Embed Size (px)
Citation preview

Camp. Biochem. Phvsiol. Vol. Printed in Great B&in
19A, No. 2, pp. 275-282, 1984 0300-9629/84 $3.00 + 0.00 0 1984 Pergamon Press Ltd
RESPIRATION OF CRABS IN AIR AND WATER
PATRICIA M. O’MAHONEY* and ROBERT J. FULL Department of Biological Sciences, State University of New York at Buffalo, Buffalo,
NY 14260, USA
(Received 31 January 1984)
Abstract-l. Oxygen consumption (i702), ventilation volume (ir) and 0, extraction (Ext) were measured for three species of crabs: the terrestrial Gecarcinus later&, the amphibious Curdisoma guunhumi, and the aquatic Cailinectes sapidus. In air the vo* of the crabs was ranked Gecarcinus > Cardisoma > Callinectes. In water the ir,, was Callinectes > Cardisoma > Gecarcinus.
2. When terrestrial Gecarcinus was submerged in water, ir,, fell to l/7 of its aerial value because both 0 and Ext fell. When aquatic Callinectes was exposed to air, voo, fell to l/3 of its aquatic value because both ir and Ext fell. Only the amphibious Cardisoma maintained iro2 constant in both media; in water v was increased to offset a fall in Ext.
3. All species behaved as %‘o, conformers when they were exposed to short-term hypoxia; the effect was more pronounced in the aquatic environment.
4. Severe short-term hypercapnia caused no change in the aquatic environment; but it stimulated increased ir and decreased Ext in air.
5. Such effects are consistent with vertebrate responses: 0, seems to be the most important regulatory factor of respiration in water and CO, is the most important in air.
INTRODUCTION
Out of several million species living on earth, few can survive in both air and water. The crustaceans are members of this elite group, and they deserve particu- lar attention for most rely on the same respiratory structure, the gill, in both media (Wolvenkamp and Waterman, 1960). Moreover, the gill chamber is ventilated in basically the same way in both water and air; the tiny respiratory appendage, the scaph- ognathite, beats water or air through the gill cham- ber, drawing the fluid in along the edge of the carapace and expelling it out of openings near the mouth. Only in highly specialized air breathers does the lining of the gill chamber begin to take on the characteristics of a lung (Diaz and Rodriguez, 1977; Taylor and Greenway, 1979).
Respiration in water and air presents different problems. Oxygen uptake is considerably more ex- pensive in water because of the medium’s density, viscosity and low capacitance for 02, demanding high ventilation requirements (Dejours, 1981). Carbon dioxide removal is accomplished more readily in water because of the gas’s high solubility in the medium. This results in a lower Pco2 in water breath- ers and a different strategy of acid-base balance than in air (Cameron, 1979; Dejours, 1978; Rahn, 1966). Respiratory control differs as well. Water breathers
are sensitive to changes in ambient 0, while air breathers respond more readily to changes in COZ. These principles have emerged largely from in- vestigation of vertebrates (Rahn and Howell, 1976). Studies on crustacean respiration tend to support their general application (see Cameron, 198lb and Taylor, 1982).
*Present address: Department of Biological Sciences, Uni- versity of Southern Maine, Portland, Maine, USA
In this paper, we examine both the aquatic and
aerial respiration of three crab species which vary in their degree of terrestrial adaptation. One species, the blue crab, Callinectes sapidus, is highly specialized for aquatic life. Another species, Gecarcinus lateralis, is highly terrestrial, returning to water only briefly for reproduction. The third species, Cardisoma guan- humi, is an amphibious crab existing in moist land burrows along the seashore and fresh water canals of the tropics. To improve understanding of the control of respiration in both media, the present study pro- vides a direct comparison of the crab species to respiratory stress (hypoxia and hypercapnia). This presentation clearly highlights the respiratory limi- tations and adaptations of crustaceans in the water/air transition.
MATERIALS AND METHODS
Animals
Three species of decapod crustaceans were used as experi- mental animals: Callinectes sapidus, the marine blue crab (88-147 g); Curdisoma guanhumi, a semi-terrestrial crab (83-176 g) and Gecurcinus lateralis, a land crab (51-75 g). All animals were obtained from southern Florida.
The crabs were housed in aquaria at a temperature of 2426°C and were fed a combination of raw clams, fish, dog food, egg shell and lettuce. The aquaria of the marine crab contained sea water at pH = 8 and specific gravity = 1.025 to 1.030 units, while the two more terrestrial species had access to a large pan of 50% sea water.
Respiratory experiments
Experiments in this paper were designed to measure 0, consumption (oo,), ventilation volume (v) and oxygen extraction (Ext) of crabs as they were exposed to different respiratory media (air vs water) during hypoxia and hyper- capnea. In all tests we used a respiratory mask, impermeable
to gas and water. It was sealed over the mouth with hot glue and wax and separated the inhalant respiratory current at the base of the legs from the exhalant current near the
275

276 P~IKK’IA X4. O’MAHOWZY and ROHEK~ J. FI'LI
mouth (Herreid et ul.. lY7Y). The mask did not obstruct free movements of any appendages. Leaks in the respiratory mask were discovered by putting the animals mto a tank of water and looking for streams of hubbies. When a leak was detected. the mask was resealed and tested again.
In aquatic experiments the water a crab ventilated left the mask by a tube and was emptied into a bottle. For the experiments conducted in air, gas ventilated from the mask was captured in balloons. Both methods suffer from two potential disadvantages. the chance of sustained reversed ventilation and an increased vent&tory resistance (Jo- hansen L’I al.. 1970). Each problem would result in an underestinlate in ii. Reversed ventilation by the animals in the aquatic and aerial experiments couid be readily observed by noting the movement of exhaled air or water. A reversal of ventilation produced an abrupt interruption of flow. No sustained periods of reversed ventilation occurred except in the cases mentioned. Infrequent and brief periods of re- versed ventilation would not cause significant errors in measurement of V. Resistance to ventilation could not be completely eliminated, but every effort was made to reduce flow resistance. This mcluded the use of short, large di- ameter tubing along with receptacles producing only small pressure gradients against ventilatory flow.
Ayurrtic, rr.spiru/ion. The experimental chamber used to determine aquatic respiratory values was a 151, or in the case of C;ccurcinu~, 6.55 I plastic box. The water level was kept constant by pumping water (Manostat Varistaltic pump) from a reservoir into the chambers at the rate of 2SOml:min and by rctumlng the excess water to the reser- voir by means of an overflow tube.
The animal exhaled via the respiratory mask through an exhalant tube (7 mm diameter: 10-l5cm in length) con- nected to a self-siphoning bottle which emptied into the reservoir. The ventilation volume was determined by know- ing the volume of the siphon bottle and the time between each emptying. We monitored the frequency of emptying by use of a thermistor probe placed into the bottle: it cooled each time the water discharged and this event was recorded on a polygraph.
Both the reservoir and the experimental chamber were aerated with air or a gas mixture supplied through a low pressure regulator (0.5-S psi). The oxygen content of the water was lowered during the hypoxia studies by bubbling the wtater with a gas mixture con~dining NZ. During the hypercapnia studies, the CO, content of the water was increased by bubbhng with a gas mixture containing CO?. The gases were mixed in a National Appliance Controlled Environmental Flow Meter Mixer. The mixed gas from the exit tube of the mixer was divided mto two streams by another small mixer and fed into two flow meters (Lab Crest Century) which were attached to air stones in the experi- mental chamber and reservoir.
The 0, content and Pc,,, were measured on inhalant samples taken from the reservoir and exhalant water sam- ples drawn from the exhalant tube leaving the respiratory mask. The 0, content was determined by the Microwinkler technique (Fox and Wingfield. 1938: Burke, 1962). In addition, in several of the experiments a calibrated YSI oxygen electrode was used and yielded comparable results. The P,,, of water samples was determined by the Astrup method (Astrup. 1956). Ph-Pco, curves were constructed by equilibrating water samples with several gases of known CO, concentration (+ 0.0 I.5 vol. cont. ‘I;,) and measuring pH at each Pcol. The P,,? of the water reservoir was determined by measuring pH and using the calibration curve. The O2 content of the water remained constant except at the highest
PcoI used (P,o2 = 100 torr) when Oz tension fell from I50 torr to I35 torr.
Procedure. Two sets of experiments were carried out to examine the response of the crab‘s respiratory system in water. At the beginning of each experiment. a crab wearing a respiratory mask, its cheiae closed with rubber bands, was
placed in the experimental chamber- .ind att,r~hcd ro II~L exhalant tube. A piece of Styrofoam hcpt the animal \irl\ merged. As the crabs were \uhmerged. air hubhlc\ LIUI’C ~~et’n leaving the gill chambers. This observatii~n \ugge\ts rhat aii retension in gill chambers was not a major prr)blenl. Laigc amounts of air were shown to be retuincd in the hranchi:il chambers of C‘urtlisornu (~ortil/e \ upon immer<;lon. IO th.tt the crab may not have been fully water-breathing for \cvcr;ii hours (Cameron. 198lb). To ensure that oui- animal\ wcrc breathing water, crabs wcrc wbjectcd to hypow clr hyper- capnia only after a 3 hr rest period. Any oxygen trapped in air bubbles would be deplcted well before the end of thi\ period.
Experiments were 9 hr in duration. An inhalant atid exhalant water sample was taken once an hour IO dctcrnlillc the oxygen content of the water. The first 3 hr of the experiment were used to determine normal or baseline dam. During the second 3 hr the gas concentration of the water was varied in a step-wise fashion by either decreasing the P,,: or increasing the P,,,, of the incoming gas: ( I ) At the end of the third hour of the experiment N, or CC), LV:I\ turned on. It required less than 15 min for the water to become equilibrated with the gas mixture. (2) At the end of the fourth hour, the Nz or CO1 was again increased. (3) The final increase in Nz and CO, took place at the end of the tifth hour of the experiment. At the end of the sixth hour of the experiment either Nz or CO, was turned oR‘ and the gas content of the water returned to normal. Data from the last 3 hr of the experiment were used to determine if the animal had recovered from the exper~mcntai handling. Ventilation volumes were recorded c~~ntlnuousl~ throughout the Y hi experiment.
Throughout all experiments. the crabs rested quietly in their chambers except during the aquatic hypoxia tests when there were 223min of sporadic struggling each time the O2 tension was lowered.
The sample sizes for the aquatic hypoxia tests were as follows: Cu1linwre.v = 8. Ccrrdkonm = IO and Gecarcinus = 6 while those for the aquatic hypercapnia experiments were: Cullinwlcs = 4, C‘trrdixmrr = I4 and Gecarcinus = 8.
Aerial txspirurion. Aerial respiration experiments were similar to the aquatic tests except for the following modifications. A large (11.25 I) plexiglass resptratory cham- ber was used for aerial experiments on C~l/lif~~~~~~.s and large specimens of Curdisomu. A smaller (3.251) chamber 01 similar design was used for Groxcims and small specimens of Cuudisnma. Gas flowed through the large chamber at the rate of 625 ml/mm. Gases introduced to the respirometer to be inhaled by a crab were lirst humidified by bubbling the gas through an Erylemeyer flask containing water.
Crabs exhaled through the respiratory mask. A short exhalant tube passed through the respiratory chamber wall and a two way stopcock. to two large balloons that functioned as miniature “Douglas Bags”. The maximum volume a balloon could hold was approximately IO I. The average volume of the balloon when emptied was lOOmI and therefore provided little resistance to ventilation. The tubing had an inner diameter of 7mm to minimiLe venli- latory resistance. The balloons were ~riodic~llly emptied. the time for filling noted and the volume of exhaled air was determined by water displacement. Exhalant gas %unples were withdrawn by syringe via a piece of polyethylene tubing in the respiratory mask. Samples were 2 4mi in volume. inhalant gas samples were obtained from near the animals’ inhalant openings. All gas samples wcrc analyrcd for 0, and CO, content on a 0.5 ml Scholander Gas Analyzer. The Scholander Micromethod is accurate to +O.Ol 5 vol. “,, (Scholander. 1947: <;audchout and Blayo, 1975). From timed measurements of gas composilion and volume, we could determine ii,,. aerial gill ventilation (Q,,) and O? Ext. Values for V,,~ were corrected to conditions ot
STPD.
Respiration of crabs in air and water 211
z 90 AIR
G 0” 360
p
% 30 2
5
o”O CALLINECTES CARDISOMA GECARCINUS
s Y * 3 60-
P ?I
H,O
z? $1 2 30- -J
AIR I- AIR
z o-
CALLINECTES CARDISOMA GECARCINUS
9-
AIR f 0” ;I AIR 36 7.7
g
13)
:: 3- AIR E c
H20 H20
O- CALLINECTES CARDISWA GECARCINUS
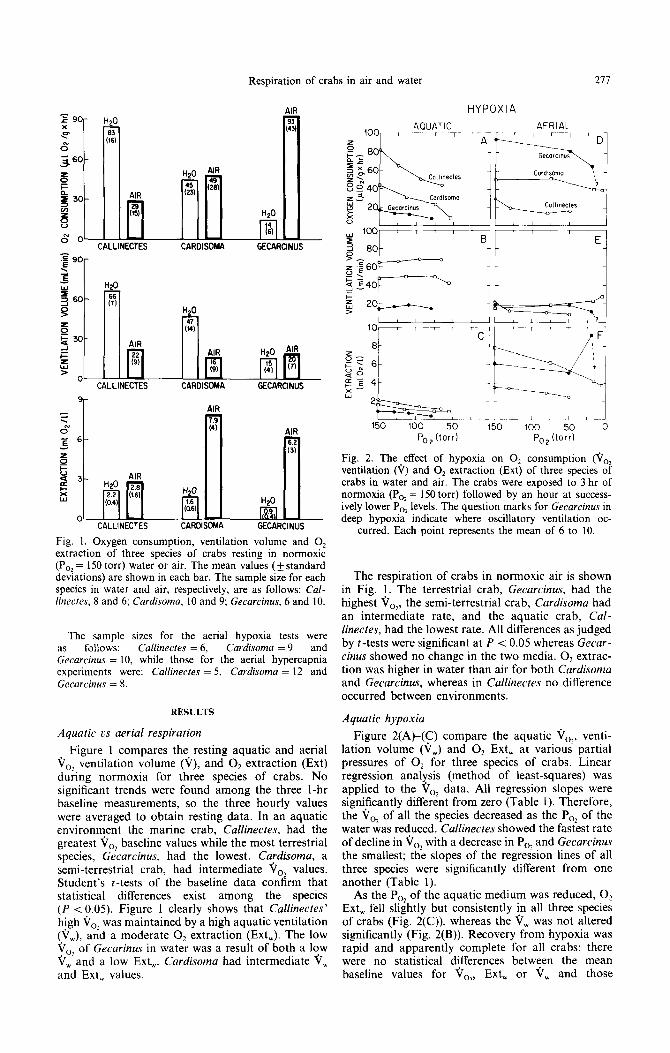
Fig. 1. Oxygen consumption, ventilation volume and 0, extraction of three species of crabs resting in normoxic (PO, = 150 torr) water or air. The mean values (+ standard deviations) are shown in each bar. The sample size for each species in water and air, respectively, are as follows: Cul- linecfes, 8 and 6; Cardisoma, 10 and 9; Gecarcinus, 6 and 10.
The sample sizes for the aerial hypoxia tests were as follows: Callinectes = 6, Cardisoma = 9 and Gecarcinus = 10, while those for the aerial hypercapnia experiments were: Callinectes = 5, Cardisoma = 12 and Gecarcinus = 8.
RESULTS
Aquatic us aerial respiration
Figure 1 compares the resting aquatic and aerial Vo, ventilation volume (V), and O2 extraction (Ext) during normoxia for three species of crabs. No significant trends were found among the three I-hr baseline measurements, so the three hourly values were averaged to obtain resting data. In an aquatic environment the marine crab, Callinectes, had the greatest V,, baseline values while the most terrestrial species, Gecarcinus, had the lowest. Cardisoma, a semi-terrestrial crab, had intermediate V,, values. Student’s t-tests of the baseline data confirm that statistical differences exist among the species (P < 0.05). Figure 1 clearly shows that Callinectes’ high Vo2 was maintained by a high aquatic ventilation (V,), and a moderate 0, extraction (Ext,). The low Vo, of Gecurinus in water was a result of both a low VW-and a low Ext,. Cardisoma had intermediate V, and Ext, values.
HYPOXIA
0
6
4
150 100 50 Pop (tow)
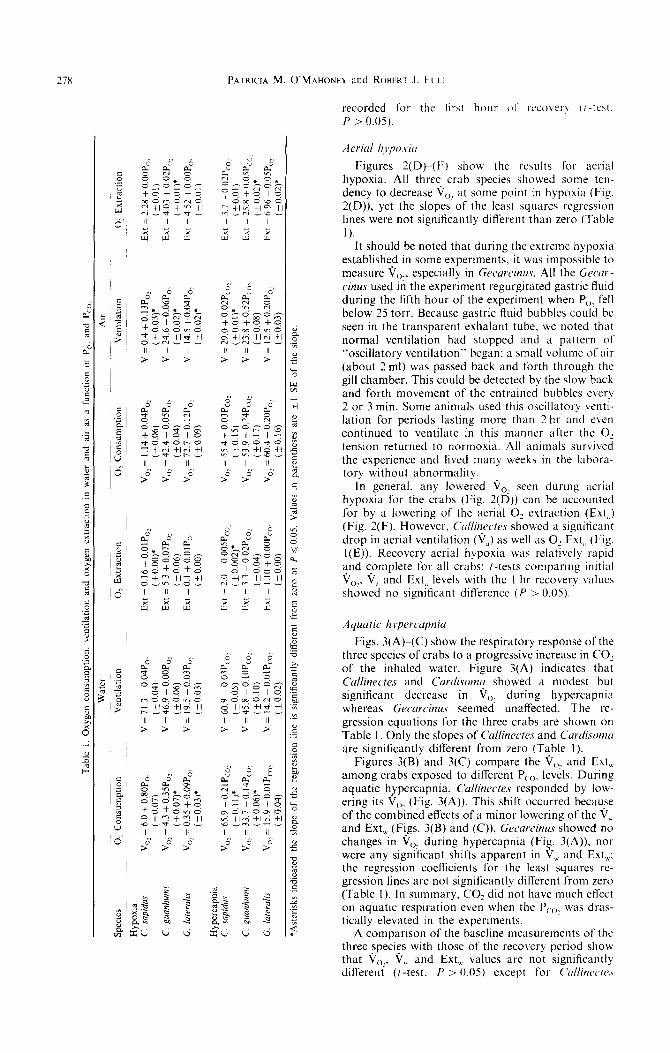
Fig. 2. The effect of hypoxia on O2 consumption (ir,, ventilation (9) and 0, extraction (Ext) of three species of crabs in water and air. The crabs were exposed to 3 hr of normoxia (PO, = 150 torr) followed by an hour at success- ively lower PO2 levels. The question marks for Gecarcinus in deep hypoxia indicate where oscillatory ventilation oc-
curred. Each point represents the mean of 6 to 10.
The respiration of crabs in normoxic air is shown in Fig. 1. The terrestrial crab, Gecarcinus, had the highest Vo2, the semi-terrestrial crab, Curdisoma had an intermediate rate, and the aquatic crab, Cal- linectes, had the lowest rate. All differences as judged by t-tests were significant at P < 0.05 whereas Gecar - cinus showed no change in the two media. 0, extrac- tion was higher in water than air for both Cardisoma and Gecarcinus, whereas in Callinectes no difference occurred between environments.
Aquatic hypoxia
Figure 2(A)-(C) compare the aquatic Vo,, venti- lation volume (9,) and O2 Ext, at various partial pressures of 0, for three species of crabs. Linear regression analysis (method of least-squares) was applied to the Vo2 data. All regression slopes were significantly different from zero (Table 1). Therefore, the V, of all the species decreased as the PO, of the water was reduced. Callinectes showed the fastest rate of decline in Vo2 with a decrease in PO1 and Gecarcinus the smallest; the slopes of the regression lines of all three species were significantly different from one another (Table 1).
As the PO* of the aquatic medium was reduced, O2 Ext, fell slightly but consistently in all three species of crabs (Fig. 2(C)), whereas the V, was not altered significantly (Fig. 2(B)). Recovery from hypoxia was rapid and apparently complete for all crabs: there were no statistical differences between the mean baseline values for V,,, Ext, or V’, and those
‘78
> > > > > >
recorded for the tirst hour 01‘ Ireco\rrb (f-[ct. P > 0.05).
Acrid li.ypo_uirr
Figures 2(D)-(F) show the results Tot- aerial hypoxia. All three. crab species showed some ten- dency to decrease V,,: at some point in hypoxia (Fig. 2(D)). yet the slopes of the least squares regression lines were not significantly different than zero (Table
1). It should be noted that during the extreme hypoxia
established in some experiments, it was impossible to measure ire,, especially in Gecurcitzus. All the Gecur - cinus used in the experiment regurgitated gastric fluid during the fifth hour of the experiment when P,,, fell below 25 torr. Because gastric fluid bubbles coulh be seen in the transparent exhalant tube, we noted that normal ventilation had stopped and a pattern of “oscillatory ventilation” began: a small volume of air (about 2ml) was passed back and forth through the gill chamber. This could be detected by the slow back and forth movement of the entrained bubbles every 2 or 3 min. Some animals used this oscillatory venti- lation for periods lasting more than 2 hr and even continued to ventilate in this manner after the 0: tension returned to normoxia. All animals survived the experience and lived many weeks in the labora- tory without abnormality.
In general. any lowered ii,? seen during aerial hypoxia for the crabs (Fig. 2(D)) can be accounted for by a lowering of the aerial 0, extraction (Ext,) (Fig. 2(F). However. C’ullincvtes showed a significant drop in aerial ventilation (V,) as well as O2 Ext,, (Fig. l(E)). Recovery aerial hypoxia was relatively rapid and complete for all crabs: /-tests comparing initial i/,,, ii,, and Ext,i levels with the I hr recovery values showed no significant difference (P > 0.05).
Aquutic h~~percupniu
Figs. 3(A)--(C) show the respiratory response of the three species of crabs to a progressive increase in CO, of the inhaled water. Figure 3(A) indicates that Cullinectes and Cutdisisomu showed a modest but significant decrease in v,), during hypercapnia whereas Grcurcinus seemed unaffected. The re- gression equations for the three crabs are shown on Table I. Only the slopes of Cullirwctes and Curdisomr are significantly different from zero (Table I).
Figures 3(B) and 3(C) compare the ii,, and Ext,, among crabs exposed to different P(o? lev&. During aquatic hypercapnia. Cullirwctr.~ responded by low- ering its Qir,,, (Fig. 3(A)). This shift occurred because of the comb:ned effects of a minor lowering of the VW and Ext, (Figs. 3(B) and (C)). Gecurcinus showed no changes m V,!, during hypercapnia (Fig. 3(A)), nor were any significant shifts apparent in V, and Ext,,; the regression coefficients for the least squares re- gression lines are not significantly different from zero (Table I). In summary, CO: did not have much efPect on aquatic respiration even when the P,.,,, was dras- tically elevated in the experiments.
A comparison of the baseline measurements of the three species with those of the recovery period show that vir,,,, ir, and Ext, values are not significantly diff‘ereni (t-test. P > 0.05) except for Ctrl/int~c~tc~.r

Respiration of crabs in air and water 279
HYPERCAPNIA AQUATIC AERIAL
I I I I I 11 1 1 1 ’
0 20 40 60 80 100 0 20 40 60 00 100 P cop (torr) P co2 (torr)
Fig. 3. The effect of hypercapnia on the 0, consumption (ire,), ventilation (v) and 0, extraction (Ext) of three species of crabs in water and air. The crabs were first exposed to 3 hr of normocapnia (Pco2 = 0.02 torr) followed by an hour of successively higher Pcol levels. Each point is the mean of
the 4 to 14 crabs.
which showed a significantly depressed vo2 due to a lowered VW even after 3 hr recovery.
Aerial hypercapnia
Figures 3(DHF) show the respiratory response of the crabs as they were exposed to increasing levels of CO* in air. Figure 2(D) shows that the initial 6,, of the three species in air were similar to one another. As the Pcoz of the environment was raised, the V,, of Callinectes showed no general trend. It is not clear whether the Callinectes’ response is due to the vagar- ies of sampling or if the crab does indeed have an elevated \jo2 at a Pco2 = 40 torr which then drops at higher ambient P,,, levels. On the other hand, Cardi- soma and Gecarcinus both displayed an apparent increase in vo, as the environmental Pco2 was in- creased. Yet the slopes of the regression equations were not different from zero (Table 1).
Examination of Figs 3(E) and 3(F). reveals the effect of increased aerial Pco, levels on V, and Ext,. In general, ventilation volumes increased with el- evated PcoZ, however this effect was only significant at the lower Pco, levels. After an ambient Pco, of 40 torr was reached, v’, stayed relatively constant. Oxygen extraction (Fig. 3(F)) showed a general and significant tendency to fall as P,,, levels increased: Gecarcinus and Cardisoma showed an abrupt drop in Ext, when Pco2 was increased above 20 torr. This was coincident with the increased 0, mentioned earlier. At Pcoz of 30 torr or higher, the Ext, in these two species remained constant. Callinectes also displayed a general decrease in Ext, with an elevated ambient
Pcol except the largest drop appeared between 40 to 80 torr.
All crabs recovered within 1 hr once they were returned to room air; i.e. no significant differences existed between their baseline and recovery values (P > 0.05). None of the crabs showed any overt signs of stress during the experiments in spite of the high levels of CO* tested.
DISCUSSION
Respiration in air and water
The blue crab, Callinectes sapidus, is primarily an aquatic species, but it can survive at least 2448 hr in air (Pearse, 1936; O’Mahoney, 1977). Our data show that such an exposure is accompanied by a decrease in vo, to about one-third of the aquatic value (Fig. 1). This decline occurs with a drop in ventilation volume. Batterton and Cameron (1978) estimated a 50% decrease in ventilation for the blue crab after 4 hr in air. It is not clear whether the fall in venti- lation was the result of a decrease in scaphognathite frequency or stroke volume; both mechanisms are possible (Wilkens, 1981). A decrease in ventilation might result in an energy savings in air. However, the possible energetic. reduction seems inadequate to ex- plain the fall in V,,, since the cost of ventilation is estimated at only l-20% of the standard ir, (Bat- terton and Cameron, 1978; Standaert, 1970). The best explanation for the fall in V, may be that ventilation efficiency is reduced in air; the blue crabs’s scaph- ognathite system may not be well designed for the air.
Callinectes did not compensate for the decrease in ventilation by increasing extraction (Fig. 1). Gill collapse probably was involved, since the gill lamellae of aquatic crabs are not rigid (Cameron, 1981a). Such collapse would sharply reduce the surface area for gas exchange. In the face of gill collapse, the extraction could only remain constant (Fig. 1) if, as we presume, circulatory adjustments were made.
The ineffectiveness of the Callinectes respiratory system in air did not cause death. This indicates Callinectes either decreased its total metabolic rate or the crabs switched to anaerobic metabolism. We lack the data to distinguish between the possibilities, but a few points are germane. Callinectes does have significant glycolytic capacity as it generates lactate readily during exercise (Booth et al., 1982). However, any prolonged anaerobic activity in this species may be limited by the acidotic effects. O’Mahoney (1977) showed a decreased hemolymph pH in Callinectes from 7.87 (water) to 7.71 (air) after 48 hr of air exposure. Hemolymph Pco? rose from 2.9 torr (water) to 8.05 torr (air) along with a corresponding increase in bicarbonate.
Cardisoma guanhumi, a semiterrestrial crab, shows a relatively constant 0, in air and water (Fig. 1). This result is consistent with Standaert’s (1970) un- published observations. The amphibious behavior is not unexpected because this crab is found on both land and in water. Moreover, water is located in the bottom of the land burrows (Gifford, 1963; Herreid and Gifford, 1963; Bliss and Mantel, 1968). A con- stant ir,, in both air and water has also been reported for other crustaceans such as the shore crab Carcinus maenus and the freshwater crayfish Austropotamobius

/~ul~ipf~.~ (Taylor and Butler, iY73: Taylor and Wheatly, lY80).
To achieve a constant Vo? in both water and air. C’. pmhmi manipulates both ventilation and 0: ex- traction In water, where 0: concentrations are low and O2 extraction falls dramatically, 0,. is increased correspondingly (Fig. 1). In air, where both 0, con- tent is high and O2 extraction rises. the V, drops. The gills of Clrrclisonru are well supported with stiffened margins of small gill leaflets (Cameron, 198 la). The branchiat lining which is reported to have extensive folds may act as a lung (Wood and Randall. I98 I ). These respiratory surfaces apparently function ade- quately in both water and air to provide a constant gas exchange.
Consistent with its amphibious nature, c’. ~uun- htlnfi maintains a constant hemolymph pH in both media. Standaert (1970) noticed that submergence of the crab caused a transitory rise in pH which may be due Co the elevation of V’, and a washout of CO?. but that pII returned to its original value within 4-6 hr. O’Mahoney (1977) measured a drop in blood Pco, but saw no difference in pH after 48 hr of sub- mergence. Forced water-breathing of another species, C~~{~i~s~~~~~ curn$r.u. showed similar results: a hypo- capnic alkalosis followed by a metabolic acidosis which restored pH (Cameron, 1.98 I h).
Gerarciws ~~t~ru~i.~. a terrestrial species, had an average aerial ii,? of 93 ~1 O$g,hr and a V, of 20 mI/min (Fig. 1). These values are comparable to rates reported by Cameron (1’375) but are inexplic- ably higher than those observed by Taylor and Davies (1981). When G. /rrrerrr/i.s was submerged in water, where O2 content is relatively low, the O? extraction plummeted to one-seventh of its aerial vatue (Fig. I). There was no ventilatory compen- sation so ii,,, dropped one-seventh of that in air.
G. ~a~~~~~~l~.s cannot survive prolonged submersion. O’Mahoney ( 1977) found only one individual of the four tested that could manage 18 hr exposure. The species appears specialized for land existence; its gilt surface area is reduced to 60”, of the area of Cur&- soma and only IS’,, of that of CuiJinecte,u (Bliss and Mantel, 1968). In addition. this species has rows of finger-like projections between gill leaflets for support in air (Cameron, 1981a). Even though G. laterulis can respire through its highly vascularized gill chamber lining (Diaz and Rodriquez, 1977). gas exchange by this route in water may be limited. The obvious conclusion seems that neither gills nor branchial lining allow the necessary 0, exchange for survival. Acid-base balance is profollndly affected in water. O’Mahoney (1977) submerged G. ~~~~~~~~~.~ for I2 hr and noticed hemolymph pII increased significantly from 7.38 to 7.54, while P,,,, shifted from 8.3 to 6.4 torr. In three animals that survived 18 hr ex- posure, the pH had dropped to 7.00 just before death. Such wild oscillations of acid -base balance may contribute to death.
Control of’ respiration
Hyperupniu. Theoretical arguments suggest that the control of respiration differs in air and water (see Qejours, 198 If. Vertebrate air-breathers are most sensitive to en~~jronmentai CO,. regardless of the
degree of terrestriality. while ~~~lter-br~~~tt~~r~ do titri respond to elevated CC+ by an increased ventilation Similar responses have reported for :I number of crustaceans in water (Batterton and Cameron. lY7S: Dejours and Beekenkamp, 1977) and air (Cameron, 1975, 1981b: Cameron and Mecklenburg, 1973). Yet. there are exceptions to this generalization am~g
other crustaceans (e.g. Arudpragasam and Naylor. 1964: Larimer. 1961; Massabuau t’f crl.. 1980). For exampie, OZ appears to control ~;ent~lati~3~~ rate in the terrestria! anomuran, ~.~~~,~~~)~i~ii ~~~p~i~~tz~~~ (~cMahon and Burggren. 1979) For crabs in water. all subject to the same experimental regime. wc noted that even drastically elevated CO, levels did not trigger compensatory increases in ventilation (Fig. 3(C)), whereas in air they did (Fig. 3(E)).
Fiypercapnia does not usually stimulate hyper- ventilation in water breathers, since blood P, ,), levels do not normally become significantly elevated. Be- cause of the large ventilation requirement of water breathers and the medium’s high CO? capacitance the difference between inspired and expired P,,,, is usu- ally small. Corr~spondingly~ there arc only small changes in arterial I?,,,, with veIltilati~~n. So venti- lation cannot serve as an effective method for controi- ting blood Pt.,,. or pH (Rahn. 1966: Dejours, IY78). Instead. in crabs acid-base regulation during aquatic hypercapnia is mainly carried out by gill ion exchange (Cameron. 197X; DeFur (‘f cri.. 1980: fruchot. 1979).
Hypercapnia in air-breathing vertebrates often stimulates hyperventilation, which in turn minimizes increases in arterial P,.,,, and pH (Dejours. 198 I), The situation seems similar in the three crab species depicted in Fig. 3(E) where aerial hypercapnia in- creased ventilation. Other authors report comparable results for G. l[~t~~r~~~i.~. f‘. ~~l~~i~~il~~~li. ~~(~~t~~l~j,su~~~l tr~n,~~‘~r~u and the coconut crab. 3irgrc.s inrro (Cam- eron, 1975. 198 I b; Cameron and ~~ecklenburg. I W?: Greenway (?I crf., 1983). However. Batterton and Cameron (1978) found no change rn the ventilation of C. sqG&.s in air; their results may differ from ours on the same species because we used higher levels of co*.
In summary, high P,.,,: in air stimulates crustacean ventilation just as it does for vertebrates. even though crabs may use the shell carbonate as a buffer to compensate for acid-base problems as well (DeFur rt al.. 1980; Cameron, 198lb; Henry c’t nl., 1981).
Hypoxia. A range of respiratory responses have been observed in crustaceans, as well as in other invertebrates during hypoxia (Herreid. 19801. At one extreme one Ends oxygen regulation and at the other extreme oxygen conformity. The same individual may operate in either mode depending upon iab stress, time of day, temperature. salinity, activity, rate of hypoxic exposure. etc. (Herreid. 1980: McMahon (JI uf., 1974; Taylor ct ui.. 1977; Taylor and Butler. 1973). If a strategy of regulation is adopted in the face of declining P,,, then either ventilation must increase or extraction must remain constant. When the latter occurs in hypoxia, this means a greater percentage of 0: must be removed (“,,Ext) as P,,: is decreased. If a strategy of OZ conformity is adopted. the animal must be abie to tolerate the lowered PC,-. by reducing its total energy expenditure or by shifting to anerohic metaboIjsm.

All the species of crabs we tested during aquatic from 135 to 50 torr. Certainly no significant venti- hypoxia acted as almost ideal 0, conformers (Fig. 2). latory compensation occurred at all. A similar com- When Po, was decreased to one-half its value, To2 ment might be made for the coconut crab Birgus; showed a similar decline. No compensation in V, occurred, and the fall in Vo, was accompanied by a
increased ventilation does not appear to be used until are reached (Cameron and Meck-
progressive and proportional decline in O2 extraction. very low Vol lenburg, 1973). In short, most crabs do not show
In contrast to these results, many authors report that undue dependency on PoZ in air, no doubt due to the crustaceans in water show a significant degree of 0, large absolute amounts of O2 present even at low P,,. regulation during hypoxia which is accomplished by Adjustments to a declining P,, do not seem to be an increase in V, (Batterton and Cameron, 1978; ventilatory except in the case of Cardisoma guanhumi. Butler et al., 1978; Childress, 1971; Dejours and Most crustaceans that regulate in air do so by Beekenkemp, 1977; McMahon et al., 1974; Wheatly maintaining constant O2 extraction, probably via and Taylor, 1981). However, sometimes an adjust- circulatory adjustments. ment on 0, extraction can be evident also (McMahon In conclusion, this comparative study suggests that and Wilkens, 1975; Taylor, 1976). These results re- the short-term response of decapod crustaceans to emphasize the tremendous diversity in crustacean respiratory stress (hypoxia and hypercapnia) depends response to hypoxia both within and between species on the medium, water vs air. In general oxygen (Taylor, 1982). Unfortunately, the causes underlying sensitivity dominates in the aquatic environment, this variability are unknown. while CO, or pH sensitivity in air is the most im-
Crustacean responses to hypoxia in air are less portant regulatory factor. This general principle of variable than in water (Fig. 2); generally regulation gas exchange developed from vertebrate studies has prevails, although mechanisms vary. For example, the aerial V,,
been supported for invertebrates. Further study of of Cardisoma guanhumi showed no crustacean respiration can lend information on the
change except at very low PO, (Fig. 2(D)). A similar evolutionary transition from water to land. response was reported for the same species *by Herreid et al. (1979) who observed no changes in Vo2 above 5 torr. Above this level, C. guanhumi seems to REFERENCES
compensate for a declining 0, Ext by an increase in Va.
Arudpragsam K. D. and Naylor E. (1964) Gill ventilation volumes, oxygen consumption and respiratory rhythms in
As another example, only a modest decline in aerial Carcinus meanus (L). J. exp. Biol. 41, 309-321. V?, was shown by Gercarcinus lateralis above 25 torr Astrup P. (1956) A simple electrometric technique for the
(Fig. 2(D)); but in contrast to Cardisoma, compen- determination of carbon dioxide tension in blood and
sation by ventilation did not occur (see a similar but plasm, and bicarbonate content in “separated” plasm at
irregular response reported by Cameron, 1975). a fixed carbon dioxide tension (40 mm Hg). &and. J. Clin.
Rather Gecarcinus adjusted to hypoxia by maintain- Invesl. 8, 33-43.
ing O2 extraction fairly constant or even increasing it. Batterton C. V. and Cameron J. N. (1978) Characteristics
Both these species respond to treadmill exercise in an of resting ventilation and response to hypoxia, hyper-
analogous way: Cardisoma guanhumi compensates capnia, and emersion in the blue crab Callinectes sapidus (Rathbun). J. exp. Zool. 203, 403418. _
primarily by increasing ventilation while Gecarcinus Bliss D. E. and Mantel L. H. (1968) Adantations of \ I
lateralis adjusts primarily by extraction presumably crustaceans to land: A summary and analysis of new via circulatory modifications (Herreid et al., 1979, findings. Am. Zool. 8, 673-685.
1983). Another difference in the species must be Booth C. E., McMahon B. R. and Finder A. W. (1982)
emphasized: Gecarcinus is less tolerant to hypoxia Oxygen uptake and the potentiating effects of increased
then Cardisoma. G. laterialis showed extreme stress at hemolymph lactate on oxygen transport during exercise in
Po, below 20 torr, regurgitating fluid and performing the blue crab. Callinectes sapidus. J. camp. Physiol. 143, 111-121.
an oscillatory ventilation. Only two C. guanhumi Burke J. D. (1962) Determination of oxygen in water using showed oscillatory ventilation patterns and those a IOml syringe. J. Mitchell Sot. No;.: 145-147. - appeared only at P,, = 5 torr. Cardisoma guanhumi Butler P. J.. Tavlor E. W. and McMahon B. R. (1978) appears better adapted to hypoxic conditions than G. Respiratory and circulatory changes in the lobster (Horn:
lateralis; both crabs are largely terrestrial but their arus vulgaris) during long term exposure to moderate
burrow habitats differ. C. guunhumi lives in mud hypoxia. J. exp. Biol. 73, 131-146.
burrows with water in the living chamber that smells Cameron J. N. (1975) Aerial gas exchange in the terrestrial
of H,S (Herreid and Clifford, 1963). Moreover, this Brachyura Gecarcinus lateralis and Cardisoma guanhumi.
species retires below ground and plugs its burrow for Comp. Biochem. Physiol. 52A, 129-134.
several months. Conditions are surely relatively hy- Cameron J. N. (1978) EtTects of hypercapnea on blood
poxic. G. lateralis tends to exist in shallow dry acid-base status, NaCl fluxes, and trans-gill potential in the fresh-water blue crab, Callinectes sapidus. J. camp.
burrows in open sandy soil with good air circulation. Physiol. 123, 137-141. Reflecting such ecological differences between species Cameron J. N. (1979) Excretion of CO, in water-breathing
- there are physiological differences as well; e.g. Cardi- animals-A short review. Mar. Biol.- Lett. 1, 3-13.
soma has a low hemolymph P,,, of 4.5 torr while Cameron J. N. (1981a) Brief introduction to the land crabs , Gecarcinus has a higher P,, of 18 torr (Redmond, of the Palau Islands: stages in the transition to air
1968; Taylor and Davies, 1981). breathing. J. exp. Zool. 218, 1-5.
What of Callinectes sapidus? This aquatic species Cameron J. N. (1981b) Acid-base response to changes in
showed no special response to aerial hypoxia (Fig. 2). CO, in two Pacific crabs: The coconut crab, Birgus latro,
Voz declined somewhat early in the experiment and and a mangrove crab. Cardisoma carn(fex. J. exp. Zool. 218, 65-73.
then remained constant over a broad range of P,,? Cameron J. N. and Mecklenburg A. T. (1973) Aerial gas
Respiration of crabs in air and water 281

2x2 PAIKICIA M. O’MAHOWY mci ROW.KT J. FUL.I
exchange in the coconut crab. Bir,~rc.s kltro with some notes on Grcurcinus l~l~~nd~jd. Resp. Pl7~3iol. 19, 245-26 1.
Childress J. (1971) Respiratory adaptations to the oxygen minimum layer in the bathypelagic mysid G~~~~~~~~~~~?~~.~j~~ ingens. Biol. Bull. Mar. Biol. Lob. Woorlv Hole 141,
109-121. DeFur P. L.. Wilkes P. R. H. and McMahon B. R. (1980)
Non-equilibrium acid-base status in C. productus: Role of exoskeletai carbonate buffers. Revp. Ph.vsiol. 43, I-- 17.
Dejours P. (1978) Carbon dioxide m water- and air- breathers. Rasp. Phyriof. 33, 121-128.
Dejours P. ( 198 I ) Principfes of Cornpurrrliw Respirut~r.~
Physiology. North-Holland. Amsterdam. Dejours P. and Beekenkamp H. (1977) Crayfish respiration
as a function of water oxygen, Resp. P/z~.siol. 30,241-251. Diaz H. and Rodriguez G. (1977) The branchial chamber in
terrestrial crabs: a comparative study. Biol. BUN. 15,
437-445. Fox H. M. and Wingfield G. A. (1938) A portable apparatus
for the determination of oxygen dissolved in a small volume of water. J. esp. tliol. 15, 437.-445.
Gaudebout C. and Blayo M. C. (1975) Assessment of Scholander micromethod for gas concentration vs weigh- ing method. J. uppf. Phy.riol. 38, 546549.
Gifford f. A. (1963) Some observations on the general biology of the land crab, Curdjsumfz ~uunfz~~~~?j (Latreille) in south Florida. Biui. Bull. Mur. Biot. Lab. Woo& Ho&.
123, 2077223. Greenway P.. Taylor II. H. and Bonaventura J. (1983)
Aerial gas exchange in Australian freshwater/land crabs of the genus Holthuisunu. J. esp. Biol. 103, 237-251.
Henry R. P.. Kormanik G. A., Smatresk N. J. and Cameron J. N. (1981) The role of shell dissolution as a source of HCO, for buffering hypercapnic acidosis in aquatic and terrestrial decapod crustaceans. J. e,yp. Biol. 94, 269-274.
Herreid C. F. (1980) Hypoxia in invertebrates. Cotnp. Biochrm. Ph_vsiol. 67A, 3 I I-320.
Herreid C. F. and Gifford C. A. (1963) The burrow habitat of the land crab, CarrZisornu guunhumi (Latreille). &olo)~,t~ 44, 773-775.
Herreid C. F., O’Mahoney P. M. and Shah G. M. (1979) Cardiac and respiratory response to hypoxia in the land crab, Car~fisomu guunhumi (Latreilie). c’o,n/~. Biocham.
Physiol. 63, 145-I 5 I Herreid C. F., O’Mahoney P. M. and Full R. J. (1983)
Locomotion in land crabs: respiratory and cardiac re- sponse of Gecarinus Iutwulis. Camp. Biochem. Phwiof.
74A. 117-124. Johansen K.. Lenfant C. and Mecklenbur~ T. A. (1970)
Respiration in the crab, Cancer tnu+fcr. Z. tvrgf. Phy.r-
ioi. 70, I 19. Larimer J. L. (1961) Measurement of ventilation volume in
Decdpod Crustaceans. Physiol. Zool. 34, I58 166. Massabuau J. C., Eclancher B. and Dejours P. (1980)
Respiratory responses to longterm hypoxic stress m the crayfish Ortwwc~~c~.s z+ri/i,c. J. <‘_~p. Biof. 60, 195-206.
O’Mahoney P. M. (1977) Respir~~ti(~xl and acid-base balance in barchyuran decapod crustaceans: The transition from water to land. Ph.D. dissertation. SUNY. Buffdlo.
Pearse A. S. (1936) Estuarinc animals at Beaufort. North Carolina. J. Efi.\-hu Micchrll <Sci. Sor,. 52, 174-222.
Rahn H. (1966) Aquatic gas exchange: theory. Rq. Plz~~- iof. I, l-6.
Rahn H. and Howell B. J. (I 976) Bimodai gas exchange. In Re.~piru~iun yf Anphihious Vwrrhrute~ (Edited by G. M. Hughes). Academic Press. London.
Redmond J. R. (1968) Transport of oxygen by the blood ot the land crab. Gecarc~inus 1utrruli.c. ‘4m. Zool. 8, 411-479.
Scholander P. F. (1947) Analyzer for accurate estimation of respiratory gases in one-hail cubic centimeter samples. J. hi&. Chem. 167. 235-250.
Standaert T. (1970) Aerial and aquatic respiration of the land crab. Crrrisomu ~~~~~~~~~zj. Ph.D. dissertation. Duke University.
Taylor A. C. (1976) The respiratory responses of Curcinus
maneus to declining oxygen tension. J. ~-X/I. Biof. 64,
45-57. Taylor A. C. and Davies P. S. ( I98 I ) Respiration in the land
crab. Gecurcinus fuferufb. J. e.~p. Biof. 93, 197F208.
Taylor E. W. (1982) Control and coordination of ventilation and circulation in crustceans: responses to hypoxia and exercise. J. e_xp. Biof. 100, 289-319.
Taylor E. W. and Butler P. J. (1973) Aquatic and aerial respiration in the shore crab, Curcinus ma~nu.~ (L). accli- mated to 15 C. J. camp. Plysiol. 127, 315-323.
Taylor E. W., Butler P, J. and Al-Was&~ A. (1977) Some responses of the shore crab, Ctrrcilrtrs ~~~~~,~~.~ to progressive hypoxia at different acclimation temperatures and salinities. J. c’on?~. Ph,wiol. 122, 391402.
Taylor E. W. and Whedtley M. W. (1980) Ventilation. heart rate and respiratory gas exchange in the crayfish Ausrro- po~umhius paiiipes (Lereboullet) submerged in normoxic water and following 3 hr exposure in air at IS’ C. J. rwmp.
Ph,l%ol. 138, 67778. Taylor H. H. and Greenway P. (1979) The structure of the
gdls and lungs of the arid-zone crab, ~uffh~i.~unu (AUSIYU-
thrlphusu) /runswrw (Brachyura: Sundathelphusidae) in- cluding observations on arterial vessels within the gills. .I. Zoo/. Lond. 189, 359-384.
Truchot J. P. (1979) Mechanism of the compensation of blood respiratory acid-base disturbance of the crab. Chrinus mun~u.v (L). J. evp. ZooI. 210, 407-416.
Whcatly M. W. and Taylor E. W. (1981) The effect of progressive hypoxia on the heart rate. ventilation, respira- tory gas exchange and acid-base balance in the crayfish. .~ustropotcrmohiu.c pullipe.\. J. cup. Biol. 92, 125-14 I.
Wilkens J. L. (I 98 I) Respiratory and circulatory coordi- nation in decapod crustaceans. In Locomotion und Enrr-
Ventilatory reflex response to hypoxia in the crayfish. grtics in ~rfh~~)p~~~~~ (Edited by Herreid, C. F. II and Astucus l~ufii~~,~. J. camp. Pfl~,~siul. 140, 193-19X. Fourtner C. R.). op. 277-298. Plenum Press. New York.
McMahon’ B. k. and Burggren W. W. ( 197’1) Respiration Wolvekamp H. P.’ and Waterman T. H. (1960) Respiration. and adaptation to the terrestrial habitat in the land hermit In The Ph,vsiofogy a/’ Crustuwu (Edited by Waterman T. crab, Coenohitu cfypt~afus. J. UP. Biof. 79, 265-28 I, H.) Vol. I, pp. 35.-100. Academic Press, New York.
McMahon B. R. and Wilkens J. L. (1975) Respiratory and Wood C. M. and Randall D. J. (1981) Oxygen and carbon circulatory responses to hypoxia in the lobster Homurus dioxide exchange during exercise in the land crab (Cardi- umwicunxs. J. exp. Biol. 62, 637-655. .sc)mu curni{p.y). J. cxp. Zool. 218, 7- 16.
McMahon B. R.. Burggren W. W. and Wilkens J. L. (1974)




























