Embed Size (px)
Citation preview

Response of ground beetle (Carabidae) assemblages to logging
history in northern hardwood–hemlock forests
Erika F. Latty a,c,*, Shahla M. Werner b,d, David J. Mladenoff a,Kenneth F. Raffa b, Theodore A. Sickley a
a Department of Forest Ecology and Management, University of Wisconsin-Madison, 1630 Linden Drive, Madison, WI 53706-1598, USAb Department of Entomology, University of Wisconsin-Madison, 345 Russell Laboratories, 1630 Linden Drive, Madison, WI 53706-1598, USA
c Biology Department, Hollins University, P.O. Box 9615, Roanoke, VA 24020, USAd Pennsylvania Department of Conservation and Natural Resources, 208 Airport Drive, 2nd Floor, Middletown, PA 17057-5027, USA
Received 10 March 2005; received in revised form 7 October 2005; accepted 7 October 2005
www.elsevier.com/locate/foreco
Forest Ecology and Management 222 (2006) 335–347
Abstract
We quantified differences in ground beetle (Coleoptera: Carabidae) communities in relation to forest management practices and historic forest
cover changes in hardwood–hemlock forests of the north central United States. Beetles were sampled with pitfall traps in 1996 and 1997 and
compared among three forest types: old-growth, and post-logging uneven- and even-aged forests. Non-metric multidimensional scaling ordination
was used to assess compositional differences among forest types for 39 carabid species (43,483 individuals), which revealed distinct differences in
beetle assemblages among forest types. Coarse woody debris, snag volume, gap area, understory vegetation and forest floor depth were
significantly correlated with ordination axes suggesting that these variables are critical in structuring the beetle communities. Several of the species
significantly associated with old-growth forests, like Carabus sylvosus, are known to favor forest habitats, whereas species commonly found in
open habitats, such as Carabus nemoralis, had stronger affiliations with managed forests. Comparisons of northern hardwood–hemlock forest
distribution from pre-Euroamerican settlement with current distributions reconstructed using the Forest Inventory and Analysis database indicate
that old-growth habitat has declined to <1% of its original extent in this region. At the landscape level, these data suggest that the abundance of
carabid species that prefer old-growth forest conditions has undergone broad-scale decline.
# 2005 Elsevier B.V. All rights reserved.
Keywords: Ground beetles; Northern hardwood forest; Forest management; Old-growth; Habitat heterogeneity; Forest landscape change
1. Introduction
The majority of the world’s landscapes are increasingly
devoted to human purposes, including agriculture, settlement
and industry (Dale et al., 2000). As these landscapes are
transitioning, ecologists are recognizing the significance of
these changes for a variety of ecosystem parameters (Turner
et al., 1990; Mladenoff and Pastor, 1993; Houghton and
Hackler, 2000). Such pervasive land use change can be
expected to have repercussions for the distribution and
abundance of a wide array of plant and animal species (Foster,
1992; Mladenoff et al., 1997; Ribera et al., 2001). Forests have
* Corresponding author. Tel.: +1 540 362 6415; fax: +1 540 362 6629.
E-mail addresses: [email protected], [email protected] (E.F. Latty),
[email protected], [email protected] (S.M. Werner), djmla-
[email protected] (D.J. Mladenoff), [email protected] (K.F. Raffa),
[email protected] (T.A. Sickley).
0378-1127/$ – see front matter # 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.foreco.2005.10.028
undergone particularly large changes (Houghton, 1995), and
within the United States overall forested area continues to
decrease despite significant reforestation in the eastern portion
of the country (Smith et al., 1993; Dale et al., 2000).
Detailed assessments of forest change within the Great Lakes
forests indicate that this landscape previously dominated by old-
growth hemlock and hardwood forests is now largely composed
of early successional second-growth hardwoods, with very few
small old-growth patches (Mladenoff et al., 1993). The majority
of these changes in forest cover were attributable to destructive
logging and fires around the turn of the 20th century (Stearns,
1990; Mladenoff and Pastor, 1993). Following broad-scale
afforestation in the mid 1900s forest management has again
become the driving force of landscape change.
In addition to changed landscape pattern, the historical and
recent logging practices have maintained younger forests,
reduced conifers and simplified structure including reductions
in coarse woody debris loads (Goodburn and Lorimer, 1998;

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347336
Duval and Grigal, 1999; Crow et al., 2002) and aboveground
living biomass (Hale et al., 1999; Ziegler, 2000). Modifications
of forest structure and vegetation are likely to have ramifica-
tions for faunal distributions (Niemela, 1997), which depend
upon key habitat features for shelter, nesting sites and foraging
areas (Harmon et al., 1986; Whittam et al., 2002). For example,
silvicultural practices have caused changes to forest inverte-
brate communities by altering microhabitat parameters like
coarse woody debris and local tree species (Niemela, 1997) and
in some cases, resulted in a loss of species diversity or altered
species composition (Niemela et al., 1993; Petterson et al.,
1995; Martikainen et al., 2000).
Developing an understanding of the collective response of
species to changing environmental conditions (Janzen, 1985;
Southwood, 1988; Ribera et al., 2001) inform conservation
practices that seek to maintain functional capabilities of
ecosystems. The composition of species that characterize a
given community is ultimately defined by more than simple
lists of species thus community studies should also incorporate
information about shared ecological characteristics (Whittaker,
1975; Cole et al., 2002). One way to detect ecological
similarities among species is to sort them into groups based on
life history attributes, requiring a broad understanding of the
ecology of the species (Ribera et al., 2001; Cole et al., 2002).
Such information exists for carabids (Lindroth, 1969, 1974;
Forsythe, 1987) whose sensitivity to environmental change is
widely recognized (Refseth, 1980; Gardner, 1991; Rieske and
Buss, 2001).
The majority of invertebrate community analyses have been
conducted in Europe (Niemela et al., 1993; Martikainen et al.,
2000; Jukes et al., 2001; Magura, 2002) where broad interest in
insect conservation exists and red lists of insects have been
created to integrate invertebrates in conservation planning
(Jonsell et al., 1998). For example, the UK Forestry
Commission’s Biodiversity Research Programme requires that
invertebrates be incorporated in land planning measures
(Hodge et al., 1998). In contrast, most conservation planning
in the United States rarely include invertebrates despite their
sensitivity to habitat changes and roles in vital ecosystem
processes like decomposition (Mattson and Addy, 1975), fire
cycles (McCullough et al., 1998), pollination (Westman, 1990)
and as a food source for other animals (Rosenberg et al., 1986).
Moreover, arthropods, most of which are insects, are the most
diverse and widespread of all animal phyla (Campbell, 1990).
The omission of insects from most United States conserva-
tion planning is likely due in part to a lack of regionally specific
insect data for all United States ecosystems. Carabids are the
third largest beetle family in North America with about 2600
species in 189 genera (Triplehorn and Johnson, 2005), making
this order an ideal focus for ecological studies. However, in
Great Lakes forests in particular, relatively few studies have
been conducted on habitat associations of carabids, and how
assemblages respond to widespread human-induced distur-
bances of the region, such as logging (but see Liebherr and
Mahar, 1979; Epstein and Kuhlman, 1990; Werner and Raffa,
2000; Vance and Nol, 2003). In addition, attempts to correlate
carabid assemblages in the greater United States with
ecologically meaningful management units, based on land-
scape scale parameters, such as climate and geological features,
have been only partially successful (Rykken et al., 1997; Moore
et al., 2004). Part of the challenge in relating invertebrate
communities to ecosystem management is attributable to the
dissimilar scales at which ecosystem processes occur and
invertebrate communities are defined. Ultimately the chosen
scale must have ecological meaning for diverse groups of taxa
in order to retain practicality for land managers. Using forest
stands, as the unit at which to implement invertebrate studies is
a reasonable spatial scale. The majority of United States forests
are managed at this scale and the management history can play
an important role in determining carabid composition (Werner
and Raffa, 2000; Jukes et al., 2001).
The purpose of this study was to understand the effects of
forest management practices on carabid assemblages of Great
Lakes forests. We addressed the following hypotheses. (1)
There are differences in communities of ground dwelling
Carabidae among old-growth, and younger uneven- and even-
aged northern hardwood–hemlock forests. (2) Differences in
beetle assemblages among forest types can be explained by
environmental variables including forest structure, vegetation
and soil characters. (3) If differences in beetle communities do
exist then changes in historical forest cover in northern
Wisconsin and Upper Michigan have resulted in a landscape
that favors carabid communities typical of younger forests.
Information gathered by this study can guide conservation
actions in terms of protecting beetle assemblages found in old-
growth forests, a rare landscape element, and by providing
necessary data for incorporation in forest management guide-
lines that currently overlook beetle communities in their
formulation.
2. Materials and methods
2.1. Study areas
Carabids were sampled in 22 forest sites in northern
Wisconsin and the Upper Peninsula of Michigan, USA, located
from 458410 to 468180 north latitude and 888590 to 898440 west
longitude (Fig. 1). This study was conducted as part of a
multidisciplinary effort examining the effects of forest
management practices on the flora, fauna and ecosystem
characteristics of the region (Bockheim, 1997; Goodburn and
Lorimer, 1998; Campbell and Gower, 2000; Werner and Raffa,
2000; Miller et al., 2002; Scheller and Mladenoff, 2002). Sites
were chosen to minimize floristic, soil and climatic variability
and most stands were located on the Winegar terminal moraine
(Goodburn and Lorimer, 1998). Average monthly temperatures
range from –12 8C (January) to 19 8C (July) and average annual
precipitation is 850 mm (Goodburn, 1996).
Sites are dominated by Acer saccharum Marsh. (sugar
maple), Betula alleghaniensis Britton (yellow birch), Tsuga
canadensis Carr. (eastern hemlock) and Tilia americana L.
(basswood), and other minor tree species. Three forest types
were represented: (1) managed, uneven-aged, n = 9 sites;
(2) managed, even-aged, n = 5 sites and (3) unmanaged,
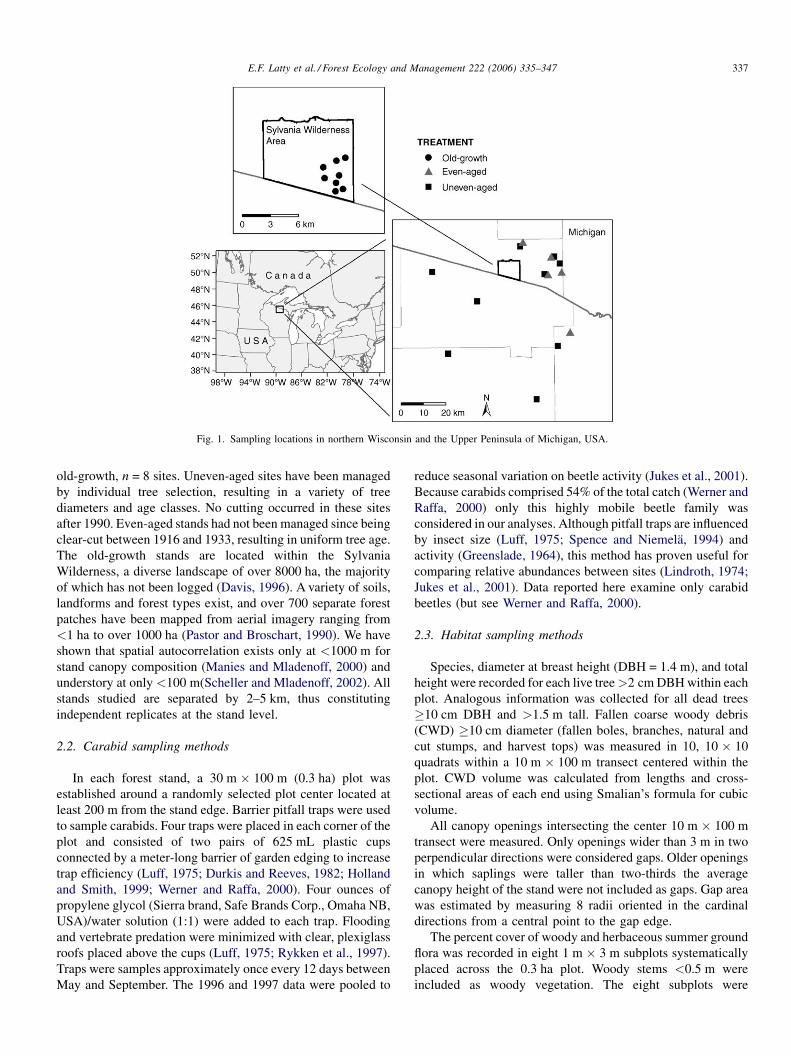
E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347 337
Fig. 1. Sampling locations in northern Wisconsin and the Upper Peninsula of Michigan, USA.
old-growth, n = 8 sites. Uneven-aged sites have been managed
by individual tree selection, resulting in a variety of tree
diameters and age classes. No cutting occurred in these sites
after 1990. Even-aged stands had not been managed since being
clear-cut between 1916 and 1933, resulting in uniform tree age.
The old-growth stands are located within the Sylvania
Wilderness, a diverse landscape of over 8000 ha, the majority
of which has not been logged (Davis, 1996). A variety of soils,
landforms and forest types exist, and over 700 separate forest
patches have been mapped from aerial imagery ranging from
<1 ha to over 1000 ha (Pastor and Broschart, 1990). We have
shown that spatial autocorrelation exists only at <1000 m for
stand canopy composition (Manies and Mladenoff, 2000) and
understory at only <100 m(Scheller and Mladenoff, 2002). All
stands studied are separated by 2–5 km, thus constituting
independent replicates at the stand level.
2.2. Carabid sampling methods
In each forest stand, a 30 m � 100 m (0.3 ha) plot was
established around a randomly selected plot center located at
least 200 m from the stand edge. Barrier pitfall traps were used
to sample carabids. Four traps were placed in each corner of the
plot and consisted of two pairs of 625 mL plastic cups
connected by a meter-long barrier of garden edging to increase
trap efficiency (Luff, 1975; Durkis and Reeves, 1982; Holland
and Smith, 1999; Werner and Raffa, 2000). Four ounces of
propylene glycol (Sierra brand, Safe Brands Corp., Omaha NB,
USA)/water solution (1:1) were added to each trap. Flooding
and vertebrate predation were minimized with clear, plexiglass
roofs placed above the cups (Luff, 1975; Rykken et al., 1997).
Traps were samples approximately once every 12 days between
May and September. The 1996 and 1997 data were pooled to
reduce seasonal variation on beetle activity (Jukes et al., 2001).
Because carabids comprised 54% of the total catch (Werner and
Raffa, 2000) only this highly mobile beetle family was
considered in our analyses. Although pitfall traps are influenced
by insect size (Luff, 1975; Spence and Niemela, 1994) and
activity (Greenslade, 1964), this method has proven useful for
comparing relative abundances between sites (Lindroth, 1974;
Jukes et al., 2001). Data reported here examine only carabid
beetles (but see Werner and Raffa, 2000).
2.3. Habitat sampling methods
Species, diameter at breast height (DBH = 1.4 m), and total
height were recorded for each live tree >2 cm DBH within each
plot. Analogous information was collected for all dead trees
�10 cm DBH and >1.5 m tall. Fallen coarse woody debris
(CWD) �10 cm diameter (fallen boles, branches, natural and
cut stumps, and harvest tops) was measured in 10, 10 � 10
quadrats within a 10 m � 100 m transect centered within the
plot. CWD volume was calculated from lengths and cross-
sectional areas of each end using Smalian’s formula for cubic
volume.
All canopy openings intersecting the center 10 m � 100 m
transect were measured. Only openings wider than 3 m in two
perpendicular directions were considered gaps. Older openings
in which saplings were taller than two-thirds the average
canopy height of the stand were not included as gaps. Gap area
was estimated by measuring 8 radii oriented in the cardinal
directions from a central point to the gap edge.
The percent cover of woody and herbaceous summer ground
flora was recorded in eight 1 m � 3 m subplots systematically
placed across the 0.3 ha plot. Woody stems <0.5 m were
included as woody vegetation. The eight subplots were

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347338
averaged to determine mean cover values per plot. Vegetation
was grouped as ferns, grasses, herbs, Lycopodia, shrubs and tree
seedlings. Goodburn (1996) and Goodburn and Lorimer (1998)
provide detailed descriptions of the methodology for assessing
structural components of the forests.
Soil sampling occurred at 10 random points within each plot,
using a 20 cm diameter circular metal frame, from which the
forest floor was removed, and a 5 cm bucket auger was used to
sample the upper 30 cm of the mineral soil. Forest floor
measurements were made at separate soil pits established at one
of the 10 sampling points. Samples were processed for particle-
size distribution, sand fractionation, calcium content, organic
carbon content, nitrogen content and cation exchange capacity
(Bockheim, 1997).
2.4. Ecological groups
Each carabid species was assigned to one guild membership
within three ecological groups based on literature reviews
(Lindroth, 1969; LaRochelle, 1972; Spence, 1990; Kavanaugh,
1992; Niemela et al., 1992, 1993). The three ecological groups
were defined by habitat preferences, diet and dispersal
strategies. Habitat preferences were classified as (1) forest
specialist, (2) open habitat specialist or (3) generalist, species
with no known specific habitat requirements. Dispersal
strategies were defined by wing development and categorized
as (1) monomorphic macropterous species that disperse by
flying, (2) monomorphic brachypterous species that disperse by
walking or (3) dimorphic where macropterous and brachypter-
ous individuals exist within the same populations. Feeding
habits were categorized as (1) carnivorous or (2) omnivorous.
Each species was also identified as native or introduced.
2.5. Past and present regional land cover
We estimated the amount of hardwood–hemlock forest prior
to Euroamerican settlement (ca. 1850) and compared this to
current (ca. 1990) estimates. Historical forest cover was
reconstructed using datasets based on the General Land Office’s
Public Lands Survey (PLS) records for Upper Michigan
(Comer et al., 1995) and northern Wisconsin (Schulte et al.,
2002). To standardize the classification schemes between
states, we aggregated all land cover types into two categories,
northern hardwood–hemlock forest and other. In Wisconsin, the
northern hardwood–hemlock cover type was defined as those
cover types in which sugar maple, eastern hemlock, yellow
birch and/or American basswood composed >50% of the
relative basal area of a PLS section (2.6 km2). The historic data
from Michigan are qualitative and cover types were typically
determined based on frequency of occurrence of a given species
(Comer et al., 1995). Therefore, in Michigan, we defined
northern hardwood–hemlock as cover types containing the
same tree species mixtures as those used to define northern
hardwood–hemlocks in Wisconsin.
The reclassified data from the two states were summarized to
a common scale, that of the PLS section. The majority land
cover class, northern hardwood–hemlocks or other, was
assigned to each PLS section. To determine the overall area
dominated by northern hardwood–hemlock forests in both
states, the reclassified and rescaled data were intersected with
ecological subsections, as defined by the U.S. Forest Service
(USFS) (Keys et al., 1995; WDNR, 1999). If >50% of a USFS
subsection was classified as northern hardwood–hemlock then
the subsection was included in the study area. Ultimately the
study area consisted of 22 ecological subsections located within
the USFS Vegetation Province 212 and totaling 63,610 km2.
To determine the current extent of northern hardwood–
hemlock forests within the same study area we used the USFS
Forest Inventory and Analysis Database (FIADB) because
reliable spatial data describing detailed forest cover classes at
fine spatial scales in Wisconsin do not exist. Using the 1996
survey data in Wisconsin and those from 1993 in Michigan, we
extracted the FIADB plots that occurred within the USFS
subsections comprising the study area. In each of the extracted
FIADB plots the relative basal area of all species was calculated.
The total amount of northern hardwood–hemlock forest was
determined by summing the areas represented by each FIADB
plot classified as that land cover type within the study area.
To relate changes in forest cover to the forest types from
which the carabid data was collected, we made finer distinctions
within the northern hardwood–hemlock forest type, pertaining to
both the level of human influence in these forests and their age
structure. All historic forests were considered unmanaged and
further delineated as either mature, old-growth or younger,
regenerating forests. Data from the PLS surveys do not distin-
guish between old-growth and regenerating forests. However,
modeling exercises and disturbance chronologies developed for
Sylvania and two other preserves containing unmanaged forests
in Upper Michigan estimated that approximately 64% of the
historic northern hardwood–hemlock forests were old-growth,
where the canopy was dominated by mature and/or large trees
(Frelich and Lorimer, 1991). We used this percentage to estimate
the amount of northern hardwood–hemlock forest that was old-
growth versus those forests regenerating from a natural
disturbance, as calculated by difference.
For the purposes of this paper unmanaged forests are those
that have been continuously dominated by natural disturbance
regimes, whereas managed stands refer to forests first logged in
the early 1900s. The unmanaged portion of the present land-
scape (the portion that had not been logged) was determined
from published reports for the region (Davis, 1996). About 73%
of these forests were considered late-successional old-growth
forests with the remainder designated as regenerating forests,
those recovering from natural disturbances (Frelich and
Lorimer, 1991).
In contrast to the historical landscape, the current landscape
is also composed of managed forests, comprising all current
northern hardwood–hemlock forests that are not considered
unmanaged. To determine the area occupied by forests most
similar to the even- and uneven-aged forests used in the carabid
study a distinction was made between managed forests �60
years old and those �60 years. This age cut-off was used
because the FIADB does not distinguish among different types
of forest management, however, our field data indicated that the
E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347 339
predominant age class of the even- and uneven-aged stands was
at least 60 years. Using plot-level information on the average
age of the trees in the dominant size class from the FIADB, we
calculated the area of managed forests in each of the two age
classes.
2.6. Statistical analyses
Species diversity was assessed by the Shannon (H0) and
Simpson’s (D) diversity indices per plot as calculated by a
jacknife estimator in the free software EstimateS (Colwell,
1997). Differences among forest types were assessed using
general linear model procedures (SAS, 1999). Compositional
differences in carabid communities between old-growth,
uneven-aged forests and even-aged forests were determined
by non-metric multidimensional scaling ordination, which is
well suited for non-normal data typical of species abundance
datasets (McCune and Mefford, 1999). A multi-response
permutation procedure (MRPP) was used to test for differences
in species assemblages between management histories. Species
exclusive to only one stand were removed from the analysis.
One old-growth site was removed as an outlier due to structural
evidence of a former lodge and heavy recreational activities
nearby. Sørensen’s coefficient was used to calculate the
community distance matrix for the remaining 22 sites using
PC-ORD software (McCune and Mefford, 1999). All data were
relativized by site prior to ordination analysis. Variation in
beetle assemblages was related to environmental variables and
the ordination axes using general linear models (SAS, 1999).
To find carabid indicator species and species assemblages
characteristic of each management type an Indicator Species
Analysis was performed using PC-ORD (Dufrene and
Legendre, 1997; McCune and Mefford, 1999). This method
produces indicator values for each species using information
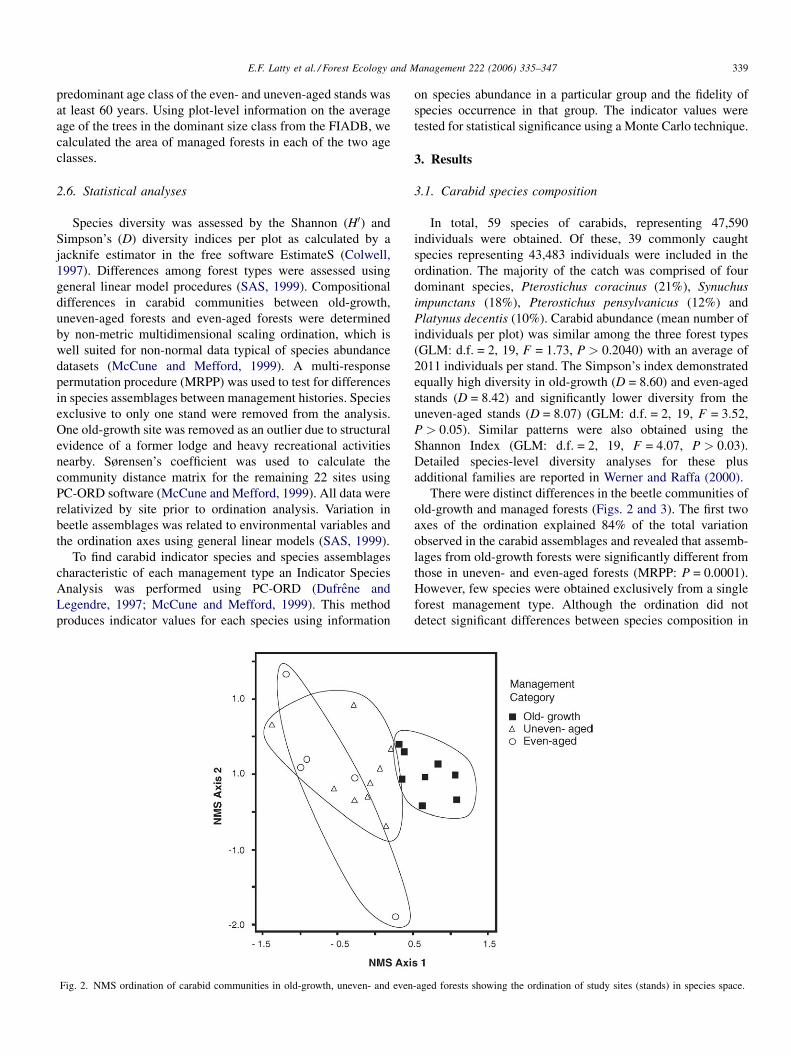
Fig. 2. NMS ordination of carabid communities in old-growth, uneven- and even
on species abundance in a particular group and the fidelity of
species occurrence in that group. The indicator values were
tested for statistical significance using a Monte Carlo technique.
3. Results
3.1. Carabid species composition
In total, 59 species of carabids, representing 47,590
individuals were obtained. Of these, 39 commonly caught
species representing 43,483 individuals were included in the
ordination. The majority of the catch was comprised of four
dominant species, Pterostichus coracinus (21%), Synuchus
impunctans (18%), Pterostichus pensylvanicus (12%) and
Platynus decentis (10%). Carabid abundance (mean number of
individuals per plot) was similar among the three forest types
(GLM: d.f. = 2, 19, F = 1.73, P > 0.2040) with an average of
2011 individuals per stand. The Simpson’s index demonstrated
equally high diversity in old-growth (D = 8.60) and even-aged
stands (D = 8.42) and significantly lower diversity from the
uneven-aged stands (D = 8.07) (GLM: d.f. = 2, 19, F = 3.52,
P > 0.05). Similar patterns were also obtained using the
Shannon Index (GLM: d.f. = 2, 19, F = 4.07, P > 0.03).
Detailed species-level diversity analyses for these plus
additional families are reported in Werner and Raffa (2000).
There were distinct differences in the beetle communities of
old-growth and managed forests (Figs. 2 and 3). The first two
axes of the ordination explained 84% of the total variation
observed in the carabid assemblages and revealed that assemb-
lages from old-growth forests were significantly different from
those in uneven- and even-aged forests (MRPP: P = 0.0001).
However, few species were obtained exclusively from a single
forest management type. Although the ordination did not
detect significant differences between species composition in
-aged forests showing the ordination of study sites (stands) in species space.
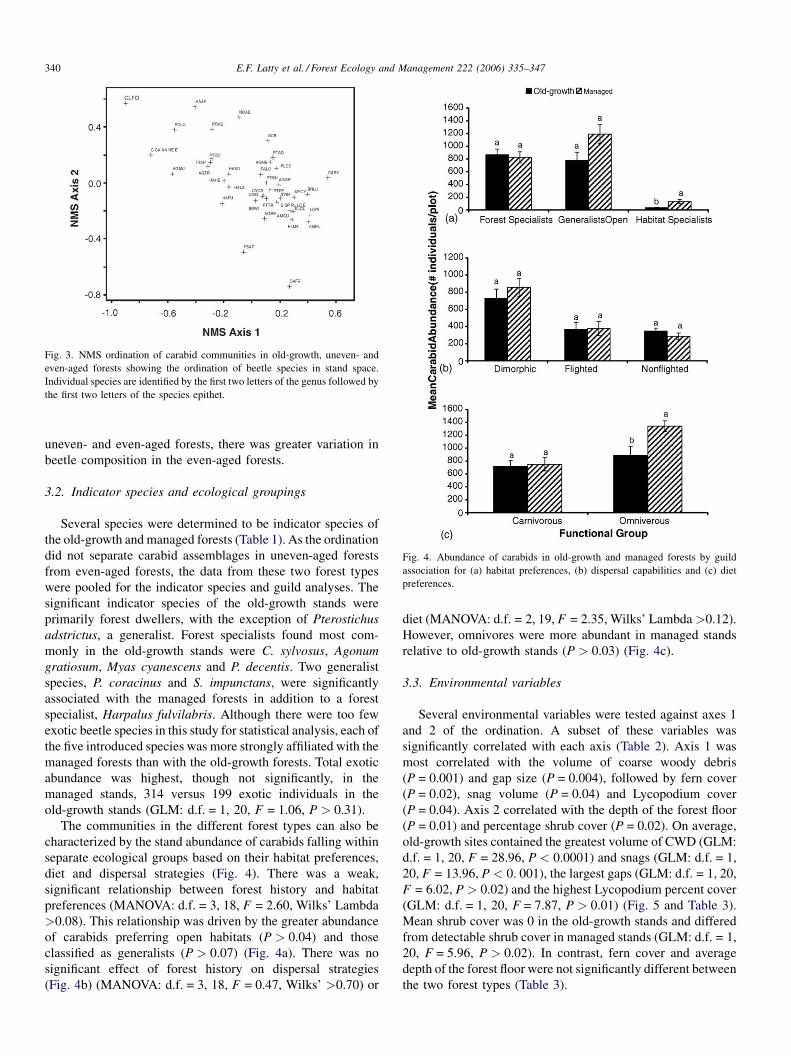
E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347340
Fig. 3. NMS ordination of carabid communities in old-growth, uneven- and
even-aged forests showing the ordination of beetle species in stand space.
Individual species are identified by the first two letters of the genus followed by
the first two letters of the species epithet.
Fig. 4. Abundance of carabids in old-growth and managed forests by guild
association for (a) habitat preferences, (b) dispersal capabilities and (c) diet
preferences.
uneven- and even-aged forests, there was greater variation in
beetle composition in the even-aged forests.
3.2. Indicator species and ecological groupings
Several species were determined to be indicator species of
the old-growth and managed forests (Table 1). As the ordination
did not separate carabid assemblages in uneven-aged forests
from even-aged forests, the data from these two forest types
were pooled for the indicator species and guild analyses. The
significant indicator species of the old-growth stands were
primarily forest dwellers, with the exception of Pterostichus
adstrictus, a generalist. Forest specialists found most com-
monly in the old-growth stands were C. sylvosus, Agonum
gratiosum, Myas cyanescens and P. decentis. Two generalist
species, P. coracinus and S. impunctans, were significantly
associated with the managed forests in addition to a forest
specialist, Harpalus fulvilabris. Although there were too few
exotic beetle species in this study for statistical analysis, each of
the five introduced species was more strongly affiliated with the
managed forests than with the old-growth forests. Total exotic
abundance was highest, though not significantly, in the
managed stands, 314 versus 199 exotic individuals in the
old-growth stands (GLM: d.f. = 1, 20, F = 1.06, P > 0.31).
The communities in the different forest types can also be
characterized by the stand abundance of carabids falling within
separate ecological groups based on their habitat preferences,
diet and dispersal strategies (Fig. 4). There was a weak,
significant relationship between forest history and habitat
preferences (MANOVA: d.f. = 3, 18, F = 2.60, Wilks’ Lambda
>0.08). This relationship was driven by the greater abundance
of carabids preferring open habitats (P > 0.04) and those
classified as generalists (P > 0.07) (Fig. 4a). There was no
significant effect of forest history on dispersal strategies
(Fig. 4b) (MANOVA: d.f. = 3, 18, F = 0.47, Wilks’ >0.70) or
diet (MANOVA: d.f. = 2, 19, F = 2.35, Wilks’ Lambda >0.12).
However, omnivores were more abundant in managed stands
relative to old-growth stands (P > 0.03) (Fig. 4c).
3.3. Environmental variables
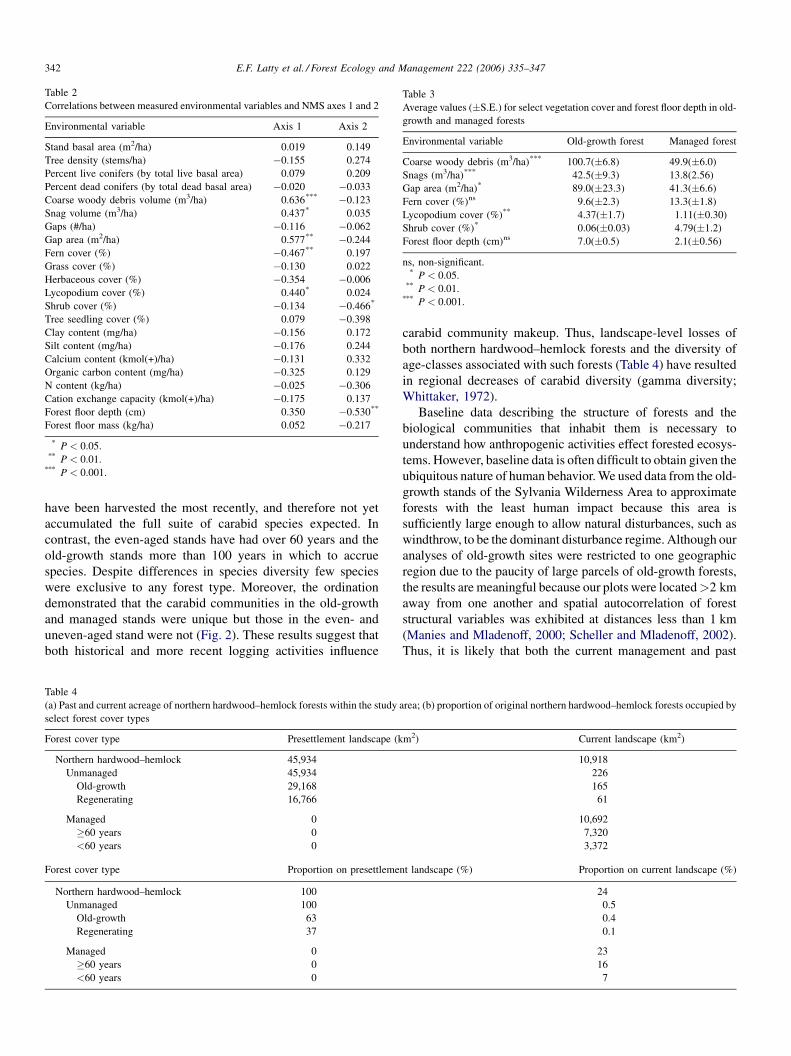
Several environmental variables were tested against axes 1
and 2 of the ordination. A subset of these variables was
significantly correlated with each axis (Table 2). Axis 1 was
most correlated with the volume of coarse woody debris
(P = 0.001) and gap size (P = 0.004), followed by fern cover
(P = 0.02), snag volume (P = 0.04) and Lycopodium cover
(P = 0.04). Axis 2 correlated with the depth of the forest floor
(P = 0.01) and percentage shrub cover (P = 0.02). On average,
old-growth sites contained the greatest volume of CWD (GLM:
d.f. = 1, 20, F = 28.96, P < 0.0001) and snags (GLM: d.f. = 1,
20, F = 13.96, P < 0. 001), the largest gaps (GLM: d.f. = 1, 20,
F = 6.02, P > 0.02) and the highest Lycopodium percent cover
(GLM: d.f. = 1, 20, F = 7.87, P > 0.01) (Fig. 5 and Table 3).
Mean shrub cover was 0 in the old-growth stands and differed
from detectable shrub cover in managed stands (GLM: d.f. = 1,
20, F = 5.96, P > 0.02). In contrast, fern cover and average
depth of the forest floor were not significantly different between
the two forest types (Table 3).

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347 341
Table 1
Ecological associations of Carabidae by affiliation with forest type as determined by indicator values
Species Indicator value Habitat preference Dispersal mechanism Diet Origin
Old-growth forest
Carabus sylvosus 86.7** Forest Brachypterous Carnivore Native
Agonum gratiosum 70.3* Open Brachypterous Carnivore Native
Myas cyanescens 68.4* Forest Macropterous NDa Native
Platynus decentis 67.5** Forest Brachypterous Omnivore Native
Pterostichus adstrictus 65.3* Generalist Macropterous Carnivore Native
Sphaeroderus lecontei 61.1 Forest ND Carnivore ND
Bradycellus lugubris 57.9 Generalist Macropterous Carnivore Native
Pterostichus pensylvanicus 55.7 Forest Dimorphic Carnivore Native
Calosoma frigidum 55.3 Forest Macropterous Carnivore Native
Cymindis cribicollis 51.6 Open Dimorphic Omnivore Native
Patrobus longicornis 49.9 Generalist Dimorphic Omnivore Native
Agonum melanarium 42.1 Generalist ND Carnivore Native
Loricera pilicornis 31.3 Forest Macropterous Omnivore Native
Agonum trigeminum 28.9 Open Macropterous ND Native
Platynus mannerheimii 20.5 Forest Brachypterous Carnivore Native
Elaphrus clairvillei 19.9 Forest Macropterous Carnivore Native
Amara cupreolata 19.7 Open Brachypterous Omnivore Native
Agonum placidum 15.8 Open Macropterous Omnivore Native
Harpalus laticeps 15.5 Generalist Macropterous ND Native
Managed forest
Harpalus fulvilabris 84.1*** Forest Dimorphic Carnivore Native
Carabus nemoralis 70.9 Open Brachypterous Omnivore Introduced
Pterostichus coracinus 66.9** Generalist ND Omnivore Native
Synuchus impunctatus 57.9 Generalist Dimorphic Omnivore Native
Pterostichus mutus 54.8 Generalist Macropterous Carnivore Native
Agonum retractum 54.4 Forest Dimorphic Carnivore Native
Pterostichus melanarius 54.0 Generalist Macropterous Omnivore Introduced
Pterostichus tristis 50.0 Forest ND Omnivore Native
Clivina fossor 50.1 Open Dimorphic Omnivore Introduced
Pseudamara aenaria 50.0* Forest Macropterous Carnivore Native
Calathus ingrates 49.0 Generalist Dimorphic Carnivore Native
Notiophilus aeneus 25.5 Forest ND Carnivore Native
Harpalus somnulentis 24.2 Open Macropterous Carnivore Introduced
Poecilus lucublandus 23.7 Open Brachypterous ND Native
Scaphinotus bilobus 21.4 Forest Brachypterous Carnivore Native
Harpalus herbivagous 18.5 Open Macropterous Omnivore Native
Trechus apicalis 14.7 Forest Dimorphic ND Native
Bembidion wingatei 14.4 Generalist Brachypterous ND Native
Agonum muelleri 14.3 Open Macropterous Carnivore Introduced
Harpalus affinis 14.3 Open Macropterous Omnivore Native
a ND, no data.* P < 0.05.
** P < 0.01.*** P < 0.001.
3.4. Regional land cover changes
The reconstruction of historic vegetation shows that
northern hardwood–hemlock forests were abundant in both
northern Wisconsin and western Upper Michigan (Fig. 5). This
forest cover type originally encompassed 72% of the defined
study area and now comprises 17%, more than a four-fold loss.
Although spatial data with similar resolution for current forest
cover types are not available, three areas containing major
parcels of old-growth northern hardwood–hemlock forests are
outlined (Fig. 5) to illustrate their spatial relationship to the
present landscape.
The historic landscape was dominated by unmanaged
hardwood–hemlock forests, whereas remaining northern hard-
wood–hemlock forests are mostly managed (Table 4). Approxi-
mately 24% of the historically defined northern hardwood–
hemlock forest is currently comprised of this forest type
(Table 4b). Within that 24%, most (98%) falls within the
managed forest type and over half of that is more than 60 years
old (Table 4a). The most dramatic losses have been to
unmanaged forests, with only about 0.5% remaining on the
current landscape (Table 4b).
4. Discussion
Species diversity and composition differed among forest
stand types. A possible explanation for the relatively low
species diversity of the uneven-aged stands is that these stands
E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347342
Table 2
Correlations between measured environmental variables and NMS axes 1 and 2
Environmental variable Axis 1 Axis 2
Stand basal area (m2/ha) 0.019 0.149
Tree density (stems/ha) �0.155 0.274
Percent live conifers (by total live basal area) 0.079 0.209
Percent dead conifers (by total dead basal area) �0.020 �0.033
Coarse woody debris volume (m3/ha) 0.636*** �0.123
Snag volume (m3/ha) 0.437* 0.035
Gaps (#/ha) �0.116 �0.062
Gap area (m2/ha) 0.577** �0.244
Fern cover (%) �0.467** 0.197
Grass cover (%) �0.130 0.022
Herbaceous cover (%) �0.354 �0.006
Lycopodium cover (%) 0.440* 0.024
Shrub cover (%) �0.134 �0.466*
Tree seedling cover (%) 0.079 �0.398
Clay content (mg/ha) �0.156 0.172
Silt content (mg/ha) �0.176 0.244
Calcium content (kmol(+)/ha) �0.131 0.332
Organic carbon content (mg/ha) �0.325 0.129
N content (kg/ha) �0.025 �0.306
Cation exchange capacity (kmol(+)/ha) �0.175 0.137
Forest floor depth (cm) 0.350 �0.530**
Forest floor mass (kg/ha) 0.052 �0.217
* P < 0.05.** P < 0.01.
*** P < 0.001.
Table 3
Average values (�S.E.) for select vegetation cover and forest floor depth in old-
growth and managed forests
Environmental variable Old-growth forest Managed forest
Coarse woody debris (m3/ha)*** 100.7(�6.8) 49.9(�6.0)
Snags (m3/ha)*** 42.5(�9.3) 13.8(2.56)
Gap area (m2/ha)* 89.0(�23.3) 41.3(�6.6)
Fern cover (%)ns 9.6(�2.3) 13.3(�1.8)
Lycopodium cover (%)** 4.37(�1.7) 1.11(�0.30)
Shrub cover (%)* 0.06(�0.03) 4.79(�1.2)
Forest floor depth (cm)ns 7.0(�0.5) 2.1(�0.56)
ns, non-significant.* P < 0.05.
** P < 0.01.*** P < 0.001.
have been harvested the most recently, and therefore not yet
accumulated the full suite of carabid species expected. In
contrast, the even-aged stands have had over 60 years and the
old-growth stands more than 100 years in which to accrue
species. Despite differences in species diversity few species
were exclusive to any forest type. Moreover, the ordination
demonstrated that the carabid communities in the old-growth
and managed stands were unique but those in the even- and
uneven-aged stand were not (Fig. 2). These results suggest that
both historical and more recent logging activities influence
Table 4
(a) Past and current acreage of northern hardwood–hemlock forests within the study
select forest cover types
Forest cover type Presettlement landscape (k
Northern hardwood–hemlock 45,934
Unmanaged 45,934
Old-growth 29,168
Regenerating 16,766
Managed 0
�60 years 0
<60 years 0
Forest cover type Proportion on presettleme
Northern hardwood–hemlock 100
Unmanaged 100
Old-growth 63
Regenerating 37
Managed 0
�60 years 0
<60 years 0
carabid community makeup. Thus, landscape-level losses of
both northern hardwood–hemlock forests and the diversity of
age-classes associated with such forests (Table 4) have resulted
in regional decreases of carabid diversity (gamma diversity;
Whittaker, 1972).
Baseline data describing the structure of forests and the
biological communities that inhabit them is necessary to
understand how anthropogenic activities effect forested ecosys-
tems. However, baseline data is often difficult to obtain given the
ubiquitous nature of human behavior. We used data from the old-
growth stands of the Sylvania Wilderness Area to approximate
forests with the least human impact because this area is
sufficiently large enough to allow natural disturbances, such as
windthrow, to be the dominant disturbance regime. Although our
analyses of old-growth sites were restricted to one geographic
region due to the paucity of large parcels of old-growth forests,
the results are meaningful because our plots were located>2 km
away from one another and spatial autocorrelation of forest
structural variables was exhibited at distances less than 1 km
(Manies and Mladenoff, 2000; Scheller and Mladenoff, 2002).
Thus, it is likely that both the current management and past
area; (b) proportion of original northern hardwood–hemlock forests occupied by
m2) Current landscape (km2)
10,918
226
165
61
10,692
7,320
3,372
nt landscape (%) Proportion on current landscape (%)
24
0.5
0.4
0.1
23
16
7

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347 343
Fig. 5. Spatial extent of the study area in northern Wisconsin and the Upper Peninsula, Michigan and the portion defined as northern hardwood–hemlock forests prior
to Euroamerican settlement. Public lands containing the largest old-growth parcels are shown in thick, black outline (1) Porcupine Wilderness State Park, (2) The
Huron Mountain Reserve Area and (3) The Sylvania Wilderness Area.
history of these stands have influenced the present carabid
communities. Similar differences in community assemblages
have been detected at equivalent spatial scales in Canada and
may reflect stand-level parameters like the amount of large
diameter woody material (Hammond et al., 2004) and understory
plant cover (Niemela and Spence, 1994).
Compositional differences in carabid assemblages in the
present study were most strongly correlated with variables
related to forest structure and groundlayer vegetation. Beetle
assemblages were related to coarse woody debris (Table 1),
which varied significantly among forest types (Table 3). Carabid
assemblages have been related to the presence of woody debris in
a variety of habitats, including Canadian pine and aspen forests
(Carcamo and Parkinson, 1999; Hammond et al., 2004), boreal
forests of Finland (Martikainen et al., 2000; Sippola et al., 2002),
Norway (Økland et al., 1996; Sverdrup-Thygeson, 2001) and
Australian hardwood forests (Oliver et al., 2000). Aside from
providing physical habitat, downed woody debris also influences
microhabitat parameters like soil moisture and local soil nutrient
content (Harmon et al., 1986) that in turn affect the abundance
and diversity of beetle species. Woody debris also augments
temporal and spatial habitat heterogeneity because dead wood
offers varied substrates to invertebrates, which change over time
as the wood decays. For example, the diversity of dead tree parts
and the presence of large diameter dead wood can increase the
abundance of many saproxylic (insects inhabiting dead wood)
beetle species (Økland et al., 1996). Therefore stands>100 years
old should be retained in order to conserve entire saproxylic
communities (Hammond et al., 2004). The characteristic carabid
assemblages of old-growth forests in this study were associated
with the highest amounts of woody debris (Table 3). Collectively,
these observations suggest that long-term reductions in the
quantity of coarse woody debris due to forest management may
have detrimental consequences for invertebrate communities,
particularly rare and saproxylic species (Sippola et al., 2002).
Understory vegetation structure, particularly Lycopodium,
fern and shrub cover, also influenced beetle assemblages
(Table 3). Several studies have linked vegetation structure with
carabid distribution (Niemela et al., 1992; Verschoor and
Krebs, 1995; Butterfield, 1997), although few have found
species-specific effects of plants on the beetle communities.
The structural characteristics of plant cover are likely to have
the greatest influence on both stand-level and microhabitat
features, and thus exert greater control over carabid distribution
than plant identity (Spence et al., 1996). The old-growth sites
were characterized by the highest percentage of Lycopodium
cover and the lowest shrub cover, creating a patchy understory
environment. The even-aged sites, in particular, had relatively
homogenous understory structure characterized by uniformly
dense fern cover and small amounts of dead wood.
With the exception of forest floor depth, we did not detect
significant correlations between soil parameters and beetle
distributions. Other studies have correlated soil characters like
organic matter content (Ings and Hartley, 1999), pH and
calcium availability (Carcamo and Parkinson, 1999) with the
composition of carabid communities when such data were
analyzed at fine spatial scales (1–100 m2). Although local
variation in soil characters is likely to influence beetle
assemblages at fine scales, we propose that at the stand-level,
structural characteristics of the forest are more likely to

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347344
influence beetle assemblages because these variables are
relatively consistent within stands yet exhibit moderate
amounts of stand-to-stand variation. We focused on stand-
level interactions of habitat variables and carabid assemblages
because this is the scale at which forest management generally
occurs.
Beetle communities were further characterized by groups of
indicator species. Several species had significant affiliations with
the old-growth forests including, C. sylvosus, A. gratiosum, M.
cyanescens and P. decentis. All Harpalus species and Synuchus
impunctatus were most strongly associated with the younger,
managed forests, similar to the findings of other studies (Niemela
et al., 1993; Werner and Raffa, 2000; Moore et al., 2004). The
large bodied Calosoma frigida was most strongly associated with
the late-successional, old-growth stands (IV = 55.3, Table 1).
This result is in keeping with a Canadian study that found the
greatest abundance of C. frigida in older stands than in those that
had been recently logged (Vance and Nol, 2003). In addition to
specific indicator species, carabids most strongly affiliated with
either old-growth or managed forests were marked by different
habitat preferences and diets. The managed forests supported
relatively high numbers of generalists and open habitat
specialists (Fig. 4a). More omnivores were also characteristic
of managed stands (Fig. 4b) and may reflect less specialized diet
preferences or requirements of species that have generalist
habitat affiliations.
In contrast, predominant modes of dispersal did not appear
to be related to the management history of the forest. According
to the habitat templet theory (Southwood, 1977, 1988),
dispersal capabilities should correlate with site disturbance,
such that species with poor dispersal abilities (e.g., brachypter-
ous species) should be more common in constant environments
than in disturbed environments. The opposite also holds and
high dispersal power is expected in populations living in highly
stochastic environments. Given this theory we expected
brachypterous species to dominate in the least disturbed
forests, the old-growth forests and macropterous species to be
more common in the post-logging forests. However, high
dispersal power also may be selected for in suitable but too
small environments (Den Boer, 1990). The remaining old-
growth forest in this region is highly fragmented and the parcels
that do exist are quite small relative to the entire landscape
(Fig. 5; Mladenoff et al., 1993). Moreover, there is some
evidence from other studies that carabids from heterogeneous
habitats have higher dispersal abilities than those from more
homogenous habitats (Den Boer, 1990; De Vries et al., 1996).
Thus conflicting selective forces, forces that favor both high
and low dispersal abilities, may be at work within forest types
thereby explaining the lack of differences in dispersal power
between old-growth and managed forests. Alternatively, the
selective forces influencing dispersal ability may be stronger
between land cover types, such as croplands versus forests, than
between different forest cover types.
Few introduced species were collected in this study,
however, those that were had stronger affiliations with managed
than old-growth forests (Table 1). Although introduced species
occurred with low frequency in this region, these data suggest
that old-growth forests may be one of the few places where
studies of relatively intact native beetle communities may still
be conducted. Our data do not indicate if this is due to habitat
characteristics or reflect that fact that old-growth stands are
generally less accessible than managed stands, and therefore
less likely to be invaded by non-indigenous species. Although
data from remaining old-growth forests are not likely to be
exact descriptions of carabid assemblages in presettlement
forests, they can inform conservation goals that seek to restore
species assemblages typical of the least human-disturbed
forests.
At a regional scale, our data suggest that the influence of past
and present forest management practices on beetle community
composition and ecological groups have had consequences for
their overall abundance and distribution. Under historic
conditions most northern hardwood–hemlock forests were in
a mature, old-growth state (Table 4). Approximately
45,934 km2 were potential habitat for carabid assemblages
characteristic of old-growth forests as opposed to 226 km2 that
exist currently. Post-logging forests have supplanted the
structurally diverse, old-growth forests of the historical
landscape and now constitute the dominant landscape matrix
of this region (Table 4, Mladenoff et al., 1993). Maturing
second-growth forests (>60 years old) are particularly common
and a significantly greater number of open habitat specialists
and more exotics reside in these managed forests, suggesting
that at the landscape level, there is a dramatically higher
proportion of species preferring open habitats than prior to
1850. This combined with the fact that those species most
closely affiliated with the old-growth forests were forest
specialists implies that conservation guidelines for this region
should emphasize the creation of old-growth characters in
remaining forests to insure forest structural diversity, and
therefore a greater diversity of insect fauna.
4.1. Implications for conservation and management
Data presented here suggest that forest management has
influenced the composition of beetle communities at the stand
and landscape scales and the abundance of species sharing
common ecological traits at the landscape scale. Despite
community level differences, of the 39 most commonly caught
carabid species, none were exclusive to a forest type. Thus, we
agree with other researchers that the use of single carabid
species as bioindicators of forest disturbance is of limited value
(Rykken et al., 1997), particularly at large geographic scales
that contain many forest types (Moore et al., 2004). The relative
abundances of groups of species proved most sensitive to past
forest management practices.
The beetle assemblages in each forest type shared some
ecological traits such as a greater number of forest specialists in
the old-growth stands. By necessity we limited our assessment
of the ecological associations of carabids to those for which we
could assemble reliable information. It is likely that other
important ecological groups may show affiliations with forests
of varying histories and future research should be directed
toward developing both a more comprehensive understanding

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347 345
of invertebrate ecology and how it is affected by human-
induced habitat changes. In particular, habitat heterogeneity
appears critical in structuring the composition of beetle
communities. The environmental variables most correlated
with beetle assemblages added habitat complexity in time and
space and have been shown to be systematically affected by
forest management practices (Goodburn and Lorimer, 1998;
Scheller and Mladenoff, 2002). Overall increases in land cover
types in the study region also yields a certain amount of
landscape heterogeneity that likely helps to maintain beetle
species diversity within the larger region (Niemela et al., 1996;
Romero-Alcaraz and Avila, 2000). However, given forest
reduction at the landscape-scale and the nearly 100% loss of
old-growth forest we suggest that the conservation of carabid
diversity is dependent on maintaining forests in a variety of age-
classes including late-successional stages.
The important role of old-growth forests in maintaining
specific carabid assemblages requires more investigation into
how old-growth character development may be accelerated
through management prescriptions. Such stands are not easily
created and require time to mature. Longer rotation times may
hasten this process but management plans should also
incorporate other techniques, such as retaining key structural
features during harvesting (Franklin et al., 1997) that maintain
habitat heterogeneity. This is also important because composi-
tional and structural characteristics may persist for several 100
years in managed forest landscapes (He and Mladenoff, 1999).
The links between beetle communities, environmental characters
and forest management suggest that old-growth forest provide a
useful surrogate for historical data describing carabid commu-
nities and may be used to inform current conservation practices.
Acknowledgements
We thank John Goodburn and James Bockheim for their data
on forest structure, vegetation cover and soil characters in the
sampled stands. This project was funded by the Wisconsin
Department of Natural Resources. We also thank two
anonymous reviewers for providing critical comments on the
manuscript.
References
Bockheim, J.G., 1997. Soils in a hardwood–hemlock ecosystem mosaic in the
Southern Lake Superior Uplands. Can. J. Forest Res. 27, 1147–1153.
Butterfield, J., 1997. Carabid community succession during the forestry cycle in
conifer plantations. Ecography 20, 614–625.
Campbell, N.A., 1990. Biology, 2nd ed. The Benjamin/Cummings Publishing
Company, Inc., Redwood City, CA, pp. 626–628.
Campbell, J.L., Gower, S.T., 2000. Detritus production and soil N transforma-
tions in old-growth eastern hemlock and sugar maple stands. Ecosystems 3,
185–192.
Carcamo, H.A., Parkinson, D., 1999. Distribution of ground beetle assemblages
(Coleoptera Carabidae) around sour gar processing plants in western
Canada. Pedobiologia 43, 160–173.
Cole, L.J., McCracken, D.I., Dennis, P., Downie, I.S., Griffin, A.L., Foster,
G.N., Murphy, K.J., Waterhouse, T., 2002. Relationships between agricul-
tural management and ecological groups of ground beetles (Coleoptera:
Carabidae) on Scottish farmland. Agric. Ecosyst. Environ. 93, 323–336.
Colwell, R.K., 1997. EstimateS. Version 5. User’s Guide and Application.
[Online, URL: http://viceroy.eeb.uconn.edu/estimates].
Comer, P.J., Albert, D.A., Wells, H.A., Hart, B.L., Raab, J.B., Price, D.L.,
Kashian, D.M., Corner, R.A., Schuen, D.W., 1995. Michigan’s Native
Landscape, as Interpreted from the General Land Office Surveys 1816–
1856. Michigan Natural Features Inventory, Lansing, MI, 78 pp.
Crow, T.R., Buckley, D.S., Nauertz, E.A., Zasada, J.C., 2002. Effects of
management on the composition and structure of northern hardwood forests
in Michigan. Forest Sci. 48, 129–145.
Dale, V.H., Brown, S., Haeuber, R.A., Hobbs, N.T., Huntly, N., Naiman, R.J.,
Riebsame, W.E., Turner, M.G., Valone, T.J., 2000. Ecological principles and
guidelines for managing the use of land. Ecol. Appl. 10, 639–670.
Davis, M.B., 1996. Extent and location. In: Davis, M.B. (Ed.), In Eastern Old-
growth Forests: Prospects for Rediscovery and Recovery. Island Press,
Washington, DC, USA, pp. 18–34.
Den Boer, P.J., 1990. The survival value of dispersal in terrestrial arthropods.
Biol. Conserv. 54, 175–192.
De Vries, H.H., Den Boer, P.J., van Dijk, T.S., 1996. Ground beetle species in
heathland fragments in relation to survival, dispersal, and habitat prefer-
ence. Oecologia 107, 332–342.
Dufrene, M., Legendre, P., 1997. Species assemblages and indicator species:
the need for a flexible asymmetrical approach. Ecol. Monogr. 67, 345–
366.
Durkis, T.J., Reeves, R.M., 1982. Barriers increase efficiency of pitfall traps.
Entomol. News 93, 9–13.
Duval, M.D., Grigal, D.F., 1999. The effects of timber harvesting on coarse
woody debris in red pine forests across the Great Lakes states, USA. Can. J.
Forest Res. 29, 1926–1934.
Epstein, M.E., Kuhlman, H.M., 1990. Habitat distribution and seasonal occur-
rence of carabid beetles in east-central Minnesota. Am. Midland Nat. 123,
209–225.
Forsythe, T.G., 1987. The relationship between body form and habit in some
Carabidae (Coleoptera). J. Zool. Soc. Lond. 211, 643–666.
Foster, D.R., 1992. Land-use history (1730–1990) and vegetation dynamics in
central New England, USA. J. Ecol. 80, 753–772.
Franklin, J.F., Rae Berg, D., Thornburgh, D.A., Tappeiner, J.C., 1997. Alter-
native silvicultural approaches to timber harvesting: variable retention
harvest systems. In: Kohm, K., Franklin, J.F. (Eds.), Creating a Forestry
for the 21st Century. Island Press, Washington, DC, pp. 111–139.
Frelich, L.E., Lorimer, G.C., 1991. A simulation of landscape-level stand
dynamics in the northern hardwood region. J. Ecol. 79, 223–233.
Gardner, S.M., 1991. Ground beetle (Coleoptera: Carabidae) communities on
upland heath and their association with heathland flora. J. Biogeogr. 18,
281–289.
Goodburn, J.M., Lorimer, C.L., 1998. Cavity tree and coarse woody debris in
old-growth and managed northern hardwood forests in Wisconsin and
Michigan. Can. J. Forest Res. 28, 427–438.
Goodburn, J.M., 1996. Comparison of forest habitat structure and composition
in old-growth and managed northern hardwoods in Wisconsin and Michi-
gan. M.S. Thesis. University of Wisconsin, Madison.
Greenslade, P.J.M., 1964. Pitfall trapping as a method for studying populations
of Carabidae (Coleoptera). J. Anim. Ecol. 33, 301–310.
Hale, C.M., Pastor, J., Rusterholz, K.A., 1999. Comparison of compositional
characteristics in old-growth and mature, managed hardwood forests of
Minnesota, USA. Can. J. Forest Res. 29, 1479–1489.
Hammond, H.E.J., Langor, D.W., Spence, J.R., 2004. Saproxylic beetles
(Coleoptera) using Populus in boreal aspen stands of western Canada:
spatiotemporal variation and conservation of assemblages. Can. J. Forest
Res. 34, 1–19.
Harmon, M.E., Franklin, J.F., Swanson, F.J., Sollins, P., Gregory, S.V., Lattin,
J.D., Anderson, N.H., Cline, S.P., Aumen, N.G., Sedell, J.R., Lienkaemper,
G.W., Cromack Jr., K., Cummins, K.W., 1986. Ecology of coarse woody
debris in temperate ecosystems. Adv. Ecol. Res. 15, 133–302.
He, H.S., Mladenoff, D.J., 1999. The effects of seed dispersal on the simulation
of long-term forest landscape change. Ecosystems 2, 308–319.
Hodge, S.J., Patterson, G., McIntosh, R., 1998. The approach of the British
Forestry Commission to the conservation of forest biodiversity. In: Bach-
mann, P., Kohl, M., Paivinen, R. (Eds.), Assessment of Biodiversity for

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347346
Improved Forest Planning. European Forest Institute Proceedings No. 18.
Kluwer Academic Publishers, Dordrecht, pp. 91–101.
Holland, J.M., Smith, S., 1999. Sampling epigeal arthropods: an evaluation of
fenced pitfall traps using mark-release-recapture and comparisons to
unfenced pitfall traps in arable crops. Entomol. Exp. App. 91, 347–357.
Houghton, R.A., 1995. Land-use change and the carbon cycle. Global Change
Boil. 1, 275–287.
Houghton, R.A., Hackler, J.L., 2000. Changes in terrestrial carbon storage in the
United States. I: the roles of agriculture and forestry. Global Ecol. Biogeogr.
9, 125–144.
Ings, T.C., Hartley, S.E., 1999. The effect of habitat structure on carabid
communities during the regeneration of a native Scottish forest. Forest
Ecol. Manag. 119, 123–136.
Janzen, D., 1985. On ecological fitting. Oikos 45, 308–310.
Jonsell, M., Weslien, J., Ehnstrom, B., 1998. Substrate requirements of red-
listed saproxylic invertebrates in Sweden. Biodivers. Conserv. 7, 749–764.
Jukes, M.R., Peace, A.J., Ferris, R., 2001. Carabid beetle communities asso-
ciated with coniferous plantations in Britain: the influence of site, ground
vegetation and stand structure. Forest Ecol. Manag. 148, 271–286.
Kavanaugh, D., 1992. Carabid beetles (Insecta: Coleoptera: Carabidae) of the
Queen Charlotte Islands, British Columbia.
Keys Jr., J., Carpenter, C., Hooks, S., Koenig, F., McNab, W.H., Russel, W.,
Smith, M.L., 1995. Ecological units of the eastern United States – first
approximation. Map. USDA Forest Service, Washington, DC.
Liebherr, J., Mahar, J., 1979. The carabid fauna of the upland oak forest in
Michigan: survey and analysis. Coleoptera Bull. 33, 183–197.
Lindroth, C.H., 1969. The ground-beetles of Canada and Alaska, Parts 1–6.
Opuscular Entomol. (Suppl. 1), 192.
Lindroth, C.H., 1974. Handbook for the Identification of British Insects, vol. IV,
Part 2: Coleoptera, Carabidae. Royal Entomological Society of London,
London.
LaRochelle, A., 1972. Notes on the food of Cychrini (Coleoptera: Carabidae).
Great Lakes Entomologist 5, 81–83.
Luff, M.L., 1975. Some features influencing the efficiency of pitfall traps.
Oecologia 19, 345–357.
Magura, T., 2002. Carabids and forest edge: spatial pattern and edge effect.
Forest Ecol. Manag. 157, 23–37.
Manies, K.L., Mladenoff, D.J., 2000. Testing methods to produce landscape-
scale presettlement vegetation maps from the US public land survey records.
Landscape Ecol. 15, 741–754.
Martikainen, P., Siitonen, J., Punttila, P., Kaila, L., Rauh FJ., 2000. Species
richness of Coleoptera in mature managed and old-growth boreal forests in
southern Finland. Biol. Conserv. 94, 199–209.
Mattson, W.J., Addy, N.D., 1975. Phytophagous insects as regulators of forest
primary production. Science 190, 515–522.
McCullough, D.G., Werner, R.A., Neumann, D., 1998. Fire and insects in
northern and boreal forest ecosystems of North America. Annu. Rev.
Entomol. 43, 107–127.
McCune, B., Mefford, M.J., 1999. PC-ORD. Multivariate Analysis of Ecological
Data, Version 4. MJM Software Design, Gleneden Beach, Oregon, USA.
Miller, T.F., Mladenoff, D.J., Clayton, M.K., 2002. Spatial autocorrelation and
patterns of understory vegetation and environment in old-growth northern
hardwood forests. Ecol. Monogr. 72, 487–503.
Mladenoff, D.J., Pastor, J., 1993. Sustainable forest ecosystems in the northern
hardwood and conifer forest region: concepts and management. In: Aplet,
G.A., Olson, J.T., Johnson, N., Sample, V.A. (Eds.), In Defining Sustainable
Forestry. Island Press, Washington, DC, pp. 145–190.
Mladenoff, D.J., White, M.A., Pastor, J., Crown, T.R., 1993. Comparing spatial
pattern in unaltered old-growth and disturbed forest landscapes. Ecol. Appl.
3, 294–306.
Mladenoff, D.J., Haight, R.G., Sickley, T.A., Wydeven, A.P., 1997. Causes and
implications of species restoration in altered ecosystems. Bioscience 47,
21–31.
Moore, J., Oiumet, R., Houle, D., Camire, C., 2004. Effects of two silvicultural
practices on ground beetles (Coleoptera: Carabidae) in a northern hardwood
forest, Quebec Canada. Can. J. Forest Res. 34, 959–968.
Niemela, J., 1997. Invertebrates and boreal forest management. Conserv. Biol.
11, 601–610.
Niemela, J., Spence, J.R., Spence, D.H., 1992. Habitat associations and seasonal
activity of ground beetles (Coleoptera: Carabidae) in Central Alberta. Can.
Entomol. 124, 521–540.
Niemela, J., Langor, D., Spence, J.R., 1993. Effects of clear-cut harvesting on
boreal ground-beetle assemblages (Coleoptera: Carabidae) in Western
Canada. Conserv. Biol. 7, 551–561.
Niemela, J., Spence, J.R., 1994. Distribution of forest dwelling carabids
(Coleoptera) – spatial scale and the concept of communities. Ecography
17, 166–175.
Niemela, J., Haila, Y., Punttila, P., 1996. The importance of small-scale
heterogeneity in boreal forests: variation in diversity in forest-floor inverte-
brates across the successional gradient. Ecography 19, 352–368.
Økland, B., Bakke, A., Hagvar, S., Kvamme, T., 1996. What factors influence
the diversity of saproxylic beetles? A multiscaled study from a spruce forest
in southern Norway. Biodivers. Conserv. 5, 75–100.
Oliver, I., Mac Nally, R., York, A., 2000. Identifying performance indicators of
the effects of forest management on ground-active arthropod biodiversity
using hierarchical partitioning and partial canonical correspondence ana-
lysis. Forest Ecol. Manag. 139, 21–40.
Pastor, J., Broschart, M., 1990. The spatial pattern of a northern conifer-
hardwood landscape. Landscape Ecol. 4, 55–68.
Petterson, R.B., Ball, J.P., Renhorn, K.E., Esseen, P.A., Sjoberg, K., 1995.
Invertebrate communities in boreal forest canopies as influenced by forestry
and lichens with implications for passerine birds. Biol. Conserv. 74,
57–63.
Refseth, D., 1980. Ecological analysis of carabid communities – potential use in
biological classification for nature conservation. Biol. Conserv. 17, 131–
141.
Ribera, I., Doledec, S., Downie, I.S., Foster, G.N., 2001. Effect of land
disturbance and stress on species traits of ground beetle assemblages.
Ecology 82, 1112–1129.
Rieske, L.K., Buss, L.J., 2001. Influence of site on diversity and abundance of
ground- and litter-dwelling Coleoptera in Appalachian oak-hickory forests.
Environ. Entomol. 30, 484–494.
Romero-Alcaraz, E., Avila, J.M., 2000. Landscape heterogeneity and relation to
variations in epigaeic beetle diversity of a Mediterranean ecosystem.
Implications for conservation. Biodivers. Conserv. 9, 985–1005.
Rosenberg, D.M., Danks, H.V., Lemkuhl, D.M., 1986. Importance of insects in
environmental impact assessment. Environ. Manag. 10, 773–783.
Rykken, J.J., Capen, D.E., Mahabir, S.P., 1997. Ground beetles as indicators of
land type diversity in the green mountains of Vermont. Conserv. Biol. 11,
522–530.
SAS, 1999. SAS/STAT User’s Guide, Version 7. SAS Institute, Cary, North
Carolina, USA.
Scheller, R.M., Mladenoff, D.J., 2002. Understory species patterns and diversity
in old-growth and managed northern hardwood forests. Ecol. Appl. 12,
1329–1343.
Schulte, L.A., Mladenoff, D.J., Nordheim, E.V., 2002. Quantitative classifica-
tion of a historic northern Wisconsin (U.S.A.) landscape: mapping forests at
regional scales. Can. J. Forest Res. 32, 1616–1638.
Sippola, A.L., Siitonen, J., Puntila, P., 2002. Beetle diversity in timberline
forests: a comparison between old-growth and regeneration areas in Finnish
Lapland. Ann. Zool. Fennici 39, 69–86.
Smith, B.E., Marks, P.L., Gardescu, S., 1993. Two hundred years of forest cover
changes in Tompkins County, New York. Bull. Torrey Bot. Club 120, 229–
247.
Southwood, T.R.E., 1977. Habitat, the templet for ecological strategies. J.
Anim. Ecol. 46, 337–365.
Southwood, T.R.E., 1988. Tactics, strategies and templets. Oikos 52, 3–18.
Spence, J.R., 1990. Success of European carabid species in western Canada:
preadaptation for synanthropy? In: Stork, N.E. (Ed.), The Role of Ground
Beetles in Ecological and Environmental Studies. Intercept, Andover, UK,
pp. 129–141.
Spence, J.R., Niemela, J.K., 1994. Sampling carabid assemblages with pitfall
traps: the madness and the method. Can. Entomol. 126, 881–894.
Spence, J.R., Langor, D.W., Niemela, J., Carcamo, H.A., Currie, C.R., 1996.
Northern forestry and carabids—the case for concern about old-growth
species. Ann. Zool. Fennoscandia 33, 173–184.

E.F. Latty et al. / Forest Ecology and Management 222 (2006) 335–347 347
Stearns, F., 1990. Forest history and management in the northern Midwest. In:
Sweeney, J.M. (Ed.), Management of Dynamic Ecosystems. North Central
Section, The Wildlife Society, West Lafayette, IN, pp. 107–122.
Sverdrup-Thygeson, A., 2001. Can ‘continuity indicator species’ predict spe-
cies richness or red-listed species of saproxylic beetles? Biodivers. Con-
serv. 10, 815–832.
Triplehorn, C.A., Johnson, N.F., 2005. Borror and Delong’s Introduction to the
Study of Insects, seventh ed. Thomson Brooks/Cole, Belmont, California,
pp. 401–402.
Turner, B.L., Clark, W.C., Kates, R.W., Richards, J.F., Mathews, J.T., Meyer,
W.B., 1990. The Earth as Transformed by Human Action. Cambridge
University, Cambridge, UK.
Vance, C.C., Nol, E., 2003. Temporal effects of selection logging on ground
beetle communities in northern hardwood forests of eastern Canada.
Ecoscience 10, 49–56.
Verschoor, B.C., Krebs, B.P.M., 1995. Diversity changes in a plant and carabid
community during early succession in an embanked saltmarsh area. Ped-
obiologia 39, 405–416.
Wisconsin Department of Natural Resources (WDNR), 1999. Ecological
Units of Wisconsin – First Approximation, Including LandType Associa-
tions (LTAs). Wisconsin Department of Natural Resources, Madison, WI.
Werner, S.M., Raffa, K.F., 2000. Effects of forest management practices
on the diversity of ground-occurring beetles in mixed northern hardwood
forests of the Great Lakes Region. Forest Ecol. Manag. 139, 135–
155.
Westman, W.E., 1990. Managing for biodiversity. Bioscience 35, 26–33.
Whittaker, R.H., 1972. Evolution and measurement of species diversity. Taxon
2/3, 151–213.
Whittaker, R.H., 1975. Communities and Ecosystems. MacMillan, New York.
Whittam, R.M., McCracken, J.D., Francis, C.M., Gartshore, M.E., 2002. The
effects of selective logging on nest-site selection and productivity of hooded
warblers. Can. J. Zool. 80, 644–654.
Ziegler, S.S., 2000. A comparison of structural characteristics between
old-growth and postfire second-growth hardwood–hemlock forests in
Adirondack Park, New York, USA. Global Ecol. Biogeogr. 9, 373–
389.





























