Embed Size (px)
Citation preview

The I'eteHnaU.flmrna11997, 154, 11-34
Review
+
Modes of Action of Anthelmintic Drugs
R.J. MARTIN
Depa,/men/ o/ l',e,li,i,al Vete,'i,m,y .Sciences. R. CDOS. V.S., Su,n,,,e,hall. University' of Edinbu,gh, Edinburgh Etq9 I QH, UK
SUMMARY
Modes of action of anthelmintic drugs are described. Some anthelmintic drugs act rapidly and selectively on neuronmscular transmission of nematodes. Levamisole, pyrantel and morantel are agonists at nicotinic acetylcholine receptors of nematode muscle and cause spastic paralysis. Dichlorvos and haloxon are organophosphorus cholinesterase antagonists. Piperazine is a GABA (T-amino-bu~ric acid) agonist at receptors on nematode muscles and causes flaccid paralysis. The avermectins increase the opening of glutanmte-gated chh)ride (GltlC1) channels and produce paralysis of phmTngeal pumping. Praziquantel has a selective efl'ect on the tegument of trematodes and increases permeability of calcium. Other anthehnintics have a biochemical mode of action. The benzimidazole drugs bind selectively to ~-tubulin of nematodes, cestodes and fluke, and inhibit nficrotubule formation. The salicylanilides: rafoxanide, oxych)zanide, brotianide and closantel and the substituted phenol, nitroxynil, are proton ionophores. Clorsuhm is a selective antagonist of fluke phosphoglycerate kinase and mutase. Diethylcarbamazine blocks host, and possibly parasite, enzymes involved in arachidonic acid metabolism, and enhances the innate, nonspecific immune system. Kx-~WOnl)S: Mode of action; anthehnintics; levamisole; dichlorvos; piperazine; avermectins; praziquantel; benzimidazoles; salicylanilides; clorsulon; diethylcarbamazine.
I N T R O D U C T I O N
Parasitic nematodes affect animals and man caus- ing considerable suffering and poor growth. Effective anthehnintic drugs, used to treat and control these infestations, must have selective toxic effects on these parasites. Unfortunately with the increased use of these compounds, anthelmintic resistance has appeared and increased in fl-equency (Prichard, 1994). If resist- ance to a particular anthehnintic has occurred, it is likely that another anthelmintic with the same mode of action will also be ineffective although other anthehnintics with another mode of action, may still be effective. Clearly then, it is important to have an understanding of the mode of action of anthehnintics in order to inform the selection of effective therapeutic agents. This rexqew describes modes of action of anthelmintic drugs (Table I).
NICOTINIC AGONISTS
Levamisole, lmtamisole, pyrantel, morantel, oxantel, bephenium and thenium
Fig. 1 shows the chemical structures of nicotinic anthelmintics. There are the imidazothiazoles (levamisole and butamisole); the tetrahydropyrim- idines (pyrantel, morantel and oxantel); the quat- ernat T ammonium salts (bephenium and theniuna) and tile pyrimidines (methyridine). These compounds act selectively as agonists at syn- aptic and extrasynaptic nicotinic acetTlcholine receptors on nematode muscle cells (Fig. 2) and produce contraction and spastic paralysis. The electrophysiological effects of levamisole, pyrantel, morantel and oxantel have been studied in greatest detail.
1 (190-0233/t)7/04(1011-24/S 12.00/0 © 1997 Bailli6re Tindall

12 THE VEH.:RIN..\RY .IOL'RNAI.. 154, I
Table I. Summary table o f the m o d e s o f action o f anthelmintic drugs
(;e,eric drueff n a m e
Nicotinic agonists
Acetylcholinesterase inhibitors
GABA agonist
GiuCI potentiators
Calcium permeability increase
[~-tubulin binding
Proton ionophores
Inhibition of malate metabolism
Inhibition of phosphoglycerate kinase and mutase
Inhibitor of arachidonic acid metabolism and stimulation of innate immunity
I cvan l iso le I l l l l an l i so l t ' pV la t l te l ii1( ii+~lll tcl I ) c p h c n h l i l i l h t ' l l i l l n i nlcthvr id inc
11;llOXOll dichhwvos
p ipura/h ic
ivt ' l+l l l t 'Cli l l a l )a l l l cc l i l l d~WalllCCli l l inox idcci in ni i l l )c invcin l)
praziqu,in tel
thial)cnda]<dc Cal l l i ) t , l ld~izt i l t + ~lxil)cndaz<dc all)cndazolc albcndazoic sulphoxidc I{ 'nl)cndazoic tlxl{.lldazoh. lllt'l)('llda/i lit ' Ihd)cndazole I~'bantcl nclo l ) i ln in lh ioj)hanaic tr ic labcnda/olc
chls<tntcl i-al i lyanidt + oxvch izi.illidc I)r(>tianidc ni l l~,xvni l n ic lopholan ] l t 'xachlorophcnt. d i l ) r l l m o s a l a n nich;samidc
dia lnphent ' lh idc
C](II'SII](III
dicthvlcarl)amazine
Iqxam/de,~ ,1 ,w,me (". K. & l '.&A. Trade .\'mm,.s
Nilverm Slyq l l i l l Stl( ingid P;u'alct ' l
( :an l l l )a r
Mulitwttrma
End. r id .
[V(IIIR'C
l)t~('tOlll;tX ( :v(I t 'ct i l l
l) l~mcit
Th ibcnz . l c Equibcn, N~vil)cn koditac Valb,lzcn Rvc<d)cn Pilll[I('lll" Svstamcx Tchniu Fhil)vn~l ~alVVtTIII I Icpadcx \V< wlnal:tc, Ncmal,tx Fasincx
Flukivcr Flukanidc. Ranidu Zanil In Fhlkombin and Vermadcx Trodax, l)ov<.'n ix BilevolL Dist<don ( :oopaphcnc, l)ismdin
( '.ol-il)an
In h'omec Super
(:aricide,Filaricide
Electrophysiological effects of nicotinic anthelmint ics in nematodes
Intracelhllar recordings made with micropip-
e t tes f l 'mn Ascaris suum b o d y m u s c l e s have shown tha t b a t h a p p l i c a t i o n o f t e t r a m i s o l e , t he D/l. r a c e m i c m i x t u r e o f l evamiso le , p r o d u c e s n t em-
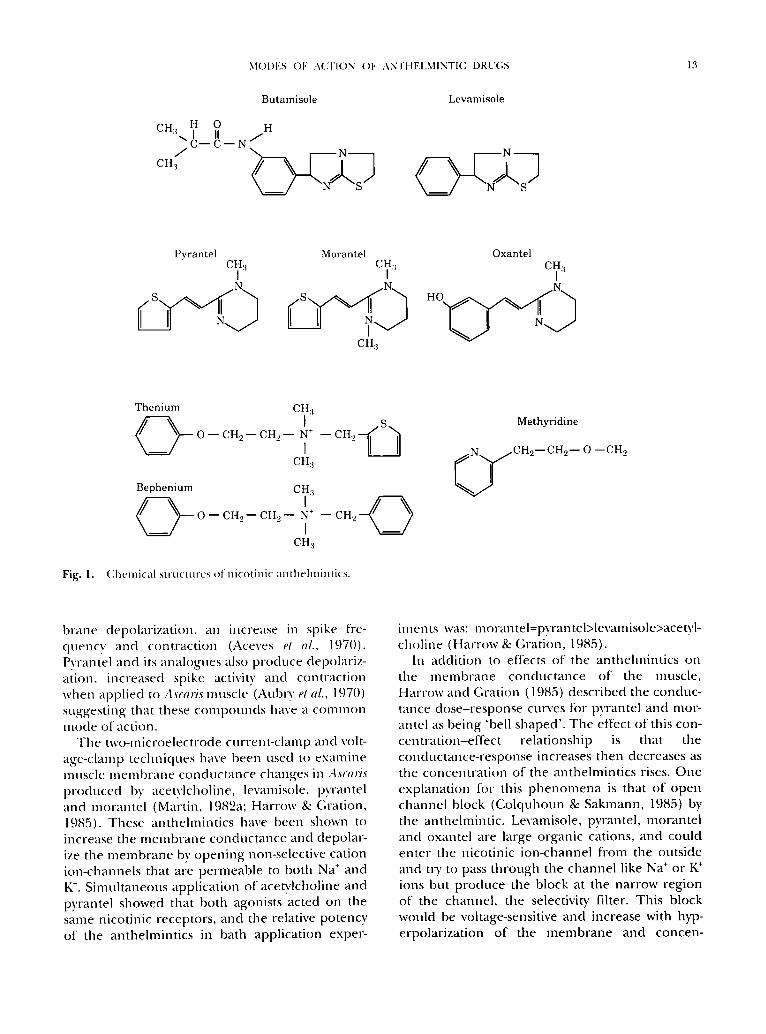
M()DES OF ACTION ()F ANTHELMINTIC DRU(;S 13
Butamisole
CH H 0 a / H N / H
CHa
S
Levamisole
N
Pyrantel Morantel Oxantel CHal CIita CI~:,
N N N
%) CHa
Fig. 1.
Thenium CH:I
0 - - CH 2 - - CH, 2 - - N + - - CH._,
I CHa
Bephenium CH a
C > ' C? O - - C H 2 - - C H 2 - - N + - - C H 2
1 CHa
Chmnica l s t ruc tures of n icot inic an thchn in t i cs .
Methyridine
~ / C H 2 - CH 2 - O - - CH 2
brahe del)olarization, an increase in spike fl'e- qnency and contraction (Aceves el al., 1970). Pvrantel and its analogues also produce depolariz- ation, increased spike actMtv and contl-action when applied to Ascaris muscle (Aubr)' el al., 1970) suggesting that these compounds have a common mode of action.
The two-microelectrode currelat-clamp and voh- age-clamp teclmiques have been used to exalnine m t t s c l e n l e m b r a n e conductance changes in Ascaris produced by acetylcholine, levamisole, pyrantel and morantel (Martin, 1982a; Harrow & Gration, 1985). These anthehnintics have I)een shown to increase the meml)rane conductance and depolar- ize the membrane by opening non-selective cation ion-channels that are permeable to both Na + and K +. Simultaneous application of acetylcholine and pyrantel showed that both agonists acted on the same nicotinic receptors, and the relative potency of the anthehnintics in bath application exper-
inlents was: morantel=pyrantel>levamisole>acetyl- choline (Harrow & Gration, 1985).
In addition to effects of tile anthehnintics on the membrane conductance of the muscle, Harrow and Gration (1985) described the conduc- tance dose-response curves for pyrantel and mor- antel as being 'bell shaped'. The effect of this con- centration-effect relationship is that the conductance-response increases then decreases as the concentration of the anthelmintics rises. One explanation tor this phenomena is that of open channel block (Colquhoun & Sakmann, 1985) by the anthelmintic. Levamisole, pyrantel, morantel and oxantel are large organic cations, and could enter the nicotinic ion-channel from the outside and tl-y to pass through the channel like Na + or K + ions but produce the block at the narrow region of the channel, the selectivity filter. This block would be voltage-sensitive and increase with hyp- erpolarization of the membrane and concen-

14 THE VETERINARY .JOURNAL 154, 1
(a)
eao ~ ' r y Arm Syncytium
s cord
I J 200 I.tm
(c) ~ t ~ ~ ~ Dorsal nelve cord
~ Syncylium
1 mm Ventral nerve cord
(b) Poster ior Anterior
cor
DE3
Left ./ commissure
Dorsal T ~ -YY Y V Y Y Y nerve .%
J. ~ k v .t ,k 2~ ,k
v . , o w v v v Y Y Y Y Y Y Y Y Y Y Y Y Y Y Y
7 T
Y g Y g 7
7 V T
Ventral nerve cord
Fig. 2. Diagrams of tile n e u r o n m s c u l a r o rgan iza t ion of n e m a t o d e somat ic muscle. (a) Diagram of a s(mmlic muscle cell showing: the cont rac t i le sp indle region; tile ba l loon-shaped bag that con ta ins tile nuc leus and glycogen granules ; tile a rm which is a process of tile muscle and reaches to one of tile nerve cords where i, divides into t]ngers Ihal I()lnl all electr ical ly-coupled com pl ex with the f ingers of ad jacent muscle cells and collectiveh' is known as the svncvtitml. T h e muscle cell possesses synaptic n icot in ic acetylchol ine receptors , and synaptic GABA recep tors at tile svncvtial region and extrasynapt ic ace ty lchol ine and G,MIA receptors over tile surface of the muscle. (b) Diagram of the dorsal and ventra l nerve cord showing the cell r ep r e s en t ed in a segment . All tile cell bodies of tile m o t o r ne t t rones are c o n t a i n e d in tile ventral nerve cord. Each segmen t has th ree r igh t -hand commissures and o n e lef t -hand commissure . Each commissu re is made up of o n e or two axons or dend r i t e s of tile m o t o r n e u r o n e s tha t pass r o u n d the body to c o n n e c t tile ventra l and dorsal nerve cord. With in each segmen t there are 11 m o t o r nero-ones tha t are divided in to seven ana tomica l types (DEI, DE2, DE3: dorsal excitato~3.' m o t o r neu rones . DI: dorsal inh ib i to ry m o t o r neu rones . VI: ventral inh ib i to ry m o t o r n e u r o n e . VI & V2: ventral excitatocv n to to r neu rones . T h e r e m a i n i n g axons in the ventral nelwe cord are in t e r segmen ta l neu rones . Tile exci tatory m o t o r n e u r o n e s are cho l inerg ic and tile inh ib i tory m o t o r n e u r o n e s are GABAergic. (c) Diagrmn of a cross sect ion of the body jus t caudal to the phau , ngeal muscle. It shows the relative locat ions of tile nerve cords, lateral lines, gut and muscle cells.
tration of the anthelmintic. This property of pro- ducing open channel block has been established using single-channel recording techniques (Robertson & Martin, 1993).
Single-channel currents activated by nicotinic anthelmintics
Initially, levamisole-activated channel currents [Fig. 3(a)] were recorded fi'om the muscle vesicle
preparation (Robertson & Martin, 1993) using the patch-clamp technique. The ion-chalmel currents activated by low levamisole concentrations were shown to cart 3 , cations and to have similar kinetics to acetylcholine-activated channels. The levami- sole activated channels have a mean open time of 1.34 ms and a conductance in the range 20-45 pS. At higher concentrations of levamisole, a flicker- ing open channel block that increases on hyper-

MODES OF ACTION OF ANTHELMINTI(" DRUGS 15
polarization is obsel-ved [Fig. 3(b)] . At -50 mV the dissociation constant for the channel block was 123 ~,xl. In addit ion to the flickering block, clusters of openings were also observed at high levamisole concent ra t ions where openings were separated by long (seconds) closed times: this chlstering behaviour is similar to that asstuned to be desensitization in vertebrates but was more voltage-sensitive and more p rominen t at hyperpol- arized potentials in Ascaris.
Pyrantel-activated channels [Fig. 3(c)] have also been r eco rded using the same preparat ion (Robertson el al., 1992). Pvrantel-activated chan- nel showed at least two distinguishable conduc- tance levels: the main condnc tance level was near 40 pS and the smaller conduc tance level was near '22 pS, suggesting the presence of more than one type of nicotinic recep tor in the membrane . Again, the channels o p e n e d by pyrantel were shown to be pernmable to monovalent cations and channel-block occur red at hyperpolar ized poten- tials. The channel block occur red more readily with pyrantel than with levamisole as shown by the tact that the dissociation constant , /x]~, for levami- sole was laigher than that of pyrantel (levamisole ~ at -50 ntV 123 ~txl; pyrantel K]~ at -50 naV 37 btxl).
Tal)le II summarizes the channel I)locking proper t ies of levamisole and pyrantel and, in addit ion, morante l and oxantel . Some general points may be made by examining Table II. It can be seen that all the nicotinic anthehnint ics pro- duce open channel-block, a form of self antagon- isnt, and that levamisole is the least po ten t at blocking its own channel . The signiticance of the open channel block ntay relate to the ability of some nematodes to resist the effects of nicotinic anthehninit ics: channel block may occur at such low anthelmint ic concent ra t ions that normal con- centra t ions of anthelmint ic are ineffective.
In Ascmis, the channel-blocking ability of pyrantel is greater than levamisole, but the greatest channel-block is p roduced by morante l
with oxantel, occupying an in termedia te position between morante l and oxantel . Th e least po ten t agonist in Ascaris is oxantel which produces the lowest probabil i t T of channel open ing even at high concent ra t ions (Dale & Martin, 1995). hater- estingly, oxantel is not effective therapeutical ly against ascariasis but is used instead to treat 7)'ichuris infections. Th e efficacy of oxantel against 7)ichuds but not against AscaHs may be due to dif- ferences in the nicotinic receptors of the two species of nematode: oxantel may be effective and p roduce open ing of the TtJchuris nicotinic acetyl- chol ine recep to r but not so effective on the Ascaris receptor . Th e concent ra t ions of oxantel and pyrantel along the intestine may also be a factor affecting efficacies of the drugs.
Non-nicotinic or 'muscarinic ' choliner~c receptor~ in nematodes and resistance to anthelmintics
In a n u m b e r of ver tebrate preparat ions, acetyl- choline is able to alter the probabili ty of" open ing of vol tage-dependent channels by acting via G-pro- tein-coupled muscarinic receptors. An action in nematodes at acetylcholine receptors analogous to ver tebrate muscarinic receptors , modula t ing voltage-sensitive ion channels has yet to be fully repor ted .
Th e re is biochemical exidence for the presence of 'muscarinic ' receptors in Ascaris muscle: (1) Donahue et aL (1982) have obsevved that
effects of chol inergic stimulation inchtdes increases in levels of cyclic-AMP;
(2) Arevalo and Saz (1992) have observed that acetylcholine increases levels of phosphoD, l- choline, 1,2-diacylglycerides and phosphat id ic acid, and demons t ra t ed the presence of phos- pholipase C activit T. D o n ah u e et al. (1982) and Arevalo and Saz (1992) did not dis- tinguish between 'muscarinic ' or 'nicot inic ' chol inergic receptors .
Previous studies on nematodes have shown that resistance to nicotinic anthelmint ics may take two forms: (1) Th e selection of genetically resistant
Table II Voltage sensitivity o f dissociation constants o f o p e n channel block by the nicotinic anthelmintics
Membrane I¢33 l.evamisole 2KI, l5,rantel Kl~ Morantel KR Oxantel
-50 mV 123 I.tXJ 37 IJ.Xl 12 p.xt 18.5 I.tM -75 mV 46 IJM 20 I.t~l 1 ~,xl 7.5 law Voltage-sensitivlty of h]~ e-fold every 20 mV e-fold eve D' 40 mV e-fbld every 22 mV e-fold ever), 29 mV
/~ is the dissociation constant: the lower the concentration of this constant the more potent it is. At the concentration of the KB the channel is blocked for 50% of its open time so tile response would be reduced by a half.
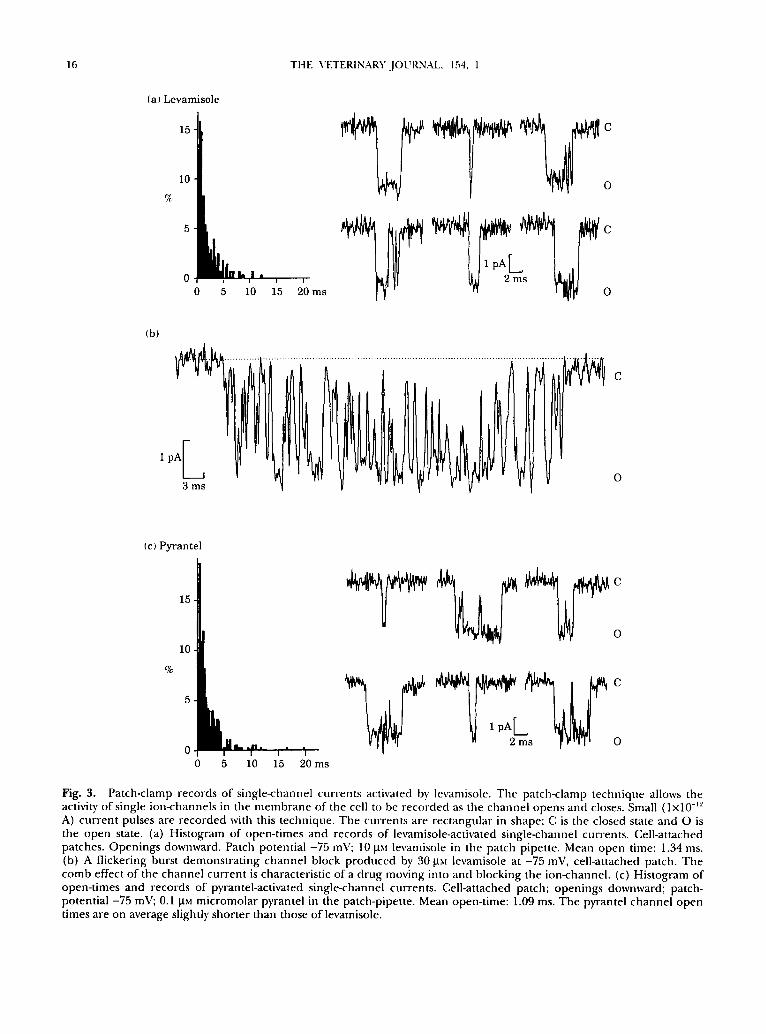
16 THE VETERINARY JOURNAL. 154, 1
(a } Levamisole
15
% 10
5
0 5 10 15 20 ms
0
1 pA L
0
(b~
. . . . . . . . . . . . i ............................................ i ...... C 1 PA3~ms ~ ~ 0
(c) Pyrantel
%
15-
10-
0- 0 5 10 15 20 ms
C
O
C
O
Fig. 3. Patch-clamp records of silagle-channel currents activated by levamisole. The patch-clamp technique allows the activit 3' of single ion-channels in the membrane of the cell to be recorded as the channel opens and closes. Small ( l x l0 -v-' A) current pulses are recorded with this technique. The currents are rectangular in shape; C is the closed state and O is the open state. (a) Histogram of open-times and records of levamisole-activated single-channel currents. Cell-attached patches. Openings downward. Patch potential -75 mV; 10 laM levamisole in the patch pipette. Mean open time: 1.34 ms. (b) A flickering burst demonstrating channel block produced by 30 last levamisole at -75 mV, cell-attached patch. The comb effect of the channel current is characteristic of a drug moving into and blocking the ion-channel. (c) Histogram of open-times and records of pyrantel-activated single-channel currents. Cell-attached patch; openings downward; patch- potential -75 mV; 0.1 I.tM micromolar pyrantel in the patch-pipette. Mean open-time: 1.09 ms. The pyrantel channel open times are on average slightly shorter than those of levamisole.

MODES OF ACTION OF ANTHELMINTIC DRUGS 17
mutants with modified nicotinic acetylcholine receptors on muscle (Lewis el al., 1980; Lewis et al., 1992). (2) Accomnaodation and recovery of parasites after long periods of exposure to the anthelmintics (Lewis et al., 1980; Lewis et al., 1992), and which may be explained by nicotinic receptor desensitization.
These resistant or recovered nematodes no longer respond to the nicotinic anthehnintics, but interestingly, may still respond to acetylcholine (Coles et al., 1975). One explanation for these resuhs is that in addition to tile nicotinic receptor on nematode muscle, there are other non-nic- otinic cholinergic receptors present on muscle not stimulated by the nicotinic anthelmintics. Such receptors may facilitate contraction by being coupled via G-proteins to vohage-activated channels.
Dichlo,wos
o H CH30 \ l i I C]
/ p - - o - - c ~ - c / CH30 C1
Haloxon
0
C1CH2CH2 0 /
CH3
Fig. 4. ( :heroical s t ruc ture o fd i ch lo rvos and ha loxon .
Genetics of resislance to nicotinic anlhelmintics Tile genetics of resistance to the anthelmintic
levamisole has been studied in the laboratory using the small fi'ee-living soil nematode, Caenor- habdili.~ eh,gans. Several hundred levamisole-resist- ant alleles have been identified that were isolated ill several screens (Lewis el al., 1980; Lewis e/ al., 1992). The genes responsible tbr this resistance inclucled: h,v-1, unc-29 and unc-38 that encode tile proteins subunits that make up the nicotinic ion channels of the nematode. In addition, other genes including: un, c-50, unc-63 and unc-74 that are believed to be involved in receptor biosynth- esis have been identified.
Only two strains of lev-1 (×21 and x61) were dominant when crossed over with wild-types (non- resistant) (Fleming el al., 1994). The lev-1 (x21) strain only inw~lves mutation at a single amino- acid, replacing glutanaic acid in the ion pore of the receptor ion channel (ill alpha-7 at position 237) with a positively-charged lysine. This change is believed to be sufficient to change the ion chan- nel from cationic to anionic, i.e., to convert tile receptor from an excitatory channel to all inhibi- tory one. The ~,-1 (x61) swain contains tile amino-acid leucine ill the pore of tile ion-channel (in alpha-7 at position 247): this point mutation is expected to produce increased desensitization and reduced affinity for levamisole, rendering lev- anfisole less potent as an agonist.
The genetics of levamisole resistance in para- sitic nematodes remains to be studied.
O R G A N O P H O S P O R U S C H O L I N E S T E R A S E I N H I B I T O R S
Dichlmvos, haloxon Compounds like dichlorvos and haloxon are
selective organophosphorus anti-cholinesterases (Fig. 4), and have an anthehnintic action, as well as all insecticidal, action. Dichlorvos and haloxon can control insect parasites, as well as hehninth parasites. The mode of action of these compounds is to block tile action of the parasite enzyme, ace- tylcholinesterase, leading to the excessive build up of tile neurotransmitter, acetylcholine. This mode of action also predisposes towards toxicity in the host animal where acetylcholinesterase enzymes are also present. Because more selective combined anthehnintic+insecticidal agents (avermectins and milbemycin) are available, the organophosphorus compounds are now used less frequently.
The existence of cholinesterase, the enzyme that breaks down acetylcholine, was first described in Ascmis by Bueding (1952). The distribution of cholinesterase was described by Lee (1962) who used histochemical techniques. In the head region of Ascmis, most of the enzyme activity is associated with the contractile spindle region of the muscle and is in the extracellular matrix. Lee (1962) also described cholinesterase activity on the muscle arms near their endings on the nerve cords but not on the bag region of the muscle.
In C. elegans, three classes of acetylcholinester- ase are recognized: class A, class B, and class C,

GABA , / ’ \
,‘A, COOH ‘\ NH;?
H
Piperazine
J 2 pA
100 ms
and are known to be products of three separate genes: owl, uw2 cind are-3 (Opperman & Chang, 1992). The three classes of acetylcholinesterase are sepal-able by their solubility in Triton, their temperature stability and sensitivity to cholinester- ase antagonists. In C. elegcr??.r, the major form is class B which is distributed in the head and body; classes A and C have a distribution biased towards the head. Extraction of the different classes for biochemical characterization has produced evi- dence that the enzyme may exist as a monomer, dimer and tetramer. Defects in the ace-1, m-e-2 mcl
ace-j genes suggest that the functions of the diff- erent classes are supplementary but that if there are defects in all of the genes, then elevated levels of acetylcholine occur in C. eleguns and there is motor incoordination (Opperman & Chang, 1992).
Although acetylcholinesterase is responsible for the breakdown of acehllcholine and involvecl in motor action in nematodes, it is also secretecl into the external environment in large quantities bv ‘4. .s717077 and other parasitic nematodes. The fun&on of llle secretecl aceh~lcholinesterase ma), be to reduce the effects of host acetylcholine in the intestine, perhaps clecreasing niucosal glanclulai secretion by the host. The original hypothesis of a biochemical ‘hold-fast’ effect (recluced acetylcho- line in the host intestine blocking peristalsis of the gut) has now been rqjectecl. The identification of the function of secreted cholinesterase hy para- sitic nematodes, ancl the abilitv to antagonize this enz\me, mav lead to the increased use of‘ anti- cholinesterases in the future to facilitate renio\xl of gut nematodes.
GABA AGONIST
Piperazine has a heteroq~lic ring structure (Fig. .5), ancl unlike y-amino-butyric acid (GABA), lacks a carboxvl group. Howe\*er. these t~vo con~pouncls act on th’e same receptor that is a ligancl-galecl Cl- channel found on the synaptic ancl extrasynaptic membrane of nematocle muscle (Martin, 1980; h4artin. 1982). Bath application of GABA or pip- erazine increases the opening of the muscle mem- brme <:I- channels, hvperpolarizes the membrane potential, increases tile membrane conductance and produces spastic paralysis. Piperazine appears to ha\pe a selective effect on large nematocles of the gastrointestinal tract, perhaps because the high CO, environment allows CO, to mimic the missing carboxy] group of piperzine. Piperazine appears less active against nematodes that occup! a high 0, environment.
Piperazine- and GABA-activated channel cur- rents (Fig. 5) ha\re been recorded using cell- attachecl and isolated outside-out patches (Martin, 1985). Although the channels may have more than one conductance level, the most frequently observed is 22 pS for both agonists. However, the mean open time of the channels for piperazine was shorter, 14 ms, than that produced by GABA, which produced channels that had mean open times of 32 ms. GABA is more than lo-100 times more potent than piperazine when conductance dose-response relationships are examined cluring bath application of the agonists (Martin, 1985).
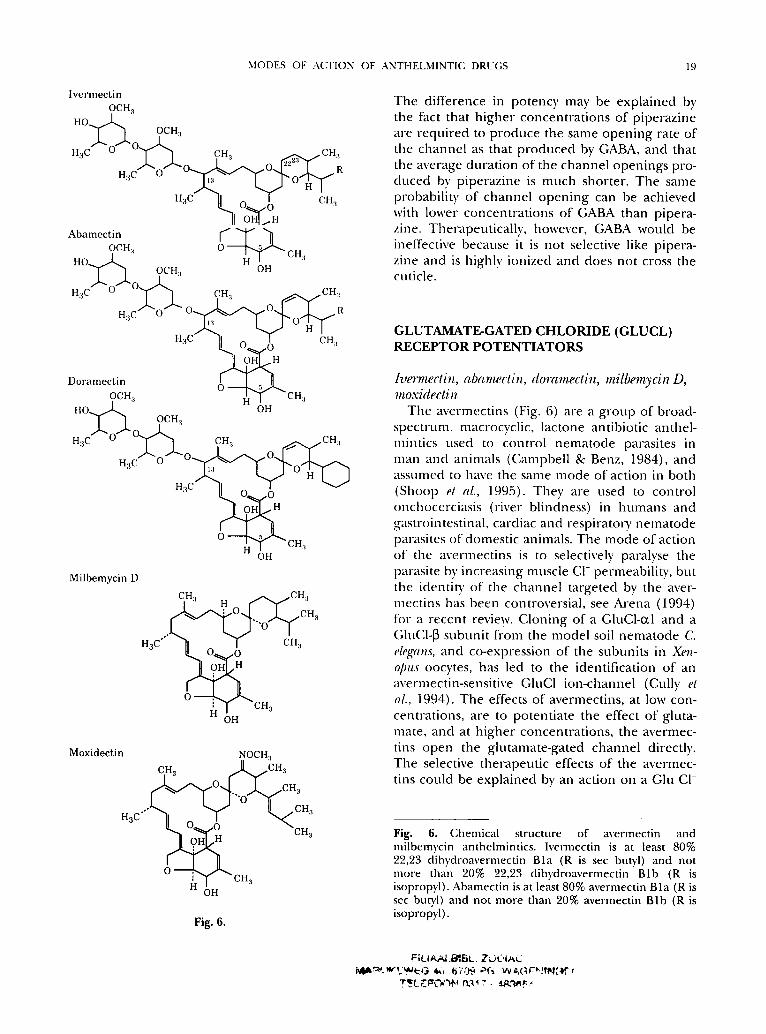
MODES OF ACTION OF ANTHELMINTI(" DRUGS 19
Ivermectin
H O ~ a ~OCH'~
H3 C O 0...1~ ~ CHa A .-CHa
IlL o_._A h-o j.
Abamectin ~" " ~ OCH.~ 0 ~ CH
HO_ 2-. hl : '
"~ OCH a OH
HaC/k" 0 ' '~ 0 ~ [ ' ~ " CH, CHa R
l! o._.'o
Doramectin ; ]__ I ~
HO.~ ~ . , . CHa OH
0 H I - . . ~ CH:, OH
Milbemyein D
CHa H A ~CHa
H3C" "]1 [ CHa
I OH
Moxidectin
c_Y 0 ~ C H a
Fig. 6.
The di f ference in potency may be explained by the fact that h igher concent ra t ions of piperazine are requi red to p roduce the same open ing rate of the channel as that p roduced by GABA, and that the average dura t ion of the channel openings pro- duced by piperazine is much shorter. Th e same probabili ty of channel open ing can be achieved with lower concent ra t ions of GABA than pipera- zine. Therapeut ical ly , however, GABA would be ineffective because it is not selective like pipera- zine and is highly ionized and does not cross the cuticle.
GLUTAMATE-GATED CHLORIDE (GLUCL) RECEPTOR POTENTIATORS
Ivermectin, abamectin, doramectin, milbemycin D, moxidectin
The avermectins (Fig. 6) are a g roup of broad- spectruna, macrocyclic, lactone antibiotic anthel- mintics used to control nema tode parasites in man and animals (Campbell & Benz, 1984), and assumed to have the same mode of action in both (Shoop et al., 1995). They are used to control onchocerciasis (river blindness) in humans and gastrointestinal, cardiac and respirato~-y nema tode parasites of domestic animals. The mode of action of the avermectins is to selectively paralyse the parasite by increasing muscle CI- permeabili ty, but the identity of the channel targeted by the aver- mectins has been controversial, see Arena (1994) for a recent review. Cloning of a GluCl-otl and a GIu('I-~3 subuni t fi'om the model soil n em a to d e C elegans, and co-expression of the subunits in Xen- opus oocytes, has led to the identification of an avermectin-sensitive GIuC1 ion-channel (Cully et al., 1994). Th e effects of avermectins, at low con- centrations, are to potent ia te the effect of gluta- mate, and at h igher concentra t ions , the avermec- tins open the glutanmte-gated channel directly. The selective therapeut ic effects of the avennec- tins could be expla ined by an action on a Glu CI-
Fig. 6. Chmnical structure of avermectin and milbenlycin anthelmintics. Ivermectin is at least 80% 22,23 dihydroavermectin Bla (R is sec butyl) and not more than 20% 22,23 dihydroavermectin Blb (R is isopropyl). Abamectin is at least 80% avermectin Bla (R is sec butTI) and not more than 20% avermectin Blb (R is isopropyl).
FI LI/,q.~.fiIIBL. Z~t)le, C
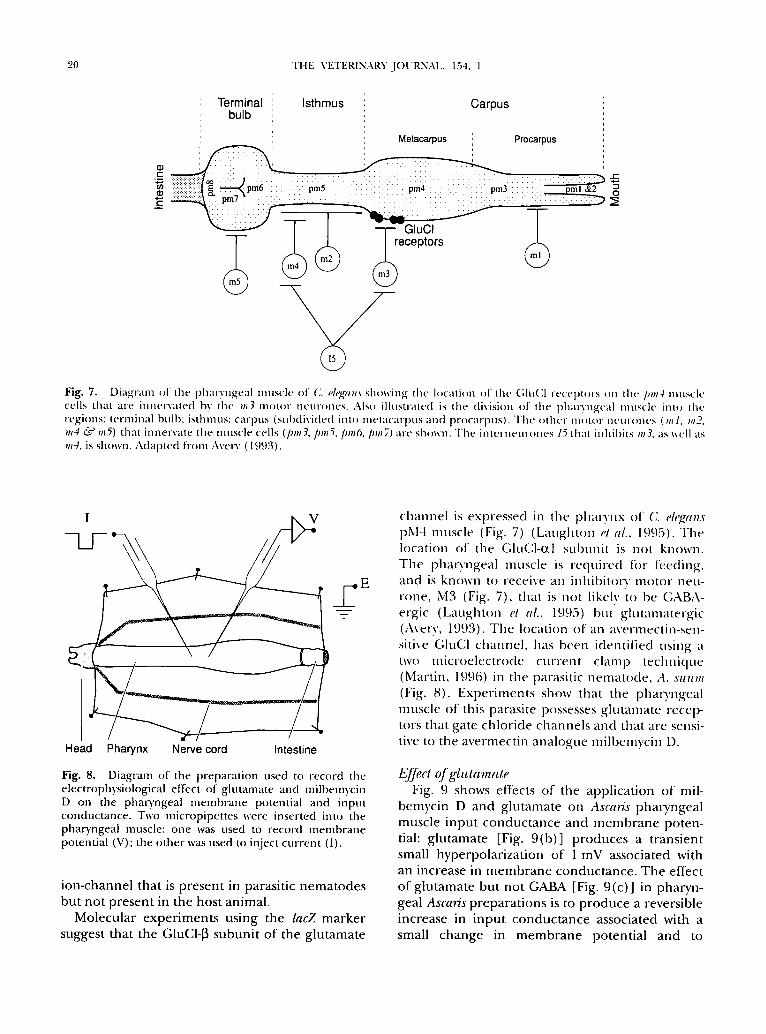
20 THE VETERINARY JOURNAL, 154, 1
- - - :::::::::::::::::::::::
i i::ii!iiiiiiii::i i i! E ~
Terminal i Isthmus : Carpus bulb :
Metacarpus : Procarpus
~ p m o ::.::::.: pm3 • :: pm4 • : pm3 :.::::.: ~ 2
Fig. 7. Diagram of the phalyngeal muscle of (J. eh'gan,~ showing tile location ~f the GIuC1 receptors ~m the pro4 muscle ceils that are innervated by the m3 motor neurones. Also illustrated is the division o1 the pha l )ngea l muscle into Ihe regions: lerminal bulb: isthmus: carpus (sul)divided into metacarpus and procarpus) . The o lhe l l l l ( l l ( } l " I l e l l l ' ( } l l e s (Ill l , m2, m4 ~" mS) that innervate the muscle cells (pro3, proS, pro6, pro7) are shown. The in te rneurones 15 thai inhibits m3, as well as m4, is shown. Adapted t iom Avery (I ~).t)~:~).
I V
Head Pharynx Nerve cord Intestine
z E
Fig. 8. Diagram of the preparat ion used to record the electrophysiological effect of glutamate and milbemycin D on the phaiTngeal m e m b r a n e potential and input conductance . Two micropipet tes were inserted into the pharyngeal muscle: one was used to record m e m b r a n e potential (V); the o ther was used to inject current (I).
ion-channel that is present in parasitic nematodes but not present in the host animal.
Molecular experiments using the lacZ marker suggest that the GluC1-]3 subunit of the glutamate
channel is exl)ressed ill tile I)halo'nx of C. e/egans pM4 muscle (Fig. 7) (Laughton el al., 1995). The location of the Glu(]l-0tl suhui-lit is n o t known. The phai)'ngeal muscle is required ti)r feeding and is known to receive an inhihitorv motor nett- tone, M3 (Fig. 7), that is not likely to be GABA- ergic (Laughton et al., 1995) but glutamatergic (Avel)', 1993). The location of an avermectin-sen- sitive (;luC1 channel, has heen identilied using a two microelectrode current clamp technique (Martin, 1996) in the parasitic nematode, A. Mtttm (Fig. 8). Experiments show that the pharyngeal muscle of this parasite possesses glutamate recep- tors that gate chloride channels and that are sensi- tive to the avermectin analogue milhemycin D.
Effect o f glutamate Fig. 9 shows effects of the application of mil-
bemycin D and glutamate oll Ascaris phal3,ngeal muscle input conductance and membrane poten- tial: glutamate [Fig. 9(b)] produces a transient small hyperpolarization of 1 mV associated with an increase in membrane conductance. The effect of glutamate but not GABA [Fig. 9(c)] in pharyn- geal AscaTis preparations is to produce a reversible increase in input conductance associated with a small change in membrane potential and to
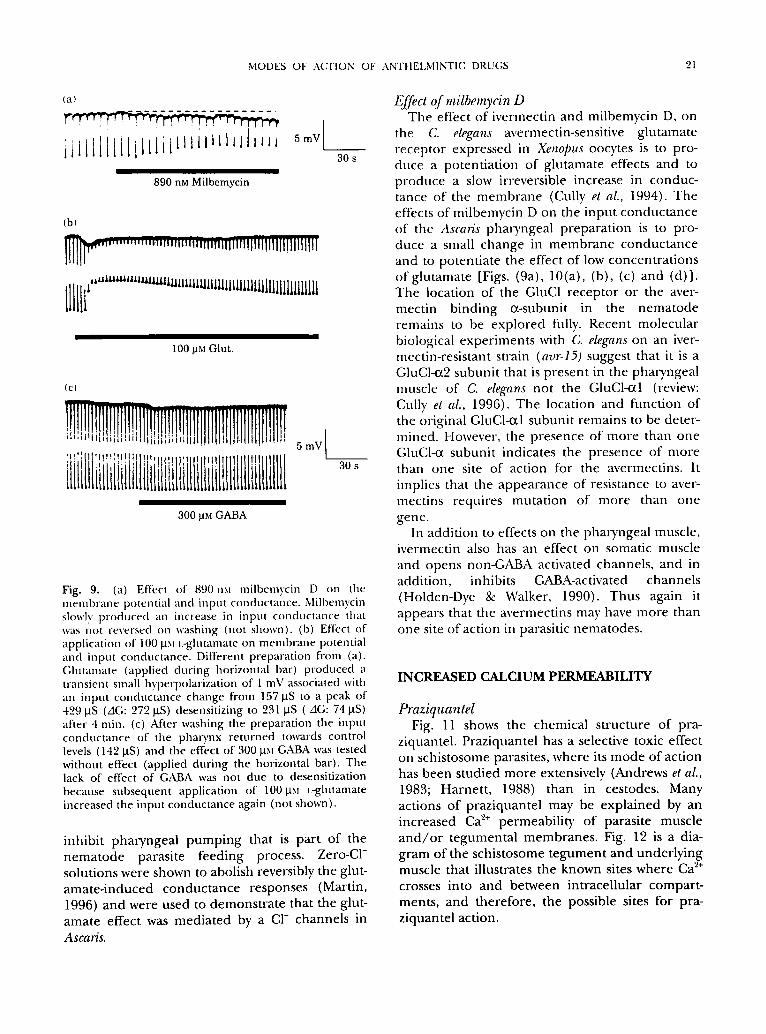
MODES OF ACTION OF ANTHELMINTIC DRUGS 21
(a)
r" - 7 - . . . . - - - . . . . . - -
iillI[llililliitlllll llll " 890 nM Milbemycin
5 mV _ _
30s
(b)
. . . . . :,,,ii',iiii,,.,?d.Tifflntitl'tl[lillllllllillilil
100 laM Glut.
(c)
llllllIHltli! ll Tiii, l ! ilt I,[!iI L!!I Nl!lI,E!ll! tilhliltttt tttlillhlll,Ll,li&L,ll[tl[,tlktdlt[
300 ~IM GABA
5mVl__ 30 s
Fig. 9. (a) Effect of 890rim milbemycin D on the memb,ane potential and input conductance. Milbemycin slowly p,oduccd an increase in input conductance that was not reversed on washing (not shown). (b) Effect of application of 100 ItM t.-glutamate on membrane potential and input conductance. Different preparation fi'om (a). Glutamate (applied during horizontal bar) produced a transient small hyperpolarization of 1 mV associated with an input conductance change from 157 ItS to a peak of 429 ItS (AG: 272 ItS) desensitizing to 231 gS ( AG: 74 ItS) after 4 rain. (c) After washing the preparation the input conductance of the pharynx returned towards control levels (142 ItS) and the effect of 300 gM GABA was tested without effect (applied during the horizontal bar). The lack of effect of GABA was not due to desensitization because subsequent application of 100~xt L-glutamate increased the input conductance again (not shown).
inhibi t pharyngea l p u m p i n g that is pa r t of the n e m a t o d e parasi te feeding process. Zero-C1- solut ions were shown to abolish reversibly the glut- ama te - induced c o n d u c t a n c e responses (Martin, 1996) and were used to d e m o n s t r a t e that the glut- ama te effect was m ed i a t ed by a CI- channels in
Ascam.
Effect of milbemycin D T h e effect o f ivermect in and mi lbemycin D, on
the C. elegans avermectin-sensit ive g lu tamate r ecep to r expressed in Xenopus oocytes is to pro- duce a poten t ia t ion of g lu tamate effects and to p roduce a slow irreversible increase in conduc- tance of the m e m b r a n e (Cully et al., 1994). T h e effects o f mi lbemycin D on the input conduc tance of the Ascaris pharyngea l p repa ra t ion is to pro- duce a small change in m e m b r a n e conduc t ance and to po ten t ia te the effect o f low concen t ra t ions of g lu tamate [Figs. (9a), 10(a), (b), (c) and (d)] . T h e locat ion of the GIuC1 recep to r or the aver- mect in b ind ing 0t-subunit in the n e m a t o d e remains to be exp lo red fully. Recent molecu la r biological expe r imen t s with C. elegans on an iver- mect in-resis tant strain (avr-15) suggest that it is a GluCl-0t2 subuni t that is p resen t in the pharyngea l muscle of C. elegans not the GluCl-otl (review: Cully el al., 1996). The location and funct ion of the original GluCl-cd subuni t r emains to be deter- mined. However, the p resence of m o r e than one GluCl-ot subuni t indicates the p resence of m o r e than one site of action for the avermect ins . It implies that the a p p e a r a n c e of resistance to aver- mect ins requires muta t ion of m o r e than one gene.
In addi t ion to effects on the pharyngea l muscle, ivermectin also has an effect on somatic muscle and opens non-GABA activated channels , and in addit ion, inhibits GABA-activated channels (Holden-Dye & Walker, 1990). Thus again it appears that the avermect ins may have m o r e than one site o f act ion in parasitic nematodes .
I N C R E A S E D C A L C I U M P E R M E A B I L I T Y
Praziquantel Fig. 11 shows the chemical su-ucture of pra-
ziquantel . Praziquantel has a selective toxic effect on schis tosome parasites, where its m o d e of act ion has been s tudied m o r e extensively (Andrews et al., 1983; Harne t t , 1988) than in cestodes. Many actions of praz iquante l may be exp la ined by an increased Ca ̀ '+ permeabi l i ty of parasi te muscle a n d / o r t egumen ta l m e m b r a n e s . Fig. 12 is a dia- g ram of the schis tosome t e g u m e n t and under ly ing muscle that illustrates the known sites where Ca ̀ '+ crosses into and be tween int racel lular compar t - ments , and therefore , the possible sites for pra- ziquantel action.
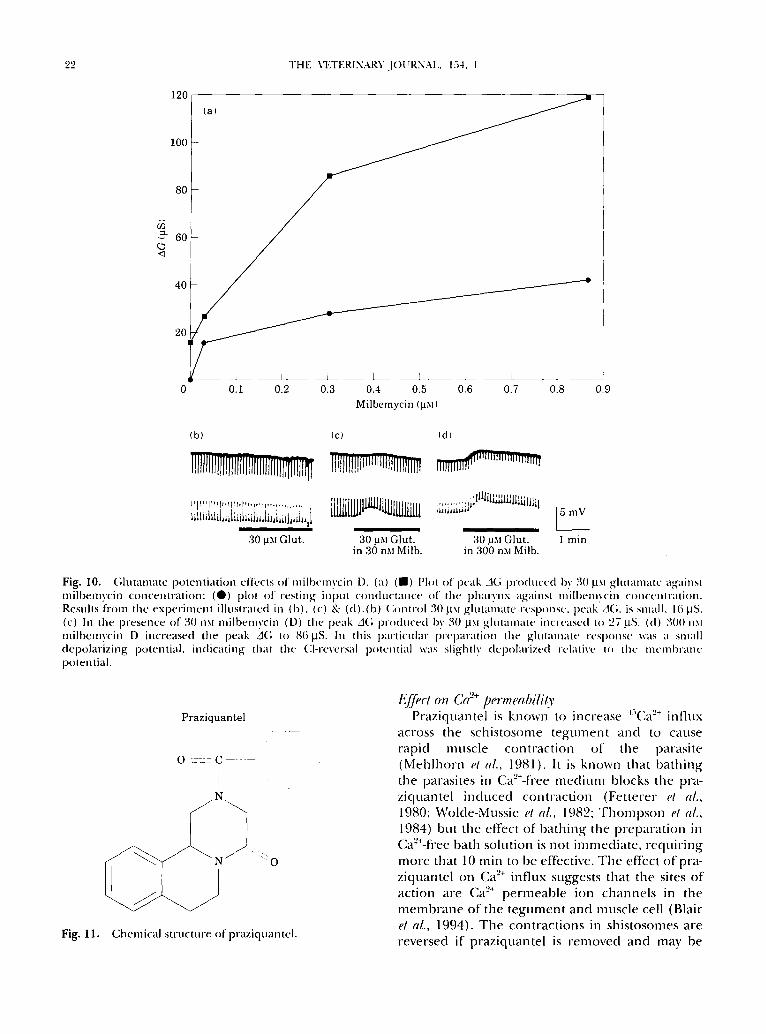
'22 ]'HE VETERINARVJOURNAI~, 154. 1
12° f 100 ~-
6 0 -
4O
2O
(a) /
/
I I I l ~ I 0.1 0.2 0.3 0.4 0 .5 0 .6 0 .7
Milbemycin (prq) 0.8 0.9
(b) (c) (d)
30 JaM Glut. 30 p.xl Glut. 30 p.~ Glut. in 30 nM Milb. in 300 nM Milb.
5 mV
1 r a i n
Fig. 10. (;luta,mlte potentiation effects ()f milbcmvcin D. (a) ( | ) Pl()l of peak A(; produced by. 30 p_~l ghmm)aw againsl milbemvcin co,lcentnuioq: (O) plot ()f resting input conductance ()f the phalynx agains! mill)cmvcin c() .ccn,al ion. Resuhs fiom the experiment illustrated in (b), (c) & (d).(b) ('ontrol 30 ~.txl glutamate vcsp()nsc, peak A(;, is small. 16 ~.tS. (c) In the pvescace of 30 .xt milbemvcin (D) the" peak A(; produced by 30 ~Xl gltmtmatc i.cvcased t() 27 l.tS. (d) 300 nxl milbemvcin D increased the peak A(; to 86 laS. I . this pavticttlar prel)aration the gltmlmatt" resp().se was a small depolarizing potential, indicating that the (:l-reversal p()te.tial was slightly depolarized relative, to the' mcmbra .c potential.
F i g . 11 .
P r a z i q u a n t e l
.N
£ \
Chemical structure of praziqtmn tel.
Effect o'n Ca "-'+ permeabi l i t y Praz iquan te l is k n o w n to increase ' 'Ca '-'+ inf lux
across the sch i s tosome t e g u m e n t and to cause rap id musc le c o n t r a c t i o n o f the paras i te ( M e h l h o r n el al., 1981). It is k n o w n that b a t h i n g the parasi tes in Ca'-'+-fi'ee mediuna blocks the pra- z iquante l i n d u c e d c o n t r a c t i o n (Fet tever el al., 1980; Wolde-Muss ie et al., 1982; T h o m p s o n el al., 1984) bu t the effect o f b a t h i n g the p r e p a r a t i o n in Ca"+-fi'ee ba th so lu t ion is no t immed ia t e , r e q u i r i n g m o r e that 10 rain to be effective. T h e effect o f pra- z iquante l on Ca ̀ -'+ inf lux suggests tha t the sites o f ac t ion are Ca '-'+ p e r m e a b l e ion c h a n n e l s in the m e m b r a n e o f the t e g u m e n t a n d musc le cell (Blair et aL, 1994). T h e c o n t r a c t i o n s in sh i s tosomes are reversed if p r az iquan t e l is r e m o v e d and may be

MODES OF ACTION OF ANTHELMINTIC DRUGS 23
D o u b l e H o s t ( B l o o d ) m e m b r a n e - 2+ C a 2+ ua
Ca 2+ 5 \ 1 ,-, I , - , "~ Ca2+ - 2+ - Ca2+ - • z, Ida 4 b
I ' V . , . • " . " " - " • • V ' " N a + • _ " . . 7 " • • • . . • '
Fig. 12. Diagram of 1he s t ruc ture of the body wall of Schi.~losoma ma,soni showing the possible sites of act ion of p raz iquante l and meclaanisms of Ca'-" t ranspor t into and out o t the t egument . 1: Vohage-act ivated Ca +-'+ channe l . 2: II'Jll'~lct'lhll~tl" i]lessell~el activated Ca +-'" chanl le l . 3: Ext race lhdar r ecep to r o p e r a t e d Ca ~" channe l . 4: Non-selective cat ion c h a n n e l also al lowing t+ntz-v of Ca'-". 5 : N a ' / C a e+ exchanger . 6: Ca ̀ '+ ATPase pt, m p i n g out ( ' J* . 7: In t r a t eg tmlen ta l Ca e+ buffers. 8: IP:~ re leasable store f rom the sarcol)lasmic rc t icuhtm. 9: Ca*-'" i nduced Ca'-" release c h a n n e l (CICR c h a n n e l ) . I0: (;~(-'" ATPase p t tmp: savcoplasmic endop lasmic re t ic tdum Ca'-'- (SERCA). 11: Electrical j t m c t i o n be tween mttscle cell and tegumet l t . 12: Electrical j u n c t i o n s be tween tnuscle cells. If praziqt tantel acts like caffeine it would act on site 9. 13: A p r o t o n I)tttnp may set tq) a pH g rad ien t actress the t egumen t and be r equ i red for qormal t i tnct ion. A p r o t o n i o n o p h o r e wc+uld ttl)Set tiffs g rad ien t and lead to the demise c~t + the organism.
blocked by Mg '-'+, or La :~+ but not I)y Ni '-'+ Co e+ 01- the calcium-channel blocker D-600 (Fetterer et al., 1980; Wolde-Mussie et al., 1982). The application of a high K + solution to depolarize the schisto- some preparat ion results in a rapid depolariz- ation, but the application of praziquantel resultsin a slower onset depolarization. From these observa- tions, it may be concluded that praziquantel some- how results in an increase in Ca ̀-'+ permeability across parasite membranes via channels that are not activated by depolarization. These channels must be pharmacologically different fi'om those of the host animal or the praziquantel would affect the host animal.
Experiments on a snail smooth muscle prep- aration (Gardner & Brezden, 1984) have illus- trated that caffeine mimics the effect of pra- ziquantel in producing contraction, and that prior t rea tment of the muscle preparation with caffeine
will eliminate the effect of praziquantel on the muscle preparat ion in Ca'-'+-free solutions. These experiments suggest that caffeine and praziquan- tel share the same site of action.
In vertebrate smooth muscle preparations, caf- feine acts to stimulate the opening of a large cat- ion channel known as the CICR channel (calcium- induced calcium release channel) (Herrmann- Frank et al., 1991) in the sarcoplasmic reticulum. This channel is activated by cytosolic rises in Ca '-'+, adenosine t r iphosphate (ATP) and caffeine but is inhibited by Mg '-'+. The CICR channel p roduces regenerative Ca ̀-'+ release from the sarcoplasmic reticulum following entry of Ca ̀-'+ through voltage- activated channels (Gregoire et al., 1989). Large cation channels, which might be similar to the CICR channel , have been observed in the tegu- mental membrane of schistosomes (Day et al., 1992).

24 THE VETERINARY .]OURNAL. 154, I
Parasite antigen exposure It has also been found that curative doses of
praziquantel in infected animals resuhs in antigens of schistosomes being exposed and binding and penetration of host antigen cells to the parasite after 17h treatment (Harnett & Knsel, 1986; Brindley & Sher, 1987). The immune system of the host, therefore, plays an important role in the praziquantel-induced death of the parasite. One explanation for the exposure of the parasite antigen may be that increased cvtosolic Ca ̀-'+ causes phospholipase C activation and then activation of protein kinase C (Berridge & h-vine, 1984; Sigiya & Furuyanaa, 1990). The phosphoDq- ation of proteins by protein kinase C could then destabilize the tegument causing the observed vacuolation and antigen exposure (Wiest el al., 1992). Once again the mode of action of pra- ziquantel relates to an effect on Ca ̀-'+ permeability of the tegumental nmmhranes. Praziquantel is used fi'equently to treat tapeworm infestations but less is known of the mode of action in this group of parasites. It is presumed that there is a common mode of action against trematodes and cestodes.
INHIBITION OF MICROTUBULE FORMATION, ~-TUBULIN BINDING
Benzimidazoles: thiabendazole, cambendazole, mebendazole, fenbendazole, oxibe~Mazole, oxfendazole, albendazole, albendazo& sulphoxide, parbendazole, flubendazole, t~4clabendazole & pro- drugs: netobimin, febantel, thiophanate
Fig. 13 illustrates the chemical structures of the benzimidazole anthehnintics and prodrugs. Pro- drugs are converted to active benzimidazoles by metabolic processes in the host animal so that it is the active metabolites that are responsible for the anthelmintic action. Triclahendazole, the flukici- dal compound, is assumed to act hy the same mechanism as the other benzimidazoles but a clear explanation for its selective effect against fluke is not known (McKellar & Kinabo, 1991) but may be related to its high level of plasma protein binding.
fl-tulmlin binding Mebendazole (Borgers et al., 1975; Van den
Bossche & De Nollin, 1973) and flubendazole (Van den Bossche & De Nollin, 1973) induce the loss of cytoplasmic microtubules of the tegu- mental and intestinal cells of cestodes and nema- todes, and this is followed by loss of transport of
secreta D, vesicles, a decreased glucose uptake and an increased utilization of stored glycogen. In Ascaris, mebendazole is taken up I)y phaiTngeal and intestinal cells where it is tound in the cyto- plasnaic fi'action bound to proteins with molecular weights of about 50 and 100 kDa that represent mollonlel-s and dimers of tubulin. O t l r current understanding of the mode of action of the benzi- midazoles thus started when it was recognized that the effect of mebendazole on Ascads was to dis- rupt the microtubules of the intestinal cells pro- ducing an inability to take up ghtcose (Van den Bossche, 1972; Van den Bossche el al., 1982). It was found that benzimidazole anthelmintic com- peted with the binding site for [H :~] colchicine on 13-tuhulin and that potency of the I)enzimidazole antlmhnintics correlated with the dissociation con- stant for hinding to nematode I]-mbulin (Lacey, 1990; Sangster et al., 1985).
Micrombules are intracellular organelles that serve a variety of fimctions including movement of chromosomes during cell division; providing the structural skeleton to the cell; movement of intra- cellular particles including e n e r ~ naetaholites: and exocvtosis. They are lound in both animal, plant, fungi and some bacterial cells (SnTer, 1995).
Microtul3ules are composed of t w o 450 amino- acid proteins known as c~-tubulin and I]-tt!bulin. Fig. 14 is a diagrammatic representation of [3-tuhtt- lin. It is composed of three domains: domain 1 is 34 kDa in size; domain 2 is 19 kDa in size and the tail is 2 kDa in size. There are GTP binding sites present which bind to sites I, II, III, and IV on the I]-tubulila. The 13-tul)ulin is folded so that the GTP- hinding sites form a pocket.
Formation of microtubules The formation of mio'otubnles is a dynamic
process (Fig. 14.) Formation involves the polynmr- ization of tubulin at one end (the positive pole) and the depolymerization at tile other end (the negative pole). The microtubules that tbrm are made up of 13 tubulin molecule rings (6 0f tubulin+7 13-tubulin alternating with 7 0t-tubulin+6 [3-tubulin rings). Seventy-five to 85% of the mass of microtubules is composed of the tubtdin pro- teins but in addition, microtubule-associated pro- teins (MAPs) are present stabilizing the structure. A number of factors favour polymerization includ- ing GTP, Mg '-'÷, and an increase in temperature (to 37°C). A decrease in temperature (to 4°C), the presence of Ca ̀-'+ or cahnodulin will favour depoly-

MODES OF ACTION OF ANTHELMINTIC DRUGS 25
BENZIMIDAZOLES
l "I N
N i L
S
H
Thiabendazole: R = H - -
C a m b e n d a z o l e : R = (CHa)2CHOCONH--
P R O D R U G
NHCO2CH3 N ~ C
NHCO2CH3
NHCOCH2OCH 3
Febantel: converts to fenbendazole
BENZIMIDAZOLE CARBAMATES
R
[
i
.N .
N
NHCO2CH a
H
TRICLABENDAZOLE
CI CI
' ' I "~ SCH 3
CH 3 " N
Oxibendazole: R = CH~CHzCH20--
Albendazole: R = CHaCH2CH2S--
R = (/,, - - ~'~ Fenbendazole:
Oxfendazole: R = @ Mebendazole: R = @
S - -
O
S - - -
O El C m
Flubendazole: R = F
0 @, S m
H
Fig. 13. Chemical structure of benzinfid~oles, benzimidazole carbamates, prodrug febantel and the antifluke drug triclabendazole.
mer iza t ion . In teres t ingly , c~- a nd [3-tubulin will pol) ,merize in to a n u m b e r o f shapes (r ings and sheets) in add i t ion to m i c r o t u b u l e s when in vitro p r e p a r a t i o n s are made . T h e t b r m a t i o n o f micro- tubules may be inh ib i t ed by subs tances that b ind to the l ead ing e d g e ( the positive pole) o f po lymer- ization. This process o f inh ib i t ion is known as ' c a p p i n g ' , and co lch ic ine , vinblast ine, vincrist ine,
the mi tot ic inh ib i tors and benz imidazo le s can do tiffs by b i n d i n g to ~-tubulin molecules .
T h u s the m o d e o f ac t ion o f the b e n z i m i d a z o l e an the lmin t i c s is the selective b i n d i n g to n e m a t o d e ~-tubulin, a n d c o n s e q u e n t inh ib i t ion o f micro- tubu le f o r m a t i o n . T h e effects are, t he re fo re , s lower in onse t t han the an the lmin t i c s tha t act as n e u r o t r a n s m i t t e r agonists , a n d inc lude slow onse t
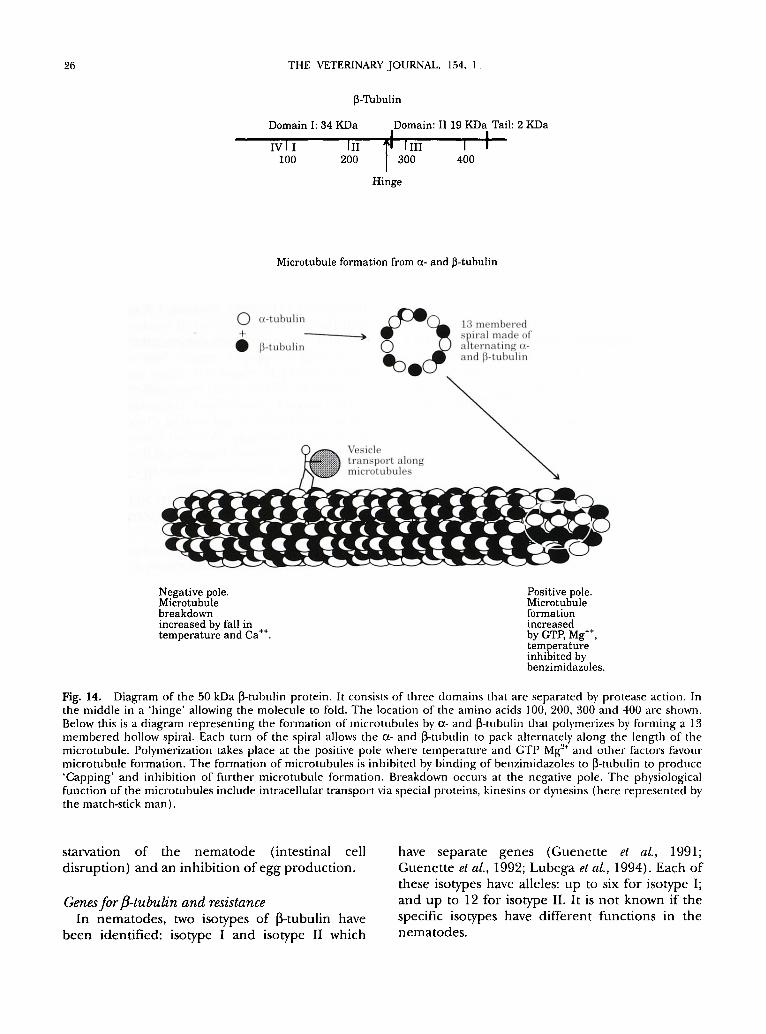
26 THE VETERINARY JOURNAL. 154, 1.
[3-Tubulin
Domain I: 34 KDa Domain: II 19 KDa Tail: 2 KDa
IH TI im i i 100 200 300 400
Hinge
Microtubule formation from cx- and ~-tubulin
Q) a-tubulin ~ ( ~ 13 membered + ~ ~ D ~ : spiral made of • ~-tubulin alternating ~-
and [3-tubulin
~ Vesicle transport along
Negative pole. Microtubule breakdown increased by fall in temperature and Ca ++ .
Positive pole. Microtubule formation increased by GTP, Mg +÷, temperature inhibited by benzimidazoles.
Fig. 14. Diagram of the 50 kDa ~-tubulin protein. It consists of three domains that are separated by protease action. In the middle in a 'hinge' allowing the molecule to fold. The location of the amino acids 100, 200, 300 and 400 are shown. Below this is a diagram representing the formation of microtubules by 0t- and ~-tubulin that polymerizes by forming a 13 membered hollow spiral. Each turn of the spiral allows the 0t- and ~-tubulin to pack ahernately along the length of the microtubule. Polymerization takes place at the positive pole where temperature and GTP Mg ''÷ and other factors favour microtubule formation. The formation of microtubules is inhibited by binding of benzimidazoles to ~-tubulin to produce 'Capping' and inhibition of further microtubule formation. Breakdown occurs at the negative pole. The physiological function of the microtubules include intracellular transport via special proteins, kinesins or dynesins (here represented by the match-stick man).
starvation of the nematode (intestinal cell disruption) and an inhibition of egg production.
Genes for fl-tubulin and resistance In nematodes, two isotypes of [3-tubulin have
been identified: isotype I and isotype II which
have separa te g e n e s ( G u e n e t t e et aL, 1991; G u e n e t t e et aL, 1992; L u b e g a et al., 1994). Each o f these isotypes have alleles: up to six for isotype I; a n d up to 12 fo r isotype II. I t is n o t k n o w n if the specific isotypes have d i f f e ren t f unc t i ons in the n e m a t o d e s .

MODES OF ACTION OF ANTHEI.MINTIC DRUGS 27
A reduct ion in the nunfl)er of isotype alleles fi)r ]3-mbulin is associated with the appearance of benzimidazole resistance (Roos e/ al., 1995). Resistance is associated with a progressive loss of alleles of isotype 1 and a total loss of isot?'pe 2 fi'om the ne ma tode populat ion. A resistant popu- lation of Haemonchus co~tlorlus has been charac- terized by only the presence of a single [3-tubulin allele re fe r red to as allele 200. The appearance of resistance can be expla ined by a loss of susceptible phenotypes and the st, rvival of a resistant pheno- type and its increased representa t ion in the remaining populat ion.
In fungi, benzimidazole resistance is associated with the appearance of a different form of [3-tubu- lin (Fujinmra el al., 1992; Jung el al., 1992) which is character ized by the appearance of tvrosine instead of phenyla lanine in position 200. Because mammal ian [3-tubulins have also tvrosine present in the 200 amino acid position (Lewis el al., 1985) it is unlikely that benzimidazole resistance may be overcome by changes in drug chemistvv. It would not be possible to design a selectively-toxic agent against the ['ungi because the fungi and host ~-tub- ulins would both bind the benzimidazole to the same degree and st) be loxic to both species. A similar i )he lmmenon may occur with nematode pm-asites.
P R O T O N I O N O P H O R E S : SALICYLANILIDES AND S U B S T I T U T E D PHENOLS.
Salic~,lanilides: closantel, raJbxanide, o.~yclozanide, bro/i'anide a~d substituted phenols: nitroxynil, niclopholan, hexachorophene dibromsalan & niclosa mide
The plaarmacolog, 3, of a range of anti-fluke drugs have been reviewed (McKellar & I<Anabo, 1991). In tiffs section, the mode of action of the p ro ton ionophores (often called oxidative phos- pholwlase uncouplers ) are considered. The chemi- cal s t ructure salicylanilides and nitroxynil illus- t,-ates that each anthelmint ic molecule possesses a de tachable p ro ton (Fig. 15). These molecules are vm T lipophilic and may shuttle protons across membranes , particularly the inner mi tochondr ia l membrane . Fig. 16 represents the process by which ATP produc t ion occurs in the mi tochond- ria, that is coup led to the p ro ton gradient across the inner mi tochondr ia l membrane . Oxidative p h o s p b o u l a t i o n may be summarized as electrons fi'om NADH or FADH being conveyed th rough a
series of prote in complexes on the inner mito- chondria l mmnbrane . This process leads to pro- tons being p u m p e d out of the mi tochondr ia matrix p roduc ing a pro ton motive force that is due to the pH gradient and t r ansmembrane elec- tric potential. ATP is synthesized when the pro- tons flow back into the mi tochondr ia l matrix through an enzyme complex. This process of oxi- dative phosphoD, lation takes place in the host ani- mal (StD, er, 1995), as well as in the parasitic hel- minths (Van den Bossche, 1972; McKellar & Kinabo, 1991).
Lipid soluble substances that can car D ' pro tons may shuttle across the inner m e m b r a n e of the mi tochondr ia , and remove the pro ton gradient and uncoup le oxidative phosphoryla t ion so that the oxidation of NADH and FADH is no longer l inked to the p roduc t ion of ATP and may carry, on without its product ion . Fig. 15 shows the position of the p ro ton that is able to dissociate froni a var- iety of compounds . 2,4-Dilfitrophenol, carbonyl-p- t r i f luoro-methoxyphenylhydrazone , (Stryer, 1995) and laexachloroplaene, nitroxynil, oxvclozanide (Corbet t & Goose, 1971; Edwards el al., 1981b) are all l ipophilic c o m p o u n d s that are capable of carr}.'ing pro tons across membranes , and there- fore, may act to uncoup le oxidative pbosphoD,1- ation prevent ing the p roduc t ion of the p ro ton gradient across the inner mi tochondr ia l membrane .
The nleclaallism of action of the pro ton ionoph- ores has been assnmed to be to selectively uncouple oxidative pbosphoD'lat ion in parasite mi tochondr ia . Pax and Bennet t (1989) have exper imenta l evidence that the inner m e m b r a n e of the parasite may not be the site of action of the pro ton iol lophores. Thev observed that appli- cation of closantel to Schistosoma mansoni and Fasci- ola hepatica produces a fall in tegumenta l pH (6.8-6.5) when measured with an intracellular e lect rode, and that this change in pH occur red after 10 rain and before an), change in the pro- duct ion of ATP bv the parasite. The intra-tegu- mental [',all in pH was associated with a reduct ion in motility of the parasite and could explain the anti-parasitic action of closantel. It was conc luded by Pax and Benne t t (1989) that closantel was a m e m b r a n e active molecule and that it may affect a n u m b e r of biochemical and physiological pro- cesses. These processes are presumably sensitive to changes in intra- tegumental pH. Thus the site o f action of the p ro ton i onophore s may include the t egument ra ther than just the mi tochondr ia .
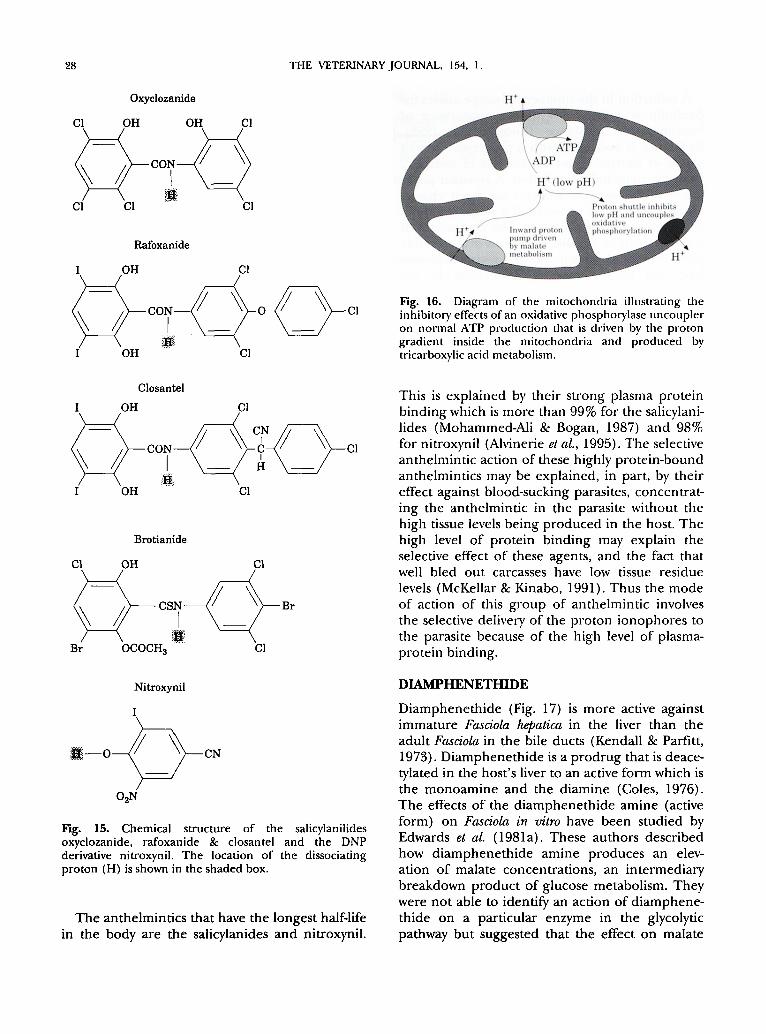
28 THE VETERINARY JOURNAL. 154, 1.
Oxyclozanide
C1 OH OH C1
CI CI CI
Rafoxanide
I OH C1
~ C O ! ~ ~ O ~ C l
I OH C1
Closantel
I OH C1
CON__// \/x_._ ?__< / \~---Cl
I OH CI
Brotianide
C1 OH CI
CS o< r Br OCOCH 3 CI
Nitroxynil
I
O2N
Fig. 15. Chemical structure of the salicylanilides oxyclozanide, rafoxanide & closantel and the DNP derivative nitroxynil. The location of the dissociating proton (H) is shown in the shaded box.
The anthelmintics that have the longest half-life in the body are the salicylanides and nitroxynil.
Fig. 16. Diagram of the mitochondria illustrating the inhibitory effects of an oxidative phosphorylase uncoupler on normal ATP production that is driven by the proton gradient inside the mitochondria and produced by tricarboxylic acid metabolism.
This is explained by their strong plasma protein binding which is more than 99% for the salicylani- lides (Mohammed-Ali & Bogan, 1987) and 98% for nitroxynil (Alvinerie et al., 1995). The selective anthehnintic action of these highly protein-bound anthelmintics may be explained, in part, by their effect against blood-sucking parasites, concentrat- ing the anthelmintic in the parasite without the high tissue levels being produced in the host. The high level of protein binding may explain the selective effect of these agents, and the fact that well bled out carcasses have low tissue residue levels (McKellar & Kinabo, 1991). Thus the mode of action of this group of anthelmintic involves the selective delivery of the proton ionophores to the parasite because of the high level of plasma- protein binding.
D I A M P H E N E T H I D E
Diamphenethide (Fig. 17) is more active against immature Fasciola hepatica in the liver than the adult Fasciola in the bile ducts (Kendall & Parfitt, 1973). Diamphenethide is a prodrug that is deace- tylated in the host's liver to an active form which is the monoamine and the diamine (Coles, 1976). The effects of the diamphenethide amine (active form) on Fasciola in vitro have been studied by Edwards et al. (1981a). These authors described how diamphenethide amine produces an elev- ation of malate concentrations, an intermediary breakdown product of glucose metabolism. They were not able to identify an action of diamphene- thide on a particular enzyme in the glycolytic pathway but suggested that the effect on malate

MODES OF ACTION OF ANTHELMINTI(" DRU(_;S 29
D i a m p h e n e t h i d e . / - - -
,,' / \
CH3CONH - - / . . . . 'i / 4 / ' :, OCHzCHzOCH2CHeO , /
.. /
N H C O C H 3
Clorsulon
C1
CI 2 = C
H2NO2S
NH 2 ~\ /
SONH 2
Fig. 17.
Diethylcarbamazine
C9H5 / \
CHaN NCON \ / .... ,
C2Hs
C h e m i c a l s t r u c t u r e o f d i a m p h e n e t h i d e , c l o r s u h m a n d d i e t h y l c a r b a m a z i n e .
was likely to be a prima D, effect because it occurred early on and before the deterioration of the whole parasite. Interestingly, in the second paper of Edwards et al. (1981h) the effect of dopa- mine, a putative neurotransmitter in Fasciola had a protective effect against diamphenethide.
The action of diamphenethide remains to be defined in greater detail but the study of Edwards el aL (1981a) showed that its biochenaical effects contrasted with the proton ionophore, oxyclozan- ide and appears to involve effects on malate metabolism in Fasciola.
INH IBITION OF PHOSPHOGLYCERATE KINASE AND MUTASE
Clorsulon Clorsulon is 4-amino-6 trichloro ethenyl 1,3-
benzenedisulphonamide (Fig. 17). Structurally, it is similar to 1,3-diphosphoglycerate (Schulman et al., 1982) and consequently, inhibits the enzymes
plmsphoglyce,'ate kinase (Schuhnan et al., 1982) and phosphoglyceromutase (Schulman & Valentino, 1982) of Fasciola and inhibits the Emden-Meyerhoff pathways in fluke. As a result, there is a selective inhibition of glucose utiliz- ation, and acetate and propionate formation. The inhibition of the Fasciola phosphoglycerate kinase is competitive (Fig. 18) with a ~ of 0.29 mM: clor- sulon competitively inhibiting the binding of ATP and 3-phosphoglycerate to the phosphoglycerate kinase. Schulman et al. (1982) suggested that the large group on the six position of clorsulon pre- vented the conformational change in the kinase enzyme required for activity. There was also a good correlation between the I~ values.for antag- onism of Fasciola phosphoglycerate kinase for a range of compounds related to clorsulon and the potency of these compounds as antifluke agents. This evidence supports the suggested mode of action.
The inhibition of Fasciola phosphoglyceromut- ase by clorsulon was studied by Schulman and

31) THE VETERINARY JOURNAl., 15-1, I
E1
E-ATP
/ \ -- I + E 3PG-E-ATP \ " /
/ , /
3PG-E
-- ADP-E-1,3DiPG "~-----~ E-3DiPG + ADP ~ - ~ - E + 1,3DiPG
Fig. 18. Diagram of the compet i t ive an tagon i sm of 19h¢lsl)hoglyctwate kiqase (El of l,'asciola hepatica by c lorsulon (1). DiphosphoiT1 glycerale: DiPG. 3-Plmsphol3l glyceratc: 3PC;.
Valentino (1982) who pointed out that this enzyme requires 2,3-diplaosphoglycerate for acti- vation and the ch)se similarity of clorsuh)n to diphosphoglycerates could explain its inhibition of the mutase enzwne. These authors puritied the enzyme and compared some of its properties with mamnaalian l~hosphoglyceromutase that were not inhibited by clorsuhm. There appears to be no recent studies on the mode of action of this series of compounds ahhough clinical trials and efficacy studies have been made.
does not appear to be involved because nucle, athvmic mice, infected with BruKia pahanffi, showed a marked reduction in microfilariae fol- lowing diethvlcarbamazinc treatment (Vickel T el al., 1986). Thus T cells and T-dependent ,'esponses (Ig(; and lgE) do not appear to be invoh'ed. The involvenlent of complement also seems unlikeh' because clearing of microfilaria ii-om the nuc(e mice by diethvlcarbanmzine was not inhibited by prio," treatment with cobra venom lactor treatment (Vicke D' el al.. 1986).
DIETHYLCARBAMAZINE
The cllelnical structure of diethylcarbamazine a piperazine derivative is shown in Fig. 17. Elect,-o- physiological experiments following bath appli- cation to A. suum of high concentrations of diethvlcarbamazine have shown that it does not mimic the action of piperazine (Martin, 1985).
Effects of diethylcarbamazine A major indication for diethylcarbamazine at
low doses is as an anti-filarial drug; it is a good microfilaricide (including microfilariae of the dog hearnvorm, Dirofilana immilis) but has limited macrofilaricidal effects. The limited action of diethylcarbamazine against some aduh hehninths (e.g. cattle lung3vorm, Dicl~,ocauhts) requires nearly 10 times the dose of the prophylactic anti- filarial dose, and may there£bre, invoh,e another mode of action. This review covers the mode of action of low closes of diethylcarbamazine; little is known of the mode of action of high doses of diethylcarbamazine.
Most studies suggest that diethylcarbamazine has no direct action on filarial parasites (Johnson el al., 1991). It appears to have no action at reasonable concentrations in vilro. In marked con- trast to in vitro experiments, it is known that in vivo diethylcarbamazine is effective within 4 rain following intravenous injection (Hawking & Laurie, 1949). The specific immune response
Fffects on arachidonic add metabolism may be the mode of attire1 of diellg, lcarbamazine
Diethylcarbanlazine has an antagonistic action of the metabolic enzymes that ,netabolize arachi- donic acid, tile products released as a resuh of phospholipase A.., action on cell membranes. Fig. 19 illustrates the sites of action of diethvlcarbama- zine proposed by Maizels and Denham (1992). Ahhough 5-1ipoxygenase is often being cited as being inhibited by dietlaylcarbamazine, tiffs may not be the case. In a mast cell line, conversion of 5-HPETE to leukotriene (LT),&~ by I.TA~ svnthet- ase was blocked by diethylcarbamazine (Mathews & Murphy, 1982). Razin el aL (1984) found that in mouse, 5-1ipoxygenase conversion of arachidonic acid to 5-HPETE was not inhibited but that LT(h and LTA_~ production was inhibited.
Diethylcarbanlazine also appears (Kanesa- Thasan el al., 1991) to inhibit endothelial pro- duction by cyclo-oxygenase of prostaglandin (PG)I._, (prostacyclin) and PGE., production but does not affect platelet production of thrombox- ane (TX)A._, by cyclo-oxygenase and the 12-1ipoxy- genase products 12- ancl 15-HPETE. Interestingly, microfilariae also produce their own PGI._, and PGE,, and like endothelial cell of blood vessels, diethylcarbamazine also inhibits the microfilarial production (Kanesa-Thasan el al., 1991). The co- administration of glucocorticoids, that are known to block phospholipase A,_, activity and the pro- duction of arachidonic acid (Fig. 19), together

M ( ) D E S O F :M71"ION O F A N T H E I . M I N T I C D R L ' ( ; S 3 1
r . . . . . . . . . . . . . . . . . . . . . . . . i i
i DIETHYLCARBAMAZINE i " - - r . . . . . . . . . . . . . . . . . . . . . .
i
LTA I SYNTHASE /
LTA 4 \ \
LTC 4 SYNTHASE
LTC.I eosinophil; mast cell
LTD4 eosinophil; mast cell
I
LTE 4 eosinophil; mast cell
MEMBRANE PHOSPHOLIPIDS
SE 1 ~- . . . . . . . . . . . . . . . . . . PLA 2 . . . . . 1 GLUCOCORTICOIDS 1
i . . . . . . . . . . . . . . . . . . . . . . . . . . .
ARACHIDONIC , DIETHYLCARBAMAZINE - 1 ACID . - " - " 1_ _ andNSAIDS
5-LIPOXYGENASE / J /
5-HPTE 5-HETE
/ /
12 and 15 HPTE/HETE
platelets; neutrophils; eosinopils.
\ \
LTB4 neutrol~hils; macropnages
~ _ , CYCLO-OXYGENASE
" " " " ' ~ PGGo and PGD~
PG-H+ '~.~"" . • mast-cells 12- and 15-
LIPOXYGENASE \ ~ , ~ PGE2
• macrophages; microfilaria \ \ .\
'\ PGI.,: PROSTACYCLIN \ vascular endothelium and \.. microfilaria
TXA2: THROMBOXAN platelets
Fig. 19. Diagram of the pr(~duction of the leukotrienes and prostaglandins via lipoxygenase or cyclo-oxygenase fi-om ar,tchidonic acid thal is produced from membrane phospholipids as a result ol phospholipase A=, acidity. The inhibitory silc.s of action of gluccwo,ticoids, non-steroidal anti-inllammatola' drugs (NSAIDs) and diethvlcarbamazine are shown. l)iethvlcmbamazine is shown inhil)iting the action of LTA, svnthase thal converts 5-hydr,>peroxyeicosatetraenoic acids (5- HPE'I:E) and 5-1avdroxveicosatet,aenoic acids (5-HETE) to the let,kotrienes I.TAa and I+TA~ to LTC; in mast cells and eosinophils. Dietl~vlcarbamazine is also shown inhibiting the cych>-oxygenase enzyme that converts a,achidonic acid to the prostaglm]dins P(~G._, and P(;H._,. The subsequent production of P(;D_,. PGE=,, PGI=, & TY~-k,_, is also inhibited in the blood vessels and ,nicrol]lari;~.
with d i e thy l ca rbam a z i ne , r e duc e s the effect o f d ie t lay lcarhamazine (Stingl el al., 1988). Thus , d i e t h y l c a r b m n a z i n e seems to inhibi t p r o d u c t i o n o f PGI=, and PGE=, hy microf i lar ia a nd endo the l i a l cells. T h e n o r m a l level o f p r o d u c t i o n o f PG1._, and PGE,_, tha t p r o d u c e s a tonic di la t ion o f b lood ves- sels a n d inhibi ts a g g r e g a t i o n o f neutroplai ls and eos inoph i l s is r e d u c e d by admin i s t r a t ion o f d i e thy l ca rhamaz ine .
A plausible e x p l a n a t i o n o f the m o d e o f ac t ion o f d i e t h y l c a r b a m a z i n e is tha t this c o m p o u n d alters the m e t a b o l i s m o f a r a c h i d o n i c acids in hos t e n d o - thelial cell a n d also in microf i la r iae tha t are sus- cep t ib le to the ac t ion o f d i e thy lca rbamaz ine . T h e r e is t h e n a vasocons t r ic t ion , ampl i f i ed e n d o - thelial a d h e s i o n and immobi l i za t i on o f the micro-
filariae and cyto toxic activig, by the hos t platelets and granulocy tes . It appea r s tha t d i e thy l ca rbama z - ine activates an inna te i m m u n e r e s p o n s e r a t h e r than an adapt ive i m m u n e r e s p o n s e (Maizels & D e n h a m , 1992). This m o d e o f ac t ion can exp la in why d i e t h y l c a r b a m a z i n e has no ac t ion in vitro agains t microf i la r iae and is effect ive in n o n - i m m u n e animals .
A C K N O W L E D G E M E N T S
I am p leased to a c k n o w l e d g e the f inancia l s u p p o r t o f the W e l l c o m e Trus t w h o have s u p p o r t e d my work on the e l ec t rophys io logy o f the p ipe raz ine ,

32 THE VETERINARY .JOURNAL, 154, 1
nicotinic an the lmint ics and the GIuCi receptors o f Asca,is.
REFERENCES
ACEVES, J., ERLUI, D. & MARTINEZ-MARNON, R. (1970). The mechanism of the paralysing action of tetramisole on Ascmqs somatic muscle. B~itish Journal of Pharmacology 38, 332-44.
Aa.VINEmE, M., Ft.oc'tl, R. & GAI.'rlER, P. (1995). Plasma protein binding of nitroxynil in several species Jour- nal of Veterinary Pha~vnacology and Therapeutics. 14, 170-3.
ANDREWS, P., TIIOSL~';, H., Pom.~, R. & SEUBERT, J. (1983). Praziquantel. Medical Research Reviews. 3, 147-200.
ARENA, J. P. (1994). Expression of caenorhabditis elegans messenger-RNA in Xeuopus oocytes: a model system to study the mechanism of action of avermectins. Pam- sitology Today 10, 35-7.
AREVa~.O, J. I. & SAZ, H.J . (1992). Effects of cholinergic agents on the metabolism of choline in muscle fi'om Ascan:s suum. Jou~al of Parasitology 78, 387-92.
ArteRY, M. L., COWELL, P., DAV~:v, M. J. & SmvD~:, S. (1970). Aspects of the pharmacology of new anthel- mintics: pyrantel. Btitish Journal of Pharmacology 38, 332-44.
AVERY, L. (1993). Motor-neuron m3 controls pharyngeal muscle-relaxation timing in Caenorhabditis elegans. Journal of Expe~4mental Biology 175, 283-97.
BERR~D~;E, M. J. & IRVlNE, R. F. (1984). Inositol tris- phosphate, a novel second messenger in cellular sig- nalling transduction. Nature312, 315.
BL~xm, I4. L., BENNett, J. u & E.xx, R. A. (1994). Schisto- soma mansoni: myogenic characteristics of phorbol ester induced muscle contraction. Expe~qmental Para- sitology 78, 302-16.
BoRc;Z~, M., DE NOLLIN, S., Dz BRABANDER, M. & THIENPO- ~NT, D. (1975). Influence of the anthelmintic meben- dazole in nematode intestinal cells. American Journal of Veterinary Research 36, 1153-66.
BmNDLEV, P.J. & SHER, A. (1987). The chemotherapeutic effect of praziquantel against Schistosoma mansoni is dependent on host antibody response. Journal of Immunology 136, 215-20.
BL~EmNG, E. (1952). Acetycholinesterase in S&istosoma mansoni. British Journal of Pharmacology 7, 563-6.
CAMPBELl., W. C. & BENZ, G. W. (1984). Ivermectin: a review of efficacy and safety. Journal of Veterinary Phar- macology and Therapeutics 7, 1-16.
COLES, G. C., EXST, J. M. & JENKINS, S. N. (1975). The mechanism of action of the anthelmintic levamisole. General Pharmacology. 6, 309-13.
CoLt.S, G. C. (1976). The inactivity of diamphenethide against liver fluke in certain species of mammals. Journal of Parasitology 20, 110-1.
COLQUHOUN, D. & SAKMANN, B. (1985). Fast events in single-channel currents activated by acetylcholine and its analogues at the frog muscle end-plate. Jour- nal of Physiology (London)369, 501-57.
COR~ETr, J. R. & GOOSE, J. (1971). The biochemical
mode of action of the tasciolicides, nitroxynil, hex- achlorophene, and oxyclozanide. Biochemical Journal 121, 41.
Cut.ix, D. F., VA~slt_.\ns, D. K., Lw, K. K., PAm-SS, P. S., V..XNDERPt.OEt;, L. H. T. & St:IIAEVFER, J. M. (1994). Clon- ing of an avermectin-sensitive glutamate-gated chlor- ide channel from Caenolhabditis elegans. Nature 371, 707-11.
CULl?,', D. F. WH.~INSON, H. VassHms, D.K. EI-II-R, A. & ARENA, J. (1996). Molecular Biology and Electrophysi- ology of Glutamate-gated Chloride Channels of Invertebrates. Parasitology 113, SXXX-SYYY.
DAI.e, V. M. E. & MARTLX, R.J. (1995). Oxantel-activated single-channel currents in the muscle membrane of Ascmis suum. Parasitology 110, 437-48.
DAL T. S., BENNE-r-r,J. L. & PAX, R. A. (1992). Shistosoma mansoni: patch-clamp study of a non-selective cation channel in the outer tegumental membrane of females. Experimental Parasitolo~, 74, 348-56.
DoYam'~:, M.J., Y.~.t:ot,u, N.J . &HAP, RIS, B. G. (1982). Correlation of muscle-activity with glycogen-metal> olism in muscle of AscaT~s suum. Ame)qcan Journal Of Physiology 242, R514-R21.
EDwam~s, S. R., CAMm~Et.L, A.J., SHVERS, M., MOORE, R.J. & MoNr.-xt;t'I-, P. E. (1981a). Studies of the effect of diamphenethide and oxyclozanide on the metal> olism of Fasciola hepatica. Molecular and Biochemical Parasitology 2, 323-38.
EmVARDS, S. R., CAMI'BeLL, A.J., SHEERS, M., MOORE, R.J. & MONTAC;t'E, P. E. (1981b). Protection of Fasciola hep- atica against flukical action of the amine of diana- phenethide in vitro. Moleodar and Biochemical Pharma- cology. 2, 339-48.
FEI-rEaER, R. H., Pax, R. A. & BENNErr,.I.L. (1980). Schis- losoma mansoni: characterization of the electrical potential fi'om the tegument of aduh males. Exper- imental Parasitolog), 49, 353-65.
FI.EMMe~'(., J. T., SQt,mE, M., SAVrZt.LE, D. B., Lzwts, J. A. (1994). Sequence Analysis of the lev-1 Dominant Alleles (x21, x61) Predict Alterations in the Ionic Sel- ectivity of the Channel. Worm Breeders Gazette. 13, 76.
Ft'jJm:~a, M., OEDA, K., Ixouz, H. & K-vro, T. (1992). A single amino acid substitution in the 13-tubulin gene of Neurospora confers both carbendazim resistance and diethofencarb sensitivity. Curreut Genetics 21, 399-404.
GARDNVg, D. R. & BREZD~:N, B. L. (1984). The sites of action of praziquantel in a smooth muscle of Lymnaea slagnalis. Canadian Journal of Physiology and Phmwta- cology. 62, 282-7.
GRrx;omz, G., LoIP,.aNO, G. & PACV~VD, P. (1989). Ca ̀-'+ and Sr '-'÷ entry induced Ca"* release from the intracellular Ca'-'* store in smooth muscle cells of rat portal vein. Journal of Physiology 474, 483-500.
GUENETTE, S., PRI'rCHARD, R. tC, ~.EIN, R. D. & I~LkTL~SHEW- SKI, G. (1991). Characterization of a ~-tubulin gene and [3-tubulin gene products of Brugia pahang~ Mol- ecular Biochemical Parasitology 44, 153-64.
GUENETrE, S., PR1TCHARD, R. K. & MATLASHEWSKI, G. (1992). Identification of a novel Brugia pahangi fl-tub- ulin gene (b2) and a 22-nucleotide spliced leader sequence on [Bl-tubulin mRNA. Moleotlar Biochemical Parasitology 50, 275-284.

MODES OF ACTION OF ANTHELMINTIC DRUGS 33
HARNETT, W. (1988). The anthehnintic action of pra- ziquantel. Parasitology Today 4, 144-6.
H..~,I~.NEVr, W. & KUSl.'I.,J. R. (1986). Increased exposure of parasite antigens at the surface of aduh male Schis- tosoma mansoni exposed to paraziquantel in vitro. Parasitology 93, 401-5.
HARROW, I. D. & GP~aTION, K. A. F. (1985). Mode of action of the anthelmintics mol-antel, pyrantel and levamisole ira the nmscle cell membrane of the nema- tode Ascaris sltum. Pesticide S¢'/eme. 16, 662-72.
Hawr;t.,4~., F. & L.~t'mv, W. (1949). Action of heu-azan on filariasis and onchocerciasis. Lancet 2, 146-7.
HH~R.XlAYX-V~Y~;, A., Dam.l.Xt,, E. & Ml-lssYzR, G. (1991). Functional characterization of the Ca ̀-'+ gated Ca ̀-'+ release channel of vascular smooth muscle sarco- plasmic reticulum. I~ugers Archives 418, 353-9.
Hol.ot.:x-m1-, L. & WaLm-~, R.J. (1990). Avermectin and avermectin derivatives are antagonists at the 4-mnino- butyric acid (GABA) receptor on the somatic mnscle- cells of Ascaris----is this the site of anthehnintic action. Parasitology 101,265-71.
.]o~txsox, P., MACKENZII-:, C. D., DF.xrIAM, D. A. & St'swll.l.O, R. R. (1991 ). The effect of diethylcarbamazine on the in vitro serum-mediated adgerence of feline granulo- cvtes to microfilariae of Brugian pahangi. Tropical iQedicine and Parasitology. 39, 291-4.
.It'.x(;, M. K., W..DV.R, I. B. & OAr~I.FV, B. R. (1992). Amino acid aherations ira the benA ([3-tubulin) gene of Aspergillus nidulans that confer benomyl resistance. Cell Motili(~' and @los]¢elelon 22, 170-4.
K~x~:sa--r~tasax, N., Dot'~,t.~s,J. G. & Kaztra, J. W. (1991). Diethylcarbamazine inhibits endothelial and microfi- larial prostanoid lnetabolisln in vitro. Molecular and Biochemical Pamsitoh)~', 49, 11-20.
Iga-:M~a~.l., S. B. & Pam=rvr, J. W. (1973). The ett'ect of diarnphenethide of Fasciola hepatica at different stages of development. Research in l'eterinmy Scieme 15, 37, 40.
L.u:~:v, E. (1990). Mode of action of benzimidazoles. Parasitology Today 6, 112-5.
k.u'~;Htox, D. L., Wotsr~-nru~..~:, A.J. & Lt'n-r, G. (1995). The beta subunit of the C. elegans inhibitor T gluta- mate receptor is expressed in the pm4 pharyngeal muscle cells. I'Vorm Breeders Gaze, tie 14, 48-.
Lt-t-, D. L. (1962). The distribution of esterase enzymes ira Asearis htmbricoides. Parasitology 52, 241-60.
Lems, J. A., Wt', C. -H., Lmn~-,J. H. & BER¢,, H. (1980). Levamisole-resitant mutants of the nematode Caenor- habditis elegans appear to lack pharmacological ace- tycholine receptors. Neuroscience 5, 967-89.
LFw~s, J. A. FLEM.X~L~t,, J. T. & BraD, D. (1992) Cloning nematode acetycholine receptor genes. In Ntntrotox '91, ed. Duce, I. R. pp. 155-64. London and New
York: Elsevier Applied Science.. L~ms, S., L~z, M. G.-S. & C o ~ y , N. J. (1985). Five
mouse tubtdin isotypes and their regulated expression during development. Jom~al of Cell Biology 101,852-61.
LUBEt;A, G. W., Ka.E~N, R. D., G~RV, T. G. & PRICHARD, R. K. (1994). Haemonchus contortus: the role of 2 beta- tubulin gene subfamilies in the resistance to benzimi- dazole anthelmintics. Biochemical Pharmacology 47, 1705-15.
Matzv:t_s, R. M. & DENHAM, D. A. (1992). Diethylcarbama- zine (DEC): immunopharmacological interactions of an anti-filarial drug. Parasitology 105, $49-$60.
MARVIN, R.J. (1980). The effect of g-aminobutyric acid in the input conductance and membrane potential of Ascaris nmscle. British Journal of Pharmacology 71, 99-106.
MaRviN, R.J. (1982). Electrophysiological effects of pip- erazine and diethylcarbamazine on Ascaris suum somatic muscle. British Journal of Pharmacology 77, 255-65.
MARTIN, R.J. (1985). y-aminobutyric acid- and pipera- zine-activated single channel cm-rents from Ascaris suum body muscle. B~itish Journal of Pharmacology 84, 445-61.
M.-~RNx, R. J. (1996). An electrophysiological prep- aration of Ascaris suum pharyngeal muscle reveals a glutamate-gated chloride channel sensitive to the avermectin analogue milbemycin D. Parasitology 147.
M-vm~.ws, W. R. & Mt'RPHV, R. C. (1982). Inhibition of leukotriene biosynthesis in mastocytoma cells by diethylcarbamazine. Biochemical Pharmacology. 31, 2129-32.
MC~HaR, Q. A. & KaNat~o, L. D. B. (1991). The pharma- colo~, of flukicidal drugs. British I/eterina O, Journal 147, 306-19.
MI.:III.IIORN, H., BFX:HEP,, B., ANDREWS, R., TIIO.XlAs, H. & Fm.:xKEI.,J. R. (1981). In vivo and in ~4tro experiments on the effect of praziquantel on Schistosoma mansoni: a light mad electron microscope study. AizneimittelFor- schung 31, 544-54.
MEi.i.xxllv, H. (1955). The identification and estimation of acetylcholine in three parasitic nematodes (Ascaris Imnbricoides, Litomosoides carinii, and the microfilar- iae of Dirofilaria repens). Parasitology 45, 287-94.
Molt.x.~lxi~:o-ALi, N. A. K. & Bot;ay, J. A. (1987). The pharmacodynamics of the flukicidal salicylanilides, rafoxanide, closantel and oxyclozanide. Jou~vml of I/et- e~ina O' Pharmacology and Thempe~ttics 10, 127-33.
OPPVR.~L~X, C. H. & CHaxt., S. (1992). Nematode acet-yl- cholinesterases-molecular-forms and their potential role ira nematode behavior. Parasitology T0dav 8, 406-11.
PAx, R. A. & BExnErr, J. L. (1989). Effect of closantel on intrategumental pH ira Sehistosoma mansoni and Fasci- ola hepatica. Journal of Parasitology 75, 169-71.
PmcH.~Rt), R. K. (1994). Anthehnintic resistance. Veterin- my Parasitology 54, 259-68.
lL~zt.,4, E., Ro~lm, L. C., KmHs, S., LIu, F.-T. & LEWIS, R. A. (1984). Pm analysis of the generation and the secretion of preformed mediators from mouse bone marrow-derrived mast cells.Journal ofhnmunology 133, 938-45.
ROBERTSON, S., J., PENNIN(;TON, A.J., Ev.~,,4s, A. M. & M..xR- wN, R. J. (1992). p)rrantel activated single-channel currents ira the nematode parasite Ascaris-suum. British Journal of Phal~nacology 107, P 154-.
ROBERTSON, S.J. 8c M:~dlTIN, R.J. (1993). Levarnisole-acti- vated single-channel currents from muscle of the nematode parasite Ascaris suum. British Journal of Phar- macology 108, 170-8.
Roos, M. H., KwA, M. S. G. & GRANT, W. N. (1995). New genetic and practical implications of selection for

34 THE \T_TERINARY.I()URNAL. 15,t. 1
anatlaelmintic resistance in parasitic nematodes. Para- sitolo#, Today 11, 148-50.
S..~X¢.STH~, N. C,,., PmTcH,~RD, R. K. & L~cl.:Y, E. (1985). Tubul in anicl benzimidazole-resistance in 7*richa~hvn- ~'ltts colublqfin'n6s (Nematoda). .Journal of Parasitoh~n' 71,645-51.
S(:IIt'I.MAN, M, D., OY, TI.INI), D. A. & \~'AI+I-XTIN(L D. (1982). Mechanism of action of ink-401 against I"as¢iola hep- a t ica- inhib idon of phosphoglyccrate kinase. Moh,cu- lar and Biochemical Parasitolo~' 5, 133-45•
SCIIt'I.MAN, ~'1. D. & VAI.ENHNO, D. (1£182). Ptu'ification, clmracterization and inhibit ion by ink-401 of Fasciola hepatica phosphoglyceromtmlse. A.loh,cular And Bio- Hlemical Parasitolo~, 5, 321-32.
S1(;1'~7% H. & Ft'Rt"~7\MA, S. (19q0). Sphingisine increases inositol t r isphosphate in rat parot id acinar-cells by a mechanism that is i ndependen t of protein kinase C but d e p e n d e n t on extracellular calcium. ('ell Calciltm 11,469-75.
SHoop, W. L., MRoztK, H. & F~sllE8, M. H. (1995). Struc- ture and activity of avermectins and mill+emycins in animal heahh, l'elerinan.' l)arasitolo~ ' 59, 139-56.
SHN(;I., P.. PIERCI-, P. F., CoNy(m, D. H., (;ll+,S()x, D, W., Sre,..\~ssL'-:, T., Ross, M. A. & thBAs. ]. L. (19881. Does dexmnethasone suppress the /Vlazzoui reaction in parients with onchocerciasis. Acta "l)opica. 45, 77-85.
SrR~l~e, L. 1995 BioHwmisth,. New York: W. H. Freeman and Company..
Tuox,,sox, D. P., P.x×, R. A. & B~:xx~:r~,.]. 1+. ( 19841. Mic-
u-oelectrode studies on tile tegument and sub-tegu- mental compar tments of male ,~'{'hi.xlO.%'Olll(I IR(IIIM)HI; [111 analysis of coxnplulmcnts of electrophysiological prol~erties, l-xperimenla! l~arasilolak.n ' 58, 314-15.
VAx DI-X Boss(:nuu<, H. 1972 Comparalive Bio¢hemish)' qf Pamsih,s. New ~)rk: Academic Press..
V.xx DP:x BosscIH:, H., Roslil.:t-I+l.:, F. & H(ml(;, (:. (1982). Mebendazole and rehtted anthelmintics. Advances in Phannacvlo,t,~' and Chemotherapy 19, 67-128.
VAN DI-N B()ss(:III.:, H. & Dl.: N()t.l.tX, S. (1973). Biochemi- cal effects o f mebendazole on Trichinella spimh,.~ lar- vae.Journal of pal:~ilolo~' 59, 970-6.
\qcK~:RY, A. C., N..wxR, J. K. & T..\xwLux, M. t+. (1986). Diet lwlcarbmnazine on microf i lar iae o f Lilomisoides carinii in viho arm in vivo. ~,itshrifi fit r Pam.~itenku ride 72, 805-13.
VCH-sJ, P. M., Li, Y., OLns. (;. R. & B()wn-x, W. D. (19921. Sehish~.~oma mansoni: characterizat ion ol + phosph,finos- itide response. Experimental IJam.~iloloLq ' 74, 38-45.
W()Lm:-Mt'ssIl.:, E., V.~xl)i--W.~..x,.]., PAx, R. A., Fi.:nqH~i.:~, R. H. & BI~XNI.71-I, .1. L. (19821. ,~¢hi.#osonta matl.xtmi: cal- cium elllux and the ell~'cts of calcium-free media on the responses of tile electrical i~otential fr(,m the tegument of adult males nltlscttlitttlrc lo pritziqtlanlcl arid o ther agents inducing contract ion. I'~xperimenlal Parasilolot,9" 53, 270-8.
(,4r¢'eph,d fi~r publical#m 21lure' 1996)











![[1].pdf - ResearchGate](https://img.dokumen.tips/doc/110x75/62129a6cbbf9242e6965a6a7/1pdf-researchgate.jpg)

















