Embed Size (px)
Citation preview

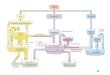
Endomembranee smistamento delle proteine

Le proteine mitocondriali sono importate: nella matrice nello spazio intermembrana.grazie a segnali di indirizzamento, posti nella porzione amino-terminale.
Nella matrice: il peptide segnale si lega ad un recettore sulla membrana mitocondriale esterna e la proteina viene traslocata grazie alla presenza di un secondo traslocatore sulla membrana interna: si crea un punto di contatto tra le due membrane.
Le proteine sono traslocate “non ripiegate”. Intervengono in questo processo le proteine chaperonine, come hsp70, che mantengono le proteine srotolate. L’idrolisi dell’ATP viene usata per staccare l’hsp70 e fornire energia motrice per importare la proteina attraverso il traslocatore.

Organizzazione dei mitocondri
Sequenza segnale per l’importazione di proteine mitocondriali

Tre proteine traslocatrici mitocondriali

Importazione di proteine nei mitocondri, la sequenza segnale è riconosciuta dalla proteina TOM.

Ruolo dell’energia nell’importazione di proteine nella matrice mitocondriale
1) La proteina hsp70 citosolica attaccata è rilasciata dalla proteina in un passaggio che Diprende Dall’idrolisi di ATP
2) Traslocazione nella matrice grazie ad un gradiente protonico
3) l’hsp mitocondriale si lega alla catena polipetidica in regioni strategiche trascinandola Nella Matrice ed è poi rimossa grazie ad idrolisi di ATP

la sequenza segnale
una sequenza di aa idrofobica dopo il peptide segnale, che funziona da nuovo peptide segnale quando il primo è stato tagliato via dalla peptidasi (meccanismo simile alla traslocazione nell’ER)
La traslocazione delle proteine nella membrana mitocondriale interna e nello spazio intermembrana richiede:

Endomembrane della cellula eucariotica

Membrane biologiche e loro interazioni

Nucleo
PoroNucleare
RE




Meccanismo di trasporto attivo attraverso i pori nucleari

Traffico di Materiale attraverso
I Pori Nucleari

Reticolo
Endoplasmatico




LE MEMBRANE DEL SER STBILISCONO CON GRANDE EFFICIENZA
GRADIENTI DI CA++
CHE SVOLGONO UN RUOLO DECISIVO NELLA
TRASDUZIONE DEI SEGNALI
NELLO STIMOLO NERVOSO E
NELLA CONTRAZIONE MUSCOLARE

IL SER E’ RESPONSABILE DELLA SINTESI DI
TRIACILGLICEROLI CHE VENGONO ACCUMULATI NEL LUME,
CON PRODUZIONE DI GOCCIOLINE LIPIDICHE.
NEGLI ADIPOCITI LE GOCCIOLINE LIPIDICHE OCCUPANO QUASI TUTTO LO SPAZIO DEL CITOPLASMA

IL SER PARTECIPA ALLA SINTESI DI STEROLI
COME IL COLESTEROLO
IL COLESTEROLO VIENE SINTETIZZATO ATTRAVERSO UNA VIA METABOLICA A PIU’ STADI ALLA QUALE PARTECIPANO ANCHE ENZIMI PRESENTI NEL CITOSOL,
OLTRE A QUELLI DEL SER

LA SPECIE UMANA SINTETIZZA UNA GRAN PARTE DEL SUO
COLESTEROLO
TUTTE LE CELLULE SONO CAPACI DI PRODURLO, MA LA MAGGIOR PARTE DI ESSO VIENE SINTETIZZATO DALLE
CELLULE EPATICHE
IL FEGATO UTILIZZA IL COLESTEROLO PER FORMARE
GLI ACIDI BILIARI

NELLA CORTECCIA SURRENALE E NELLE GONADI (REGIONE INTERSTIZIALE DEL
TESTICOLO)
IL COLESTEROLO E’ UTILIZZATO NELLA
PRODUZIONE DEGLI ORMONI STEROIDEI

IL SER INTERVIENE NEL PRODUZIONE DEGLI ORMONI STEROIDEI
NEI MITOCONDRI VIENE SCISSA LA CATENA LATERALE
DEL COLESTEROLO, IMPORTATO DAL CITOSOL
IL PRODOTTO COSI’ FORMATOLASCIA I MITOCONDRI E PASSA NEL SER DOVE VIENE
DI NUOVO MODIFICATO.
INFINE, TALE PRODOTTO DAL SER TORNA DI NUOVO NEI MITOCONDRI PER LE M0DIFICAZIONI FINALI
COSI’ QUESTI DUE ORGANELLI GIOCANO UNA SORTA DI “PALLA A VOLO” PER PRODURRE L’ORMONE
STEROIDEO

IL SER SINTETIZZA I FOSFOLIPIDI DELLE MEMBRANE CELLULARI
IL FOSFOLIPIDE PRINCIPALE PRODOTTO E’
FOSFATIDILCOLINA O LECITINA
PUO’ ESSERE FORMATO IN TRE PASSAGGI DA COLINA, DUE ACIDI
GRASSI E GLICEROLO FOSFATO


IL SER CONTIENE UN ENZIMA GLUCOSIO-6-FOSFATASI
G6Pase
ESSENZIALE PER L’OMEOSTASI DEL GLUCOSIO
NEL SANGUE,L’ENZIMA E’ PRESENTE SOLO
NELLE CELLULE PARENCHIMALI DEL FEGATO

LA DETOSSIFICAZIONE E’ SVOLTA DA UN SISTEMA DI ENZIMI CHE TRASFERISCONO
L’OSSIGENO (MONOSSIGENASI) E IL CUI COMPONENTE PRINCIPALE E’ IL
CITOCROMO P-450
IL SISTEMA DELLE MONOSSIGENASI P-450 E’ FORMATO DA NUMEROSI ISOENZIMI (50-
100) DEL CITOCROMO P-450 (EMO-PROTEINA DI TIPO b),
DA UNA SINGOLA REDUTTASI E FOSFOLIPIDE.

QUESTI ENZIMI SONO PROTEINE INTEGRALI DI MEMBRANA E SONO PREMINENTI NEL SER, MA PRESENTI
ANCHE NEI MITOCONDRI
IL CITOCROMO P-450 E’ UBIQUITARIO, MA SI TROVA IN CONCENTRAZIONI MOLTO
ELEVATE NEL SER DELLE CELLULE EPATICHE
RAPPRESENTA CIRCA IL 20% DELLE PROTEINE DEL ER E IL 2-3% DELLE
PROTEINE TOTALI DELLA CELLULA


QUESTI ENZIMI SONO CAPACI DI OSSIDARE MIGLIAIA DI COMPOSTI
IDROFOBICI DIFFERENTI CHE CONVERTONO IN DERIVATI PIU’
IDROFILICI E, QUINDI, PIU’ FACILMENTE SECRETI
I SUBSTRATI DI TALI ENZIMI SONO IN GRADO DI INDURRE PROLIFERAZIONE
DEL SER E CONSEGUENTE AUMENTO DELL’ATTIVITA’ DEGLI ENZIMI

SONNIFERI A BASE DI FENOBARBITAL (C OSI’ COME CENTINAIA DI ALTRE SOSTANZE CHIMICHE) INDUCONO
PROLIFERAZIONE DEL SER E AUMENTO DELL’ATTIVITA’ DELLE MONOSSIDASI
NELLE CELLULE EPATICHE
IL CONSEGUENTE AUMENTO DELLA CAPACITA’ DI DEGRADARE IL
FENOBARBITAL SPIEGA PER QUALE MOTIVO I CONSUMATORI DI SONNIFERO
DEVONO ASSUMERNE DOSI SEMPRE PIU’ ELEVATE

PERTANTO L’ASSUNZIONE CRONICA DI FENOBARBITAL (O ALTRE SOSTANZE)
AUMENTA L’EFFICIENZA DI DEGRADAZIONE, DA PARTE DEL FEGATO,
DI MOLTI ALTRI FARMACI, TRA CUI SOSTANZE TERAPEUTICAMENTE UTILI,
COME ANTIBIOTICI, STEROIDI, ANTICOAGULANTI E NARCOTICI
COMUNQUE L’AUMENTATO SVILUPPO DEL SER NON COMPORTA AUMENTO
EQUIVALENTE DI TUTTI GLI ENZIMI

MENTRE L’AZIONE DEL COMPLESSO P-450 E’ NEL
COMPLESSO FAVOREVOLE DAL PUNTO DI VISTA
FISIOLOGICO, CERTI EVENTI METABOLICI BASATI SUL P-
450 POSSONO PORTARE A SERIE PATOLOGIE

RETICOLO ENDOPLASMATICO RUGOSO
RER


Sintesi di proteine a livello del RER

Sintesi proteica a livello del Reticolo endoplasmatico rugoso

Modalità di inserzione di una proteina nel bilayer fosfolipidico del RER

Modalità di inserzione di una proteina nel bilayer fosfolipidico del RER

Conclusione:Le modalità di inserzione delleproteine nei bilayer fosfolipidicivengono giàdefinite nel RER.

Glicosilazione a livello del RER


Membrane biologiche e loro interazioni


Il percorso delle proteine destinate alla secrezione



Vescicole di trasporto e proteine associate

TrafficoVescicolareTra RE e
Apparato diGolgi

Meccanismo usato per trattenere le proteine residenti nel RER

Apparato di Golgi

Compartimentalizzazione
funzionale del Golgi

Elementi di Transizione e del reticolo cis del Golgi


Trasporto selettivo mediato da vescicole rivestite

Modello del meccanismo di approdo delle vescicole di trasporto

Fusione di vescicole di trasporto


Ruolo delle proteine di fusione nell’internalizzazione di virus


Modello di recupero per proteine residenti nell’ER

LisosomaI lisosomi: punti di incontro ove convergono le sostanze che devono essere digerite.
Enzimi lisosomali: idrolasi lisosomali
Vie per arrivare al lisosoma:
Autofagia (ER si trasforma in lisosoma)
Fagocitosi
Endosoma precoci e tardivi
Alcune proteine sono trasportate nel lisosoma direttamente con l’uso di un segnale lisosomale: KFERQ (Lys, Phe, Glut, Arg, glutammina)

Le idrolasi lisosomiali sono caratterizzate dalla presenza di un gruppo specifico, il mannosio-6-fosfato (M6P), che serve a farle riconoscere da un recettore specifico presente sul trans-Golgi.
L’M6P è aggiunto agli oligosaccaridi legati all’asparagina, nel reticolo cis del Golgi.
Il recettore è una proteina transmembrana localizzata sul trans Golgi. L’unione con l’idrolasi inizia la formazione di vescicole e si verifica a pH 7. Le idrolasi si dissociano dai recettori negli endosomi tardivi a pH 6.
I recettori sono riciclati verso il trans Golgi.


Riconoscimento di una idrolasi lisosomale

Trasporto di idrolasi lisosomali di nuova sintesi ai lisosomi

Tre vie di degradazione dei lisosomi

Malattie da accumulo lisosomale: le idrolasi lisosomali sono alterate per difetti genetici recessivi.
Malattia di Hurler
I-cell disease
I substrati delle idrolasi si accumulano non digeriti nel lisosomi, per cui si formano inclusioni nelle cellule.
Le idrolasi alterate sfuggono ai lisosomi, sono secrete dalla via di default e si accumulano nel sangue: il difetto è dovuto alla mancanza o al difetto di una fosfotransferasi GlcNAc. Le idrolasi sono secrete.
Non funziona la via di scavenger (raccolta dei rifiuti) di endocitosi e si accumulano.
Nel fegato via alternativa.

Endocitosi: Dalla superficie cellulare verso i lisosomi.
Fagocitosi di grandi particelle (250 nm) svolta da cellule specializzate tramite fagosomi.
Le particelle si devono legare ai fagociti: recettori attivati scatenano la risposta . Normalmente sono gli anticorpi che riconoscono le particelle estranee e le presentano ai fagociti.
Proteine Fc o complemento.

Fagocitosi da parte di un
macrofago di due globuli
rossi chimicamente
alterati
Frecce rosse = pseudopodi

Fosse rivestite di clatrina: 2% dell’area di membrana
La clatrina forma un cesto o una gabbia sotto la membrana e dà origine ad una vescicola rivestita di clatrina: è un processo rapido e continuo in molti tipi cellulari.
Le vescicole si fondono poi negli endosomi precoci.
Servono per l’endocitosi in fase fluida e per l’assunzione di specifiche macromolecole dall’esterno.
Questo processo è chiamato endocitosi mediata da recettori: processo selettivo che permette di assumere grandi quantità di macromolecole. Es. trasporto del colesterolo. Le cellule assumono colesterolo che serve per assemblare le membrane.
Se si blocca l’assunzione, il colesterolo si accumula nel sangue e può formare placche aterosclerotiche.
Il colesterolo è trasportato nel sangue associato a proteine LDL (lipoproteine a basse densità).


Ogni LDL contiene 1500 mol colesterolo esterificato ad acidi grassi

Le cellule presentano dei recettori per le LDL, che si associano alle zone di membrana ricche in clatrina. L’LDL si lega al recettore e si formano vescicole rivestite che sono internalizzate.
Le vescicole perdono il rivestimento in clatrina e si fondono negli endosomi precoci. Le LDL sono portate agli endosomi tardivi ed ai lisosomi, dove gli esteri di colesterolo sono idrolizzati ed il colesterolo liberato per l’incorporazione nelle membrane.
Se c’è troppo colesterolo, viene spenta la sintesi di colesterolo e anche di recettori per LDL, per non importarlo più e così il colesterolo si accumula nel sangue.
Geni difettosi per le LDL: mancata assunzione del colesterolo, aterosclerosi in età giovanile.

Struttura della clatrina



Interazione clatrina-adattatore

Recettori delle LDL normali e mutanti

I recettori per le LDL sono riciclati dagli endosomi e riportati in membrana per essere riutilizzato.

Deposito di proteine della membrana plasmatica in endosomi riciclati


Endocitosi di LDL mediata da recettori

Endocitosi mediata da recettori

Le vie meglio conosciute di smistamento delle proteine
nel reticolo trans del Golgi

Vie di endocitosi costitutiva e regolata

Compartimenti intracellulari coinvolti nella vie secretorie e endocitotica