Embed Size (px)
Citation preview

REPÚBLICA BOLIVARIANA DE VENEZUELA UNIVERSIDAD NACIONAL EXPERIMENTAL DE GUAYANA
VICERRECTORADO ACADÉMICO COORDINACIÓN GENERAL DE INVESTIGACIÓN Y POSTGRADO COORDINACIÓN DEL POSTGRADO EN CIENCIAS AMBIENTALES
PROGRAMA DE MAESTRÍA EN CIENCIAS AMBIENTALES- MENCIÓN ECOLOGÍA APLICADA
Estado poblacional, distribución geoespacial, aspectos reproductivos y uso consuntivo de Paleosuchus trigonatus (Crocodylia: Alligatoridae) en la Cuenca
del Río Erebato, Alto Caura, Venezuela.
Trabajo de Grado presentado ante el Comité Académico del Postgrado en Ciencias Ambientales de la Universidad Nacional Experimental de Guayana, como requisito
parcial para optar al título de Magister Scientarum en Ciencias Ambientales- mención Ecología Aplicada.
Por: Lcdo. Carlos Valeris
Tutor: Hernán Castellanos Ph.D.
Co-tutor: MSc. Robinson Botero-Arias
Puerto Ordaz, Abril de 2016.


ii
DEDICATORIA
de la Cuenca del Río Caura

iii
AGRADECIMIENTOS
En primer lugar, quiero dar gracias a Dios todopoderoso por el aliento y
motivación que fueron muy necesarias para realizar y culminar este proyecto. Por la
oportunidad de recorrer los ríos, caños, bosques y conocer la fauna de la cuenca del
río Caura y por compartir tantos momentos especiales con su gente.
A la Universidad Nacional Experimental de Guayana (UNEG), la Coordinación
General de Investigación y Postgrado, la Coordinación del Postgrado en Ciencias
Ambientales, al Centro de Investigaciones Ecológicas de Guayana (CIEG) y al
Departamento Hombre y Ambiente, por todo el apoyo institucional recibido.
a las comunidades: Entreríos, Sayusodiña, Dakudakuña, Dadiña, Ayawaña, Yudiña,
Troncón, Madoña, Anadekeña y Jüwütüña por su receptividad y colaboración.
A la Organización de los Pueblos Indígenas de la Cuenca del Río Caura
KUYUJANI por el apoyo institucional y al Ministerio del Poder Popular para el
Ambiente por la autorización de caza con fines científicos expedida para el proyecto.
A Wildlife Conservation Society (WCS) por el préstamo de equipos y apoyo
logístico y financiero. Al Grupo de Especialistas en Cocodrilos de la UICN por el
apoyo financiero. Muy especialmente a la Fundación Rufford (Small Grant Rufford
Foundation) por el apoyo financiero asignado (Proyecto Nº 12964-1), que contribuyó
de forma muy significativa al desarrollo de las actividades en campo. Al Fondo
Nacional para la Ciencia y Tecnología (FONACYT) del Ministerio del Poder Popular
para la Ciencia y Tecnología por su apoyo financiero para la asistencia a cursos de
entrenamiento en Brasil relacionados con el proyecto, en 2011 y 2013.
Especial mención para los parabiólogos indígenas que participaron durante todo el
desarrollo del proyecto: Williams Sarmiento, Elso Espinoza y Orlando Pastai
Rodríguez, a quienes agradezco infinitamente todo el duro trabajo que hizo posible
que este proyecto se realizará. A los parabiólogos y monitores comunitarios: Narcio
Rodríguez , César Jokomosoma, Ramón Jasatao, Manuel Asatali, Jhonny Linarez,

iv
Jimmy Rodríguez Espinoza
Rodríguez, Kevin Apiama, Jesús García, Miguel Rodríguez, Jovani Castro, Romeo
Payena, Water Espinoza y Mario Asatali. A Dolinta Dominguez y Naileth Martínez
por su apoyo logístico durante las actividades de campo.
A Lucy Perera-Romero por su incondicional apoyo durante el desarrollo del
proyecto, como colega y amiga. A los biólogos: Igor Castillo y Luis Sibira por su
importante participación en la colección de datos, su compromiso y contribución al
proyecto. A Raquel Kassalian por su participación en el registro de la nidada. Al
Abog. Alberto Rodríguez, al Sr. Mayraleno Cortéz, al Sr. Cayetano y al Sr. Jesús
García por su ayuda, interés e intermediación con las comunidades de la cuenca del
río Erebato. Al equipo del Centro de Investigaciones Ecológicas de Guayana,
especialmente a Jesús Pérez, Solciret Perales, Gicela González, Judith Rosales, Luz
Delgado y a Marianela La Grave (WCS) por el apoyo administrativo y logístico.
A mi tutor, Dr. Hernán Castellanos, por su confianza, amistad y ayuda. Por
compartir conmigo su entusiasmo por el Caura y su gente. A mi co-tutor MSc.
Robinson Botero-Arias, MSc. Boris Marioni y Dr. Ronis Da Silveira por todas sus
enseñanzas, consejos, asesorías y amistad. A Guillermo Freire, Raiclicia Bizerril,
Jean Samonek y Diogo Araújo, por su apoyo y motivación.
A Carolina Bertsch por la oportunidad que me permitió conocer la cuenca del río
Caura Pio Colmenares y a la familia
Marioni- Bertsch por su amistad y apoyo durante mis estancias en Manaos.
Al
Sanoja, Dra. Nalúa Silva, Dra. Judith Rosales, Dr. Alexander Mansutti, MSc. Sara
Julia Leal, Dra. Aixa Vieira, Dra. Juana Figueroa y MSc. Aracélis Narayán por
contribuir con mi formación académica.
Al Cdte. Johnny Bompal y al Cuerpo de Bomberos de la Universidad Nacional
Experimental de Guayana, por su ayuda durante los momentos más difíciles que nos
tocaron vivir a Jesús Pérez y a mí durante este proyecto.
A mi familia y especialmente a mi esposa: Johanna Hernández por estar siempre,
por motivarme y creer en mí.

v
ÍNDICE DE CONTENIDO
pp. ÍNDICE DE TABLA vii
xii
. xiv
. 16 CAPÍTULO
18 18 20
20 21
Limitaci
23
23 Características del Género Paleosuchus 25 Distribución 26
27 28 29
Cacería de subsistencia y otros usos de la fauna silvestre en la cuenca del r 30 Teoría fuente- 32
33
33 38
53
54
Distribución geoespacial, posición en el hábitat y características del micro- 61

vi
Aspectos reproductiv 70 80
Recomendaciones para la conservación y manejo de Paleosuchus trigonatus . 89
4
94 95
97
105

vii
ÍNDICE DE TABLAS pp. Tabla 1. Características físico-químicas y estructurales de algunos caños y ríos de la Cuenca del Río Erebato donde se identificaron P. trigonatus 35 Tabla 2. Registros de consumo consultados y entrevistas realizadas por comunidad. Fuente: 1. Perera-Romero (datos no publicados); 2. Datos
propios; 3. Castellanos et al. (2010); 4. WCS-Programa Caura y Organización Kuyujani (datos no publicados)
Tabla 3. Conteos nocturnos en el caño Ajuju, con los valores de abundancia relativa para cada recorrido 55 Tabla 4. Conteos nocturnos en el río Madajano, con los valores de abundancia
relativa para cada recorrido Tabla 5. Esfuerzo de búsqueda para P. trigonatus en 8 localidades de la CRE. Los
valores para cada localidad representan el menor esfuerzo registrado en un recorrido entre los conteos nocturnos realizados, salvo aquellas localidades donde se efectuó un solo recorrido. Los valores son expresados en horas: 1= 60 minutos; 0,5= 30 minutos; 0,25= 15 minutos, necesarios para la observación de un individuo Tabla 6. Posición en el hábitat ocupada por P. trigonatus para cada categoría de
tamaño. N=66. SO: Sobre orilla; IAT: Interfase Agua-Tierra; EA: En el agua; SU: Sumergidos
Tabla 7. Detalle de la talla (largo y ancho), ancho de la banda y peso de los huevos
intactos 72 Tabla 8. Resumen de las características del nido y su micro-hábitat 73 Tabla 9. Características de los nidos registrados en la Cuenca del Río Erebato entre
2012 y 2013. Fuentes: 1 Sr. Sarmiento (Comunidad Anadekeña); 2 César Jokomosoma (Comunidad Yudiña); 3 Datos propios; 4 Carmelo Estaba (Comunidad Yudiña) 5 Sr. Fernancio (Comunidad Dadiña) 6
Tabla 10. Distribución temporal de los registros de nidos, hembras con huevos y
grupos de neonatos referidos para la Cuenca del Río Caura y Erebato. Las celdas sombreadas indican el periodo estimado de incubación de la nidada basado en Vásquez- Ruesta (1982) y Magnusson (1990). Abreviaturas relativas a los registros: END: Embriones no desarrollados; EVA:

viii
Embriones avanzados; BVI: Banda visible; HCH: Hembra con huevos en oviducto. *Datos propios 9
Tabla11. Registros de cacería de P. trigonatus en distintos sectores de la CRE.
Fuentes: 1. Castellanos et al. (2010); 2. WCS-Programa Caura/ Organización Kuyujani (datos no publicados); 3. Perera-Romero (datos
no publicados) 80 Tabla 12.Número de entrevistas y de P. trigonatus cazados en la CRE 81 Tabla 13. Número de P. trigonatus cazados por comunidad, localidad y año, según los entrevistados. Datos propios 82 Tabla 14. Relación de P. trigonatus cazados y la duración de las faenas de cacería en la CRK en los periodos 2004-2005 y 2008-2009. Fuentes: 1. Castellanos et al., (2010) y 2. WCS- Programa Caura 7 Tabla 15. Relación de P. trigonatus cazados y los días de registro de cacería en la
CRK en los periodos 2004-2005 y 2008-2009. Fuentes: 1. Castellanos et al., (2010) y 2. WCS- Programa Caura 7 Tabla 16. Resumen de las principales variables de cacería relacionadas con P.
trigonatus en comunidades de la CRE. Fuente: Perera-Romero (datos no publicados). Leyenda: 1 Especies de vertebrados; 2 Las especies cazadas fueron.3Números de P. trigonatus cazados días de registro de cacería;4
Con relación al número de individuos cazados;5 Número de P. trigonatus cazados por jornadas de cacería 8

ix
ÍNDICE DE FIGURAS
pp.
Figura 1. Cuenca del río Erebato y los sectores evaluados. Mapa elaborado por: Lucy Perera-Romero 35
Figura 2. Alto Erebato, a la altura de la comunidad Jüwütüña durante el periodo de aguas bajas (abril de 2009). Foto: Carolina Bertsch 36 Figura 3. Río Erebato, en las adyacencias de la comunidad Ayawaña, durante
periodo de aguas bajas. Foto: Carlos Valeris, 2013 36 Figura 4. Río Yudi. Foto: Carlos Valeris, 2013 37 Figura 5. Río Madajano con tepuy Jaua al fondo. Foto: Carlos Valeris, 2012 . 37 Figura 6.
2008 39 Figura 7. Valeris, 2009 39 Figura 8. Comunidad sanema Dadiña. Foto: Carlos Valeris, 2012 40 Figura 9. Vista parcial de la comunidad sanema de Ayawaña. Foto: Carlos Valeris, 2008 40 Figura 10.
comunal al fondo. Foto: Carlos Valeris, 2012 41 Figura 11. Bertsch, 2009 .. 41 Figura 12. Nomenclatura de las escamas caudales dobles y simples de P. trigonatus. Se señalan los códigos correspondientes a cada escama. Se omiten las escamas S6, S7, S8 y S9 por las características de la imagen. Foto: Lucy Perera-Romero, 2013. Diseño: Carlos Valeris 43 Figura 13. Paleosuchus trigonatus
2013 54 Figura 14. Estructura poblacional de P. trigonatus
Sólo se incluyen a los individuos estimados en tamaño 61

x
Figura 15. Registros de P. trigonatus en el sector Alto Erebato. Los puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios 62 Figura 16. Registros de P. trigonatus en el sector Erebato medio. Los Puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios 63 Figura 17. Registros de P. trigonatus en el sector Yudi. Los puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios 64 Figura 18. Registros de P. trigonatus rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios 65 Figura 19. Caño Ñayudi. Foto: Carlos Valeris, 2012 65 Figura 20. Registros de P. trigonatus en el sector Madajano. Los puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios 66 Figura 21. Caño Ajuju, desembocadura en el río Madajano. Foto: Carlos Valeris, 2009 67 Figura 22. Nido de P. trigonatus hallado en afluente del caño Washadi, mayo de 2013. Foto: Raquel Kassalian 71 Figura 23. Caño afluente del caño Washadi. En sus orillas se halló el nido de P. trigonatus referido. Foto: Raquel Kassalian, 2013 72 Figura 24. Huevos fértiles de P. trigonatus. Se observa cubierta de barro de termitas en algunos de los huevos. Foto: Raquel Kassalian, 2013 74 Figura 25. Distribución de los nidos registrados de P. trigonatus en la CRE. Las estrellas amarillas representan su ubicación relativa. La estrella en el sector Yudi corresponde a dos nidos en la misma localidad. Mapa elaborado por: Lucy Perera- Romero. Datos de los registros propios 75

xi
Figura 26. Sitios donde se registraron nidos de P. trigonatus: 1. Rama caño Madijisi (Sector Alto Caura); 2. Rama caño Wadishidi (Erebato medio); 3. Caño Sajme (Erebato medio); 4. Caño Wakalama (Erebato medio) y 5. Rama caño Sisipö (Yudi). Fotos: 1 y 3. Carlos Valeris; 2. Raquel Kassalian; 4 y 5. Igor Castillo 77 Figura 27. Distribución anual de las precipitaciones en dos localidades de la CRE: Jüwütüña y Entreríos. Fuente: Departamento de Gestión Ambiental CVG-Edelca 79 Figura 28. Estructura de tamaños de los P. trigonatus cazados en la Cuenca en 2008 y 2009. Se obtuvo con base a 13 individuos. Fuente de datos: WCS-Programa Caura (datos no publicados) 85 Figura 29. Productos de cacería ahumados con piezas de P. trigonatus, ©WCS, 2009 86

xii
UNIVERSIDAD NACIONAL EXPERIMENTAL DE GUAYANA COORDINACIÓN GENERAL DE INVESTIGACIÓN Y POSTGRADO COORDINACIÓN DEL POSTGRADO EN CIENCIAS AMBIENTALES MAESTRÍA EN CIENCIAS AMBIENTALES- MENCIÓN ECOLOGÍA
APLICADA
ESTADO POBLACIONAL, DISTRIBUCIÓN GEOESPACIAL, ASPECTOS REPRODUCTIVOS USO CONSUNTIVO DE PALEOSUCHUS
TRIGONATUS (CROCODYLIA: ALLIGATORIDAE) EN LA CUENCA DEL RÍO EREBATO, ALTO CAURA, VENEZUELA.
Autor: Carlos Valeris
Tutor: Hernán Castellanos Co-tutor: Robinson Botero-Arias Abril de 2016.
RESUMEN
Se evaluó el estado poblacional, la distribución geoespacial, el uso del hábitat, los eventos reproductivos y el uso consuntivo de P. trigonatus en tres sectores de la Cuenca del Río Erebato, Alto Caura. Se realizaron conteos nocturnos en curiaras con remo en caños y con motor en ríos, y recorridos diurnos para la búsqueda de nidos y rastros. Cada individuo observado o capturado fue registrado y su posición, georeferenciada. Se evaluaron los elementos más relevantes del micro-hábitat y la posición de los ejemplares al momento de la observación. Se registraron los eventos de cacería en 8 comunidades de la Cuenca del Río Erebato (CRE) a través de entrevistas y registros directos de consumo. La abundancia relativa en Caño Ajuju y Río Madajano se registró en 2,05 y 0,64 ind/Km lineal de cuerpo de agua respectivamente. La única especie identificada en los sectores evaluados fue P. trigonatus y fue avistada con mayor frecuencia en caños que en ríos. El 41,5% de los registros corresponden a individuos entre los 30 y 39,9 cm de Longitud Rostro-Cloaca posterior. Esta especie construye nidos de tipo montículo en áreas no inundables en bosques de tierra firme, cercanos a caños de aguas transparentes,

xiii
generalmente asociados a la base de un árbol con diámetros a la altura del pecho mayor a 70 cm y a termiteros u hormigueros activos y el periodo de nidificación abarca la época de lluvia y sequía en la CRE. En todas las comunidades ysanema se lleva a cabo la caza de P. trigonatus y el aprovechamiento de sus nidadas con motivo de subsistencia. En la comunidad Yudiña se acordaron las primeras medidas para la conservación y manejo de P. trigonatus, que incluyen no cazar a los neonatos ni aprovechar las nidadas. Se recomienda hacer seguimiento a las poblaciones de esta especie en los sectores más vulnerables de la CRE. Palabras clave: Paleosuchus trigonatus, estado poblacional, aspectos reproductivos, consumo, cuenca del río Erebato.

xiv
UNIVERSIDAD NACIONAL EXPERIMENTAL DE GUAYANA COORDINACIÓN GENERAL DE INVESTIGACIÓN Y POSTGRADO COORDINACIÓN DEL POSTGRADO EN CIENCIAS AMBIENTALES MAESTRÍA EN CIENCIAS AMBIENTALES- MENCIÓN ECOLOGÍA
APLICADA
POPULATION STATUS, ESPATIAL DISTRIBUTION, NESTING AND CONSUNTIVE USE OF PALEOSUCHUS TRIGONATUS (CROCODYLIA:
ALLIGATORIDAE) IN EREBATO RIVER BASIN, UPPER CAURA, VENEZUELA.
Autor: Carlos Valeris Tutor: Hernán Castellanos
Co-tutor: Robinson Botero-Arias April, 2016
ABSTRACT
The population status, spatial distribution, habitat use, reproductive events and the consumptive use of P. trigonatus in three sectors of Erebato River Basin (ERB), Upper Caura, were evaluated. Night count surveys were carried out in canoes along creeks and rivers, and diurnal trips in search of nests and tracks. Each individual observed or captured was registered and its position georeferenced. The most important elements of micro-habitat and its position at the time of observation were evaluated. Hunting events were recorded in 8 communities of ERB through interviews and direct consumption records. The relative abundance in caño Ajuju and Madajano River were 2,05 and 0,64 ind/linear Km of water body respectively. The only species identified in evaluated sectors was P. trigonatus and that species was spotted more often in creeks than in rivers. 41.5% of the records correspond to individuals between 30 and 39,9 cm LRCp. This species builds mound type nests in non-flooded areas in upland forests, near transparent water creeks, usually associated with the base of a tree with DAP greater than 70 cm and termite or ant assets. Nesting period covers the dry and wet season in the ERB. In all ye'kwana and sanema

xv
communities subsistence hunting of P. trigonatus and use of their clutches was recorded. In Yudiña community the first measures for the conservation and management of P. trigonatus which includes an agreement of not to hunt neonates neither collect the eggs. It is recommended to monitor the populations of this species in the most vulnerable sectors of the ERB.
Key words: Paleosuchus trigonatus, population status, reproductive aspects, consumption, Erebato River Basin.

INTRODUCCIÓN
En Venezuela, hay un escaso conocimiento científico sobre la biología, estado
poblacional y distribución de los cocodrilianos del género Paleosuchus (Seijas,
2007), al igual que en el resto de los países en donde están presentes (Magnusson y
Campos, 2010 a y b). Sin embargo, las comunidades indígenas de la Cuenca del Río
Erebato (CRE), en el sur de Venezuela, conocen aspectos muy detallados sobre la
vida de éstos cocodri
et al., 2015) y
dan a estas especies un uso consuntivo (Castellanos et al., 2010), lo que ha ocurrido y
ocurre en otras regiones donde estas especies habitan (Medem, 1958; Magnusson,
1990; Thorbjarnarson, 1992; Rivera, 2013).
En este sentido, los miembros de varias comunidades indígenas asentadas en la
cuenca media y alta del río Erebato han notado, en los últimos años, que las
poblaciones de Paleosuchus sp. han ido en franco descenso, debido a la dificultad que
supone encontrarlos en lugares donde antes eran considerados más abundantes
(Perera-Romero y Palma, 2012 a, b y c). Por ello y por el desconocimiento de muchos
aspectos de la ecología de este grupo, fue necesario evaluar el estado actual de sus
poblaciones y aspectos básicos de su ecología, para contribuir, en consecuencia, con
su conservación en la cuenca.
Este trabajo está estructurado en cinco capítulos: el Capítulo I incluye el
planteamiento del problema, las preguntas de investigación, las predicciones, los
objetivos, la justificación e importancia y las limitaciones de la investigación. El
Capítulo II contiene los fundamentos teóricos que dan soporte a la investigación,
haciendo énfasis en los antecedentes y los conceptos clave. El Capítulo III describe el
área de estudio y los métodos empleados para la obtención de los datos y análisis de
los mismos. El Capítulo VI muestra los resultados obtenidos y se discute su
significado, mientras que el Capítulo V expone las conclusiones y muestra las
recomendaciones derivadas de este estudio.

17
Los instrumentos empleados para la colección de datos e información sobre las
localidades evaluadas se incluyen dentro de los apéndices. La lista de referencias
contiene sólo las publicaciones citadas en el documento.

18
CAPÍTULO I
EL PROBLEMA
Planteamiento del problema
Los cocodrilianos constituyen especies clave en todos los hábitats que ocupan,
ejerciendo la función de depredadores tope (Ross, 1998), lo que favorece el
mantenimiento de las dinámicas poblacionales de las presas que forman parte de su
dieta. Además, su presencia es vital en algunos sistemas acuáticos, porque
contribuyen con la movilización de nutrientes y en el caso de los grandes
cocodrilianos, con la formación y mantenimiento de pozos dentro de cuerpos de agua
permanentes y corrientes intermitentes (Rueda-Almonacid et al., 2007). Por otra
parte, son fuente de alimento para las comunidades indígenas y criollas,
especialmente en áreas rurales, y forman parte del comercio local e internacional,
legal e ilegal de fauna silvestre para: consumo de carne, huevos, pieles y como
mascotas (Magnusson y Campos, 2010 a y b).
En Venezuela, cinco especies de cocodrilianos están presentes: Crocodylus
intermedius (el Caimán del Orinoco), Crocodylus acutus (el Caimán de la costa),
Caiman crocodilus (la Baba), Paleosuchus palpebrosus (el Babo morichalero) y
Paleosuchus trigonatus (el Babo negro) (Seijas, 2011). No obstante, su situación
poblacional difiere entre especies: C. intermedius está en peligro crítico de extinción;
C. acutus, en peligro de extinción (Rodríguez y Rojas-Suárez, 2008), Caiman
crocodilus (ambas subespecies: C. c. crocodilus Baba y C. c. fuscus- Babilla) es
considerada estable y algunas poblaciones están sujetas a manejo (Velasco, 2005).

19
Con respecto a Paleosuchus, no hay datos suficientes que permitan evidenciar el
estado de sus poblaciones, a pesar de su utilización como fuente de alimento por
algunas comunidades indígenas y criollas y de su amplio ámbito de distribución
geográfica (Thorbjarnarson, 1992; Seijas, 2011).
Con relación a las especies del género Paleosuchus, la ausencia de informaciones
atinentes al estado poblacional, las características del hábitat, la dieta y reproducción,
entre otros, podrían afectar la definición de estrategias efectivas para su conservación
y manejo a nivel local y regional o propiciar la implementación de estrategias poco
claras (Thorbjarnarson, 1992). Actualmente, ambas especies de Paleosuchus están
incluidas a nivel internacional en el Apéndice II de la Convención Internacional para
el Tráfico y Comercio de Fauna Silvestre (CITES) y consideradas en el estatus de no
amenazadas, dentro de la categoría Preocupación menor (Least Concern- LC) dentro
de las listas rojas de la Unión Internacional para la Conservación de la Naturaleza
(UICN). A pesar de tal consideración, la UICN reconoce la ausencia de datos actuales
sobre su estado poblacional (Rueda-Almonacid et al., 2007; Magnusson y Campos,
2010 a y b). En Venezuela, ambas especies están incluidas en el Libro Rojo de la
Fauna dentro de la categoría «Datos insuficientes» (DD) (Rodríguez y Rojas-Suárez,
2008 y 2015), lo que sugiere la ausencia de información sobre su estado poblacional y
de distribución.
No obstante, miembros de las comunidades indígenas que pueblan el Alto Caura,
al sur de Venezuela, han notado con preocupación la disminución del recurso
«yadiwe», las especies Paleosuchus en lengua ye kwana o «iatamö», como se conoce
en lengua sanema. Al respecto, Castellanos et al. (2010) refieren la cosecha y el uso
consuntivo de P. palpebrosus l río
anema, aunque registros comprobados de
caza y de observaciones más recientes de P. trigonatus en la misma localidad (WCS-
Programa Caura, 2009), sugieren que la especie cazada, reportada por Castellanos et
al., (2010) pudiera ser en realidad P. trigonatus, ambas especies en muchas zonas de
su distribución suelen ser simpátricas o simpátridas -término acuñado por Reig
(1983), como por ejemplo en Brasil en la estación ecológica de Maraca y en algunos

20
lugares de la cuenca del río Branco, próximos a Boa Vista en el estado de Roraima
(Muniz et al. 2015). No obstante, el estado poblacional de los cocodrilianos aún
permanece desconocido en esa cuenca y en cuencas adyacentes, lo que hace
imposible evaluar la sostenibilidad de su aprovechamiento.
La importancia de este recurso para las comunidades indígenas del Alto Caura,
sumado a la ausencia de información ecológica referida al estado poblacional,
distribución, hábitat y reproducción de ambas especies en la Guayana venezolana,
hace necesaria la evaluación de sus poblaciones bajo presión de cacería, a fin de
contribuir con la instrumentación de un plan de manejo que permita utilizar el recurso
de manera sostenible para el beneficio de las comunidades, del ecosistema y de estos
cocodrilianos.
Preguntas de investigación
Las preguntas de investigación que orientaron esta propuesta científica fueron:
1. ¿Será la población de P. trigonatus más abundante que P. palpebrosus en
la cuenca media del río Erebato?
2. ¿Será P. trigonatus más abundante en las cabeceras de los ríos y caños que
en el canal principal del río y desembocadura de los caños?
3. ¿Estarán los eventos reproductivos de P. trigonatus restringidos a las áreas
no inundables de caños y ríos?
Objetivos
Objetivo General
Evaluar el estado poblacional, la distribución geoespacial, el uso del hábitat, los
eventos reproductivos y el aprovechamiento de los cocodrilianos del género
Paleosuchus, en la cuenca del río Erebato y proponer medidas para su conservación y
manejo.

21
Objetivos específicos
1. Determinar el estado poblacional de P. trigonatus.
2. Establecer la distribución geoespacial y el uso del hábitat para la especie
en cinco sectores de la cuenca del río Erebato: Alto Erebato, Erebato
medio, cuenca del río Yudi, Bajo cuenca del río Madajano.
3. Caracterizar los nidos y eventos reproductivos.
4. Analizar los eventos de cacería y el aprovechamiento de nidadas en las
comunidades indígenas y sanema de la cuenca del río Erebato.
5. Proponer medidas para la conservación y el manejo de P. trigonatus en los
sectores de la cuenca de río Erebato evaluados.
Justificación e importancia de la investigación
Este proyecto de investigación surgió para dar respuesta a una inquietud de los
pobladores de las comunidades indígenas Ayawaña, Anadekeña y Jüwütüña, en la
cuenca del río Erebato, quienes han percibido el descenso de las poblaciones de
cocodrilianos (Paleosuchus sp.) en varias localidades donde practican la cacería de
subsistencia. Este hecho fue evidenciado en los talleres comunitarios propiciados por
el Programa Caura de la Sociedad para la Conservación de la Vida Silvestre (WCS-
por sus siglas en inglés) y la Organización Indígena del Caura KUYUJANI (Perera-
Romero, 2012).
Esta preocupación, por parte de la comunidad, propició el desarrollo de la
investigación para conocer aspectos de las poblaciones y de la biología de las
especies de cocodrilianos del género Paleosuchus, como un prerrequisito
fundamental para el desarrollo de propuestas de conservación y manejo. Los
resultados estarán disponibles para las comunidades indígenas, quienes podrán

22
utilizarlo como insumo para la construcción colectiva de un plan de manejo y
conservación que contribuya con el mantenimiento de las poblaciones de esta especie.
Por otra parte, al ser este grupo de cocodrilianos uno de los menos estudiados en el
Neotrópico, los aportes de esta investigación contribuirán con información ecológica
importante, con especial énfasis en la ecología reproductiva y aprovechamiento, lo
que incrementará el conocimiento disponible para la especie, sobre todo para
Venezuela y la Guayana venezolana, dónde las últimas investigaciones que aportan
datos ecológicos sobre P. trigonatus, datan de 1986 (Gorzula y Paolillo, 1986; Seijas,
2007).
Limitaciones
El desarrollo de este trabajo de investigación presentó limitaciones, debido a lo
extenso del área de estudio, lo remoto de las localidades evaluadas, las condiciones
ambientales y las características propias del bosque tropical. Muchos de los cuerpos
de agua sólo pudieron ser evaluados parcialmente debido a la limitada navegabilidad
de éstos por el bajo nivel de las aguas, la presencia de raudales y de árboles caídos en
su curso.
Lo remoto del área de estudio implicó un importante esfuerzo logístico, que
sumado a la problemática existente en la cuenca del río Caura, como el aumento de la
actividad minera ilegal en el río Yuduani, la limitada oferta de combustible en Maripa
y los altos costos para su traslado entre El Playón y Kuyuwi, incrementó
significativamente los costos operativos y exigió aumentar las medidas de seguridad
para proteger la integridad de las personas y equipos durante las realización de las
actividades del proyecto. Muchas de estas limitaciones fueron superadas gracias al
apoyo de las organizaciones que financiaron el proyecto y al respaldo de las
comunidades indígenas de la cuenca, quienes fueron muy receptivas y apoyaron en
aspectos logísticos y técnicos durante las actividades de campo.

23
CAPÍTULO II
MARCO REFERENCIAL
Antecedentes
La mayor parte de las investigaciones sobre la ecología de P. trigonatus y su
congénere P. palpebrosus se han realizado en Brasil, en la cuenca del río Amazonas.
Una de las más relevantes fue realizada por Magnusson, Lima y Sampaio (1985),
quienes estudiaron la ecología reproductiva de P.trigonatus y evaluaron las fuentes de
calor en sus nidos. Los autores señalan que las colonias de termitas asociadas con los
nidos de P. trigonatus son una fuente de calor importante durante la incubación de la
nidada. Magnusson y Lima, en 1991, publicaron uno de los trabajos más completos
sobre la ecología de la misma especie en la amazonia brasilera. Señalan que los
adultos son sedentarios y territoriales y estimaron la edad reproductiva de las hembras
en 11 años y en 20 años para los machos.
Botero-Arias (2007) evaluó los patrones de movimiento, micro-hábitat y dieta en
una población de P. palpebrosus en el Amazonas central, concluyendo que es una
especie generalista, basado en la dieta y el uso del hábitat y con tendencia a migrar
localmente en las áreas inundadas. Más recientemente Villamarín et al. (2010)
estudiaron los movimientos, abundancia y crecimiento de una población de P.
trigonatus en un fragmento de bosque en la ciudad de Manaos, utilizando el método
de captura-recaptura y Martínez-Freire (2016) comparó los movimientos de P.
trigonatus entre ejemplares residentes y liberados en la misma área de estudio. Los
primeros autores refieren abundancias entre 2 y 9,5 ind/Km de cuerpo de agua, tasas
de re-captura de 14% a lo largo de 4 años de estudio y movimientos lineales de hasta

24
600 metros para un individuo joven. Por su parte, Martínez-Freire (2016) refiere
patrones de movimiento discretos para hembras de P. trigonatus, las cuales en dos de
los tres casos estudiados permanecieron en el mismo cuerpo de agua, moviéndose
aguas arriba y aguas abajo, coincidiendo con lo publicado por Magnusson y Lima
(1991) sobre el menor tamaño territorial de las hembras comparada con el tamaño del
territorio de los machos.
No obstante, pocos trabajos de investigación han sido desarrollados con estas
especies en Venezuela, debido probablemente por ocupar lugares prístinos, remotos y
poco poblados, por ser de pequeño tamaño y a su escaso o nulo valor comercial
(Seijas, 2007). Seijas (2007) publicó una valiosa contribución a la actualización de la
información morfológica y de distribución de los Paleosuchus catalogados en las
colecciones científicas nacionales, señalando una aparente diferencia en la
distribución entre P. trigonatus y P. palpebrosus. La primera es restringida a bosques
y con mayor amplitud en la franja altitudinal (300 a 1200 m.s.n.m.), mientras que P.
palpebrosus ocupa tierras bajas, con altitudes menores a los 450 m.s.n.m, incluyendo
sabanas, morichales y bosques, donde puede ocupar ríos de mayor orden con relación
a su cóngenere (op. cit.) Por otra parte, ofrece una guía que contribuye
substancialmente con minimizar los errores de identificación entre estas especies
recurrentes en la literatura.
Otros aportes sobre la distribución de este grupo de cocodrilos en el país fueron
realizados por Godshalk (1982), quien hizo referencia a nuevas localidades donde
registraron estas especies, entre las cuales destacó un cráneo de P. trigonatus en el río
Erebato, el cual es uno de los pocos registros dentro del área de estudio previo a esta
investigación y Gorzula y Paolillo (1986) quienes refieren datos de abundancia en
varios ríos de la guayana venezolana. Gorzula y Señaris (1998), resumen las
localidades donde han sido registradas ambas especies de Paleosuchus, destacándose
una localidad para P. trigonatus en la Sierra de Lema a 1340 m.s.n.m., que es la
mayor altitud en la que se haya observado a esta especie (Gorzula, Paolini y
Thorbjarnarson, 1987).

25
En la cuenca del río Paragua, Señaris, Lasso y Flores (2008) registraron la
presencia de P. trigonatus (Babo negro) en el cauce principal del río y en caños
tributarios en el área Bajo Paragua-Karún, donde también señalaron la presencia de
Caiman c. crocodilus en las lagunas de inundación. No obstante, estos registros de
Paleosuchus son imprecisos, debido a la aparición de P. palpebrosus en los apéndices
de especies coleccionadas e identificadas y al uso del mismo nombre común: Babo
negro, en la mencionada publicación. Otro registro impreciso de Paleosuchus fue
documentado por Barrios-Amorós y Brewer-Carias (2008), refiriendo el hallazgo de
un cráneo incompleto de un Paleosuchus sp. cazado en la comunidad Kanadakuni, en
Alto Caura. Otra localidad guayanesa donde se han identificado Paleosuchus fue
recientemente señalada por Barrio-Amorós, Brewer-Carias y Fuentes-Ramos (2011),
quienes refirieron la presencia de P. trigonatus en la sección occidental de Sierra de
Lema (Cerro Santa Rosa), otra localidad dentro de la Guayana venezolana.
Sobre la abundancia de este grupo en Venezuela, la información es igualmente
escasa. Los únicos datos disponibles para P. trigonatus fueron publicados por
Gorzula y Paolillo en 1986 (Seijas, 2007). Ellos incluyeron siete localidades en la
Guayana venezolana. Pacheco (2009) por su parte, evaluó a las poblaciones de P.
palpebrosus en tres ríos de los llanos orientales venezolanos. En ambos casos, fueron
considerados poco abundantes, con índices de abundancia entre 0,8 y 2 ind/Km de
cuerpo de agua. Sobre la densidad, Magnusson y Lima (1991) registraron altas
densidades de P. trigonatus en una localidad de la amazonia brasileña y estimaron su
biomasa entre 34,4 y 59,6 Kg/Km2, en bosques de tierra firme.
Características de las especies del género Paleosuchus
El género Paleosuchus está representado por dos especies: P. trigonatus y P.
palpebrosus, los cuales difieren del resto de los miembros de la Familia Alligatoridae
por sus características anatómicas externas y craneales (Medem, 1967; Magnusson,
1990). Una de ellas es la gran cantidad de osteodermos que poseen en su piel, que

26
cubren incluso la zona ventral, lo que les confiere una fuerte coraza protectora
(Magnusson, 1990; Rueda-Almonacid et al., 2007). Sin embargo, las características
más notables guardan relación con la anatomía craneal, en la que se observa la
ausencia de las fosas supratemporales (Medem, 1958).
Ambas especies se consideran las más pequeñas entre los cocodrilianos.
P. palpebrosus tiene menor talla con una longitud rostro-cloaca (LRC) máxima
señalada de 95cm para los individuos adultos machos (Campos y Magnusson, 2013),
mientras que P. trigonatus es un poco mayor (LRC 136cm) (Magnusson, 1990).
Seijas (2007) destacó que P. palpebrosus posee un cráneo más robusto y el hocico
más levantado que P. trigonatus; la variación numérica del patrón de las escamas
nucales, inguinales y ventrales, incluso a nivel intraespecífico. El mismo autor indicó
que P. trigonatus posee una coloración más oscura que P. palpebrosus y una banda
de color negro en la parte media del hocico, la cual es una característica diagnóstica.
Distribución geográfica
Ambas especies tienen extensiones de distribución solapada. P. trigonatus se
distribuye en las cuencas de los ríos Amazonas y Orinoco y en aquellos que drenan
hacia el océano Atlántico en las Guyanas. Por su parte, la distribución geográfica de
P. palpebrosus es más amplia, incluye las cuencas de los ríos Paraná y Paraguay, al
norte de Paraguay y suroeste de Brasil, y la porción norte de los llanos venezolanos.
Sin embargo y pese a compartir gran parte de sus ámbitos de distribución, pocas
veces se han observado compartiendo el mismo hábitat (Medem, 1967; Magnusson,
1990; Magnusson y Campos, 2010 a y b).
En Venezuela, Rivas et al. (2012), mencionan la presencia de P. trigonatus en dos
biorregiones: Guayana y Amazonia (en los estados Bolívar y Amazonas), mientras
que P. palpebrosus, ocupa además la biorregión de los Llanos y el Delta del Orinoco.
Con respecto a la distribución altitudinal, en la Guayana venezolana, Gorzula y

27
Paolillo (1986) registraron individuos de P. trigonatus a 1.300 m.s.n.m (Magnusson,
1990). En la cuenca del río Erebato, sólo Godshak (1982) refiere la presencia de P.
trigonatus por el hallazgo de un cráneo y Castellanos et al. (2010) de P. palpebrosus
en la cuenca del r (tributario del río Erebato). Este último dato, sin
embargo, es considerado P. trigonatus por la identificación de esta especie en años
recientes y la ausencia de registros, en ese mismo periodo, de P. palpebrosus en esa
cuenca.
Hábitat
La mayoría de los autores indican que ambas especies ocupan hábitats diferentes y
que raras veces son sintópicas; entiéndase por este término los lugares donde dos
especies simpátricas (especies que comparten sus ámbitos de distribución geográfica)
coexisten. Generalmente P. trigonatus está asociada con las corrientes de agua
menores (caños, quebradas y similares), dentro de los bosques de tierra firme,
mientras que P. palpebrosus es más adaptable; habita ríos, lagunas y caños en
bosques, sabanas, morichales, bosques de galería y áreas de várzea (Medem, 1958,
1967; Magnusson, 1990; Magnusson y Campos, 2010 a y b; Seijas, 2011).
Ese uso del espacio coincide con observaciones realizadas sobre otras especies de
cocodrilianos simpátricas, donde se observa una segregación de hábitat (Medem,
1981), lo que implica que la especie dominante ocupe el hábitat principal (e. g. canal
del río), mientras que la especie no dominante, las zonas marginales (e. g. tributarios
pequeños). En este sentido, cuando ambas especies coexisten, una de ellas ha
resultado ser más abundante que la otra, como lo reseña Medem (1967) en el río
Apaporis en Colombia, siendo P. palpebrosus más abundante en la cuenca baja del
río y P. trigonatus más abundante en la cuenca alta. Esto sugiere que el hábitat
óptimo de ambas especies puede diferir, aún en los mismos sistemas fluviales.
Pese a su importancia, el estudio ecológico de los cocodrilianos en simpatría no ha
recibido la atención necesaria en Venezuela (Seijas, 1988, 1996). Algunos autores

28
han indicado que, cuando existe simpatría, las especies ocupan un ambiente distinto o
que el grado de simpatría varía temporalmente (Medem, 1983; Webb, Manolis y
Sack, 1983). Al respecto, Pearcy (2011) evaluó las relaciones ecológicas y
segregación de nichos entre especies de cocodrilianos que coexisten, apoyando la
idea que la morfología craneal tenga relación directa con el hábitat, lo que se refleja
en el uso y segregación del nicho.
No obstante, esto puede no ser tan obvio con las especies de Paleosuchus, al ser
ambas de tamaño pequeño y morfología bucal similar, lo que no ocurre con la
mayoría de las especies de cocodrilianos simpátricas (e. g. Crocodylus. intermedius
con Caiman c. crocodilus y Crocodylus acutus con Caiman crocodilus fuscus). Por
otra parte, hay algunos elementos que distinguen a estas especies, como el
comportamiento críptico de P. trigonatus (Magnusson y Lima, 1991), que implica
grandes diferencias en la ocupación, uso de los hábitat y en la dieta, con respecto al
resto de las especies de cocodrilianos (Magnusson, 1990). Este último hecho, ha
representado una limitación importante para estudiar esta especie en su hábitat, lo que
obliga a establecer estrategias metodológicas diferentes a las aplicadas típicamente
para el estudio de los cocodrilianos en el Neotrópico (Magnusson y Lima, 1991).
Recomiendan el desarrollo de estudios a largo plazo (de 8 años o más), el muestreo
intensivo y periódico de las localidades evaluadas y el seguimiento de individuos
utilizando técnicas de marcado y radio-telemetría (op. cit).
Aspectos reproductivos
Medem (1967) refirió información sobre eventos reproductivos de Paleosuchus.
Los nidos de ambas especies son de tipo montículo aunque hay todavía muchos
vacíos de información, sobre todo para P. trigonatus (Rivas, Aktay y Owens, 2001).
Ambas especies construyen los nidos durante la época seca y los nacimientos
acontecen al inicio de la temporada lluviosa en la Amazonía central, que inicia en
diciembre y culmina en junio (Magnusson, 1990; Campos et al., 2015). Sobre los

29
nidos de P. palpebrosus, Campos et al. (2015) señala una diferencia entre las
temporadas de anidación a nivel geográfico, que incluye la temporada seca en la
Amazonía central y la temporada de lluvias en el Pantanal. La primera coincide con el
calurosa y lluviosa, pero los sitios de los nidos, en este caso, no estaban en las áreas
inundables.
El periodo de incubación es bastante largo en comparación con otras especies de
cocodrilianos, P. palpebrosus entre 90 y 114 días y P. trigonatus mayor a 100 días
(Medem, 1981; Magnusson, 1990; Rueda- Almonacid et al., 2007). El tamaño de la
nidada varía entre los 16 y 22 huevos, los cuales tienen en promedio 70 gramos de
peso (Rueda- Almonacid et al., 2007). Una vez nacidos, los neonatos tienden a
dispersarse rápidamente (White y Rivas, 2003) y el cuidado parental no está muy
desarrollado (Campos et al., 2012). Al igual que el resto de las especies de
cocodrilianos, la mortalidad de los neonatos y jóvenes es alta, mientras que los
adultos tienen pocos depredadores naturales (Magnusson y Lima, 1991), aunque
ambas especies son pequeñas y en el caso de P. trigonatus exhibe comportamiento
críptico.
Al ser territoriales, los individuos jóvenes se dispersan en búsqueda de nuevos
territorios. Éstos, en el caso de los P. trigonatus machos, pueden solaparse con
territorios menores de las hembras (Magnusson, 1990; Magnusson y Lima, 1991).
Este tipo de comportamiento es común en varias especies de cocodrilianos como una
estrategia que disminuye la competencia intraespecífica.
Comunidades y kwana y sanema de la cuenca del río Erebato
Los y y sanema son dos de los grupos indígenas que pueblan la cuenca del
río Erebato y también los más numerosos. Ambos grupos son cazadores- recolectores,
aunque también cultivan la yuca (elemento básico de su dieta) y algunas especies
frutales en las proximidades de sus comunidades (conucos) (Silva-Monterrey, 1997a

30
y b; Colchester, 1997). Cada grupo posee elementos culturales que condicionan su
relación con el entorno, aunque en las últimas décadas, el intercambio comercial y
cultural con los criollos incorporó elementos nuevos, como la cacería con armas de
fuego (escopetas) y la pesca con redes de nylon, lo que sumado a un cambio en la
dinámica de asentamiento mediada por la evangelización durante los años
que el patrón de uso de recursos también cambiara (op. cit.).
populosas como Jüwütuña y Anadekeña, esto ha ocasionado el agotamiento de los
recursos en ciertos lugares, sobre todo aquellos próximos a las comunidades (Perera-
Romero, 2012), lo que podría amenazar la sostenibilidad de la cacería de subsistencia
y la misma existencia de estas comunidades y sus modo de vida tradicionales.
Cacería de subsistencia y otros usos de la fauna silvestre en la cuenca del río
Erebato
Las comunidades indígenas y sanema asentadas en la CRE practican la
cacería de subsistencia, la pesca, la recolección y la agricultura de conuco (Silva-
Monterrey, 1997 a y b; Colchester, 1997). En el caso de la cuenca del r
(CRK), tributario del río Erebato, no hay asentamientos permanentes de comunidades
debido a las características físico-ambientales particulares, pero el área es
comúnmente utilizada por las comunidades de la cuenca del río Erebato como zona
de pesca y caza (zona multiuso) (Castellanos et al., 2010).
Entre los productos de caza, están los Paleosuchus, tanto los individuos adultos
como los jóvenes. No obstante, la identificación de los individuos resulta dudosa
debido a la forma de preparación de las presas (ahumadas) aunque los individuos
frescos fueron identificados como P. trigonatus (WCS-Programa Caura, 2009). Se
estima que entre abril de 2008 y julio de 2009 se capturaron en la CRK al menos 42
P. trigonatus; la mayoría de ellos capturados con lanzas en caños principales durante
las faenas de caza y pesca nocturnas. Una de las jornadas de cacería más intensivas

31
ocurrió en 2009 e incluyó a 9 individuos cazados, en una semana por un mismo grupo
de cazadores, con longitudes rostro-cloaca media de 40 cm (op. cit.). Este evento
incluyó un número mayor de especies de caza y pesca, y se registró como el mayor
evento en su tipo (en número de ejemplares cazados/pescados y por biomasa
estimada) para la CRK del que se tenga información. Sin embargo, no hay
información del estado de las poblaciones de este grupo en la zona, ni en áreas
circundantes, por lo que no era posible prever el efecto que esta extracción tendría
sobre la estructura de las poblaciones y su impacto a mediano y largo plazo.
Una de las consecuencias de la cacería sobre las poblaciones de cocodrilianos
guarda una estrecha relación con el comportamiento de los individuos, los cuales
tienden a ser más cautelosos (Verdade, 1996). Así mismo, la presión de caza puede
afectar la persistencia del cuidado parental, resultando en un descenso en su
proporción o interrupción del mismo por las hembras, lo que pudiere incidir en una
disminución del éxito reproductivo (op. cit.). Esto último puede ser no relevante para
las especies de Paleosuchus, debido a que no exhiben cuidado parental prolongado
(Magnusson, 1990; Campos et al., 2012). Sobre este particular, el aprovechamiento
de nidos por comunidades humanas en la CRE no había sido registrado
científicamente, ni tampoco en otra localidad del país.
Otro comportamiento que pudiere ser afectado por las actividades de caza es la
territorialidad. Al disminuir la densidad de los individuos y al afectar la estructura de
tamaños, el espacio disponible para los individuos dominantes supervivientes
aumentaría y, en consecuencia, ellos podrían ampliar sus territorios, favoreciendo una
interacción negativa con los individuos sub-adultos, que aparecerían tras la
recuperación de su población a causa de los eventos de caza prolongados
(Magnusson, 1986). Aunque es aún incierto que el comportamiento de Paleosuchus
sp. esté siendo afectado por la actividad de caza en la CRE, la percepción sobre las
poblaciones de Paleosuchus sp anema son claras:
los yadiwe- iatamö (Paleosuchus sp.) son uno de los animales más amenazados, junto
a los crácidos (paujíes y pavas), en las áreas de influencias de las comunidades del
Alto Caura (Perera-Romero, 2012).

32
Teoría Fuente-Sumidero La teoría fuente-sumidero propone que la dinámica poblacional de las especies
ocurre entre un hábitat fuente, donde la tasa de natalidad excede la mortalidad, y uno
sumidero, donde esta relación es inversa (Pulliam, 1988). En este caso, la persistencia
de la población en esos hábitat depende de la emigración de individuos desde los
hábitat fuente por lo que la dinámica poblacional de cada hábitat es balanceada por la
dispersión de los individuos (Dias, 1996).
Uno de los mecanismos por lo cual dicho proceso ocurre es a través de la
competencia intra-específica. En este caso, los hábitats de mejor calidad reproductiva
son ocupados por los individuos dominantes. Una vez que la densidad se expresa
como un factor limitante, los individuos subordinados se dispersan hacia los hábitats
periféricos, que funcionan como sumideros (Dias, 1996; Fryxell, 2001).
Este hecho podría estar ocurriendo con las poblaciones de Paleosuchus en la CRE,
al estar compuestas por individuos territoriales con preferencia por hábitats asociados
con los caños en bosques de tierra firme. Por otra parte, la actividad de caza asociada
con el canal principal del río y con los caños de mayor tamaño podría estar
s s individuos que
emigran de los hábitats fuente. Esto, asumiendo que los hábitats asociados con el
canal principal del río sean óptimos para la especie, aumentaría la probabilidad de
éxito para la dispersión de los individuos que buscan nuevos territorios (Fryxell,
2001).
Algunos autores consideran esta teoría para el manejo y conservación de
cocodrilianos (Da Silveira y Thorbjarnarson, 1999), por lo que es fundamental
conocer a profundidad los hábitats reproductivos de estas especies a fin de contribuir
con una protección oportuna y con un manejo adecuado (Dias, 1996). No obstante, la
protección y el manejo de las especies no deben basarse sólo en las áreas fuente
(Howe, Davis y Mosca, 1991 citado por Amarasekare, 2004).

33
CAPÍTULO III
MARCO METODOLÓGICO
Área de estudio
El área de estudio comprende la cuenca del río Erebato, uno de los tributarios
principales del río Caura. Administrativamente forma parte de los municipios Sucre y
Cedeño del estado Bolívar, Venezuela. El régimen hídrico incluye cuatro fases
hidrológicas: aguas bajas (enero a abril), subida de aguas (mayo a junio), aguas altas
(julio y agosto) y bajada de aguas (septiembre a diciembre). Sus principales
tributarios son los ríos Ka kada, Yudi y Chajura.
El to orden, de aguas negras, ácidas y poco turbias
(Echevarría, González y Mass, 2011), que nace en la meseta Jaua. Su cuenca posee
una extensión de 3.163Km2 (Perera-Romero, Polisar y Maffei, 2015). La época de
aguas bajas coincide con el período más seco del año (febrero- abril) y la dinámica
hidrológica está condicionada por el represamiento que el río Erebato ocasiona
(Castellanos et al., 2010). No hay comunidades humanas permanentes asentadas en
sus riberas y es utilizada por las comunidades ye kwana y sanema como un área de
caza, pesca y recolección (op. cit).
El río Yudi es de 4to orden, de aguas claras, nace en la Serranía de Maigualida
(Monumento Natural Serranía de Maigualida). En sus riberas, se asientan dos
comunidades sanema: Yudiña y Salasalaña. En su curso principal hay abundantes
raudales que dificultan la navegación y que representan obstáculos importantes
durante la época de subida de aguas.

34
Se establecieron 5 sectores dentro de la CRE (ver figura 1):
1) Alto Erebato: comprende la zona alta del río Erebato. En este sector están las
populosas de la cuenca. Se evaluaron los caños: Sajme, Jaku, Jadia, Masümü y
el canal principal del río (ver figura 2).
2) Erebato medio: esta sección incluye el tramo del río Erebato, entre la
comunidad sanema de Ayawaña y la comunidad Sayusodiña. En este
sector se evaluaron los caños Dadi, Washadi y Jadana/Akudu (ver figura 3).
3) Yudi: comprenden la zona media del río Yudi, el río Manasi (Kuyuwi), y la
comunidad sanema Yudiña (ver figura 4).
4)
desembocadura en el río Erebato y los caños: Suajadito, Widiki, Kajioko,
Dojüjano, Kaduaimudü.
5) Madajano: comprende la sub-
desde la base del Tepuy Jaua hasta la d
caño Ajuju, su tributario más importante (ver figura 5).
(Bajo Erebato), fue incluida sólo para la
evaluación de uso consuntivo de P. trigonatus a través de la aplicación de entrevistas,
pero no pertenece a ninguno de los sectores mencionados.
Los caños y tramos de los ríos evaluados fueron seleccionados con base a los
registros de caza y recolección de huevos, que los miembros de las comunidades
ye kwana y sanema suelen realizar periódicamente, con especial énfasis en los
registros recientes (2010-2013). En la tabla 1 se muestran las características físico-
químicas y estructurales de algunos ríos y caños del área de estudio donde se
identificaron P. trigonatus.

35
Figura 1.Cuenca del río Erebato y los sectores evaluados. Mapa elaborado por: Lucy Perera- Romero. Tabla 1. Características físico-químicas y estructurales de algunos caños y ríos de la CRE, donde se identificaron P. trigonatus. Datos propios.
Variable Caño Ajuju Río Madajano Río Yudi
Caño Manasi
Ancho promedio (m) 20 60 50 35 25
Profundidad canal medio (m)
2 3 5 4 3
Temperatura del agua (ºC)
25 22 25 27 26
Cobertura del dosel (%)
40 5 5 5 10
Velocidad de la corriente(m/seg)
3,5 2 1,5 1 2
pH 5 5,1 5,2 5,2 5,7

36
Figura 2. Alto Erebato, a la altura de la comunidad Jüwütüña durante el periodo de aguas bajas (abril de 2009). Foto: Carolina Bertsch.
Figura 3. Río Erebato, en las adyacencias de la comunidad Ayawaña, durante periodo de aguas bajas. Foto: Carlos Valeris, 2013.

37
Figura 4. Río Yudi. Foto: Carlos Valeris, 2013.
Figura 5. Río Madajano con tepuy Jaua al fondo. Foto: Carlos Valeris, 2012.

38
Métodos
Los datos fueron registrados entre abril de 2012 y junio de 2014, durante 150 días
de trabajo en campo, que incluyeron las fases: aguas bajas, subida de aguas y bajada
de aguas. En la colección de datos participaron 4 biólogos, 8 parabiólogos y 7
monitores comunitarios. Para la evaluación del uso consuntivo se visitaron 8
comunidades y utilizaron datos de Castellanos et al. (2010), WCS (2009) y Perera-
Romero (datos no publicados). Las comunidades visitadas fueron:
Entreríos:
Erebato, en la desembocadura con el río Caura. Posee una pista de aterrizaje para
avionetas y funcionó como estación hidro-metereológica de Edelca (ver figura 6).
Sayusodiña: del Erebato medio, próxima a la boca del río
Cusime. Es la comunidad más populosa de este sector de la CRE (ver figura 7).
Dakudakuña:
Dadiña: pequeña comunidad sanema establecida en la margen derecha del río
Erebato, a 500 metros aguas debajo de la desembocadura del año Dadi (ver figura 8).
Ayawaña: comunidad sanema del Erebato medio, emplazada en la margen
izquierda del río Erebato, a la altura del raudal Ayawa (ver figura 9).
Yudiña: comunidad sanema establecida en la margen izquierda del río Yudi. Es la
comunidad más numerosa de esta cuenca.
Anadekeña:
división de la comunidad de Jüwütüña (ver figura 10).
Jüwütüña:
María de Erebato. Es junto con Anadekeña una de las comunidades con mayor
número de habitantes de la CRE (ver figura 11).
Las comunidades sanemas Ayawaña y Yudiña poseen entre 250 y 300 habitantes,
habitantes respectivamente (Perera-Romero et al., 2015).

39
Figura 6. 2008.
Figura 7. 2009.

40
Figura 8. Comunidad sanema Dadiña. Foto: Carlos Valeris, 2012.
Figura 9. Vista parcial de la comunidad sanema de Ayawaña. Foto: Carlos Valeris, 2008.

41
Figura 10. comunal al fondo. Foto: Carlos Valeris, 2012.
Figura 11. 2009.

42
Estado poblacional Se evaluó el estado poblacional con base a tres variables fundamentales:
abundancia relativa, esfuerzo de búsqueda y estructura de tamaños.
Abundancia poblacional Se estimó por conteos nocturnos en bote y a pie. Los primeros se realizaron con la
ayuda de linternas de frente y faros piloto, mientras que los segundos consistieron en
caminatas a lo largo de caños y cuerpos de aguas menores. Los conteos se realizaron
en curiaras de 12, 10 y 6 metros (según el tamaño y accesibilidad del cuerpo de agua)
a remo a lo largo de los caños y en algunas secciones del canal principal del río,
cuando la corriente lo permitió, durante las fases hidrológicas: bajada de aguas, aguas
bajas y subida de aguas. El motor fuera de borda se utilizó en las secciones más
amplias del río, que tenían una corriente moderada a fuerte.
Una vez observado un individuo, se realizó un acercamiento para asegurar, en
primer lugar, la identificación correcta de la especie. Posteriormente, se estimó la
longitud total (LT). La abundancia relativa para P. trigonatus se obtuvo con base al
número de animales observados por kilómetro lineal de cuerpo de agua muestreado.
Esfuerzo de búsqueda Se refiere como el tiempo de búsqueda necesario para avistar individuos de P.
trigonatus en cada localidad evaluada, utilizando conteos nocturnos (curiara a remo
en los recorridos fluviales y a pie en trayectos por tierra). Se registró la hora de inicio
y final de cada recorrido y el número de individuos observados y utilizó la fórmula:
Esfuerzo de búsqueda (n/t) = Número de individuos observados (n) Tiempo de búsqueda (t)
Se compararon únicamente las localidades donde se detectaron individuos de
P. trigonatus.

43
Estructura poblacional Se estimó la longitud total (LT) de cada individuo observado, acercándose lo
máximo posible. Se procuró la captura de los ejemplares que permitieron una
aproximación al alcance de lazos corredizos o de la mano del observador. La captura
diurna de los ejemplares en caños se realizó por medio de la búsqueda sub-acuática
con máscaras de buceo y su captura con lazos corredizos, en las zonas identificadas
como hábitat potencial y en los lugares donde se registró su presencia a través de
huellas, cuevas o avistamiento directo.
Los ejemplares capturados fueron marcados por medio del corte de las escamas
caudales, con un código único (Campos, Zucco y Batista, 2007). El código empleado
se basó en el corte de las escamas caudales simples (S) y dobles. Para las escamas
caudales dobles se identificó siempre el lado: D (derecha) e I (Izquierda). El número
asignado a las escamas simples inició desde la porción proximal hacia la distal (S1 a
S9). Para las escamas dobles, desde la porción más distal hacia la proximal (ver
figura 12) (e. g.: S1: individuo con la 1era escama simple cortada; S1D1: individuo
con la primera escama simple cortada y la primera escama doble derecha cortada).
Figura 12. Nomenclatura de las escamas caudales dobles y simples de P. trigonatus. Se señalan los códigos correspondientes a cada escama. Se omiten las escamas S6, S7, S8 y S9 por las características de la imagen. Foto: Lucy Perera-Romero, 2013. Diseño: Carlos Valeris.

44
Las medidas morfométricas se tomaron con un calibrador o una cinta métrica
metálica y flexible según el tamaño del animal. Para cada individuo, se registró las
siguientes medidas:
Longitud total (LT): comprende la distancia entre el extremo más anterior del
hocico y el extremo más posterior de la cola, tomada en línea recta de forma ventral.
Longitud rostro-cloaca posterior (LRCp): medida desde el extremo anterior del
hocico hasta la porción posterior de la abertura cloacal.
Largo de la cabeza (LC): distancia comprendida entre el extremo anterior del
hocico y la porción posterior de la cabeza, indicada por el límite posterior de los
huesos supraoccipitales.
Ancho de la cabeza (AC): medido a la altura de la unión mandibular.
Distancia inter-ocular (DIO): corresponde a la distancia entre los bordes
proximales de los ojos, medido con calibrador.
Peso: obtenido con balanza de 20 Kg de capacidad y de una apreciación de 100
gramos.
También se registraron características distintivas como cicatrices, heridas,
presencia de parásitos externos y patrón de las escamas nucales de los individuos
capturados. Los individuos que, a pesar de haber sido identificados, no fueron
estimados en tamaño se registraron como no estimados (NE).
La estructura poblacional, referida a la relación del tamaño corporal de los P.
trigonatus observados y cazados se basó en la LRCp. Para ello, las LRCp fueron
obtenidas al utilizar las relaciones morfométricas LT-LRCp derivadas a partir de la
regresión lineal de las tallas conocidas de LT y LRCp de 12 individuos capturados y
2 individuos cazados (n=14), productos de las capturas realizadas durante el proyecto,
sin distinción entre sexos, resultando la ecuación:
LRCp= 0,5681(LT) + 0,1042 con R2 = 0,9858.
El valor del coeficiente de determinación fue alto a pesar del tamaño de muestra,
lo que implica una relación lineal entre ambas medidas morfométricas, permitiendo el
uso de estas medidas para calcular una a partir de la otra con un margen de error
desestimable.

45
Distribución geoespacial, posición en el hábitat y características del micro-hábitat
Una vez observado un individuo, se registró la coordenada UTM y describieron
los aspectos más relevantes de la posición y del micro-hábitat que ocupaba (Espinosa-
Blanco y Seijas, 2010), así como las características del cuerpo de agua y el bosque
circundante como: tipo de agua (transparente, negra o blanca); ancho del cauce del río
o caño y la cobertura del dosel. Para evaluar la distribución, se incluyeron los
registros de huellas, rastros y nidos observados durante los recorridos diurnos y los
ejemplares cazados, sólo cuando el cazador reconoció el sitio exacto donde el
ejemplar fue cazado o refirió detalles precisos de una localidad ya conocida. Todos
los individuos, rastros, huellas y nidos registrados fueron representados en un Sistema
de Información Geográfico, a través del Programa ArcGIS v.8.2.
Con base a las observaciones obtenidas, se establecieron cinco categorías de
posición en el hábitat:
1. Sobre la orilla (SO): cuando los individuos estaban completamente fuera del
agua.
2. Entre vegetación en el agua (EVA): cuando los individuos estuviesen en el
agua, entre o sobre ramas, troncos, lianas o árboles caídos.
3. En el agua (EA): cuando los ejemplares estuviesen nadando o flotando en el
agua, sin contacto con el fondo, la orilla o vegetación.
4. En interfase agua-tierra (IAT): cuando los ejemplares estuviesen en la
orilla, en contacto con el agua o dentro del agua en contacto con el fondo.
5. Sumergido (SU): cuando los ejemplares fuesen observados en reposo, en el
lecho o dentro de una cueva subacuática en el caño o río, totalmente
sumergidos.
Las características del micro-hábitat fueron evaluadas en campo a través de
medición y estimación de las categorías cuantitativas y cualitativas.

46
1. Tamaño del cuerpo de agua: Se obtuvo a través del promedio de la
medición, con cinta métrica flexible, del ancho del cuerpo del agua en cuatro
puntos dentro del sector evaluado. Se establecieron tres categorías:
a. Caño principal: caños con un ancho igual o superior a los 10 metros, pero
menor a 30 metros.
b. Caño secundario: caños con un ancho menor a 10 metros.
c. Río: incluye a todos los ríos, con ancho igual o mayor a 30 metros.
2. Tipo de cuerpo de agua: Se refiere a la característica del agua (blanca, negra
o clara) en ríos y caños. En el área de estudio, se identificaron cuatro tipos de
cuerpo de agua:
a. Caño de aguas claras: caños donde es posible ver el fondo a través del
agua.
b. Caño de aguas blancas: aguas con sedimentos en suspensión.
c. Río de aguas negras: ríos con aguas color té.
d. Río de aguas blancas: ríos con aguas que transportan gran cantidad de
sedimentos en suspensión.
3. Tipo de orilla más próxima: Se basó en el ángulo de inclinación estimado de
la orilla. Se identificaron tres tipos de orilla según la pendiente:
a. Playa: tipo de orilla con pendiente suave, estimada entre los 10° y 45º, que
incluyen sustrato arenosos o fangoso, generalmente desprovista de
vegetación herbácea o arbustiva.
b. Barrancada: orilla de pendiente estimada entre 45º y 80º.
c. Barranco: orilla de corte abrupto con pendiente estimada en mayor a 80º.
4. Cobertura del dosel: La cobertura del dosel fue estimada a través de la
relación de sombra del dosel sobre el canal principal del caño o río (diferencia
entre el ancho del río o caño y la proporción de la sombra sobre el río o caño

47
entre las 12:00 y 13:00 horas), en tres secciones dentro del sector evaluado. Se
establecieron cuatro categorías de cobertura del dosel:
a. Cobertura total: Entre 100 y 75% de cobertura.
b. Cobertura moderada: Entre 74 y 50% de cobertura.
c. Cobertura parcial: Entre 49 y 25% de cobertura.
d. Poca cobertura: Menos de 25% de cobertura.
Debido al escaso número de observaciones obtenidas para cada tamaño de LRCp,
para el análisis de la posición y micro-hábitat, se agruparon a los ejemplares en tres
categorías:
Categoría I: incluye a los ejemplares neonatos y jóvenes, con tallas entre los 10 y
35 cm de LRCp.
Categoría II: corresponden a individuos sub-adultos, con LRCp mayores a 35 y
hasta 65 cm.
Categoría III: corresponden a ejemplares adultos, LRCp mayor a 65cm.
Las categorías mencionadas fueron establecidas con referencia en Magnusson y
Lima (1991), quienes establecen la edad adulta para P. trigonatus con base en
estimados de edad relativa y crecimiento a partir de individuos re-capturados.
Para establecer si los datos de ubicación espacial y de tamaño de los ejemplares
registrados presentaban o no autocorrelación espacial, se calculó la semivarianza
(Legendre, 1993), utilizando el programa GS+ Versión 10. Una vez comprobada que
no había autocorrelación, se realizó un análisis de varianza (ANOVA) con el
programa estadístico R, para determinar si existe relación entre la categoría de
tamaño del ejemplar y la posición que éste ocupaba en el hábitat. Se utilizó el Método
de Bonferroni, Prueba de Bartlett del Programa Minitab Express Versión 1.4.0 para el
calculó la homogeneidad de la varianza. Esta prueba se aplicó a fin de comprobar si
existía variación entre las posiciones ocupadas por los ejemplares y las características
del micro-hábitat evaluadas y entre su categoría de tamaño y las características del
micro-hábitat.

48
Aspectos reproductivos
Se realizaron búsquedas sistemáticas de nidos de P. trigonatus en 27 localidades
dentro de los cinco sectores de la CRE evaluados. La búsqueda se realizó por los
márgenes de los caños principales y sus tributarios, considerando especialmente
aquellos que estuviesen en hábitat reproductivos potenciales, según la información
obtenida de la literatura disponible y del conocimiento tradicional de los miembros de
las comunidades y parabiólogos indígenas. El esfuerzo se cuantificó con base a los
kilómetros recorridos con relación a las horas/hombre de búsqueda.
Una vez ubicado el nido, se registraron las coordenadas UTM de la localidad y
evaluaron las características del hábitat como sigue:
Distancia con relación al cuerpo de agua: medida en línea recta con cinta
flexible desde el punto central del nido hasta la orilla más próxima del cuerpo de
agua.
Etno-especie del árbol más cercano: identificación de la especie reconocida por
anema del árbol más cercano al nido.
Distancia con relación al árbol más cercano: medida en línea recta desde el
punto central del nido hasta la base del tronco del árbol más próximo.
Diámetro a la altura del pecho (DAP) del árbol más cercano: se midió con
cinta métrica flexible a 1,3 metros sobre el suelo.
Pendiente del terreno: se estimó la pendiente del terreno en un radio de 50
metros.
Ancho del cuerpo de agua: se midió el ancho del cuerpo del agua en tres puntos.
En el punto más próximo a la nidada, a 25 metros aguas arriba y 25 metros aguas
abajo y se calculó la media.
Tipo de cuerpo de agua: se describió con relación al tipo de agua: transparente,
aguas blancas, aguas negras y si era caño o río.
Profundidad del cuerpo de agua: se registraron cinco medidas de profundidad
en el plano transversal del cuerpo de agua (ancho), la primera en la orilla donde el
agua está en contacto, la segunda entre la primera medida y el canal medio del cuerpo
de agua, la tercera en el canal medio del cuerpo de agua, la cuarta entre el canal

49
medio y la orilla opuesta y la quinta en la orilla opuesta. Estas mediciones se
repitieron a 10 metros aguas arriba y 10 metros aguas abajo. En cada sector, se
promedió la profundidad y luego se promedió entre los tres sectores.
Para describir el nido, se registraron las siguientes características:
Altura del montículo: se obtuvo midiendo la distancia entre la base y el punto
más prominente del montículo.
Ancho y largo del montículo: se midió la sección más ancha y la sección más
larga del montículo con cinta métrica.
Profundidad de la cámara: se obtuvo midiendo la distancia entre la base de la
cámara y el punto más alto sobre ella.
Presencia de hormigas o termitas: se registró la presencia de hormigas y termitas
en el nido al momento del hallazgo, así como la presencia de hormigueros y
termiteros en las proximidades de nidos, en un radio de 5 metros.
Se cuantificó el número de huevos y registró el largo y ancho máximo de cada
huevo intacto con un calibrador de metal de alta precisión (± 0,01mm), así como el
ancho de la banda. La biomasa de cada huevo y de la nidada se determinó con una
Pesola® de 50mg de capacidad (± 0,1g de precisión) (Campos y Sanaiotti, 2006).
Para el conjunto de datos, se calculó la media y la desviación estándar.
Registro de nidadas halladas por miembros de las comunidades indígenas
Cincuenta y ocho entrevistas fueron realizadas a informantes clave de ocho
comunidades y se les consultó sobre el hallazgo de nidadas, grupos de neonatos y
hembras con huevos en sus oviductos. A partir de la información obtenida, se
visitaron cinco localidades donde cazadores refieren haber encontrado nidos entre los
años 2012 y 2014. En cada sitio, fueron evaluadas las características del hábitat y
registrado su posición.
Al localizar los nidos mencionados, se analizaron las características biofísicas más
relevantes del micro-hábitat donde los nidos estaban emplazados. La temporalidad de

50
la nidada se basó en el estado de los huevos y en la información aportada por los
recolectores (cazadores), siempre que recordaran con seguridad la fecha de la
recolección de la nidada. El desarrollo de la nidada se estimó con base en Magnusson
(1990) y Vásquez- Ruesta (1983). Los sitios de las nidadas se representaron en un
SIG utilizando el programa ArGIS v.8.2.
Presión de uso Para evaluar la presión de uso para P. trigonatus, se utilizaron dos fuentes de
información: entrevistas y registros de consumo.
Las entrevistas fueron aplicadas por biólogos y parabiólogos entre 2012 y 2014 a
informantes clave sobre el uso consuntivo de P. trigonatus: número de ejemplares
cazados, época del año, lugares de caza; su reproducción: temporalidad y sitio de
nidadas, hallazgo de grupos de neonatos; sobre la percepción de la abundancia de la
especie: lugares donde son abundantes, lugares dónde son escasos y las razones de
ello (ver Apéndice 1). En la tabla 2, se muestran las distintas fuentes de información
analizadas para evaluar la presión de uso por cada comunidad.
De estas comunidades, cinco son ye kwana: Anadekeña, Dakudakuña, Entreríos,
Jüwütüña y Sayusodiña, cuatro sanema: Ayawaña, Dadiña, Troncón, Yudiña.
Campamento Ratón es un asentamiento permanente ocupado por una sola familia
sanema.
Los registros de consumos fueron obtenidos de Castellanos et al., (2010) en 2004
y 2005; el Programa Caura de WCS y la Organización Indígena Kuyujani (2008 y
2009) (Perera-Romero, 2012) y por Perera-Romero entre los años 2013 y 2014 (datos
no publicados), éste último en el marco del proyecto: Comportamiento y patrones
espaciales de la cacería alrededor de anema, financiado
por el Programa de apoyo a investigadores de WCS.
Estos registros fueron realizados por Parabiólogos (años 2004, 2005, 2008 y 2009)
(Castellanos et al., 2010; Perera-Romero, 2012) y Monitores comunitarios de

51
recursos (2013 y 2014) (Perera-Romero, datos no publicados), quienes registraron
cada evento considerando, siempre que fue posible, las variables: fecha, lugar y hora
de la cacería, LT y LRCp (medidas con cinta métrica), sexo, peso, preparación, arte
de caza, características del hábitat, características del grupo de cazadores: número,
tipo de embarcación utilizada; el esfuerzo de caza, entre otros datos de interés. Los
datos obtenidos por el Programa Caura de WCS y la Organización Kuyujani, aquí
analizados, forman parte de una iniciativa de seguimiento de la cacería en la CRK
(Perera-Romero, 2012).
Tabla 2. Registros de consumo consultados y entrevistas realizadas por comunidad. Fuente: 1. Perera-Romero (datos no publicados); 2. Datos propios; 3. Castellanos et al., (2010); 4. WCS- Programa Caura y Organización Kuyujani (datos no publicados).
Comunidad Tipo de registro Año Fuente
Anadekeña Registro de consumo
2013-2014 1
Entrevistas 2014 2
Ayawaña Registros de consumo
2013-2014 1
Entrevistas 2014 2
Dadiña Entrevistas 2012 2
Dakudakuña Entrevistas 2013 2
Entreríos Entrevistas 2013-2014 2
Jüwütüña Entrevistas 2014 2
Sayusodiña Entrevistas 2013 2
Yudiña Registro de consumo
2013-2014 1
Entrevistas 2013-2014 2
Troncón Entrevista 2014 2
Campamento Ratón Entrevista 2014 2
Cuenca del Río
Registro de consumo
2004-2005 3
Registro de consumo
2008-2009 4

52
Los datos referidos fueron analizados con base a: número de ejemplares cazados,
frecuencia de caza, esfuerzo de búsqueda e importancia relativa de la especie con
relación al resto de especies cazadas, por número y biomasa. Se analizó la estructura
de tamaños de los ejemplares cazados y el esfuerzo de búsqueda durante las jornadas
de caza.
Recomendaciones para la conservación y manejo de P. trigonatus
Analizados los resultados, se elaboró una lista de recomendaciones para la
conservación y manejo de P. trigonatus en los sectores de la cuenca evaluados. Las
recomendaciones se basaron en los criterios de conservación: A. Protección de las
áreas con mayor abundancia, consideradas zonas fuentes. B. Protección de áreas para
cada sector de las cuencas evaluadas; C. Resguardo de las áreas reproductivas y
manejo: D. Exclusión de ejemplares según su talla.
Se presentaron los mismas en la comunidad sanema Yudiña, con el objetivo que
acordaran (si así lo consideraban), medidas para la conservación y uso sostenible de
P. trigonatus en las áreas de influencia de sus comunidades. La actividad consistió en
un Taller participativo donde se congregaron los ancianos, cazadores, mujeres, niños
de la comunidad, seis ancianos y cazadores de la comunidad Ayawaña. En el taller, se
presentaron los resultados obtenidos por sector y comunidad, a la vez que se
promovió la participación de los ancianos y cazadores. Se presentan los acuerdos
alcanzados en el taller.

53
CAPÍTULO IV
RESULTADOS Y DISCUSIÓN
La única especie de Crocodylia identificada en los sectores de la CRE evaluados
fue Paleosuchus trigonatus -
fronted caiman) (figura 13). Esta identificación se corresponde con la distribución
esperada de P. trigonatus, en ríos y caños de cuencas altas al sur del río Orinoco,
aunque sólo hubo un registro previo para el río Erebato (Godshalk, 1982) y 38 para
Venezuela (Seijas, 2007). Aunque Castellanos et al. (2010) mencionan el uso
consuntivo de P. palpebrosus para la CRK, esta especie no se registró en ésta cuenca,
ni en los sectores evaluados. Tampoco se ha documentado la presencia de P.
palpebrosus en otras localidades de la cuenca alta del río Caura. Los registros de P.
palpebrosus de Castellanos et al. (2010) pueden considerarse como P. trigonatus,
debido a una errónea identificación de los ejemplares revisados en registros de
consumo en la CRK entre 2008 y 2009 (WCS- Programa Caura, datos no publicados;
Perera-Romero, 2012). Las localidades más próximas donde se han registrado a esta
especie son la cuenca baja del río Caroní y en las porciones media y baja del río
Orinoco (Seijas, 2007), que distan más de 200 Km lineales del área de estudio.

54
Figura 13. Paleosuchus trigonatus . Foto: Igor Castillo, 2013. Estado poblacional
En todas las localidades evaluadas, no siempre fue posible estimar la abundancia
de P. trigonatus, debido a las difíciles condiciones de acceso (caños no navegables
por la topografía o por la gran cantidad de árboles caídos). En estos casos, sólo fueron
posibles efectuar recorridos parciales en curiara y, en algunos casos, a pie a lo largo
de las orillas de los cuerpos de agua. En esas localidades, se comprobó la presencia de
P. trigonatus a través de los registros directos durante los recorridos diurnos y
nocturnos, e indirectos (huellas, rastros, nidos). Se obtuvieron sólo datos de
abundancia relativa (ind/Km lineal de cuerpo de agua) para el sector Madajano (caño
Ajuju y río Madajano) y el sector Erebato medio (caño Dadi).

55
Abundancia relativa
Resumidos en la tabla 3, están los tres recorridos nocturnos realizados en el caño
Ajuju, afluente del río Madajano. El segundo recorrido incluyó el mismo sector
evaluado en 2012, mientras que el tercero incluyó aquellos sectores evaluados en los
recorridos previos. En todos los conteos realizados, se observaron ejemplares de P.
trigonatus.
Tabla 3. Conteos nocturnos en el caño Ajuju, con los valores de abundancia relativa para cada recorrido.
Localidad
Caño Ajuju
Fecha
(Bajada de aguas)
Kilómetros recorridos
Nº de P. trigonatus observados
Abundancia relativa (ind/Km)
1er Conteo 25/11/2012 7,3 15 2,05
2do Conteo 03/11/2013 12 18 1,5
3er Conteo 05/11/2013 14,4 13 0,9
Como se aprecia en la tabla 3, la abundancia relativa máxima registrada fue 2,05
ind/Km lineal del cuerpo de agua y la mínima, de 0,9 ind/Km. Cabe destacar que,
para este cálculo, se consideran los ejemplares observados sólo durante los conteos
nocturnos. Esta localidad es referida por miembros de las comunidades y por
cazadores como la localidad con mayor abundancia de yadiwes (P. trigonatus) de la
CRK, donde ocasionalmente realizan jornadas de caza y pesca. Sin embargo, es una
localidad muy remota, donde pocos grupos de cazadores llegan. En ese caño, se

56
observaron cuatro P. trigonatus durante el recorrido de reconocimiento en horas
matutinas y se registraron seis huellas y rastros en orillas lodosas y barrancadas en
2012. En el río Madajano, se realizaron cuatro recorridos, dos por sección del río. En
la tabla 4, están representados los valores de abundancia obtenidos.
Tabla 4. Conteos nocturnos en el río Madajano, con los valores de abundancia relativa para cada recorrido. Localidad
Río Madajano
Fecha (Bajada de
aguas)
Kilómetros recorridos
Nº de P. trigonatus
observados
Abundancia relativa (ind/Km)
1er Conteo (sección 2) 22/11/2012 9,1 1 0,1
1er Conteo (sección 1) 23/11/2012 6,2 4 0,64
2do Conteo (sección 1) 06/11/2013 6,2 3 0,48
2do Conteo (sección 2) 07/11/2013 9,1 1 0,1
Al comparar las abundancias relativas máximas obtenidas en el caño Ajuju y el río
Madajano, el caño Ajuju registró una abundancia 3,2 veces más alta, que aquella del
río Madajano. Todo parece indicar que P. trigonatus pudiera tener mayor preferencia
por el caño que por el canal principal del río. Los únicos datos de abundancia para
esta especie en Venezuela, aportados por Gorzula y Paolillo (1986), refieren una
abundancia relativa de 0,2 y 1,6 ind/Km en ríos de la Guayana venezolana, con sólo
un dato para caños de 7,5 ind/Km. Aunque este último dato se obtuvo con base a la
observación de 3 ejemplares en los 0,4 Km de cuerpo de agua evaluados, en la
porción próxima a la desembocadura (Seijas, 2007), por lo que no es representativo.
Aunque el río Madajano y el caño Ajuju tienen sus nacientes muy próximas,
algunas de las características hidrológicas y estructurales de estos cuerpos de agua

57
difieren (ver tabla 1). El río Madajano es un río de aguas negras y un ancho de 60
metros en promedio, mientras que el caño Ajuju transporta gran cantidad de
sedimentos en suspensión y tiene un ancho promedio de 20 metros. Ambos son
corrientes de aguas frías (temperaturas de 20ºC en noviembre), debido a la cercanía
de la Meseta Jaua, donde tienen sus nacientes.
El hecho que no se hayan identificado a otras especies de cocodrilianos en la
cuenca permite excluir la competencia inter-específica como causa de la relativa alta
o baja abundancia poblacional de P. trigonatus en la CRE. No así la cacería de
subsistencia, que se ha practicado y practica en ese sector de la cuenca. Con relación
a los estudios poblacionales de P. trigonatus, Magnusson y Lima (1991) refieren que
la condición críptica y las características del hábitat que ocupa, representan
limitaciones a los métodos convencionales utilizados para estudiar a esta especie de
cocodriliano.
Además del caño Ajuju y el río Madajano (ambas localidades del sector
Madajano), se evaluaron parcialmente 11 caños y 9 secciones de los ríos: Ka kada
(3), Yudi (1) y Erebato (5), en el resto de los sectores del área de estudio: Alto
. En todos los sectores, se comprobó la
presencia de P. trigonatus (ver Apéndice 2), identificándose en 40% de las
localidades evaluadas. Debido a la evaluación incompleta de los caños, como
consecuencia del bajo nivel de las aguas y la gran cantidad de árboles caídos, no es
posible descartar la presencia de P. trigonatus en éstos, aunque no hayan sido
detectados durante los recorridos.
Esfuerzo de búsqueda El esfuerzo de búsqueda varió entre 0,25 y 3,37 horas por individuo entre las
localidades evaluadas (ver tabla 5). Estos valores refieren a una medida indirecta
basada en la probabilidad de avistamiento, lo que podría relacionarse con los índices
de cacería y esfuerzo de caza. Las localidades donde se observaron más individuos
por unidad de esfuerzo fueron: caño Ajuju y río Madajano (ambas dentro del sector

58
Madajano). En este último, se capturó el ejemplar de mayor tamaño (ejemplar macho
de 91 cm de LRCp). El aislamiento y difícil acceso a estos cuerpos de agua, pudieran
explicar la facilidad de detección de los individuos, los cuales fueron avistados en
horas diurnas inclusive y no mostraron un comportamiento cauteloso al momento del
avistamiento en horas nocturnas.
Tabla 5. Esfuerzo de búsqueda para P. trigonatus en 8 localidades de la CRE. Los valores para cada localidad representan el menor esfuerzo registrado en un recorrido entre los conteos nocturnos realizados, salvo aquellas localidades donde se efectuó un solo recorrido. Los valores son expresados en horas: 1= 60 minutos; 0,5= 30 minutos; 0,25= 15 minutos, necesarios para la observación de un individuo. Localidad Número
de conteos
Fecha Tiempo de búsqueda
(horas)
Número de individuos observados
Esfuerzo de búsqueda (h/ ind)
Esfuerzo de búsqueda (min/ ind)
Caño Suajadito 2 19/04/2012 1 1 1 60
Caño Kajioko 2 20/04/2012 2,1 1 2,1 126
Río Ka kada 4 23/04/2012 2 2 1 60
Caño Dadi 1 30/04/2012 7 4 1,75 105
Río Madajano 4 23/11/2012 2,85 4 0,71 42
Caño Ajuju 3 03/11/2013 4,42 18 0,25 15
Caño Kadujaimudu 2 16/11/2013 3,37 1 3,37 202
Caño Jadia 1 27/04/2014 1,74 1 1,74 104
En el sector Yudi, no se observaron P. trigonatus durante los conteos nocturnos.
Esta cuenca está poblada por dos comunidades sanema: Yudiña y Salasalaña. La

59
primera está localizada en la cuenca media y la segunda, en la cuenca baja próxima a
la desembocadura del río Yudi en el río Erebato. La mayoría de las localidades
evaluadas son de fácil acceso, lo que favorece una mayor presión de cacería en esa
cuenca por miembros de las comunidades mencionadas. No obstante, la captura de
una hembra grávida (LRCp= 76 cm; peso= 10,5 Kg), en caño Wali, tributario del río
Manasi, en marzo de 2013 (durante un recorrido diurno) y los resultados de
entrevistas y registro de cacería (Perera-Romero, datos no publicados), evidencian la
presencia de ejemplares adultos y jóvenes y de actividad reproductiva en la zona.
En el sector Alto Erebato se registró sólo un ejemplar en caño Jadia. No se
observaron ejemplares en el canal principal del río. Esto, como lo reseñaron Medem
(1967) y Magnusson (1990), puede deberse a que P. trigonatus prefiere los caños y
corrientes de agua menores dentro del bosque tropical. Aunque los mismos autores
señalan que los machos adultos y los jóvenes en búsqueda de territorios pueden
observarse en corrientes de agua mayores, como lo sería el río Erebato. Otra posible
causa de la ausencia de registros en este sector del río Erebato, pudiera deberse a la
constante actividad de caza y pesca de subsistencia practicada por miembros de las
comunidades Jüwütüña y Anadekeña y el tráfico frecuente de embarcaciones con
motor fuera de borda a lo largo de esta arteria fluvial, lo que podría alejar a los P.
trigonatus o hacerlos más cautelosos.
En el Erebato medio, se registró a P. trigonatus en el caño Dadi, donde se
observaron 4 ejemplares en los 9 Km evaluados (0,44 ind/Km). En ese caño,
miembros de la comunidad Dadiña informaron la caza de ejemplares adultos y el
hallazgo de nidos. También hacen referencia a que un mayor número de individuos
están presentes en las cabeceras, a más de 20 Km aguas arriba de la desembocadura.
En este sector, tampoco se registró la presencia de la especie en el canal principal del
río Erebato.

60
Estructura poblacional
Se basa en 60 individuos observados a los que se les estimó su talla, que
representan el 71,4% de todos los animales avistados en la CRE. Trece de ellos
fueron capturados. En la figura 14, se agrupan las observaciones en intervalos de 10
cm con relación a sus valores de LRCp. En la misma están representados los
individuos de todas las tallas, con excepción de los neonatos, que no fueron
observados durante los conteos nocturnos.
El 31,67% de los registros corresponden a individuos entre los 30 y 39,9 cm de
LRCp y el 21,67% por ejemplares de 40 a 49,9 cm de LRCp. Este grupo de tamaño lo
integran animales sub-adultos (Magnusson, 1990). Un 18,33% de las observaciones
corresponden a individuos entre los 50 y 59,9 cm. El resto de las categorías de
tamaño combinadas (60 a 110 cm) equivalen a 16,67%. Los ejemplares de menor
tamaño observado, entre los 20 y 29,9 cm de LRCp, corresponden a 11,67% de todos
los registros. Los estimados se asemejan a los tamaños referidos de los P. trigonatus
cazados entre 2008 y 2009 en la CRK (WCS-Programa Caura, datos no publicados),
sugiriendo que este grupo es, al menos, el más accesible para los cazadores en ese
sector. No hay datos publicados sobre la estructura poblacional de P. trigonatus en
localidades venezolanas, lo que no hace posible comparar este resultado con el de
otras poblaciones (o período) en el país.
La cacería de P. trigonatus por parte de los miembros de las comunidades
, basada en la amplia selección de
intervalos de tamaño de los yadiwe (iatamö) cazados. Los ejemplares de P. trigonatus
cazados en las comunidades de Anadekeña, Ayawaña y Yudiña entre 2013 y 2014
totalizaron 25 individuos, cuyos tamaños mínimo y máximo eran de 28 y de 84 cm de
LRCp (Perera-Romero, datos no publicados) (ver presión de uso).
Estos resultados pueden estar más relacionados con el comportamiento críptico de
P. trigonatus y su detectabilidad. Los ejemplares adultos generalmente son más
esquivos y sedentarios, mientras que los más jóvenes tienden a dispersarse en procura
de establecer sus territorios (Magnusson y Lima, 1991).

61
Figura 14. Estructura poblacional de P. trigonatus en la Cuenca del Río Ka kada. Sólo se incluyen a los individuos estimados en tamaño.
Distribución geoespacial, posición en el hábitat y características del micro-hábitat
Se obtuvieron en total 95 registros de P. trigonatus por la identificación o captura
de los ejemplares en las adyacencias. Las huellas, rastros, nidos y observaciones
casuales incluyeron 56 registros, que fueron confirmados en gran medida por la
identificación o captura de P. trigonatus en las adyacencias. Las entrevistas sumaron
116 registros y la caza de 25 P. trigonatus en las comunidades (Perera-Romero, datos
no publicados). Los registros derivados de los conteos nocturnos, capturas y registros
indirectos, tienen información geográfica asociada (coordenada UTM), mientras que
las entrevistas y los registros de caza refieren una localidad o sector.
La distribución de P. trigonatus abarca todos los sectores de la CRE evaluados
(ver figuras 15 a 20). En Alto Erebato se detectó la presencia de P. trigonatus en los
caños Sajme, Jadia, Masümü y Madisiji (ver figura 15). Dos de los registros son de
nidos, uno de ellos aprovechado (caño Madijisi) y el otro hallado posterior a la
eclosión (caño Sajme).

62
Figura 15. Registros de P. trigonatus en el sector Alto Erebato. Los puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios.
En el Erebato medio, se comprobó la presencia de P. trigonatus en el caño Dadi,
donde también se registró el hallazgo y aprovechamiento de un nido (ver figura 16).
En el resto de las localidades evaluadas en este sector: caño Jadana/Akudu y canal
principal del río Erebato, no se detectó a la especie. Sin embargo, los registros de
cacería de la comunidad Ayawaña refieren la caza de P. trigonatus en los caños
Ayawa y Silakasötu y en el campamento Jutumatapata, localidades próximas a la
comunidad.

63
Figura 16. Registros de P. trigonatus en el sector Erebato medio. Los puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios.
En Yudi, se detectó la presencia de P. trigonatus en los caños Wali, Juti, Sisipö y
Tomasito, lo que confirma la presencia de esta especie en este sector (ver figura 17), a
pesar de la actividad de cacería asociada con las comunidades sanema Yudiña y
Salasalaña. También se registró un nido aprovechado y el hallazgo de dos nidos no
activos (ver Aspectos reproductivos).

64
Figura 17. Registros de P. trigonatus en el sector Yudi. Los puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios.
de P. trigonatus de P. trigonatus se detectó en el canal
principal del río
y Kaduaimudu (ver figura 18). La mayoría de los registros se realizaron en caño
Ñayudi, alejados del canal principal del río. Ñayudi posee aguas claras, cobertura de
80% y pendientes entre 30 y 60º de inclinación en su zona media y alta (ver figura
19). Esto sugiere que esta especie se distribuye en toda la cuenca baja del río
, con mayor presencia en caños.

65
Figura 18. Registros de P. trigonatus representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios.
Figura 19. Caño Ñayudi. Foto: Carlos Valeris, 2012.

66
En el sector Madajano, se registraron P. trigonatus en la sección navegable del río
Madajano y en el caño Ajuju. En esta última localidad se registró el mayor número
ejemplares durante los conteos nocturnos (ver figura 20). Este caño posee un canal
amplio (de aproximadamente 20 mts de ancho) y una cobertura de dosel de 40% en
promedio (ver figura 21).
En la CRK, las diferencias de abundancia entre localidades puede estar
condicionada por la presión de caza, mayor en (ver registro de
cacería), y en el canal principal del río y caños navegables, que en los caños pequeños
y en las áreas más distantes a éste. En el resto de los sectores evaluados, las áreas más
alejadas de las comunidades son las que concentran la mayoría de los registros de
caza.
Figura 20. Registros de P. trigonatus en el sector Madajano. Los puntos rojos representan registros de conteos nocturnos y los puntos amarillos registros diurnos directos e indirectos. Mapa elaborado por: Lucy Perera-Romero. Datos de los registros propios.

67
Al observar las figuras 18 y 20, se aprecia la concentración de los registros de P.
trigonatus en la cuenca del r con una proporción menor en las localidades
próximas a las comunidades indígenas. Esta cuenca no posee asentamientos humanos
permanentes y es considerada un área multiuso por las comunidades y
(Castellanos et al., 2010: Perera-Romero et al., 2015), lo que podría constituirse en
un área clave para la zonificación y planificación de las actividades de caza, pesca y
recolección. Los resultados tienen similitud a los obtenidos durante las entrevistas de
percepción sobre el estado de esta especie por parte de las comunidades indígenas
(Castillo et al., 2015), lo que refleja la difícil detección o escasa presencia de P.
trigonatus en algunos sectores y su fácil detección o abundancia relativa mayor en
otros.
Figura 21. Caño Ajuju, desembocadura en el río Madajano. Foto: Carlos Valeris, 2009.

68
El resultado de la evaluación de la semivarianza de los registros directos de P.
trigonatus en la CRE indica que los datos no están autocorrelacionados entre sí y
están definidos por un modelo lineal (Robertson, 2008), lo que sugiere un patrón de
distribución en gradiente. La no autocorrelación de los datos permitió el uso de
pruebas estadística paramétricas, para los análisis de las variables de interés asociados
a los registros directos (Legendre, 1993).
Posición y características del micro-hábitat
Los registros de observaciones de P. trigonatus realizados durante los conteos
nocturnos, incluyeron cinco posiciones en el hábitat que los ejemplares ocupaban al
momento de la observación. El análisis de varianza ANOVA entre las categorías de
los ejemplares (jóvenes, sub-adultos y adultos), basado en su LRCp y la posición que
ocupaban resultó ser no significativo.
Para P. trigonatus, el tipo de cuerpo de agua: caño o río, es un factor determinante
en la posición que éste ocupa en esos ambientes. En los caños, los ejemplares ocupan
todos los espacios disponibles, sin mayores limitaciones ambientales. En los ríos de
canal amplio, con raudales y corrientes más fuertes, los ejemplares tienden a ocupar
las orillas y zonas con rápidos y saltos, con vegetación en las riberas (Medem, 1967).
Con relación al análisis de homogeneidad de la varianza de Bonferroni, método
Barlett, entre la posición ocupada y el tipo de cuerpo de agua, resultó ser significativa
(F= 23,66; g. l.= 65; P = 0,0086). Sobre las características de los cuerpos de agua que
ocupa P. trigonatus en la cuenca del río Caroní, Gorzula et al. (1987) refieren que
éstos son de temperaturas frías (media 22 °C), sus aguas ácidas (pH entre 4 y 6) y
baja conductividad. En esta cuenca, P. trigonatus se ha registrado en un amplio
gradiente altitudinal (100 - 1340 m.s.n.m). En la CRE, los cuerpos de agua evaluados
presentaron valores similares de temperatura y pH (ver tabla 1, p. 68), en un gradiente
entre los 300 y 400 m.s.n.m).
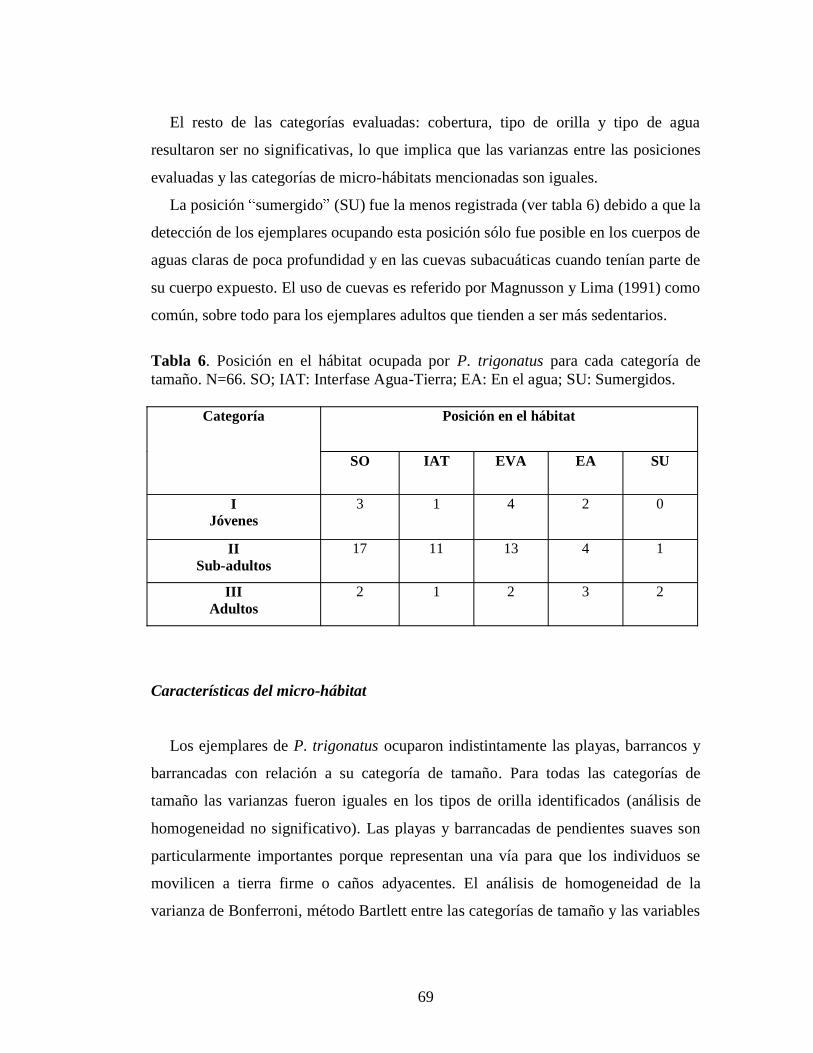
69
El resto de las categorías evaluadas: cobertura, tipo de orilla y tipo de agua
resultaron ser no significativas, lo que implica que las varianzas entre las posiciones
evaluadas y las categorías de micro-hábitats mencionadas son iguales.
La posición sumergido (SU) fue la menos registrada (ver tabla 6) debido a que la
detección de los ejemplares ocupando esta posición sólo fue posible en los cuerpos de
aguas claras de poca profundidad y en las cuevas subacuáticas cuando tenían parte de
su cuerpo expuesto. El uso de cuevas es referido por Magnusson y Lima (1991) como
común, sobre todo para los ejemplares adultos que tienden a ser más sedentarios.
Tabla 6. Posición en el hábitat ocupada por P. trigonatus para cada categoría de tamaño. N=66. SO; IAT: Interfase Agua-Tierra; EA: En el agua; SU: Sumergidos.
Categoría Posición en el hábitat
SO IAT EVA EA SU
I Jóvenes
3 1 4 2 0
II Sub-adultos
17 11 13 4 1
III Adultos
2 1 2 3 2
Características del micro-hábitat
Los ejemplares de P. trigonatus ocuparon indistintamente las playas, barrancos y
barrancadas con relación a su categoría de tamaño. Para todas las categorías de
tamaño las varianzas fueron iguales en los tipos de orilla identificados (análisis de
homogeneidad no significativo). Las playas y barrancadas de pendientes suaves son
particularmente importantes porque representan una vía para que los individuos se
movilicen a tierra firme o caños adyacentes. El análisis de homogeneidad de la
varianza de Bonferroni, método Bartlett entre las categorías de tamaño y las variables

70
del micro-hábitat evaluadas resultó ser significativo para: tipo de cuerpo de agua (F=
26,02; g.l.= 65; P = 0,005), cobertura (F= 21,17; g.l.= 65; P = 0,0067) y tipo de agua
(F= 22,66; g.l.= 65; P = 0,0009).
Con relación al tipo de cuerpo de agua, cobertura y tipo de agua, los resultados
indican variación en la presencia de P. trigonatus con relación a la categoría de
tamaño de los ejemplares. Los caños han sido identificados como hábitats
reproductivos, por lo que las hembras y machos adultos ocupan estos espacios y
establecen allí sus territorios (Medem, 1967; Magnusson, 1990; Magnusson y Lima,
1991).
En la CRE, los individuos adultos ocupan tanto caños como ríos, mientras que los
jóvenes y sub-adultos fueron comúnmente observados ocupando los caños. Aunque
esto puede estar relacionado con los patrones de dispersión de los individuos, también
puede ser consecuencia de la intensidad de cacería, mayor en los ríos por ser la vía
fluvial más utilizada por las comunidades y menor en los caños más remotos e
inaccesibles. Los ejemplares adultos son más cautelosos y en consecuencia difíciles
de cazar, lo que puede explicar su presencia en algunos sectores de los ríos y caños de
canal amplio, donde se registra actividad de caza (ver Presión de uso).
Aspectos reproductivos
Sólo se registró un nido activo en el sector Erebato medio y un nido viejo (sin
huevos) en Alto Erebato (ver figura 22). El esfuerzo de búsqueda fue de 60 Km
recorridos y 135 horas/hombre de búsqueda. La dificultad que representó identificar
un nido de P. trigonatus en su hábitat, incluso para los pobladores locales, exige un
mayor esfuerzo de búsqueda e incluir las zonas más remotas de caños (nacientes),
donde miembros de las comunidades indígenas refieren el hallazgo de nidos en otros
sectores de la CRE.
En mayo de 2013, se realizó el hallazgo del primer y único nido de P. trigonatus
evaluado en Venezuela a la fecha, en un tributario del caño Washadi, afluente del río

71
Erebato en su sección media (ver figura 23) (Valeris et. al., 2014). El nido estaba
parcialmente depredado por Tupinambis teguixini (Mato de agua), suposición basada
en el tamaño del agujero y la disposición de las cáscaras. Es probable que la
depredación haya estado asociada con el hallazgo del nido, una semana antes de la
evaluación. Campos et al. (2016) documentaron la depredación de huevos bien
desarrollados de P. trigonatus por parte de Priodontes maximus (Cuspón), Dasypus
novemcinctus (Cachicamo montañero), Eyra barbara (Hurón), Nasua nasua (Coatí) y
T. teguixini. En dos de los cuatro nidos referidos, las hembras no defendieron la
nidada (op. cit).
Al momento del hallazgo, la presunta hembra estaba en el caño a
aproximadamente 2 mts de distancia, dentro de una cueva con la cola expuesta, pero
no defendió el nido y no fue hallada al momento de la evaluación de éste. Se contaron
13 huevos, 8 intactos con una biomasa media de 85,81 gr y dimensiones de 70,56 mm
de largo y 45,44 mm de ancho (ver tabla 7). Todos los huevos intactos presentaron
una banda central de 39,78 mm en promedio y la mayoría estaba total o parcialmente
cubierto por una mezcla de barro elaborada por las termitas asociadas al nido (ver
figura 24). La biomasa estimada de la nidada fue 1.115,56 gr (Valeris et al., 2014).
Figura 22. Nido de P. trigonatus hallado en afluente del caño Washadi, mayo de 2013. Foto: Raquel Kassalian.

72
Tabla 7. Detalle de la talla (largo y ancho), ancho de la banda y peso de los huevos intactos.
Huevo N°
Largo (mm) Ancho (mm) Ancho de
banda(mm) Peso (gr)
1 69,92 46,18 36,9 85,5
2 71,7 45,68 38,64 85,6
3 71,14 46,2 44,92 83,3
4 69,92 45,8 38,04 83,6
5 69,16 46,3 38,72 85,1
6 70,42 46,08 39,36 82
7 71,38 41 41,52 89,9
8 70,88 46,28 40,16 91,5
Media 70,56 45,44 39,78 85,81
D.E. 0,86 1,81 2,49 3,28
Figura 23. Caño afluente del caño Washadi. En sus orillas se halló el nido de P. trigonatus referido. Foto: Raquel Kassalian, 2013.
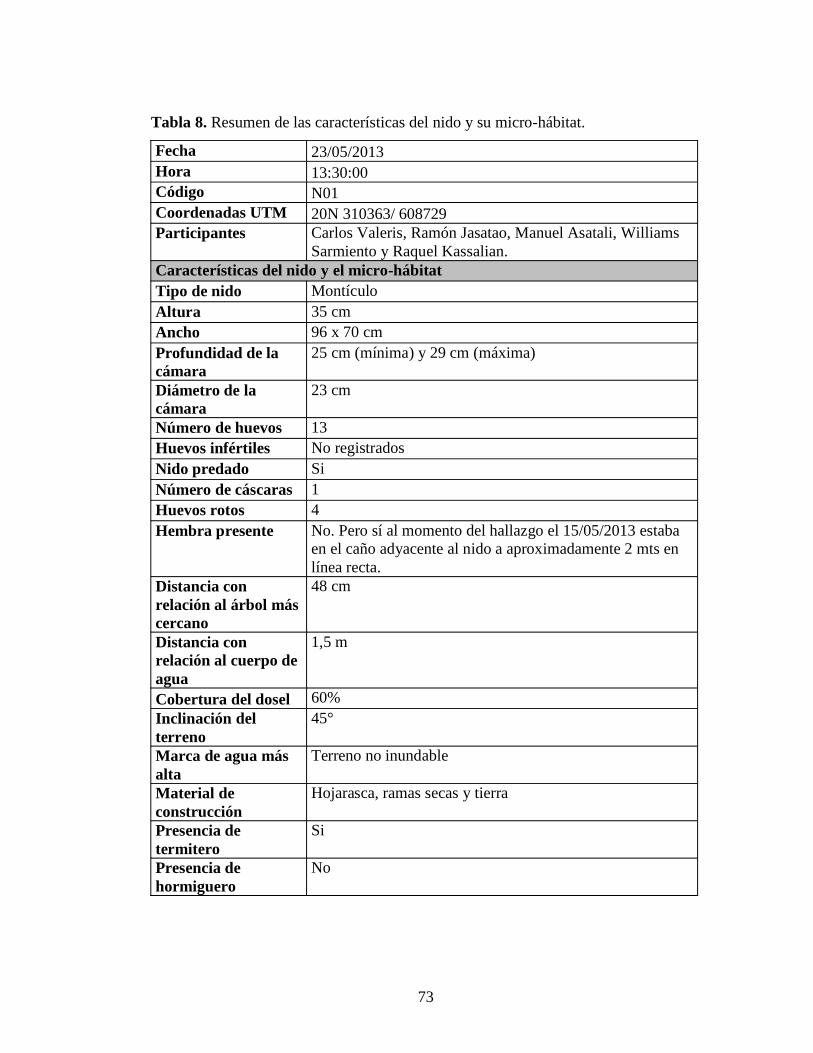
73
Tabla 8. Resumen de las características del nido y su micro-hábitat.
Fecha 23/05/2013 Hora 13:30:00 Código N01 Coordenadas UTM 20N 310363/ 608729 Participantes Carlos Valeris, Ramón Jasatao, Manuel Asatali, Williams
Sarmiento y Raquel Kassalian. Características del nido y el micro-hábitat Tipo de nido Montículo Altura 35 cm Ancho 96 x 70 cm Profundidad de la cámara
25 cm (mínima) y 29 cm (máxima)
Diámetro de la cámara
23 cm
Número de huevos 13 Huevos infértiles No registrados Nido predado Si Número de cáscaras 1 Huevos rotos 4 Hembra presente No. Pero sí al momento del hallazgo el 15/05/2013 estaba
en el caño adyacente al nido a aproximadamente 2 mts en línea recta.
Distancia con relación al árbol más cercano
48 cm
Distancia con relación al cuerpo de agua
1,5 m
Cobertura del dosel 60% Inclinación del terreno
45°
Marca de agua más alta
Terreno no inundable
Material de construcción
Hojarasca, ramas secas y tierra
Presencia de termitero
Si
Presencia de hormiguero
No

74
Figura 24. Huevos fértiles de P. trigonatus. Se observa cubierta de barro de termitas en algunos de los huevos. Foto: Raquel Kassalian, 2013.
Durante la evaluación de otras localidades de la CRE, entre 2012 y 2013 se geo-
referenciaron cinco localidades, donde miembros de las comunidades indígenas
informaron haber encontrado y aprovechado nidos de P. trigonatus (ver figura 25 y
tabla 9). Dos de los registros corresponden a una misma localidad, donde cazadores
reportan haber hallado restos de cáscaras de P. trigonatus y dos nidadas en años
distintos: una en 2010 y otra en 2013. Esto sugiere la posibilidad de fidelidad de sitio
para la construcción de los nidos o bien la selección del mismo lugar por dos hembras
distintas, hecho registrado para Alligator mississipiensis (Elsey, Trosclair y Glenn,
2008).

75
Figura 25. Distribución de los nidos registrados de P. trigonatus en la CRE. Las estrellas amarillas representan su ubicación relativa. La estrella en el sector Yudi corresponde a dos nidos en la misma localidad. Mapa elaborado por: Lucy Perera- Romero. Datos de los registros propios.
Los sitios donde los nidos estaban emplazados mostraron gran similitud en cuatro
elementos del hábitat: cercanía al cuerpo de agua, construidos en la ribera de caños no
navegables en bosques de tierra firme, asociado con una orilla tipo barranco con un
desnivel superior a 1 metro con relación al nivel del agua del caño y la proximidad a
la base de un árbol de diámetro mayor a 70 cm (DAP) (ver figura 26). Rivas et al.,
(2001) y Vásquez-Ruesta (1983) refieren tres de éstos elementos durante el hallazgo
de un nido de P. trigonatus en la Amazonía ecuatoriana y otro en la Amazonía
peruana: la construcción del nido en la base de un árbol, próximo a un caño no
navegable (de 1 metro de ancho), en un área no inundable (bosque de tierra firme).
También señalaron la presencia de un claro en el dosel que permitía el paso de luz
solar sobre el montículo del nido. Todos los informantes mencionaron la asociación
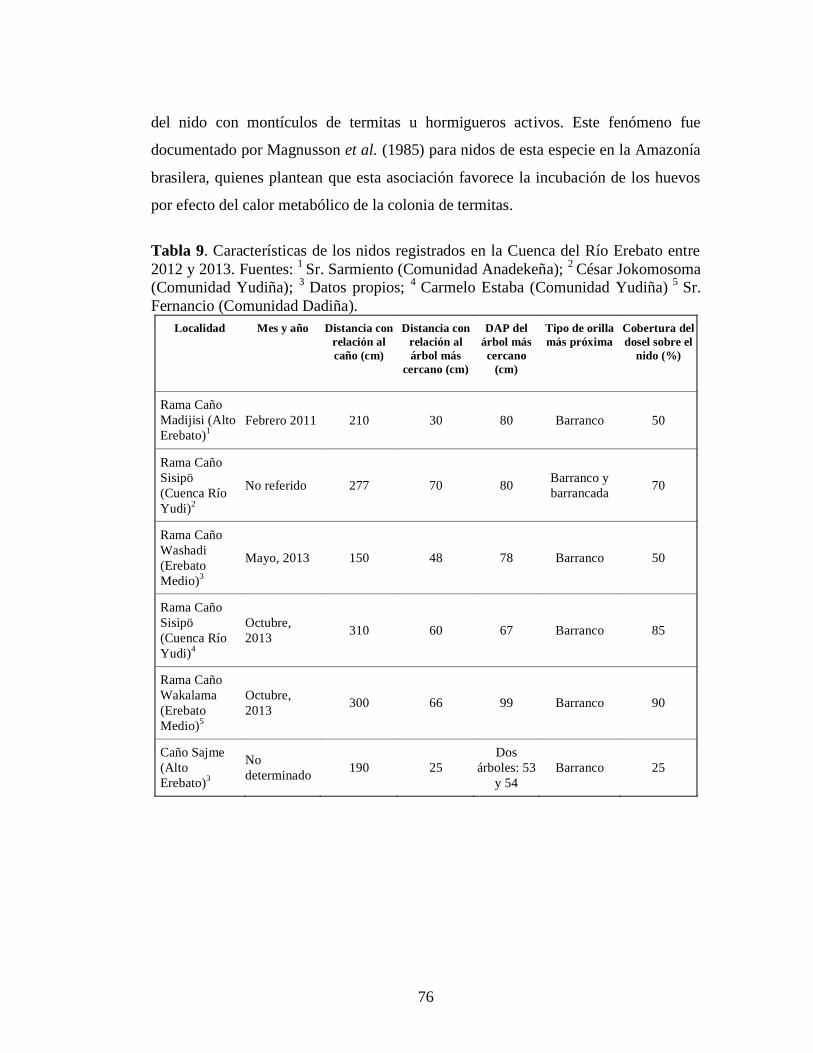
76
del nido con montículos de termitas u hormigueros activos. Este fenómeno fue
documentado por Magnusson et al. (1985) para nidos de esta especie en la Amazonía
brasilera, quienes plantean que esta asociación favorece la incubación de los huevos
por efecto del calor metabólico de la colonia de termitas.
Tabla 9. Características de los nidos registrados en la Cuenca del Río Erebato entre 2012 y 2013. Fuentes: 1 Sr. Sarmiento (Comunidad Anadekeña); 2 César Jokomosoma (Comunidad Yudiña); 3 Datos propios; 4 Carmelo Estaba (Comunidad Yudiña) 5 Sr. Fernancio (Comunidad Dadiña).
Localidad Mes y año Distancia con relación al caño (cm)
Distancia con relación al árbol más
cercano (cm)
DAP del árbol más cercano
(cm)
Tipo de orilla más próxima
Cobertura del dosel sobre el
nido (%)
Rama Caño Madijisi (Alto Erebato)1
Febrero 2011 210 30 80 Barranco 50
Rama Caño Sisipö (Cuenca Río Yudi)2
No referido 277 70 80 Barranco y barrancada 70
Rama Caño Washadi (Erebato Medio)3
Mayo, 2013 150 48 78 Barranco 50
Rama Caño Sisipö (Cuenca Río Yudi)4
Octubre, 2013 310 60 67 Barranco 85
Rama Caño Wakalama (Erebato Medio)5
Octubre, 2013 300 66 99 Barranco 90
Caño Sajme (Alto Erebato)3
No determinado 190 25
Dos árboles: 53
y 54 Barranco 25

77
Figura 26. Sitios donde se registraron nidos de P. trigonatus: 1. Rama caño Madijisi (Sector Alto Caura); 2. Rama caño Wadishidi (Erebaro medio); 3. Caño Sajme (Erebato medio); 4. Caño Wakalama (Erebato medio) y 5. Rama caño Sisipö (Yudi). Fotos: 1 y 3. Carlos Valeris; 2. Raquel Kassalian; 4 y 5. Igor Castillo.
Uno de los aspectos más resaltantes sobre la reproducción de P. trigonatus en la
CRE y en el resto de las localidades donde se ha estudiado a esta especie es la
temporalidad de las nidadas. Magnusson (1990) sugiere que en la Amazonía brasilera
esta especie construye sus nidos en la temporada seca y los nacimientos ocurren al
inicio de la temporada lluviosa. No obstante, el mismo autor destaca la poca
información disponible. Por su parte, Medem (1958 y 1981) publicó algunos datos
sobre la reproducción de las especies del género Paleosuchus e indicó relatos y
anécdotas de nidos hallados en localidades de la Amazonía colombiana, durante la

78
el apareamiento y el
período de ovoposición se extienden a lo largo del año, con excepción del período
máximo de lluvia, cuando los nidos no pueden ser construidos en las áreas
inundables Vásquez-Ruesta (1983) halló un nido de P. trigonatus en temprano
estado de desarrollo, en septiembre de 1978 en un tributario de la cuenca del río
Ucayali, en la Amazonía peruana, durante la época seca. Rivas et al., (2001)
encontraron un nido de P. trigonatus el 30 de septiembre de 1999 y el nacimiento
ocurrió cerca del 27 de noviembre, durante un segundo período de lluvia.
La información obtenida de los registros y observaciones propias, indica que la
reproducción de esta especie en la CRE no se concentra en una temporada en
particular, sino que ocurre a lo largo del año, abarcando las temporadas de lluvia y
sequía (ver tabla 10 y figura 27). Este hecho no ha sido registrado para otra especie de
cocodriliano. Los cocodrilianos comúnmente concentran su esfuerzo reproductivo en
determinadas temporadas, según las característica de la nidada (hoyo o montículo) y
del hábitat (e. g. Crocodylus intermedius, construye sus nidos en la temporada seca
cuando las playas y bancos de arena están expuestos y Caiman c. crocodilus en la
lluviosa, en sitios sobre la línea de inundación).
Las características de P. trigonatus: tamaño pequeño, crecimiento lento, bajo
número de huevos por nidada, sumado a la considerable homogeneidad del hábitat
(temperatura y humedad), pudiera explicar la no concentración de la actividad
reproductiva en periodos determinados. Este planteamiento está apoyado por
afirmaciones fehacientes hechas por miembros de las comunidades ye kwana y
sanema de la CRE y forma parte del conocimiento tradicional de ambos grupos
humanos, principales usuarios de la especie en esa cuenca (Castillo et al., 2015).
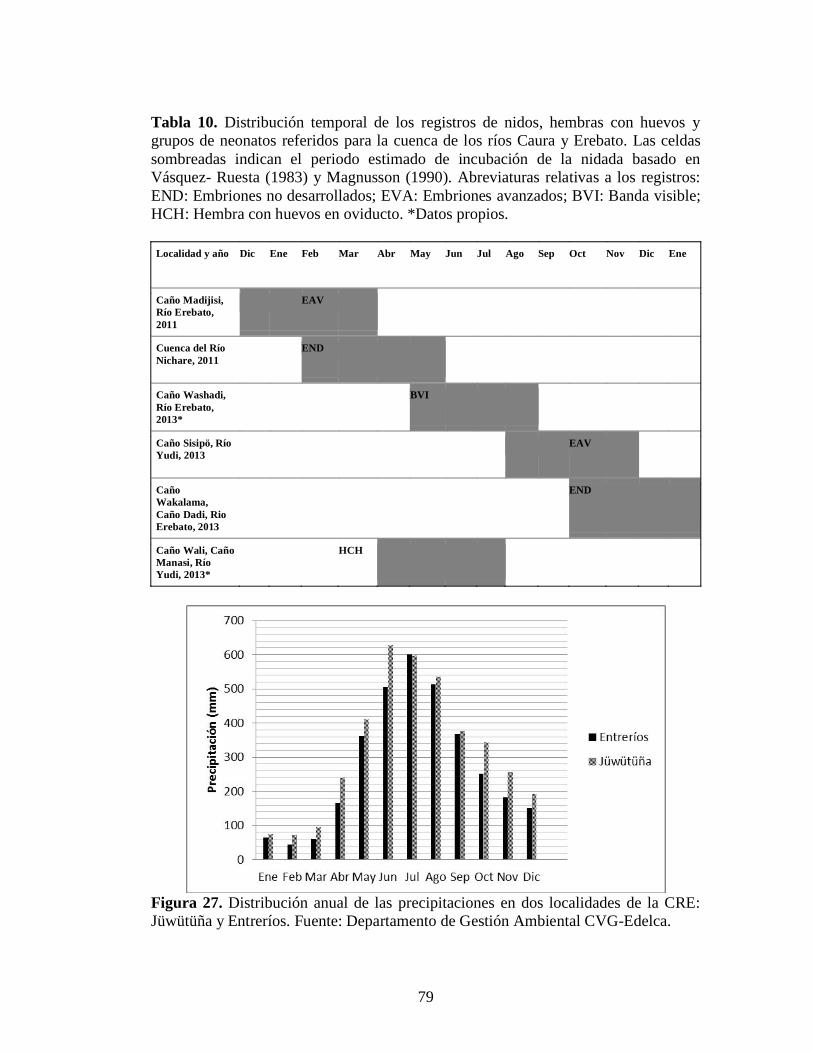
79
Tabla 10. Distribución temporal de los registros de nidos, hembras con huevos y grupos de neonatos referidos para la cuenca de los ríos Caura y Erebato. Las celdas sombreadas indican el periodo estimado de incubación de la nidada basado en Vásquez- Ruesta (1983) y Magnusson (1990). Abreviaturas relativas a los registros: END: Embriones no desarrollados; EVA: Embriones avanzados; BVI: Banda visible; HCH: Hembra con huevos en oviducto. *Datos propios. Localidad y año Dic Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic Ene
Caño Madijisi, Río Erebato, 2011
EAV
Cuenca del Río Nichare, 2011 END
Caño Washadi, Río Erebato, 2013*
BVI
Caño Sisipö, Río Yudi, 2013 EAV
Caño Wakalama, Caño Dadi, Rio Erebato, 2013
END
Caño Wali, Caño Manasi, Río Yudi, 2013*
HCH
Figura 27. Distribución anual de las precipitaciones en dos localidades de la CRE: Jüwütüña y Entreríos. Fuente: Departamento de Gestión Ambiental CVG-Edelca.
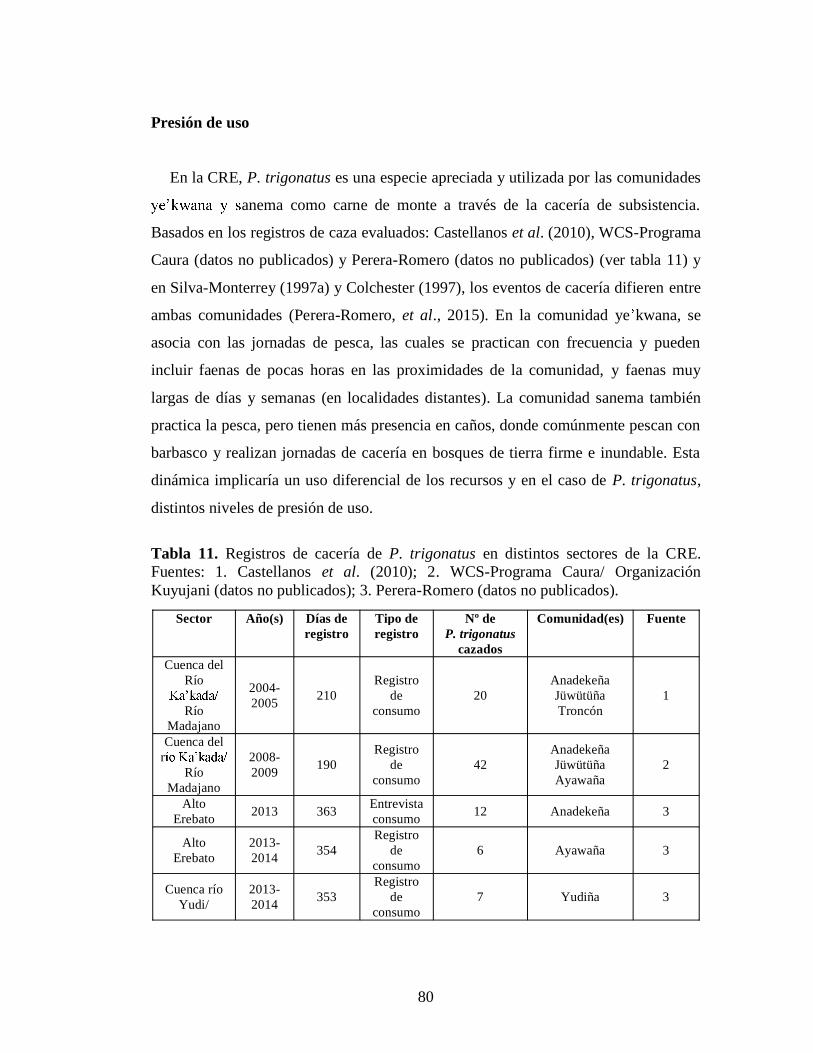
80
Presión de uso
En la CRE, P. trigonatus es una especie apreciada y utilizada por las comunidades
anema como carne de monte a través de la cacería de subsistencia.
Basados en los registros de caza evaluados: Castellanos et al. (2010), WCS-Programa
Caura (datos no publicados) y Perera-Romero (datos no publicados) (ver tabla 11) y
en Silva-Monterrey (1997a) y Colchester (1997), los eventos de cacería difieren entre
ambas comunidades (Perera-Romero, et al., 2015). En la comunidad ye kwana, se
asocia con las jornadas de pesca, las cuales se practican con frecuencia y pueden
incluir faenas de pocas horas en las proximidades de la comunidad, y faenas muy
largas de días y semanas (en localidades distantes). La comunidad sanema también
practica la pesca, pero tienen más presencia en caños, donde comúnmente pescan con
barbasco y realizan jornadas de cacería en bosques de tierra firme e inundable. Esta
dinámica implicaría un uso diferencial de los recursos y en el caso de P. trigonatus,
distintos niveles de presión de uso.
Tabla 11. Registros de cacería de P. trigonatus en distintos sectores de la CRE. Fuentes: 1. Castellanos et al. (2010); 2. WCS-Programa Caura/ Organización Kuyujani (datos no publicados); 3. Perera-Romero (datos no publicados).
Sector Año(s) Días de registro
Tipo de registro
Nº de P. trigonatus
cazados
Comunidad(es) Fuente
Cuenca del Río
Río Madajano
2004-2005 210
Registro de
consumo 20
Anadekeña Jüwütüña Troncón
1
Cuenca del r
Río Madajano
2008-2009 190
Registro de
consumo 42
Anadekeña Jüwütüña Ayawaña
2
Alto Erebato 2013 363 Entrevista
consumo 12 Anadekeña 3
Alto Erebato
2013-2014 354
Registro de
consumo 6 Ayawaña 3
Cuenca río Yudi/
2013-2014 353
Registro de
consumo 7 Yudiña 3

81
Entrevistas Las entrevistas a 58 miembros de las comunidades informaron la caza de 116 P.
trigonatus, 82 en 2013 y 34 hasta junio de 2014, en 33 localidades dentro de los cinco
sectores evaluados de la CRE y dos localidades de la cuenca del río Caura,
adyacentes a la comunidad Entreríos. Las comunidades donde se realizaron las
entrevistas, el número de P. trigonatus cazados por comunidad y las localidades, se
resumen en las tablas 12 y 13.
Tabla 12.Número de entrevistas y de P. trigonatus cazados en la CRE.
Comunidad Nº de entrevistados Nº de P. trigonatus cazados
Jüwütüña 8 18
Anadekeña 8 6
Dakudakuña 2 4
Sayusodiña 5 20
Entreríos 9 31
Madoña 2 0
Troncón 1 0
Campamento Ratón 2 4
Ayawaña 2 6
Yudiña 19 27
Total 58 116
Los entrevistados de la comunidad Sayusodiña informaron el mayor número de P.
trigonatus cazados con relación a la cantidad de entrevistados. Otra comunidad donde
se dio caza a un número significativo de P. trigonatus fue Entreríos, en localidades de
la cuenca del río Caura, próximas a la comunidad. Las localidades con mayores
informes de P. trigonatus cazados fueron: caño Sudumo (cuenca río Caura), caño
Wakalama (sector Yudi), caño Washadi (Erebato medio), caño Jasa y Madudaimö
(cuenca río Caura) (ver Tabla 12).
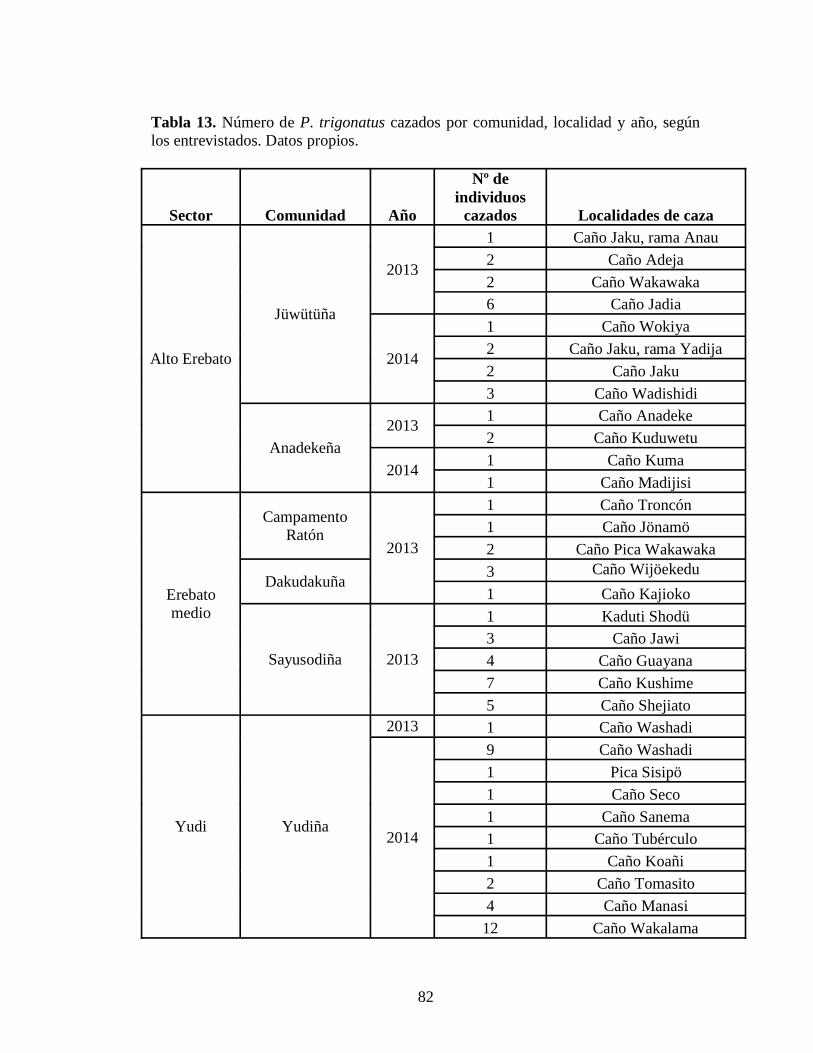
82
Tabla 13. Número de P. trigonatus cazados por comunidad, localidad y año, según los entrevistados. Datos propios.
Sector Comunidad Año
Nº de individuos
cazados Localidades de caza
Alto Erebato
Jüwütüña
2013
1 Caño Jaku, rama Anau 2 Caño Adeja 2 Caño Wakawaka 6 Caño Jadia
2014
1 Caño Wokiya 2 Caño Jaku, rama Yadija 2 Caño Jaku 3 Caño Wadishidi
Anadekeña 2013 1 Caño Anadeke
2 Caño Kuduwetu
2014 1 Caño Kuma 1 Caño Madijisi
Erebato medio
Campamento Ratón
2013
1 Caño Troncón 1 Caño Jönamö 2 Caño Pica Wakawaka
Dakudakuña 3 Caño Wijöekedu 1 Caño Kajioko
Sayusodiña 2013
1 Kaduti Shodü 3 Caño Jawi 4 Caño Guayana 7 Caño Kushime 5 Caño Shejiato
Yudi Yudiña
2013 1 Caño Washadi
2014
9 Caño Washadi 1 Pica Sisipö 1 Caño Seco 1 Caño Sanema 1 Caño Tubérculo 1 Caño Koañi 2 Caño Tomasito 4 Caño Manasi
12 Caño Wakalama

83
Tabla 13. Continuación.
Sector Comunidad Año Nº de individuos
cazados
Localidades de caza
Bajo Erebato/ Cuenca del Río Caura
Entreríos 2013 9 Caño Jasa 9 Caño
Madudaimö 12 Caño Sudumo
El bajo número de individuos cazados en la comunidad Yudiña (la mayor
comunidad de la cuenca del río Yudi), puede estar relacionado con la poca presencia
de cazadores durante este periodo, debido a los constantes viajes de éstos a las minas
del río Yuduani (río Caura) o, como comentan los entrevistados, a la intensa cacería
hace difícil
encontrar los Iatamö- Yadiwe P.
trigonatus ocurrida en febrero en el caño Wakalama, cabecera del río Yudi. Según
relatos de los entrevistados, eran muy fáciles de cazar. Un mínimo estimado de 12
individuos fue cazado, en su mayoría con máscara de buceo y lanzas bajo el agua
(algunos entrevistados refirieron que el número era mayor).
La técnica de caza y peca con máscaras de buceo, aunque no es tradicional, es
popular entre los miembros de las comunidades, quienes comúnmente pescan de este
modo a las aimaras (Hoplias aimara), pero no había sido referida para la caza de P.
trigonatus. Hames (1979) señaló que la incorporación de nuevas técnicas e
instrumentos de caza en
cacería. Sin embargo, el costo que implica la obtención de ésta hace que los
cazadores tengan que ejercer otras actividades económicas que les permita su
adquisición. Los sanemas por su parte, tienen menos acceso a estas tecnologías, por
lo que sus jornadas de cacería y pesca son más frecuentes e intensivas (op. cit.).
Los entrevistados también mencionaron el aprovechamiento de un grupo de 9
neonatos durante el mes de marzo de 2014 (aguas bajas) en las cabeceras del caño
Washadi, CRE. El grupo de neonatos fue hallado mientras los entrevistados se
disponían a pescar con barbasco en un caño de cabecera. Un ejemplar fue preparado

84
, mientras eran
mantenidos en cautiverio.
Estas informaciones sugieren que, en las cabeceras del río Yudi, aún hay presencia
P. trigonatus. Es probable que estos episodios de
cacería tan intensivos pudieran perturbar a poblaciones locales de esta especie.
Aunque estas jornadas no sean comunes, la necesidad constante de procurar carne
puede hacer que nuevos grupos busquen visitar esa localidad.
Análisis de los registros de cacería
Durante 2008-2009, la mayoría de los ejemplares fueron cazados en caños y
desembocaduras (bocas) de caño: 84,4%, mientras que el resto: 15,6% en distintos
sectores del canal principal del río, en su zona baja. La alta proporción de P.
trigonatus cazados en caños o en su desembocadura refleja la tendencia registrada en
la CRK y en otros sectores de la CRE: P. trigonatus
observan con menores esfuerzos en caños que en ríos (ver Abundancia poblacional y
Esfuerzo de búsqueda).
La estructura de tamaños, basada únicamente en los ejemplares que fueron
medidos: LT, LRCp, LC u otra medida que permitiera estimar el tamaño real
aproximado, es similar a la obtenida durante los conteos nocturnos en distintos
sectores de la CRK (ver Figura 28). Los ejemplares entre los 34 y 45 cm de LRCp
fueron los más cazados. Sin embargo, también existen registros de ejemplares de gran
tamaño (LRCp mayor a 85 cm) cazados y una muerte accidental asociadas con el arte
de pesca (un ejemplar adulto murió ahogado en un tren de pesca). Esto refiere la
predominancia de la caza por un grupo de tamaño particular, aunque este hecho está
relacionado con la abundancia, detectabilidad y/o accesibilidad de los ejemplares sub-
adultos y no con una cacería selectiva a priori, como lo demuestra el amplio intervalo
de tamaños de los P. trigonatus cazados.
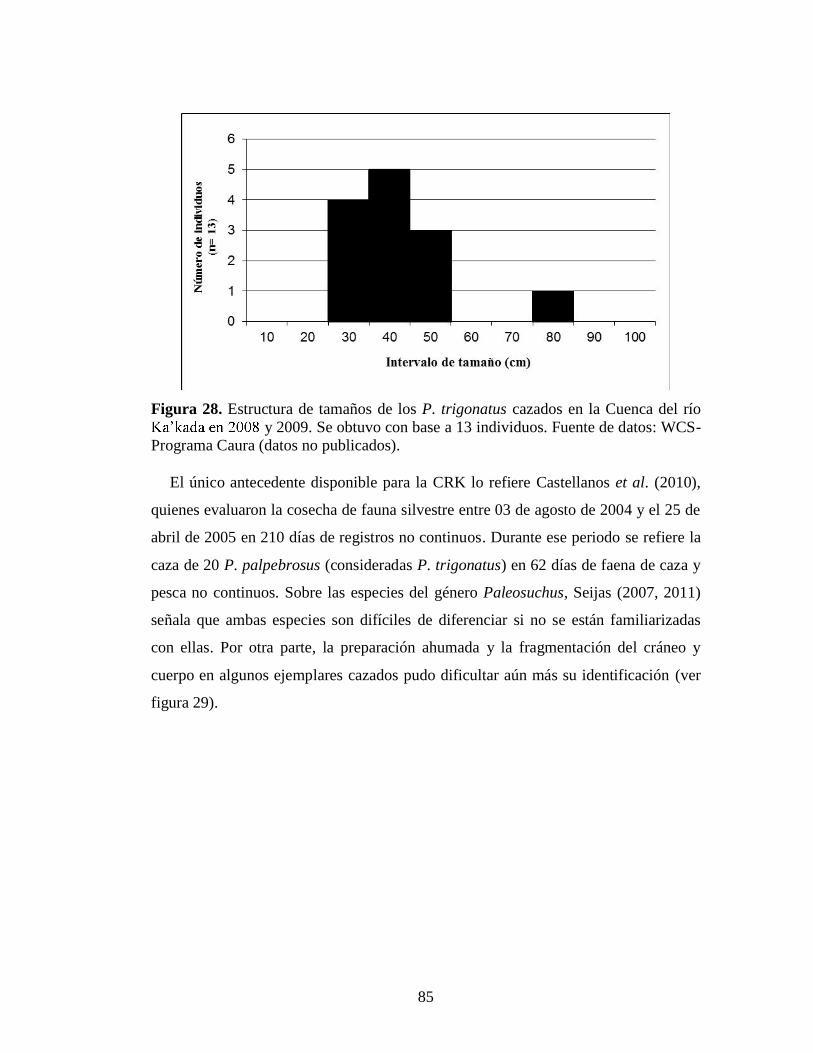
85
Figura 28. Estructura de tamaños de los P. trigonatus cazados en la Cuenca del río
y 2009. Se obtuvo con base a 13 individuos. Fuente de datos: WCS-Programa Caura (datos no publicados).
El único antecedente disponible para la CRK lo refiere Castellanos et al. (2010),
quienes evaluaron la cosecha de fauna silvestre entre 03 de agosto de 2004 y el 25 de
abril de 2005 en 210 días de registros no continuos. Durante ese periodo se refiere la
caza de 20 P. palpebrosus (consideradas P. trigonatus) en 62 días de faena de caza y
pesca no continuos. Sobre las especies del género Paleosuchus, Seijas (2007, 2011)
señala que ambas especies son difíciles de diferenciar si no se están familiarizadas
con ellas. Por otra parte, la preparación ahumada y la fragmentación del cráneo y
cuerpo en algunos ejemplares cazados pudo dificultar aún más su identificación (ver
figura 29).

86
Figura 29. Productos de cacería ahumados con piezas de P. trigonatus, registrados en
Al comparar los registros de cacería disponibles para la CRK puede apreciarse una
disminución leve en el número de ejemplares cazados con relación a las jornadas de
caza y pesca (Castellanos et al., 2010; WCS- Programa Caura, datos no publicados)
(ver tabla 14). Sin embargo, al comparar los ejemplares cazados considerando los
días de registro, se observa un aumento significativo en el número de jornadas de
caza y número de P. trigonatus cazados entre ambos periodos (ver tabla 15). La casi
totalidad de las jornadas de caza y pesca registradas, fueron realizadas por miembros
de las comunidades ye kwanas Jüwütüña y Anadekeña, ambas establecidas en el
sector Alto Erebato. Estas comunidades son las más numerosas de la CRE (Perera-
Romero et al., 2015) y poseen, al compararlas con las comunidades sanema, mayor
acceso a motores fuera de borda, escopetas y combustible, lo que implica los
miembros de estas comunidades tengan la posibilidad de visitar lugares remotos con
abundantes recursos de fauna silvestre y acuática, como el río Madajano y caño
Ajuju.
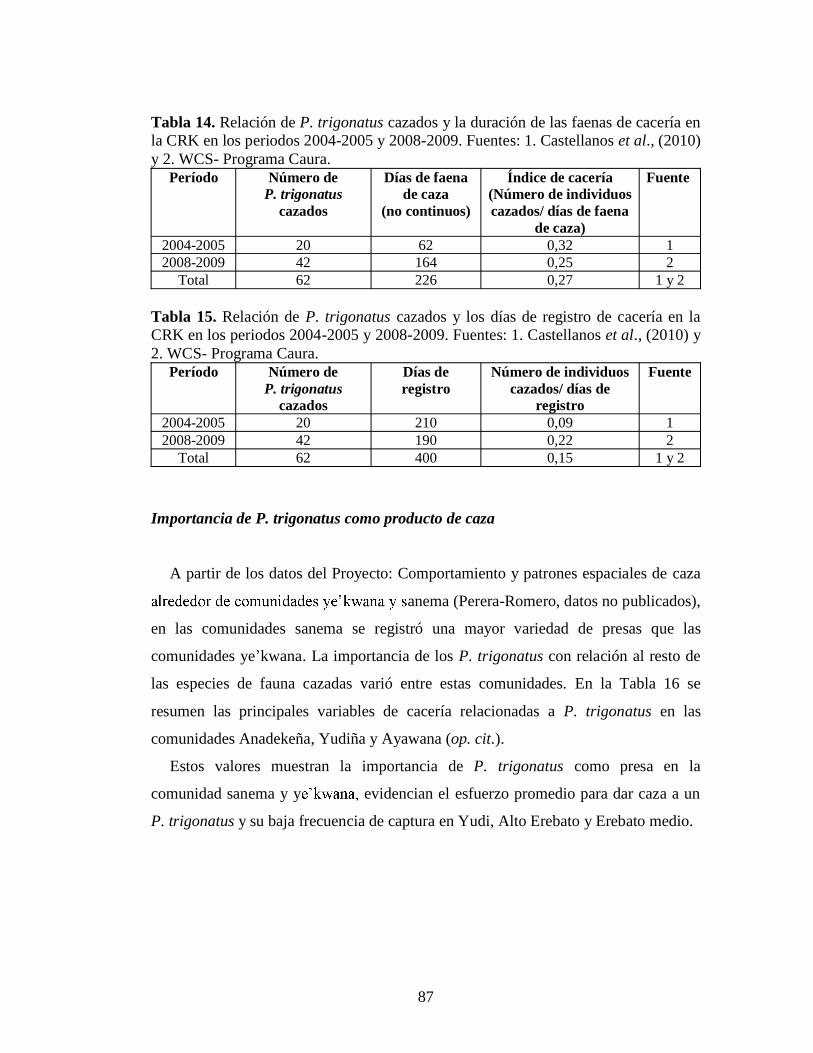
87
Tabla 14. Relación de P. trigonatus cazados y la duración de las faenas de cacería en la CRK en los periodos 2004-2005 y 2008-2009. Fuentes: 1. Castellanos et al., (2010) y 2. WCS- Programa Caura.
Período Número de P. trigonatus
cazados
Días de faena de caza
(no continuos)
Índice de cacería (Número de individuos cazados/ días de faena
de caza)
Fuente
2004-2005 20 62 0,32 1 2008-2009 42 164 0,25 2
Total 62 226 0,27 1 y 2 Tabla 15. Relación de P. trigonatus cazados y los días de registro de cacería en la CRK en los periodos 2004-2005 y 2008-2009. Fuentes: 1. Castellanos et al., (2010) y 2. WCS- Programa Caura.
Período Número de P. trigonatus
cazados
Días de registro
Número de individuos cazados/ días de
registro
Fuente
2004-2005 20 210 0,09 1 2008-2009 42 190 0,22 2
Total 62 400 0,15 1 y 2
Importancia de P. trigonatus como producto de caza
A partir de los datos del Proyecto: Comportamiento y patrones espaciales de caza
anema (Perera-Romero, datos no publicados),
en las comunidades sanema se registró una mayor variedad de presas que las
comunidades ye kwana. La importancia de los P. trigonatus con relación al resto de
las especies de fauna cazadas varió entre estas comunidades. En la Tabla 16 se
resumen las principales variables de cacería relacionadas a P. trigonatus en las
comunidades Anadekeña, Yudiña y Ayawana (op. cit.).
Estos valores muestran la importancia de P. trigonatus como presa en la
comunidad sanema y y evidencian el esfuerzo promedio para dar caza a un
P. trigonatus y su baja frecuencia de captura en Yudi, Alto Erebato y Erebato medio.

88
Tabla 16. Resumen de las principales variables de cacería relacionadas con P. trigonatus en comunidades de la CRE. Fuente: Perera-Romero (datos no publicados). Leyenda: 1Especies de vertebrados; 2Las especies cazadas fueron: P. trigonatus, Chelonoidis denticulata, Podocnemis sp. y Eunectes murinus. 3Números de P. trigonatus cazados por días de registro de cacería; 4Con relación al número de individuos cazados; 5Número de P. trigonatus cazados por jornadas de cacería. Comunidad Nº de sp.
cazadas1/ Número
sp. de reptiles2
Nº de ejemplares
cazados
Frecuencia de caza3
Importancia relativa4
(posición)
Esfuerzo de caza (horas/cazador/P.
trigonatus)
Tasa de captura5
Anadekeña
17/1 3006 0,07 5ta 39,9 0,07
Yudiña
23/3 97 0,08 3era 57,4 0,11
Ayawaña
24/4 209 0,04 8va 64,5 0,05
Los mayores esfuerzos de caza se registraron en Ayawaña y los menores en
Anadekeña. Esto puede estar relacionado con las características de la actividad de
cacería de cada comunidad. La cacería y pesca que practican las comunidades
ye kwana y sanema se diferencian en que las primeras tienen un mayor campo de
acción, debido al uso de curiaras con motor fuera de borda, escopetas y grupos de
caza mayores (familias completas), pero la cacería está casi siempre limitada a las
riberas y canal principal de los ríos y caños navegables. Por su parte, los sanema
viajan en curiaras a remo y a pie, en grupos más pequeños, por lo que no son
comunes las expediciones a lugares remotos, pero su cacería incluye el recorrido de
extensas zonas en las inmediaciones de sus comunidades, en áreas de tierra firme,
donde los P. trigonatus anidan. Esto podría tener efectos diferentes sobre las
poblaciones de esta especie.
La importancia relativa de P. trigonatus entre las presas identificadas (Perera-
Romero, datos no publicados) indican que es un recurso utilizado y apreciado por las
comunidades ye kwana y sanema, por lo que conviene hacer seguimiento a su caza
para evaluar su sostenibilidad a mediano y largo plazo. En toda su distribución
geográfica, P. trigonatus es una especie utilizada para su consumo de carne. Rivera
(2013) señala que los Matsé en la Amazonía peruana tienen preferencia por su carne
sobre otras especies de cocodrilos como C. c. crocodilus y Melanosuchus niger. Esto

89
también fue mencionado por Medem (1967) para comunidades indígenas en
Colombia.
En la CRE, la comunidad ye kwana comúnmente utiliza a P. trigonatus
considerando pautas relacionadas a tabúes alimentarios y creencias que condiciona el
uso diferencial de la especie, al igual que para el resto de las especies de caza (Silva-
Monterrey, 1997a). Cazadores con hijos recién nacidos o menores de 1 año, o con su
mujer embarazada, no cazan ni pueden tocar a un P. trigonatus, debido a que podría
traer consecuencias negativas para su hijo. Estas pautas fueron incluso aplicadas por
los Parabiólogos ye kwana que participaron en el proyecto, quienes en esos casos no
participaron en la captura ni manipulación de los P. trigonatus, lo que demuestra un
fuerte arraigo cultural. Para la comunidad ye kwana, los yadiwes poseen dueños
conocidos como Wiyu o gente de agua, que ejercen venganza sobre los hijos por
nacer o recién nacidos de quienes dan caza a un yadiwe. También hay lugares
sagrados donde está prohibida la caza como en los pozos profundos de caños y ríos.
Estas prácticas culturales son muy importantes y contribuyen con un manejo
tradicional de las especies de caza (Silva-Monterrey, 1997a), pero no se conocen
prácticas de este tipo en la cultura sanema.
Recomendaciones para la conservación y manejo de P. trigonatus en la cuenca del río Erebato
A continuación se enumeran algunas de las recomendaciones para la conservación
y manejo de las poblaciones de P. trigonatus en la CRE:
1. Establecer por consenso áreas donde la cacería de P. trigonatus sea permitida
y áreas en las cuales no: esto permitiría conservar la población de algunas
localidades importantes del río en su zona alta, media y baja, lo que
favorecerá el reclutamiento de individuos desde estos lugares hacia los
sectores afectados por la caza, comúnmente desocupados. Para la cuenca del

90
río Ka kada, se recomiendan incluir en la zona de protección en sector
Madajano: caño Ajuju y río Madajano (desde la cabecera hasta la confluencia
con el caño Ajuju); : caño Kaduaimüdü, Ñayudi,
Ama na, Kajioko, Widiki y Suajadito. La cacería podría ser permitida en el
canal principal del río Ka kada, y en algunos caños, hasta un máximo de 2Km
aguas adentro desde la desembocadura.
2. Evitar la cazar de los individuos con longitud total mayores a 90 cm o 60 cm
(LRCp) y excluir de la caza a neonatos. Esto permitiría que los animales
alcancen el tamaño reproductivo (establecida por Magnusson (1990) para las
hembras en 90 cm y para los machos en 100cm de LT).
3. No aprovechar ni alterar los nidos, ni a las hembras durante el proceso de
incubación de las nidadas. Los nidos y sus huevos constituyen la forma como
la población recluta nuevos individuos que permite la variación genética de la
misma. El discreto tamaño de la nidada hace que la pérdida constante de nidos
afecte a largo plazo el reclutamiento de nuevos miembros para una población.
La periodicidad de las posturas por parte de las hembras de P. trigonatus se ha
estimado en una por cada tres años (Magnusson, 1990) por lo que esta especie
posee un potencial reproductivo menor que la mayoría de las especies de
cocodrilianos neotropicales (Magnusson, Lima y Sampaio, 1991). En este
sentido, conviene proteger las localidades donde ya han sido encontrados o
aprovechados los nidos y las hembras, bajo el supuesto de fidelidad de sitios
de nidadas y porque ya habría evidencia que son hábitat idóneos para la
reproducción de la especie.
4. Apoyar las iniciativas de monitoreo de caza en las distintas comunidades: la
información de uso y ecológica obtenida es vital para la revisión y adecuación
de las pautas de manejo acordadas, así como para evaluar su viabilidad. En
este sentido conviene conservar el cráneo o algún hueso de los miembros:
húmero o fémur en buen estado para estimar con precisión el tamaño de cada
individuo cazado, además de la localidad, hora y fecha de la captura, biomasa

91
y medidas biométricas. Esto último sería en caso que en la comunidad esté
presente un monitor comunitario de uso de recursos o un parabiólogo.
sanema de la cuenca del río Caura han participado y
reflexionado sobre sus recursos y la forma de utilizarlos de manera sostenible (Silva-
Monterrey, Rodríguez y Castellanos, 2012). Esto a través de talleres participativos,
proyectos y asambleas comunitarias. Al respecto, han identificado problemas
asociados a la cacería de subsistencia como:
a. La poca abundancia de presas.
b. La caza de individuos muy pequeños, jóvenes y hembras.
c. La cacería reiterada en el mismo lugar.
d. Cazar animales preñados.
e. Utilizar las crías como mascotas.
f. No cumplir con los rituales tradicionales y tabúes alimenticios, entre
otros. (op. cit).
También han propuesto recomendaciones para el manejo de la cacería:
g. No ir de caza a los mismos lugares.
h. Cazar sólo lo necesario.
i. Cazar de forma selectiva, no cazar hembras ni animales muy jóvenes.
j. Proteger el hábitat de las presas.
k. Cumplir con los cantos sagrados, rituales y tabúes relacionados con la
cacería, entre otros. (op. cit).
Estas experiencias y nivel de conciencia comunitaria son elementos importantes
que pudieran servir de base para iniciativas de manejo y conservación de las especies
sujetas a cacería de subsistencia, entre ellas los yadiwes (iatamö). Sin embargo, en las
es algo opcional que
cada miembro decide o no cumplir, sin riesgo de ser juzgado o expulsado de su
comunidad o cultura. Un caso similar, ocurre actualmente con la participación de
muchos miembros de estas comunidades en actividades de minería ilegal de oro, a

92
pesar de las recomendaciones de ancianos y asambleas comunitarias de no participar
en la minería, ana y
sanema.
Taller participativo en la Comunidad Yudiña Durante el Taller participativo del Proyecto Yadiwe, realizado en la comunidad
Yudiña el 19 de junio de 2014, se reunieron un grupo de veintidós hombres y sus
familias en la churuata central de la comunidad. Allí, por espacio de dos horas se
presentaron los resultados obtenidos para la cuenca del río Yudi, que incluyeron las
entrevistas y el registro de cacería relativo a los P. trigonatus. Luego de la
presentación, varios ancianos opinaron sobre la necesidad de proteger a esta especie
para los jóvenes y niños de la comunidad. En consenso, se acordaron cuatro medidas
para la conservación y manejo de P. trigonatus (Iatama en lengua sanema) en la
cuenca del río Yudi:
1. Iatama sipönapiipia söti: No cazar hembras ovadas de P. trigonatus.
2. Iatama osewai söti: No cazar a las crías de P. trigonatus.
3. Iatama tadaka suati: No colectar los huevos de P. trigonatus.
4. Iatama pe epi a sösisa 1 metro: Cazar sólo a los P. trigonatus mayores a 1
metro de largo total.
La primera medida, sin embargo, es de difícil aplicación debido a la dificultad que
supone establecer con certeza si una hembra posee huevos en su interior a simple
vista. No se acordaron medidas para proteger a P. trigonatus a través de la
prohibición de la caza en determinadas localidades ni épocas del año. Las medidas
acordadas son un hecho trascendental al constituirse como las primeras medidas
conocidas que una comunidad indígena de Venezuela acuerda para la conservación y
manejo de P. trigonatus en su área de influencia. No obstante, es necesario el

93
seguimiento de la caza de P. trigonatus en la comunidad Yudiña para evaluar el éxito
de estos acuerdos y sus implicaciones reales para las poblaciones de P. trigonatus en
el mediano y largo plazo.
Aunque P. trigonatus está considerado a nivel global por la IUCN como
Preocupación menor (LC) y en la versión más reciente del Libro Rojo de la Fauna
Venezolana se mantiene en estatus Datos insuficientes (DD) (Rodríguez y Rojas-
Suárez, 2015), la caza de subsistencia permanente en algunas de las localidades de la
CRE y la probable contaminación por metilmercurio (aún no evaluada) representan a
nivel local, amenazas a mediano y largo plazo para esta especie de cocodriliano. En
consecuencia, conviene hacer seguimiento de su uso y de las poblaciones en los
sectores donde persisten año Ajuju (cuenca del río
Ka kada) y caño Wakalama (cuenca del río Yudi) y apoyar a las comunidades
indígenas en la consolidación de planes de conservación y manejo de ésta y otras
especies que son vitales para su subsistencia y la de su cultura en la cuenca del río
Erebato.

94
CAPÍTULO V
CONCLUSIONES Y RECOMENDACIONES
Conclusiones
En los sectores evaluados de la cuenca del río Erebato, incluido sus dos tributarios
más importantes: el río Ka kada y el río Yudi, sólo se registró una especie de
Crocodylia: P. trigonatus. Así mismo, no se registró la caza de otra especie en las
comunidades de la cuenca donde se evaluó ésta, entre 2008 y 2014. En la cuenca del
río Ka kada (Sector Madajano), el caño Ajuju es la localidad con mayor abundancia
relativa de P. trigonatus (2,05 ind/Km). Dentro de la cuenca se detectó la presencia
de la especie en 60% de las localidades evaluadas, de las cuales 90% corresponden a
caños. Los esfuerzos de búsqueda variaron entre 4,42 y 1 P. trigonatus observados
por hora de búsqueda en las localidades evaluadas.
La población de P. trigonatus en la cuenca del río Erebato está dominada por
individuos sub-adultos, con LRCp entre 30 y 39,9cm, con 41,5% de los registros
ubicados en esta clase de tamaños. No hubo diferencias significativas entre las
categorías de tamaño evaluadas y la posición que estas ocupaban dentro del hábitat,
pero si las hubo entre éstas y las variables del micro-hábitat: cobertura, tipo de cuerpo
de agua y tipo de agua, lo que indica una relación entre el tamaño corporal de los
ejemplares y los tipos de micro-hábitat que ocupan, con excepción del tipo de orilla.
La temporada de nidificación de P. trigonatus en la cuenca del río Erebato incluye
el periodo seco y de lluvia y las fases hidrológicas: aguas bajas, bajadas de aguas y
subidas de aguas. P. trigonatus construye nidos de tipo montículo en áreas no
inundables en bosques de tierra firme, cercanos a caños de aguas claras, generalmente

95
asociados con la base de un árbol con DAP mayor a 70 cm y a termiteros u
hormigueros activos.
El análisis de las fuentes de información utilizadas: Castellanos et al. (2010);
WCS-Programa Caura (datos no publicados) y Perera-Romero (datos no publicados)
y los registros propios evidencian la caza de P. trigonatus con motivo de subsistencia,
en todas las comunidades ye kwana y sanema de la cuenca del río Erebato visitadas.
Los intervalos de tamaños de los ejemplares cazados son muy variables entre los 38,2
y 81,3 cm de LRCp. También se registró el aprovechamiento de nidadas y neonatos,
por lo que la caza no es selectiva. Aunque P. trigonatus es apreciado por su carne, las
comunidades ye kwana consideran su caza sólo si tienen oportunidad, si lo avistan
durante sus jornadas nocturnas de caza y pesca. La comunidad sanema lo busca
comúnmente en los caños próximos y alejados de sus comunidades. El
aprovechamiento de las nidadas, aunque no es común, ocurre siempre que encuentran
un nido.
En la comunidad sanema de Yudiña, se acordaron en asamblea cuatro propuestas
para la conservación y manejo de P. trigonatus, basado en la cacería selectiva de
ejemplares adultos con LT mayores a 1 metro y la exclusión de las nidadas, neonatos,
hembras ovadas de los eventos de cacería.
Recomendaciones
Se recomienda:
1. Proponer, en el seno de cada comunidad, un plan de conservación y manejo
para P. trigonatus, basado en la propuesta de zonas de exclusión de caza y en
la aplicación de tallas mínimas para dar caza a un ejemplar.
2. La conservación de la cuenca alta del río Ka kada: caño Ajuju y río Madajano,
caño Washadi, caño Manasi (aguas arriba de Hoti Pola), caño Sudumo (río
Caura), caño Dadi, caño Sisipö y caño Jaku.
3. Cazar P. trigonatus sólo fuera de los lugares protegidos.

96
4. No colectar los huevos ni cazar a las hembras durante el proceso de cuidado
de la nidada y registrar de forma precisa su hallazgo para evaluar la selección
del hábitat reproductivo y temporalidad.
5. Registrar en cada comunidad todos los ejemplares cazados y las localidades
de procedencia a fin de hacer seguimiento a la caza y disponer de información
base para la adecuación de las medidas de conservación acordadas de forma
bianual.
6. Rescatar el conocimiento tradicional, leyendas y cuentos de las culturas
ye kwana y sanema referidos a P. trigonatus.
7. Estudiar lo relativo al ámbito de hogar, territorialidad, ciclo reproductivo,
dieta y concentración de metilmercurio de P. trigonatus en la cuenca del río
Erebato.
8. Establecer un plan de seguimiento anual para las poblaciones de P. trigonatus
en el caño Ajuju, Alto Manasi, caño Sudumo y caño Washadi.
9. Evaluar a las poblaciones de P. trigonatus en la cuenca del río Medewadi y
Bajo Caura.

97
REFERENCIAS BIBLIOGRÁFICAS
Amarasekare P. (2004). The role of density-dependent dispersal in source-sink
dynamics. Journal of Theoretical Biology, 226, 159-168. Consultado el 16 de marzo de 2012. Disponible en: http://www.eeb.ucla.edu/Faculty/amarasekare/papers/amarasekare_2004_JTB.pdf
Barrio-Amorós C. y C. Brewer-Carias (2008) Herpetological results of the 2002 expedition to Sarisariñama, a tepui in Venezuelan Guayana, with the description of five new species Zootaxa 1942. 68 pp.
Barrio-Amorós C., Brewer-Carias C. y O. Fuentes-Ramos. (2011). Aproximación preliminar a la herpetocenosis de un bosque pluvial en la sección occidental de la Sierra de Lema, Guayana venezolana. Rev. Ecol. Lat. Am., 16 (1), 1-46.
Botero-Arias, R. (2007). Padrões de movimiento, uso de microhábitat e dieta do Jacaré-Paguá, Paleosuchus palpebrosus (Crocodilia: Alligatoridae), em uma floresta de paleovárzea au sul do rio Solimões, Amazônia Central, Brasil. Dissertação para obtenção do título de Mestre em Ecologia INPA/UFAM, Manaus- Brazil.
Campos, Z., Muniz, F., Desbiez, A. y W. E. Magnusson (2016). Predation on eggs of Paleosuchus trigonatus (Schneider, 1807), by
armadillos and other predators. Natural history Vol. N°. pp. . Campos, Z. y T. Sanaiotti, (2006). Paleosuchus palpebrosus (Dwarf caimán) Nesting.
Herpetological Review, 37 (1), 81pp. Campos, Z., Sanaiotti, T., Marques, V. y W. E. Magnusson. (2015). Geographic
variation in clutch size and reproductive season of the Dwarf Caiman, Paleosuchus palpebrosus, in Brazil. Journal of Herpetology. Vol. 49. N°1. pp. 95-98.
Campos Z., Sanaiotti T., Muniz F., Farias I. y W. E. Magnusson. (2012). Parental
care in the dwarf caiman, Paleosuchus palpebrosus Cuvier, 1807 (Reptilia: Crocodilia: Alligatoridae). Journal of Natural History. Volume 46, Issue 47-48. pp. 2979-2984. Consultado el: 01 de marzo de 2016. Disponible en: http://www.tandfonline.com/doi/abs/10.1080/00222933.2012.724723.
Campos Z. y W. E. Magnusson. (2013). Thermal relations of Dwarf caiman,
Paleosuchus palpebrosus, in a hillside stream: evidence for an unusual thermal niche among crocodilians. Journal of Thermal Biology. 38. Pp. 20-23.

98
Campos Z., C. Zucco y G. Batista. (2007). Registro de ocorrência de Jacaré- paguá (Paleosuchus palpebrosus), na RPPN Engenheiro Eliezer Batista, Pantanal, Brasil. Corumbá: Embrapa. Comunicado Técnico 60. Ministério da Agricultura, Pecuária e Abastecimento. 4 p.
Castellanos H., Bertsch C., Veit A., Valeris C., Sarmiento W. y F. Rodríguez. (2010).
Cosecha de fauna silvestre y acuática por comunidades Ye kwana y Sanema del Alto río Caura. Simposio: Investigación y manejo de fauna silvestre en Venezuela en Homenaje al Dr. Juhani Ojasti. Caracas- Venezuela. 276 p.
Castillo, I., Perera-Romero, L., Bertsch, C., Espinoza, E., Sarmiento, W., Rodríguez,
O., Villasmil, I. 2015. Conocimiento tradicional de la ecología del Yadiwe (Paleosuchus trigonatus) en las comunidades indígenas del Río Erebato, Alto Caura. Boletín divulgativo Nº 10. Programa de Conservación de la Cuenca del Río Caura. Wildlife Conservation Society. Venezuela.
Colchester, M. (1997). La ecología social de los indígenas Sanema. Scientia
Guaianae. Nº 7: 111-140. En: Huber, O. y J. Rosales (Eds). (1997). Ecología de la cuenca del Río Caura, Venezuela. Nº 2. 473 pp.
Dias P. (1996). Sources and sinks in population biology. Tree, 11(8), 326-330.
Consultado el 16 de marzo de 2012. Disponible en: http://www.digeo.cl/asignaturas/ecologia/Dias%201996.pdf
Da Silveira R. y J. Thorbjarnarson. (1999). Conservation implications of commercial
hunting of black and spectacled caiman in the Mamirauá Sustainable Development Reserve, Brazil. Biological Conservation. Nº 88: 103-109.
Echevarría G., González N. y F. Mass. (2011). Aspectos reproductivos de la
comunidad de peces de la cuenca alta del río Caura, Venezuela. Ciencia, 19 (1), 25-40. Maracaibo- Venezuela. Consultado el 15 de noviembre de 2011. Disponible en: http://www.revistas.luz.edu.ve/index.php/cien/article/viewFile/7845/7511.
Elsey R., P. L. Trosclair y T. C. Glenn. (2008). Nest-site fidelity in American
alligators in a Lousiana coastal marsh. Southeastern Naturalist.Vol 7 (4):737 743. Espinosa-Blanco, A. y A. E. Seijas. (2010). Uso de hábitat entre crocodílidos en el
sistema del río Cojedes, Venezuela. Rev. Lat. Cons. Vol. 1 (2): 112 119. Fryxell J. (2001). Habitat suitability and source-sink dynamics of beavers. Journal of
Animal Ecology, 70, 310-316. Consultado el 16 de marzo de 2012. Disponible en: http://breb.ro/Publicatii/habitat_suitability_and_source.pdf
Godshalk R. (1982). The habitat and distribution of Paleosuchus in Venezuela. In:
Crocodiles: Proceedings of the 5th Working Meeting of the Crocodile Specialist

99
Group. (pp. 31- 38). International Union for Conservation of Nature and Natural Resources, Gland- Switzerland. Consultado el 15 de diciembre de 2011. Disponible en: http://books.google.co.ve/
Gorzula, S. y A. Paolillo. (1986). La ecología y el estado actual de los aligatóridos de
la Guyana venezolana. En: Crocodiles. Proceedings of the 7th Working Meeting of the IUCN/SSC Crocodile Specialist Group. (pp. 37- 54). IUCN/SSC. Gland- Switzerland.
Gorzula, S., Paolini, J. y J. Thorbjarnarson. (1987). Some hydrochemical and
hydrological characteristics of crocodilians habitats. Trop. Freshwat. Biol. 0795- 0101. pp. 50- 61.
Gorzula, S. y J. C. Señaris. (1998). Contribution to the herpetofauna of the
Venezuelan Guayana. I. A data base. Scientia Guaianae. Nº 8. 302pp. Hames, R. (1979) A comparison of the efficiencies of the shotgun and the bow in the
neotropical forest hunting. Human Ecology. Nº7, Vol. 3. pp. 219-252. Legendre, P. (1993). Spatial autocorrelation: trouble or new paradigm? Ecology. Vol.
74, No. 6, pp. 1659-1673. Consultado el 01 de febrero de 2016. Disponible en: http://www.jstor.org/stable/1939924.
Magnusson, W. E. (1986). The peculiarities of crocodiles population dynamics and
their possible importance for management strategies. In: Crocodiles: Proceedings of the 7th Meeting of Crocodile Specialist Group/ Species Survival Commission IUCN. (pp. 434- 442). IUCN. Gland, Switzerland.
Magnusson, W.E. (1990). Paleosuchus. En: Crocodiles, their Ecology, Management
and Conservation. (pp.101-109). A Special Publication of the Crocodile Specialist Group. IUCN. Gland, Switzerland.
-fronted Caiman
Paleosuchus palpebrosus. In Manolis S.C. and C. Stevenson (Eds.), Crocodiles. Status Survey and Conservation Action Plan. Third Edition. Crocodile Specialist Group (pp. 40-42). Darwin- Australia.
Magnusson, W.E., Campos, Z. (2010b). -fronted Caiman
Paleosuchus trigonatus. In Manolis S.C. and C. Stevenson (Eds.), Crocodiles. Status Survey and Conservation Action Plan. Third Edition. Crocodile Specialist Group (pp. 43-45). Darwin- Australia.
Magnusson W, A. Lima y R Sampaio (1985). Sources of heat for nest of Paleosuchus
trigonatus and a review of crocodilian nest temperatures. Journal of Herpetology, 19 (2), 199- 207.

100
Magnusson, W.E., Lima, A.P. (1991). The Ecology of a Criptic Predator, Paleosuchu strigonatus, in a Tropical Rainforest. Journal of Herpetology, 25(1), 41-48.
Martínez-Freire G. (2016). -
fronted caiman (Paleosuchus trigonatus) in protected habitats in Amazonia. Final report Student Research Assistance Scheme (SRAS) for International Union for Conservation of Nature (IUCN)-Crocodile Specialist Group (CSG). 14 p.
Medem, F. (1958). The crocodilian genus Paleosuchus. Fieldiana- Zoology. Chicago
Natural History Museum. 39 (21), 227-247. Medem, F. (1967). El género Paleosuchus en la Amazonia. Atas Simp. Biota
Amazonica. Vol. 3 (Limnología), 141-162. Medem, F. (1981). Los Crocodylia de Sur América. Los Crocodylia de Colombia.
Vol. I. Editorial Carrera 7a. Bogotá. Colombia. 354 p. Medem, F. (1983). Los Crocodylia de Sur América. Vol. II. Universidad Nacional de
Colombia. Bogotá. Colombia. 270 p. Muniz, F., Senna-Bittencourt, P., Pires-Farias, I., Hrbek, T. y Z. Campos. (2015).
New records on occurrence of Paleosuchus in the Branco River Basin, Roraima State, Brazil. Crocodile Specialist Group Newsletter, Vol. 34, N° 4. pp. 8-11.
Pacheco, A. (2009). Distribución, abundancia y estructura poblacional del Babo
Morichalero (Paleosuchus palpebrosus) en los Llanos Orientales del Estado Anzoátegui. Tesis de Licenciatura. Universidad Central de Venezuela, Caracas, Venezuela. p.102.
Pearcy A. (2011). Implications of skull shape for the ecology and conservation
biology of crocodiles. Doctoral Dissertation unpublished. Universiteit Leiden. p.78. Consultada el: 01 de marzo de 2012. Disponible en: https://openaccess.leidenuniv.nl/bitstream/handle/1887/17708/PhDDissertation12nieuwste%20versie.pdf?sequence=2
Perera-Romero, L. (2012). Manejo de fauna silvestre en comunidades indígenas del
Alto Caura. Acciones y logros del período 2007-2012 del Programa de Conservación de la Cuenca del Río Caura. Boletín divulgativo Nº 7. Wildlife Conservation Society. Venezuela.
Perera-Romero, L., Polisar, J. Maffei, L. (2015). Grandes vertebrados terrestres en
tierras ancestrales indígenas del alto Caura: importancia de los acuerdos comunitarios para la conservación del Escudo Guayanés venezolano. Capítulo 10. pp. 191-208. En: Payan, E., Lasso C. y Castaño-Uribe, C. (Editores). (2015). Conservación de Grandes vertebrados en áreas no protegidas de Colombia,

101
Venezuela y Brasil. Serie Editorial Fauna Silvestre Neotropical. Instituto de investigación de Recursos Biológicos Alexander von Humbolt (IAvH). Bogotá D. C., Colombia.
Perera-Romero, L. (Datos no publicados). Hunting behavior and hunting spatial
Society Research Fellowship- Year 2012. Perera-Romero, L., Palma, Z., (2012a). Construcción colectiva de medidas de manejo
de la caza y la pesca en la comunidad Asakositö uli (Ayawaña), Alto Caura. Relatoría de taller comunitario. Programa de Conservación de la Cuenca del Río Caura, Wildlife Conservation Society.
Perera-Romero, L., Palma, Z., (2012b). Construcción colectiva de medidas de manejo
de la caza y la pesca en la comunidad Anadekeña, Alto Caura. Relatoría de taller comunitario. Programa de Conservación de la Cuenca del Río Caura, Wildlife Conservation Society.
Perera-Romero, L., Palma, Z., (2012c). Construcción colectiva de medidas de manejo
de la caza y la pesca en la comunidad Jüwütüña, Alto Caura. Relatoría de taller comunitario. Programa de Conservación de la Cuenca del Río Caura, Wildlife Conservation Society.
Pulliam H. R. (1988). Sources, sinks, and population regulation. The American
Naturalist, 132(5), 652-661. Consultado el: 16 de marzo de 2012. Disponible en: http://www.montana.edu/hansen/documents/labreadings2011/Pulliam%201988.pdf
Reig, O. (1983). Estado actual de la teoría de la formación de las especies animales.
En: Aguilar, P (Eds). Informe final del IX Congreso Latinoamericano de Zoología. Arequipa, Perú. pp. 37-57. 9 al 15 de octubre 1983.
Rivas G., C. Molina, G. Ugueto, T. Barros, C. Barrio-Amorós y P. Kok. (2012).
Reptiles of Venezuela: an updated and commented cheklist. Zootaxa 3211. pp. 1-64. Consultado el 16 de marzo de 2012. Disponible en: http://www.terrario.org.ve/Rivasandet2012.ReptilesofVenezuelaanupdatedandcommentedchecklist.pdf.
Rivas, J.A., Aktay, S.A., Owens, R.Y. (2001). Paleosuchus trigonatus
smooth-fronted caiman) nesting. Herpetological Review 32 (4), 251.
Rivera D. (2013). Influencia del aprovechamiento de subsistencia y variables ambientales sobre tres especies de caimanes en la Comunidad Nativa Matsés y la Reserva Nacional Matsés en Loreto, Perú. Trabajo de Grado no publicado. Universidad Nacional- Costa Rica. Heredia, Costa Rica.

102
Robertson, G.P. (2008). GS+: Geostatistics for the Environmental Sciences. Gamma Design Software, Plainwell, Michigan USA.
Rodríguez J. P. y F. Rojas-Suárez (Eds.). (2008). Libro rojo de la fauna venezolana.
Caracas: Provita- Shell. Rodríguez J. P. y F. Rojas-Suárez (Eds.). (2015). Libro rojo de la fauna venezolana.
Caracas: Provita- Shell. Ross, J.P. (1998). Crocodiles: Status Survey and Conservation Action Plan. IUCN
Publications. Gland, Switzerland. Rueda-Almonacid J., J. Carr, R. Mittermeier, J. Rodríguez- Mahecha, R. Mast, R.
Vogt, A. Rhodin, J. de la Ossa- Velásquez, J. Rueda y C. Goettsch Mittermeier. (2007). Las tortugas y los cocodrilianos de los países andinos del Trópico. Serie de guías de campo N°6. Conservación Internacional. Editorial Panamericana, Formas e Impresos. Bogotá, Colombia. 538 p.
Seijas, A. (1986). Situación actual de las poblaciones de babas y babillas (Caiman
crocodilus) en la región norte-costera de Venezuela. Informe Técnico del Servicio Nacional de Fauna Silvestre. Ministerio del Ambiente y de los Recursos Naturales Renovables. (MARNR). Maracay, Venezuela.
Seijas, A. (1988). Habitat use by the American crocodile and the spectacled caiman
coexisting along the Venezuelan coastal region. Tesis de Doctorado no publicada. University of Florida, Miami.
Seijas, A. (1996). Coexistencia de babas y caimanes en la región costera venezolana.
En: Péfaur, E. (1997) (Comp.). Herpetología Neotropical, Volumen II, (pp. 217- 253). Universidad de Los Andes. Mérida, Venezuela.
Seijas A. (2007). Características distintivas y estado del conocimiento de las especies
del género Paleosuchus (Crocodylia; Alligatoridae) en Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales, 166, 29-46. Consultado el 11 de noviembre. Disponible en: http://www2.scielo.org.ve/pdf/mem/v66n166/art03.pdf.
Seijas A. (2011). Los Crocodylia de Venezuela: Ecología y conservación. Colección
Estudios Divulgación Científica y Tecnológica. Academia de Ciencias Físicas, Matemáticas y Naturales. Venezuela.
Señaris J. C., C. A. Lasso y A. L. Flores (eds). (2008). Evaluación Rápida de la
Biodiversidad de los Ecosistemas Acuáticos de la Cuenca Alta del río Paragua, Estado Bolívar, Venezuela. RAP Bulletin of Biological Assessment 49. Conservation International, Arlinton, VA, USA.

103
Silva-Monterrey, N. (1997a Scientia Guaianae. Nº7: 65-84. En: Huber, O. y J. Rosales (Eds). (1997). Ecología de la cuenca del Río Caura, Venezuela. Nº 2. 473 pp.
Silva-Monterrey, N. (1997b). Utilización alimentaria de los recursos naturales entre . Scientia Guaianae. Nº7: 85-107. En: Huber, O. y J. Rosales (Eds).
(1997). Ecología de la cuenca del Río Caura, Venezuela. Nº 2. 473 pp. Silva-Monterrey, N. Rodríguez A. y H. Castellanos (Eds). (2012). Pautas para el
. Forest People Programme. 126 pp.
Thorbjarnarson, J. (Comp.). (1992). Crocodiles. An action plan for their conservation. International Union for Conservation of Nature and Natural Resources (IUCN). Gland- Switzerland.
Valeris C., Perera-Romero L., Jasatao R., Asatali M. y H. Castellanos. (2014). Primer registro de anidación de Paleosuchus trigonatus (Crocodylia: Alligatoridae) para Venezuela. Bol. Acad. C. Fís. Mat. y Nat. Vol. LXXIV Nº. 2: 9-14.
Vásquez-Ruesta, P. G. (1983). Descripción del desarrollo embrionario de
Paleosuchus trigonatus Schneider en Requena, Loreto. Revista Forestal del Perú. Vol. 11 (1-2): 1-6.
Velasco, A. (2005). La conservación de los cocodrilos en América Latina y el Caribe.
En: Reunión Regional de América Latina y el Caribe del Grupo de Especialistas en Cocodrilos (CSG/SSC/IUCN) 17 al 20 de mayo 2005, Santa Fe, Argentina. pp. 75- 87. Consultado el 28 de mayo de 2012. Disponible en: http://www.iucncsg.org/365_docs/attachments/protarea/Regi-b60dee69.pdf
Verdade, L. (1996). The influence of hunting pressure on the social behavior of
vertebrates. Rev. Brasil. Biol., 56 (1), 1-13. Consultada el 01 de marzo de 2012. Disponible en: http://www.esalq.usp.br/docentes/lea/Artigos_pdf/Verdade%201996.PDF
Villamarín F., G. Freire, G. Da Silva, J. Lacerda, B. Marioni, A. Nobre, M. Ribeiro,
E. Silva, A. Gómez y R. Da Silveira. (2010). Monitoring the Paleosuchus trigonatus population of a large brazilian amazon urban forest fragment. 20th Working Meeting of the Crocodile Specialist Group IUCN/SSC.Manaus- Brazil.
WCS-Programa Caura. (2009). Informe del Componente de Ecosistemas Terrestres.
Ciudad Guayana: Valeris C. y Veit A.

104
Webb, G., Manolis, S. C.y Sack, G. C. (1983). Crocodylus johnstoni and C. porosus coexisting in a Tidal River. Australian Wildlife Research 10, 639 650.
White J y Rivas J. (2003). Paleosuchus trigonatus (Dwarf Caiman). Neonate time
budget. Herpetological Review, 34 (2), 141.

105
Apéndice 1. Modelo de entrevista para evaluar el uso consuntivo y aspectos reproductivos de P. trigonatus en comunidades de la Cuenca del Río Erebato. ____________________________________________________________________ Fecha: Localidad/ Comunidad: Entrevistado: Edad (años): Sexo: Entrevistador:
1. ¿Ha cazado yadiwes durante este año? Si: _______ No:_______ 1.1. ¿Cuántos? 1.2. ¿Dónde? 1.3. ¿Cómo era el hábitat? Río, Bosque de tierra firme u otro. 1.4. ¿Recuerda el sexo y tamaño? 1.5. ¿Cómo lo cazó? ¿Qué armas utilizó? 1.6. ¿Cómo lo preparó? Asado, cocido, u otro.
2. ¿Ha visto nidos de yadiwe? Si: _______ No:_______ 2.1. ¿Dónde y cuándo? 2.2. ¿Cuántos huevos encontró en el nido?
3. ¿Ha visto crías de yadiwe? ¿en qué mes? 4. ¿En qué lugares hay más yadiwes? ¿Por qué considera que hay muchos?
5. ¿En qué lugares hay pocos yadiwes? ¿Por qué considera que hay pocos?
6. ¿Algún comentario adicional?
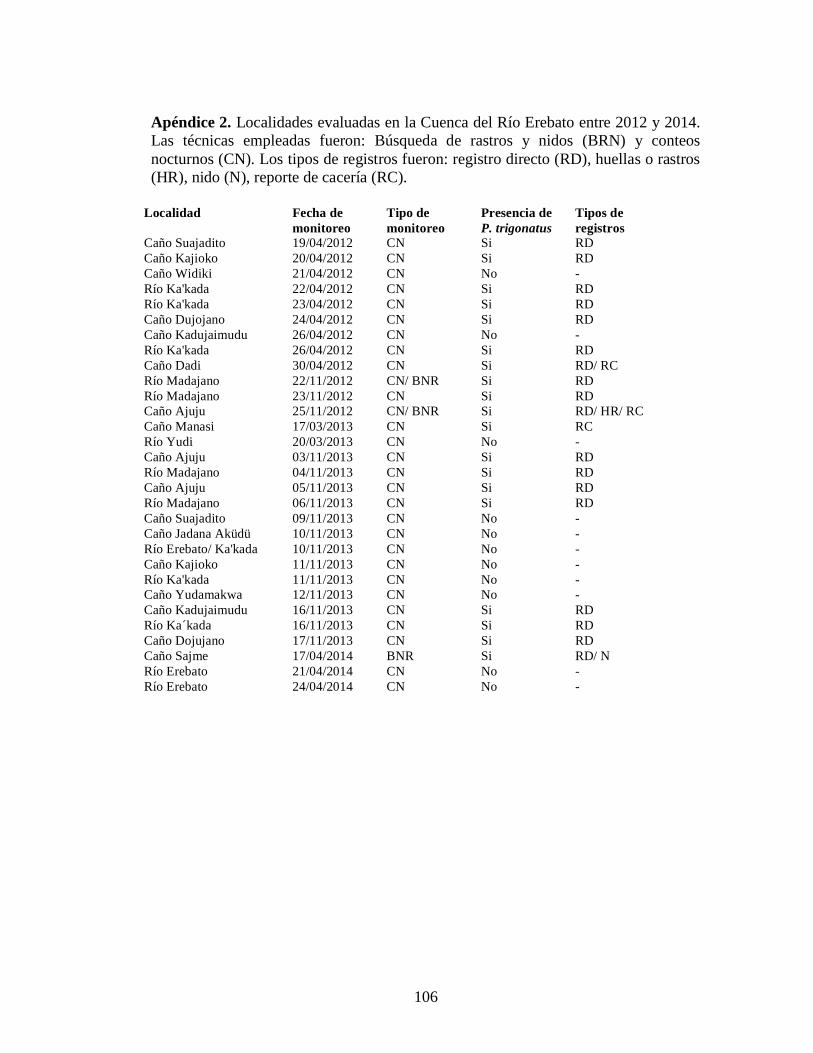
106
Apéndice 2. Localidades evaluadas en la Cuenca del Río Erebato entre 2012 y 2014. Las técnicas empleadas fueron: Búsqueda de rastros y nidos (BRN) y conteos nocturnos (CN). Los tipos de registros fueron: registro directo (RD), huellas o rastros (HR), nido (N), reporte de cacería (RC).
Localidad Fecha de monitoreo
Tipo de monitoreo
Presencia de P. trigonatus
Tipos de registros
Caño Suajadito 19/04/2012 CN Si RD Caño Kajioko 20/04/2012 CN Si RD Caño Widiki 21/04/2012 CN No - Río Ka'kada 22/04/2012 CN Si RD Río Ka'kada 23/04/2012 CN Si RD Caño Dujojano 24/04/2012 CN Si RD Caño Kadujaimudu 26/04/2012 CN No - Río Ka'kada 26/04/2012 CN Si RD Caño Dadi 30/04/2012 CN Si RD/ RC Río Madajano 22/11/2012 CN/ BNR Si RD Río Madajano 23/11/2012 CN Si RD Caño Ajuju 25/11/2012 CN/ BNR Si RD/ HR/ RC Caño Manasi 17/03/2013 CN Si RC Río Yudi 20/03/2013 CN No - Caño Ajuju 03/11/2013 CN Si RD Río Madajano 04/11/2013 CN Si RD Caño Ajuju 05/11/2013 CN Si RD Río Madajano 06/11/2013 CN Si RD Caño Suajadito 09/11/2013 CN No - Caño Jadana Aküdü 10/11/2013 CN No - Río Erebato/ Ka'kada 10/11/2013 CN No - Caño Kajioko 11/11/2013 CN No - Río Ka'kada 11/11/2013 CN No - Caño Yudamakwa 12/11/2013 CN No - Caño Kadujaimudu 16/11/2013 CN Si RD Río Ka´kada 16/11/2013 CN Si RD Caño Dojujano 17/11/2013 CN Si RD Caño Sajme 17/04/2014 BNR Si RD/ N Río Erebato 21/04/2014 CN No - Río Erebato 24/04/2014 CN No -

107
RESEÑA CURRICULAR
Carlos Eduardo Valeris Chacín nació en Maracaibo, estado Zulia, Venezuela, el 03 de junio de 1981. Es hijo de Wintila Chacín de Valeris y Rafael Valeris. Sus estudios de educación básica, secundaria y diversificada los realizó en la Unidad Educativa
tulo de Bachiller en Ciencias en 1998. En 1999 inicia sus estudios en la Universidad del Zulia (LUZ), donde cursó la Licenciatura en Biología, culminando en 2007, con la presentación del trabajo especial de grado: Estado poblacional, distribución y hábitat del Caimán de la costa (Crocodylus acutus) en el río Santa Rosa, Zulia, Venezuela, obteniendo la mención honorífica. Durante sus estudios ejerció el cargo de Preparador docente en la asignatura Zoología de Vertebrados desde el año 2004 al 2006. También se desempeñó como asistente ad honorem en el Museo de Biología de la Universidad del Zulia (MBLUZ) donde colaboró en las secciones de Herpetología y Ornitología. En 2007 participó como asistente de campo con Idaho Bird Observatory (IBO) - Boise State University (BSU) y Hagerman Fossils Beds National Monument (National Park Service) en Estados Unidos. En 2008 se une al Programa Caura de Wildlife Conservation Society (WCS), donde se desempeñó como investigador de campo del componente de ecosistemas terrestres, que incluyó la formación de
Venezuela. Desde 2009 ejerce el cargo de docente-investigador en la Universidad Nacional Experimental de Guayana (UNEG) y está adscrito al Centro de Investigaciones Ecológicas de Guayana (CIEG) en la Línea de investigación: Ecología y Manejo de fauna. En la UNEG ha participado como profesor en las asignaturas: Fundamentos Bio-ecológicos, Zoología Agrícola, Biología y Excursionismo Ecológico. En 2016 culmina la Maestría en Ciencias Ambientales- mención: Ecología Aplicada, en la UNEG, con el trabajo de grado: Estado poblacional, distribución geoespacial, aspectos reproductivos y uso consuntivo de Paleosuchus trigonatus (Crocodylia: Alligatoridae) en la cuenca del río Erebato, Alto Caura, Venezuela, obteniendo mención publicación. Este proyecto fue financiado por WCS, el Grupo de Especialistas en Cocodrilos de la Unión Internacional para la Conservación de la Naturaleza (UICN) y Small Grant Rufford Foundation. Desde 2014 participa como investigador principal en el proyecto: Ecología y conservación del Semillero de carrizales (Amaurospiza carrizalensis Lentino y Restall, 2003) en la cuenca baja del río Caroní, Venezuela, financiado por The Mohamed bin Zayed Species Conservation Fund, Neotropical Bird Club y Auckland Zoo. En 2016 obtiene una beca de la Comisión Nacional de Investigación Científica y Tecnológica (CONICYT) del Gobierno de Chile, para estudiar el Doctorado en Ciencias- mención: Ecología y Biología Evolutiva en la Universidad de Chile.











