Embed Size (px)
Citation preview

www.elsevier.com/locate/clinchim
Clinica Chimica Acta 342 (2004) 1–12
Review
Peripheral markers of blood–brain barrier damage
Nicola Marchia, Marco Cavagliaa, Vincent Fazioa, Sunil Bhudiab,Kerri Hallenea, Damir Janigroa,*
aDepartment of Neurological Surgery, Cerebrovascular Research Center, NB20, The Cleveland Clinic Foundation,
9500 Euclid Avenue, Cleveland, OH 44195, USAbDepartment of Thoracic and Cardiac Surgery, The Cleveland Clinic, Cleveland, OH 44199, USA
Received 1 September 2003; received in revised form 3 December 2003; accepted 4 December 2003
Abstract
Neurological diseases are often associated with cerebrovascular dysfunction and changes in blood–brain barrier (BBB)
function. This is important for two seemingly conflicting reasons. On the one hand, a leaky BBB may lead to brain disease by
allowing extravasation of cells and molecules normally segregated in the periphery, while on the other hand an intact BBB may
hamper drug delivery to the ailing brain. Under both circumstances, it would be desirable to follow closely over time BBB
‘‘tightness’’. Several lines of evidence have suggested that the astrocytic protein S100h is a potentially useful peripheral marker
of BBB permeability. Other markers of brain-to-blood barriers have been recently discovered by a proteomic approach. These
proteins are virtually absent in normal blood, appear in serum from patients with cerebral lesions, and can be easily detected. We
will present clinical and laboratory evidence supporting the use of these markers as modern neurodiagnostic tools.
D 2004 Elsevier B.V. All rights reserved.
Keywords: S100h; BBB (blood–brain barrier); Blood–CSF barrier; Endothelium; Cerebral ischemia; MRI (magnetic resonance imaging);
Neurological disorders; NSE (neuron-specific enolase); TTR (transthyretin); Neurodiagnostic tools
The discovery that the entry of many substances Not all the blood vessels in the brain constitute a
into the brain is restricted has been generally attribut-
ed to Paul Ehrlich, who observed in the early 1880s
that aniline dyes injected into animals intravenously
colored all the organs except for the brain and spinal
cord [1]. The singular failure of the brain to be stained
at autopsy during jaundice must have been observed
before Ehrlich’s work, but apparently did not steer
inquiry into its mechanisms.
0009-8981/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.cccn.2003.12.008
* Corresponding author. Tel.: +1-216-445-0561; fax: +1-216-
445-1466.
E-mail address: [email protected] (D. Janigro).
blood–brain barrier (BBB): only capillary vessels are
endowed with a full-blown BBB phenotype. Vessels
of increasing diameter have comparably increasing
levels of leakiness and thus superficial vessels of large
diameter are the leakiest while penetrating pial vessels
and descending penetrating vessels tend to have an
intermediate barrier function. Since most animals,
including vertebrates, have some form of barrier
separating their blood circulation from the brain or
the central nervous system, it has been speculated that
profound evolutionary pressure existed to create such
a complex organ [2,13]. At present, however, it is not
clear why such sophistication exists, and we can only

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–122
speculate on the teleological pressure that manifests in
the specialization observed in endothelial cells (EC) of
the brain.
The ontogeny of the mammalian blood–brain bar-
rier may be related to the absolute need for controlled
homeostasis of the central nervous system. It has been
known for many years that the ionic and molecular
composition of the cerebrospinal fluid (CSF) differs
considerably from the composition of the plasma and
remains remarkably constant in spite of intravascular
changes. For example, proteins are few in the CSF, and
ionic gradients for potassium and calcium ions exist
across the blood–brain barrier. Glucose constitutes the
main energy supply to the brain and is avidly con-
sumed by neuronal cells. To ensure a constant supply
of glucose to the brain, even under conditions where
plasma glucose is low, the blood–brain barrier main-
tains nearly a 10-fold gradient for glucose entry into
the brain [3]. Other substances are actively transported
into and out of the brain, including amino acids,
metallic ions, etc.
Homeostatic control of the brain parenchyma by
vascular endothelial cells is also achieved by a func-
tion that is seemingly opposed to their ‘‘barrier’’
function. For example, the abluminal membrane of
the endothelium facing the glia end feet is endowed
with a high concentration of sodium/potassium trans-
porter molecules that permit absorption of excess
potassium from the brain into the blood [4–8]. Failure
of such a mechanism could lead to an abnormal
accumulation of potassium into the brain leading to
neurological disorders such as seizures. Control of
parenchymal pH may also depend on the integrity of
brain endothelial cells.
1. Anatomy and physiology of the normal blood–
brain barrier
The BBB is primarily composed of microvascular
EC linked by tight junctions that largely prevent
molecular communication between the plasma and
the brain or central nervous system [9–12]. Only
capillary vessels have complete BBB properties. As
vessel diameter increases, leakiness also increases:
superficial vessels of large diameter are the leakiest,
and penetrating pial vessels and descending penetrat-
ing vessels have intermediate degrees of barrier func-
tion. Surprisingly, in spite of the large body of
evidence on the neurophysiological properties of the
central nervous system, comparatively little is known
about the physiology and pathophysiology of micro-
vascular EC.
The long axis of these cells is perpendicular to
blood flow, and they are exquisitely sensitive to shear
stress. Recent evidence has shown that shear stress
promotes the expression of numerous genes involved
in various aspects of endothelial cell function [12,14–
16]. It is unknown whether flow-induced changes in
EC affect neighboring glia, and whether these changes
affect expression of astrocytic proteins that may be
detected in plasma. Some of the unique properties of
the BBB are induced by the perivascular glia. This is
known from two lines of evidence. First, it is evident
that shear stress alone cannot be responsible for BBB
properties in EC, because systemic microvascular EC
are exposed to comparable levels of shear stress.
Second, glia cells have been shown to induce
blood–brain barrier properties in vessels of systemic
origin [17,18]. Thus, the blood–brain barrier is com-
monly understood to be constituted of both endothe-
lial cells and glial end feet. Perivascular pericytes and
microglia may also be considered active components
of the blood–brain barrier.
The microvascular endothelium shares a common
basement membrane with astrocytes and pericytes.
Beyond the basement membrane in the parenchymal
vessels of the brain lies a close investment of end feet
from neuroglial cells, predominantly astrocytes.
Astrocytes and their processes invest more than 90%
of endothelial capillaries, and their end feet are
projected tightly around the endothelial cells. There-
fore, the glial end feet are a natural candidate for
mediating communication between neurons and cap-
illaries. Astrocytic proteins are synthesized and re-
leased next to capillaries, but it is thought that they
extravasate into the plasma only when the BBB is
breached.
2. Protein permeability at the BBB
Brain-derived proteins may be useful markers of
BBB integrity because they have several possible
mechanisms of passage across the BBB. Under
physiologic conditions, the production of CSF from

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–12 3
plasma involves efficient removal of plasma pro-
teins by filtering mechanisms in the choroid plexus.
The resulting CSF contains extremely small quan-
tities of proteins [19]. However, many neurological
disorders are associated with elevated CSF pro-
tein levels. Furthermore, CSF-specific proteins exist
[20].
Fig. 1. Gradients and topography of peripheral markers of BBB/brain dam
immunopositive astrocytes and NSE immunoreactivity in human neocorte
highly concentrated in proximity to blood vessels (indicated by small arro
indicate 20 Am. n = neuron, a = astrocyte.
Proteins in CSF can be detected by directly sam-
pling CSF, which requires invasive techniques such as
lumbar puncture or intraoperative sampling from
ventricles or subarachnoid space. BBB integrity can
also be assessed by contrast-enhanced computed to-
mography or MRI (see above). An obvious limitation
of intrathecal detection methods is that they are
age. See text for details. The micrographs show GFAP and S100hx (temporal lobe). Note that GFAP and S100h immunoreactivity is
ws). A large NSE+ neuron neighboring a vessel is also shown. Bars

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–124
invasive and that the sample itself may be contami-
nated by the procedure. Accurate non-invasive tech-
niques would clearly be preferable, particularly to
analyze multiple longitudinal samples. A small group
of proteins are found exclusively or almost exclusive-
ly in the cerebrospinal fluid. Any disruption in BBB
integrity may allow protein leakage in both directions
(see Fig. 1). Thus, testing serum levels of CSF
proteins represents a non-invasive means for evaluat-
ing BBB integrity that may be of diagnostic value
[21]. Fig. 2 shows the predicted behavior of proteins
in conditions of intact vs. impaired BBB function.
Scenarios A and B demonstrate the observed elevation
in S100h serum levels as the result of BBB opening
following and prior to neuronal damage (respective-
Fig. 2. Possible mechanisms of release of putative markers of neuronal func
dots) may be released after neuronal injury; (B) before injury across a leak
the absence of neuronal damage. Hypothesis A predicts that a marker tha
plasma compartment only after brain injury. In contrast, if BBB dysfunctio
under normal conditions, appearance in plasma will be temporally relate
release due to increased synthesis or release in the CSF. However, if the
availability in the CNS from neuronal damage, the appearance in plasma
ly). Scenario C exemplifies the use of a chemical
means (mannitol infusion) that has been used to
produce a transient opening of the BBB without
producing accompanying neuronal damage.
3. Blood–brain barrier in neurological disorders
Substantial progress has been made in the under-
standing of the pathophysiology and mechanisms
involved in the attenuation of BBB permeability. In
many diseases that affect the brain, the cerebral
endothelium plays an active part in the disease process
with the BBB becoming disrupted, or modified, in
such a way that there is a dramatic increase in vascular
tion or blood–brain barrier damage across the BBB. (A) Marker (red
y blood–brain barrier, with additional release after injury; or (C) in
t is released or synthesized in response to injury will appear in the
n precedes injury and if the marker is present in CSF but not plasma
d to opening of the blood–brain barrier, with a possible additional
BBB opening is not followed by significant increase in marker’s
will be brief in duration.

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–12 5
permeability. Theoretically, several ways exist in
which various molecules can pass the endothelium.
These include intercellular routes, vesicular transport
or direct transcellular penetration through damaged
endothelium. BBB dysfunction may be a cause or
consequence of a particular disease process. Cerebral
vessels from pathological brain tissue often resemble
peripheral endothelium, with significant increases in
vesicular transport and abnormal tight junctions. Dis-
eases in which increased BBB permeability have been
reported include neoplasia, ischemia, hypertension,
dementia, epilepsy, infection, multiple sclerosis and
trauma. The effect of a disease on BBB function will
secondarily affect the cerebral blood flow and vascu-
lar tone in the brain, which further influences transport
across the BBB. Besides the effects of increased
vascular permeability on the brain parenchyma, a
question of great significance is whether, in certain
neuropathological conditions, the BBB disturbance
constitutes the main pathogenic factor itself, which
then triggers a sequence of events molding the final
pathological state.
While loss of blood brain barrier (BBB) function is
an etiologic component of many neurological diseases
an intact BBB may restrict the delivery of certain
therapeutic substances to the brain. Thus, measuring
BBB function may be important to diagnose disease
progression and monitor time-dependent changes in
BBB integrity when chemotherapic penetration may
be enhanced. At present, only invasive and expensive
techniques such as contrast-enhanced magnetic reso-
nance imaging, CT-scan and lumbar puncture are
available to clinically assess BBB integrity. An alter-
native approach has been proposed, consisting of
detection of changes in blood composition that indi-
cate BBB disruption [21].
Current BBB assessment by imaging or cerebro-
spinal fluid sampling is based on direct or indirect
determination of protein permeability across the
BBB. CNS proteins are normally asymmetrically
distributed, with generally much higher concentration
in plasma than in CSF. Thus, the appearance of
plasma proteins in CSF is a hallmark of numerous
CNS disorders with presumed or overt BBB disrup-
tion. Only a few proteins are synthesized exclusively
by, or are present in higher concentrations in CSF or
interstitial compartment compared to the blood.
These CSF markers may appear or increase their
plasma concentration after passage across a failed
BBB. Therefore, measuring levels of CSF proteins in
plasma may be a reliable way to monitor blood to
CNS barrier integrity without the use of invasive
methods.
4. Neuronal damage vs. glial/BBB damage
Most research into brain damage has focused on
neuronal damage, because this is the cause of most
deficits from neurological disease. In fact, ‘‘brain
damage’’ has often been used as a synonym for
neuronal damage or death. This research has shown
that neuronal sensitivity to insult is region-and dis-
ease-specific. As example, ischemic insult selectively
affects the CA1 region of the hippocampus, leaving
the neighboring dentate gyrus and CA3 practically
intact [22]. In contrast, damage resulting from epilep-
tic seizures is more prominent in CA3 hippocampal
subfield [23]. In addition to these patterns of speci-
ficity, it has also been observed that neuronal cell
death does not occur concomitantly with the insult,
but rather after a delay.
In acute insults such as ischemia [24,25], the delay
provides a potential therapeutic window for neuro-
protective intervention. In chronic and progressive
neurological diseases, such as multiple sclerosis, the
delay may be even longer. Ideal markers of BBB
permeability and of neuronal damage share several
characteristics (Table 1): both should be virtually
undetectable in normal subjects and should show
distinct alterations in response to insults that are
correlated with the severity of the damage.
Distinguishing between BBB defects and neuronal
damage has enormous clinical relevance. For exam-
ple, in acute CNS disturbances such as ischemic
stroke, the delay between insult and irreversible
neuronal cell death offers a window of therapeutic
opportunity. If BBB openings develop early after the
initial arterial occlusion [26–32], clinicians would
have a unique opportunity to administer drugs that
are normally BBB-impermeant (e.g., nerve growth
factors) before neurons were damaged. The duration
of these openings may be unpredictable, so a periph-
eral, non-invasive, easily repeatable test would be
extremely useful. In chronic neurological diseases,
such as multiple sclerosis, BBB openings may have

Table 1
Properties of selected proteins and their use as potential markers
BBB integrity
S100h NSE GFAP
(1) Low or
undetectable
levels in plasma
in normal subjects
yes no no
(2) Normally
present in CSF
yes yes yes
(3) [Plasma] < [CSF] yes no no
(4) Increase [CSF] in
response to insult
yes yes yes
(5) Normally excluded
by BBB
yes yes yes
(6) Expected flux
after BBBD
blood-to-
brain
brain-to-
blood
brain-to-
blood
(1) Stroke 28 (1997) 1956–1960; (2) Clin. Chem. 43 (1997) 541–
543; (3) Clin. Chem. 46, (2000) 993–996; (4) Clin. Chem. 44
(1998) 1056–1058; (5) Stroke 31 (2000) 2670–2677; (6) Perfusion
12 (1997) 171–177; (7) Glia 22 (1997) 1–11.
N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–126
both therapeutic and etiologic significance. Severity
of symptoms has been suggested to correlate with
BBB function in these conditions, and promising
therapies using brain-derived proteins have failed
largely because the compounds are poorly transported
across the BBB. When a patient experiences both
blood–brain barrier opening and neuronal damage,
plasma levels of both markers would be expected to
exceed normal levels [54]. Also, as we will discuss
below, S100 protein levels appear to be directly corre-
lated with BBB integrity rather than with neuronal
damage, while another neuronal protein, monomeric-
transthyretin (TTR) represents a potential marker of
opening to the blood–CSF barrier [48].
Because of this focus on neuronal damage, much
of the previous research on biochemical markers has
focused on markers that measure neuronal damage.
However, most neurologic diseases are accompanied
by increased BBB permeability, and thus the markers
thought to indicate neuronal damage may in fact
indicate BBB defects. Marker proteins under investi-
gation have included neuron-specific enolase (NSE),
glial fibrillary acidic protein (GFAP), NSE and S100h(see Fig. 1). S100h seems particularly promising. In
normal subjects, NSE is more concentrated in plasma
and S100h is primarily present in central nervous
system fluids [33]. Thus, opening the blood–brain
barrier in the absence of neuronal damage would be
expected to markedly increase plasma S100h levels
while leaving NSE levels unchanged.
5. Markers of blood–brain barrier integrity:
S100B
S100B is primarily synthesized in the brain by the
end feet process of the astrocytes and is quickly
released from the brain in the blood when the BBB
is disrupted [21,34–37]. S100B has also been found
in other tissues but at lower concentrations [38–40].
While S100B appearance in plasma correlated well
with BBB openings, S100B has been shown to in-
crease in plasma, CSF or both as a consequence of
other pathologies not limited to the CNS. S100B may
also detect brain damage, or indicate advanced me-
tastasis in melanoma patients [41–45].
The fact that S100B can increase in serum inde-
pendent of brain (or neuronal) damage was demon-
strated indirectly by a team of scientists who studied
the affects of boxing as well as other high cardiovas-
cular output activities on the levels of S100B in serum.
Interestingly, this study found that a significant in-
crease in S100B was observed in serum of subjects
undertaking activities that involved repetitive, jarring
movement or contact to the head (such as boxing,
sparring, running and jogging), but essentially no
increase was observed in persons exerting themselves
through exercise on a stationary bicycle. Clearly, these
activities did neither cause nor promoted brain dam-
age but the rise in S100B protein in running activities
may be due to astroglial activation, astroglial destruc-
tion or blood–brain barrier disruption or a combina-
tion of the three. This finding also indicated that the
source of S-100B was not influenced greatly from
excretion of the protein from extra-cranial tissue.
Another study demonstrated dissociation between
serum S100B and brain damage. S100B was measured
in sera from patients diagnosed with major depression
[46]. This study discovered a positive correlation of
the severity of depression with S100B levels; patients
given antidepressive therapy (prescription anti-depres-
sants) experienced significantly decreased levels of
serum S100B.
A direct demonstration proving that S100B may
increase in serum in spite of an obvious lack of

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–12 7
additional brain damage was provided by studies
where S100B was measured at short intervals
(minutes) in patients undergoing osmotic opening of
the blood–brain barrier [21,34,47,48]. These authors
demonstrated that S100B elevation correlated tempo-
rally with BBB disruption; these findings were also
confirmed radiologically by Kanner et al. [47]. To test
the connection between S100B and blood–brain bar-
rier integrity, NSE and S100B were measured in
serum of patients with primary central nervous lym-
phoma who underwent blood–brain barrier disruption
by intra-arterial mannitol infusion before receiving
methotrexate infusion. Mean serum levels of S100B
increased significantly after mannitol infusion and
again after methotrexate infusion, and they remained
elevated through recovery. However, NSE serum
levels remained constant throughout the procedure
and during recovery. Previous investigators have
shown that blood–brain barrier disruption with intra-
arterial methotrexate does not lead to brain damage
[49–51]. To rule out the possibility that the increased
plasma S100B levels were caused by the methotrex-
ate, the authors measured S100B and NSE in the
blood of three patients who were given methotrexate
Fig. 3. Different distribution between S-100h and TTR in the brain. In the
the BBB, whereas TTR is synthesized by the choroid plexuses and is foun
the different roles of these markers.
without blood–brain barrier disruption. Under these
conditions (no BBB disruption) the levels of both
markers remained within normal ranges. It was con-
cluded that the increase in S100B immediately after
the blood–brain barrier disruption was almost cer-
tainly too soon to be the result of synthesis and
release from ‘‘reactive’’ glia and that S100B may be
an early marker of blood–brain barrier disruption that
is not necessarily related to either neuronal or glial
brain damage. These findings of course do not chal-
lenge the traditional understanding that NSE is related
to neuronal damage.
In addition to S100B, another putative marker of
blood–brain barrier function was unveiled in these
studies. The CSF protein, monomeric-TTR was also
temporally linked to S100B movement from brain to
serum [48]. However, since the authors observed a
slight, but significant delay (within minutes) between
the peak serum elevation of S100B (immediately
following chemical opening of the BBB) and TTR
(occurring shortly thereafter following the administra-
tion of chemotherapy), it was concluded that TTR was
a marker of blood-to-CSF rather than a BBB marker
(see Fig. 3).
brain, S-100h is primarily synthesized by the astrocytes surrounded
d in the ventricular CSF. This topographic segregation may explain
N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–128
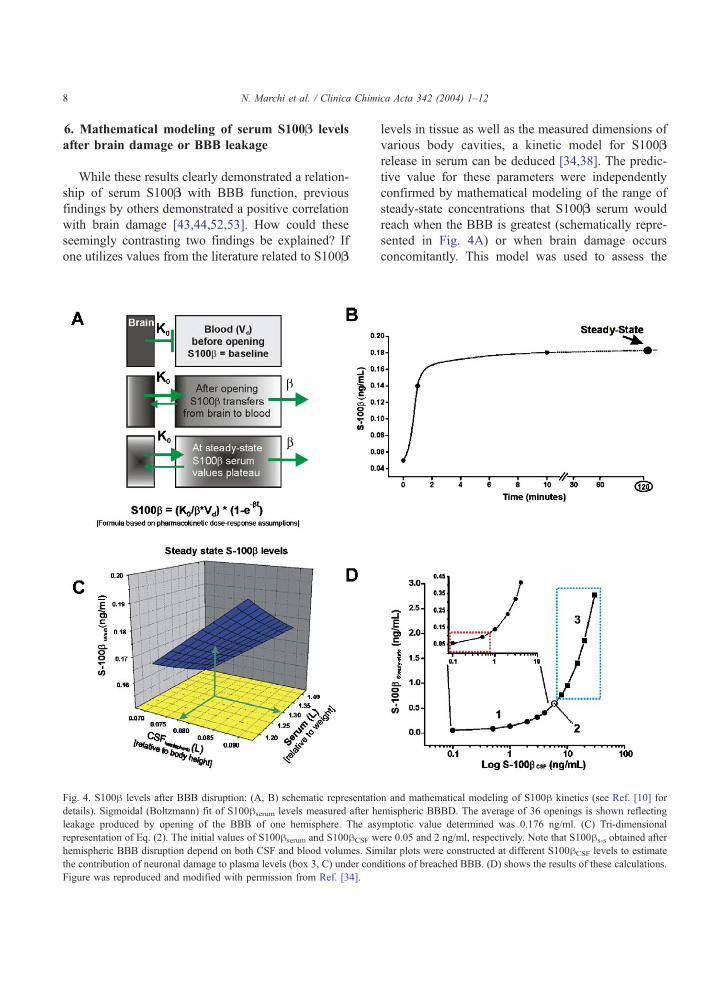
6. Mathematical modeling of serum S100B levels
after brain damage or BBB leakage
While these results clearly demonstrated a relation-
ship of serum S100B with BBB function, previous
findings by others demonstrated a positive correlation
with brain damage [43,44,52,53]. How could these
seemingly contrasting two findings be explained? If
one utilizes values from the literature related to S100B
Fig. 4. S100h levels after BBB disruption: (A, B) schematic representatio
details). Sigmoidal (Boltzmann) fit of S100hserum levels measured after h
leakage produced by opening of the BBB of one hemisphere. The as
representation of Eq. (2). The initial values of S100hserum and S100hCSF w
hemispheric BBB disruption depend on both CSF and blood volumes. Sim
the contribution of neuronal damage to plasma levels (box 3, C) under con
Figure was reproduced and modified with permission from Ref. [34].
levels in tissue as well as the measured dimensions of
various body cavities, a kinetic model for S100B
release in serum can be deduced [34,38]. The predic-
tive value for these parameters were independently
confirmed by mathematical modeling of the range of
steady-state concentrations that S100B serum would
reach when the BBB is greatest (schematically repre-
sented in Fig. 4A) or when brain damage occurs
concomitantly. This model was used to assess the
n and mathematical modeling of S100h kinetics (see Ref. [10] for
emispheric BBBD. The average of 36 openings is shown reflecting
ymptotic value determined was 0.176 ng/ml. (C) Tri-dimensional
ere 0.05 and 2 ng/ml, respectively. Note that S100hs-s obtained after
ilar plots were constructed at different S100hCSF levels to estimate
ditions of breached BBB. (D) shows the results of these calculations.

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–12 9
dependency of S100B steady state on serum and CSF
volumes as well as CSF levels of the protein. The
initial values used were derived from the literature
(e.g., S100B CSF= 2 ng/ml and S100B serum = 0.05
ng/ml). Data were fitted according to the following
equation:
S100hs�s ¼ ½ðS100hCSF*1=2CSFvolÞ þ ðS100hser
*SerumvolÞ�=½1=2CSFvol þ Serumvol�ð1Þ
where S100hs-s is the steady-state serum concentration
after hemispheric opening of the barrier; S100hser/CSF
are the reference concentrations of S100h in serum and
CSF expressed in ng/ml; CSFvol and serumvol are
volumes of these compartments expressed in liters.
The resulting three-dimensional plot is shown in Fig.
4C to demonstrate the dependence of S100hs-s on CSF
and blood volume. As expected, the peak levels of
S100hs-s are achieved when CSF volume is greatest
and serum lowest. As predicted by our direct experi-
mental observation and fitting, these values were again
close to 0.18 ng/ml (green arrow), which closely
parallels the amount leaked from the CNS after hemi-
spheric BBB disruption. These data and fits were
based on CSF S100h levels typical of uninjured brain.
To estimate the steady-state values of S100h serum at
different S100h CSF and under condition of bi-hemi-
spheric BBB damage, we used the following equation
(Fig. 4D):
S100hs�s ¼ ½ðCSFvol*S100hCSFÞ þðserumvol*0:05Þ�=½CSFvol þ serumvol� ð2Þ
Values for CSF and serum volume were arbitrarily set
at 0.15 and 1.5 l, respectively, to reflect the average
volumes for serum and cerebrospinal fluid. The red
region in Fig. 4D represents S100hs-s within a range
that includes normal values and levels that may be
achieved by breaching the BBB in absence of damage
(our findings). The data point 2 refers to data from
Buttner et al. [55], where experimentally measured
CSF values of 6 Ag/l corresponded to serum levels of
around 0.6 Ag/l. Note that these values were identicalto those predicted by our model. The boxed blue region
(3) represents S100hCSF levels measured in patients
affected by a variety of neurological diseases
[38,43,44,52,54–60]. These levels were compared in
the same study with S100serum. Again, a perfect
correlation of CSF/serum ratios with data obtained
from our model was found.
Taken together, these experimental results and
mathematical modeling demonstrate that the maximal
levels of S100hs-s achievable after BBB failure are
around 0.34 ng/ml (Fig. 4C). Thus, levels of S100hs-s
exceeding this value may be due to other factors, such
as non-CNS release [57], synthesis ex novo due to
damage, or other mechanisms.
7. Conclusions
S100h, neuron-specific enolase and other putative
markers of brain damage have been shown to corre-
late with outcome in a variety of neurological dis-
orders [57–59,61,62]. The interpretation of the
clinical significance of the appearance of S100h in
serum of neurological patients is complicated by the
fact that the cerebral circulation, unlike the coronary
vascular network, is characterized by tight junctions
between endothelial cells. The presence of tight junc-
tions is the molecular basis of the so-called blood–
brain barrier, a specialized endothelial structure effec-
tively shielding the brain from systemic influences.
The presence of this endothelial barrier minimizes the
extravasation of a variety of molecules including CSF
(or serum) S100h. Thus, detection of passage of
albumin from serum to brain is the preferred clinical
method to evaluate BBB intactness by either direct
measurements (lumbar puncture) or contrast-enhanced
CT-MRI where albumin is chemically linked to radio-
opaque ions (e.g., gadolinium). The opposite ap-
proach, detection of S100h protein in serum, is also
possible in virtue of the fact that this protein is almost
exclusively present in brain astrocytes.
The fact that S100hserum may be used as marker of
BBB integrity is not necessarily in disagreement with
the notion that S100h is a marker of brain damage,
since both phenomena (BBB failure and brain dam-
age) are temporally and topographically associated.
A possible explanation of the dual message that
levels of S100hserum may convey was derived mathe-
matically [34]. According to these authors, low levels
of S100h are normally present at the blood-to-brain
interface and in the CSF while disruption of the BBB

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–1210
will result in sudden appearance of cerebral S100h in
serum. It was possible to estimate the steady-state
levels of S100h that are when (1) The BBB is com-
pletely leaky; (2) levels of S100hCSF do not increase
over time due to neuroglial damage; and (3) CSF and
serum concentrations are constant. Furthermore, simi-
lar analysis was performed for S100hCSF levels typical
of a broad range of cerebral dysfunction. Serum levels
of S100h exceeding the ‘‘BBB ceiling’’ may implicate
brain damage or release from non-CNS sources.
In conclusion, interpretation of recent results and
existing literature compelled us to reinterpret the sig-
nificance of S100h as marker of brain damage. Exper-
imental, clinical and theoretical data show that: (1)
S100h is a marker of both BBB and neuronal damage;
(2) threshold serum values indicating brain damage can
be estimated; (3) conditions exist when S100hserum is
low in spite of massive brain damage; and (4) detection
of slightly elevated levels of S100hserum may be an
early sign of future neuronal damage, triggered or
accompanied by blood–brain barrier failure.
Acknowledgements
The authors would like to thank Anne-Charlotte
Aronsson for her helpful comments and support. This
work was supported by NIH-2RO1 HL51614, NIH-
RO1 NS 43284 and NIH-RO1 NS38195 to Damir
Janigro.
References
[1] Ehrlich BE, Diamond JM, Braun LD, Cornford EM, Olden-
dorf WH. Effects of lithium on blood–brain barrier transport
of the neurotransmitter precursors choline, tyrosine and tryp-
tophan. Brain Res 1980;193:604–7.
[2] Abbott NJ, Pichon Y. The glial blood–brain barrier of crus-
tacea and cephalopods: a review. J Physiol 1987;82:304–13.
[3] McAllister MS, Krizanac-Bengez L, Macchia F, et al. Mecha-
nisms of glucose transport at the blood–brain barrier: an in
vitro study. Brain Res 2001;904:20–30.
[4] Stummer W, Betz AL, Keep RF. Mechanisms of brain
ion homeostasis during acute and chronic variations of
plasma potassium. J Cereb Blood Flow Metab 1995;15:
336–44.
[5] D’Ambrosio R, Wenzel J, Schwartzkroin PA, Janigro D. Func-
tional specialization and topographic segregation of hippo-
campal astrocytes. J Neurosci 1998;18:1–14.
[6] D’Ambrosio R, Maris DO, Grady MS, Winn HR, Janigro D.
Impaired K homeostasis and altered electrophysiological
properties of post-traumatic hippocampal glia. J Neurosci
1999;19:8152–62.
[7] McKhann GM, D’Ambrosio R, Janigro D. Heterogeneity of
astrocyte resting membrane potentials revealed by whole cell
and gramicidin-perforated patch recordings from cultured neo-
cortical and hippocampal slice astrocytes. J Neurosci 1997;
17:6850–63.
[8] Betz AL, Keep RF, Beer ME, Ren XD. Blood–brain barrier
permeability and brain concentration of sodium, potassium,
and chloride during focal ischemia. J Cereb Blood Flow
Metab 1994;14:29–37.
[9] Neuwelt EA, Abbott NJ, Drewes L, et al. Cerebrovascular
biology and the various neural barriers: challenges and future
directions. Neurosurgery 1999;44:604–9.
[10] Abbott NJ. Comparative physiology of the blood–brain
barrier. In: Bradbury MW, editor. Physiology and pharma-
cology of the blood–brain barrier. Berlin: Springer; 1992.
p. 373–96.
[11] Grant GA, Abbott NJ, Janigro D. Understanding the physio-
logy of the blood–brain barrier: in vitro models. News Physiol
Sci 1998;13:287–93.
[12] Janigro D, Grant GA. The blood–brain barrier. In: Winn HR,
editor. Youman’s neurosurgery. 5th ed. Philadelphia: Saun-
ders; 2003. p. 153–74.
[13] Butt AM, Jones HC, Abbott NJ. Electrical resistance across
the blood–brain barrier in anaesthetized rats: a developmental
study. J Physiol (Lond) 1990;429:47–62.
[14] Ballermann BJ, Dardik A, Eng E, Liu A. Shear stress and
the endothelium. Kidney Int Suppl 1998;67:S100–8.
[15] Desai SY, Marroni M, Cucullo L, et al. Mechanisms of
endothelial survival under shear stress. Endothelium 2002;
9:89–102.
[16] Krizanac-Bengez L, Kapural M, Parkinson F, et al. Effects of
transient loss of shear stress on blood–brain barrier endotheli-
um: role of nitric oxide and IL-6. Brain Res 2003;977: 239–46.
[17] Cancilla PA, Bready J, Berliner J. Brain endothelial–astrocyte
interactions. In: Pardridge WM, editor. The blood–brain bar-
rier cellular and molecular biology. New York: Raven Press;
1993. p. 25–47.
[18] Stanness KA, Westrum LE, Fornaciari E, et al. Morphological
and functional characterization of an in vitro blood–brain
barrier model. Brain Res 1997;771:329–42.
[19] Reiber H, Peter JB. Cerebrospinal fluid analysis: disease-re-
lated data patterns and evaluation programs. J Neurol Sci
2001;184:101–22.
[20] Davson H, Segal MB. The proteins and other macromolecules
of the CSF. In: Davson H, Segal MB, editors. Physiology of
the CSF and of the blood–brain barrier. New York, NY: CRC
Press; 1995. p. 573–606.
[21] Kapural M, Bengez L, Barnett G, et al. S-100B as a possible
serum marker for disruption on the blood–brain barrier. Brain
Res 2002;940/1–2:102–4.
[22] Kirino T, Tamura A, Sano K. Selective vulnerability of the
hippocampus to ischemia: reversible and irreversible types of
ischemic cell damage. Progr Brain Res 1985;63:39–58.

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–12 11
[23] Babb TL. Synaptic reorganizations in human and rat hippo-
campal epilepsy. In: Delgado-Escueta AV, Wilson WA, Olsen
RW, Porter RJ, editors. Japser’s basic mechanisms of the ep-
ilepsies. Third edition. Philadelphia, PA: Lippincott Williams
and Wilkins; 1999. p. 763–79.
[24] Endres M, Namura S, Shimizu-Sasamata M, et al. Attenuation
of delayed neuronal death after mild focal ischemia in mice by
inhibition of the caspase family. J Cereb Blood Flow Metab
1998;18:238–47.
[25] Fink K, Zhu J, Namura S, et al. Prolonged therapeutic window
for ischemic brain damage caused by delayed caspase activa-
tion. J Cereb Blood Flow Metab 1998;18:1071–6.
[26] Ling BN, O’Neill WC. Ca(2+)-dependent and Ca(2+)-perme-
able ion channels in aortic endothelial cells. Am J Physiol
1992;263:H1827–38.
[27] Mossakowski MJ, Lossinsky AS, Pluta R, Wisniewski HM.
Abnormalities of the blood–brain barrier in global cerebral
ischemia in rats due to experimental cardiac arrest. Acta Neu-
rochir Suppl 1994;60:274–6.
[28] Pluta R, Lossinsky AS, Wi’sniewski HM, Mossakowski MJ.
Early blood–brain barrier changes in the rat following tran-
sient complete cerebral ischemia induced by cardiac arrest.
Brain Res 1994;633:41–52.
[29] Nakamura Y, Iga K, Shibata T, Shudo M, Kataoka K. Glial
plasmalemmal vesicles: a subcellular fraction from rat hippo-
campal homogenate distinct from synaptosomes. Glia 1993;9:
48–56.
[30] Rosenberg GA, Estrada EY, Dencoff JE. Matrix metalloprotei-
nases and TIMPs are associated with blood–brain barrier open-
ing after reperfusion in rat brain. Stroke 1998;29: 2189–95.
[31] Aschner M, Vrana KE, Zheng W. Manganese uptake and
distribution in the central nervous system (CNS). Neuroto-
xicology 1999;20:173–80.
[32] Del Bigio MR, Yan HJ, Buist R, Peeling J. Experimental
intracerebral hemorrhage in rats. Magnetic resonance ima-
ging and histopathological correlates. Stroke 1996;27:
2312–20.
[33] Reiber H. Cerebrospinal fluid-physiology, analysis and inter-
pretation of protein patterns for diagnosis of neurological di-
seases. Mult Scler 1998;4:99–107.
[34] Marchi N, Rasmussen PA, Kapural M, Fazio V, Cavaglia M,
Janigro D. Peripheral markers of brain damage and blood–
brain barrier dysfunction. Restor Neurol Neurosci 2003;21:
109–21.
[35] Buccoliero AM, Caldarella A, Noccioli B, Fiorini P, Taddei A,
Taddei GL. Brain heterotopia in pharyngeal region. A mor-
phological and immunohistochemical study. Pathol Res Pract
2002;198:59–63.
[36] Dyck RH, Van Eldik LJ, Cynader MS. Immunohistochemical
localization of the S-100 beta protein in postnatal cat visual
cortex: spatial and temporal patterns of expression in corti-
cal and subcortical glia. Brain Res Dev Brain Res 1993;
72:181–92.
[37] Mercier F, Hatton GI. Immunocytochemical basis for a menin-
geo-glial network. J Comp Neurol 2000;420:445–65.
[38] Jonsson H, Johnsson P, Alling C, Backstrom M, Bergh C,
Blomquist S. S100B after coronary artery surgery: release
pattern, source of contamination, and relation to neuropsycho-
logical outcome. Ann Thorac Surg 1999;68: 2202–8.
[39] Mrak RE, Flanigan S, Collins CL. Malignant acoustic
schwannoma. Arch Pathol Lab Med 1994;118:557–61.
[40] Rickmann M, Wolff JR. Modifications of S100-protein immu-
noreactivity in rat brain induced by tissue preparation. Histo-
chem Cell Biol 1995;103:135–45.
[41] Brochez L, Naeyaert JM. Serological markers for melanoma.
Br J Dermatol 2000;143:256–68.
[42] Chakrabarty A, Franks AJ. Meningioangiomatosis: a case re-
port and review of the literature. Br J Neurosurg 1999;13:
167–73.
[43] Grocott HP, Arrowsmith JE. Serum S100 protein as a marker
of cerebral damage during cardiac surgery. Br J Anaesth 2001;
86:289–90.
[44] Ingebrigtsen T, Romner B, Marup-Jensen S, et al. The clinical
value of serum S-100 protein measurements in minor head
injury: a Scandinavian multicentre study. Brain Inj 2000;14:
1047–55.
[45] Jonsson H, Johnsson P, Alling C, Westaby S, Blomquist S.
Significance of serum S100 release after coronary artery by-
pass grafting. Ann Thorac Surg 1998;65:1639–44.
[46] Rothermundt M, Arolt V, Wiesmann M, et al. S-100B is in-
creased in melancholic but not in non-melancholic major de-
pression. J Affect Disord 2001;66:89–93.
[47] Kanner AA, Marchi N, Fazio V, et al. Serum S100beta:
a noninvasive marker of blood–brain barrier function and
brain lesions. Cancer 2003;97:2806–13.
[48] Marchi N, Fazio V, Cucullo L, et al. Serum transthyretin as a
possible marker of blood-to-CSF barrier disruption. J Neurosci
2003;23:1949–55.
[49] Crossen JR, Goldman DL, Dahlborg SA, Neuwelt EA. Neu-
ropsychological assessment outcomes of nonacquired immu-
nodeficiency syndrome patients with primary central nervous
system lymphoma before and after blood–brain barrier dis-
ruption chemotherapy [see comments]. Neurosurgery 1992;
30:23–9.
[50] Kroll RA, Neuwelt EA. Outwitting the blood–brain barrier
for therapeutic purposes: osmotic opening and other means.
Neurosurgery 1998;42:1083–99.
[51] Neuwelt EA, Frenkel EP, Diehl J, Vu LH, Rapoport SI, Hill
SA. Reversible osmotic blood–brain barrier disruption in
humans: implications for the chemotherapy of brain tumors.
Neurosurgery 1980;7:44–52.
[52] Ingebrigtsen T, Waterloo K, Jacobsen EA, Langbakk B,
Romner B. Traumatic brain damage in minor head injury:
relation of serum S-100 protein measurements to magnetic
resonance imaging and neurobehavioral outcome. Neurosur-
gery 1999;45:468–75.
[53] Romner B, Ingebrigtsen T, Kongstad P, Borgesen SE. Traumat-
ic brain damage: serum S-100 protein measurements related to
neuroradiological findings. J Neurotrauma 2000;17: 641–7.
[54] Barregard L, Wikkelso C, Rosengren LE, et al. Cerebrospinal
fluid proteins in men with chronic encephalopathy after expo-
sure to organic solvents. Scand J Work Environ Health 1990;
16:423–7.
[55] Buttner T, Weyers S, Postert T, Sprengelmeyer R, Kuhn W. S-

N. Marchi et al. / Clinica Chimica Acta 342 (2004) 1–1212
100 protein: serum marker of focal brain damage after ische-
mic territorial MCA infarction. Stroke 1997;28:1961–5.
[56] Fassbender K, Schmidt R, Schreiner A, et al. Leakage of
brain-originated proteins in peripheral blood: temporal profile
and diagnostic value in early ischemic stroke. J Neurol Sci
1997;148:101–5.
[57] Grocott HP, Laskowitz DT, Newman MF. Markers of cerebral
injury. In: Newman SP, Harrison MJG, editors. The brain and
cardiac surgery. Amsterdam, NL: Harwood Academic Pub-
lishers; 2001. p. 113–42.
[58] Johnsson P, Lundqvist C, Lindgren A, Ferencz I, Alling C,
Stahl E. Cerebral complications after cardiac surgery assessed
by S-100 and NSE levels in blood. J Cardiothorac Vasc
Anesth 1995;9:694–9.
[59] Vermuyten K. Determination of glial fibrillary acidic protein,
S100, myelin basic protein and neuron specific enolase in
cerebrospinal fluid from patients suffering from dementia.
Acta Neurol Belg 1989;89:318.
[60] Westaby S, Johnsson P, Parry AJ, et al. Serum S100 protein:
a potential marker for cerebral events during cardiopulmonary
bypass. Ann Thorac Surg 1996;61:88–92.
[61] Zimmer DB, Cornwall EH, Landar A, Song W. The S100
protein family: history, function, and expression. Brain Res
Bull 1995;37:417–29.
[62] Soler Federsppiel BS, Karcher D, Lowenthal A. Blood and
cerebrospinal fluid anomalies in brain ageing and Alzheimer’s
disease. Gerontology 1987;33:193–6.





























![Beyond the Blood-Brain Barrier - UCLA CTSI · Beyond the Blood-Brain Barrier: ... Circumventing the blood-brain barrier ... K30 presentation final clean.ppt [Read-Only] Author:](https://img.dokumen.tips/doc/110x75/5b0543887f8b9a0a548e9fa1/beyond-the-blood-brain-barrier-ucla-ctsi-the-blood-brain-barrier-circumventing.jpg)