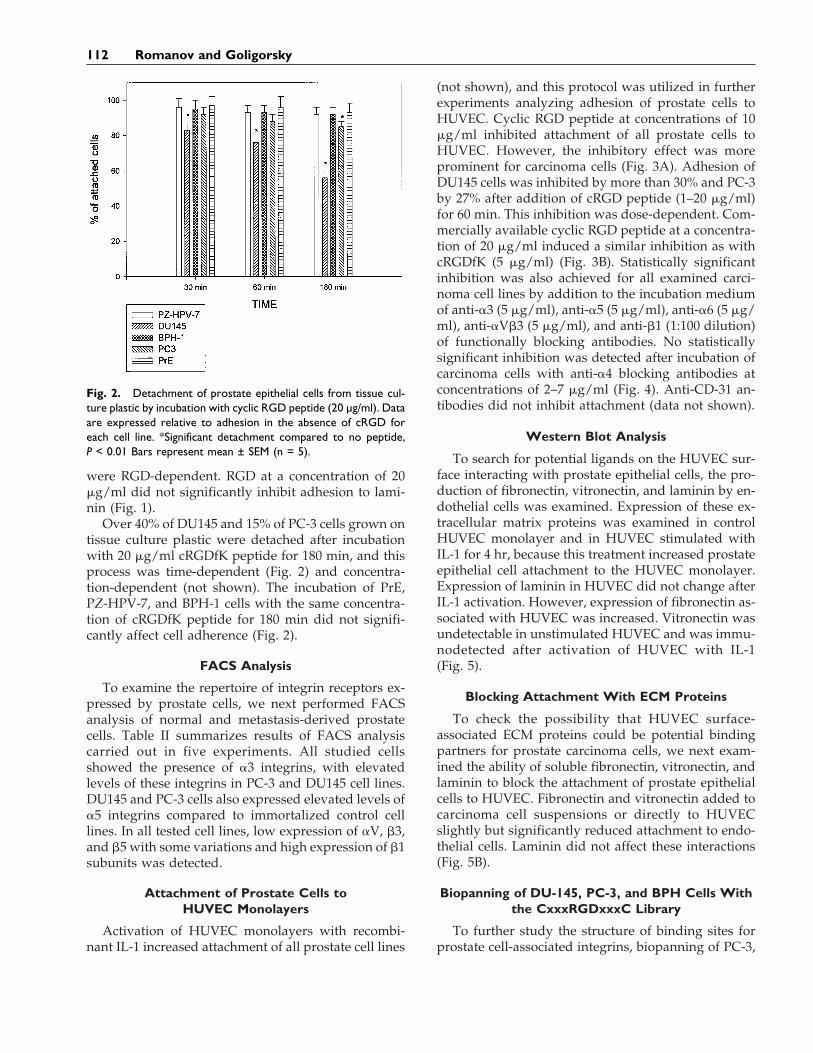
Over 40% of DU145 and 15% of PC-3 cells grown ontissue culture plastic were detached after incubationwith 20 mg/ml cRGDfK peptide for 180 min, and thisprocess was time-dependent (Fig. 2) and concentra-tion-dependent (not shown). The incubation of PrE,PZ-HPV-7, and BPH-1 cells with the same concentra-tion of cRGDfK peptide for 180 min did not signifi-cantly affect cell adherence (Fig. 2).
FACS Analysis
To examine the repertoire of integrin receptors ex-pressed by prostate cells, we next performed FACSanalysis of normal and metastasis-derived prostatecells. Table II summarizes results of FACS analysiscarried out in five experiments. All studied cellsshowed the presence of a3 integrins, with elevatedlevels of these integrins in PC-3 and DU145 cell lines.DU145 and PC-3 cells also expressed elevated levels ofa5 integrins compared to immortalized control celllines. In all tested cell lines, low expression of aV, b3,and b5 with some variations and high expression of b1subunits was detected.
Attachment of Prostate Cells toHUVEC Monolayers
Activation of HUVEC monolayers with recombi-nant IL-1 increased attachment of all prostate cell lines
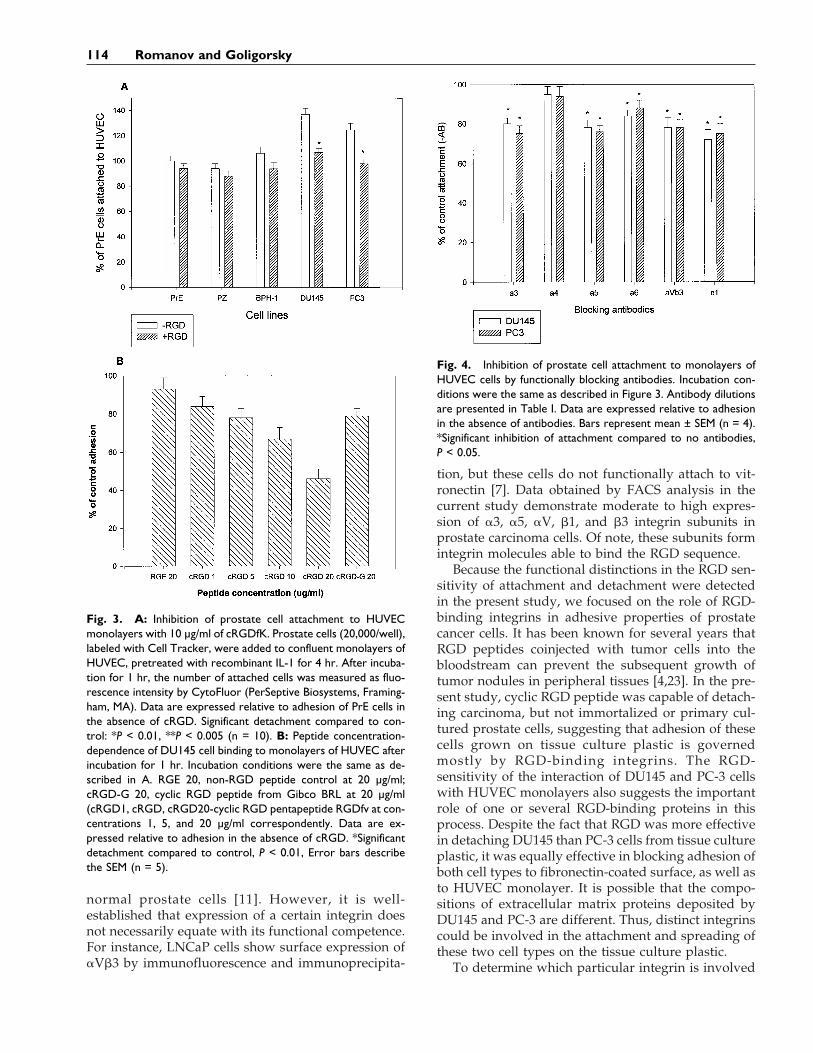
(not shown), and this protocol was utilized in furtherexperiments analyzing adhesion of prostate cells toHUVEC. Cyclic RGD peptide at concentrations of 10mg/ml inhibited attachment of all prostate cells toHUVEC. However, the inhibitory effect was moreprominent for carcinoma cells (Fig. 3A). Adhesion ofDU145 cells was inhibited by more than 30% and PC-3by 27% after addition of cRGD peptide (1–20 mg/ml)for 60 min. This inhibition was dose-dependent. Com-mercially available cyclic RGD peptide at a concentra-tion of 20 mg/ml induced a similar inhibition as withcRGDfK (5 mg/ml) (Fig. 3B). Statistically significantinhibition was also achieved for all examined carci-noma cell lines by addition to the incubation mediumof anti-a3 (5 mg/ml), anti-a5 (5 mg/ml), anti-a6 (5 mg/ml), anti-aVb3 (5 mg/ml), and anti-b1 (1:100 dilution)of functionally blocking antibodies. No statisticallysignificant inhibition was detected after incubation ofcarcinoma cells with anti-a4 blocking antibodies atconcentrations of 2–7 mg/ml (Fig. 4). Anti-CD-31 an-tibodies did not inhibit attachment (data not shown).
Western Blot Analysis
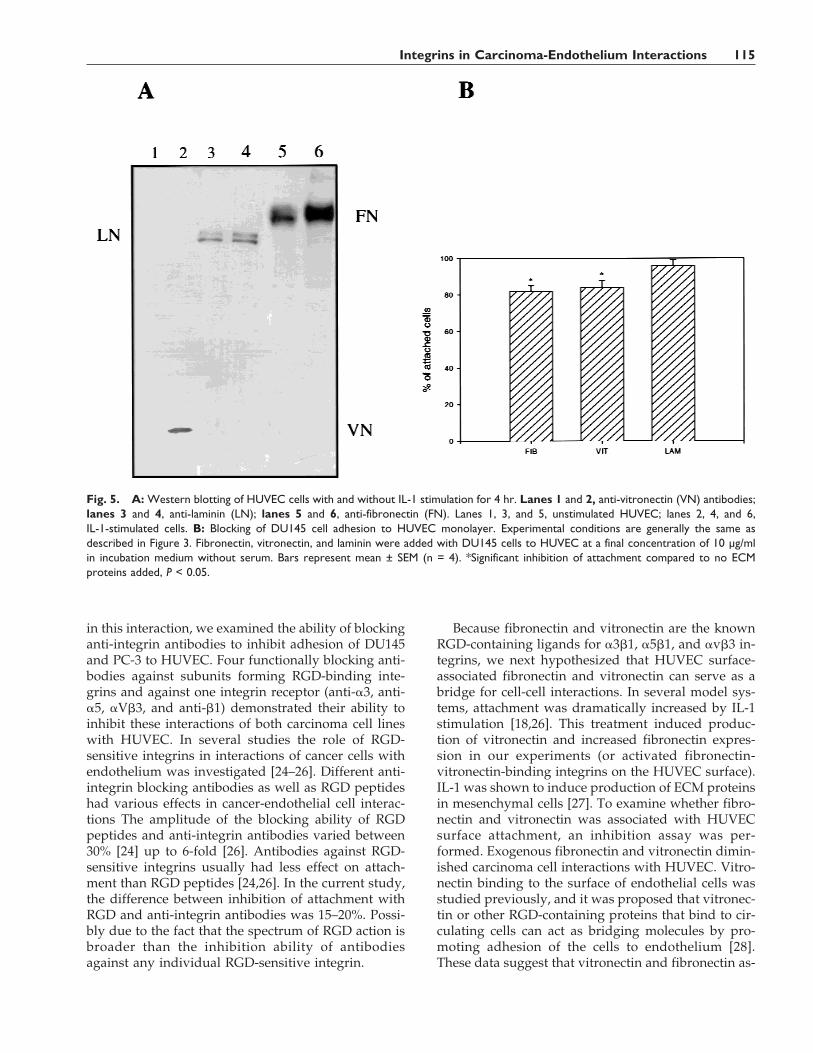
To search for potential ligands on the HUVEC sur-face interacting with prostate epithelial cells, the pro-duction of fibronectin, vitronectin, and laminin by en-dothelial cells was examined. Expression of these ex-tracellular matrix proteins was examined in controlHUVEC monolayer and in HUVEC stimulated withIL-1 for 4 hr, because this treatment increased prostateepithelial cell attachment to the HUVEC monolayer.Expression of laminin in HUVEC did not change afterIL-1 activation. However, expression of fibronectin as-sociated with HUVEC was increased. Vitronectin wasundetectable in unstimulated HUVEC and was immu-nodetected after activation of HUVEC with IL-1(Fig. 5).
Blocking Attachment With ECM Proteins
To check the possibility that HUVEC surface-associated ECM proteins could be potential bindingpartners for prostate carcinoma cells, we next exam-ined the ability of soluble fibronectin, vitronectin, andlaminin to block the attachment of prostate epithelialcells to HUVEC. Fibronectin and vitronectin added tocarcinoma cell suspensions or directly to HUVECslightly but significantly reduced attachment to endo-thelial cells. Laminin did not affect these interactions(Fig. 5B).
Biopanning of DU-145, PC-3, and BPH Cells Withthe CxxxRGDxxxC Library
To further study the structure of binding sites forprostate cell-associated integrins, biopanning of PC-3,
Fig. 2. Detachment of prostate epithelial cells from tissue cul-ture plastic by incubation with cyclic RGD peptide (20 µg/ml). Dataare expressed relative to adhesion in the absence of cRGD foreach cell line. *Significant detachment compared to no peptide,P < 0.01 Bars represent mean ± SEM (n = 5).
112 Romanov and Goligorsky

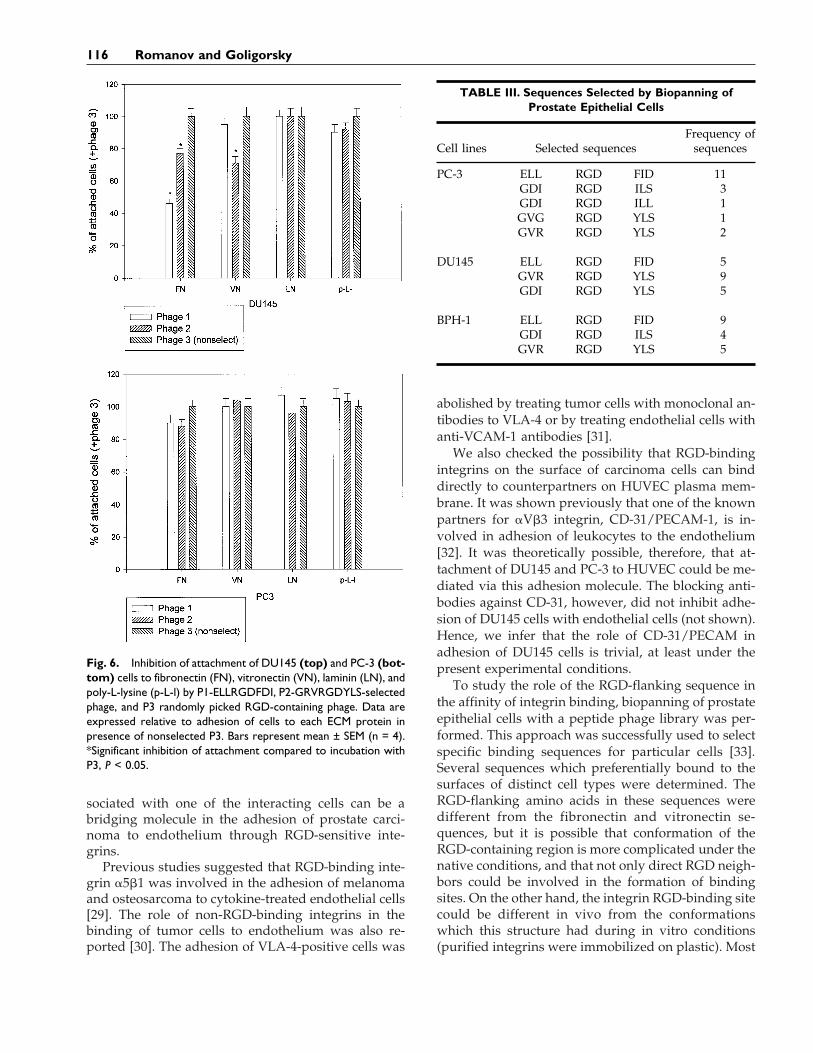
DU145, and BPH-1 cells with the peptide phage li-brary was performed. This library was constructed tostudy the role of RGD-flanking amino acids in inter-actions of RGD-containing ECM proteins with inte-grins. The number of transducing units eluted fromthe cell surface increased in second and third roundsof biopanning. After three rounds of biopanning, 20selected clones were randomly picked up for sequenc-ing. Several major motifs were present for all exam-ined cells. ELLRGDFID motif appeared preferentially(11/18) in the PC-3 cell line, and the GRVRGDYLSmotif with some variations was more often found inphage eluted from DU145 cells (14/19). Both se-quences and some derivatives were present equally inphage eluted from BPH-1 cells (Table III).
To study the specificity of selected phages, we usedthe two most frequently present sequences for block-ing the adhesion of all cell lines to fibronectin, laminin,vitronectin, and poly-L-lysine. Nonselected RGD-containing phage was used as a negative control. At-tachment of BPH-1 cells to all matrix proteins and topoly-L-lysine was slightly dependent on phage pres-ence in incubation media (data not shown). Phage 1was able to inhibit attachment of DU145 cells to fibro-nectin (56%) and to a lesser degree to vitronectin, butdid not inhibit attachment of PC3 to any ECM pro-teins. Phage 2 decreased attachment of DU145 to fi-bronectin and vitronectin and PC-3 to fibronectinonly. Nonselected RGD-containing phage did not sig-nificantly affect cell-matrix interactions and was usedas the 100% standard. None of the three phages waseffective in blocking attachment of all examined cellsto laminin (Fig. 6).
DISCUSSION
The data reported here provide functional evidencefor the distinct adhesive properties of cancer and nor-mal prostate epithelial cells. Two of the best-exploredtransformed prostate epithelial cell lines, two immor-talized normal prostate epithelial cell lines, and two
primary cultures of prostate were used in this study.DU145 and PC-3 cells are derived from prostate cancermetastasis, and both present tumorigenic activity invitro and in vivo as well as the ability to producemetastasis in nude mice [20]. Primary epithelial cellsderived from normal prostate as well as previouslycharacterized immortalized cells were used for com-parison. While cancer cells showed sensitivity to theeffects of cyclic RGD peptide, implying the expressionand engagement of RGD-recognizing integrins in theadhesion of DU145 and PC-3 cells to ECM compo-nents and HUVEC, the cells derived from the normalgland revealed much lower sensitivity to this peptide.
Generally, the pattern of integrins expressed intransformed cultured cells differed from the integrinrepertoire in normal cell lines, and also differed intumors compared with corresponding normal tissues.For instance, changes in the expression of fibronectin-binding integrins have been observed in some types oftransformed fibroblasts, while changes in the expres-sion of other integrins have been observed in variousmalignant cells [21]. Despite the absence of a strongdirect correlation between expression of particular in-tegrins and the aggression of tumor cells, there aresome general observations related to individual inte-grins. The a5- and a30-containing integrins have a ten-dency to be reduced in transformed cells, but upregu-lation of aVb3 accompanies some forms of malignantprogression [22].
Several studies compared integrin expression incells of prostatic origin [6–12]. Despite the general re-duction of integrin expression in prostate carcinomas,some malignant forms conserve moderate expressionof several integrins, e.g., a3, a5, and b1 [6]. Compari-son of the integrin profile of nontumorigenic (immor-talized cells or primary cultures) and cancerous celllines undertaken in other studies demonstrated an in-crease in the expression of several integrin subunits intumor cell lines. Expression of a2-, a3-, a5-, a6-, b1-,and b4-containing integrins, for instance, increased inDU145 cells compared with PNT-1A immortalized
TABLE II. FACS Analysis of a and b Integrin Subunits on HumanProstate Cell Lines*
Cell lines
Integrin subunits (median channel of peak fluorescence)
a3 a5 aV b1 b3 b5
PZ-HPV-7 11.3 7.4 6.1 64.3 2.1 1.3PrE 14.8 NA 7.3 91.3 2.8 NABPH-1 19.1 1.7 2.3 25.8 6.9 0.9PC-3 64.4 10.6 4.8 76.9 11.3 2.4DU145 40.9 23.7 7.1 68.4 3.1 1.9
*NA, data not available.
Integrins in Carcinoma-Endothelium Interactions 113
normal prostate cells [11]. However, it is well-established that expression of a certain integrin doesnot necessarily equate with its functional competence.For instance, LNCaP cells show surface expression ofaVb3 by immunofluorescence and immunoprecipita-
tion, but these cells do not functionally attach to vit-ronectin [7]. Data obtained by FACS analysis in thecurrent study demonstrate moderate to high expres-sion of a3, a5, aV, b1, and b3 integrin subunits inprostate carcinoma cells. Of note, these subunits formintegrin molecules able to bind the RGD sequence.
Because the functional distinctions in the RGD sen-sitivity of attachment and detachment were detectedin the present study, we focused on the role of RGD-binding integrins in adhesive properties of prostatecancer cells. It has been known for several years thatRGD peptides coinjected with tumor cells into thebloodstream can prevent the subsequent growth oftumor nodules in peripheral tissues [4,23]. In the pre-sent study, cyclic RGD peptide was capable of detach-ing carcinoma, but not immortalized or primary cul-tured prostate cells, suggesting that adhesion of thesecells grown on tissue culture plastic is governedmostly by RGD-binding integrins. The RGD-sensitivity of the interaction of DU145 and PC-3 cellswith HUVEC monolayers also suggests the importantrole of one or several RGD-binding proteins in thisprocess. Despite the fact that RGD was more effectivein detaching DU145 than PC-3 cells from tissue cultureplastic, it was equally effective in blocking adhesion ofboth cell types to fibronectin-coated surface, as well asto HUVEC monolayer. It is possible that the compo-sitions of extracellular matrix proteins deposited byDU145 and PC-3 are different. Thus, distinct integrinscould be involved in the attachment and spreading ofthese two cell types on the tissue culture plastic.
To determine which particular integrin is involved
Fig. 3. A: Inhibition of prostate cell attachment to HUVECmonolayers with 10 µg/ml of cRGDfK. Prostate cells (20,000/well),labeled with Cell Tracker, were added to confluent monolayers ofHUVEC, pretreated with recombinant IL-1 for 4 hr. After incuba-tion for 1 hr, the number of attached cells was measured as fluo-rescence intensity by CytoFluor (PerSeptive Biosystems, Framing-ham, MA). Data are expressed relative to adhesion of PrE cells inthe absence of cRGD. Significant detachment compared to con-trol: *P < 0.01, **P < 0.005 (n = 10). B: Peptide concentration-dependence of DU145 cell binding to monolayers of HUVEC afterincubation for 1 hr. Incubation conditions were the same as de-scribed in A. RGE 20, non-RGD peptide control at 20 µg/ml;cRGD-G 20, cyclic RGD peptide from Gibco BRL at 20 µg/ml(cRGD1, cRGD, cRGD20-cyclic RGD pentapeptide RGDfv at con-centrations 1, 5, and 20 µg/ml correspondently. Data are ex-pressed relative to adhesion in the absence of cRGD. *Significantdetachment compared to control, P < 0.01, Error bars describethe SEM (n = 5).
Fig. 4. Inhibition of prostate cell attachment to monolayers ofHUVEC cells by functionally blocking antibodies. Incubation con-ditions were the same as described in Figure 3. Antibody dilutionsare presented in Table I. Data are expressed relative to adhesionin the absence of antibodies. Bars represent mean ± SEM (n = 4).*Significant inhibition of attachment compared to no antibodies,P < 0.05.
114 Romanov and Goligorsky
in this interaction, we examined the ability of blockinganti-integrin antibodies to inhibit adhesion of DU145and PC-3 to HUVEC. Four functionally blocking anti-bodies against subunits forming RGD-binding inte-grins and against one integrin receptor (anti-a3, anti-a5, aVb3, and anti-b1) demonstrated their ability toinhibit these interactions of both carcinoma cell lineswith HUVEC. In several studies the role of RGD-sensitive integrins in interactions of cancer cells withendothelium was investigated [24–26]. Different anti-integrin blocking antibodies as well as RGD peptideshad various effects in cancer-endothelial cell interac-tions The amplitude of the blocking ability of RGDpeptides and anti-integrin antibodies varied between30% [24] up to 6-fold [26]. Antibodies against RGD-sensitive integrins usually had less effect on attach-ment than RGD peptides [24,26]. In the current study,the difference between inhibition of attachment withRGD and anti-integrin antibodies was 15–20%. Possi-bly due to the fact that the spectrum of RGD action isbroader than the inhibition ability of antibodiesagainst any individual RGD-sensitive integrin.
Because fibronectin and vitronectin are the knownRGD-containing ligands for a3b1, a5b1, and avb3 in-tegrins, we next hypothesized that HUVEC surface-associated fibronectin and vitronectin can serve as abridge for cell-cell interactions. In several model sys-tems, attachment was dramatically increased by IL-1stimulation [18,26]. This treatment induced produc-tion of vitronectin and increased fibronectin expres-sion in our experiments (or activated fibronectin-vitronectin-binding integrins on the HUVEC surface).IL-1 was shown to induce production of ECM proteinsin mesenchymal cells [27]. To examine whether fibro-nectin and vitronectin was associated with HUVECsurface attachment, an inhibition assay was per-formed. Exogenous fibronectin and vitronectin dimin-ished carcinoma cell interactions with HUVEC. Vitro-nectin binding to the surface of endothelial cells wasstudied previously, and it was proposed that vitronec-tin or other RGD-containing proteins that bind to cir-culating cells can act as bridging molecules by pro-moting adhesion of the cells to endothelium [28].These data suggest that vitronectin and fibronectin as-
Fig. 5. A: Western blotting of HUVEC cells with and without IL-1 stimulation for 4 hr. Lanes 1 and 2, anti-vitronectin (VN) antibodies;lanes 3 and 4, anti-laminin (LN); lanes 5 and 6, anti-fibronectin (FN). Lanes 1, 3, and 5, unstimulated HUVEC; lanes 2, 4, and 6,IL-1-stimulated cells. B: Blocking of DU145 cell adhesion to HUVEC monolayer. Experimental conditions are generally the same asdescribed in Figure 3. Fibronectin, vitronectin, and laminin were added with DU145 cells to HUVEC at a final concentration of 10 µg/mlin incubation medium without serum. Bars represent mean ± SEM (n = 4). *Significant inhibition of attachment compared to no ECMproteins added, P < 0.05.
Integrins in Carcinoma-Endothelium Interactions 115
sociated with one of the interacting cells can be abridging molecule in the adhesion of prostate carci-noma to endothelium through RGD-sensitive inte-grins.
Previous studies suggested that RGD-binding inte-grin a5b1 was involved in the adhesion of melanomaand osteosarcoma to cytokine-treated endothelial cells[29]. The role of non-RGD-binding integrins in thebinding of tumor cells to endothelium was also re-ported [30]. The adhesion of VLA-4-positive cells was
abolished by treating tumor cells with monoclonal an-tibodies to VLA-4 or by treating endothelial cells withanti-VCAM-1 antibodies [31].
We also checked the possibility that RGD-bindingintegrins on the surface of carcinoma cells can binddirectly to counterpartners on HUVEC plasma mem-brane. It was shown previously that one of the knownpartners for aVb3 integrin, CD-31/PECAM-1, is in-volved in adhesion of leukocytes to the endothelium[32]. It was theoretically possible, therefore, that at-tachment of DU145 and PC-3 to HUVEC could be me-diated via this adhesion molecule. The blocking anti-bodies against CD-31, however, did not inhibit adhe-sion of DU145 cells with endothelial cells (not shown).Hence, we infer that the role of CD-31/PECAM inadhesion of DU145 cells is trivial, at least under thepresent experimental conditions.
To study the role of the RGD-flanking sequence inthe affinity of integrin binding, biopanning of prostateepithelial cells with a peptide phage library was per-formed. This approach was successfully used to selectspecific binding sequences for particular cells [33].Several sequences which preferentially bound to thesurfaces of distinct cell types were determined. TheRGD-flanking amino acids in these sequences weredifferent from the fibronectin and vitronectin se-quences, but it is possible that conformation of theRGD-containing region is more complicated under thenative conditions, and that not only direct RGD neigh-bors could be involved in the formation of bindingsites. On the other hand, the integrin RGD-binding sitecould be different in vivo from the conformationswhich this structure had during in vitro conditions(purified integrins were immobilized on plastic). Most
Fig. 6. Inhibition of attachment of DU145 (top) and PC-3 (bot-tom) cells to fibronectin (FN), vitronectin (VN), laminin (LN), andpoly-L-lysine (p-L-l) by P1-ELLRGDFDI, P2-GRVRGDYLS-selectedphage, and P3 randomly picked RGD-containing phage. Data areexpressed relative to adhesion of cells to each ECM protein inpresence of nonselected P3. Bars represent mean ± SEM (n = 4).*Significant inhibition of attachment compared to incubation withP3, P < 0.05.
TABLE III. Sequences Selected by Biopanning ofProstate Epithelial Cells
Cell lines Selected sequencesFrequency of
sequences
PC-3 ELL RGD FID 11GDI RGD ILS 3GDI RGD ILL 1GVG RGD YLS 1GVR RGD YLS 2
DU145 ELL RGD FID 5GVR RGD YLS 9GDI RGD YLS 5
BPH-1 ELL RGD FID 9GDI RGD ILS 4GVR RGD YLS 5
116 Romanov and Goligorsky

previous panning experiments were performed underthese experimental conditions [4,34].
To examine the selectivity and specificity of definedsequences, an attachment inhibition assay was per-formed. Attachment of BPH-1 cells to all ECM pro-teins was slightly reduced with all phages (notshown). Phages 1 and 2 were able to inhibit attach-ment of PC-3 to fibronectin but not to vitronectin.However, DU145 attachment to fibronectin was inhib-ited with both phages (more effectively with phage 1).Attachment of DU145 cells to vitronectin was inhib-ited with phage 2 only. The differing effectiveness ofphages 1 and 2 in interactions of DU145 cells withfibronectin and vitronectin shows that the phageswere specifically selected as ligands for distinct inte-grin molecules. The lower effectiveness of both phagesin PC-3 interactions with fibronectin and lack of inhi-bition activity of both phages in PC-3 interactions withvitronectin can be explained by the differing func-tional status of integrins on the surface of these twocell types. Of course, it is extremely important to findthe peptide which will be equally effective in all kindsof prostate cancer. Improvement of the biopanningprocedure may help us to discover these universalpeptides. The RGD-binding motif is reasonably effec-tive at least in the interactions of prostate epithelialcells with HUVEC. The search for the most effectivepeptide(s) will continue.
The present findings suggest that RGD-binding in-tegrins expressed on the surface of prostate carcinomacells participate in interactions with HUVEC cells. Theknown ligands for these receptors could be fibronectinand vitronectin expressed on or associated with theapical cell surface and in the intercellular spaces.These data also demonstrate that the integrins respon-sible for binding of tumor cells to endothelium arespecific for individual tumor cells and can utilize dif-ferent counterparts on the surface of endothelial cells.Cyclic RGD peptide flanked by optimal amino acidscan potentially be useful as an inhibitor of these inter-actions that are crucial for the metastatic process.
ACKNOWLEDGMENTS
We are grateful to Dr. David Durand (Departmentof Pathology, State University of New York at StonyBrook) for reading the manuscript and for insightfuldiscussions, and to Dr. Horst Kessler (Institute of Bio-chemistry, Munich, Germany) for cyclic RGD peptide.
REFERENCES
1. Waltregny D, Castronovo V. Recent advances in prostate cancermetastasis. Tumori 1996;82:193–204.
2. Isaacs JT. Molecular markers for prostate cancer metastasis. AmJ Pathol 1997;150:1511–1521.
3. Honn KV, Tang DG. Adhesion molecules and tumor cell inter-actions with endothelium and subendothelial matrix. CancerMetastasis Rev 1992;11:353–375.
4. Ruoslahti E. RGD and other recognition sequences for integrins.Annu Rev Cell Dev Biol 1996;12:697–7155.
5. Varner JA, Cheresh DA. Integrins and cancer. Curr Opin CellBiol 1996;8:724–730.
6. Witkowski CM, Rabinovitz I, Nagle RB, Affinito K-SD, CressAE. Characterization of integrin subunits, cellular adhesion andtumorigenicity of four human prostate cell lines. J Cancer ResClin Oncol 1993;119:637–644.
7. Nagle RB, Knox JD, Wolf C, Bowden GT, Cress AE. Adhesionmolecules, extracellular matrix, and proteases in prostate carci-noma. J Cell Biochem [Suppl] 1994;19:232–237.
8. Knox JD, Cress AE, Clark V, Manriquez L, Affinito K-SD, DalkinBL, Nagle RB. Differential expression of extracellular matrixmolecules and the a6-integrins in the normal and neoplasticprostate. Am J Pathol 1994;145:167–174.
9. Cress AE, Rabinovitz I, Zhu W, Nagle RB. The a6b1 and a6b4integrins in human prostate cancer progression. Cancer Metas-tasis Rev 1995;14:219–228.
10. Rabinovitz I, Nagle RB, Cress AE. Integrin a6 expression inhuman prostate carcinoma cells is associated with a migratoryand invasive phenotype in vitro and in vivo. Clin Exp Metas-tasis 1995;13:481–491.
11. Haywood-Reid PL, Zipf DR, Springer WR. Quantification ofintegrin subunits on human prostatic cell lines: comparison ofnontumorigenic and tumorigenic lines. Prostate 1997;31:1–8.
12. Rokhlin OW, Cohen MB. Expression of cellular adhesion mol-ecules on human prostate tumor cell lines. Prostate 1995;26:205–212.
13. Lang SH, Clarke NW, George NJR, Testa NG. Primary prostaticepithelial cell binding to human bone marrow stroma and therole of a2b1 integrin. Clin Exp Metastasis 1997;15:218–227.
14. Weijerman PC, Konig JJ, Wong ST, Niesters HGM, Peehl DM.Lipofection-mediated immortalization of human prostatic epi-thelial cells of normal and malignant origin using human pap-illomavirus type 18 DNA. Cancer Res 1994;54:5579–5583.
15. Smith GP, Scott JK. Libraries of peptides and proteins displayedon filamentous phage. Methods Enzymol 1993;217:228–257.
16. Adley NB, Sparks AB, Beasley J, Kay BK. Construction of ran-dom peptide libraries in bacteriophage M13. In: Kay BK, WinterJ, McCafferty J, editors. Phage display of peptides and proteins.New York: Academic Press; 1996. p 67–78.
17. Romanov V, Noiri E, Czerwinski G, Finsinger D, Kessler H,Goligorsky M. Two novel probes reveal tubular and vascularArg-Gly-Asp (RGD) binding sites in the ischemic rat kidney.Kidney Int 1997;52:93–102.
18. Dejana E, Bertocchi F, Bortolami MC, Regonesi A, Tonto A,Breviario F, Giavazzi R. Interleukin 1 promotes tumor cell ad-hesion to cultured human endothelial cells. J Clin Invest 1988;82:1466–1470.
19. Bertomeu MC, Gallo S, Lauri D, Haas T, Orr FW, Bastida E,Buchanan MR. Interleukin 1-induced cancer cell/endothelialcell adhesion in vitro and its relationship to metastasis in vivo:role of vessel wall 13-HODE synthesis and integrin expression.Clin Exp Metastasis 1993;11:243–250.
20. Webber MM, Bello D, Quader S. Immortalized and tumorigenicadult human prostatic epithelial cell lines: characteristics andapplications part 2. Tumorigenic cell lines. Prostate 1997;30:58–64.
21. Giancotti FG, Mainiero F. Integrin-mediated adhesion and sig-naling in tumorigenesis. Biochim Biophys Acta 1994;1198:47–64.
Integrins in Carcinoma-Endothelium Interactions 117

22. Juliano RL, Varner JA. Adhesion molecules in cancer: the role ofintegrins. Curr Opin Cell Biol 1993;5:812–818.
23. Humphries MJ, Akiyama SK, Komoriya A, Olden K, YamadaKM. Identification of an alternative spliced site in humanplasma fibronectin that mediates cell type specific adhesion. JCell Biol 1986;103:2637–2647.
24. Bliss RD, Kirby JA, Browell DA, Lennard TWJ. The role of b1integrins in adhesion of two breast carcinoma cell lines to amodel endothelium. Clin Exp Metastasis 1995;13:173–183.
25. Fornaro M, Tallini G, Bofetiado CJM, Bosari S, Languino LR.Down-regulation of b1C integrin, an inhibitor of cell prolifera-tion in prostate carcinoma. Am J Pathol 1996;149:765–773.
26. Lauri D, Bertomeu M-C, Orr FW, Bastida E, Sauder D, BuchananMR. Interleukin-1 increases tumor cell adhesion to endothelialcells through an RGD dependent mechanism: in vitro and invivo studies. Clin Exp Metastasis 1990;8:27–32.
27. Ebert EC. Mechanisms of colon cancer binding to substratumand cells. Dig Dis Sci 1996;41:1551–1556.
28. Zanetti A, Conforti G, Hess S, Martin-Padura I, Ghibaudi E,Preissner KT, Dejana E. Clustering of vitronectin and RGD pep-tides on microspheres leads to engagement of integrins on theluminal aspect of endothelial cell membrane. Blood 1994;84:1116–1123.
29. Lauri D, Martin-Padura I, Biondelli T. Role of b1 integrins intumor cell adhesion to cultured human endothelial cells. LabInvest 1991;65:525–531.
30. Rice GE, Gimbrone MA, Bevilacqua MP. Tumor cell-endothelialinteractions. Increased adhesion of human melanoma cellsto activated vascular endothelium. Am J Pathol 1988;133:204–210.
31. Martin-Padura J, Mortarini R, Lauri D, Bernasconi S, Sanches-Madrid F, Parmiani G, Mantovani A, Anichini A, Dejana E.Heterogenity in human melanoma cell adhesion to cytokine ac-tivated endothelial cells correlates with VLA-4 expression. Can-cer Res 1991;51:2239–2241.
32. Pauli L, Hammel P, Uherek C, Bachmann F, Gisler RH, DunonD. CD31/PECAM-1 is a ligand for aVb3 integrin involved inadhesion of leukocytes to endothelium. J Cell Biol 1995;130:451–460.
33. Barry MA, Dower WJ, Johnston SA. Toward cell-targeting genetherapy vectors: selection on cell-binding peptides from randompeptide-presenting phage libraries. Nat Med 1996;2:299–305.
34. Koivunen E, Wang B, Dickinson CD, Ruoslahti E. Peptides incell adhesion research. Methods Enzymol 1994;245:346–369.
118 Romanov and Goligorsky








![Effects of the RGD loop and C-terminus of rhodostomin on … · 2017. 5. 3. · RGD loop to regulate integrins recognition [8, 11, 16–22]. For example, Marcinkiewicz et al. reported](https://img.dokumen.tips/doc/110x75/611e0fdbc7885320dd5190dc/effects-of-the-rgd-loop-and-c-terminus-of-rhodostomin-on-2017-5-3-rgd-loop.jpg)



















