Embed Size (px)
Citation preview

Christa et al. Frontiers in Zoology 2014, 11:15http://www.frontiersinzoology.com/content/11/1/15
RESEARCH Open Access
Identification of sequestered chloroplasts inphotosynthetic and non-photosyntheticsacoglossan sea slugs (Mollusca, Gastropoda)Gregor Christa1*, Katharina Händeler1, Till F Schäberle2, Gabriele M König2 and Heike Wägele1
Abstract
Background: Sacoglossan sea slugs are well known for their unique ability among metazoans to incorporatefunctional chloroplasts (kleptoplasty) in digestive glandular cells, enabling the slugs to use these as energy sourcewhen starved for weeks and months. However, members assigned to the shelled Oxynoacea and Limapontioidea(often with dorsal processes) are in general not able to keep the incorporated chloroplasts functional. Sinceobviously no algal genes are present within three (out of six known) species with chloroplast retention ofseveral months, other factors enabling functional kleptoplasty have to be considered. Certainly, the origin of thechloroplasts is important, however, food source of most of the about 300 described species is not known so far.Therefore, a deduction of specific algal food source as a factor to perform functional kleptoplasty was still missing.
Results: We investigated the food sources of 26 sacoglossan species, freshly collected from the field, by applyingthe chloroplast marker genes tufA and rbcL and compared our results with literature data of species known fortheir retention capability. For the majority of the investigated species, especially for the genus Thuridilla, we wereable to identify food sources for the first time. Furthermore, published data based on feeding observations wereconfirmed and enlarged by the molecular methods. We also found that certain chloroplasts are most likelyessential for establishing functional kleptoplasty.
Conclusions: Applying DNA-Barcoding appeared to be very efficient and allowed a detailed insight into sacoglossanfood sources. We favor rbcL for future analyses, but tufA might be used additionally in ambiguous cases. We narroweddown the algal species that seem to be essential for long-term-functional photosynthesis: Halimeda, Caulerpa, Penicillus,Avrainvillea, Acetabularia and Vaucheria. None of these were found in Thuridilla, the only plakobranchoideangenus without long-term retention forms. The chloroplast type, however, does not solely determine functionalkleptoplasty; members of no-retention genera, such as Cylindrobulla or Volvatella, feed on the same algae as e.g.,the long-term-retention forms Plakobranchus ocellatus or Elysia crispata, respectively. Evolutionary benefits offunctional kleptoplasty are still questionable, since a polyphagous life style would render slugs more independentof specific food sources and their abundance.
Keywords: DNA-barcoding, Food analyses, Kleptoplasty, rbcL, Sacoglossa, tufA
BackgroundSacoglossa sea slugs are a relatively small group ofheterobranch gastropods with currently about 300 spe-cies described [1], comprising the shelled Oxynoaceaand the shell-less Plakobranchacea. The latter is furtherdivided into the ceras-bearing Limapontioidea (although
* Correspondence: [email protected] Forschungsmuseum Alexander Koenig, Adenauerallee 160,53113 Bonn, GermanyFull list of author information is available at the end of the article
© 2014 Christa et al.; licensee BioMed CentralCommons Attribution License (http://creativecreproduction in any medium, provided the orDedication waiver (http://creativecommons.orunless otherwise stated.
not monophyletic in phylogenetic analyses) and the para-podia bearing Plakobranchoidea [2-4]. Because of theirability to incorporate functional chloroplasts and subse-quently use them for sustenance during starvation overweeks and months Sacoglossa fascinated scientists overdecades. This feature is unique among metazoan lifeforms [3,5-8] and only known elsewhere in members ofthe Foraminifera (Rhizaria) [9], Ciliophora (Alveolata) [10]and in Dinoflagellata (Alveolata) [11]. Within Sacoglossausually three states of kleptoplasty are differentiated, either
Ltd. This is an Open Access article distributed under the terms of the Creativeommons.org/licenses/by/2.0), which permits unrestricted use, distribution, andiginal work is properly credited. The Creative Commons Public Domaing/publicdomain/zero/1.0/) applies to the data made available in this article,

Christa et al. Frontiers in Zoology 2014, 11:15 Page 2 of 12http://www.frontiersinzoology.com/content/11/1/15
based on measurements of functional chloroplasts withPulse Amplitude Modulated (PAM) Fluorometry or onCO2 fixation experiments [3,12,13]. Following the clas-sification of Händeler et al. [3], specimens that are notable to incorporate chloroplasts functional are consid-ered as no-retention forms (NR); specimens that areable to incorporate functional chloroplasts for up totwo weeks of starvation are called short-term-retentionforms (StR) and those incorporating functional chloro-plasts for over 20 days during starvation are called long-term-retention forms (LtR). The fact that chloroplastssurvive for weeks and months in the slug’s digestive systemand perform functional photosynthesis, despite the absenceof the algal nucleus, has led to the intriguing hypothesisthat a horizontal gene transfer must have occurred fromthe algal organisms into the metazoan life form [7,12,14].Up to now, a few single genes amplified from Elysia chlo-rotica, Gould [15] were interpreted to encode proteinsrelevant for photosynthesis, such as Lhc (light harvestingcomplex), fcp (fucoxanthin protein), psbO (manganese sta-bilizing protein), and others [7,12,16,17].Analyzing transcriptomic data for the first time, Wägele
et al. [18] found no evidence for a horizontal gene transferin two sacoglossan species known to maintain chloro-plasts for several months, Elysia timida Risso [19] andPlakobranchus ocellatus van Hasselt [20]. Similar negativeresults were obtained later for Elysia chlorotica by tran-scriptomic and genomic data [21,22]. Conflicting resultsof Pierce et al. [23] that suggested a gene transfer basedon few single reads in their transcriptomic data, have mostrecently been interpreted in a different way [24]. Thus, inour view the hypothesis that slugs are able to activelysupport the kleptoplasts via the translational products oftransferred algal genes is rejected and the focus for under-standing long-term incorporation of kleptoplasts has toinclude other factors, like properties of chloroplasts andtheir origin with regard to functional photosynthesis.We clearly demonstrated this recently by investigatingPlakobranchus ocellatus during various starvation pe-riods [25]: Despite a broad documented food range infresh caught animals, only chloroplasts from the ulvo-phycean Halimeda macroloba Decaisne [26] remainedin the digestive gland after a starvation period of twomonths. Unfortunately, our knowledge on sacoglossanfood preferences is still scarce. In general the slugs areconsidered to be stenophagous and mainly sequestermembers of the Ulvophyceae sensu Floyd & O’Kelly[22,27-33]. Few species (Elysia crispata, E. clarki andP. ocellatus) are known to feed on a high variety of algae[18,25,30,34-37]. Only some species are recorded to feedon specific members of Rhodophyta (e.g., Hermaea bifida)[38,39], Heterokontophyta (e.g., Elysia chlorotica) or seagrasses (Elysia serca, Marcus [40]) [3,30,31,41]. Foodsources were usually identified by observation, feeding
experiments [28,33,42], or by electron microscopicallystudies of chloroplast types [43,44]. But it is obvious that,especially in potential polyphagous sea slugs, not everyfood alga may be detected by feeding experiments, inparticular when not knowing, which ones to offer. On theother hand, slugs may feed on alternative food sourcesduring food limitation rather than having these as hostalgae. Molecular barcoding has proved to be a high effi-cient method to identify algal food sources even whenusing a single barcoding-marker [30,34-37], instead of two[25]. This now well-established method opens the oppor-tunity to study sacoglossan food sources and origin ofkleptoplasts in a highly reliable mode. To find a pattern inchloroplast origin and functional kleptoplasty, a profounddatabase on sequestered chloroplasts for all sacoglossangroups, and especially those taxa hardly studied at all, e.g.,the genus Thuridilla [18,30], is needed. Following themethods introduced previously [30,34,35] we investigatedchloroplast origin in 26 non-starved sacoglossan species,including NR, StR and LtR forms, by DNA-barcodingusing the chloroplast markers tufA and rbcL to enlargeour insight in, and state more precisely, sacoglossan foodspectrum. We combined the identified food sources withliterature data of species for which retention ability is doc-umented and analyzed in combination with this informa-tion if there is a correlation between food sources andretention-form. DNA-barcoding has become an importantmethod in identifying plastid origin in Sacoglossa, but dueto varying results and minor pitfalls in the application ofrbcL, we compared reliability of this gene.
ResultsBarcoding of tufA and rbcLWe successfully identified food sources for 30 sacoglossanspecimens (overall 26 species) by at least applying onebarcoding marker (Additional file 1). For 19 of the 26species included here we were able to identify the foodsources for the first time. For the remaining seven species,we could confirm literature data or even enlarged the foodrange (Additional file 1). Yet, no rbcL amplification prod-ucts were obtained for four specimens and no tufA prod-uct was obtained for seven specimens (Additional file 1).TufA sequences obtained from Elysia amakusana (703)Baba [45], Elysia sp. (841), Cyerce nigra (860) Bergh [46]and Costasiella sp. (863) exclusively represented sequen-ces of bacterial origin.We were able to identify 14 different algal genera com-
bining both markers: eleven genera can be assigned toBryopsidophyceae, two to Dasycladophyceae and one toUlvophyceae. RbcL revealed about two times more diffe-rent haplotypes than tufA (30 for rbcL and 14 for tufA,respectively, Figures 1 and 2), though the number ofgenera is equal (9 for rbcL and 10 for tufA, respectively).Nine haplotypes for rbcL and three haplotypes for tufA

could not be assigned to a certain algal genus, becauseof missing reference sequences in GenBank. We are notable to clarify if some of the unidentified haplotypes ofrbcL and tufA actually represent the same algal species.Especially for rbcL almost always a higher number ofalgal haplotypes for a distinct sacoglossan species wasrevealed, with the highest number of haplotypes for anunknown sister clade of Pseudochlorodesmis (7) and forBryopsis (6) (Additional file 1, Figures 1 and 2).
Food sources and retention formWe combined the information obtained from barcodingand literature data, and analyzed this information withregard to the according retention-form. We found thatthe food of NR forms covers a broad spectrum of ulvophy-cean algae and includes items that are not consumed byStR or LtR forms: Boodlea, Chlorodesmis, Ulva, Urospora,
red algae, angiosperms and eggs of other sacoglossans(Figure 3). In contrast, algal taxa like Poropsis, Rhipidosi-phon, Rhipocephalus, Udotea and an unidentified memberof the Ulvophyceae were not identified so far as foodsources for NR-forms.LtR forms consume various ulvophycean algal species,
but prefer Halimeda, Caulerpa, Penicillus and Avrainvillea,the dasycladalen Acetabularia and the heterokontophyteVaucheria (Figure 3). However, some NR and LtR formsbelonging to different sacoglossan families [5] seques-ter the same algal species (e.g. Halimeda, Caulerpa,Avrainvillea, Vaucheria); therefore a correlation bet-ween food sources and functional kleptoplasty cannotbe solely algal based. Interestingly, Thuridilla speciesthat only comprise StR forms rarely feed on plastidssequestered by LtR forms of Elysia and Plakobranchusocellatus (Figure 3).
Figure 1 Food sources identified using tufA as barcode marker. ML tree of identified algal haplotypes in Sacoglossa by using tufA (highlighted ingreen). Identified haplotype is noted on the right side. When sequence match was < 99%, higher taxon name of the algae that formed a monophyleticgroup with the corresponding haplotype was used. Haplotypes with no monophyletic grouping are named “Ulvophyceae sp.”.
Christa et al. Frontiers in Zoology 2014, 11:15 Page 3 of 12http://www.frontiersinzoology.com/content/11/1/15

Within sacoglossan clades, differences in preferredfood items exist. All investigated members of theOxynoacea are NR forms and stenophagous. This isin contrast to all other (non oxynoacean) sacoglossangenera that include polyphagous species (Figure 3).Cylindrobulla is the only member of the Oxynoaceathat exclusively feeds on Halimeda, whereas the membersof the remaining Oxynoacea specialized on Caulerpaspecies, with one exception that the rbcL analysis inLobiger viridis revealed additional Halimeda as food source(Additional file 1, Figure 3). Within Limapontioidea, sev-eral species have a broad food spectrum, like Costasiellasp. (864) with at least four and Cyerce nigra Bergh [46]with at least five different food sources, respectively. How-ever, Halimeda and Caulerpa, the major oxynoacean food,
is scarcely represented amongst the food items ofLimapontioidea and only found in Cyerce antillensis andPolybranchia viridis, respectively. Interestingly, Costasiellaocellifera Simroth [48], the only non-plakobranchoid LtRform ([49], unpublished data), feeds exclusively on Avrain-villea nigricans.Four genera of the Plakobranchoidea that all in-
clude functional retention forms, are investigated. Boselliamimetica is stenophagous, feeding on Halimeda, contraryto the polyphagous Plakobranchus ocellatus that feeds onseven different algae (including Halimeda, Caulerpa andUdotea) exhibiting the most diverse food spectrum of allinvestigated sacoglossans so far (Figure 3); however,these data are obtained from different specimens of differ-ent localities [25,37]. The geographically wide distributed
Figure 2 Food sources identified using rbcL as barcode marker. ML tree of identified algal haplotypes in Sacoglossa by using rbcL(highlighted in blue). Identified haplotype is noted on the right side. When sequence match was < 99%, higher taxon name of the algae that formeda monophyletic group with the corresponding haplotype was used. Haplotypes with no monophyletic grouping are named “Ulvophyceae sp.”.
Christa et al. Frontiers in Zoology 2014, 11:15 Page 4 of 12http://www.frontiersinzoology.com/content/11/1/15
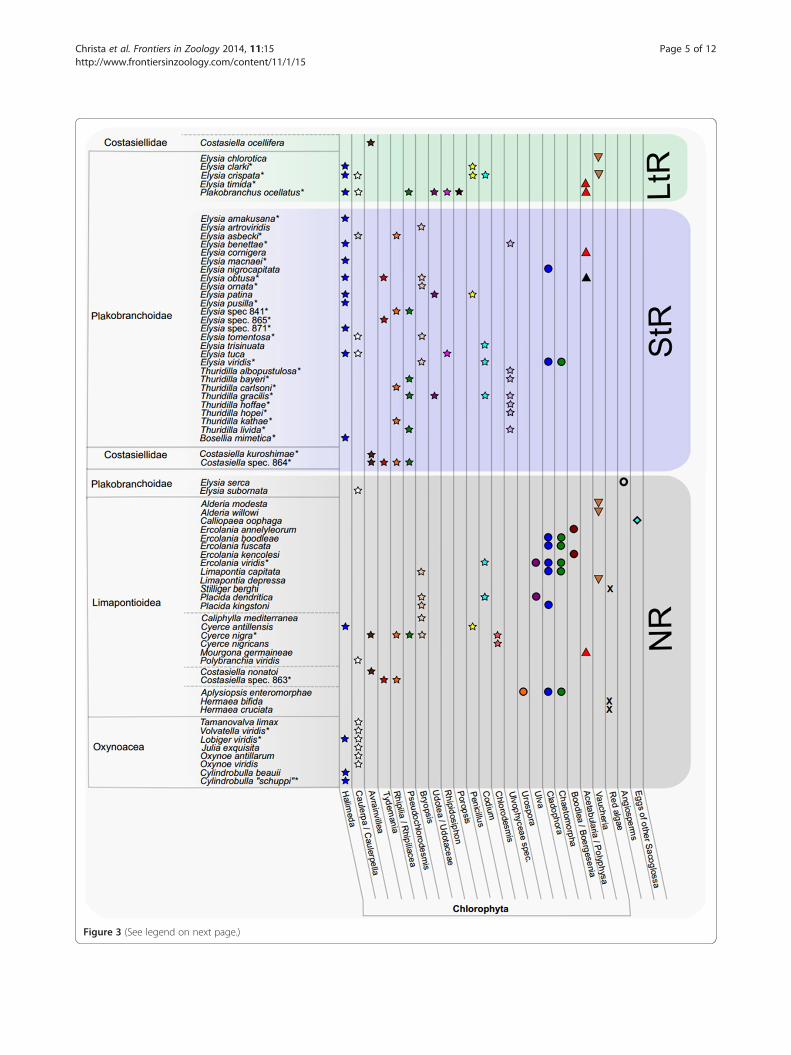
Figure 3 (See legend on next page.)
Christa et al. Frontiers in Zoology 2014, 11:15 Page 5 of 12http://www.frontiersinzoology.com/content/11/1/15

(See figure on previous page.)Figure 3 Food spectrum of Sacoglossa compared to functional retention form. Food sources of 68 species of Sacoglossa are shown. Theclassification of Sacoglossa is based on works of [3,4,46]. Asterisk indicates molecular identification of food (overall 33 species). Food sources wereeither obtained in this study or taken out of literature [3,4,18,25,30,34-37,80-83]. Classification of retention ability was done according to literaturedata [3,18,47,49,80-84]. LtR= Long-term-retention, StR= short-term-retention, NR= no retention. Stars indicate members of the Bryopsidales, filled circlesmembers of the Ulvophyceae other than Bryopsidales; triangles represent Dasycladales, inverted triangles Heterokontophyta, X red algae, circle sea grassand diamond eggs of other Sacoglossa.
Christa et al. Frontiers in Zoology 2014, 11:15 Page 6 of 12http://www.frontiersinzoology.com/content/11/1/15
taxon Plakobranchus might show geographic differencesin the consumed food items. Furthermore, cryptic spe-ciation with already ecological differences in feeding canalso not be ruled out [50]. Based on the molecular phyl-ogeny of Händeler et al. [3], Thuridilla is the sister taxonto Plakobranchus, its members principally feeding on avariety of food sources, though Thuridilla species neitherconsume Halimeda, nor Caulerpa or Poropsis (food sour-ces of Plakobranchus), but a bryopsidophycean genusrelated to Pseudochlorodesmis (Figure 2). This peculiarfood item could not be identified more specifically due tolack of reference sequences in GenBank. Members of thefourth investigated genus, Elysia, consume food itemsrecorded also for the shelled sacoglossans, Halimeda andCaulerpa. Additionally, many Elysia species feed on thesame bryopsidophycean genera similar to those observedin Thuridilla (exceptions see above) or Plakobranchus.Some species became very specialized. Elysia chloro-tica feeds on Vaucheria (like the limapontioidean genusAlderia), whereas its sister taxon, Elysia serca, is reportedto feed solely on higher plants (i.e., sea grasses [31]), aunique feature within Sacoglossa. Several species are con-firmed in their specific narrow food spectrum, e.g., Elysiatimida, which exclusively feeds on the dasycladophyceanAcetabularia acetabulum P.C. Silva [51].
DiscussionFood sources and retention formWe analyzed a wide spectrum of sacoglossan sea slugswith regard to their food preferences and plastid originby using a combination of two barcoding markers, rbcLand tufA, instead of one [3,18,34-37]. This method provedto be more rapid and more precise in identification offood sources of a large set of Sacoglossa sea slugs, com-pared to former feeding experiments or feeding observa-tions. Although not every algal food source is identifiedon species level, the information we provide here increasesour knowledge considerably and can now be used forfurther ecological or behavioral studies.We found that the ability to perform photosynthesis
for at least several days and weeks is established withcertain food sources (members of the genera Elysia, Boselliaand Plakobranchus, see Figure 3 and [3]). At least one ofthe following six algae, the bryopsidophycean Halimeda,Caulerpa, Penicillus and Avrainvillea, the heterokontophyteVaucheria, or the dasycladophycean Acetabularia, seem to
be essential for establishing long-term-retention. However,non-retention forms also feed on these specific algae andare not able to perform photosynthesis. Both aspectsindicate that a special type of chloroplast is not sufficientto establish functional kleptoplasty and intrinsic factors ofthe slugs also contribute to a functional photosynthesis.Here, we pave the way for further research on these spe-cific algae consumed by LtR forms to better understandwhich genetically and physiological commonalities thesetarget plastids may have. Pelletreau et al. [21] recentlyconsidered special abilities of chloroplasts from respectivehost algae as relevant for long-term incorporation, anoption already mentioned earlier [52,53]. A factor nowfound to possible enhance plastid longevity is ftsH, aprotein important for the repair of Photosystem II. It isplastid encoded in the food algae of the two LtR formsElysia chlorotica and Plakobranchus ocellatus [54]. Withthe information of this study we are now able to specific-ally examine the presence of ftsH in plastids of the identi-fied food sources of other retention forms and correlateplastid origin, photosynthetic capability and plastid lon-gevity. Interestingly, Thuridilla species – as close relativesof Plakobranchus and Elysia – and which exhibit extre-mely short chloroplast retention [3,55], did not reveal anyof these six algal species mentioned above (Additionalfile 1, Figure 3).Besides plastid origin, several other factors surely in-
fluence survival of chloroplasts and render them nutri-tional at least in sacoglossans in one way or the other. Ahorizontal gene transfer as a factor was excluded notonly for the sea slugs Plakobranchus ocellatus, Elysiatimida and E. chlorotica [18,22], but also for the Foram-inifera Elphidium margaritaceum and most likely for thedinoflagellate Dinophysis acuminate [56,57]. Common toall three different systems is that there is no mechanicalgrinding before chloroplast uptake. E.g., the sacoglossanspierce the cell wall with their teeth and then suck outthe cytoplasm of the algae, which is then processed inthe digestive gland. Second, the intracellular digestion ofplastids seems to be stopped or regulated in a way yet tobe discovered.Händeler et al. [3] have shown in their phylogenetic
analysis by using ancestral character state reconstruc-tion, that the ancestor of the Plakobranchoidea mostlikely developed an unknown mechanism, which hin-ders direct digestion of chloroplasts. However, the genus

Christa et al. Frontiers in Zoology 2014, 11:15 Page 7 of 12http://www.frontiersinzoology.com/content/11/1/15
Costasiella was not included in their analysis, which isnow verified in comprising both, LtR and StR species([49], [58], unpublished data). Thus, at least in Costasiella,this mechanism must be developed as well. Evertsen &Johnsen [59] showed that in Elysia viridis starch grainspersist throughout degradation of plastids and are notbroken up, as was the case in the NR form Placida dendri-tica (Alder & Hancock) [60]. If the slugs cannot access thephotosynthetic produced starch of functional chloroplastsin the first place, the benefit, under starvation conditions,has to arise from another process. In this case the accu-mulated photosynthates would only become accessible forslug metabolism after the complete degradation of thechloroplast. The nutritional benefit of the chloroplastswould lie in the presence of an additional food reservoir(starch) that becomes available only after degradation ordigestion of the chloroplasts. This is in contrast to formerhypotheses that usually assume a continuous supply ofphotosynthates and therefore a continuous exchange bet-ween chloroplast and slug’s cytosol [61,62].
General food source of SacoglossaOur results in combination with literature data show thatsacoglossans generally feed on a high variety of algal spe-cies, although the majority prefers bryopsidophycean taxa.Jensen [4] assumed Halimeda as the ancestral food ofSacoglossa based on her observations on Cylindrobulla.We can confirm here by molecular analyses of the gutcontent that this species exclusively feeds on Halimeda,whereas all other oxynoacean species feed on Caulerpa,though Lobiger viridis seems also to at least feed additionalon Halimeda. The position of Cylindrobulla within theOxynoacea is not resolved yet, thus the ancestral foodsource of this clade cannot be deduced [3,63-65]. Futurephylogenetic studies are needed to address these questions.All higher sacoglossan taxa switched to other food sourcesthan Halimeda (especially members of the Limapon-tioidea), or broadened their food spectrum (many Elysiaspecies).Why some species feed on several different algal spe-
cies and others specialized on just one prey species, isdifficult to explain. Further information on biology hasto be considered, e.g., incorporation of defensive com-pounds obtained via sequestration of toxic algae, or denovo synthesis (rendering the slug independent fromalgal toxins). We have to emphasize that we were notable to study all species on a broad scale by includingspecimens from various geographic areas or seasons.Food availability and seasonality, and/or intraspecific orinterspecific competition might also force specimensfrom the same species to switch to other food items andthus influences the finding of certain food items in saco-glossan sea slugs. Further starvation studies on polypha-gous species (see e.g. [25]) will verify if these species have
a preferred food source, as was shown for Plakobranchusocellatus and these investigations will certainly contributeto our understanding of polyphagous strategies [37].Jensen [22,53] studied morphological differentiation of
the radula teeth in connection with algal food. Based onher findings she suggested that certain radula teeth arecorrelated with the polysaccharide of cell walls of therespective food sources. However, Händeler & Wägele[23] mapped radula teeth forms on their molecular basedcladogram and were not able to confirm this previouslysuggested correlation. We therefore compared three un-usual food switches within the Sacoglossa with regard to apossible correlation of food source and radula tooth shapei) The NR form Hermaea bifida (Montagu) [66] feeds onthe red algae Griffithsia with cell walls composed of cellu-lose. However, the radula of H. bifida is not different fromspecies feeding on the green alga Caulerpa with cell wallscomposed of xylan [67]. ii) Elysia serca, also a NR form[68], presents a unique switch to sea grasses with cell wallscomposed of cellulose [28,29], but the radula is similar tospecies feeding on Halimeda with cell walls composed ofxylan. iii) The NR forms Alderia modesta and A. willowi,as well as the LtR form Elysia chlorotica, feed on theheterokontophyte genus Vaucheria composed of cellulose-based cell walls. The radulae of Alderia species and Elysiachlorotica are similar to species feeding on the green algaCodium composed of mannan-based cell walls. Thus, wethink that different radula shapes did not force or influencea host switch in these three examples, though it cannot beruled out for others that are not examined here.Whether functional kleptoplasty is of higher evolution-
ary benefit than e.g., feeding on a wide spectrum of algaemay be reconsidered. A polyphagous strategy –in contrastto stenophagous strategies – would render the slugs moreindependent of seasonality or general availability of theirhost algae. LtR forms like Costasiella ocellifera, Elysiachlorotica and Elysia timida feed on one algal species.Acetabularia acetabulum, the only natural food of E.timida, calcifies in fall, and therefore probably forcesElysia timida to rely on its incorporated chloroplasts [69].Unfortunately we have no similar information on season-ality of Avrainvillea and Vaucheria, the major food itemsof C. ocellifera and E. chlorotica, respectively. Plakobran-chus ocellatus on the other side has a broad foodspectrum. As Maeda et al. [37] showed in a subtle experi-ment, the algae show a high seasonality and the slugscombine here functional kleptoplasty and multiple foodsources that certainly enhances survival.
Usefulness of barcoding markersRbcL and tufA are now state of the art markers for bar-coding algal species [70] and are therefore intensively usedfor identification of sequestered chloroplasts in Sacoglossa[25,30,34-37]. So far it is not possible to identify the food
Christa et al. Frontiers in Zoology 2014, 11:15 Page 8 of 12http://www.frontiersinzoology.com/content/11/1/15
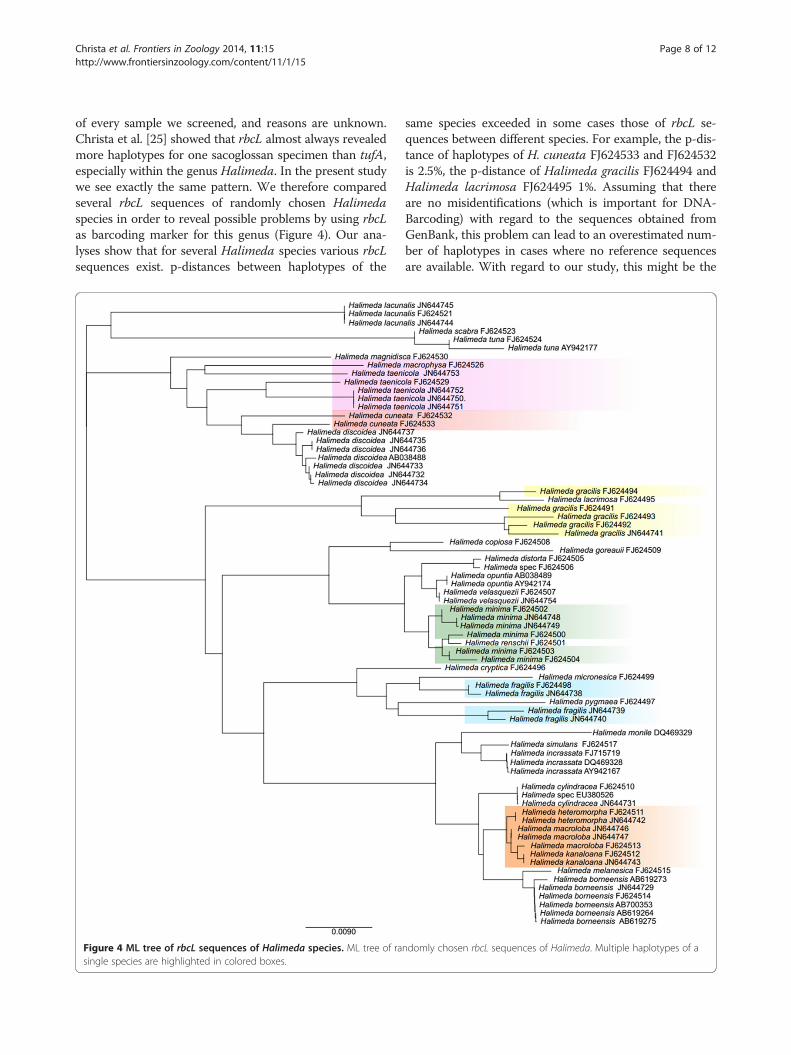
of every sample we screened, and reasons are unknown.Christa et al. [25] showed that rbcL almost always revealedmore haplotypes for one sacoglossan specimen than tufA,especially within the genus Halimeda. In the present studywe see exactly the same pattern. We therefore comparedseveral rbcL sequences of randomly chosen Halimedaspecies in order to reveal possible problems by using rbcLas barcoding marker for this genus (Figure 4). Our ana-lyses show that for several Halimeda species various rbcLsequences exist. p-distances between haplotypes of the
Figure 4 ML tree of rbcL sequences of Halimeda species. ML tree of rasingle species are highlighted in colored boxes.
same species exceeded in some cases those of rbcL se-quences between different species. For example, the p-dis-tance of haplotypes of H. cuneata FJ624533 and FJ624532is 2.5%, the p-distance of Halimeda gracilis FJ624494 andHalimeda lacrimosa FJ624495 1%. Assuming that thereare no misidentifications (which is important for DNA-Barcoding) with regard to the sequences obtained fromGenBank, this problem can lead to an overestimated num-ber of haplotypes in cases where no reference sequencesare available. With regard to our study, this might be the
ndomly chosen rbcL sequences of Halimeda. Multiple haplotypes of a

Christa et al. Frontiers in Zoology 2014, 11:15 Page 9 of 12http://www.frontiersinzoology.com/content/11/1/15
case for several algal sequences obtained from Thuridilla,which are now assigned as seven different unidentifiedhaplotypes. It is very likely that these problems do notonly occur in the taxon Halimeda, but also in other ulvo-phyceaen taxa where detailed studies are still missing.Händeler et al. [30] demonstrated for Caulerpa that iden-tification and annotation using tufA on species level couldalso be problematic due to sequence similarities. Hanyudaet al. [71] reported introns in rbcL sequences frommembers of the Caulerpaceae, a problem we faced inCaulerpa sequences obtained from Volvatella viridis.These sequences cannot be aligned with the remainingrbcL sequences, and were therefore not included in ouranalyses here.An additional problem for identifying food organisms
in Sacoglossa is connected to the lack of references forboth genes in the databases. Available information doesnot cover many of the obtained haplotypes; therefore anextension of algal taxon sampling in the future by algalspecialists is absolutely necessary. Here we show that theuse of two markers for plastid identification in Sacoglossaonly leads to a more detailed identification of ingestedplastids in some species. Yet, we suggest that, based onthe higher reliability and despite possible multiple haplo-types for some genera, rbcL should be used preferentially.TufA may be added in cases where an identification ofrbcL failed or sequence variability of rbcL hinders unam-biguous identification. Unfortunately both primer pairsare still not applicable at the moment for Cladophoraceae[71,72]. Constructing new primers for both genes basedon chloroplast genomic data probably could solve thisproblem as soon as a cladophoraceaen plastid genome isavailable. Until then it has to be kept in mind that barcod-ing results may cover only a reduced food range. Evenwith the limitations mentioned above, we consider DNA-Barcoding a more accurate and effective method thanfeeding observations, especially in polyphagous specimens.
ConclusionsDNA-Barcoding of sequestered chloroplasts in Sacoglossaled to a more specific insight in food sources of sacoglos-san sea slugs than by direct observation and is certainlymuch more efficient. RbcL should be used primarily be-cause of higher amplification and sequencing success,while tufA should be added in analyses with ambiguousresults.Our results revealed food items for many species not
investigated before and additionally confirmed literaturedata. We could show that food sources are highly variablein some species and morphological features such as radulashape have probably only little impact on food preferencesor food switches. There seem to be certain food sourcesthat are essential for functional kleptoplasty. LtR formspreferably consume algal species belonging to the genera
Halimeda, Caulerpa, Avrainvillea, Acetabularia or Vau-cheria. Kleptoplasty is only established in slugs feeding ongreen algae and heterokontophytes, but not in slugs feedingon rhodophytes and sea grasses. NR forms may also feedon algae that are known to be the sole food of LtR forms.Factors like physiology of food items, genetic and physio-logical properties of the plastids and digestion properties inslugs need to be more investigated in future studies toreveal principles of establishing functional kleptoplasty. Theevolutionary benefit of kleptoplasty is still enigmatic, sincea polyphagous life style may lead to more independencefrom specific food source’s seasonality and abundance.
MethodsSacoglossan specimens used in this study were identifiedby morphological examination using a taxonomic speciesfile based on original literature ([73] on Thuridilla),as well as identification books (e.g., [74]) and the seaslug forum (http://www.seaslugforum.net). These iden-tified specimens, covering NR, StR and LtR forms, arelisted in Additional file 1 and represent those for whichDNA-Barcoding of food source was successful, in order todetermine the minimal food items of these species. Speci-mens were fixed in 96%-EtOH immediately after collec-tion. Slug parts containing digestive glandular tissuewere cut off and subsequently DNA extracted usingthe DNeasy® Blood & Tissue Kit (Qiagen, Germany) follo-wing manufacturer’s instructions and stored at -20°C.
DNA amplificationStandard PCR reactions for rbcL and tufA as reportedelsewhere [25,30,34-37] were performed using a touch-down protocol and ulvophycean specific primers. 2.5 μlof genomic DNA was used as template in a 20 μl finalvolume reaction supplied with 5.5 μl sterilized water,2 μl Qiagen® Q-Solution, 10 μl of double concentratedQIAGEN® Multiplex PCR Master Mix and 1 μl of 5 pmol/μlconcentrated primer each. PCR for amplification of rbcLwas performed with primer pairs rbcL 1 ([36] and rbcL R,5′-CCA WCG CAT ARA NGG TTG HGA-3′ ([25] mo-difid after [36]) by an initially denaturation for 15 min at95°C, followed by 9 touch-down cycles at 94°C for 45 s,53°C (-1°C per cycle) for 45 s, 72°C for 90 s, followed by25 standard cycles (94°C for 45 s, 45°C for 45 s and 72°Cfor 90 s.) and a final extension at 72°C for 10 min. tufAamplification was performed with primer pair tufAF andtufAR [75] by an initially denaturation for 15 min at 95°C,followed by 9 touch-down cycles at 94°C for 45 s, 57°C(-1°C per cycle) for 45 s, 72°C for 90 s, followed by 25standard cycles (94°C for 45 s, 48°C for 45 s and 72°C for90 s.) and a final extension at 72°C for 10 min. PCRproducts were size-fractionated in a 1.5% agarose gelfor 90 min at 70 V and bands according to desired gene-fragment length and subsequently gel-extracted using

Christa et al. Frontiers in Zoology 2014, 11:15 Page 10 of 12http://www.frontiersinzoology.com/content/11/1/15
Machery-Nagel NucleoSpin® Extract II (Düren, Germany)kit following manufacturer’s instructions. Isolated frag-ments were ligated into pGEM T-easy Vector (Promega,Germany) and cloned into competent E. coli XL1-bluecells from Stratagene (Heidelberg, Germany). For eachspecimen 12 clones were sequenced by Macrogen Inc,Amsterdam.
Sequence analysisSequence identity of tufA and rbcL of every clone wasverified by BLAST search using Geneious (BiomattersLtd, New Zealand, v. 6.0.3). Consensus sequences of oneslug specimen were created when sequence divergenceof chloroplast genes was lower than 1%, as introducedby Händeler et al. [30] (see Additional files 2, 3 and 4).All gained sequences were again verified by BLASTsearch and the first top 5 BLAST results of each sequencewere taken to create a dataset of overall 54 sequences fortufA (688 bp) and 94 sequences for rbcL (561 bp). Bothdatasets were aligned with MAFFT plugin as implementedin Geneious (v6.814b [76]). A maximum likelihood treefor each dataset was calculated using PhyML plugin [77]implemented in Geneious with GTR + I + R as substitu-tion model. For final identifying sequence origin we useda combination of similarity-based and tree-based DNA-barcoding approaches [78,79]: Sequences were assigned toa certain algal species by a similarity-based approach usingBLAST analysis when sequence similarity was 99-100%.We included a 99% match as positive identification of analgal species, thus allowing a small amount of ambiguitynucleotides within our consensus sequences which are notidentified as similarity by the BLAST search in NCBI(www.ncbi.nlm.nih.gov). When multiple positive identi-fications were obtained, sequences were assigned to thecorresponding genus based on the position within the MLtree. When not able to define on species level (BLASTsimilarity <99%), sequences were assigned to higher taxabased on their position within monophyletic algal groupsin the ML tree analysis. When sequences obtained in ourstudy did not group within distinct algal taxa, we assignedthem to the taxon level Ulvophyceae.
Sequence analysis of rbcL sequences of HalimedaRandomly chosen rbcL sequences of Halimeda specieswere downloaded from Genbank to analyze sequencesvariability. Overall 72 sequences were used, representing32 species. Dataset was aligned with MAFFT plugin asimplemented in Geneious (v6.814b [76]). A maximumlikelihood tree for the dataset was calculated using PhyMLplugin [77] implemented in Geneious with GTR + I + Ras substitution model. P-distances of sequences werecalculated using HyPhy (v 2.1). On the basis of thesep-distances the sequence variability of haplotypes of thesame algal species were then analyzed.
Comparison of literature data and new barcoding dataDNA-Barcoding results of the present study were com-bined with results taken from literature to compare foodsources of different retention forms (LtR, StR, NR).Literature data on food sources and retention form weretaken from [3,4,18,25,30,34-37,80-84] (Additional files 1and 5). The retention ability of the majority of thespecies investigated here was measured by means of aPAM. Therefore, we used the classification scheme ofHändeler et al. [3]. Results of Clark et al. [68] based on 14Cmethods were re-assigned, when new data were available.For example, Mourgona germaineae Marcus, 1970 isclassified as StR form by Clark et al. [68], however PAMmeasurements revealed no fluorescence at all (unpub-lished data). We therefore set 14C obtained by valuessimilar to M. germaineae as “no retention”, e.g. Caliphyllamediterranea Costa [85], Hermaea cruciata, Placida king-stoni Thompson [86] and Elysia serca. Most recentlySchmitt et al. [87] could show that E. timida can be artifi-cially raised on Acetabularia peniculus, an algal species thatnaturally does not co-occur with the slugs. Similar reten-tion of plastids was observed as from the natural food A.
Additional files
Additional file 1: Origin of specimen and identified food sourceswith comparison of literature data. In Table S1 we provide a list ofSacoglossa specimens and species analyzed with regard to food items.Table S2 displays the identified food sources in Sacoglossa specimens byanalyzing the chloroplast markers tufA and rbcL. The number ofhaplotypes per gene obtained from each slug specimen is listed in thelast two columns. * tufA sequences of Thuridilla hopei (809), Vérany werepublished previously [18]. Table S3 shows a comparison of chloroplastorigin by feeding observations (4th column) and identified by DNA-Barcoding(this study and literature data – last column). When not specified specifically,information on retention-form is taken from Händeler et al. [3]. Food sourcesidentified by using tufA are indicated with 1, by rbcL with 2 in the lastcolumn. Literature data on food sources base on the review of Händeler &Wägele [29] or is indicated otherwise.
Additional file 2: Sequence affinity based on BLAST search.
Additional file 3: Alignment of tufA sequences used for consensussequences.
Additional file 4: Alignment of rbcL sequences used for consensussequences.
Additional file 5: Classification on retention form in Sacoglossa.
Competing interestsThe authors declare that they have no competing interests.
Authors’ contributionsHW, KH and GC conceived and designed the experiment. GC, TS and KHperformed the experiments. HW, GC and KH analyzed the data. GC, TS, GK andHW wrote the paper. All authors read and approved the final manuscript.
AcknowledgmentsWe thank all collectors Ingo Burghardt, Yvonne Grzymbowski, AnnetteKlussmann-Kolb, Patrick Krug, Wolfgang Loch, Tane Sinclair, Kristina Stemmer,Carmen Zinßmeister, and the directors of the Lizard Island Research Station,Anne Hoggett and Lyle Vail and the director of the Marine Laboratory,Guam, Peter Schupp, providing us with material. Emilie Goralski and ClaudiaEtzbauer helped with the molecular work. Anonymous reviewers provided

Christa et al. Frontiers in Zoology 2014, 11:15 Page 11 of 12http://www.frontiersinzoology.com/content/11/1/15
valuable information and helped considerably in improving the first versionof this manuscript. We thank the German Science Foundation (DFG) forfinancial support to HW (Wa 618/8 and Wa 618/12). GC was partlyfinanced by an ERC grant to W. Martin (Düsseldorf, Germany): NetworkoriginsProj. Ref. 232975.
Author details1Zoologisches Forschungsmuseum Alexander Koenig, Adenauerallee 160,53113 Bonn, Germany. 2Institute for Pharmaceutical Biology, University ofBonn, Nussallee 6, 53115 Bonn, Germany.
Received: 9 October 2013 Accepted: 6 February 2014Published: 21 February 2014
References1. Jensen KR: Biogeography of the Sacoglossa (Mollusca, Opisthobranchia).
Bonn Zool Beitr 2007, 55:255–281.2. Kohnert P, Brenzinger B, Jensen KR, Schrödl M: 3D- microanatomy of
the semiterrestrial slug Gascoignella aprica Jensen, 1985—a basalplakobranchacean sacoglossan (Gastropoda, Panpulmonata). Org DiversEvol 2013, 13:583–603.
3. Händeler K, Grzymbowski YP, Krug PJ, Wägele H: Functional chloroplasts inmetazoan cells - a unique evolutionary strategy in animal life. Front Zool2009, 6:28.
4. Jensen KR: Evolution of the Sacoglossa (Mollusca, Opisthobranchia) andthe ecological associations with their food plants. Evol Ecol 1997,11:301–335.
5. Muscatine L, Greene RW: Chloroplasts and algae as symbionts inmolluscs. Int Rev Cytol 1973, 36:137–169.
6. Trench RK: Of “leaves that crawl”: functional chloroplasts in animal cells.Sym Soc Exp Biol 1975, 29:229–265.
7. Rumpho ME, Summer EJ, Manhart JR: Solar-powered sea slugs.Mollusc/algal chloroplast symbiosis. Plant Physiol 2000, 123:29–38.
8. Wägele H, Johnson G: Observations on the histology and photosyntheticperformance of “solar-powered” opisthobranchs (Mollusca, Gastropoda,Opisthobranchia) containing symbiotic chloroplasts or zooxanthellae.Org Divers Evol 2001, 1:193–210.
9. Lee JJ: Algal symbiosis in larger Foraminifera. Symbiosis 2006, 42:63–75.10. McManus GB: Chloroplast symbiosis in a marine ciliate: ecophysiology
and the risks and rewards of hosting foreign organelles. Front Microbiol2012, 3:1–9.
11. Gast RJ, Moran DM, Dennett MR, Caron DA: Kleptoplasty in an Antarcticdinoflagellate: caught in evolutionary transition? Environ Microbiol 2007,9:39–45.
12. Rumpho ME, Pelletreau KN, Moustafa A, Bhattacharya D: The making of aphotosynthetic animal. J Exp Biol 2011, 214:303–311.
13. Wägele H, Martin WF: Endosymbioses in sacoglossan seaslugs:plastid-bearing animals that keep photosynthetic organelles withoutborrowing genes. In Endosymbiosis. Edited by Löffelhardt W. Vienna:Springer Vienna; 2013:291–324.
14. Pierce S, Biron R, Rumpho M: Endosymbiotic chloroplasts in molluscancells contain proteins synthesized after plastid capture. J Exp Biol 1996,199:2323–2330.
15. Gould AA: Report on the Invertebrata of Massachusetts. 2nd edition. Boston:Wright and Potter, State Printers; 1870.
16. Schwartz JA, Curtis NE, Pierce SK: Using algal transcriptome sequences toidentify transferred genes in the sea slug, Elysia chlorotica. Evol Biol 2010,37:29–37.
17. Rumpho ME, Worful JM, Lee J, Kannan K, Tyler MS, Bhattacharya D,Moustafa A, Manhart JR: Horizontal gene transfer of the algalnuclear gene psbO to the photosynthetic sea slug Elysia chlorotica.Proc Natl Acad Sci USA 2008, 105:17867–17871.
18. Wägele H, Deusch O, Händeler K, Martin R, Schmitt V, Christa G, Pinzger B,Gould SB, Dagan T, Klussmann-Kolb A, Martin W: Transcriptomic evidencethat longevity of acquired plastids in the photosynthetic slugs Elysiatimida and Plakobranchus ocellatus does not entail lateral transfer ofalgal nuclear genes. Mol Biol Evol 2011, 28:699–706.
19. Risso A: Mémoire sur quelques gastéropodes nouveaux, nudibrancheset tectibranches observés dans la Mer de Nice. J Phys Chim Hist Nat Arts1818, 87:368–377.
20. Hasselt JCV: Extrait d’une lettre du Dr JC van Hasselt au Prof. vanSwinderen, sur mollusques de Java. Bull Sci Nat Géol 1824, 2:237–245.
21. Pelletreau KN, Bhattacharya D, Price DC, Worful JM, Moustafa A, Rumpho ME:Sea slug kleptoplasty and plastid maintenance in a metazoan. Plant Physiol2011, 155:1561–1565.
22. Bhattacharya D, Pelletreau KN, Price DC, Sarver KE, Rumpho ME: Genomeanalysis of Elysia chlorotica egg DNA provides no evidence for horizontalgene transfer into the germ line of this kleptoplastic mollusc. Mol BiolEvol 2013, 8:1843–1852.
23. Pierce SK, Fang X, Schwartz JA, Jiang X, Zhao W, Curtis NE, Kocot KM, Yang B,Wang J: Transcriptomic evidence for the expression of horizontallytransferred algal nuclear genes in the photosynthetic sea slug, Elysiachlorotica. Mol Biol Evol 2012, 29:1545–1556.
24. Christa G, Zimorski V, Woehle C, Tielens AGM, Wägele H, Martin WF, Gould SB:Plastid-bearing sea slugs fix CO2 in the light but do not requirephotosynthesis to survive. Proc R Soc B 2014, 281:20132493.
25. Christa G, Wescott L, Schäberle TF, König GM, Wägele H: What remainsafter 2 months of starvation? Analysis of sequestered algae in aphotosynthetic slug, Plakobranchus ocellatus (Sacoglossa,Opisthobranchia), by barcoding. Planta 2013, 237:559–572.
26. Decaisne J: Plantes de l’Arabie Heureuse, receuillies par MP-E. Botta.Archs Mus. natn. Hist nat Paris 1842, 2:89–193.
27. Taylor DL: Chloroplasts as symbiotic organelles in the digestive gland ofElysia viridis (Gastropoda, Opisthobranchia). J Mar Biol Ass 1968, 48:1–15.
28. Jensen KR: Morphological adaptations and plasticity of radular teeth ofthe Sacoglossa (= Ascoglossa) (Mollusca: Opisthobranchia) in relation totheir food plants. Biol J Linn Soc 1993, 48:135–155.
29. Händeler K, Wägele H: Preliminary study on molecular phylogeny ofSacoglossa and a compilation of their food organisms. Bonn Zool Beitr2007, 55:231–254.
30. Händeler K, Wägele H, Wahrmund U, Rüdinger M, Knoop V: Slugs’ lastmeals: molecular identification of sequestered chloroplasts fromdifferent algal origins in Sacoglossa (Opisthobranchia, Gastropoda).Mol Ecol Resour 2010, 10:968–978.
31. Jensen KR: A review of sacoglossan diets, with comparative notes onradular and buccal anatomy. Malacol Rev 1980, 13:55–77.
32. Thompson TE, Jarman GM: Nutrition of Tridachia crispata (Mörch)(Sacoglossa). J Moll Stud 1989, 55:239–244.
33. Williams SI, Walker DI: Mesoherbivore–macroalgal interactions: feedingecology of sacoglossan sea slugs (Mollusca, Opisthobranchia) and theireffects on their food algae. Oceanogr Mar Biol Annu Rev 1999, 37:87–138.
34. Curtis NE, Massey SE, Schwartz JA, Maugel TK, Pierce SK: The intracellular,functional chloroplasts in adult sea slugs (Elysia crispata) come fromseveral algal species, and are also different from those in juvenile slugs.Microsc Microanal 2005, 11:1195–1196.
35. Curtis NE, Massey SE, Pierce SK: The symbiotic chloroplasts in thesacoglossan Elysia clarki are from several algal species. Invertebr Biol 2006,125:336–345.
36. Pierce SK, Curtis NE, Massey SE, Bass AL, Karl SA, Finney CM: Amorphological and molecular comparison between Elysia crispata and anew species of kleptoplastic sacoglossan sea slug (Gastropoda:Opisthobranchia) from the Florida Keys, USA. Mollusc Res 2006, 26:23–38.
37. Maeda T, Hirose E, Chikaraishi Y, Kawato M, Takishita K, Yoshida T,Verbruggen H, Tanaka J, Shimamura S, Takaki Y, Tsuchiya M, Iwai K,Maruyama T: Algivore or phototroph? Plakobranchus ocellatus(Gastropoda) continuously acquires kleptoplasts and nutrition frommultiple algal species in nature. PLoS ONE 2012, 7:e42024.
38. Taylor DL: Photosynthesis of symbiotic chloroplasts in Tridachia crispata(Bergh). Comp Biochem Physiol 1971, 38:233.
39. Vogel RM: The biology and a redescription of the opisthobranch molluskHermaea cruciata Gould, from Chesapeake Bay (Maryland). Veliger 1971,14:155–157.
40. Marcus E: Opisthobranchia from Brazil. Bol Fac Filos, Cienc Letr 1955, 20:89–261.41. Krug PJ: Bet-hedging dispersal strategy of a specialist marine herbivore:
a settlement dimorphism among sibling larvae of Alderia modesta.Mar Ecol Prog Ser 2001, 213:177–192.
42. Jensen KR: Observations on feeding methods in some FloridaAscoglossans. J Moll Stud 1981, 47:190–199.
43. Kawaguti S, Yamasu T: Electron microscopy on the symbiosis between anelysioid gastropod and chloroplasts of a green alga. Biol J Okayama Univ1965, 11:57–65.

Christa et al. Frontiers in Zoology 2014, 11:15 Page 12 of 12http://www.frontiersinzoology.com/content/11/1/15
44. Hirose E: Digestive system of the sacoglossan Plakobranchus ocellatus(Gastopoda: Opishtobranchia): Light and electron-microscopicobservations with remarks on chloroplast retention. Zool Sci 2005,22:905–916.
45. Baba K: Opisthobranchia of Kii, Middle Japan. J Dep Agriculture, Kyushu ImpUniv 1938, 6:1–19.
46. Bergh L: Malacologische Untersuchungen. In Reisen im Archipel derPhilippinen. Edited by Semper CG. Wissenschaftliche Resultate; 1870. 1:1–30.
47. Christa G, Gould SB, Franken J, Vleugels M, Karmeinski D, Händeler K, MartinWF, Wägele H: Functional kleptoplasty in a limapontioid genus:Phylogeny, food preferences and photosynthesis in Costasiella withfocus on C. ocellifera (Gastropoda, Sacoglossa). J Mollusc Stud (in press)
48. Simroth H: Opisthobranchia Gymnobranchia. Die Gastropoden der PlanktonExpedition 1895, 2:1–206.
49. Clark KB, JK R, Hugh SM, Fermin C: Chloroplast symbiosis in anon-elysiid mollusc, Costasiella lilianae Marcus (Hermaeidae: Ascoglossa(= Sacoglossa): Effects of temperature, light intensity, and starvation oncarbon fixation rate. Biol Bull 1981, 160:43–54.
50. Krug PJ, Händeler K, Vendetti J: Genes, morphology, development andphotosynthetic ability support the resurrection of Elysia cornigera(Heterobranchia:Plakobranchoidea) as distinct from the “solar-powered”sea slug, E. timida. Invertebr Syst 2011, 25:477.
51. Silva PC: A review of nomenclatural conservation in the algae fromthe point of view of the type method. Univ Calif Publ Botany 1952,25:241–323.
52. Wright SW, Grant BR: Properties of chloroplasts isolated from siphonousalgae: effects of osmotic shock and detergent treatment on intactness.Plant Physiol 1978, 61:768–771.
53. Trench RK, Boyle JE, Smith DC: The association between chloroplasts ofCodium fragile and the mollusc Elysia viridis. I. Characteristics of isolatedCodium chloroplasts. Proc R Soc B 1973, 184:51–61.
54. de Vries J, Habicht J, Woehle C, Changjie H, Christa G, Wägele H, Nickelsen J,Martin WF, Gould SB: Is ftsH the key to plastid longevity in sacoglossanslugs? Genome Biol Evol 2013, 5:2540–2548.
55. Ventura P, Calado G, Jesus B: Photosynthetic efficiency and kleptoplastpigment diversity in the sea slug Thuridilla hopei (Vérany, 1853).J Exp Mar Biol Ecol 2013, 441(C):105–109.
56. Pillet L, Pawlowski J: Transcriptome analysis of foraminiferan Elphidiummargaritaceum questions the role of gene transfer in kleptoplastidy.Mol Biol Evol 2013, 30:66–69.
57. Wisecaver JH, Hackett JD: Transcriptome analysis reveals nuclear-encodedproteins for the maintenance of temporary plastids in the dinoflagellateDinophysis acuminata. BMC Genomics 2010, 11:366.
58. Händeler K: Evolution of Sacoglossa (Opisthobranchia) with Emphasis on theirFood (Ulvophyceae) and the Ability to Incorporate Kleptoplasts, Doctoralthesis. University Bonn; 2011:1–134.
59. Evertsen J, Johnsen G: In vivo and in vitro differences in chloroplastfunctionality in the two north Atlantic sacoglossans (Gastropoda,Opisthobranchia) Placida dendritica and Elysia viridis. Mar Biol 2009,156:847–859.
60. Alder J, Hancock A: A Monograph of the British Nudibranchiate Mollusca withFigures of all the Species. London: The Ray Society; 1845.
61. Trench RK, Gooday GW: Incorporation of [H-3]-leucine into protein byanimal tissues and by endosymbiotic chloroplasts in Elysia viridisMontagu. Comp Biochem Physiol 1973, 44:321.
62. Gallop A: Evidence for the presence of a “factor” in Elysia viridis whichstimulates photosynthate release from its symbiotic chloroplasts.New Phytol 1974, 73:1111–1117.
63. Maeda T, Kajita T, Maruyama T, Hirano Y: Molecular phylogeny of thesacoglossa, with a discussion of gain and loss of kleptoplasty in theevolution of the group. Biol Bull 2010, 219:17–26.
64. Jörger KM, Stöger I, Kano Y, Fukuda H, Knebelsberger T, Schrödl M: On theorigin of Acochlidia and other enigmatic euthyneuran gastropods, withimplications for the systematics of Heterobranchia. BMC Evol Biol 2010,10:323.
65. Neusser TP, Fukuda H, Jörger KM, Kano Y, Schrödl M: Sacoglossa orAcochlidia? 3D reconstruction, molecular phylogeny and evolution ofAitengidae (Gastropoda: Heterobranchia). J Moll Stud 2011, 77:332–350.
66. Montagu A: An account of some new and rare British shells and animals.Transac Linn Soc Lond 1815, 11:179–204.
67. Thompson TE: Eastern Mediterranean Opisthobranchia: oxynoidae,Polybranchiidae, Stiligeridae (Sacoglossa). J Moll Stud 1988, 54:157–172.
68. Clark KB, Jensen KR, Stirts HM: Survey for functional kleptoplasty amongwest Atlantic Ascoglossa (= Sacoglossa) (Mollusca: Opisthobranchia).Veliger 1990, 33:339–345.
69. Marìn A, Ros J: Ultrastructural and ecological aspects of the developmentof chloroplast retention in the sacoglossan gastropod Elysia timida.J Moll Stud 1993, 59:95–104.
70. Saunders GW: Applying DNA barcoding to red macroalgae: a preliminaryappraisal holds promise for future applications. Philos T R Soc B 2005,360:1879–1888.
71. Hanyuda T, Arai S, Ueda K: Variability in the rbcL introns of Caulerpaleanalgae (Chlorophyta, Ulvophyceae). J Plant Res 2000, 113:403–413.
72. Verbruggen H, Ashworth M, LoDuca ST, Vlaeminck C, Cocquyt E, Sauvage T,Zechman FW, Littler DS, Littler MM, Leliaert F: A multi-locus time-calibratedphylogeny of the siphonous green algae. Mol Phylogenet Evol 2009,50:642–653.
73. Gosliner TM: The genus Thuridilla (Opisthobranchia: Elysiidae) from thetropical Indo-Pacific, with a revision of the phylogeny of the Elysiidae.In Proceedings of the California Academy of Sciences. 1995, 49:1-54.
74. Gosliner TM, Behrens DW, Valdés Á: Indo-Pacific Nudibranchs and Sea Slugs: aField Guide to the World’s Most Diverse Fauna. San Francisco: California Academyof Sciences/Sea Challengers Natural History Books; 2008:1–426.
75. Famà P, Wysor B, Kooistra WH, Zuccarello GC: Molecular phylogeny of thegenus Caulerpa (Caulerpales, Chlorophyta) inferred from chloroplast tufAgene. J Phycol 2002, 38:1040–1050.
76. Katoh K, Misawa K, Kuma K-I, Miyata T: MAFFT: a novel method forrapid multiple sequence alignment based on fast Fourier transform.Nucleic Acids Res 2002, 30:3059–3066.
77. Guindon S, Gascuel O: A simple, fast, and accurate algorithm to estimatelarge phylogenies by maximum likelihood. Syst Biol 2003, 52:696–704.
78. Tanabe AS, Toju H: Two New computational methods for universal DNAbarcoding: a benchmark using barcode sequences of bacteria, archaea,animals, fungi, and land plants. PLoS ONE 2013, 8:e76910.
79. Little DP: DNA barcode sequence identification incorporating taxonomichierarchy and within taxon variability. PLoS ONE 2011, 6:e20552.
80. Yamamoto YY, Yusa Y, Yamamoto S, Hirano Y, Hirano Y, Motomura T,Tanemura T, Obokata J: Identification of photosynthetic sacoglossansfrom Japan. Endocyt Cell Res 2009, 19:112–119.
81. Klochkova TA, Han J-W, Kim J-H, Kim K-Y, Kim G-H: Feeding specificity andphotosynthetic activity of Korean sacoglossan mollusks. Algae 2010,25:217–227.
82. Klochkova TA, Han J-W, Chah K-H, Kim RW, Kim J-H, Kim K-Y, Kim G-H:Morphology, molecular phylogeny and photosynthetic activity of thesacoglossan mollusc, Elysia nigrocapitata, from Korea. Mar Biol 2012,160:155–168.
83. Wägele H, Stemmer K, Burghardt I, Haendeler K: Two new sacoglossan seaslug species (Opisthobranchia, Gastropoda): Ercolania annelyleorum sp.nov. (Limapontioidea) and Elysia asbecki sp. nov. (Plakobranchoidea),with notes on anatomy, histology and biology. Zootaxa 2010, 2676:1–28.
84. Evertsen J, Burghardt I, Johnsen G, Wägele H: Retention of functionalchloroplasts in some sacoglossans from the Indo-Pacific and Mediterranean.Mar Biol 2007, 151:2159–2166.
85. Costa A: Illustrazione di due generi di Molluschi Nudibranchi. Rendi AccadSci Fis Mat Napoli 1867, 6:136–137.
86. Thompson TE: Jamaican opisthobranch molluscs I. J Moll Stud 1977,43:93–140.
87. Schmitt V, Händeler K, Gunkel S, Escande ML, Menzel D, Gould SB, Martin WF,Wägele H: Chloroplast incorporation and long-term photosynthetic per-formance through the life cycle in laboratory cultures of Elysia timida(Sacoglossa, Heterobranchia). Front Zool 2014, 11:5.
doi:10.1186/1742-9994-11-15Cite this article as: Christa et al.: Identification of sequestered chloroplasts inphotosynthetic and non-photosynthetic sacoglossan sea slugs (Mollusca,Gastropoda). Frontiers in Zoology 2014 11:15.





























