Embed Size (px)
Citation preview

Review ArticleLong Noncoding RNAs in Metabolic SyndromeRelated Disorders
Magdalena Losko Jerzy Kotlinowski and Jolanta Jura
Faculty of Biochemistry Biophysics and Biotechnology Department of General Biochemistry Jagiellonian University Krakow Poland
Correspondence should be addressed to Jolanta Jura jolantajuraujedupl
Received 12 May 2016 Accepted 5 October 2016
Academic Editor Teresa Zelante
Copyright copy 2016 Magdalena Losko et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Ribonucleic acids (RNAs) are very complex and their all functions have yet to be fully clarified Noncoding genes (noncoding RNAsequences and pseudogenes) comprise 67 of all genes and they are represented by housekeeping noncoding RNAs (transferRNA (tRNA) ribosomal RNA (rRNA) small nuclear RNA (snRNA) and small nucleolar RNA (snoRNA)) that are engaged inbasic cellular processes and by regulatory noncoding RNA (short and long noncoding RNA (ncRNA)) that are important for geneexpressiontranscript stability In this review we summarize data concerning the significance of long noncoding RNAs (lncRNAs)in metabolic syndrome related disorders focusing on adipose tissue and pancreatic islands
1 Introduction
Recent genome-wide transcriptome studies have revealedthat the majority of the mammalian genome is transcribedthereby giving rise to a range of coding and noncodingRNA (ncRNA) transcripts [1] The latest Human GENCODERelease (version 24) presented by the Encyclopedia of DNAElements Consortium (ENCODE) showed that the humangenome is composed of 60554 genes (Figure 1) Amongthose 19815 are protein-coding genes whereas the remainingtwo-thirds of the genome represents noncoding RNA genes(15941 long noncoding RNA genes 9882 small noncodingRNA genes and 14505 pseudogenes) Noncoding RNAs arepredominant in the genome and are classified as either house-keeping or regulatory noncoding RNAs Housekeeping non-coding RNAs are constitutively expressed and they functionas key regulatory molecules of many cellular processes Thisgroup includes ribosomal (rRNA) transfer (tRNA) smallnuclear (snRNA) and small nucleolar RNAs (snoRNAs)Thesecond group regulatory noncoding RNAs is divided intoshort ncRNAs (lt200 nucleotides) or long noncoding RNAs(lncRNAs gt200 nucleotides) Short ncRNAs are representedby microRNAs (miRNAs) small interfering RNAs (siRNAs)and Piwi-associated RNAs (piRNAs) and in few cases by
antisense RNAs and enhancer RNAs (eRNAs) Long non-coding RNAs group includes antisense RNAs and enhancerRNAs (eRNAs) [1ndash3] (Figure 2)
In this review we focus on the broadest class of ncRNAas represented by lncRNAs This class of ncRNAs was firstdiscovered in the mouse during large-scale sequencing offull-length cDNA libraries [4] Generally lncRNAs werecharacterized as transcripts longer than 200 nucleotides withsome features typical for protein-coding mRNA They aretranscribed by RNA-polymerase (Pol) II [5] capped onthe 51015840 end polyadenylated and commonly expressed asalternatively spliced variants Moreover lncRNAs were foundto have the same trimethylation marks of H3K4 and H3K36at their promoters and transcribed regions [1] Howeverthey possess unique characteristics that distinguish themfrommRNA In contrast to mRNA lncRNAs usually containintronexon structure they are expressed at lower levels oftenin a tissue-specific manner and their sequence is poorlyevolutionary conserved [6] Additionally they do not possessopen reading frames (ORFs) 31015840 UTR and terminationregions thus the majority have limited coding potentialNevertheless recent studies have demonstrated that somelncRNAs can be associated with polysomes and ribosomessuggesting that they may act as coding transcripts and give
Hindawi Publishing CorporationMediators of InflammationVolume 2016 Article ID 5365209 12 pageshttpdxdoiorg10115520165365209
2 Mediators of Inflammation
Protein-codinggenes33
Long noncodingRNA genes
Small noncodingRNA genes
Pseudogenes
Noncodinggenes67
Human genesHuman GENCODE release (version 24)
27
16
24
Figure 1 Schematic illustration of human genome presented by theEncyclopedia of DNA Elements Consortium (ENCODE) Detaileddescription of gene representation in the main text
rise to small peptides [7] An emerging number of studiesrelated to the discovery of new lncRNAs and their position tothe nearest protein-coding genes have resulted in the intro-duction of a new nomenclature for these molecules Longnoncoding RNAs may be classified into five categories (1)sense lncRNAs that overlap the nearest protein-coding genesat the same strand (2) antisense lncRNAs located across theexons of protein-coding genes from the opposite strand (3)bidirectional lncRNAs transcribed on the opposite strandwithin 1 kb from the nearest protein-coding gene (4) introniclncRNAs that overlap intronic regions of coding genes ineither the sense or antisense orientation (5) intergeniclncRNAs that represent the largest group of lncRNA theyare located between protein-coding genes but are at least 1 kbaway from the nearest protein-coding gene [3] (Figure 3)
2 Biological Function of LongNoncoding RNAs
In recent years numerous studies have aimed to iden-tify functions of newly discovered long noncoding RNAsAlthough the primary role of lncRNAs was their epigeneticregulation of protein-coding gene expression only a limitednumber of such long noncoding transcripts have been iden-tified [21] It was soon discovered that lncRNAs play a rolein variety of biological process acting in the nucleus or inthe cytoplasm or in both [22] Functions of lncRNAs aremanifested by three types of interaction RNA-RNA RNA-DNA and RNA-protein all of which depend on the site ofaction [23]
In the nucleus for example the main role of lncRNAsis assumed to be epigenetic imprinting One of the best-described mechanisms of lncRNAs action during the epige-netic regulation is the X chromosome inactivation in whichX chromosome inactive-specific transcript (XIST) plays akey role [24] Recent studies demonstrated that lncRNAscan interact with chromatin remodeling complexes includingpolycomb repressive complex (PRC2) 2 TrithoraxMLL andH3K9 methyltransferase G9a acting as docking platforms tospecific genomic loci [4] These protein complexes modifyDNAmethylation patterns via trimethylation leading to tran-scriptional repression (PRC2) or activation (TrithoraxMLL)of the lncRNA target genes Numerous lncRNAs have been
recognized as scaffolds for inhibitory PRC2 including XISTKCNQ1OT1 andHOTAIR [8 25]Other lncRNAs likeHOT-TIP directly bind to protein complexes of TrithoraxMLL [9]In these situations lncRNAs have been shown to drive orrepress gene expression respectively Moreover it has beenpostulated that these lncRNAs are guides for chromatin-modifying complexes where they first recognize the targetlocalizations at the chromatin bind to them and then formdocking platforms for protein partners [26]
In addition to regulation of genomic imprinting lncR-NAs are also involved in the control of gene expressionat the transcriptional level lncRNAs can directly interactwith transcription factors acting as decoys or inhibitors fortheir binding to the target DNA sequence For exampletwo lncRNA molecules Lethe and p50-associated Cox-2extragenic RNA (PACER) have been shown to interact withdifferent subunits of NF-120581B thus preventing it from bindingto the promoter region of the target gene [10 27] More-over lncRNAs exert their regulatory function during mRNAprocessing and stability interacting with the heterogeneousnuclear ribonucleoproteins (hnRNPs) An example of thistype of RNA-protein linkage is demonstrated for severallncRNAs including lincRNA-p21 [28] lincRNA-Cox2 [29] orTHRIL [11] thus they affect either activation or repressionof target genes Other actions of lncRNA in the nucleusare manifested by alternative splicing regulation [12] andsubnuclear compartment formation [30] (Figure 4)
In the cytosol lncRNAs exert their function by interact-ing with target mRNAs (or miRNAs) through base-pairingIn this manner lncRNAs may either stabilize (eg BACE1-AS prevents miRNA-induced repression of BACE1 transcript[31]) or decay (eg 12-sbsRNAs bind to the Alu elementof SMD target mRNAs within the 31015840 UTR region [32])target transcripts Moreover lncRNAs promote translation oftranscripts (eg antisense Uchl1 interacts with Uchl1 mRNAresulting in recruitment of ribosomes [13]) or repress thisprocess (eg lincRNA-p21 binds to target mRNA causingrecruitment of translation repressors [14]) (Figure 4)
3 Inflammation and Its Significance inMetabolic Syndrome
Inflammation is a complex process aiming to defend thebody against harmful agents by removing or neutralizingthem in order to restore tissue homeostasis [33] The com-plexity of this process results from the broad spectrum ofinflammatory pathways various inflammatory inducers sen-sors andmediators [34] In the classic view the inflammatoryresponse is induced immediately after stimulation by theinflammatory stimuli such as a bacterial pathogen [33] Theforeign antigen is recognized by the sensors of the hostdefense cell Cells such as neutrophils dendritic cells (DCs)and macrophages express evolutionarily conserved Toll-likereceptors (TLRs) on their surface The TLR family is com-posed of several classes of transmembrane proteins and rec-ognizes specific structures present on the microbes knownas pathogen-associated molecular patterns (PAMPs) [35]Induction of TLRs triggers a multistep signaling cascadethereby resulting in the modulation of gene expression
Mediators of Inflammation 3
NoncodingRNAs
HousekeepingncRNAs
asRNAseRNAs
miRNAssiRNAspiRNAs
rRNA tRNA snRNA snoRNA
RegulatoryRNAs
ShortncRNAslt200nt
LongncRNAsgt200nt
Figure 2 Types of noncoding RNAs (ncRNAs) Noncoding RNAs are classified into housekeeping and regulatory noncoding RNAsHousekeeping ncRNAs include ribosomal (rRNA) transfer (tRNA) small nuclear (snRNA) and small nucleolar RNAs (snoRNAs)Regulatory noncoding RNAs are divided into short ncRNAs (lt200 nucleotides) or long ncRNAs (gt200 nucleotides) including microRNAs(miRNAs) small interfering RNAs (siRNAs) Piwi-associated RNAs (piRNAs) antisense RNAs (asRNAs) and enhancer RNAs (eRNAs)
BidirectionallncRNA
SenselncRNA
Intronic lncRNA
Intronic lncRNA
AntisenselncRNA
Intergenic lncRNA
Protein-coding gene
Figure 3 Classification of long noncoding RNAs lncRNAs (shownas red boxes) may be classified into five categories sense lncRNAsantisense lncRNAs bidirectional lncRNAs (transcribed on theopposite strand within 1 kb from the nearest protein-coding gene)intronic lncRNAs and intergenic lncRNA (transcribed betweenprotein-coding genes but they are located at least 1 kb away from thenearest protein-coding gene) [3] Detailed descriptions of lncRNAclasses are in the main text
involved in the immune response Key processes regulatedby TLRs are related to the secretion of various mediators byimmune cells phagocytosis cell migration metabolic repro-gramming and tissue repair [36]
In general based on the length the inflammatory res-ponse can be classified into acute or chronic Acute inflamma-tion usually develops locally and its role is to eliminate toxicagents repair damaged tissue and restore homeostasis Insome circumstances local inflammation might be greatertherefore called a ldquogeneralized inflammatory responserdquowhich is associated with the response of the whole organism
[37] Although the acute phase leads to the beneficial recoveryof the injured tissue and healing this nonspecific immuneprocess could have also negative effects Without the properresolution phase it can easily turn into a chronic stateChronic inflammation may appear after the switch from theacute phase and it is characterized by the specific long-termcellular and humoral immune response present at the site oftissue injury [38] In consequence it may significantly con-tribute to dangerous pathophysiological changes and initiatethe development of chronic diseases
Each inflammatory stage is subjected to precise controlby several transcription factors involved in the immune res-ponse [39] One such factor is NF-120581B (nuclear factor kappa-light-chain-enhancer of activated B cells) representing afamily of transcription factors NF-120581B plays an important rolein gene expression regulation of multiple factors involved inimmune acute phase and inflammatory responses [40] NF-120581B in addition to controlling the development and functionof the immune system is required in other physiologicalprocesses such as suppression of apoptosis and cell prolif-eration and differentiation In mammals the active formof NF-120581B is composed of homo- and heterodimers of theNF-120581B family members which can be divided into fivedifferent subunits p50 p52 p65 (RelA) c-Rel and RelB Allof these share homology of the specific N-terminal domainresponsible for the dimerization and binding to target DNAsequences Depending on the local cytokines produced bythe immune cells NF-120581B also controls macrophage relocal-ization activation and differentiation into two different phe-notypes proinflammatory (M1) or anti-inflammatory (M2)In response to proinflammatory cytokines NF-120581B is alsoinvolved in the activation of T lymphocytes which prolife-rate and secrete cytokines such as interferon gamma (IFN120574)interleukin 6 (IL-6) or TNF-120572The result of NF-120581B activationis stimulation of proinflammatory cytokine production by
4 Mediators of Inflammation
SRSRSR
SR
MALAT1
P PP
PP
PPP
P
P
Trx HOTTIPHOTAIR
H3K4me3H3K27me3
Nucleus CytosolPRC1
PRC2 MLL
Pre-mRNA
mRNA
(A) Epigenetic imprinting
(B) Transcription
(C) Alternative splicing
(D) miRNA sponge
(E) mRNA stabilitydegradation
(F) Translation
Uchl1 lincRNA-p21
MIAT
BACE1-AS
miR-150-5p
p50PACER
THRILhnRNP
COX-2p65
Transcriptionalactivation
Transcriptional repression
TNF-120572
Figure 4 Biological role of lncRNAs Role of lncRNAs has been implicated in the regulation of diverse processes manifested by three waysof interactions RNA-RNA RNA-DNA and RNA-protein both in the nucleus and in the cytoplasm (A) In the nucleus lncRNAs have beenshown to play a key role in imprinting control lncRNAs may act as docking platforms for the chromatin remodeling complex polycombrepressive complex (PRC2) 2 (eg HOTAIR) which methylates histone H3 at lysine 27 (H3K27me3) or with the heterogeneous group ofTrithoraxMLL proteins (eg HOTTIP) leading to a repression or gain of transcriptional activity respectively [8 9] (B) Other regulatoryroles of lncRNAs in the nucleus include transcriptional regulation by interactingwith transcription factors (eg PACERdirectly interacts withthe repressive subunit of NF-120581B p50 thus preventing it from binding to the Cox-2 promoter [10]) heterogeneous nuclear ribonucleoproteins(hnRNPs) (eg THRIL together with hnRNPL acts as RNA-protein complex and binds to TNF-120572 promoter and mediates the induction ofTNF-120572 expression [11]) (C)Moreover lncRNAsmay regulate the pre-mRNA alternative splicing (eg MALAT1) which acts as scaffold for SRproteins present in nuclear speckles andmodulates their phosphorylation [12] In the cytosol lncRNAs exert their function by interactingwithtarget transcripts and miRNAs through base-pairing (D) For miRNAs lncRNAs may act as a molecular sponge preventing specific miRNAsfrom binding to their target mRNAs (eg MIAT binds to miR-150-5p thereby upregulating the level of miR-150-5p target gene) (E) lncRNAbinding to mRNA may stabilize (eg BACE1-AS prevents miRNA-induced repression of BACE1 transcript) or decay target transcripts (F)Moreover lncRNAs promote (eg antisense Uchl1 interacts with Uchl1 mRNA resulting in recruitment of ribosomes [13]) or repress (eglincRNA-p21 which binds to target mRNA causing recruitment of translation repressors [14]) translation of transcripts This figure wasproduced using modified elements from the Servier Medical Art (httpwwwserviercom)
macrophages and T lymphocytes thereby enhancing theinflammatory response [41]
One disease closely linked to a chronic inflammatory stateis obesity It is characterized by an excessive fat accumu-lation and enhanced production of proinflammatory cyto-kineschemokines by adipose tissues It is commonly causedby a combination of excessive food intake lack of physicalactivity and genetic susceptibility [42] A great majority ofobese individuals present features of themetabolic syndrome(a) increased waist circumference (b) insulin resistance(c) hyperglycemia (d) hypertension and (e) hypertriglyc-eridemia [15] Appearance of these symptoms is associated
with a high risk of serious disease development includingtype 2 diabetes cardiovascular diseases [18] nonalcoholicfatty liver disease (NAFLD) [17] and even some cancers [19]Themechanismofmetabolic disorder development is still notfully understood nevertheless numerous studies have sug-gested an adipocyte dysfunction as a main cause of metabolicsystem failure [43] In this pathological state adipose tissuesecretes various mediators including cytokines inducing achronic low-grade inflammatory response that results inrecruitment of immune cells into the tissue
The excessive accumulation of tissue fat leads to distur-bance both in the secretory and in the storage functions of
Mediators of Inflammation 5
adipocytes It is postulated that the excessive fat mass inducesthe production of reactive oxygen species and the develop-ment of chronic oxidative stress in patients with obesity andmetabolic dysfunction [44 45] In adipose tissue of obesepatients is observed severe inflammatory response charac-terized by high production of cytokines and increased infil-tration of immune cells including monocytes stimulated todifferentiate into macrophages and lymphocytes T activatedto proliferate and secrete cytokines [46 47] Cytokinessecreted by activated macrophages act on TLRs present onthe surface of adipocytes stimulating them to produce adipo-kines and cytokines [48] Based on these observations it hasbeen found that the reactions of both the immune and meta-bolic systems are closely connected and they play an impor-tant role in the development of metabolic disorders
4 lncRNA in Obesity andAdipogenesis Control
Adipocytes play a central role in energy homeostasis byfine-tuning the equilibrium between nutrient deposition(white adipose tissue WAT) and energy expenditure (brownadipose tissue BAT) Additionally adipose tissue acts asan endocrine organ by secreting factors (adipokines) thatregulate whole body energy and glucose homeostasis [49]Adipogenesis is a complex process governed by a network oftranscription factors cofactors and signaling intermediatesfrom numerous pathways [50] The transcriptional cascadethat regulates adipocyte differentiation is predominantlydriven by peroxisome proliferator-activated receptor gamma(PPAR120574) shown to be both necessary and sufficient foradipogenesis [51] Importantly adipogenesis is highly regu-lated not only by PPAR120574 but also by the coordinated effectsof other transcription factors including CCAATenhancer-binding proteins (CEBPs) Kruppel-like factors (KLFs) orWingless proteins (Wnt) and transcriptional cofactors [52ndash54]
In recent years there has been growing attention paid tononcoding RNAs as a novel cell intrinsic regulatory mecha-nism Specifically lncRNAs are now emerging as importantregulators of gene expression both on the transcriptional andon the posttranscriptional levels Long noncoding RNAs arebeing discovered using modern techniques of microarray ornext-generation sequencing However only small portionshave been fully characterized In the case of adipogenesisthere is wide expression of lncRNAs controlling a varietyof genes involved in the formation differentiation andactivation of adipocytes (Figure 5)
Steroid receptor RNA Activator (SRA) lncRNA was des-cribed for the first time as a molecule regulating adipo-genesis [55] It was shown that lncRNA SRA initially identi-fied as a coactivator of steroid receptors also acts as a tran-scriptional coactivator of PPAR120574 [55 56] lncRNA SRA pro-motes adipocyte differentiation by binding to the N-terminalportion of PPAR120574 and enhances its transcriptional activ-ity Additionally its overexpression significantly increasedmRNA and protein levels of adipocyte master regulatorsPPAR120574 and CEBP120572 as well as PPAR120574 target genes Con-versely knockdown of endogenous lncRNA SRA resulted in
inhibition of 3T3-L1 preadipocyte differentiation Moreovergene expression profiling showed that lncRNA SRA regulatesexpression of genes related to various cellular processesincluding cell cycle and insulin transduction pathways [55]Further experiments proved that lncRNA SRA overexpres-sion inhibits phosphorylation of p38 mitogen activated pro-tein kinase (MAPK) and c-JunNH2-terminal kinase (JNK) inthe early differentiation of ST2 mesenchymal precursor cellsIn contrast knockdown of lncRNA SRA increased p38 andJNK phosphorylation while reducing insulin receptormRNAand protein levels which led to decreased downstreamsignaling [57] Supporting that data SRA knock-out mice(SRAminusminus) are resistant to high fat diet (HFD) induced obesityAfter 14 weeks of HFD SRAminusminus mice are characterized byreduced WAT mass and decreased expression of adipocytegenes such as adiponectin or fatty acid-binding protein 4Thelean phenotype is also associated with smaller adipocytes inWAT compared to WT mice reduced liver mass fewer lipiddroplets in the liver and decreased expression of lipogenesis-associated genes Finally SRAminusminus animals have improvedinsulin sensitivity [58]
In a recent paper Xiao and coworkers [59] identifiedlncRNA ADINR (adipogenic differentiation induced non-coding RNA) that in cis transcriptionally activates CEBP120572It was upregulated 20ndash30-fold during the course of adipoge-nesis and was transcribed from a position sim450 bp upstreamof the CEBP120572 gene Knockdown of lncRNA ADINR withsiRNA resulted in a dramatic adipogenic defect as shown bya decreased number of Oil Red O positive cells and reducedadipogenic transcripts CEBP120572 PPAR120574 fatty acid-bindingprotein 4 and lipoprotein lipase Importantly inhibition ofadipogenesis caused by depletion of lncRNA ADINR wasrescued by overexpression of CEBP120572 The lncRNA ADINRmechanism of action relies on its binding to PA1 followed byrecruitment ofMLL34 histonemethyltransferase complexesIn turn that causes an increase inH3K4me3 and a decrease inH3K27me3 histone modification in the CEBP120572 locus duringadipogenesis [59]
Another lncRNA that induces adipogenesis in ciswas dis-covered within themouse PU1 locus Expression of the PU1gene gives rise to both the mRNA encoding PU1 proteinand antisense (AS) lncRNA originating from a promoter inthe antisense strand of intron 3 [60] The protein PU1 wasoriginally demonstrated to be a key transcription factor regu-lating hematopoiesis but recent studies revealed that it is alsoexpressed in 3T3-L1 preadipocytes and murine adipocytesisolated from WAT [60 61] Gain and loss of functionstudies established the PU1 protein as an inhibitor of 3T3-L1 preadipocyte differentiation acting via downregulation ofPPAR120574 [62] On the other hand PU1 AS lncRNA promotesadipogenesis through preventing PU1 mRNA translationby binding to PU1 mRNA to form an mRNAAS lncRNAduplex [60] Formation of this sense-antisense RNA duplexwas also confirmed in porcine adipocytes by an endogenousribonuclease protection assay combinedwithRT-PCR In linewith the previous data expression of PPAR120574 and fatty acidsynthase (FASN)was significantly upregulated in the lncRNAPU1 shRNA treated group [63]
6 Mediators of Inflammation
Diabetic retinopathy
CVDs
Type 2 diabetes
Cancers
NAFLD
ObesitySRA
ADINRNEAT1lncRAPs
MALAT1MIAT
HI-LNC25KCNQ1OT1LOC283177
Risk factors
(i) Abdominal obesity
(ii) Insulin resistance
(iii) Hyperglycemia
(iv) Hypertriglyceridemia
(v) Hypertension
uarr 535 lncRNAsdarr 1200 lncRNAs
Figure 5 Development of metabolic syndrome The common clinical symptoms of metabolic disorders include abdominal obesity insulinresistance hyperglycemia hypertriglyceridemia and hypertension [15] Their appearance is associated with a high risk for development ofserious diseases including type 2 diabetes diabetes-related diabetic retinopathy [16] and nonalcoholic fatty liver disease (NAFLD) [17] as wellas cardiovascular diseases (CVDs) [18] and even some cancers including liver pancreas and colon [19] (CVDs and cancers are not discussedin this review) Selected lncRNAs involved in the control of metabolic system activity (eg adipogenesis SRA ADINR NEAT1 and lncRAPs)and pathology of metabolic diseases (eg type 2 diabetes HI-LNC25 KCNQ1OT1 LOC283177 diabetic retinopathy MALAT1 and MIAT)are marked with the red color In NAFLD 535 lncRNAs are upregulated and 1200 are downregulated however these sequences are not yetwell characterized [20] This figure was produced using modified elements from the Servier Medical Art (httpwwwserviercom)
In a number of studies global transcriptome profilingwasused to select specific lncRNAs either involved in adipogen-esis or characteristic for adipocytes specifically isolated fromWAT or BAT In a seminal study done by Sun and cowork-ers [64] the transcriptome of primary brown and whiteadipocytes preadipocytes and cultured adipocytes was pro-filed Scientists identified 175 lncRNAs that were specificallyregulated during adipogenesis To further validate lncRNAsthat are functionally important for adipogenesis researchersselected the top 20 lncRNA genes based on the followingcriteria (i) significant upregulation in both brown and whitefat cultures (ii) binding to PPAR120574 or CEPB120572 promoters and(iii) independent validation of adipose-specific expressionBy using RNAi-mediated loss of function the top 10 mostimportant lncRNAs were selected and called Regulated inAdiPogenesis (lncRAPs) in order to highlight their key rolein proper differentiation of adipocyte precursors [64]
In comparison to white adipocytes brown adipocyteshave a higher mitochondrial content and express uncouplingprotein 1 (UCP1) which disperses chemical energy throughheat production Brown adipose tissue helps to regulateenergy expenditure in rodents and newborn babies butit has been considered to have no physiologic relevancein adults [65 66] However recent studies demonstratedthat metabolically functional BAT is also present in adulthumans As a result an understanding of BAT physiologymight provide an effective treatment of obesity or othermetabolic disorders [67] Identification of the lncRNAs cru-cial for BAT differentiation and function was done usingwhole transcriptome RNA sequencing Next gain and loss offunction studies established lncRNA Blnc1 as a potent acti-vator of thermogenic adipocyte differentiation It functionsupon formation of RNA-protein complex with the early Bcell factor 2 (EBF2) transcription factor to stimulate BAT
Mediators of Inflammation 7
formation Additionally lncRNA Blnc1 itself is a target ofEBF2 thereby forming a feed-forward regulatory loop Onthe other hand despite its stimulatory effects on brown pre-adipocyte differentiation lncRNA Blnc1 failed to promotedifferentiation of 3T3-L1 cells into UCP1 positive adipocytes[68] Another global profiling of gene expression duringmouse brown fat cell differentiation followed by candidatelncRNAs genomic context analysis pathway analysis andgene ontology enrichment of their associated protein-codinggenes was recently described In this study scientists identi-fied three lncRNAs (Gm15051 Tmem189 and Cebpd) associ-ated with their flanking coding genes (Hoxa1 CEBP120573 andCEBP120575) that participated in adipose commitment [69]
More recently RNA-seq analysis of murine brown ingui-nal white and epididymal white fat allowed for identificationof lncRNA cluster enriched in BAT Among them thelncRNA BATE1 was induced during brown adipose tissuedifferentiation Inhibition of BATE1 led to decreased expres-sion of brown fat markers and mitochondrial genes Authorsshowed that BATE1 binds to the heterogeneous nuclear ribo-nucleoprotein U a factor also required for BAT formation[70]
Finally interactions of lncRNAs with microRNAs in thecontext of adipogenesis were also described As demonstratedby Gernapudi and coworkers [71] the miR-140lncRNANEAT1 signaling network is necessary for adipogenesisAdipocyte derived stem cells isolated from miR-140 knock-out mice had dramatically decreased adipogenic capabilitiesfound to be associated with the downregulation of lncRNANEAT1 expression Overexpression of NEAT1 in such cellswas sufficient enough to restore their differentiation capabil-ities Authors identified a miR-140 binding site in the NEAT1sequence They also found that the physical interaction withmiR-140 in nucleus led to increased expression of NEAT1[71]
Excessive fat accumulation not only causes pathologicalexpansion of adipose tissue but also leads to triglyceridedeposition in the liver Presence of lipid droplets in excessof 5 of total hepatocytes is a diagnostic hallmark of non-alcoholic fatty liver disease (NAFLD) [72] Although mostpatients suffer from a mild course of illness approximately25 of cases progress and can subsequently lead to thedevelopment of steatohepatitis hepatic fibrosis liver cirrho-sis and hepatoma [73] Scientists and physicians estimatethat between 20 and 30 of Westrsquos general populationsuffers from NAFLD which makes this affects more than abillion people worldwide [74] To address questions aboutthe global expression pattern and functional contribution oflncRNAs during NAFLD Sun and coworkers [20] analyzedmicroarray expression using RNA extracted from liver biop-sies of healthy patients and those suffering from NAFLDIn this analysis 535 lncRNAs were found upregulated inNAFLD samples compared with controls and 1200 weredownregulated in NAFLD (Figure 5) Out of these sevenNAFLD samples that had highly up- or downregulatedgene expression were validated by quantitative real-timePCR (qPCR) Three lncRNA results obtained by qPCR weredifferent in comparison to the microarray data showing nochange between analyzed groups However changes in other
selected hits were confirmed highlighting the importance ofmicroarray data validation [20]
5 lncRNA in Diabetes and PancreaticFunctionGlucose Homeostasis
Pancreatic islets through the secretion of endocrinehormones such as insulin and glucagon play a key role inmetabolic homeostasis However one of the most commonmalfunctions of 120573-cells is caused by lack of insulin leading tothe development of diabetes [75] Additionally diabetes maydevelop as a result of insulin resistance a condition in whichthe body cannot effectively use the insulin produced by 120573-cells As a result blood sugar levels are deregulated and vari-ous tissues are exposed to prolonged hyperglycemia that overtime causes serious damage to many of the bodyrsquos systemsIt should be emphasized that hyperglycemia a hallmark ofdiabetes can cause both acute and long-term complicationslike cardiovascular disease stroke foot ulcers ketoacidosisor hyperosmolar coma [76] Based on etiology WHOclassifies diabetes into four categories but the vast majorityof cases fall into two broad etiopathogenetic groups type 1diabetes mellitus (T1DM) or type 2 diabetes mellitus (T2DM)[77]
Accounting for 5ndash10 of all cases T1DM is characterizedby the loss of insulin producing120573-cells due to an autoimmunereaction Autoimmune destruction of 120573-cells is unrelated tolifestyleThere are genetic variants known to increase the riskof T1DM development Type 2 diabetes mellitus represents90ndash95 of all cases and unlike T1DM is strongly associatedwith patientsrsquo lifestyle [78] A number of lifestyle factors areknown to be important for development of T2DM includingobesity (BMI above 30) lack of physical activity poor dietor stress [79] In fact most patients suffering from T2DM areobese It is a chronic disease that begins with insulin resist-ance a condition in which cells (mostly adipocytes hepato-cytes andmuscle cells) fail to properly absorb andmetabolizeglucose As a result 120573-cells increase insulin production toovercome hyperglycemia but this compensatory mechanismis transient and generally fails over time [77] Intensifiedmetabolism of 120573-cells leads to a gradual loss of their endo-crine function and finally causes 120573-cell apoptosis Similar toobesity diabetes is a worldwide problemAccording toWHOin 2014 the global prevalence of diabetes was estimated tobe 9 among adults aged over 18 years and it is expectedto double before 2030 [75 80] Additionally experts projectthat in 2030 diabetes will be the 7th leading cause of death[75]
Initial studies aimed at identification of specific lncRNAsexpressed in murine and human pancreatic islets were donein 2012 [81 82] By using next-generation sequencing scien-tists revealed a large collection of 1359 intergenic lncRNAsexpressed in murine 120573-cells (Figure 5) Many of them werehighly tissue-specific but their function was not evaluatedand future experiments are still needed [81] Similar numbersof 1128 intergenic and antisense lncRNAs were identifiedin human 120573-cells and many of them (eg HI-LNC12 andHI-LNC77) were highly tissue-specific [82] AdditionallylncRNA-encoding genes were preferentially located near
8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Mediators of Inflammation
Protein-codinggenes33
Long noncodingRNA genes
Small noncodingRNA genes
Pseudogenes
Noncodinggenes67
Human genesHuman GENCODE release (version 24)
27
16
24
Figure 1 Schematic illustration of human genome presented by theEncyclopedia of DNA Elements Consortium (ENCODE) Detaileddescription of gene representation in the main text
rise to small peptides [7] An emerging number of studiesrelated to the discovery of new lncRNAs and their position tothe nearest protein-coding genes have resulted in the intro-duction of a new nomenclature for these molecules Longnoncoding RNAs may be classified into five categories (1)sense lncRNAs that overlap the nearest protein-coding genesat the same strand (2) antisense lncRNAs located across theexons of protein-coding genes from the opposite strand (3)bidirectional lncRNAs transcribed on the opposite strandwithin 1 kb from the nearest protein-coding gene (4) introniclncRNAs that overlap intronic regions of coding genes ineither the sense or antisense orientation (5) intergeniclncRNAs that represent the largest group of lncRNA theyare located between protein-coding genes but are at least 1 kbaway from the nearest protein-coding gene [3] (Figure 3)
2 Biological Function of LongNoncoding RNAs
In recent years numerous studies have aimed to iden-tify functions of newly discovered long noncoding RNAsAlthough the primary role of lncRNAs was their epigeneticregulation of protein-coding gene expression only a limitednumber of such long noncoding transcripts have been iden-tified [21] It was soon discovered that lncRNAs play a rolein variety of biological process acting in the nucleus or inthe cytoplasm or in both [22] Functions of lncRNAs aremanifested by three types of interaction RNA-RNA RNA-DNA and RNA-protein all of which depend on the site ofaction [23]
In the nucleus for example the main role of lncRNAsis assumed to be epigenetic imprinting One of the best-described mechanisms of lncRNAs action during the epige-netic regulation is the X chromosome inactivation in whichX chromosome inactive-specific transcript (XIST) plays akey role [24] Recent studies demonstrated that lncRNAscan interact with chromatin remodeling complexes includingpolycomb repressive complex (PRC2) 2 TrithoraxMLL andH3K9 methyltransferase G9a acting as docking platforms tospecific genomic loci [4] These protein complexes modifyDNAmethylation patterns via trimethylation leading to tran-scriptional repression (PRC2) or activation (TrithoraxMLL)of the lncRNA target genes Numerous lncRNAs have been
recognized as scaffolds for inhibitory PRC2 including XISTKCNQ1OT1 andHOTAIR [8 25]Other lncRNAs likeHOT-TIP directly bind to protein complexes of TrithoraxMLL [9]In these situations lncRNAs have been shown to drive orrepress gene expression respectively Moreover it has beenpostulated that these lncRNAs are guides for chromatin-modifying complexes where they first recognize the targetlocalizations at the chromatin bind to them and then formdocking platforms for protein partners [26]
In addition to regulation of genomic imprinting lncR-NAs are also involved in the control of gene expressionat the transcriptional level lncRNAs can directly interactwith transcription factors acting as decoys or inhibitors fortheir binding to the target DNA sequence For exampletwo lncRNA molecules Lethe and p50-associated Cox-2extragenic RNA (PACER) have been shown to interact withdifferent subunits of NF-120581B thus preventing it from bindingto the promoter region of the target gene [10 27] More-over lncRNAs exert their regulatory function during mRNAprocessing and stability interacting with the heterogeneousnuclear ribonucleoproteins (hnRNPs) An example of thistype of RNA-protein linkage is demonstrated for severallncRNAs including lincRNA-p21 [28] lincRNA-Cox2 [29] orTHRIL [11] thus they affect either activation or repressionof target genes Other actions of lncRNA in the nucleusare manifested by alternative splicing regulation [12] andsubnuclear compartment formation [30] (Figure 4)
In the cytosol lncRNAs exert their function by interact-ing with target mRNAs (or miRNAs) through base-pairingIn this manner lncRNAs may either stabilize (eg BACE1-AS prevents miRNA-induced repression of BACE1 transcript[31]) or decay (eg 12-sbsRNAs bind to the Alu elementof SMD target mRNAs within the 31015840 UTR region [32])target transcripts Moreover lncRNAs promote translation oftranscripts (eg antisense Uchl1 interacts with Uchl1 mRNAresulting in recruitment of ribosomes [13]) or repress thisprocess (eg lincRNA-p21 binds to target mRNA causingrecruitment of translation repressors [14]) (Figure 4)
3 Inflammation and Its Significance inMetabolic Syndrome
Inflammation is a complex process aiming to defend thebody against harmful agents by removing or neutralizingthem in order to restore tissue homeostasis [33] The com-plexity of this process results from the broad spectrum ofinflammatory pathways various inflammatory inducers sen-sors andmediators [34] In the classic view the inflammatoryresponse is induced immediately after stimulation by theinflammatory stimuli such as a bacterial pathogen [33] Theforeign antigen is recognized by the sensors of the hostdefense cell Cells such as neutrophils dendritic cells (DCs)and macrophages express evolutionarily conserved Toll-likereceptors (TLRs) on their surface The TLR family is com-posed of several classes of transmembrane proteins and rec-ognizes specific structures present on the microbes knownas pathogen-associated molecular patterns (PAMPs) [35]Induction of TLRs triggers a multistep signaling cascadethereby resulting in the modulation of gene expression
Mediators of Inflammation 3
NoncodingRNAs
HousekeepingncRNAs
asRNAseRNAs
miRNAssiRNAspiRNAs
rRNA tRNA snRNA snoRNA
RegulatoryRNAs
ShortncRNAslt200nt
LongncRNAsgt200nt
Figure 2 Types of noncoding RNAs (ncRNAs) Noncoding RNAs are classified into housekeeping and regulatory noncoding RNAsHousekeeping ncRNAs include ribosomal (rRNA) transfer (tRNA) small nuclear (snRNA) and small nucleolar RNAs (snoRNAs)Regulatory noncoding RNAs are divided into short ncRNAs (lt200 nucleotides) or long ncRNAs (gt200 nucleotides) including microRNAs(miRNAs) small interfering RNAs (siRNAs) Piwi-associated RNAs (piRNAs) antisense RNAs (asRNAs) and enhancer RNAs (eRNAs)
BidirectionallncRNA
SenselncRNA
Intronic lncRNA
Intronic lncRNA
AntisenselncRNA
Intergenic lncRNA
Protein-coding gene
Figure 3 Classification of long noncoding RNAs lncRNAs (shownas red boxes) may be classified into five categories sense lncRNAsantisense lncRNAs bidirectional lncRNAs (transcribed on theopposite strand within 1 kb from the nearest protein-coding gene)intronic lncRNAs and intergenic lncRNA (transcribed betweenprotein-coding genes but they are located at least 1 kb away from thenearest protein-coding gene) [3] Detailed descriptions of lncRNAclasses are in the main text
involved in the immune response Key processes regulatedby TLRs are related to the secretion of various mediators byimmune cells phagocytosis cell migration metabolic repro-gramming and tissue repair [36]
In general based on the length the inflammatory res-ponse can be classified into acute or chronic Acute inflamma-tion usually develops locally and its role is to eliminate toxicagents repair damaged tissue and restore homeostasis Insome circumstances local inflammation might be greatertherefore called a ldquogeneralized inflammatory responserdquowhich is associated with the response of the whole organism
[37] Although the acute phase leads to the beneficial recoveryof the injured tissue and healing this nonspecific immuneprocess could have also negative effects Without the properresolution phase it can easily turn into a chronic stateChronic inflammation may appear after the switch from theacute phase and it is characterized by the specific long-termcellular and humoral immune response present at the site oftissue injury [38] In consequence it may significantly con-tribute to dangerous pathophysiological changes and initiatethe development of chronic diseases
Each inflammatory stage is subjected to precise controlby several transcription factors involved in the immune res-ponse [39] One such factor is NF-120581B (nuclear factor kappa-light-chain-enhancer of activated B cells) representing afamily of transcription factors NF-120581B plays an important rolein gene expression regulation of multiple factors involved inimmune acute phase and inflammatory responses [40] NF-120581B in addition to controlling the development and functionof the immune system is required in other physiologicalprocesses such as suppression of apoptosis and cell prolif-eration and differentiation In mammals the active formof NF-120581B is composed of homo- and heterodimers of theNF-120581B family members which can be divided into fivedifferent subunits p50 p52 p65 (RelA) c-Rel and RelB Allof these share homology of the specific N-terminal domainresponsible for the dimerization and binding to target DNAsequences Depending on the local cytokines produced bythe immune cells NF-120581B also controls macrophage relocal-ization activation and differentiation into two different phe-notypes proinflammatory (M1) or anti-inflammatory (M2)In response to proinflammatory cytokines NF-120581B is alsoinvolved in the activation of T lymphocytes which prolife-rate and secrete cytokines such as interferon gamma (IFN120574)interleukin 6 (IL-6) or TNF-120572The result of NF-120581B activationis stimulation of proinflammatory cytokine production by
4 Mediators of Inflammation
SRSRSR
SR
MALAT1
P PP
PP
PPP
P
P
Trx HOTTIPHOTAIR
H3K4me3H3K27me3
Nucleus CytosolPRC1
PRC2 MLL
Pre-mRNA
mRNA
(A) Epigenetic imprinting
(B) Transcription
(C) Alternative splicing
(D) miRNA sponge
(E) mRNA stabilitydegradation
(F) Translation
Uchl1 lincRNA-p21
MIAT
BACE1-AS
miR-150-5p
p50PACER
THRILhnRNP
COX-2p65
Transcriptionalactivation
Transcriptional repression
TNF-120572
Figure 4 Biological role of lncRNAs Role of lncRNAs has been implicated in the regulation of diverse processes manifested by three waysof interactions RNA-RNA RNA-DNA and RNA-protein both in the nucleus and in the cytoplasm (A) In the nucleus lncRNAs have beenshown to play a key role in imprinting control lncRNAs may act as docking platforms for the chromatin remodeling complex polycombrepressive complex (PRC2) 2 (eg HOTAIR) which methylates histone H3 at lysine 27 (H3K27me3) or with the heterogeneous group ofTrithoraxMLL proteins (eg HOTTIP) leading to a repression or gain of transcriptional activity respectively [8 9] (B) Other regulatoryroles of lncRNAs in the nucleus include transcriptional regulation by interactingwith transcription factors (eg PACERdirectly interacts withthe repressive subunit of NF-120581B p50 thus preventing it from binding to the Cox-2 promoter [10]) heterogeneous nuclear ribonucleoproteins(hnRNPs) (eg THRIL together with hnRNPL acts as RNA-protein complex and binds to TNF-120572 promoter and mediates the induction ofTNF-120572 expression [11]) (C)Moreover lncRNAsmay regulate the pre-mRNA alternative splicing (eg MALAT1) which acts as scaffold for SRproteins present in nuclear speckles andmodulates their phosphorylation [12] In the cytosol lncRNAs exert their function by interactingwithtarget transcripts and miRNAs through base-pairing (D) For miRNAs lncRNAs may act as a molecular sponge preventing specific miRNAsfrom binding to their target mRNAs (eg MIAT binds to miR-150-5p thereby upregulating the level of miR-150-5p target gene) (E) lncRNAbinding to mRNA may stabilize (eg BACE1-AS prevents miRNA-induced repression of BACE1 transcript) or decay target transcripts (F)Moreover lncRNAs promote (eg antisense Uchl1 interacts with Uchl1 mRNA resulting in recruitment of ribosomes [13]) or repress (eglincRNA-p21 which binds to target mRNA causing recruitment of translation repressors [14]) translation of transcripts This figure wasproduced using modified elements from the Servier Medical Art (httpwwwserviercom)
macrophages and T lymphocytes thereby enhancing theinflammatory response [41]
One disease closely linked to a chronic inflammatory stateis obesity It is characterized by an excessive fat accumu-lation and enhanced production of proinflammatory cyto-kineschemokines by adipose tissues It is commonly causedby a combination of excessive food intake lack of physicalactivity and genetic susceptibility [42] A great majority ofobese individuals present features of themetabolic syndrome(a) increased waist circumference (b) insulin resistance(c) hyperglycemia (d) hypertension and (e) hypertriglyc-eridemia [15] Appearance of these symptoms is associated
with a high risk of serious disease development includingtype 2 diabetes cardiovascular diseases [18] nonalcoholicfatty liver disease (NAFLD) [17] and even some cancers [19]Themechanismofmetabolic disorder development is still notfully understood nevertheless numerous studies have sug-gested an adipocyte dysfunction as a main cause of metabolicsystem failure [43] In this pathological state adipose tissuesecretes various mediators including cytokines inducing achronic low-grade inflammatory response that results inrecruitment of immune cells into the tissue
The excessive accumulation of tissue fat leads to distur-bance both in the secretory and in the storage functions of
Mediators of Inflammation 5
adipocytes It is postulated that the excessive fat mass inducesthe production of reactive oxygen species and the develop-ment of chronic oxidative stress in patients with obesity andmetabolic dysfunction [44 45] In adipose tissue of obesepatients is observed severe inflammatory response charac-terized by high production of cytokines and increased infil-tration of immune cells including monocytes stimulated todifferentiate into macrophages and lymphocytes T activatedto proliferate and secrete cytokines [46 47] Cytokinessecreted by activated macrophages act on TLRs present onthe surface of adipocytes stimulating them to produce adipo-kines and cytokines [48] Based on these observations it hasbeen found that the reactions of both the immune and meta-bolic systems are closely connected and they play an impor-tant role in the development of metabolic disorders
4 lncRNA in Obesity andAdipogenesis Control
Adipocytes play a central role in energy homeostasis byfine-tuning the equilibrium between nutrient deposition(white adipose tissue WAT) and energy expenditure (brownadipose tissue BAT) Additionally adipose tissue acts asan endocrine organ by secreting factors (adipokines) thatregulate whole body energy and glucose homeostasis [49]Adipogenesis is a complex process governed by a network oftranscription factors cofactors and signaling intermediatesfrom numerous pathways [50] The transcriptional cascadethat regulates adipocyte differentiation is predominantlydriven by peroxisome proliferator-activated receptor gamma(PPAR120574) shown to be both necessary and sufficient foradipogenesis [51] Importantly adipogenesis is highly regu-lated not only by PPAR120574 but also by the coordinated effectsof other transcription factors including CCAATenhancer-binding proteins (CEBPs) Kruppel-like factors (KLFs) orWingless proteins (Wnt) and transcriptional cofactors [52ndash54]
In recent years there has been growing attention paid tononcoding RNAs as a novel cell intrinsic regulatory mecha-nism Specifically lncRNAs are now emerging as importantregulators of gene expression both on the transcriptional andon the posttranscriptional levels Long noncoding RNAs arebeing discovered using modern techniques of microarray ornext-generation sequencing However only small portionshave been fully characterized In the case of adipogenesisthere is wide expression of lncRNAs controlling a varietyof genes involved in the formation differentiation andactivation of adipocytes (Figure 5)
Steroid receptor RNA Activator (SRA) lncRNA was des-cribed for the first time as a molecule regulating adipo-genesis [55] It was shown that lncRNA SRA initially identi-fied as a coactivator of steroid receptors also acts as a tran-scriptional coactivator of PPAR120574 [55 56] lncRNA SRA pro-motes adipocyte differentiation by binding to the N-terminalportion of PPAR120574 and enhances its transcriptional activ-ity Additionally its overexpression significantly increasedmRNA and protein levels of adipocyte master regulatorsPPAR120574 and CEBP120572 as well as PPAR120574 target genes Con-versely knockdown of endogenous lncRNA SRA resulted in
inhibition of 3T3-L1 preadipocyte differentiation Moreovergene expression profiling showed that lncRNA SRA regulatesexpression of genes related to various cellular processesincluding cell cycle and insulin transduction pathways [55]Further experiments proved that lncRNA SRA overexpres-sion inhibits phosphorylation of p38 mitogen activated pro-tein kinase (MAPK) and c-JunNH2-terminal kinase (JNK) inthe early differentiation of ST2 mesenchymal precursor cellsIn contrast knockdown of lncRNA SRA increased p38 andJNK phosphorylation while reducing insulin receptormRNAand protein levels which led to decreased downstreamsignaling [57] Supporting that data SRA knock-out mice(SRAminusminus) are resistant to high fat diet (HFD) induced obesityAfter 14 weeks of HFD SRAminusminus mice are characterized byreduced WAT mass and decreased expression of adipocytegenes such as adiponectin or fatty acid-binding protein 4Thelean phenotype is also associated with smaller adipocytes inWAT compared to WT mice reduced liver mass fewer lipiddroplets in the liver and decreased expression of lipogenesis-associated genes Finally SRAminusminus animals have improvedinsulin sensitivity [58]
In a recent paper Xiao and coworkers [59] identifiedlncRNA ADINR (adipogenic differentiation induced non-coding RNA) that in cis transcriptionally activates CEBP120572It was upregulated 20ndash30-fold during the course of adipoge-nesis and was transcribed from a position sim450 bp upstreamof the CEBP120572 gene Knockdown of lncRNA ADINR withsiRNA resulted in a dramatic adipogenic defect as shown bya decreased number of Oil Red O positive cells and reducedadipogenic transcripts CEBP120572 PPAR120574 fatty acid-bindingprotein 4 and lipoprotein lipase Importantly inhibition ofadipogenesis caused by depletion of lncRNA ADINR wasrescued by overexpression of CEBP120572 The lncRNA ADINRmechanism of action relies on its binding to PA1 followed byrecruitment ofMLL34 histonemethyltransferase complexesIn turn that causes an increase inH3K4me3 and a decrease inH3K27me3 histone modification in the CEBP120572 locus duringadipogenesis [59]
Another lncRNA that induces adipogenesis in ciswas dis-covered within themouse PU1 locus Expression of the PU1gene gives rise to both the mRNA encoding PU1 proteinand antisense (AS) lncRNA originating from a promoter inthe antisense strand of intron 3 [60] The protein PU1 wasoriginally demonstrated to be a key transcription factor regu-lating hematopoiesis but recent studies revealed that it is alsoexpressed in 3T3-L1 preadipocytes and murine adipocytesisolated from WAT [60 61] Gain and loss of functionstudies established the PU1 protein as an inhibitor of 3T3-L1 preadipocyte differentiation acting via downregulation ofPPAR120574 [62] On the other hand PU1 AS lncRNA promotesadipogenesis through preventing PU1 mRNA translationby binding to PU1 mRNA to form an mRNAAS lncRNAduplex [60] Formation of this sense-antisense RNA duplexwas also confirmed in porcine adipocytes by an endogenousribonuclease protection assay combinedwithRT-PCR In linewith the previous data expression of PPAR120574 and fatty acidsynthase (FASN)was significantly upregulated in the lncRNAPU1 shRNA treated group [63]
6 Mediators of Inflammation
Diabetic retinopathy
CVDs
Type 2 diabetes
Cancers
NAFLD
ObesitySRA
ADINRNEAT1lncRAPs
MALAT1MIAT
HI-LNC25KCNQ1OT1LOC283177
Risk factors
(i) Abdominal obesity
(ii) Insulin resistance
(iii) Hyperglycemia
(iv) Hypertriglyceridemia
(v) Hypertension
uarr 535 lncRNAsdarr 1200 lncRNAs
Figure 5 Development of metabolic syndrome The common clinical symptoms of metabolic disorders include abdominal obesity insulinresistance hyperglycemia hypertriglyceridemia and hypertension [15] Their appearance is associated with a high risk for development ofserious diseases including type 2 diabetes diabetes-related diabetic retinopathy [16] and nonalcoholic fatty liver disease (NAFLD) [17] as wellas cardiovascular diseases (CVDs) [18] and even some cancers including liver pancreas and colon [19] (CVDs and cancers are not discussedin this review) Selected lncRNAs involved in the control of metabolic system activity (eg adipogenesis SRA ADINR NEAT1 and lncRAPs)and pathology of metabolic diseases (eg type 2 diabetes HI-LNC25 KCNQ1OT1 LOC283177 diabetic retinopathy MALAT1 and MIAT)are marked with the red color In NAFLD 535 lncRNAs are upregulated and 1200 are downregulated however these sequences are not yetwell characterized [20] This figure was produced using modified elements from the Servier Medical Art (httpwwwserviercom)
In a number of studies global transcriptome profilingwasused to select specific lncRNAs either involved in adipogen-esis or characteristic for adipocytes specifically isolated fromWAT or BAT In a seminal study done by Sun and cowork-ers [64] the transcriptome of primary brown and whiteadipocytes preadipocytes and cultured adipocytes was pro-filed Scientists identified 175 lncRNAs that were specificallyregulated during adipogenesis To further validate lncRNAsthat are functionally important for adipogenesis researchersselected the top 20 lncRNA genes based on the followingcriteria (i) significant upregulation in both brown and whitefat cultures (ii) binding to PPAR120574 or CEPB120572 promoters and(iii) independent validation of adipose-specific expressionBy using RNAi-mediated loss of function the top 10 mostimportant lncRNAs were selected and called Regulated inAdiPogenesis (lncRAPs) in order to highlight their key rolein proper differentiation of adipocyte precursors [64]
In comparison to white adipocytes brown adipocyteshave a higher mitochondrial content and express uncouplingprotein 1 (UCP1) which disperses chemical energy throughheat production Brown adipose tissue helps to regulateenergy expenditure in rodents and newborn babies butit has been considered to have no physiologic relevancein adults [65 66] However recent studies demonstratedthat metabolically functional BAT is also present in adulthumans As a result an understanding of BAT physiologymight provide an effective treatment of obesity or othermetabolic disorders [67] Identification of the lncRNAs cru-cial for BAT differentiation and function was done usingwhole transcriptome RNA sequencing Next gain and loss offunction studies established lncRNA Blnc1 as a potent acti-vator of thermogenic adipocyte differentiation It functionsupon formation of RNA-protein complex with the early Bcell factor 2 (EBF2) transcription factor to stimulate BAT
Mediators of Inflammation 7
formation Additionally lncRNA Blnc1 itself is a target ofEBF2 thereby forming a feed-forward regulatory loop Onthe other hand despite its stimulatory effects on brown pre-adipocyte differentiation lncRNA Blnc1 failed to promotedifferentiation of 3T3-L1 cells into UCP1 positive adipocytes[68] Another global profiling of gene expression duringmouse brown fat cell differentiation followed by candidatelncRNAs genomic context analysis pathway analysis andgene ontology enrichment of their associated protein-codinggenes was recently described In this study scientists identi-fied three lncRNAs (Gm15051 Tmem189 and Cebpd) associ-ated with their flanking coding genes (Hoxa1 CEBP120573 andCEBP120575) that participated in adipose commitment [69]
More recently RNA-seq analysis of murine brown ingui-nal white and epididymal white fat allowed for identificationof lncRNA cluster enriched in BAT Among them thelncRNA BATE1 was induced during brown adipose tissuedifferentiation Inhibition of BATE1 led to decreased expres-sion of brown fat markers and mitochondrial genes Authorsshowed that BATE1 binds to the heterogeneous nuclear ribo-nucleoprotein U a factor also required for BAT formation[70]
Finally interactions of lncRNAs with microRNAs in thecontext of adipogenesis were also described As demonstratedby Gernapudi and coworkers [71] the miR-140lncRNANEAT1 signaling network is necessary for adipogenesisAdipocyte derived stem cells isolated from miR-140 knock-out mice had dramatically decreased adipogenic capabilitiesfound to be associated with the downregulation of lncRNANEAT1 expression Overexpression of NEAT1 in such cellswas sufficient enough to restore their differentiation capabil-ities Authors identified a miR-140 binding site in the NEAT1sequence They also found that the physical interaction withmiR-140 in nucleus led to increased expression of NEAT1[71]
Excessive fat accumulation not only causes pathologicalexpansion of adipose tissue but also leads to triglyceridedeposition in the liver Presence of lipid droplets in excessof 5 of total hepatocytes is a diagnostic hallmark of non-alcoholic fatty liver disease (NAFLD) [72] Although mostpatients suffer from a mild course of illness approximately25 of cases progress and can subsequently lead to thedevelopment of steatohepatitis hepatic fibrosis liver cirrho-sis and hepatoma [73] Scientists and physicians estimatethat between 20 and 30 of Westrsquos general populationsuffers from NAFLD which makes this affects more than abillion people worldwide [74] To address questions aboutthe global expression pattern and functional contribution oflncRNAs during NAFLD Sun and coworkers [20] analyzedmicroarray expression using RNA extracted from liver biop-sies of healthy patients and those suffering from NAFLDIn this analysis 535 lncRNAs were found upregulated inNAFLD samples compared with controls and 1200 weredownregulated in NAFLD (Figure 5) Out of these sevenNAFLD samples that had highly up- or downregulatedgene expression were validated by quantitative real-timePCR (qPCR) Three lncRNA results obtained by qPCR weredifferent in comparison to the microarray data showing nochange between analyzed groups However changes in other
selected hits were confirmed highlighting the importance ofmicroarray data validation [20]
5 lncRNA in Diabetes and PancreaticFunctionGlucose Homeostasis
Pancreatic islets through the secretion of endocrinehormones such as insulin and glucagon play a key role inmetabolic homeostasis However one of the most commonmalfunctions of 120573-cells is caused by lack of insulin leading tothe development of diabetes [75] Additionally diabetes maydevelop as a result of insulin resistance a condition in whichthe body cannot effectively use the insulin produced by 120573-cells As a result blood sugar levels are deregulated and vari-ous tissues are exposed to prolonged hyperglycemia that overtime causes serious damage to many of the bodyrsquos systemsIt should be emphasized that hyperglycemia a hallmark ofdiabetes can cause both acute and long-term complicationslike cardiovascular disease stroke foot ulcers ketoacidosisor hyperosmolar coma [76] Based on etiology WHOclassifies diabetes into four categories but the vast majorityof cases fall into two broad etiopathogenetic groups type 1diabetes mellitus (T1DM) or type 2 diabetes mellitus (T2DM)[77]
Accounting for 5ndash10 of all cases T1DM is characterizedby the loss of insulin producing120573-cells due to an autoimmunereaction Autoimmune destruction of 120573-cells is unrelated tolifestyleThere are genetic variants known to increase the riskof T1DM development Type 2 diabetes mellitus represents90ndash95 of all cases and unlike T1DM is strongly associatedwith patientsrsquo lifestyle [78] A number of lifestyle factors areknown to be important for development of T2DM includingobesity (BMI above 30) lack of physical activity poor dietor stress [79] In fact most patients suffering from T2DM areobese It is a chronic disease that begins with insulin resist-ance a condition in which cells (mostly adipocytes hepato-cytes andmuscle cells) fail to properly absorb andmetabolizeglucose As a result 120573-cells increase insulin production toovercome hyperglycemia but this compensatory mechanismis transient and generally fails over time [77] Intensifiedmetabolism of 120573-cells leads to a gradual loss of their endo-crine function and finally causes 120573-cell apoptosis Similar toobesity diabetes is a worldwide problemAccording toWHOin 2014 the global prevalence of diabetes was estimated tobe 9 among adults aged over 18 years and it is expectedto double before 2030 [75 80] Additionally experts projectthat in 2030 diabetes will be the 7th leading cause of death[75]
Initial studies aimed at identification of specific lncRNAsexpressed in murine and human pancreatic islets were donein 2012 [81 82] By using next-generation sequencing scien-tists revealed a large collection of 1359 intergenic lncRNAsexpressed in murine 120573-cells (Figure 5) Many of them werehighly tissue-specific but their function was not evaluatedand future experiments are still needed [81] Similar numbersof 1128 intergenic and antisense lncRNAs were identifiedin human 120573-cells and many of them (eg HI-LNC12 andHI-LNC77) were highly tissue-specific [82] AdditionallylncRNA-encoding genes were preferentially located near
8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 3
NoncodingRNAs
HousekeepingncRNAs
asRNAseRNAs
miRNAssiRNAspiRNAs
rRNA tRNA snRNA snoRNA
RegulatoryRNAs
ShortncRNAslt200nt
LongncRNAsgt200nt
Figure 2 Types of noncoding RNAs (ncRNAs) Noncoding RNAs are classified into housekeeping and regulatory noncoding RNAsHousekeeping ncRNAs include ribosomal (rRNA) transfer (tRNA) small nuclear (snRNA) and small nucleolar RNAs (snoRNAs)Regulatory noncoding RNAs are divided into short ncRNAs (lt200 nucleotides) or long ncRNAs (gt200 nucleotides) including microRNAs(miRNAs) small interfering RNAs (siRNAs) Piwi-associated RNAs (piRNAs) antisense RNAs (asRNAs) and enhancer RNAs (eRNAs)
BidirectionallncRNA
SenselncRNA
Intronic lncRNA
Intronic lncRNA
AntisenselncRNA
Intergenic lncRNA
Protein-coding gene
Figure 3 Classification of long noncoding RNAs lncRNAs (shownas red boxes) may be classified into five categories sense lncRNAsantisense lncRNAs bidirectional lncRNAs (transcribed on theopposite strand within 1 kb from the nearest protein-coding gene)intronic lncRNAs and intergenic lncRNA (transcribed betweenprotein-coding genes but they are located at least 1 kb away from thenearest protein-coding gene) [3] Detailed descriptions of lncRNAclasses are in the main text
involved in the immune response Key processes regulatedby TLRs are related to the secretion of various mediators byimmune cells phagocytosis cell migration metabolic repro-gramming and tissue repair [36]
In general based on the length the inflammatory res-ponse can be classified into acute or chronic Acute inflamma-tion usually develops locally and its role is to eliminate toxicagents repair damaged tissue and restore homeostasis Insome circumstances local inflammation might be greatertherefore called a ldquogeneralized inflammatory responserdquowhich is associated with the response of the whole organism
[37] Although the acute phase leads to the beneficial recoveryof the injured tissue and healing this nonspecific immuneprocess could have also negative effects Without the properresolution phase it can easily turn into a chronic stateChronic inflammation may appear after the switch from theacute phase and it is characterized by the specific long-termcellular and humoral immune response present at the site oftissue injury [38] In consequence it may significantly con-tribute to dangerous pathophysiological changes and initiatethe development of chronic diseases
Each inflammatory stage is subjected to precise controlby several transcription factors involved in the immune res-ponse [39] One such factor is NF-120581B (nuclear factor kappa-light-chain-enhancer of activated B cells) representing afamily of transcription factors NF-120581B plays an important rolein gene expression regulation of multiple factors involved inimmune acute phase and inflammatory responses [40] NF-120581B in addition to controlling the development and functionof the immune system is required in other physiologicalprocesses such as suppression of apoptosis and cell prolif-eration and differentiation In mammals the active formof NF-120581B is composed of homo- and heterodimers of theNF-120581B family members which can be divided into fivedifferent subunits p50 p52 p65 (RelA) c-Rel and RelB Allof these share homology of the specific N-terminal domainresponsible for the dimerization and binding to target DNAsequences Depending on the local cytokines produced bythe immune cells NF-120581B also controls macrophage relocal-ization activation and differentiation into two different phe-notypes proinflammatory (M1) or anti-inflammatory (M2)In response to proinflammatory cytokines NF-120581B is alsoinvolved in the activation of T lymphocytes which prolife-rate and secrete cytokines such as interferon gamma (IFN120574)interleukin 6 (IL-6) or TNF-120572The result of NF-120581B activationis stimulation of proinflammatory cytokine production by
4 Mediators of Inflammation
SRSRSR
SR
MALAT1
P PP
PP
PPP
P
P
Trx HOTTIPHOTAIR
H3K4me3H3K27me3
Nucleus CytosolPRC1
PRC2 MLL
Pre-mRNA
mRNA
(A) Epigenetic imprinting
(B) Transcription
(C) Alternative splicing
(D) miRNA sponge
(E) mRNA stabilitydegradation
(F) Translation
Uchl1 lincRNA-p21
MIAT
BACE1-AS
miR-150-5p
p50PACER
THRILhnRNP
COX-2p65
Transcriptionalactivation
Transcriptional repression
TNF-120572
Figure 4 Biological role of lncRNAs Role of lncRNAs has been implicated in the regulation of diverse processes manifested by three waysof interactions RNA-RNA RNA-DNA and RNA-protein both in the nucleus and in the cytoplasm (A) In the nucleus lncRNAs have beenshown to play a key role in imprinting control lncRNAs may act as docking platforms for the chromatin remodeling complex polycombrepressive complex (PRC2) 2 (eg HOTAIR) which methylates histone H3 at lysine 27 (H3K27me3) or with the heterogeneous group ofTrithoraxMLL proteins (eg HOTTIP) leading to a repression or gain of transcriptional activity respectively [8 9] (B) Other regulatoryroles of lncRNAs in the nucleus include transcriptional regulation by interactingwith transcription factors (eg PACERdirectly interacts withthe repressive subunit of NF-120581B p50 thus preventing it from binding to the Cox-2 promoter [10]) heterogeneous nuclear ribonucleoproteins(hnRNPs) (eg THRIL together with hnRNPL acts as RNA-protein complex and binds to TNF-120572 promoter and mediates the induction ofTNF-120572 expression [11]) (C)Moreover lncRNAsmay regulate the pre-mRNA alternative splicing (eg MALAT1) which acts as scaffold for SRproteins present in nuclear speckles andmodulates their phosphorylation [12] In the cytosol lncRNAs exert their function by interactingwithtarget transcripts and miRNAs through base-pairing (D) For miRNAs lncRNAs may act as a molecular sponge preventing specific miRNAsfrom binding to their target mRNAs (eg MIAT binds to miR-150-5p thereby upregulating the level of miR-150-5p target gene) (E) lncRNAbinding to mRNA may stabilize (eg BACE1-AS prevents miRNA-induced repression of BACE1 transcript) or decay target transcripts (F)Moreover lncRNAs promote (eg antisense Uchl1 interacts with Uchl1 mRNA resulting in recruitment of ribosomes [13]) or repress (eglincRNA-p21 which binds to target mRNA causing recruitment of translation repressors [14]) translation of transcripts This figure wasproduced using modified elements from the Servier Medical Art (httpwwwserviercom)
macrophages and T lymphocytes thereby enhancing theinflammatory response [41]
One disease closely linked to a chronic inflammatory stateis obesity It is characterized by an excessive fat accumu-lation and enhanced production of proinflammatory cyto-kineschemokines by adipose tissues It is commonly causedby a combination of excessive food intake lack of physicalactivity and genetic susceptibility [42] A great majority ofobese individuals present features of themetabolic syndrome(a) increased waist circumference (b) insulin resistance(c) hyperglycemia (d) hypertension and (e) hypertriglyc-eridemia [15] Appearance of these symptoms is associated
with a high risk of serious disease development includingtype 2 diabetes cardiovascular diseases [18] nonalcoholicfatty liver disease (NAFLD) [17] and even some cancers [19]Themechanismofmetabolic disorder development is still notfully understood nevertheless numerous studies have sug-gested an adipocyte dysfunction as a main cause of metabolicsystem failure [43] In this pathological state adipose tissuesecretes various mediators including cytokines inducing achronic low-grade inflammatory response that results inrecruitment of immune cells into the tissue
The excessive accumulation of tissue fat leads to distur-bance both in the secretory and in the storage functions of
Mediators of Inflammation 5
adipocytes It is postulated that the excessive fat mass inducesthe production of reactive oxygen species and the develop-ment of chronic oxidative stress in patients with obesity andmetabolic dysfunction [44 45] In adipose tissue of obesepatients is observed severe inflammatory response charac-terized by high production of cytokines and increased infil-tration of immune cells including monocytes stimulated todifferentiate into macrophages and lymphocytes T activatedto proliferate and secrete cytokines [46 47] Cytokinessecreted by activated macrophages act on TLRs present onthe surface of adipocytes stimulating them to produce adipo-kines and cytokines [48] Based on these observations it hasbeen found that the reactions of both the immune and meta-bolic systems are closely connected and they play an impor-tant role in the development of metabolic disorders
4 lncRNA in Obesity andAdipogenesis Control
Adipocytes play a central role in energy homeostasis byfine-tuning the equilibrium between nutrient deposition(white adipose tissue WAT) and energy expenditure (brownadipose tissue BAT) Additionally adipose tissue acts asan endocrine organ by secreting factors (adipokines) thatregulate whole body energy and glucose homeostasis [49]Adipogenesis is a complex process governed by a network oftranscription factors cofactors and signaling intermediatesfrom numerous pathways [50] The transcriptional cascadethat regulates adipocyte differentiation is predominantlydriven by peroxisome proliferator-activated receptor gamma(PPAR120574) shown to be both necessary and sufficient foradipogenesis [51] Importantly adipogenesis is highly regu-lated not only by PPAR120574 but also by the coordinated effectsof other transcription factors including CCAATenhancer-binding proteins (CEBPs) Kruppel-like factors (KLFs) orWingless proteins (Wnt) and transcriptional cofactors [52ndash54]
In recent years there has been growing attention paid tononcoding RNAs as a novel cell intrinsic regulatory mecha-nism Specifically lncRNAs are now emerging as importantregulators of gene expression both on the transcriptional andon the posttranscriptional levels Long noncoding RNAs arebeing discovered using modern techniques of microarray ornext-generation sequencing However only small portionshave been fully characterized In the case of adipogenesisthere is wide expression of lncRNAs controlling a varietyof genes involved in the formation differentiation andactivation of adipocytes (Figure 5)
Steroid receptor RNA Activator (SRA) lncRNA was des-cribed for the first time as a molecule regulating adipo-genesis [55] It was shown that lncRNA SRA initially identi-fied as a coactivator of steroid receptors also acts as a tran-scriptional coactivator of PPAR120574 [55 56] lncRNA SRA pro-motes adipocyte differentiation by binding to the N-terminalportion of PPAR120574 and enhances its transcriptional activ-ity Additionally its overexpression significantly increasedmRNA and protein levels of adipocyte master regulatorsPPAR120574 and CEBP120572 as well as PPAR120574 target genes Con-versely knockdown of endogenous lncRNA SRA resulted in
inhibition of 3T3-L1 preadipocyte differentiation Moreovergene expression profiling showed that lncRNA SRA regulatesexpression of genes related to various cellular processesincluding cell cycle and insulin transduction pathways [55]Further experiments proved that lncRNA SRA overexpres-sion inhibits phosphorylation of p38 mitogen activated pro-tein kinase (MAPK) and c-JunNH2-terminal kinase (JNK) inthe early differentiation of ST2 mesenchymal precursor cellsIn contrast knockdown of lncRNA SRA increased p38 andJNK phosphorylation while reducing insulin receptormRNAand protein levels which led to decreased downstreamsignaling [57] Supporting that data SRA knock-out mice(SRAminusminus) are resistant to high fat diet (HFD) induced obesityAfter 14 weeks of HFD SRAminusminus mice are characterized byreduced WAT mass and decreased expression of adipocytegenes such as adiponectin or fatty acid-binding protein 4Thelean phenotype is also associated with smaller adipocytes inWAT compared to WT mice reduced liver mass fewer lipiddroplets in the liver and decreased expression of lipogenesis-associated genes Finally SRAminusminus animals have improvedinsulin sensitivity [58]
In a recent paper Xiao and coworkers [59] identifiedlncRNA ADINR (adipogenic differentiation induced non-coding RNA) that in cis transcriptionally activates CEBP120572It was upregulated 20ndash30-fold during the course of adipoge-nesis and was transcribed from a position sim450 bp upstreamof the CEBP120572 gene Knockdown of lncRNA ADINR withsiRNA resulted in a dramatic adipogenic defect as shown bya decreased number of Oil Red O positive cells and reducedadipogenic transcripts CEBP120572 PPAR120574 fatty acid-bindingprotein 4 and lipoprotein lipase Importantly inhibition ofadipogenesis caused by depletion of lncRNA ADINR wasrescued by overexpression of CEBP120572 The lncRNA ADINRmechanism of action relies on its binding to PA1 followed byrecruitment ofMLL34 histonemethyltransferase complexesIn turn that causes an increase inH3K4me3 and a decrease inH3K27me3 histone modification in the CEBP120572 locus duringadipogenesis [59]
Another lncRNA that induces adipogenesis in ciswas dis-covered within themouse PU1 locus Expression of the PU1gene gives rise to both the mRNA encoding PU1 proteinand antisense (AS) lncRNA originating from a promoter inthe antisense strand of intron 3 [60] The protein PU1 wasoriginally demonstrated to be a key transcription factor regu-lating hematopoiesis but recent studies revealed that it is alsoexpressed in 3T3-L1 preadipocytes and murine adipocytesisolated from WAT [60 61] Gain and loss of functionstudies established the PU1 protein as an inhibitor of 3T3-L1 preadipocyte differentiation acting via downregulation ofPPAR120574 [62] On the other hand PU1 AS lncRNA promotesadipogenesis through preventing PU1 mRNA translationby binding to PU1 mRNA to form an mRNAAS lncRNAduplex [60] Formation of this sense-antisense RNA duplexwas also confirmed in porcine adipocytes by an endogenousribonuclease protection assay combinedwithRT-PCR In linewith the previous data expression of PPAR120574 and fatty acidsynthase (FASN)was significantly upregulated in the lncRNAPU1 shRNA treated group [63]
6 Mediators of Inflammation
Diabetic retinopathy
CVDs
Type 2 diabetes
Cancers
NAFLD
ObesitySRA
ADINRNEAT1lncRAPs
MALAT1MIAT
HI-LNC25KCNQ1OT1LOC283177
Risk factors
(i) Abdominal obesity
(ii) Insulin resistance
(iii) Hyperglycemia
(iv) Hypertriglyceridemia
(v) Hypertension
uarr 535 lncRNAsdarr 1200 lncRNAs
Figure 5 Development of metabolic syndrome The common clinical symptoms of metabolic disorders include abdominal obesity insulinresistance hyperglycemia hypertriglyceridemia and hypertension [15] Their appearance is associated with a high risk for development ofserious diseases including type 2 diabetes diabetes-related diabetic retinopathy [16] and nonalcoholic fatty liver disease (NAFLD) [17] as wellas cardiovascular diseases (CVDs) [18] and even some cancers including liver pancreas and colon [19] (CVDs and cancers are not discussedin this review) Selected lncRNAs involved in the control of metabolic system activity (eg adipogenesis SRA ADINR NEAT1 and lncRAPs)and pathology of metabolic diseases (eg type 2 diabetes HI-LNC25 KCNQ1OT1 LOC283177 diabetic retinopathy MALAT1 and MIAT)are marked with the red color In NAFLD 535 lncRNAs are upregulated and 1200 are downregulated however these sequences are not yetwell characterized [20] This figure was produced using modified elements from the Servier Medical Art (httpwwwserviercom)
In a number of studies global transcriptome profilingwasused to select specific lncRNAs either involved in adipogen-esis or characteristic for adipocytes specifically isolated fromWAT or BAT In a seminal study done by Sun and cowork-ers [64] the transcriptome of primary brown and whiteadipocytes preadipocytes and cultured adipocytes was pro-filed Scientists identified 175 lncRNAs that were specificallyregulated during adipogenesis To further validate lncRNAsthat are functionally important for adipogenesis researchersselected the top 20 lncRNA genes based on the followingcriteria (i) significant upregulation in both brown and whitefat cultures (ii) binding to PPAR120574 or CEPB120572 promoters and(iii) independent validation of adipose-specific expressionBy using RNAi-mediated loss of function the top 10 mostimportant lncRNAs were selected and called Regulated inAdiPogenesis (lncRAPs) in order to highlight their key rolein proper differentiation of adipocyte precursors [64]
In comparison to white adipocytes brown adipocyteshave a higher mitochondrial content and express uncouplingprotein 1 (UCP1) which disperses chemical energy throughheat production Brown adipose tissue helps to regulateenergy expenditure in rodents and newborn babies butit has been considered to have no physiologic relevancein adults [65 66] However recent studies demonstratedthat metabolically functional BAT is also present in adulthumans As a result an understanding of BAT physiologymight provide an effective treatment of obesity or othermetabolic disorders [67] Identification of the lncRNAs cru-cial for BAT differentiation and function was done usingwhole transcriptome RNA sequencing Next gain and loss offunction studies established lncRNA Blnc1 as a potent acti-vator of thermogenic adipocyte differentiation It functionsupon formation of RNA-protein complex with the early Bcell factor 2 (EBF2) transcription factor to stimulate BAT
Mediators of Inflammation 7
formation Additionally lncRNA Blnc1 itself is a target ofEBF2 thereby forming a feed-forward regulatory loop Onthe other hand despite its stimulatory effects on brown pre-adipocyte differentiation lncRNA Blnc1 failed to promotedifferentiation of 3T3-L1 cells into UCP1 positive adipocytes[68] Another global profiling of gene expression duringmouse brown fat cell differentiation followed by candidatelncRNAs genomic context analysis pathway analysis andgene ontology enrichment of their associated protein-codinggenes was recently described In this study scientists identi-fied three lncRNAs (Gm15051 Tmem189 and Cebpd) associ-ated with their flanking coding genes (Hoxa1 CEBP120573 andCEBP120575) that participated in adipose commitment [69]
More recently RNA-seq analysis of murine brown ingui-nal white and epididymal white fat allowed for identificationof lncRNA cluster enriched in BAT Among them thelncRNA BATE1 was induced during brown adipose tissuedifferentiation Inhibition of BATE1 led to decreased expres-sion of brown fat markers and mitochondrial genes Authorsshowed that BATE1 binds to the heterogeneous nuclear ribo-nucleoprotein U a factor also required for BAT formation[70]
Finally interactions of lncRNAs with microRNAs in thecontext of adipogenesis were also described As demonstratedby Gernapudi and coworkers [71] the miR-140lncRNANEAT1 signaling network is necessary for adipogenesisAdipocyte derived stem cells isolated from miR-140 knock-out mice had dramatically decreased adipogenic capabilitiesfound to be associated with the downregulation of lncRNANEAT1 expression Overexpression of NEAT1 in such cellswas sufficient enough to restore their differentiation capabil-ities Authors identified a miR-140 binding site in the NEAT1sequence They also found that the physical interaction withmiR-140 in nucleus led to increased expression of NEAT1[71]
Excessive fat accumulation not only causes pathologicalexpansion of adipose tissue but also leads to triglyceridedeposition in the liver Presence of lipid droplets in excessof 5 of total hepatocytes is a diagnostic hallmark of non-alcoholic fatty liver disease (NAFLD) [72] Although mostpatients suffer from a mild course of illness approximately25 of cases progress and can subsequently lead to thedevelopment of steatohepatitis hepatic fibrosis liver cirrho-sis and hepatoma [73] Scientists and physicians estimatethat between 20 and 30 of Westrsquos general populationsuffers from NAFLD which makes this affects more than abillion people worldwide [74] To address questions aboutthe global expression pattern and functional contribution oflncRNAs during NAFLD Sun and coworkers [20] analyzedmicroarray expression using RNA extracted from liver biop-sies of healthy patients and those suffering from NAFLDIn this analysis 535 lncRNAs were found upregulated inNAFLD samples compared with controls and 1200 weredownregulated in NAFLD (Figure 5) Out of these sevenNAFLD samples that had highly up- or downregulatedgene expression were validated by quantitative real-timePCR (qPCR) Three lncRNA results obtained by qPCR weredifferent in comparison to the microarray data showing nochange between analyzed groups However changes in other
selected hits were confirmed highlighting the importance ofmicroarray data validation [20]
5 lncRNA in Diabetes and PancreaticFunctionGlucose Homeostasis
Pancreatic islets through the secretion of endocrinehormones such as insulin and glucagon play a key role inmetabolic homeostasis However one of the most commonmalfunctions of 120573-cells is caused by lack of insulin leading tothe development of diabetes [75] Additionally diabetes maydevelop as a result of insulin resistance a condition in whichthe body cannot effectively use the insulin produced by 120573-cells As a result blood sugar levels are deregulated and vari-ous tissues are exposed to prolonged hyperglycemia that overtime causes serious damage to many of the bodyrsquos systemsIt should be emphasized that hyperglycemia a hallmark ofdiabetes can cause both acute and long-term complicationslike cardiovascular disease stroke foot ulcers ketoacidosisor hyperosmolar coma [76] Based on etiology WHOclassifies diabetes into four categories but the vast majorityof cases fall into two broad etiopathogenetic groups type 1diabetes mellitus (T1DM) or type 2 diabetes mellitus (T2DM)[77]
Accounting for 5ndash10 of all cases T1DM is characterizedby the loss of insulin producing120573-cells due to an autoimmunereaction Autoimmune destruction of 120573-cells is unrelated tolifestyleThere are genetic variants known to increase the riskof T1DM development Type 2 diabetes mellitus represents90ndash95 of all cases and unlike T1DM is strongly associatedwith patientsrsquo lifestyle [78] A number of lifestyle factors areknown to be important for development of T2DM includingobesity (BMI above 30) lack of physical activity poor dietor stress [79] In fact most patients suffering from T2DM areobese It is a chronic disease that begins with insulin resist-ance a condition in which cells (mostly adipocytes hepato-cytes andmuscle cells) fail to properly absorb andmetabolizeglucose As a result 120573-cells increase insulin production toovercome hyperglycemia but this compensatory mechanismis transient and generally fails over time [77] Intensifiedmetabolism of 120573-cells leads to a gradual loss of their endo-crine function and finally causes 120573-cell apoptosis Similar toobesity diabetes is a worldwide problemAccording toWHOin 2014 the global prevalence of diabetes was estimated tobe 9 among adults aged over 18 years and it is expectedto double before 2030 [75 80] Additionally experts projectthat in 2030 diabetes will be the 7th leading cause of death[75]
Initial studies aimed at identification of specific lncRNAsexpressed in murine and human pancreatic islets were donein 2012 [81 82] By using next-generation sequencing scien-tists revealed a large collection of 1359 intergenic lncRNAsexpressed in murine 120573-cells (Figure 5) Many of them werehighly tissue-specific but their function was not evaluatedand future experiments are still needed [81] Similar numbersof 1128 intergenic and antisense lncRNAs were identifiedin human 120573-cells and many of them (eg HI-LNC12 andHI-LNC77) were highly tissue-specific [82] AdditionallylncRNA-encoding genes were preferentially located near
8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
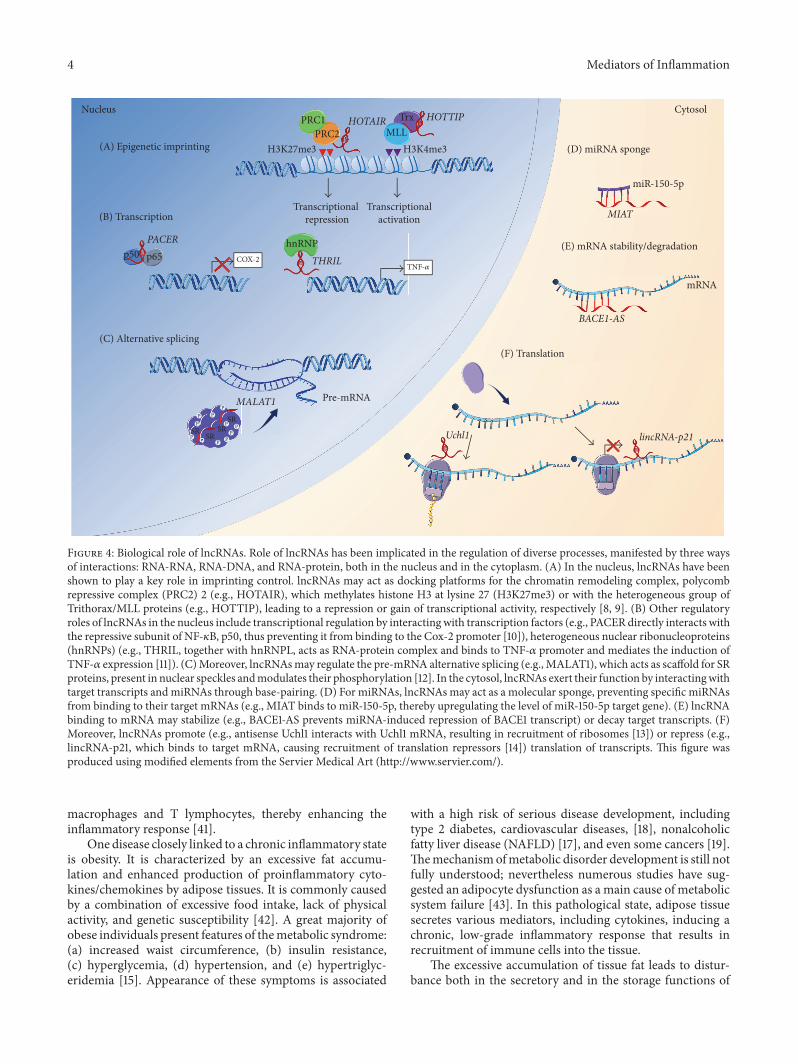
4 Mediators of Inflammation
SRSRSR
SR
MALAT1
P PP
PP
PPP
P
P
Trx HOTTIPHOTAIR
H3K4me3H3K27me3
Nucleus CytosolPRC1
PRC2 MLL
Pre-mRNA
mRNA
(A) Epigenetic imprinting
(B) Transcription
(C) Alternative splicing
(D) miRNA sponge
(E) mRNA stabilitydegradation
(F) Translation
Uchl1 lincRNA-p21
MIAT
BACE1-AS
miR-150-5p
p50PACER
THRILhnRNP
COX-2p65
Transcriptionalactivation
Transcriptional repression
TNF-120572
Figure 4 Biological role of lncRNAs Role of lncRNAs has been implicated in the regulation of diverse processes manifested by three waysof interactions RNA-RNA RNA-DNA and RNA-protein both in the nucleus and in the cytoplasm (A) In the nucleus lncRNAs have beenshown to play a key role in imprinting control lncRNAs may act as docking platforms for the chromatin remodeling complex polycombrepressive complex (PRC2) 2 (eg HOTAIR) which methylates histone H3 at lysine 27 (H3K27me3) or with the heterogeneous group ofTrithoraxMLL proteins (eg HOTTIP) leading to a repression or gain of transcriptional activity respectively [8 9] (B) Other regulatoryroles of lncRNAs in the nucleus include transcriptional regulation by interactingwith transcription factors (eg PACERdirectly interacts withthe repressive subunit of NF-120581B p50 thus preventing it from binding to the Cox-2 promoter [10]) heterogeneous nuclear ribonucleoproteins(hnRNPs) (eg THRIL together with hnRNPL acts as RNA-protein complex and binds to TNF-120572 promoter and mediates the induction ofTNF-120572 expression [11]) (C)Moreover lncRNAsmay regulate the pre-mRNA alternative splicing (eg MALAT1) which acts as scaffold for SRproteins present in nuclear speckles andmodulates their phosphorylation [12] In the cytosol lncRNAs exert their function by interactingwithtarget transcripts and miRNAs through base-pairing (D) For miRNAs lncRNAs may act as a molecular sponge preventing specific miRNAsfrom binding to their target mRNAs (eg MIAT binds to miR-150-5p thereby upregulating the level of miR-150-5p target gene) (E) lncRNAbinding to mRNA may stabilize (eg BACE1-AS prevents miRNA-induced repression of BACE1 transcript) or decay target transcripts (F)Moreover lncRNAs promote (eg antisense Uchl1 interacts with Uchl1 mRNA resulting in recruitment of ribosomes [13]) or repress (eglincRNA-p21 which binds to target mRNA causing recruitment of translation repressors [14]) translation of transcripts This figure wasproduced using modified elements from the Servier Medical Art (httpwwwserviercom)
macrophages and T lymphocytes thereby enhancing theinflammatory response [41]
One disease closely linked to a chronic inflammatory stateis obesity It is characterized by an excessive fat accumu-lation and enhanced production of proinflammatory cyto-kineschemokines by adipose tissues It is commonly causedby a combination of excessive food intake lack of physicalactivity and genetic susceptibility [42] A great majority ofobese individuals present features of themetabolic syndrome(a) increased waist circumference (b) insulin resistance(c) hyperglycemia (d) hypertension and (e) hypertriglyc-eridemia [15] Appearance of these symptoms is associated
with a high risk of serious disease development includingtype 2 diabetes cardiovascular diseases [18] nonalcoholicfatty liver disease (NAFLD) [17] and even some cancers [19]Themechanismofmetabolic disorder development is still notfully understood nevertheless numerous studies have sug-gested an adipocyte dysfunction as a main cause of metabolicsystem failure [43] In this pathological state adipose tissuesecretes various mediators including cytokines inducing achronic low-grade inflammatory response that results inrecruitment of immune cells into the tissue
The excessive accumulation of tissue fat leads to distur-bance both in the secretory and in the storage functions of
Mediators of Inflammation 5
adipocytes It is postulated that the excessive fat mass inducesthe production of reactive oxygen species and the develop-ment of chronic oxidative stress in patients with obesity andmetabolic dysfunction [44 45] In adipose tissue of obesepatients is observed severe inflammatory response charac-terized by high production of cytokines and increased infil-tration of immune cells including monocytes stimulated todifferentiate into macrophages and lymphocytes T activatedto proliferate and secrete cytokines [46 47] Cytokinessecreted by activated macrophages act on TLRs present onthe surface of adipocytes stimulating them to produce adipo-kines and cytokines [48] Based on these observations it hasbeen found that the reactions of both the immune and meta-bolic systems are closely connected and they play an impor-tant role in the development of metabolic disorders
4 lncRNA in Obesity andAdipogenesis Control
Adipocytes play a central role in energy homeostasis byfine-tuning the equilibrium between nutrient deposition(white adipose tissue WAT) and energy expenditure (brownadipose tissue BAT) Additionally adipose tissue acts asan endocrine organ by secreting factors (adipokines) thatregulate whole body energy and glucose homeostasis [49]Adipogenesis is a complex process governed by a network oftranscription factors cofactors and signaling intermediatesfrom numerous pathways [50] The transcriptional cascadethat regulates adipocyte differentiation is predominantlydriven by peroxisome proliferator-activated receptor gamma(PPAR120574) shown to be both necessary and sufficient foradipogenesis [51] Importantly adipogenesis is highly regu-lated not only by PPAR120574 but also by the coordinated effectsof other transcription factors including CCAATenhancer-binding proteins (CEBPs) Kruppel-like factors (KLFs) orWingless proteins (Wnt) and transcriptional cofactors [52ndash54]
In recent years there has been growing attention paid tononcoding RNAs as a novel cell intrinsic regulatory mecha-nism Specifically lncRNAs are now emerging as importantregulators of gene expression both on the transcriptional andon the posttranscriptional levels Long noncoding RNAs arebeing discovered using modern techniques of microarray ornext-generation sequencing However only small portionshave been fully characterized In the case of adipogenesisthere is wide expression of lncRNAs controlling a varietyof genes involved in the formation differentiation andactivation of adipocytes (Figure 5)
Steroid receptor RNA Activator (SRA) lncRNA was des-cribed for the first time as a molecule regulating adipo-genesis [55] It was shown that lncRNA SRA initially identi-fied as a coactivator of steroid receptors also acts as a tran-scriptional coactivator of PPAR120574 [55 56] lncRNA SRA pro-motes adipocyte differentiation by binding to the N-terminalportion of PPAR120574 and enhances its transcriptional activ-ity Additionally its overexpression significantly increasedmRNA and protein levels of adipocyte master regulatorsPPAR120574 and CEBP120572 as well as PPAR120574 target genes Con-versely knockdown of endogenous lncRNA SRA resulted in
inhibition of 3T3-L1 preadipocyte differentiation Moreovergene expression profiling showed that lncRNA SRA regulatesexpression of genes related to various cellular processesincluding cell cycle and insulin transduction pathways [55]Further experiments proved that lncRNA SRA overexpres-sion inhibits phosphorylation of p38 mitogen activated pro-tein kinase (MAPK) and c-JunNH2-terminal kinase (JNK) inthe early differentiation of ST2 mesenchymal precursor cellsIn contrast knockdown of lncRNA SRA increased p38 andJNK phosphorylation while reducing insulin receptormRNAand protein levels which led to decreased downstreamsignaling [57] Supporting that data SRA knock-out mice(SRAminusminus) are resistant to high fat diet (HFD) induced obesityAfter 14 weeks of HFD SRAminusminus mice are characterized byreduced WAT mass and decreased expression of adipocytegenes such as adiponectin or fatty acid-binding protein 4Thelean phenotype is also associated with smaller adipocytes inWAT compared to WT mice reduced liver mass fewer lipiddroplets in the liver and decreased expression of lipogenesis-associated genes Finally SRAminusminus animals have improvedinsulin sensitivity [58]
In a recent paper Xiao and coworkers [59] identifiedlncRNA ADINR (adipogenic differentiation induced non-coding RNA) that in cis transcriptionally activates CEBP120572It was upregulated 20ndash30-fold during the course of adipoge-nesis and was transcribed from a position sim450 bp upstreamof the CEBP120572 gene Knockdown of lncRNA ADINR withsiRNA resulted in a dramatic adipogenic defect as shown bya decreased number of Oil Red O positive cells and reducedadipogenic transcripts CEBP120572 PPAR120574 fatty acid-bindingprotein 4 and lipoprotein lipase Importantly inhibition ofadipogenesis caused by depletion of lncRNA ADINR wasrescued by overexpression of CEBP120572 The lncRNA ADINRmechanism of action relies on its binding to PA1 followed byrecruitment ofMLL34 histonemethyltransferase complexesIn turn that causes an increase inH3K4me3 and a decrease inH3K27me3 histone modification in the CEBP120572 locus duringadipogenesis [59]
Another lncRNA that induces adipogenesis in ciswas dis-covered within themouse PU1 locus Expression of the PU1gene gives rise to both the mRNA encoding PU1 proteinand antisense (AS) lncRNA originating from a promoter inthe antisense strand of intron 3 [60] The protein PU1 wasoriginally demonstrated to be a key transcription factor regu-lating hematopoiesis but recent studies revealed that it is alsoexpressed in 3T3-L1 preadipocytes and murine adipocytesisolated from WAT [60 61] Gain and loss of functionstudies established the PU1 protein as an inhibitor of 3T3-L1 preadipocyte differentiation acting via downregulation ofPPAR120574 [62] On the other hand PU1 AS lncRNA promotesadipogenesis through preventing PU1 mRNA translationby binding to PU1 mRNA to form an mRNAAS lncRNAduplex [60] Formation of this sense-antisense RNA duplexwas also confirmed in porcine adipocytes by an endogenousribonuclease protection assay combinedwithRT-PCR In linewith the previous data expression of PPAR120574 and fatty acidsynthase (FASN)was significantly upregulated in the lncRNAPU1 shRNA treated group [63]
6 Mediators of Inflammation
Diabetic retinopathy
CVDs
Type 2 diabetes
Cancers
NAFLD
ObesitySRA
ADINRNEAT1lncRAPs
MALAT1MIAT
HI-LNC25KCNQ1OT1LOC283177
Risk factors
(i) Abdominal obesity
(ii) Insulin resistance
(iii) Hyperglycemia
(iv) Hypertriglyceridemia
(v) Hypertension
uarr 535 lncRNAsdarr 1200 lncRNAs
Figure 5 Development of metabolic syndrome The common clinical symptoms of metabolic disorders include abdominal obesity insulinresistance hyperglycemia hypertriglyceridemia and hypertension [15] Their appearance is associated with a high risk for development ofserious diseases including type 2 diabetes diabetes-related diabetic retinopathy [16] and nonalcoholic fatty liver disease (NAFLD) [17] as wellas cardiovascular diseases (CVDs) [18] and even some cancers including liver pancreas and colon [19] (CVDs and cancers are not discussedin this review) Selected lncRNAs involved in the control of metabolic system activity (eg adipogenesis SRA ADINR NEAT1 and lncRAPs)and pathology of metabolic diseases (eg type 2 diabetes HI-LNC25 KCNQ1OT1 LOC283177 diabetic retinopathy MALAT1 and MIAT)are marked with the red color In NAFLD 535 lncRNAs are upregulated and 1200 are downregulated however these sequences are not yetwell characterized [20] This figure was produced using modified elements from the Servier Medical Art (httpwwwserviercom)
In a number of studies global transcriptome profilingwasused to select specific lncRNAs either involved in adipogen-esis or characteristic for adipocytes specifically isolated fromWAT or BAT In a seminal study done by Sun and cowork-ers [64] the transcriptome of primary brown and whiteadipocytes preadipocytes and cultured adipocytes was pro-filed Scientists identified 175 lncRNAs that were specificallyregulated during adipogenesis To further validate lncRNAsthat are functionally important for adipogenesis researchersselected the top 20 lncRNA genes based on the followingcriteria (i) significant upregulation in both brown and whitefat cultures (ii) binding to PPAR120574 or CEPB120572 promoters and(iii) independent validation of adipose-specific expressionBy using RNAi-mediated loss of function the top 10 mostimportant lncRNAs were selected and called Regulated inAdiPogenesis (lncRAPs) in order to highlight their key rolein proper differentiation of adipocyte precursors [64]
In comparison to white adipocytes brown adipocyteshave a higher mitochondrial content and express uncouplingprotein 1 (UCP1) which disperses chemical energy throughheat production Brown adipose tissue helps to regulateenergy expenditure in rodents and newborn babies butit has been considered to have no physiologic relevancein adults [65 66] However recent studies demonstratedthat metabolically functional BAT is also present in adulthumans As a result an understanding of BAT physiologymight provide an effective treatment of obesity or othermetabolic disorders [67] Identification of the lncRNAs cru-cial for BAT differentiation and function was done usingwhole transcriptome RNA sequencing Next gain and loss offunction studies established lncRNA Blnc1 as a potent acti-vator of thermogenic adipocyte differentiation It functionsupon formation of RNA-protein complex with the early Bcell factor 2 (EBF2) transcription factor to stimulate BAT
Mediators of Inflammation 7
formation Additionally lncRNA Blnc1 itself is a target ofEBF2 thereby forming a feed-forward regulatory loop Onthe other hand despite its stimulatory effects on brown pre-adipocyte differentiation lncRNA Blnc1 failed to promotedifferentiation of 3T3-L1 cells into UCP1 positive adipocytes[68] Another global profiling of gene expression duringmouse brown fat cell differentiation followed by candidatelncRNAs genomic context analysis pathway analysis andgene ontology enrichment of their associated protein-codinggenes was recently described In this study scientists identi-fied three lncRNAs (Gm15051 Tmem189 and Cebpd) associ-ated with their flanking coding genes (Hoxa1 CEBP120573 andCEBP120575) that participated in adipose commitment [69]
More recently RNA-seq analysis of murine brown ingui-nal white and epididymal white fat allowed for identificationof lncRNA cluster enriched in BAT Among them thelncRNA BATE1 was induced during brown adipose tissuedifferentiation Inhibition of BATE1 led to decreased expres-sion of brown fat markers and mitochondrial genes Authorsshowed that BATE1 binds to the heterogeneous nuclear ribo-nucleoprotein U a factor also required for BAT formation[70]
Finally interactions of lncRNAs with microRNAs in thecontext of adipogenesis were also described As demonstratedby Gernapudi and coworkers [71] the miR-140lncRNANEAT1 signaling network is necessary for adipogenesisAdipocyte derived stem cells isolated from miR-140 knock-out mice had dramatically decreased adipogenic capabilitiesfound to be associated with the downregulation of lncRNANEAT1 expression Overexpression of NEAT1 in such cellswas sufficient enough to restore their differentiation capabil-ities Authors identified a miR-140 binding site in the NEAT1sequence They also found that the physical interaction withmiR-140 in nucleus led to increased expression of NEAT1[71]
Excessive fat accumulation not only causes pathologicalexpansion of adipose tissue but also leads to triglyceridedeposition in the liver Presence of lipid droplets in excessof 5 of total hepatocytes is a diagnostic hallmark of non-alcoholic fatty liver disease (NAFLD) [72] Although mostpatients suffer from a mild course of illness approximately25 of cases progress and can subsequently lead to thedevelopment of steatohepatitis hepatic fibrosis liver cirrho-sis and hepatoma [73] Scientists and physicians estimatethat between 20 and 30 of Westrsquos general populationsuffers from NAFLD which makes this affects more than abillion people worldwide [74] To address questions aboutthe global expression pattern and functional contribution oflncRNAs during NAFLD Sun and coworkers [20] analyzedmicroarray expression using RNA extracted from liver biop-sies of healthy patients and those suffering from NAFLDIn this analysis 535 lncRNAs were found upregulated inNAFLD samples compared with controls and 1200 weredownregulated in NAFLD (Figure 5) Out of these sevenNAFLD samples that had highly up- or downregulatedgene expression were validated by quantitative real-timePCR (qPCR) Three lncRNA results obtained by qPCR weredifferent in comparison to the microarray data showing nochange between analyzed groups However changes in other
selected hits were confirmed highlighting the importance ofmicroarray data validation [20]
5 lncRNA in Diabetes and PancreaticFunctionGlucose Homeostasis
Pancreatic islets through the secretion of endocrinehormones such as insulin and glucagon play a key role inmetabolic homeostasis However one of the most commonmalfunctions of 120573-cells is caused by lack of insulin leading tothe development of diabetes [75] Additionally diabetes maydevelop as a result of insulin resistance a condition in whichthe body cannot effectively use the insulin produced by 120573-cells As a result blood sugar levels are deregulated and vari-ous tissues are exposed to prolonged hyperglycemia that overtime causes serious damage to many of the bodyrsquos systemsIt should be emphasized that hyperglycemia a hallmark ofdiabetes can cause both acute and long-term complicationslike cardiovascular disease stroke foot ulcers ketoacidosisor hyperosmolar coma [76] Based on etiology WHOclassifies diabetes into four categories but the vast majorityof cases fall into two broad etiopathogenetic groups type 1diabetes mellitus (T1DM) or type 2 diabetes mellitus (T2DM)[77]
Accounting for 5ndash10 of all cases T1DM is characterizedby the loss of insulin producing120573-cells due to an autoimmunereaction Autoimmune destruction of 120573-cells is unrelated tolifestyleThere are genetic variants known to increase the riskof T1DM development Type 2 diabetes mellitus represents90ndash95 of all cases and unlike T1DM is strongly associatedwith patientsrsquo lifestyle [78] A number of lifestyle factors areknown to be important for development of T2DM includingobesity (BMI above 30) lack of physical activity poor dietor stress [79] In fact most patients suffering from T2DM areobese It is a chronic disease that begins with insulin resist-ance a condition in which cells (mostly adipocytes hepato-cytes andmuscle cells) fail to properly absorb andmetabolizeglucose As a result 120573-cells increase insulin production toovercome hyperglycemia but this compensatory mechanismis transient and generally fails over time [77] Intensifiedmetabolism of 120573-cells leads to a gradual loss of their endo-crine function and finally causes 120573-cell apoptosis Similar toobesity diabetes is a worldwide problemAccording toWHOin 2014 the global prevalence of diabetes was estimated tobe 9 among adults aged over 18 years and it is expectedto double before 2030 [75 80] Additionally experts projectthat in 2030 diabetes will be the 7th leading cause of death[75]
Initial studies aimed at identification of specific lncRNAsexpressed in murine and human pancreatic islets were donein 2012 [81 82] By using next-generation sequencing scien-tists revealed a large collection of 1359 intergenic lncRNAsexpressed in murine 120573-cells (Figure 5) Many of them werehighly tissue-specific but their function was not evaluatedand future experiments are still needed [81] Similar numbersof 1128 intergenic and antisense lncRNAs were identifiedin human 120573-cells and many of them (eg HI-LNC12 andHI-LNC77) were highly tissue-specific [82] AdditionallylncRNA-encoding genes were preferentially located near
8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 5
adipocytes It is postulated that the excessive fat mass inducesthe production of reactive oxygen species and the develop-ment of chronic oxidative stress in patients with obesity andmetabolic dysfunction [44 45] In adipose tissue of obesepatients is observed severe inflammatory response charac-terized by high production of cytokines and increased infil-tration of immune cells including monocytes stimulated todifferentiate into macrophages and lymphocytes T activatedto proliferate and secrete cytokines [46 47] Cytokinessecreted by activated macrophages act on TLRs present onthe surface of adipocytes stimulating them to produce adipo-kines and cytokines [48] Based on these observations it hasbeen found that the reactions of both the immune and meta-bolic systems are closely connected and they play an impor-tant role in the development of metabolic disorders
4 lncRNA in Obesity andAdipogenesis Control
Adipocytes play a central role in energy homeostasis byfine-tuning the equilibrium between nutrient deposition(white adipose tissue WAT) and energy expenditure (brownadipose tissue BAT) Additionally adipose tissue acts asan endocrine organ by secreting factors (adipokines) thatregulate whole body energy and glucose homeostasis [49]Adipogenesis is a complex process governed by a network oftranscription factors cofactors and signaling intermediatesfrom numerous pathways [50] The transcriptional cascadethat regulates adipocyte differentiation is predominantlydriven by peroxisome proliferator-activated receptor gamma(PPAR120574) shown to be both necessary and sufficient foradipogenesis [51] Importantly adipogenesis is highly regu-lated not only by PPAR120574 but also by the coordinated effectsof other transcription factors including CCAATenhancer-binding proteins (CEBPs) Kruppel-like factors (KLFs) orWingless proteins (Wnt) and transcriptional cofactors [52ndash54]
In recent years there has been growing attention paid tononcoding RNAs as a novel cell intrinsic regulatory mecha-nism Specifically lncRNAs are now emerging as importantregulators of gene expression both on the transcriptional andon the posttranscriptional levels Long noncoding RNAs arebeing discovered using modern techniques of microarray ornext-generation sequencing However only small portionshave been fully characterized In the case of adipogenesisthere is wide expression of lncRNAs controlling a varietyof genes involved in the formation differentiation andactivation of adipocytes (Figure 5)
Steroid receptor RNA Activator (SRA) lncRNA was des-cribed for the first time as a molecule regulating adipo-genesis [55] It was shown that lncRNA SRA initially identi-fied as a coactivator of steroid receptors also acts as a tran-scriptional coactivator of PPAR120574 [55 56] lncRNA SRA pro-motes adipocyte differentiation by binding to the N-terminalportion of PPAR120574 and enhances its transcriptional activ-ity Additionally its overexpression significantly increasedmRNA and protein levels of adipocyte master regulatorsPPAR120574 and CEBP120572 as well as PPAR120574 target genes Con-versely knockdown of endogenous lncRNA SRA resulted in
inhibition of 3T3-L1 preadipocyte differentiation Moreovergene expression profiling showed that lncRNA SRA regulatesexpression of genes related to various cellular processesincluding cell cycle and insulin transduction pathways [55]Further experiments proved that lncRNA SRA overexpres-sion inhibits phosphorylation of p38 mitogen activated pro-tein kinase (MAPK) and c-JunNH2-terminal kinase (JNK) inthe early differentiation of ST2 mesenchymal precursor cellsIn contrast knockdown of lncRNA SRA increased p38 andJNK phosphorylation while reducing insulin receptormRNAand protein levels which led to decreased downstreamsignaling [57] Supporting that data SRA knock-out mice(SRAminusminus) are resistant to high fat diet (HFD) induced obesityAfter 14 weeks of HFD SRAminusminus mice are characterized byreduced WAT mass and decreased expression of adipocytegenes such as adiponectin or fatty acid-binding protein 4Thelean phenotype is also associated with smaller adipocytes inWAT compared to WT mice reduced liver mass fewer lipiddroplets in the liver and decreased expression of lipogenesis-associated genes Finally SRAminusminus animals have improvedinsulin sensitivity [58]
In a recent paper Xiao and coworkers [59] identifiedlncRNA ADINR (adipogenic differentiation induced non-coding RNA) that in cis transcriptionally activates CEBP120572It was upregulated 20ndash30-fold during the course of adipoge-nesis and was transcribed from a position sim450 bp upstreamof the CEBP120572 gene Knockdown of lncRNA ADINR withsiRNA resulted in a dramatic adipogenic defect as shown bya decreased number of Oil Red O positive cells and reducedadipogenic transcripts CEBP120572 PPAR120574 fatty acid-bindingprotein 4 and lipoprotein lipase Importantly inhibition ofadipogenesis caused by depletion of lncRNA ADINR wasrescued by overexpression of CEBP120572 The lncRNA ADINRmechanism of action relies on its binding to PA1 followed byrecruitment ofMLL34 histonemethyltransferase complexesIn turn that causes an increase inH3K4me3 and a decrease inH3K27me3 histone modification in the CEBP120572 locus duringadipogenesis [59]
Another lncRNA that induces adipogenesis in ciswas dis-covered within themouse PU1 locus Expression of the PU1gene gives rise to both the mRNA encoding PU1 proteinand antisense (AS) lncRNA originating from a promoter inthe antisense strand of intron 3 [60] The protein PU1 wasoriginally demonstrated to be a key transcription factor regu-lating hematopoiesis but recent studies revealed that it is alsoexpressed in 3T3-L1 preadipocytes and murine adipocytesisolated from WAT [60 61] Gain and loss of functionstudies established the PU1 protein as an inhibitor of 3T3-L1 preadipocyte differentiation acting via downregulation ofPPAR120574 [62] On the other hand PU1 AS lncRNA promotesadipogenesis through preventing PU1 mRNA translationby binding to PU1 mRNA to form an mRNAAS lncRNAduplex [60] Formation of this sense-antisense RNA duplexwas also confirmed in porcine adipocytes by an endogenousribonuclease protection assay combinedwithRT-PCR In linewith the previous data expression of PPAR120574 and fatty acidsynthase (FASN)was significantly upregulated in the lncRNAPU1 shRNA treated group [63]
6 Mediators of Inflammation
Diabetic retinopathy
CVDs
Type 2 diabetes
Cancers
NAFLD
ObesitySRA
ADINRNEAT1lncRAPs
MALAT1MIAT
HI-LNC25KCNQ1OT1LOC283177
Risk factors
(i) Abdominal obesity
(ii) Insulin resistance
(iii) Hyperglycemia
(iv) Hypertriglyceridemia
(v) Hypertension
uarr 535 lncRNAsdarr 1200 lncRNAs
Figure 5 Development of metabolic syndrome The common clinical symptoms of metabolic disorders include abdominal obesity insulinresistance hyperglycemia hypertriglyceridemia and hypertension [15] Their appearance is associated with a high risk for development ofserious diseases including type 2 diabetes diabetes-related diabetic retinopathy [16] and nonalcoholic fatty liver disease (NAFLD) [17] as wellas cardiovascular diseases (CVDs) [18] and even some cancers including liver pancreas and colon [19] (CVDs and cancers are not discussedin this review) Selected lncRNAs involved in the control of metabolic system activity (eg adipogenesis SRA ADINR NEAT1 and lncRAPs)and pathology of metabolic diseases (eg type 2 diabetes HI-LNC25 KCNQ1OT1 LOC283177 diabetic retinopathy MALAT1 and MIAT)are marked with the red color In NAFLD 535 lncRNAs are upregulated and 1200 are downregulated however these sequences are not yetwell characterized [20] This figure was produced using modified elements from the Servier Medical Art (httpwwwserviercom)
In a number of studies global transcriptome profilingwasused to select specific lncRNAs either involved in adipogen-esis or characteristic for adipocytes specifically isolated fromWAT or BAT In a seminal study done by Sun and cowork-ers [64] the transcriptome of primary brown and whiteadipocytes preadipocytes and cultured adipocytes was pro-filed Scientists identified 175 lncRNAs that were specificallyregulated during adipogenesis To further validate lncRNAsthat are functionally important for adipogenesis researchersselected the top 20 lncRNA genes based on the followingcriteria (i) significant upregulation in both brown and whitefat cultures (ii) binding to PPAR120574 or CEPB120572 promoters and(iii) independent validation of adipose-specific expressionBy using RNAi-mediated loss of function the top 10 mostimportant lncRNAs were selected and called Regulated inAdiPogenesis (lncRAPs) in order to highlight their key rolein proper differentiation of adipocyte precursors [64]
In comparison to white adipocytes brown adipocyteshave a higher mitochondrial content and express uncouplingprotein 1 (UCP1) which disperses chemical energy throughheat production Brown adipose tissue helps to regulateenergy expenditure in rodents and newborn babies butit has been considered to have no physiologic relevancein adults [65 66] However recent studies demonstratedthat metabolically functional BAT is also present in adulthumans As a result an understanding of BAT physiologymight provide an effective treatment of obesity or othermetabolic disorders [67] Identification of the lncRNAs cru-cial for BAT differentiation and function was done usingwhole transcriptome RNA sequencing Next gain and loss offunction studies established lncRNA Blnc1 as a potent acti-vator of thermogenic adipocyte differentiation It functionsupon formation of RNA-protein complex with the early Bcell factor 2 (EBF2) transcription factor to stimulate BAT
Mediators of Inflammation 7
formation Additionally lncRNA Blnc1 itself is a target ofEBF2 thereby forming a feed-forward regulatory loop Onthe other hand despite its stimulatory effects on brown pre-adipocyte differentiation lncRNA Blnc1 failed to promotedifferentiation of 3T3-L1 cells into UCP1 positive adipocytes[68] Another global profiling of gene expression duringmouse brown fat cell differentiation followed by candidatelncRNAs genomic context analysis pathway analysis andgene ontology enrichment of their associated protein-codinggenes was recently described In this study scientists identi-fied three lncRNAs (Gm15051 Tmem189 and Cebpd) associ-ated with their flanking coding genes (Hoxa1 CEBP120573 andCEBP120575) that participated in adipose commitment [69]
More recently RNA-seq analysis of murine brown ingui-nal white and epididymal white fat allowed for identificationof lncRNA cluster enriched in BAT Among them thelncRNA BATE1 was induced during brown adipose tissuedifferentiation Inhibition of BATE1 led to decreased expres-sion of brown fat markers and mitochondrial genes Authorsshowed that BATE1 binds to the heterogeneous nuclear ribo-nucleoprotein U a factor also required for BAT formation[70]
Finally interactions of lncRNAs with microRNAs in thecontext of adipogenesis were also described As demonstratedby Gernapudi and coworkers [71] the miR-140lncRNANEAT1 signaling network is necessary for adipogenesisAdipocyte derived stem cells isolated from miR-140 knock-out mice had dramatically decreased adipogenic capabilitiesfound to be associated with the downregulation of lncRNANEAT1 expression Overexpression of NEAT1 in such cellswas sufficient enough to restore their differentiation capabil-ities Authors identified a miR-140 binding site in the NEAT1sequence They also found that the physical interaction withmiR-140 in nucleus led to increased expression of NEAT1[71]
Excessive fat accumulation not only causes pathologicalexpansion of adipose tissue but also leads to triglyceridedeposition in the liver Presence of lipid droplets in excessof 5 of total hepatocytes is a diagnostic hallmark of non-alcoholic fatty liver disease (NAFLD) [72] Although mostpatients suffer from a mild course of illness approximately25 of cases progress and can subsequently lead to thedevelopment of steatohepatitis hepatic fibrosis liver cirrho-sis and hepatoma [73] Scientists and physicians estimatethat between 20 and 30 of Westrsquos general populationsuffers from NAFLD which makes this affects more than abillion people worldwide [74] To address questions aboutthe global expression pattern and functional contribution oflncRNAs during NAFLD Sun and coworkers [20] analyzedmicroarray expression using RNA extracted from liver biop-sies of healthy patients and those suffering from NAFLDIn this analysis 535 lncRNAs were found upregulated inNAFLD samples compared with controls and 1200 weredownregulated in NAFLD (Figure 5) Out of these sevenNAFLD samples that had highly up- or downregulatedgene expression were validated by quantitative real-timePCR (qPCR) Three lncRNA results obtained by qPCR weredifferent in comparison to the microarray data showing nochange between analyzed groups However changes in other
selected hits were confirmed highlighting the importance ofmicroarray data validation [20]
5 lncRNA in Diabetes and PancreaticFunctionGlucose Homeostasis
Pancreatic islets through the secretion of endocrinehormones such as insulin and glucagon play a key role inmetabolic homeostasis However one of the most commonmalfunctions of 120573-cells is caused by lack of insulin leading tothe development of diabetes [75] Additionally diabetes maydevelop as a result of insulin resistance a condition in whichthe body cannot effectively use the insulin produced by 120573-cells As a result blood sugar levels are deregulated and vari-ous tissues are exposed to prolonged hyperglycemia that overtime causes serious damage to many of the bodyrsquos systemsIt should be emphasized that hyperglycemia a hallmark ofdiabetes can cause both acute and long-term complicationslike cardiovascular disease stroke foot ulcers ketoacidosisor hyperosmolar coma [76] Based on etiology WHOclassifies diabetes into four categories but the vast majorityof cases fall into two broad etiopathogenetic groups type 1diabetes mellitus (T1DM) or type 2 diabetes mellitus (T2DM)[77]
Accounting for 5ndash10 of all cases T1DM is characterizedby the loss of insulin producing120573-cells due to an autoimmunereaction Autoimmune destruction of 120573-cells is unrelated tolifestyleThere are genetic variants known to increase the riskof T1DM development Type 2 diabetes mellitus represents90ndash95 of all cases and unlike T1DM is strongly associatedwith patientsrsquo lifestyle [78] A number of lifestyle factors areknown to be important for development of T2DM includingobesity (BMI above 30) lack of physical activity poor dietor stress [79] In fact most patients suffering from T2DM areobese It is a chronic disease that begins with insulin resist-ance a condition in which cells (mostly adipocytes hepato-cytes andmuscle cells) fail to properly absorb andmetabolizeglucose As a result 120573-cells increase insulin production toovercome hyperglycemia but this compensatory mechanismis transient and generally fails over time [77] Intensifiedmetabolism of 120573-cells leads to a gradual loss of their endo-crine function and finally causes 120573-cell apoptosis Similar toobesity diabetes is a worldwide problemAccording toWHOin 2014 the global prevalence of diabetes was estimated tobe 9 among adults aged over 18 years and it is expectedto double before 2030 [75 80] Additionally experts projectthat in 2030 diabetes will be the 7th leading cause of death[75]
Initial studies aimed at identification of specific lncRNAsexpressed in murine and human pancreatic islets were donein 2012 [81 82] By using next-generation sequencing scien-tists revealed a large collection of 1359 intergenic lncRNAsexpressed in murine 120573-cells (Figure 5) Many of them werehighly tissue-specific but their function was not evaluatedand future experiments are still needed [81] Similar numbersof 1128 intergenic and antisense lncRNAs were identifiedin human 120573-cells and many of them (eg HI-LNC12 andHI-LNC77) were highly tissue-specific [82] AdditionallylncRNA-encoding genes were preferentially located near
8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
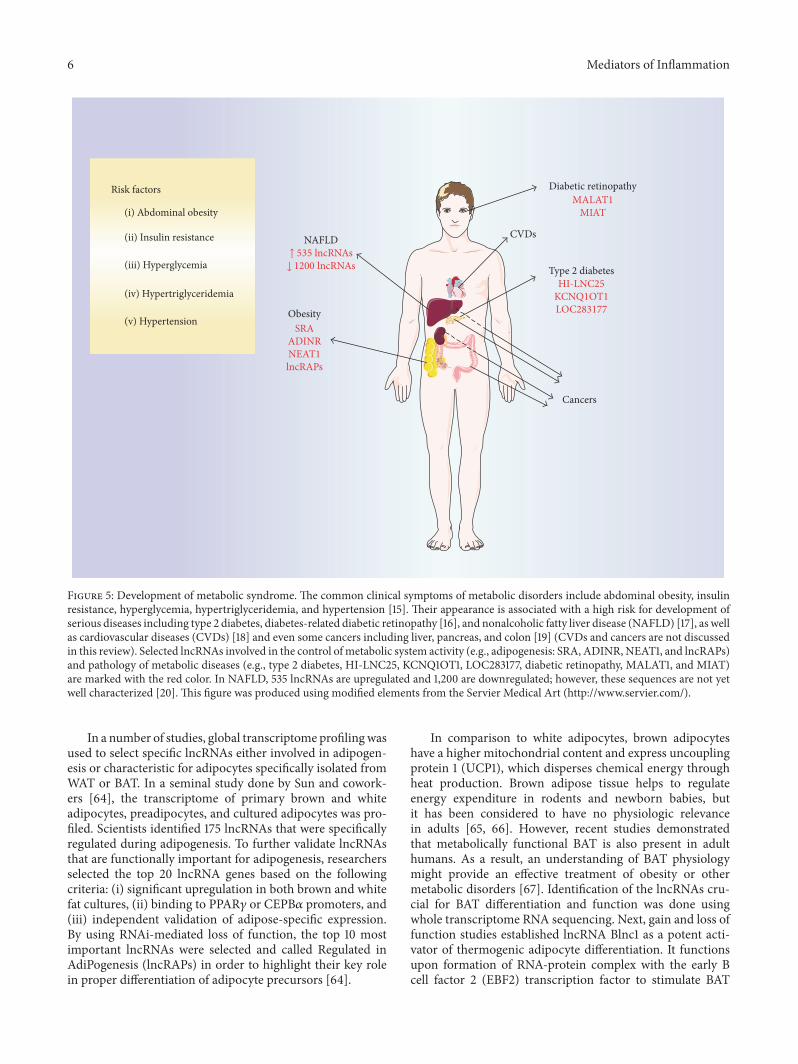
6 Mediators of Inflammation
Diabetic retinopathy
CVDs
Type 2 diabetes
Cancers
NAFLD
ObesitySRA
ADINRNEAT1lncRAPs
MALAT1MIAT
HI-LNC25KCNQ1OT1LOC283177
Risk factors
(i) Abdominal obesity
(ii) Insulin resistance
(iii) Hyperglycemia
(iv) Hypertriglyceridemia
(v) Hypertension
uarr 535 lncRNAsdarr 1200 lncRNAs
Figure 5 Development of metabolic syndrome The common clinical symptoms of metabolic disorders include abdominal obesity insulinresistance hyperglycemia hypertriglyceridemia and hypertension [15] Their appearance is associated with a high risk for development ofserious diseases including type 2 diabetes diabetes-related diabetic retinopathy [16] and nonalcoholic fatty liver disease (NAFLD) [17] as wellas cardiovascular diseases (CVDs) [18] and even some cancers including liver pancreas and colon [19] (CVDs and cancers are not discussedin this review) Selected lncRNAs involved in the control of metabolic system activity (eg adipogenesis SRA ADINR NEAT1 and lncRAPs)and pathology of metabolic diseases (eg type 2 diabetes HI-LNC25 KCNQ1OT1 LOC283177 diabetic retinopathy MALAT1 and MIAT)are marked with the red color In NAFLD 535 lncRNAs are upregulated and 1200 are downregulated however these sequences are not yetwell characterized [20] This figure was produced using modified elements from the Servier Medical Art (httpwwwserviercom)
In a number of studies global transcriptome profilingwasused to select specific lncRNAs either involved in adipogen-esis or characteristic for adipocytes specifically isolated fromWAT or BAT In a seminal study done by Sun and cowork-ers [64] the transcriptome of primary brown and whiteadipocytes preadipocytes and cultured adipocytes was pro-filed Scientists identified 175 lncRNAs that were specificallyregulated during adipogenesis To further validate lncRNAsthat are functionally important for adipogenesis researchersselected the top 20 lncRNA genes based on the followingcriteria (i) significant upregulation in both brown and whitefat cultures (ii) binding to PPAR120574 or CEPB120572 promoters and(iii) independent validation of adipose-specific expressionBy using RNAi-mediated loss of function the top 10 mostimportant lncRNAs were selected and called Regulated inAdiPogenesis (lncRAPs) in order to highlight their key rolein proper differentiation of adipocyte precursors [64]
In comparison to white adipocytes brown adipocyteshave a higher mitochondrial content and express uncouplingprotein 1 (UCP1) which disperses chemical energy throughheat production Brown adipose tissue helps to regulateenergy expenditure in rodents and newborn babies butit has been considered to have no physiologic relevancein adults [65 66] However recent studies demonstratedthat metabolically functional BAT is also present in adulthumans As a result an understanding of BAT physiologymight provide an effective treatment of obesity or othermetabolic disorders [67] Identification of the lncRNAs cru-cial for BAT differentiation and function was done usingwhole transcriptome RNA sequencing Next gain and loss offunction studies established lncRNA Blnc1 as a potent acti-vator of thermogenic adipocyte differentiation It functionsupon formation of RNA-protein complex with the early Bcell factor 2 (EBF2) transcription factor to stimulate BAT
Mediators of Inflammation 7
formation Additionally lncRNA Blnc1 itself is a target ofEBF2 thereby forming a feed-forward regulatory loop Onthe other hand despite its stimulatory effects on brown pre-adipocyte differentiation lncRNA Blnc1 failed to promotedifferentiation of 3T3-L1 cells into UCP1 positive adipocytes[68] Another global profiling of gene expression duringmouse brown fat cell differentiation followed by candidatelncRNAs genomic context analysis pathway analysis andgene ontology enrichment of their associated protein-codinggenes was recently described In this study scientists identi-fied three lncRNAs (Gm15051 Tmem189 and Cebpd) associ-ated with their flanking coding genes (Hoxa1 CEBP120573 andCEBP120575) that participated in adipose commitment [69]
More recently RNA-seq analysis of murine brown ingui-nal white and epididymal white fat allowed for identificationof lncRNA cluster enriched in BAT Among them thelncRNA BATE1 was induced during brown adipose tissuedifferentiation Inhibition of BATE1 led to decreased expres-sion of brown fat markers and mitochondrial genes Authorsshowed that BATE1 binds to the heterogeneous nuclear ribo-nucleoprotein U a factor also required for BAT formation[70]
Finally interactions of lncRNAs with microRNAs in thecontext of adipogenesis were also described As demonstratedby Gernapudi and coworkers [71] the miR-140lncRNANEAT1 signaling network is necessary for adipogenesisAdipocyte derived stem cells isolated from miR-140 knock-out mice had dramatically decreased adipogenic capabilitiesfound to be associated with the downregulation of lncRNANEAT1 expression Overexpression of NEAT1 in such cellswas sufficient enough to restore their differentiation capabil-ities Authors identified a miR-140 binding site in the NEAT1sequence They also found that the physical interaction withmiR-140 in nucleus led to increased expression of NEAT1[71]
Excessive fat accumulation not only causes pathologicalexpansion of adipose tissue but also leads to triglyceridedeposition in the liver Presence of lipid droplets in excessof 5 of total hepatocytes is a diagnostic hallmark of non-alcoholic fatty liver disease (NAFLD) [72] Although mostpatients suffer from a mild course of illness approximately25 of cases progress and can subsequently lead to thedevelopment of steatohepatitis hepatic fibrosis liver cirrho-sis and hepatoma [73] Scientists and physicians estimatethat between 20 and 30 of Westrsquos general populationsuffers from NAFLD which makes this affects more than abillion people worldwide [74] To address questions aboutthe global expression pattern and functional contribution oflncRNAs during NAFLD Sun and coworkers [20] analyzedmicroarray expression using RNA extracted from liver biop-sies of healthy patients and those suffering from NAFLDIn this analysis 535 lncRNAs were found upregulated inNAFLD samples compared with controls and 1200 weredownregulated in NAFLD (Figure 5) Out of these sevenNAFLD samples that had highly up- or downregulatedgene expression were validated by quantitative real-timePCR (qPCR) Three lncRNA results obtained by qPCR weredifferent in comparison to the microarray data showing nochange between analyzed groups However changes in other
selected hits were confirmed highlighting the importance ofmicroarray data validation [20]
5 lncRNA in Diabetes and PancreaticFunctionGlucose Homeostasis
Pancreatic islets through the secretion of endocrinehormones such as insulin and glucagon play a key role inmetabolic homeostasis However one of the most commonmalfunctions of 120573-cells is caused by lack of insulin leading tothe development of diabetes [75] Additionally diabetes maydevelop as a result of insulin resistance a condition in whichthe body cannot effectively use the insulin produced by 120573-cells As a result blood sugar levels are deregulated and vari-ous tissues are exposed to prolonged hyperglycemia that overtime causes serious damage to many of the bodyrsquos systemsIt should be emphasized that hyperglycemia a hallmark ofdiabetes can cause both acute and long-term complicationslike cardiovascular disease stroke foot ulcers ketoacidosisor hyperosmolar coma [76] Based on etiology WHOclassifies diabetes into four categories but the vast majorityof cases fall into two broad etiopathogenetic groups type 1diabetes mellitus (T1DM) or type 2 diabetes mellitus (T2DM)[77]
Accounting for 5ndash10 of all cases T1DM is characterizedby the loss of insulin producing120573-cells due to an autoimmunereaction Autoimmune destruction of 120573-cells is unrelated tolifestyleThere are genetic variants known to increase the riskof T1DM development Type 2 diabetes mellitus represents90ndash95 of all cases and unlike T1DM is strongly associatedwith patientsrsquo lifestyle [78] A number of lifestyle factors areknown to be important for development of T2DM includingobesity (BMI above 30) lack of physical activity poor dietor stress [79] In fact most patients suffering from T2DM areobese It is a chronic disease that begins with insulin resist-ance a condition in which cells (mostly adipocytes hepato-cytes andmuscle cells) fail to properly absorb andmetabolizeglucose As a result 120573-cells increase insulin production toovercome hyperglycemia but this compensatory mechanismis transient and generally fails over time [77] Intensifiedmetabolism of 120573-cells leads to a gradual loss of their endo-crine function and finally causes 120573-cell apoptosis Similar toobesity diabetes is a worldwide problemAccording toWHOin 2014 the global prevalence of diabetes was estimated tobe 9 among adults aged over 18 years and it is expectedto double before 2030 [75 80] Additionally experts projectthat in 2030 diabetes will be the 7th leading cause of death[75]
Initial studies aimed at identification of specific lncRNAsexpressed in murine and human pancreatic islets were donein 2012 [81 82] By using next-generation sequencing scien-tists revealed a large collection of 1359 intergenic lncRNAsexpressed in murine 120573-cells (Figure 5) Many of them werehighly tissue-specific but their function was not evaluatedand future experiments are still needed [81] Similar numbersof 1128 intergenic and antisense lncRNAs were identifiedin human 120573-cells and many of them (eg HI-LNC12 andHI-LNC77) were highly tissue-specific [82] AdditionallylncRNA-encoding genes were preferentially located near
8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 7
formation Additionally lncRNA Blnc1 itself is a target ofEBF2 thereby forming a feed-forward regulatory loop Onthe other hand despite its stimulatory effects on brown pre-adipocyte differentiation lncRNA Blnc1 failed to promotedifferentiation of 3T3-L1 cells into UCP1 positive adipocytes[68] Another global profiling of gene expression duringmouse brown fat cell differentiation followed by candidatelncRNAs genomic context analysis pathway analysis andgene ontology enrichment of their associated protein-codinggenes was recently described In this study scientists identi-fied three lncRNAs (Gm15051 Tmem189 and Cebpd) associ-ated with their flanking coding genes (Hoxa1 CEBP120573 andCEBP120575) that participated in adipose commitment [69]
More recently RNA-seq analysis of murine brown ingui-nal white and epididymal white fat allowed for identificationof lncRNA cluster enriched in BAT Among them thelncRNA BATE1 was induced during brown adipose tissuedifferentiation Inhibition of BATE1 led to decreased expres-sion of brown fat markers and mitochondrial genes Authorsshowed that BATE1 binds to the heterogeneous nuclear ribo-nucleoprotein U a factor also required for BAT formation[70]
Finally interactions of lncRNAs with microRNAs in thecontext of adipogenesis were also described As demonstratedby Gernapudi and coworkers [71] the miR-140lncRNANEAT1 signaling network is necessary for adipogenesisAdipocyte derived stem cells isolated from miR-140 knock-out mice had dramatically decreased adipogenic capabilitiesfound to be associated with the downregulation of lncRNANEAT1 expression Overexpression of NEAT1 in such cellswas sufficient enough to restore their differentiation capabil-ities Authors identified a miR-140 binding site in the NEAT1sequence They also found that the physical interaction withmiR-140 in nucleus led to increased expression of NEAT1[71]
Excessive fat accumulation not only causes pathologicalexpansion of adipose tissue but also leads to triglyceridedeposition in the liver Presence of lipid droplets in excessof 5 of total hepatocytes is a diagnostic hallmark of non-alcoholic fatty liver disease (NAFLD) [72] Although mostpatients suffer from a mild course of illness approximately25 of cases progress and can subsequently lead to thedevelopment of steatohepatitis hepatic fibrosis liver cirrho-sis and hepatoma [73] Scientists and physicians estimatethat between 20 and 30 of Westrsquos general populationsuffers from NAFLD which makes this affects more than abillion people worldwide [74] To address questions aboutthe global expression pattern and functional contribution oflncRNAs during NAFLD Sun and coworkers [20] analyzedmicroarray expression using RNA extracted from liver biop-sies of healthy patients and those suffering from NAFLDIn this analysis 535 lncRNAs were found upregulated inNAFLD samples compared with controls and 1200 weredownregulated in NAFLD (Figure 5) Out of these sevenNAFLD samples that had highly up- or downregulatedgene expression were validated by quantitative real-timePCR (qPCR) Three lncRNA results obtained by qPCR weredifferent in comparison to the microarray data showing nochange between analyzed groups However changes in other
selected hits were confirmed highlighting the importance ofmicroarray data validation [20]
5 lncRNA in Diabetes and PancreaticFunctionGlucose Homeostasis
Pancreatic islets through the secretion of endocrinehormones such as insulin and glucagon play a key role inmetabolic homeostasis However one of the most commonmalfunctions of 120573-cells is caused by lack of insulin leading tothe development of diabetes [75] Additionally diabetes maydevelop as a result of insulin resistance a condition in whichthe body cannot effectively use the insulin produced by 120573-cells As a result blood sugar levels are deregulated and vari-ous tissues are exposed to prolonged hyperglycemia that overtime causes serious damage to many of the bodyrsquos systemsIt should be emphasized that hyperglycemia a hallmark ofdiabetes can cause both acute and long-term complicationslike cardiovascular disease stroke foot ulcers ketoacidosisor hyperosmolar coma [76] Based on etiology WHOclassifies diabetes into four categories but the vast majorityof cases fall into two broad etiopathogenetic groups type 1diabetes mellitus (T1DM) or type 2 diabetes mellitus (T2DM)[77]
Accounting for 5ndash10 of all cases T1DM is characterizedby the loss of insulin producing120573-cells due to an autoimmunereaction Autoimmune destruction of 120573-cells is unrelated tolifestyleThere are genetic variants known to increase the riskof T1DM development Type 2 diabetes mellitus represents90ndash95 of all cases and unlike T1DM is strongly associatedwith patientsrsquo lifestyle [78] A number of lifestyle factors areknown to be important for development of T2DM includingobesity (BMI above 30) lack of physical activity poor dietor stress [79] In fact most patients suffering from T2DM areobese It is a chronic disease that begins with insulin resist-ance a condition in which cells (mostly adipocytes hepato-cytes andmuscle cells) fail to properly absorb andmetabolizeglucose As a result 120573-cells increase insulin production toovercome hyperglycemia but this compensatory mechanismis transient and generally fails over time [77] Intensifiedmetabolism of 120573-cells leads to a gradual loss of their endo-crine function and finally causes 120573-cell apoptosis Similar toobesity diabetes is a worldwide problemAccording toWHOin 2014 the global prevalence of diabetes was estimated tobe 9 among adults aged over 18 years and it is expectedto double before 2030 [75 80] Additionally experts projectthat in 2030 diabetes will be the 7th leading cause of death[75]
Initial studies aimed at identification of specific lncRNAsexpressed in murine and human pancreatic islets were donein 2012 [81 82] By using next-generation sequencing scien-tists revealed a large collection of 1359 intergenic lncRNAsexpressed in murine 120573-cells (Figure 5) Many of them werehighly tissue-specific but their function was not evaluatedand future experiments are still needed [81] Similar numbersof 1128 intergenic and antisense lncRNAs were identifiedin human 120573-cells and many of them (eg HI-LNC12 andHI-LNC77) were highly tissue-specific [82] AdditionallylncRNA-encoding genes were preferentially located near
8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Mediators of Inflammation
genes encoding important regulators of 120573-cell functiondevelopment and transcription One of them lncRNA HI-LNC25 conserved between mouse and human was shownto positively regulate GLIS3 mRNA Silencing of lncRNAHI-LNC25 by shRNA depleted GLIS3 mRNA GLIS3 encodesa pancreatic islet transcription factor that is mutated inmonogenic diabetes and contains T2DM risk variants [8384] In the same set of experiments scientists also showedthat selected lncRNAs (eg HI-LNC12 and HI-LNC25) arelinked to 120573-cell differentiation program as their expressionwas significantly higher in human islets when compared tothe embryonic pancreas Moreover the addition of glucose toin vitro 120573-cell culture induced expression of some lncRNAssuggesting their importance for mature islet cell physiologyFinally authors showed deregulation of isletsrsquo lncRNAs inT2DM by comparing profiles of lncRNAs in islets from 19nondiabetic and 16 T2DM donors Two lncRNAs namelyKCNQ1OT1 and HI-LNC45 were significantly increased ordecreased in T2DM islets respectively [82]
Seven lncRNAs specific for 120573-cells as reported by Moranand coworkers [82] were also discussed in another studythat analyzed pancreatic islets from 89 individuals with orwithout diabetes Microarray analysis RNA sequencing andexome sequencing methods enabled researchers to identify493 lncRNAs that were differentially expressed in the pan-creatic islets Out of those 17 long intergenic noncodingRNAs (lincRNAs) were significantly associated with donorrsquosHbA1c levels and two (LOC283177 and SNHG5) were alsoinvolved in gene expression regulation To obtain insight intofunctional target genes of lncRNAs LOC283177 and SNHG5the authors performed a coexpression analysis linking theirexpression with all other genes in pancreatic islets lncRNALOC283177 levels correlated with expression of genes thatplay a key role in islet function namely synaptotagmin 11MAP-kinase activating death domain (MADD) and pairedbox 6 (PAX6) Synaptotagmin 11 is known to regulate theexocytosis of insulin and MADD proinsulin synthesis andPAX6 is involved in development of pancreatic islets [85 86]Supporting these findings lncRNA LOC283177 expressionwas found to be directly associated with insulin exocytosis inthe islets [87]
Insulin is secreted from 120573-cells in response to glucosewhile other nutrients such as free fatty acids and amino acidscan augment glucose-induced insulin secretion In additionvarious hormones (eg leptin growth hormone glucagonlike peptide-1 and estrogen) also regulate insulin secretionAs a result 120573-cells are called a metabolic hub in the bodythat coordinate nutrient metabolism with endocrine system[88] Insufficient insulin production or insulin resistance isa prime cause of diabetic complications Among the manytissues affected by hyperglycemia endothelial cells are veryimportant because they are implicated in pathogenesis ofdiabetes-related microvascular and macrovascular complica-tions [16]
One of these complications diabetic retinopathy wasrecently shown to be influenced by lncRNAMALAT1 (meta-stasis-associated lung adenocarcinoma transcript 1) [89] (Fig-ure 5) In a mouse model of streptozotocin-induced diabe-tes scientists performed lncRNA expression profiling and
compared expression between diabetic and control reti-nas Microarray analysis followed by qPCR validation ofselected genes allowed for identification of 303 differen-tially expressed lncRNAs including 214 downregulated and89 upregulated genes in diabetic versus nondiabetic sam-ples One gene lncRNA chr19 5795689ndash5802671 a murineortholog of human lncRNA MALAT1 had a greater than 10-fold upregulation in diabetic retinas [89] lncRNA MALAT1was previously described in humans to be deregulated inseveral solid tumors and was associated with cancer metas-tasis and recurrence [90] It was shown to be significantlyupregulated in a RF6A cell model of hyperglycemia in theaqueous tumor samples and in fibrovascular membranes ofdiabetic patients [89]
To further investigate the molecular mechanism oflncRNA MALAT1 action scientists decided to analyze itsfunction in retinal vasculature and endothelial cell dysfunc-tion in diabetes mellitus [91] lncRNA MALAT1 knockdownin retinas of dbdb mice resulted in amelioration of diabeticretinopathy as manifested by reduced apoptosis of retinalcells and pericytes Additionally retinal inflammation wasalso alleviated Importantly after administration of lncRNAMALAT shRNA scientists observed decreased retinal vascu-lar permeability as measured by Evans blue leakage More-over in vitro tests showed that lncRNAMALAT1 knockdowndecreased retinal endothelial cell proliferation migrationand tube formation In RF6A endothelial cells lncRNAMALAT1 acts via induction of p38 MAPK signaling andits silencing reduced phosphorylated p38 level but had noeffect on phosphorylated ERK12 or JNK12 Pretreatment ofRF6A cells with SB203580 a p38 MAPK pathway inhibitorstrongly blocked the effect of lncRNA MALAT1-induced cellproliferation [91]
More recently the same research group reported thatlncRNA MIAT (myocardial infarction-associated transcript)also regulates diabetes mellitus microvascular dysfunctionacting as a competing endogenous RNA [92] (Figure 5)Similar to a previous report lncRNA MIAT expression wasincreased in diabetic retinas and endothelial cells culturedin high glucose medium As shown by in vivo tests lncRNAMIAT knockdown ameliorated diabetes mellitus inducedretinal microvascular dysfunction In line with in vivo datasilencing of lncRNA MIAT in endothelial cells cultured invitro led to an inhibition of proliferation migration andtube formation lncRNAs may act as an mRNA molecularsponge and regulatemiRNAs available for binding their targetmRNAs Bioinformatics suggested that the lncRNA MIATsequence contains 4 putative miRNA binding sites includingmiR-29a-3p miR-29b-3p miR-29c-3p and miR-150-5p Oneof these miR-150-5p was proven to directly target lncRNAMIAT in endothelial cell both in vitro and in vivo [92] Thisparticular miRNA is of great importance for angiogenesisbecause it regulates the expression of VEGF a key angiogenicfactor involved in physiological and pathological angiogene-sis [93] Yan and coworkers [92] proved that MIAT regulatesthe expression of miR-150-5p target gene VEGF creating acritical regulatory loop for endothelial cell function Duringangiogenesis lncRNA MIAT is significantly upregulatedwhich in turn alleviates the miR-150-5p repression effect
Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 9
thereby upregulating the level of miR-150-5p target geneVEGF [92]
In contrast to lncRNAMIAT lncRNAMEG3 (maternallyexpressed gene 3) expressionwas significantly downregulatedin the retinas and endothelial cells of STZ-induced diabeticmice upon high glucose and oxidative stress conditions [94]Its knockdown regulated retinal endothelial cell proliferationmigration and tube formation in vitro As demonstratedin RF6A endothelial cells inhibition of lncRNA MEG3 bysiRNA significantly increased levels of phosphorylated PI3Kand phosphorylated Akt at Thr308 and Ser473 However levelsof the total PI3K and total Akt were not affected Addition ofPI3K inhibitors to cell cultures abrogated the observed phe-notype proving that lncRNA MEG3 regulates hyperprolifer-ation of retinal endothelial cells through PI3KAkt signaling[94] PI3KAkt signaling is an important signaling pathwayregulating glycogen metabolism Additionally its activationinfluences endothelial cell biology by modulating angiogene-sis proliferation and microvascular permeability [95]
Additional Points
SummaryThe data demonstrated above highlights an impor-tant role of newly identified lncRNAs in the etiology andprogression of metabolic diseases Importantly new reportsabout lncRNAs such as the identification of lncRNA GAS5whose serum levels correlate with T2DM aid in leadingto a better understanding of this complex processes [96]However further cell culture and animal studies are needed tofully clarify lncRNArsquos mechanism of action Our understand-ing of lncRNA function is still very limited and importantquestions need to be addressed First how is their expressionregulated Taking into account that many lncRNAs areexpressed in a cell type specificmanner their expressionmustbe tightly regulated As a result currently available databasesof lncRNAs are likely to be incomplete Secondly little isknown about the biological functions of lncRNAs Despitelarge numbers of differentially expressedmolecules in variousdiseases only a few have been thoroughly studied For in vitrostudies although there are many utilizable techniques (egRNA-seq crosslinking immunoprecipitation or chromatinisolation by RNA purification) we still lack a proper in vivomodel Thirdly very little is known about secondary andtertiary structures of lncRNAs Such studies might help tobetter predict lncRNA function or enable a more detailedclassification Finally as mentioned previously for lncRNAGAS5 lncRNAs may serve as a prognostic or diagnosticmarker However the reliability and sensitivity of such mark-ers require careful evaluation
Competing Interests
The authors declare that they have no competing interests
Acknowledgments
This study was supported by the research grant from theNational Science Centre 201311BNZZ00125 and grantfor young scientists (BMN72016) Faculty of Biochemistry
Biophysics and Biotechnology of Jagiellonian University is apartner of the Leading National Research Center (KNOW)supported by the Ministry of Science and Higher Education
References
[1] M Sun and W L Kraus ldquoFrom discovery to function theexpanding roles of long noncoding RNAs in physiology anddiseaserdquo Endocrine Reviews vol 36 no 1 pp 25ndash64 2015
[2] J E Wilusz H Sunwoo and D L Spector ldquoLong noncodingRNAs functional surprises from the RNA worldrdquo Genes andDevelopment vol 23 no 13 pp 1494ndash1504 2009
[3] C P Ponting P L Oliver andWReik ldquoEvolution and functionsof longnoncodingRNAsrdquoCell vol 136 no 4 pp 629ndash641 2009
[4] T R Mercer M E Dinger and J S Mattick ldquoLong non-codingRNAs insights into functionsrdquoNature Reviews Genetics vol 10no 3 pp 155ndash159 2009
[5] M Guttman I Amit M Garber et al ldquoChromatin signaturereveals over a thousand highly conserved large non-codingRNAs in mammalsrdquo Nature vol 458 no 7235 pp 223ndash2272009
[6] M J Hangauer I W Vaughn and M T McManus ldquoPervasivetranscription of the human genome produces thousands ofpreviously unidentified long intergenic noncoding RNAsrdquo PLoSGenetics vol 9 no 6 Article ID e1003569 2013
[7] A Pircher J Gebetsberger and N Polacek ldquoRibosome-asso-ciated ncRNAs an emerging class of translation regulatorsrdquoRNA Biology vol 11 no 11 pp 1335ndash1339 2014
[8] RKurokawaMG Rosenfeld andCKGlass ldquoTranscriptionalregulation through noncoding RNAs and epigenetic modifica-tionsrdquo RNA Biology vol 6 no 3 pp 233ndash236 2009
[9] K C Wang Y W Yang B Liu et al ldquoA long noncodingRNA maintains active chromatin to coordinate homeotic geneexpressionrdquo Nature vol 472 no 7341 pp 120ndash124 2011
[10] M Krawczyk and B M Emerson ldquoP50-associated COX-2Extragenic RNA (pacer) activates human COX-2 gene expres-sion by occluding repressive NF-120581B p50 complexesrdquo eLife vol2014 no 3 Article ID e01776 2014
[11] Z Li T-C Chao K-Y Chang et al ldquoThe long noncoding RNATHRIL regulates TNF120572 expression through its interaction withhnRNPLrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 111 no 3 pp 1002ndash1007 2014
[12] V Tripathi J D Ellis Z Shen et al ldquoThe nuclear-retainednoncoding RNA MALAT1 regulates alternative splicing bymodulating SR splicing factor phosphorylationrdquoMolecular Cellvol 39 no 6 pp 925ndash938 2010
[13] C Carrieri L CimattiM Biagioli et al ldquoLong non-coding anti-sense RNA controls Uchl1 translation through an embeddedSINEB2 repeatrdquo Nature vol 491 no 7424 pp 454ndash457 2012
[14] J-H Yoon K Abdelmohsen S Srikantan et al ldquoLincRNA-p21suppresses target mRNA translationrdquoMolecular Cell vol 47 no4 pp 648ndash655 2012
[15] L A Barness J M Opitz and E Gilbert-Barness ldquoObesitygenetic molecular and environmental aspectsrdquoAmerican Jour-nal of Medical Genetics Part A vol 143 no 24 pp 3016ndash30342007
[16] M J Fowler ldquoMicrovascular and macrovascular complicationsof diabetesrdquo Clinical Diabetes vol 26 no 2 pp 77ndash82 2008
[17] S Stojsavljevic M Gomercic Palcic L Virovic Jukic L SmircicDuvnjak and M Duvnjak ldquoAdipokines and proinflammatory
10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Mediators of Inflammation
cytokines the key mediators in the pathogenesis of nonalco-holic fatty liver diseaserdquoWorld Journal of Gastroenterology vol20 no 48 pp 18070ndash18091 2014
[18] EGalkina andK Ley ldquoImmune and inflammatorymechanismsof atherosclerosisrdquo Annual Review of Immunology vol 27 pp165ndash197 2009
[19] A Mantovani P Allavena A Sica and F Balkwill ldquoCancer-related inflammationrdquo Nature vol 454 no 7203 pp 436ndash4442008
[20] C Sun X Liu Z Yi et al ldquoGenome-wide analysis of long non-coding RNA expression profiles in patients with non-alcoholicfatty liver diseaserdquo IUBMB Life vol 67 no 11 pp 847ndash852 2015
[21] J S Mattick P P Amaral M E Dinger T R Mercer and MF Mehler ldquoRNA regulation of epigenetic processesrdquo BioEssaysvol 31 no 1 pp 51ndash59 2009
[22] K Zhang Z-M Shi Y-N Chang Z-M Hu H-X Qi and WHong ldquoThe ways of action of long non-coding RNAs in cyto-plasm and nucleusrdquo Gene vol 547 no 1 pp 1ndash9 2014
[23] A T Willingham A P Orth S Batalov et al ldquoA strategy forprobing the function of noncoding RNAs finds a repressor ofNFATrdquo Science vol 309 no 5740 pp 1570ndash1573 2005
[24] C J Brown B DHendrich J L Rupert et al ldquoThe humanXISTgene analysis of a 17 kb inactive X-specific RNA that containsconserved repeats and is highly localized within the nucleusrdquoCell vol 71 no 3 pp 527ndash542 1992
[25] T C Roberts K V Morris and M S Weinberg ldquoPerspectiveson the mechanism of transcriptional regulation by long non-coding RNAsrdquo Epigenetics vol 9 no 1 pp 13ndash20 2014
[26] C Chu K Qu F Zhong S Artandi and H Chang ldquoGenomicmaps of long noncoding RNA occupancy reveal principles ofrna-chromatin interactionsrdquo Molecular Cell vol 44 no 4 pp667ndash678 2011
[27] N A Rapicavoli K Qu J Zhang M Mikhail R-M Labergeand H Y Chang ldquoA mammalian pseudogene lncRNA at theinterface of inflammation and antiinflammatory therapeuticsrdquoeLife vol 2013 no 2 Article ID e00762 2013
[28] M Huarte M Guttman D Feldser et al ldquoA large intergenicnoncoding RNA induced by p53 mediates global gene repres-sion in the p53 responserdquo Cell vol 142 no 3 pp 409ndash419 2010
[29] S Carpenter D AielloM K Atianand et al ldquoA long noncodingRNA mediates both activation and repression of immuneresponse genesrdquo Science vol 341 no 6147 pp 789ndash792 2013
[30] Y S Mao H Sunwoo B Zhang and D L Spector ldquoDirectvisualization of the co-transcriptional assembly of a nuclearbody by noncoding RNAsrdquo Nature Cell Biology vol 13 no 1pp 95ndash101 2011
[31] M A Faghihi M Zhang J Huang et al ldquoEvidence for naturalantisense transcript-mediated inhibition of microRNA func-tionrdquo Genome Biology vol 11 no 5 article R54 2010
[32] C Gong and L E Maquat ldquoLncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 39 UTRs via Aluelemerdquo Nature vol 470 no 7333 pp 284ndash290 2011
[33] S Akira S Uematsu and O Takeuchi ldquoPathogen recognitionand innate immunityrdquo Cell vol 124 no 4 pp 783ndash801 2006
[34] R Medzhitov ldquoInflammation 2010 new adventures of an oldflamerdquo Cell vol 140 no 6 pp 771ndash776 2010
[35] K Takeda T Kaisho and S Akira ldquoToll-like receptorsrdquoAnnualReview of Immunology vol 21 pp 335ndash376 2003
[36] S Carpenter and K A Fitzgerald ldquoTranscription of inflamma-tory genes long noncoding RNA and beyondrdquo Journal of Inter-feron and Cytokine Research vol 35 no 2 pp 79ndash88 2015
[37] J Jura and A Koj Regulatory Mechanisms Controlling Inflam-mation and Synthesis of Acute Phase Proteins INTECH OpenAccess 2011
[38] C Nathan and A Ding ldquoNonresolving inflammationrdquo Cell vol140 no 6 pp 871ndash882 2010
[39] R Medzhitov and T Horng ldquoTranscriptional control of theinflammatory responserdquoNature Reviews Immunology vol 9 no10 pp 692ndash703 2009
[40] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[41] M S Hayden and S Ghosh ldquoNF-120581B in immunobiologyrdquo CellResearch vol 21 no 2 pp 223ndash244 2011
[42] Fact sheetNo 311 ldquoObesity and overweightrdquoWHO httpwwwwhointmediacentrefactsheetsfs311en
[43] R G Baker M S Hayden and S Ghosh ldquoNF-120581B inflamma-tion and metabolic diseaserdquo Cell Metabolism vol 13 no 1 pp11ndash22 2011
[44] M I Schmidt and B B Duncan ldquoDiabesity an inflamma-tory metabolic conditionrdquo Clinical Chemistry and LaboratoryMedicine vol 41 no 9 pp 1120ndash1130 2003
[45] L F Van Gaal I L Mertens and C E De Block ldquoMechanismslinking obesity with cardiovascular diseaserdquo Nature vol 444no 7121 pp 875ndash880 2006
[46] S P Weisberg D McCann M Desai M Rosenbaum R LLeibel andAW Ferrante Jr ldquoObesity is associatedwithmacro-phage accumulation in adipose tissuerdquo The Journal of ClinicalInvestigation vol 112 no 12 pp 1796ndash1808 2003
[47] U Kintscher M Hartge K Hess et al ldquoT-lymphocyte infiltra-tion in visceral adipose tissue a primary event in adipose tissueinflammation and the development of obesity-mediated insulinresistancerdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 28 no 7 pp 1304ndash1310 2008
[48] Y Lin H Lee A H Berg M P Lisanti L Shapiro and PE Scherer ldquoThe lipopolysaccharide-activated Toll-like receptor(TLR)-4 induces synthesis of the closely related receptor TLR-2in adipocytesrdquoThe Journal of Biological Chemistry vol 275 no32 pp 24255ndash24263 2000
[49] E D Rosen and B M Spiegelman ldquoAdipocytes as regulators ofenergy balance and glucose homeostasisrdquo Nature vol 444 no7121 pp 847ndash853 2006
[50] E D Rosen and O A MacDougald ldquoAdipocyte differentiationfrom the inside outrdquoNature ReviewsMolecular Cell Biology vol7 no 12 pp 885ndash896 2006
[51] P Tontonoz R A Graves A I Budavari et al ldquoAdipocyte-specific transcription factor ARF6 is a heterodimeric complexof two nuclear hormone receptors PPAR120574 and RXR120572rdquo NucleicAcids Research vol 22 no 25 pp 5628ndash5634 1994
[52] M I Lefterova Y Zhang D J Steger et al ldquoPPAR120574 and CEBPfactors orchestrate adipocyte biology via adjacent binding on agenome-wide scalerdquo Genes and Development vol 22 no 21 pp2941ndash2952 2008
[53] Y Oishi I Manabe K Tobe et al ldquoKruppel-like transcriptionfactor KLF5 is a key regulator of adipocyte differentiationrdquo CellMetabolism vol 1 no 1 pp 27ndash39 2005
[54] S E Ross N Hemati K A Longo et al ldquoInhibition of adipo-genesis by Wnt signalingrdquo Science vol 289 no 5481 pp 950ndash953 2000
[55] B Xu I GerinHMiao et al ldquoMultiple roles for the non-codingRNA SRA in regulation of adipogenesis and insulin sensitivityrdquoPLoS ONE vol 5 no 12 article e14199 2010
Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 11
[56] R B Lanz N J McKenna S A Onate et al ldquoA steroid receptorcoactivator SRA functions as an RNA and is present in an SRC-1 complexrdquo Cell vol 97 no 1 pp 17ndash27 1999
[57] S Liu R Xu I Gerin et al ldquoSRA regulates adipogenesis bymodulating p38jnk phosphorylation and stimulating insulinreceptor gene expression and downstream signalingrdquo PLoSONE vol 9 no 4 Article ID e95416 2014
[58] S Liu L Sheng H Miao et al ldquoSRA gene knockout protectsagainst diet-induced obesity and improves glucose tolerancerdquoJournal of Biological Chemistry vol 289 no 19 pp 13000ndash130092014
[59] T Xiao L Liu H Li et al ldquoLong noncoding RNAADINR regu-lates adipogenesis by transcriptionally activatingCEBP120572rdquo StemCell Reports vol 5 no 5 pp 856ndash865 2015
[60] W-J Pang L-G Lin Y Xiong et al ldquoKnockdown of PU1 ASlncRNA inhibits adipogenesis through enhancing PU1 mRNAtranslationrdquo Journal of Cellular Biochemistry vol 114 no 11 pp2500ndash2512 2013
[61] D G Tenen R Hromas J D Licht and D-E Zhang ldquoTran-scription factors normal myeloid development and leukemiardquoBlood vol 90 no 2 pp 489ndash519 1997
[62] F Wang and Q Tong ldquoTranscription factor PU1 is expressed inwhite adipose and inhibits adipocyte differentiationrdquo AmericanJournal of Physiology-Cell Physiology vol 295 no 1 pp C213ndashC220 2008
[63] N Wei Y Wang R-X Xu et al ldquoPU1 antisense lncRNAagainst its mRNA translation promotes adipogenesis in porcinepreadipocytesrdquoAnimalGenetics vol 46 no 2 pp 133ndash140 2015
[64] L Sun L A Goff C Trapnell et al ldquoLong noncoding RNAsregulate adipogenesisrdquo Proceedings of the National Academy ofSciences of the United States of America vol 110 no 9 pp 3387ndash3392 2013
[65] J Nedergaard T Bengtsson and B Cannon ldquoUnexpected evi-dence for active brown adipose tissue in adult humansrdquo Amer-ican Journal of Physiology-Endocrinology and Metabolism vol293 no 2 pp E444ndashE452 2007
[66] A M Cypess S Lehman GWilliams et al ldquoIdentification andimportance of brown adipose tissue in adult humansrdquoThe NewEngland Journal ofMedicine vol 360 no 15 pp 1509ndash1517 2009
[67] J Nedergaard and B Cannon ldquoThe changed metabolic worldwith human brown adipose tissue therapeutic visionsrdquo CellMetabolism vol 11 no 4 pp 268ndash272 2010
[68] X-Y Zhao S Li G-X Wang Q Yu and J D Lin ldquoA long non-coding RNA transcriptional regulatory circuit drives thermo-genic adipocyte differentiationrdquo Molecular Cell vol 55 no 3pp 372ndash382 2014
[69] L H You L J Zhu L Yang et al ldquoTranscriptome analysisreveals the potential contribution of long noncoding RNAs tobrown adipocyte differentiationrdquo Molecular Genetics and Gen-omics vol 290 no 5 pp 1659ndash1671 2015
[70] J R Alvarez-Dominguez Z Bai D Xu et al ldquoDe novo recon-struction of adipose tissue transcriptomes reveals long non-coding RNA regulators of brown adipocyte developmentrdquo CellMetabolism vol 21 no 5 pp 764ndash776 2015
[71] R Gernapudi B Wolfson Y Zhang et al ldquoMicroRNA 140 pro-motes expression of long noncoding RNA NEAT1 in adipoge-nesisrdquo Molecular and Cellular Biology vol 36 no 1 pp 30ndash382016
[72] V Ratziu ldquoStarting the battle to control non-alcoholic steato-hepatitisrdquoThe Lancet vol 385 no 9972 pp 922ndash924 2015
[73] S Singh A M Allen Z Wang L J Prokop M H MuradandR Loomba ldquoFibrosis progression in nonalcoholic fatty livervs nonalcoholic steatohepatitis a systematic review and meta-analysis of paired-biopsy studiesrdquoClinical Gastroenterology andHepatology vol 13 no 4 pp 643ndash654e9 2015
[74] R Loomba and A J Sanyal ldquoThe global NAFLD epidemicrdquoNature Reviews Gastroenterology and Hepatology vol 10 no 11pp 686ndash690 2013
[75] WHO ldquoDiabetesrdquo Fact Sheet 312 2016 httpwwwwhointmediacentrefactsheetsfs312en
[76] A E Kitabchi G E Umpierrez J M Miles and J N FisherldquoHyperglycemic crises in adult patients with diabetesrdquoDiabetesCare vol 32 no 7 pp 1335ndash1343 2009
[77] American Diabetes Association ldquoDiagnosis and classificationof diabetes mellitusrdquo Diabetes Care vol 33 supplement 1 ppS62ndashS69 2010
[78] U Riserus W C Willett and F B Hu ldquoDietary fats and pre-vention of type 2 diabetesrdquo Progress in Lipid Research vol 48no 1 pp 44ndash51 2009
[79] M Prentki andC J Nolan ldquoIslet120573 cell failure in type 2 diabetesrdquoJournal of Clinical Investigation vol 116 no 7 pp 1802ndash18122006
[80] J E Shaw R A Sicree and P Z Zimmet ldquoGlobal estimates ofthe prevalence of diabetes for 2010 and 2030rdquoDiabetes Researchand Clinical Practice vol 87 no 1 pp 4ndash14 2010
[81] GM KuH Kim IW Vaughn et al ldquoResearch resource RNA-seq reveals unique features of the pancreatic 120573-cell transcrip-tomerdquo Molecular Endocrinology vol 26 no 10 pp 1783ndash17922012
[82] I Moran I Akerman M Van De Bunt et al ldquoHuman 120573cell transcriptome analysis uncovers lncRNAs that are tissue-specific dynamically regulated and abnormally expressed intype 2 diabetesrdquo Cell Metabolism vol 16 no 4 pp 435ndash4482012
[83] Y S Cho C-H Chen C Hu et al ldquoMeta-analysis of genome-wide association studies identifies eight new loci for type 2diabetes in east AsiansrdquoNatureGenetics vol 44 no 1 pp 67ndash722012
[84] V Senee C Chelala S Duchatelet et al ldquoMutations in GLIS3are responsible for a rare syndrome with neonatal diabetesmellitus and congenital hypothyroidismrdquo Nature Genetics vol38 no 6 pp 682ndash687 2006
[85] J R Huyghe A U Jackson M P Fogarty et al ldquoExomearray analysis identifies new loci and low-frequency variantsinfluencing insulin processing and secretionrdquo Nature Geneticsvol 45 no 2 pp 197ndash201 2013
[86] Y Gosmain L S Katz M H Masson C Cheyssac C Poissonand J Philippe ldquoPax6 is crucial for 120573-cell function insulin bio-synthesis and glucose-induced insulin secretionrdquo MolecularEndocrinology vol 26 no 4 pp 696ndash709 2012
[87] J Fadista P Vikman E O Laakso et al ldquoGlobal genomicand transcriptomic analysis of human pancreatic islets revealsnovel genes influencing glucose metabolismrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 111 no 38 pp 13924ndash13929 2014
[88] Z Fu E R Gilbert and D Liu ldquoRegulation of insulin synthesisand secretion and pancreatic beta-cell dysfunction in diabetesrdquoCurrent Diabetes Reviews vol 9 no 1 pp 25ndash53 2013
[89] B Yan Z-F Tao X-M Li H Zhang J Yao and Q JiangldquoAberrant expression of long noncoding RNAs in early diabeticretinopathyrdquo Investigative Ophthalmology amp Visual Science vol55 no 2 pp 941ndash951 2014
12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Mediators of Inflammation
[90] O Wapinski and H Y Chang ldquoLong noncoding RNAs andhuman diseaserdquo Trends in Cell Biology vol 21 no 6 pp 354ndash361 2011
[91] J-Y Liu J Yao X-M Li et al ldquoPathogenic role of lncRNA-MALAT1 in endothelial cell dysfunction in diabetes mellitusrdquoCell Death and Disease vol 5 no 10 Article ID e1506 2014
[92] B Yan J Yao J-Y Liu et al ldquoLncRNA-MIAT regulates micro-vascular dysfunction by functioning as a competing endoge-nous RNArdquo Circulation Research vol 116 no 7 pp 1143ndash11562015
[93] I Zachary andRDMorgan ldquoTherapeutic angiogenesis for car-diovascular disease biological context challenges prospectsrdquoHeart vol 97 no 3 pp 181ndash189 2011
[94] G-Z Qiu W Tian H-T Fu C-P Li and B Liu ldquoLongnoncoding RNA-MEG3 is involved in diabetes mellitus-related microvascular dysfunctionrdquo Biochemical and Biophysi-cal Research Communications vol 471 no 1 pp 135ndash141 2016
[95] M R Abid S Guo T Minami et al ldquoVascular endothelialgrowth factor activates PI3KAktforkhead signaling in endo-thelial cellsrdquo Arteriosclerosis Thrombosis and Vascular Biologyvol 24 no 2 pp 294ndash300 2004
[96] G Carter B Miladinovic A A Patel L Deland S Mastoridesand N A Patel ldquoCirculating long noncoding RNA GAS5 levelsare correlated to prevalence of type 2 diabetes mellitusrdquo BBAClinical vol 4 pp 102ndash107 2015
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom





























