Embed Size (px)
Citation preview

This article was downloaded by: [University of Illinois at Urbana-Champaign]On: 28 September 2013, At: 10:35Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Freshwater EcologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tjfe20
Relationships betweenmacroinvertebrate communities andland use types within different riparianwidths in three headwater streams ofTaizi River, ChinaYuan Zhang a , Rui Zhao a , Weijing Kong a , Shiwei Geng a ,Catherine N. Bentsen a & Xiaodong Qu aa State Key Laboratory of Environmental Criteria and RiskAssessment and Laboratory of Riverine Ecological Conservationand Technology, Chinese Research Academy of EnvironmentalSciences , Beijing , 100012 , P.R. ChinaPublished online: 22 May 2013.
To cite this article: Yuan Zhang , Rui Zhao , Weijing Kong , Shiwei Geng , Catherine N. Bentsen& Xiaodong Qu (2013) Relationships between macroinvertebrate communities and land use typeswithin different riparian widths in three headwater streams of Taizi River, China, Journal ofFreshwater Ecology, 28:3, 307-328, DOI: 10.1080/02705060.2013.779941
To link to this article: http://dx.doi.org/10.1080/02705060.2013.779941
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Relationships between macroinvertebrate communities and land use
types within different riparian widths in three headwater streams
of Taizi River, China
Yuan Zhang, Rui Zhao, Weijing Kong, Shiwei Geng, Catherine N. Bentsen and
Xiaodong Qu�
State Key Laboratory of Environmental Criteria and Risk Assessment and Laboratory of RiverineEcological Conservation and Technology, Chinese Research Academy of Environmental Sciences,Beijing 100012, P.R. China
(Received 21 September 2012; final version received 1 February 2013)
The effects of riparian land uses on water quality and macroinvertebrate communitieswere examined in three headwater streams of the Taizi River in northeastern China.Land uses of forest, grassland, farmland, and villages were evaluated within differentwidths of riparian corridors (length ¼ 3 km upstream; width ¼ 25, 50, 100, 200, and500 m). Overall, the results revealed a significant trend for each stream linking a rela-tively low proportion of farmland in the riparian corridor with better water quality andhealthier macroinvertebrate community. Cluster analysis confirmed significant varia-tion in the macroinvertebrate communities found among the three streams. Correlationanalysis identified significant positive correlations between the proportion of farmlandand metrics of water quality (conductivity, dissolved solids, and total nitrogen) atsmall (25–50 m) to medium (100–200 m) widths of riparian corridors. The sensitivetaxa of stoneflies and functional feeding groups of scrapers and predators were goodpredictors of anthropogenic disturbance associated with the proportional extent offarmland and village land use across most of all riparian widths. In contrast, correla-tions between biodiversity indices and proportion of villages and between bioassess-ment indices and proportion of farmland were only significant at relatively smallwidths (25–50 m) of riparian corridors. The canonical correspondence analysisrevealed that the proportions of forest (25 m), farmland (200 m), grassland (200 m),and village (100 m) showed independent effects on the spatial distribution of macroin-vertebrates in the Taizi River. The first two axes contributed 34.6% of the cumulativepercent variance of species–environment relations. This research indicates thatmanagement of riparian corridor land uses at relatively small to medium widths(25–200 m) could be effective in restoring water quality and macroinvertebratecommunities in headwater stream regions.
Keywords: macroinvertebrates; water quality; riparian corridor; headwater stream;land use
Introduction
Changes in land use, from natural forest to urban developed or agriculture, have been
shown to cause profound modifications in stream ecosystems (Allan et al. 1997; Foley
et al. 2005; V€or€osmarty et al. 2010). In headwater streams, changes in land use have
greater ecological impacts because the headwaters usually comprise a relatively fragile
ecosystem, contain more taxa of aquatic organisms that are sensitive to changes in water
*Corresponding author. Email: [email protected]
� 2013 Taylor & Francis
Journal of Freshwater Ecology, 2013
Vol. 28, No. 3, 307–328, http://dx.doi.org/10.1080/02705060.2013.779941
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

quality and habitat disturbance, and play an important role in maintaining the function,
health, and biodiversity of whole river networks (Meyer and Wallace 2001; Meyer et al.
2007; Richardson and Danehy 2007; Wipfli et al. 2007).
Many researchers have emphasized the importance of riparian corridors in influencing
the structure and composition of macroinvertebrate communities (Dodd et al. 1994;
Sponseller et al. 2001; Potter et al. 2005; Rios and Bailey 2006). Riparian vegetation
serves as an important energy subsidy to stream systems, particularly in forested head-
waters, where inputs of allochthonous material – large woody debris and leaf litter – drive
the trophic cascade (Vannote et al. 1980; Allan and Castillo 2007). Many macroinverte-
brates, particularly the shredder functional feeding group, rely on these allochthonous
inputs as a food source, contributing to the degradation of material into coarse particulate
organic matter (CPOM), fine particulate organic matter (FPOM) and other forms of or-
ganic matter (Cummins and Klug 1979). Riparian vegetation also shades the stream chan-
nel, thereby regulating stream temperature and reducing primary production, and filters
nutrients, sediments, and contaminants. These are functions that further alter the composi-
tion of producers and consumers through the food web (Allan 2004; Potter et al. 2005).
As an important functional group in stream ecosystems that sustains the stability and
complexity of aquatic communities, macroinvertebrates show predictable responses as
the quality of riparian habitat degrades (Weigel et al. 2000; Sponseller et al. 2001).
Many studies have demonstrated the impact of land use disturbances, such as forestry,
agriculture, and urbanization, on macroinvertebrate communities at different scales of
watersheds and riparian corridors. Based on 17 years of monitoring, Reid et al. (2010)
evaluated the community variation of macroinvertebrates in response to forest harvest.
The study found that the density of Diptera, Mollusca, and Oligochaeta increased, while
the density of Ephemeroptera decreased, with more pronounced negative impacts in
smaller watersheds than larger ones. Weigel et al. (2003) identified that the proportion of
forested area within the 0–30 m riparian zone showed a positive relationship with species
richness, biotic integrity, Ephemeroptera-Plecoptera-Trichoptera (EPT) taxa, and scraper
and filterer functional feeding groups, while development (commercial, residential, or ur-
ban areas) and disturbed areas (cropland, pasture, and relatively modified vegetation)
within the 30–100 m corridor contributed to erosional assemblages within the Northern
Lakes and Forests ecoregion of the Midwestern United States.
Agricultural land use contributes non-point sources of fertilizer, sediment, and pesti-
cide pollution into the stream system; land use conversion from forest to agriculture also
changes the light, temperature, and trophic regimes (Cooper 1993; Allan 2004). These ag-
ricultural effects are manifested in changes in the macroinvertebrate community, such as
reduced diversity (Allan et al. 1997; Sponseller et al. 2001). Although urban areas typi-
cally constitute a small percentage of the overall area of a watershed, they have a dispro-
portionately large influence on the stream system (Paul and Meyer 2001) by increasing
surface runoff and channel erosion, altering channel morphology, and increasing loads of
sediments, nutrients, and contaminants (Roy et al. 2003). Paul and Meyer (2001) demon-
strated that indices of macroinvertebrate diversity and biotic integrity declined even under
low levels of impervious surface cover in streams across several states in the United
States. In the Piedmont region of Georgia, changes in sediment transport, sediment size,
and increased solutes associated with expanding urbanization detrimentally affected mac-
roinvertebrate metrics of taxon richness, EPT richness, and biodiversity, while favoring
less diverse and more pollution-tolerant macroinvertebrate assemblages (Roy et al. 2003).
During the past few decades, the importance of riparian corridors in sustaining water
quality and biological integrity has been widely discussed (Stevens and Cummins 1999;
308 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Sponseller et al. 2001). Moore and Palmer (2005) offered a nuanced perspective of the in-
teraction among land uses. The researchers compared urban and agricultural headwater
streams in Maryland and found relatively high diversity among agricultural headwaters,
while urban areas exhibited predictable negative influences on the macroinvertebrate
community. The negative influence of urban headwaters was minimized, however, in
those streams that contained forested buffers. Furthermore, the authors attributed the high
macroinvertebrate diversity in the agricultural sites to agricultural best management prac-
tices (BMPs) in place in the region (Moore and Palmer 2005). Such agricultural BMPs in-
cluded no-till farming and riparian buffer strips. The protection of streamside zones
(Prud’homme and Greis 2002) through the use of BMPs is already widely used as a man-
agement practice for improving stream quality in the United States (Gergel et al. 2002;
Shandas and Alberti 2009).
In order to develop effective riparian management and restoration plans for upper
headwater streams of the Taizi River, it is necessary to quantify how land uses affect the
stream ecosystem at different riparian corridor scales. In this study, water quality and
macroinvertebrates were used as indicators due to their predictable responses and broad
application to riparian land use developments. The aims were to: (1) identify the main
effects of local scale land uses on water quality and macroinvertebrate assemblages along
riparian corridors in headwater streams, and (2) reveal the relationships between indica-
tors and dominant land uses (forest, farmland, grassland, and villages in this research) at
different corridor widths. The results are placed in a management context for use by local
environmental protection agencies.
Methods
Study area and sampling sites
The study was carried out along three headwater streams of the Taizi River, including
Taizi Bei Stream (TZB), Taizi Nan Stream (TZN), and Xiaotanghe Stream (XTH), lo-
cated in the Changbai mountain area of northeastern China. The research area is located
within China’s mid- to high-latitudes in a temperate and continental monsoon climate
zone. Sampling occurred in autumn 2009, from early September to mid-October, to mini-
mize the effects of hydrological variation from precipitation. The autumn season
(September–November) has an average air temperature of 8.0�C (minimum 6.2�C;maximum 10.0�C) and an average precipitation of 144 mm (minimum 58 mm; maximum
260 mm), as observed during the past 52 years (Liu and Zhang 2012). Autumn typically
has the most stable hydrological condition since snowmelt in the spring and relatively
high precipitation in the summer contribute to higher stream flows in those seasons.
The three streams are located in the same sub-ecoregion, characterized by different
levels and modes of riparian disturbances. The TZB Stream has the greatest area of farm-
land and village development in the stream riparian corridors. The XTH Stream has the
second highest level of forest cover and farmland development, and the lowest level of
village development. In contrast, the agricultural development of the TZN Stream is
broadly restricted for the purpose of tourism. The areas of crop farmland and villages
were lowest and second lowest, respectively, along the TZN Stream. Forest cover is pre-
dominately mixed broadleaf deciduous. Corn (Zea mays) is the predominant crop type in
the research area. Corn crops are planted in May and harvested in late October; both
planting and harvesting are done manually. After the harvest, the stalks are left in the
fields and burned as fertilizer. The village houses are clustered together along the
Journal of Freshwater Ecology 309
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

riverbank with a population density of about 50–100 people per village. Villages are sur-
rounded by agricultural land and located several kilometers away from neighboring
villages.
In this study, only the local scale was considered. In total, 43 sites were sampled from
the three streams, including 18 sites from TZB, 15 sites from TZN, and 10 sites from
XTH (Figure 1). Sites were selected based on similar stream size (first- to third-order
streams according to Strahler stream order). The sampling sites were arranged from up-
stream to downstream, with a distance of 3 km between each site, except one in the XTH
due to difficulty of access. Since the study focused on land use impacts on mesohabitats
within the stream segment, the 3-km length was determined based on an estimated
35 times the mean stream width (Cohen et al. 1998). Four sampling sites (TZB1, TZB2,
TZB3, and TZN1) were excluded from the data analysis due to extremely low and discon-
tinuous flow, as the samples were collected from isolated, stagnant pools. Another five
sampling sites (TZB5, TZB13, TZB17, TZN3, and TZN6) were also excluded due to pre-
cipitation events that occurred within 24 hours before the sampling period, so as to main-
tain consistency when comparing samples collected during base flow conditions.
Riparian land use data acquisition
Land use data were visually interpreted from SPOT 5 images with a panchromatic band of
resolution of 2.5 m and multispectral bands of 10 m. Data acquisition occurred during
Figure 1. Location of the 43 sampling sites distributed in three tributaries of Taizi River in autumn2009.
310 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

September 2010 before crops were harvested in the surrounding farmland. The panchro-
matic band and multispectral bands were geometrically corrected with a topographic map
of 1:50,000 scale. All bands were projected into Albers conical equal area projection. The
panchromatic band and multispectral bands were then fused in ERDAS to obtain images
with both high resolution and spectral information. Land use information was determined
through visual interpretation of the fused map.
Land uses in the gradient were extracted through buffer tools in ArcGIS and ground-
truthed to verify the classification during field surveys in autumn 2010 before the crops
were harvested. Eight types of land uses were classified. Forest, farmland, grassland, and
village were the dominant land use types. The remaining land cover area comprised the
river channel, floodplain, unused (e.g., wasteland or abandoned land), and pools (e.g., ar-
tificial ponds for aquaculture or irrigation alongside the stream channel). In order to un-
derstand the influence of riparian land use on water quality and macroinvertebrates, the
study focused on the four dominant land use types of forest, farmland, grassland, and vil-
lages within different widths of riparian corridors. Riparian corridors were selected at 25,
50, 10, 200, and 500 m widths for 3-km segments upstream of each sampling site
(Figure 2).
Figure 2. A representative example of an ArcGIS map with the eight land use types shown in rela-tion to the five different widths of riparian corridors.
Journal of Freshwater Ecology 311
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Water quality
Eight chemical factors were measured to quantify the effects of land use on water quality.
Water quality was sampled one time per site, concurrent with macroinvertebrate samples.
The pH was measured using a pH pen (YSI, Inc., http://www.ysi.com) instrument. The
electrical conductivity (EC), total dissolved solids (TDS), and dissolved oxygen (DO)
were measured in situ with a handheld multi-probe (Professional Plus, YSI, Inc.). Calibra-
tion of sensors was performed each day during the sampling period. Before the samples
were processed, 2 L of stream water were collected for laboratory analysis in polyethyl-
ene plastic bottles, which were pre-rinsed three times with distilled water, and transported
to the lab within 24 h on ice. In the laboratory, ammonia-nitrogen (NH3-N) was measured
with Nessler’s reagent and total phosphorus (TP) was measured with an inductively-
coupled plasma atomic emission spectrometer (ICP-AES; IRIS Intrepid II XSP DUO,
USA). Total nitrogen (TN) was determined through the alkaline potassium persulfate
oxidation–UV spectrophotometric method. The chemical oxygen demand index (CODMn)
was measured by the potassium permanganate method (MEP 2002). All the methods
followed Chinese Standard Methods for Examination of Water and Wastewater
(MEP 2002).
Benthic macroinvertebrate indices
Macroinvertebrates were sampled in targeted riffles within the 100 m sampling reach
using a Surber net (30 � 30 cm, 1 mm mesh size) with three replicates. Each replicate
was collected and preserved with 70% ethanol in separate 500-mL bottles. All the indi-
viduals were sorted, counted, and identified to the species or genus level using micro-
scopes in the lab. The Chironomidae were identified to subfamily level.
Twenty-five macroinvertebrate indices, including community structure (14), func-
tional feeding groups (6), biodiversity (4), and bioassessment (1), were calculated. Four-
teen indices were calculated to reflect the community structure: total number of
individuals; total taxa richness; Ephemeroptera-Plecoptera-Trichoptera (EPT) taxa rich-
ness; taxa richness of each Ephemeroptera, Plecoptera, and Trichoptera group; relative
EPT abundance (%); relative abundance of each Ephemeroptera, Plecoptera, and Trichop-
tera group; relative abundance of Chironomidae; and relative abundance of each Chirono-
midae subfamily, Chironominae, Orthocladiinae, and Tanypodinae. According to
identification of the mouthpart structure and literature references, taxa were classified
into six functional feeding groups: filterers, scrapers, collector-gatherers, predators (di-
vided into two groups of Tanypodinae-included and Tanypodinae-excluded), and shred-
ders (Morse et al. 1994; Merritt and Cummins 1996). Four biodiversity indices were
calculated, namely Shannon’s diversity, Margalef’s diversity, Peilou’s evenness, and
Simpson’s diversity (Magurran 2004). A bioassessment index of average score per taxon
(ASPT) was calculated according to Armitage et al. (1983).
Statistical analysis
Cluster analysis was used to classify the sampling sites based on the species composition
of macroinvertebrates using the flexible Beta linkage method (with a default value of
�0.25) with Sørensen’s distance measure (McCune and Mefford 1999). Differences
among clusters were tested by the multi-response permutation procedure (MRPP), which
312 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

is a nonparametric method for testing multivariate differences among predefined groups
(Zimmerman et al. 1985). The Sørensen coefficient was used as the distance measure
in the MRPP. The significance of the null hypothesis – that there was no difference
among groups – was tested with a Monte Carlo randomization procedure with 1000
permutations.
The differences in water quality factors and macroinvertebrate indices among differ-
ent streams were evaluated using Kruskal–Wallis (K–W) tests. Among the four main
types of land uses, the percent area of farmland and villages were selected as the main
stressors to identify the effects of land use on the degradation of water quality and macro-
invertebrate communities at different widths of riparian corridors. The Pearson correla-
tion analysis was used to determine the significance among the main land uses with water
quality factors and biological indices.
The integrated relationship between land uses at different riparian widths and macro-
invertebrates was explored using canonical correspondence analysis (CCA). Significant
correlations among the four dominant land uses and various biological indices were ana-
lyzed in the final CCA. Those land uses that had significant correlations with each other
within the same riparian widths were omitted in the final CCA. Forward selection and
Monte Carlo permutations were used to select a minimum set of variables that had signifi-
cant and independent effects on macroinvertebrate metrics. Prior to the cluster analysis
and CCA, all data were log(x þ 1) transformed in order to eliminate the influence of ex-
treme values and ensure that the data fit was normal. The cluster analysis and MRPP test
were performed using PC-ORD (Version 5.0) software. The difference analysis and corre-
lation analysis were performed using STATISTICA (Version 7.0). CCA was carried out
with CANOCO (Version 4.5).
Results
Land use properties at different riparian corridor widths
The percentages of dominant land use types were compared for different riparian widths
along the three streams (Figure 3). Farmland and villages had higher percentages along
XTH and TZB Streams than along TZN Stream at all riparian widths, except XTH Stream
at 500 m riparian width. The percentage of forest and grassland area was higher at TZN
Stream than the other two streams. The land use patterns were generally similar among
the three streams. The four main land use types did not comprise a majority of the riparian
area at the 25 m corridor width, which was instead dominated by other land use types.
Both forest and farmland dominated the main land use types as the riparian corridor wid-
ened from 100 m to 500 m at TZB Stream (Figure 3a) and TZN Stream (Figure 3b), and
from 50 m to 500 m at XTH Stream (Figure 3c). The variation in farmland and village
area confirmed that the human disturbances occurred mainly within the 500 m riparian
corridors for all three streams. When the riparian width doubled, the percentage of farm-
land and villages did not increase correspondingly. In contrast, the percentage of forest in-
creased considerably with increasing riparian width, up to 200 m, for all three streams.
Water quality
The main physical and chemical factors that reflect the water quality were compared
among the three streams (Table 1). All the water quality parameters, except for NH3-N,
Journal of Freshwater Ecology 313
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

differed significantly among the three streams. The pH values in TZN and XTH Streams
were relatively high, although they were within the expected range because the water is
naturally alkaline in the study areas. The pH in TZB Stream, however, was lower than the
normal range. The EC and TDS were significantly higher in TZB and XTH Streams. The
DO values were all nearly saturated for the three streams. Meanwhile, the organic pollu-
tion was relatively low based on the concentration of CODMn. Significant nutrient enrich-
ment, reflected in the concentrations of nitrogen and phosphorus, was confirmed in XTH
Stream. Both the TN and TP concentrations were only slightly elevated at TZB Stream.
The concentrations of NH3-N were low and showed similar ranges for each stream.
Biological indices of macroinvertebrates
Overall, 124 taxa were identified from the three streams of upper Taizi River. In TZN
Stream, the three most dominant taxa were Orthocladiinae spp. (Diptera: Chironomidae),
Ecdyonurus viridis (Ephemeroptera: Heptageniidae), and Paraleptophlebia japonica
Figure 3. Percentages of the four dominant land use types of forest, farmland, grassland, and vil-lages within different riparian widths at each stream. River channel, floodplain, pool, and unusedland types were combined into the other category. The three graphs represent (a) TZN Stream, (b)TZB Stream, and (c) XTH Stream.
314 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

(Ephemeroptera: Leptophlebiidae) with an average of 122, 107, and 100 individuals per
site, respectively. Four taxa, including Paraleptophlebia japonica (Ephemeroptera: Lepto-
phlebiidae), Antocha sp. (Diptera: Tipulidae), Epeorus latifolium (Ephemeroptera: Hepta-
geniidae), and Suwallia sp. (Plecoptera: Chloroperlidae), appeared at all sampling sites in
TZN Stream. Chironominae spp. (Diptera: Chironomidae), Orthocladiinae spp. (Diptera:
Chironomidae), and Cheumatopsyche sp. (Trichoptera: Hydropsychidae) were the three
most dominant taxa, averaging 542, 514, and 339 individuals per site, respectively, in TZB
Stream. Among the dominant taxa, the first two appeared at all sampling sites in TZB
Stream. In XTH Stream, Orthocladiinae spp. (Diptera: Chironomidae), Beatis thermicus
(Ephemeroptera: Baetidae), and Serratella rufa (Ephemeroptera: Ephemerellidae) were the
most dominant taxa with an average of 313, 78, and 32 individuals per site, respectively.
The first two taxa, in addition to Tanypodinae spp. (Diptera: Chironomidae) and Chirono-
minae spp. (Diptera: Chironomidae) appeared at all XTH Stream sampling sites.
Fourteen indices of macroinvertebrate community composition were calculated and
compared among the three streams (Table 2). Except for the percentage of Ephemerop-
tera, all of the other indices were generally significantly different among the three
streams. The total abundance was significantly higher at TZB Stream. The total taxa num-
ber and EPT taxa number were significantly higher at both the TZB and TZN Streams.
Among the EPT taxa, the Plecoptera (stoneflies) were found at higher richness at TZN
Stream, while TZB Stream had higher Ephemeroptera (mayfly) and Trichoptera (caddis-
fly) richness. The percentage of EPT was significantly greater at TZN Stream (74.8%)
compared to 50.2% and 33.7% at TZB and XTH Streams, respectively. The percentage of
Table 1. The status of water quality, biodiversity indices, and a bioassessment index for macroin-vertebrates for 43 sites sampled in the Taizi River watershed (18 sites in Taizi Bei, 15 sites in TaiziNan, and 10 sites in Xiaotanghe). Values represent mean � standard deviation. Kruskal–Wallis(K–W) tests were used to evaluate differences among the three streams. The target thresholds forChinese environmental quality standards (EQS; MEP 2002) for surface water are also shown.
Index TZB TZN XTH K–W EQS
Water qualitypH 7.73 � 0.59a 8.7 � 0.31b 9.45 � 0.38c 27.9�� 7–9EC (ms/cm) 276.29 � 186.50b 45.11 � 20.99a 133.01 � 25.20b 22.0��
TDS (mg/L) 128.08 � 14.66b 39.65 � 18.30a 101.47 � 19.60b 25.8��
DO (mg/L) 11.96 � 1.36a 12.67 � 0.59a 10.87 � 1.43b 10.4�� 6 y
TN 1.80 � 0.29a 1.35 � 0.56a 6.15 � 0.45b 21.9��
TP 0.07 � 0.15a,b 0.02 � 0.01a 0.11 � 0.11b 6.7� 0.1 y
CODMn – 1.09 � 0.91 2.25 � 0.19 4.0 y
NH3-N 0.25 � 0.14 0.25 � 0.14 0.25 � 0.19 0.3 0.5 y
Biodiversity indicesShannon’s (H´) 3.46 � 0.73a 3.78 � 0.43a 2.48 � 0.77b 13.2��
Margalef’s (M) 3.40 � 0.61a 3.47 � 0.60a 2.47 � 0.62b 10.7��
Evenness (Ev) 0.65 � 0.12a,b 0.74 � 0.07a 0.55 � 0.16b 8.5�
Simpson’s (S) 0.84 � 0.09a 0.88 � 0.05a 0.67 � 0.16b 11.5��
Bioassessment indexof macroinvertebrates
ASPT 5.03 � 0.41 5.55�0.71 4.64 � 1.07 6.4�
a,b,cDifferences among the three streams as tested with the Kruskal–Wallis test. Those values with differentsuperscripts indicate significant differences.�p < 0.05, ��p < 0.01.yValues were derived from level II limits of the environmental quality standards for surface water (MEP 2002).
Journal of Freshwater Ecology 315
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13
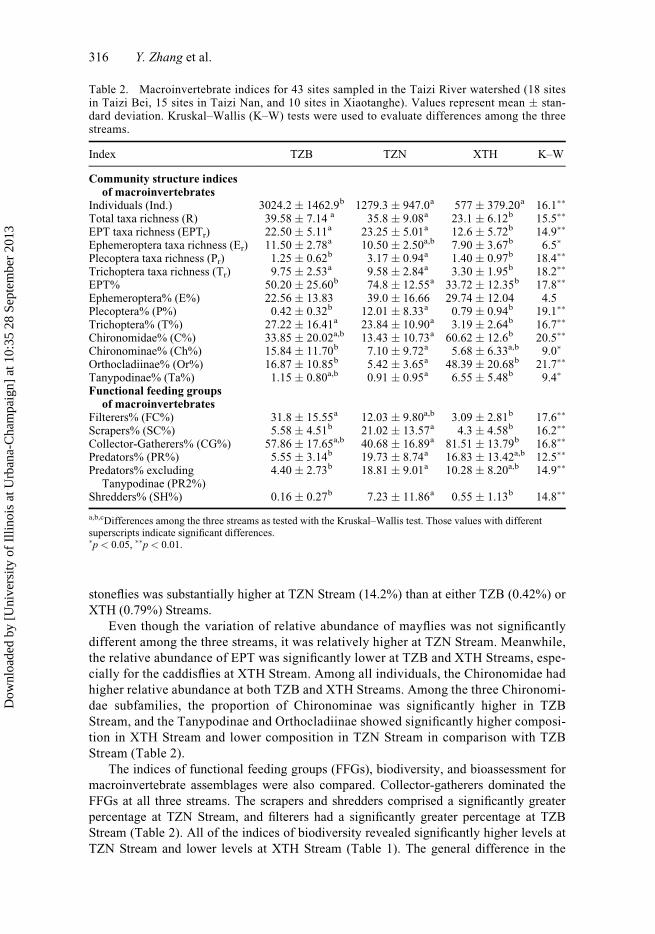
stoneflies was substantially higher at TZN Stream (14.2%) than at either TZB (0.42%) or
XTH (0.79%) Streams.
Even though the variation of relative abundance of mayflies was not significantly
different among the three streams, it was relatively higher at TZN Stream. Meanwhile,
the relative abundance of EPT was significantly lower at TZB and XTH Streams, espe-
cially for the caddisflies at XTH Stream. Among all individuals, the Chironomidae had
higher relative abundance at both TZB and XTH Streams. Among the three Chironomi-
dae subfamilies, the proportion of Chironominae was significantly higher in TZB
Stream, and the Tanypodinae and Orthocladiinae showed significantly higher composi-
tion in XTH Stream and lower composition in TZN Stream in comparison with TZB
Stream (Table 2).
The indices of functional feeding groups (FFGs), biodiversity, and bioassessment for
macroinvertebrate assemblages were also compared. Collector-gatherers dominated the
FFGs at all three streams. The scrapers and shredders comprised a significantly greater
percentage at TZN Stream, and filterers had a significantly greater percentage at TZB
Stream (Table 2). All of the indices of biodiversity revealed significantly higher levels at
TZN Stream and lower levels at XTH Stream (Table 1). The general difference in the
Table 2. Macroinvertebrate indices for 43 sites sampled in the Taizi River watershed (18 sitesin Taizi Bei, 15 sites in Taizi Nan, and 10 sites in Xiaotanghe). Values represent mean � stan-dard deviation. Kruskal–Wallis (K–W) tests were used to evaluate differences among the threestreams.
Index TZB TZN XTH K–W
Community structure indicesof macroinvertebrates
Individuals (Ind.) 3024.2 � 1462.9b 1279.3 � 947.0a 577 � 379.20a 16.1��
Total taxa richness (R) 39.58 � 7.14 a 35.8 � 9.08a 23.1 � 6.12b 15.5��
EPT taxa richness (EPTr) 22.50 � 5.11a 23.25 � 5.01a 12.6 � 5.72b 14.9��
Ephemeroptera taxa richness (Er) 11.50 � 2.78a 10.50 � 2.50a,b 7.90 � 3.67b 6.5�
Plecoptera taxa richness (Pr) 1.25 � 0.62b 3.17 � 0.94a 1.40 � 0.97b 18.4��
Trichoptera taxa richness (Tr) 9.75 � 2.53a 9.58 � 2.84a 3.30 � 1.95b 18.2��
EPT% 50.20 � 25.60b 74.8 � 12.55a 33.72 � 12.35b 17.8��
Ephemeroptera% (E%) 22.56 � 13.83 39.0 � 16.66 29.74 � 12.04 4.5Plecoptera% (P%) 0.42 � 0.32b 12.01 � 8.33a 0.79 � 0.94b 19.1��
Trichoptera% (T%) 27.22 � 16.41a 23.84 � 10.90a 3.19 � 2.64b 16.7��
Chironomidae% (C%) 33.85 � 20.02a,b 13.43 � 10.73a 60.62 � 12.6b 20.5��
Chironominae% (Ch%) 15.84 � 11.70b 7.10 � 9.72a 5.68 � 6.33a,b 9.0�
Orthocladiinae% (Or%) 16.87 � 10.85b 5.42 � 3.65a 48.39 � 20.68b 21.7��
Tanypodinae% (Ta%) 1.15 � 0.80a,b 0.91 � 0.95a 6.55 � 5.48b 9.4�
Functional feeding groupsof macroinvertebrates
Filterers% (FC%) 31.8 � 15.55a 12.03 � 9.80a,b 3.09 � 2.81b 17.6��
Scrapers% (SC%) 5.58 � 4.51b 21.02 � 13.57a 4.3 � 4.58b 16.2��
Collector-Gatherers% (CG%) 57.86 � 17.65a,b 40.68 � 16.89a 81.51 � 13.79b 16.8��
Predators% (PR%) 5.55 � 3.14b 19.73 � 8.74a 16.83 � 13.42a,b 12.5��
Predators% excludingTanypodinae (PR2%)
4.40 � 2.73b 18.81 � 9.01a 10.28 � 8.20a,b 14.9��
Shredders% (SH%) 0.16 � 0.27b 7.23 � 11.86a 0.55 � 1.13b 14.8��
a,b,cDifferences among the three streams as tested with the Kruskal–Wallis test. Those values with differentsuperscripts indicate significant differences.�p < 0.05, ��p < 0.01.
316 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

ASPT scores was that they were significant within each tributary, but not in relation to
other tributaries.
Similarity analysis for macroinvertebrates
The similarity of benthic macroinvertebrate communities was analyzed to reveal the
effects of land uses on community composition among the three streams. Thirty-four sam-
pling sites were classified into three clusters by applying the cluster analysis (Figure 4).
MRPP revealed significant differences among the three clusters (A ¼ 0.321, p < 0.001).
The classifying result showed that species composition had greater similarity within each
stream, and was significantly different among each of the three streams in general. Con-
sidering the similar climate, geomorphology, underlying geological gradient, and vegeta-
tion types of the three streams, the differences in community composition potentially
result from differing characteristics of long-term land use development.
Relationships among land use, water quality, and macroinvertebrates
The Pearson’s correlation analysis confirmed the strong relationships between land uses
and water quality (Table 3). The proportion of forest had significantly positive correla-
tions with pH values from 200 m (p < 0.05) to 500 m (p < 0.01) riparian width. The pro-
portion of forest also had significantly positive correlations with concentrations of DO
and CODMn at the largest riparian width of 500 m. The proportion of farmland had
strongly positive correlations with EC from 50 m to 200 m riparian widths (r ¼ 0.41 to
0.47, p < 0.05). The TN was also positively correlated with proportion of farmland at
25 m and 50 m riparian widths (r ¼ 0.37 to 0.39, p < 0.05). In contrast, the proportion of
grassland showed negative correlations with EC and TDS at all riparian widths. The pro-
portion of village negatively correlated with pH, DO, and CODMn at the relatively large
Figure 4. Classification of the sampling sites with the flexible Beta linkage method usingSørensen’s distance measure. Each sampling site from the three clusters is indicated in Figure 1.
Journal of Freshwater Ecology 317
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

riparian widths of 200 m and 500 m (r ¼ �0.37 to �0.57, p < 0.05). Meanwhile, the pro-
portion of village showed positive correlations with TP from 50 m to 200 m riparian
widths (r ¼ 0.39 to 0.42, p < 0.05; Table 3).
The correlation analysis broadly revealed the associations among proportions of farm-
land and village and the macroinvertebrate indices of species richness, species composi-
tion, functional feeding groups, diversity, and bioassessment (Tables 4 and 5). Stoneflies
showed broad significant correlations with all types of land uses. Both the taxa richness
and relative abundance of stoneflies were positively correlated with the proportion of for-
est and grassland and negatively correlated with the proportion of farmland and village at
most of the riparian widths, except for relative stonefly abundance at 100 m and 200 m ri-
parian widths (Table 4). The proportion of forest showed positive correlations with preda-
tors of both groups at all riparian widths. The proportion of farmland also showed
negative correlations with both predator groups at all riparian widths except 25 m. The
proportion of farmland showed significantly negative correlations with taxa richness of
EPT (r ¼ �0.37, p < 0.05) and with taxa richness of caddisflies at both 25 m and 50 m
(r ¼ �0.36 to �0.41, p < 0.05) riparian widths. The relative abundance of Chironomidae
positively correlated with the proportion of farmland also at the relatively narrow riparian
widths from 25–100 m (Table 4). The percentages of both Chironominae and Tanypodi-
nae did not have significant correlations with land uses at any widths. However, the per-
centage of Orthocladiinae showed significant correlations with the proportion of forest at
the small to medium widths (25–100 m) and with the proportion of farmland at medium
widths (100–200 m).
Except for the filterers, the composition of other FFGs showed broad correlations with
the proportion of farmland at different riparian widths. Among the FFGs, the scrapers,
Table 3. Pearson’s correlation values (r) between the proportion of main land uses within differentriparian corridor widths and water quality parameters. Significant values are indicated in bold.
Land useCorridorwidth (m) pH EC TDS DO TN TP CODMn NH3-N
Forest 25 0.19 �0.15 �0.11 0.16 �0.14 �0.08 0.12 0.0250 0.22 �0.14 �0.08 0.19 �0.10 �0.05 0.16 0.01100 0.27 �0.11 �0.04 0.22 �0.04 �0.04 0.21 0200 0.34� �0.08 0.01 0.28 0.07 �0.05 0.28 0500 0.44�� �0.05 0.07 0.38� 0.21 �0.03 0.37� 0.03
Farmland 25 0.13 0.34 0.38� 0.09 0.39� �0.18 0.01 0.0950 0.14 0.41� 0.44� 0.10 0.37� �0.14 �0.06 0.13100 0.13 0.47�� 0.48�� 0.10 0.33 �0.07 �0.15 0.18200 0.02 0.45�� 0.42� 0.01 0.21 �0.03 �0.25 0.20500 �0.22 0.33 0.24 �0.14 �0.02 0.04 �0.34 0.15
Grassland 25 �0.07 �0.40� �0.40� 0 �0.08 �0.18 0.16 �0.1450 �0.12 �0.45�� �0.45�� �0.02 �0.16 �0.21 0.13 �0.12100 �0.18 �0.51�� �0.52�� �0.06 �0.24 �0.24 0.10 �0.13200 �0.20 �0.54�� �0.55�� �0.09 �0.26 �0.24 0.10 �0.13500 �0.18 �0.49�� �0.50�� �0.08 �0.26 �0.24 0.04 �0.08
Villages 25 �0.08 �0.02 �0.03 �0.18 0.18 0.31 0.13 0.2050 �0.16 �0.05 �0.03 �0.25 0.18 0.39� 0.15 0.22100 �0.26 �0.06 �0.06 �0.31 0.14 0.42� 0.11 0.20200 �0.41� �0.09 �0.10 �0.37� �0.02 0.39� �0.08 0.06500 �0.57�� �0.16 �0.22 �0.57�� �0.33 0.17 �0.42� �0.15
�p < 0.05; ��p < 0.01.
318 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Table
4.
Pearson’s
correlationcoefficients
(r)betweenmainlandusesandmacroinvertebratescommunitystructure
indices.Index
abbreviationsaredefined
inTable2.Significantvalues
areindicated
inbold.
Landuse
Corridorwidth
(m)
Ind.
REPTr
Er
Pr
Tr
EPT%
E%
P%
T%
C%
Ch%
Or%
Ta%
Forest
25
�0.16
0.11
0.12
0.02
0.35�
0.10
0.13
0.05
0.39�
0.14
�0.17
0.06
�0.38�
0.13
50
�0.21
0.08
0.11
0.01
0.38�
0.08
0.12
0.05
0.44�
0.14
�0.23
0.09
�0.40�
0.06
100
�0.27
0.04
0.06
�0.02
0.37�
0.03
0.11
0.05
0.46��
0.10
�0.24
0.12
�0.37�
�0.01
200
�0.31
�0.02
�0.03
�0.07
0.32
�0.09
0.09
0.07
0.42�
0.01
�0.20
0.16
�0.29
�0.05
500
�0.32
�0.07
�0.13
�0.10
0.25
�0.22
0.09
0.15
0.35�
�0.10
�0.13
0.18
�0.18
�0.11
Farmland
25
0.03
�0.19
�0.37�
�0.15
�0.55��
�0.41�
�0.18
�0.05
�0.47��
�0.32
0.36�
0.22
0.32
0.23
50
0.08
�0.17
�0.32
�0.11
�0.54��
�0.36�
�0.19
�0.05
�0.51��
�0.30
0.36�
0.17
0.33
0.24
100
0.14
�0.13
�0.26
�0.04
�0.55��
�0.28
�0.16
�0.03
�0.56��
�0.25
0.34�
0.11
0.36�
0.21
200
0.22
�0.06
�0.12
0.05
�0.51��
�0.11
�0.10
�0.01
�0.55��
�0.14
0.27
00.34�
0.13
500
0.29
0.02
0.06
0.13
�0.42�
0.13
�0.06
�0.07
�0.48��
0.04
0.18
�0.10
0.25
0.12
Grassland
25
0.04
0.18
0.26
0.18
0.39�
0.23
0.10
0.10
0.35�
0.07
0.01
0.12
�0.13
0.10
50
0.06
0.20
0.29
0.18
0.41�
0.27
0.13
0.11
0.39�
0.11
�0.05
0.02
�0.14
0.02
100
0.07
0.22
0.30
0.16
0.41�
0.29
0.15
0.10
0.42�
0.14
�0.10
�0.06
�0.18
�0.02
200
0.06
0.21
0.28
0.13
0.41�
0.29
0.16
0.12
0.43�
0.14
�0.11
�0.09
�0.18
�0.05
500
0.03
0.21
0.27
0.11
0.38�
0.29
0.16
0.05
0.43�
0.16
�0.08
�0.13
�0.16
�0.04
Villages
25
0.09
�0.25
�0.22
�0.06
�0.35�
�0.26
�0.14
0.06
�0.27
�0.32
0.23
0.17
0.26
0.03
50
0.11
�0.25
�0.20
�0.07
�0.36�
�0.22
�0.17
0.03
�0.32
�0.32
0.25
0.14
0.32
�0.01
100
0.17
�0.20
�0.15
�0.02
�0.36�
�0.16
�0.15
0.03
�0.36�
�0.27
0.26
0.08
0.36�
�0.02
200
0.29
�0.09
�0.07
0.01
�0.37�
�0.05
�0.18
�0.06
�0.41�
�0.17
0.25
0.01
0.31
0.11
500
0.42�
0.11
0.07
0.04
�0.22
0.16
�0.21
�0.21
�0.32
0.03
0.14
�0.15
0.15
0.24
� p<
0.05;��p<
0.01.
Journal of Freshwater Ecology 319
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Table
5.
Pearson’s
correlationcoefficients(r)betweenmainlandusesandfunctional
feedinggroups,biodiversity,andbioassessmentindices
ofmacroinverte-
brates.Index
abbreviationsaredefined
inTables1and2.Significantvalues
areindicated
inbold.
Functionalfeedinggroupsofmacroinvertebrates
Biodiversity
andbioassessmentindices
Landuse
Corridorwidth
(m)
FC%
SC%
CG%
PR%
PR2%
SH%
H´
MEv
SASPT
Forest
25
�0.12
0.25
�0.20
0.37�
0.39�
0.23
0.24
0.23
0.25
0.19
0.08
50
�0.18
0.30
�0.26
0.42�
0.44�
0.30
0.21
0.23
0.24
0.15
0.15
100
�0.25
0.33
�0.28
0.45��
0.47��
0.33
0.17
0.20
0.21
0.10
0.15
200
�0.32
0.29
�0.21
0.43�
0.44�
0.30
0.11
0.14
0.15
0.04
0.06
500
�0.38�
0.25
�0.11
0.39�
0.40�
0.24
0.05
0.07
0.11
0.01
�0.07
Farmland
25
�0.12
�0.34�
0.35�
�0.30
�0.35�
�0.35�
�0.25
�0.28
�0.22
�0.08
�0.55��
50
�0.06
�0.34�
0.35�
�0.34�
�0.39�
�0.39�
�0.23
�0.26
�0.20
�0.05
�0.49��
100
0.04
�0.36�
0.35�
�0.40�
�0.45��
�0.43�
�0.19
�0.25
�0.18
�0.02
�0.42�
200
0.19
�0.34�
0.28
�0.43�
�0.47��
�0.40�
�0.12
�0.20
�0.13
0.03
�0.28
500
0.32
�0.30
0.17
�0.43�
�0.46��
�0.34�
�0.08
�0.13
�0.12
0.02
�0.04
Grassland
25
�0.07
0.24
0.03
0.28
0.27
0.35�
0.20
0.22
0.17
0.14
0.21
50
�0.05
0.27
�0.01
0.26
0.27
0.38�
0.23
0.24
0.19
0.16
0.22
100
�0.05
0.30
�0.06
0.24
0.27
0.40�
0.23
0.26
0.19
0.17
0.21
200
�0.04
0.29
�0.06
0.24
0.28
0.40�
0.24
0.25
0.20
0.17
0.18
500
�0.01
0.24
�0.06
0.22
0.27
0.40�
0.24
0.27
0.20
0.18
0.17
Villages
25
�0.07
�0.35�
0.24
�0.16
�0.23
�0.18
�0.28
�0.38�
�0.24
�0.20
�0.15
50
�0.09
�0.41�
0.26
�0.23
�0.30
�0.20
�0.34�
�0.40�
�0.32
�0.27
�0.09
100
�0.05
�0.43�
0.27
�0.30
�0.37�
�0.26
�0.34�
�0.36�
�0.35�
�0.28
�0.05
200
0.06
�0.39�
0.24
�0.40�
�0.46��
�0.28
�0.27
�0.28
�0.31
�0.19
�0.01
500
0.32
�0.27
0.09
�0.40�
�0.42�
�0.20
�0.07
�0.07
�0.15
0.02
0.07
� p<
0.05;��p<
0.01.
320 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

predators, and shredders showed negative correlations with the proportion of farmland,
while collector-gatherers showed positive correlations with the proportion of farmland.
The proportion of grassland showed positive correlations with shredders at all riparian
widths. The proportion of village had negative correlations with scrapers from 25 m to
200 m riparian width and negative correlations with predators, including Tanypodinae
from 200 m to 500 m and predators, excluding Tanypodinae from 100 m to 500 m ripar-
ian widths. None of the biodiversity indices showed significant correlations with propor-
tions of forest, farmland, or grassland at any riparian corridor widths. However, the
proportion of village had negative correlations with Shannon’s diversity index at 50 m
and 100 m riparian width, with Margalef’s index from 25 m to 100 m riparian width, and
with Peilou’s evenness at 100 m riparian width (r ¼ �0.40 to �0.34, p < 0.05). For the
bioassessment index, the ASPT showed significant negative correlations with the propor-
tion of farmland from 25 m to 100 m riparian widths (Table 5).
Since the four main land uses showed significant correlations with various macroin-
vertebrate indices at all riparian widths, the final CCA employed nineteen variables: the
four dominant land use types at each of the five riparian corridor widths, except for farm-
land at 500 m, since the percentages of farmland and forest were significantly correlated
at the 500-m width (R2 > 0.90, p < 0.001). The results showed that the distribution of
benthic macroinvertebrates was independently affected by the proportions of forest
(50 m), farmland (200 m), grassland (200 m), and village (100 m) in the upper streams
of the Taizi River (Monte Carlo test, p < 0.05, Figure 5). These land uses showed the co-
incident effects on the distribution of macroinvertebrates. Loadings on axis 1 (21.7%)
and 2 (12.9%) were substantially larger than that of succeeding axes, and primarily
expressed variation at the 200 m width of farmland and 25 and 50 m widths of forest
within the riparian corridors. The cumulative percentage variance of the species–environ-
ment relation was 34.6% of the first two axes (Table 6).
Figure 5. CCA ordination biplot of sampling sites of macroinvertebrate communities and fourdominant land uses within five riparian corridor widths. The sampling sites of different clusters de-fined in Figure 4 are indicated by different symbols. The first and second axes show 0.209 and0.124 eigenvalues with 21.7% and 12.9% of variance explained, respectively.
Journal of Freshwater Ecology 321
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Discussion
Two key findings highlighted the importance of riparian corridors for stream ecosystems.
Firstly, significant differences in water quality parameters (Table 1) among the three
tributaries confirmed findings from other studies that long-term land uses could alter wa-
ter quality (Scarsbrook and Halliday 1999; Kratzer et al. 2006), even with low levels of
human disturbance. Compared to the Chinese environmental quality standards for surface
water (MEP 2002), the water quality condition was relatively good compared with the
level II thresholds (Table 1). The pH values were within the acceptable range, DO was
above the threshold, and TP, CODMn, and NH3-N were all below level II thresholds, ex-
cept for the elevated levels of TP in the XTH Stream.
The relatively high percentages of Ephemeroptera, Plecoptera, and Trichoptera, and
relatively high ASPT index values in the TZB and TZN streams reflected generally healthy
macroinvertebrate communities (Tables 1and 2; Armitage et al. 1983). However, our re-
search revealed the relationship of agriculture with EC and TDS at small to medium widths
(25–200 m), and slight enrichment of nitrogen at small widths (25–50 m). The CCA also
showed the importance of the proportion of farmland and grassland at the 200 m width
(Figure 5). Our findings corroborated those of Sponseller et al. (2001) and Tran et al.
(2010), which showed that the strongest influence on stream water quality occurred within
Table 6. Weighted correlation of land uses at different riparian zone widths with CCA axis 1 andaxis 2. The eigenvalues and cumulative percentage variance of species–environment relations areshown for the first two axes. The F-values and p-values of the Monte Carlo test are also shown. Thebold values represent a minimum set of land uses that had significant and independent effects onmacroinvertebrate distribution.
Land use typeRiparian zonewidth (m) Axis 1 Axis 2
F-valuesof MonteCarlo test p-value
Forest 25 0.4219 0.5742 1.45 0.07450 0.4592 0.5256 1.53 0.036100 0.4675 0.4676 0.77 0.726200 0.4517 0.4384 1.07 0.324500 0.4442 0.3684 0.62 0.826
Farmland 25 �0.5224 0.0790 1.22 0.17450 �0.5580 0.0064 0.76 0.730100 �0.6160 �0.1012 0.69 0.824200 �0.6482 �0.2654 3.03 0.002
Grassland 25 0.4411 �0.0871 1.14 0.26850 0.4661 �0.1731 0.87 0.624100 0.4848 �0.2230 0.69 0.760200 0.4888 �0.2315 1.53 0.022500 0.4255 �0.2087 0.60 0.852
Villages 25 �0.1289 0.1398 1.43 0.08450 �0.1891 0.1174 0.98 0.432100 �0.2462 0.0481 1.61 0.036200 �0.3084 �0.0887 0.54 0.906500 �0.3309 �0.3169 1.36 0.080
Eigenvalues 0.209 0.124Cumulative percentage
variance of species–environment relation
21.7% 34.6%
322 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

200 m of the stream channel. Our study did not find a significant correlation between the
proportion of land use and phosphorus enrichment, although Norton and Fisher (2000)
found that the forest within the riparian zone acted as a total phosphorus (TP) source within
0–100 m of the channel, but acted as a TP sink at further distances of 100–300 m. In con-
trast, the villages located in the riparian corridors may be an important TP source in the
study region. Villages in China are characterized by clusters of single-family houses sur-
rounded by farmland. These villages, however, act as a nonpoint source of pollution, espe-
cially through the input of phosphorus present in untreated domestic wastewater and
sewage from households and associated livestock (Zhang et al. 2004, Yang et al. 2013).
The increasing proportion of village area from 50 m to 200 m widths likely contributed to
the high levels of phosphorus observed at the sampling sites (Table 3).
Even with low levels of human disturbances, significant variation of macroinverte-
brate composition was observed among the three tributaries (Figures 4 and 5), which con-
firmed earlier findings that long-term land uses alter the biological communities in river
ecosystems (Dance and Hynes 1980; Harding et al. 1999; Schulz and Liess 1999; Hill
et al. 2001). As the proportion of farmland and villages increased, the lack of stoneflies,
both in terms of taxa richness and relative abundance, was most significant. Stoneflies are
sensitive to the alteration of thermal patterns, especially the relationship between increas-
ing maximum in-stream temperature associated with declining forest cover (Vannote and
Sweeney 1980; Sweeney et al. 1986; Quinn et al. 1994). A similar effect has also been
observed for mayflies: past studies have found that changes in temperature of 3–5�C, as-sociated with declining riparian vegetation cover, were sufficient to negatively alter larval
recruitment and growth for many mayflies in Piedmont streams (Sweeney 1993). The
effects of riparian deforestation and other disturbances of stream thermal patterns have
been reported within 50 m (Burton and Likens 1973), 200 m (Sponseller et al. 2001), and
300 m (Storey and Cowley 1997) riparian corridors. Since most taxa of stoneflies have
relatively high sensitivity for land use changes, organic pollution, and habitat disturban-
ces, even a low level of human disturbance could cause significant degradation in stonefly
assemblages. Correspondingly, stoneflies have been widely used as a predictor of stream
health degradation (Hellawell 1986; Rosenberg and Resh 1993; Wright 1995; Barbour
et al. 1999).
The shift in macroinvertebrate FFGs from a relatively diverse composition to the pre-
dominance of collector-gatherers and filterers generally revealed the presumed nutrient
input shifts from CPOM to FPOM (Short and Ward 1980; Vallania and Corigliano 2007).
The broad significant correlations between land use change and FFGs revealed the effects
of deforestation, corresponding with farmland and grassland development, on macroin-
vertebrates in the riparian corridors (Table 5). The predominance of filterers and collec-
tor-gatherers has been well-documented in agricultural areas (Shieh et al. 2003; Utz et al.
2009). Shredders showed broad negative correlations with farmland and positive correla-
tions with grassland (Table 5). The relatively high abundance of shredders is a typical
characteristic of streams within forested regions (Vannote et al. 1980; Ward and Stanford
1983). Miserendino and Masi (2010) also found that shredder richness was consistently
higher within forest-dominated areas. The relative abundance of predators showed wide,
negative correlations with farmland and village, and positive correlations with forest at
most riparian widths (Table 5). This might be because stoneflies comprised the highest
percentages of predators, or because the legacy of human disturbance has caused the cor-
responding cascades through the food web.
Second, our study indicated that a considerable number of water quality parameters
and macroinvertebrate indices were significantly correlated with land uses at different
Journal of Freshwater Ecology 323
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

riparian corridor widths (Tables 3, 4, and 5; Figure 5). The importance of the riparian
landscape attributes that affect the water quality and macroinvertebrate communities is
well documented (Dudgeon 1988; Naiman et al. 1993; Potter et al. 2005; Rios and Bailey
2006; Hutchens et al. 2009; Miserendino and Masi 2010). Riparian revegetation has been
used as one of the main techniques for the protection of stream ecosystems (Milner and
Gloyne-Phillips 2005; Carline and Walsh 2007; Palmer et al. 2007; Becker and Robson
2009). Those findings were comparable with the results of our study as widely positive
correlations between farmland and TDS and EC, and widely negative correlations be-
tween grassland and EC and TDS (Table 3). However, the effects of forest and village on
the pH and DO were only significant at relatively large riparian widths. The effects of
farmland on TN and villages on TP were all significant at riparian corridors most closely
adjacent to the stream channel. The correlations between sediment input and substratum
characteristics and the adjacent riparian corridors have been documented for widths rang-
ing from a narrow 3 m (Robinson et al. 1996) to 30 m (Sponseller et al. 2001). Except
for the relative abundance of predators and shredders that showed wide correlations with
land uses at different riparian widths, the other indices were mostly significant at the rela-
tively small and medium sizes of riparian corridors. At 25, 50, and 100 m riparian corri-
dors, farmland was a strong indicator of taxa richness of caddisflies, relative abundance
of Chironomidae, and ASPT index (Tables 4 and 5). The general variation in the macroin-
vertebrate composition was also confirmed at relatively small (50 m) to medium widths
(200 m; Figure 5). Within a similar range of riparian corridors, the village was a strong
negative indicator for biodiversity indices of Shannon’s index, Margalef’s index, and
Pielou’s evenness. In contrast, at only the 500-m riparian corridor width were the villages
a strong indicator for total individuals and forest a strong indicator for relative abundance
of filterers.
Despite findings of broad correlations between the land uses and water quality
(Table 3) and the significant effects of land uses on macroinvertebrates (Tables 4 and 5;
Figure 5), a one-time sampling, as conducted in this study, is inadequate for assessing
long-term water quality and reflecting the seasonal variation of macroinvertebrates. In or-
der to guide local riparian restoration, long-term measurement of water quality and sea-
sonal measurement of macroinvertebrates would be necessary to determine suitable
widths for riparian buffers. However, there is still little scientific and management guid-
ance about how to restore the riparian corridors of streams. Stream restoration now repre-
sents a multibillion-dollar industry (Brooks and Lake 2007) with riparian management as
one of the most common techniques, although its effectiveness for recovery of stream bi-
ota is still uncertain (Palmer et al. 2007). Harding et al. (1998) found that 50 years of ri-
parian reforestation did not recover the stream to its pre-disturbed biotic condition in a
western North Carolina watershed. By applying riparian revegetation techniques, Becker
and Robson (2009) found that eight years was the minimum time duration necessary to
see improvements in macroinvertebrate assemblages.
In this research, the one-time sampling of water quality factors and macroinvertebrate
indices reflected significant correlations with different types of land uses from relatively
small (25–50 m) to medium widths (100–200 m). According to our analysis, the farmland
within 200 m of the stream produced the most significant changes in aquatic ecosystems,
including input of dissolved solids and nitrogen, with predictable consequences for mac-
roinvertebrate communities. These effects, however, are still extremely low according to
the national water quality standards. The CCA results indicated that 50 m of forest or
200 m of grassland may provide a suitable riparian buffer for the protection of macroin-
vertebrates. However, due to the lack of seasonal data, determination of the most suitable
324 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

widths of riparian buffers requires further research. Changes in riparian land uses in the
mountain streams are a long-term issue in China. This paper shows that while the stream
ecosystem is particularly sensitive to anthropogenic influence, the condition of the stream
system can potentially be improved with a focus on the protection and revegetation of ri-
parian corridors.
Acknowledgements
This study was supported by the Central Institute Basic Foundation of Scientific Researches forPublic Services (2010KYYW06) and Chinese Major Science and Technology Program for WaterPollution Control and Management (2012ZX07501-001). We greatly appreciate the reviews ofDr Young-Seuk Park and Dr Fengqing Li for their thoughtful and constructive suggestions for thediscussion.
References
Allan JD. 2004. Landscapes and riverscapes: the influence of land use on stream ecosystems. An-nual Review of Ecology, Evolution, and Systematics. 35:257–284.
Allan JD, Castillo MM. 2007. Stream ecology: structure and function of running waters. 2nd ed.The Netherlands: Springer.
Allan JD, Erickson DL, Fay J. 1997. The influence of catchment land use on stream integrity acrossmultiple spatial scales. Freshwater Biology. 37:149–161.
Armitage PD, Moss D, Wright JF, Furse MT. 1983. The performance of a new biological waterquality score system based on macroinvertebrates over a wide range of unpolluted running-water sites. Water Research. 17:333–347.
Barbour MT, Gerritsen J, Snyder BD, Stribling JB. 1999. Rapid bioassessment protocols for use instreams and wadeable rivers: periphyton, benthic macroinvertebrates, and fish. 2nd ed.Washington (DC): US Environmental Protection Agency, Office of Water.
Becker A, Robson BJ. 2009. Riverine macroinvertebrate assemblages up to 8 years after riparianrestoration in a semi-rural catchment in Victoria, Australia. Marine and Freshwater Research.60:1309–1316.
Brooks SS, Lake PS. 2007. River restoration in Victoria, Australia: change is in the wind, and nonetoo soon. Restoration Ecology. 15:584–591.
Burton TM, Likens GE. 1973. The effect of strip-cutting on stream temperatures in the HubbardBrook Experimental Forest, New Hampshire. Bioscience. 23:433–435.
Carline RF, Walsh MC. 2007. Responses to riparian restoration in the Spring Creek watershed, cen-tral Pennsylvania. Restoration Ecology. 15:731–742.
Chinese Ministry of Environmental Protection (MEP), P.R. China. 2002. Environmental qualitystandards for surface water (GB3838-2002).
Cohen P, Andriamahefa H, Wasson JG. 1998. Towards a regionalization of aquatic habitat: distribu-tion of mesohabitats at the scale of a large basin. Regulated Rivers: Research and Management.14:391–404.
Cooper CM. 1993. Biological effects of agriculturally derived surface water pollutants on aquaticsystems – a review. Journal of Environmental Quality. 22(3):402–408.
Cummins KW, Klug MJ. 1979. Feeding ecology of stream invertebrates. Annual Review of Ecol-ogy and Systematics. 10:147–172.
Dance KW, Hynes HBN. 1980. Some effects of agricultural land use on stream insect communities.Environmental Pollution Series A, Ecological and Biological. 22:19–28.
Dodd RC, Cunningham PA, Tippett JP, Curry RJ, Stichter SJ, McMahon G. 1994. Watershedassessment in the Albemarle-Pamlico region. Proceedings: Watershed ’93: A National Confer-ence on Watershed Management; 1993 Mar 21–24; Washington (DC): US Bureau of Reclama-tion. p. 735–741.
Dudgeon D. 1988. The influence of riparian vegetation on macroinvertebrate community structurein four Hong Kong streams. Journal of Zoology. 216:609–627.
Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, Chapin FS, Coe MT, Daily GC,Gibbs HK, et al. 2005. Global consequences of land use. Science. 22:570–574.
Journal of Freshwater Ecology 325
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Gergel SE, Turner MG, Miller JR, Melack JM, Stanley EH. 2002. Landscape indicators of humanimpacts to riverine systems. Aquatic Sciences. 64:118–128.
Harding JS, Benfield EF, Bolstad PV, Helfman GS, Jones EBD. 1998. Stream biodiversity: theghost of land use past. Proceedings of the National Academy of Sciences of the USA.95:14843–14847.
Harding JS, Young RG, Hayes JW, Shearer KA, Stark JD. 1999. Changes in agricultural intensityand river health along a river continuum. Freshwater Biology. 42:345–357.
Hellawell JM. 1986. Biological indicators of freshwater pollution and environmental management.London: Elsevier Applied Science Publications Ltd. p. 451
Hill WR, Mulholland PJ, Marzolf ER. 2001. Stream ecosystem responses to forest leaf emergencein spring. Ecology. 82:2306–2319.
Hutchens Jr JJ, Schuldt JA, Richards C, Johnson LB, Host GE, Breneman DH. 2009. Multi-scalemechanistic indicators of Midwestern USA stream macroinvertebrates. Ecological Indicators.9:1138–1150.
Kratzer EB, Jackson JK, Arscott DB, Aufdenkampe AK, Dow CL, Kaplan LA, Newbold JD,Sweeney BW. 2006. Macroinvertebrate distribution in relation to land use and water chemistryin New York City drinking-water-supply watersheds. Journal of the North American Bentho-logical Society. 25:954–976.
Liu MZ, Zhang HJ. 2012. Study on variation characteristics of climate in Benxi County,Liaoning Province in recent 52 years. Journal of Anhui Agricultural Sciences. 40:987–989.Chinese.
Magurran AE. 2004. Measuring biological diversity. Oxford (UK): Blackwell Publishing. p. 260.McCune B, Mefford MJ. 1999. PC-ORD. Multivariate analysis of ecological data [M], Version 4.
Oregon: MjM Software Design.Merritt RW, Cummins KW. 1996. An introduction to the aquatic insects of North America. 3rd ed.
Dubuque (IA): Kendall/Hunt Publishing Company. p. 862.Meyer JL, Strayer DL, Wallace JB, Eggert SL, Helfman GS, Leonard NE. 2007. The contribution of
headwater streams to biodiversity in river networks. Journal of the American Water ResourcesAssociation. 43:86–103.
Meyer JL, Wallace JB. 2001. Lost linkages in lotic ecology: rediscovering small streams. In: PressMC, Huntly N, Levin S, editors. Ecology: achievement and challenge. The 41st Symposium ofthe British Ecological Society sponsored by the Ecological Society of America; 2000 Apr10–13; Orlando(FL): Blackwell Science, p. 295–317.
Milner M, Gloyne-Phillips IT. 2005. The role of riparian vegetation and woody debris in the develop-ment of macroinvertebrate assemblages in streams. River Research and Applications. 21:403–420.
Miserendino ML, Masi CI. 2010. The effects of land use on environmental features and functionalorganization of macroinvertebrate communities in Patagonian low order streams. EcologicalIndicators. 10:311–319.
Moore AA, Palmer MA. 2005. Invertebrate biodiversity in agricultural and urban headwaterstreams: implications for conservation and management. Ecological Applications. 15(4):1169–1177.
Morse JC, Yang LF, Tian LX. 1994. Aquatic insects of China useful for monitoring water quality.Nanjing: Hohai University Press. p. 570.
Naiman RJ, Decamps H, Pollock M. 1993. The role of riparian corridors in maintaining regionalbiodiversity. Ecological Applications. 3:209–212.
Norton MM, Fisher TR. 2000. The effects of forest on stream water quality in two Coastal Plainwatersheds of the Chesapeake Bay. Ecological Engineering. 14:337–362.
Palmer M, Allan JD, Meyer J, Bernhardt ES. 2007. River restoration in the twenty-first century: dataand experiential knowledge to inform future efforts. Restoration Ecology. 15(3):472–481.
Paul MJ, Meyer JL. 2001. Streams in the urban landscape. Annual Review of Ecology and System-atics. 32:333–365.
Potter KM, Cubbage FW, Schaberg RH. 2005. Multiple-scale landscape predictors of benthicmacroinvertebrate community structure in North Carolina. Landscape and Urban Planning.71:77–90.
Prud’homme BA, Greis JG. 2002. Best management practices in the South. In: Wear, DN, Greis JG,editors. Southern forest resource assessment. General Technical Report SRS-53. Asheville(NC): US Department of Agriculture, Forest Service, Southern Research Station. p. 519–535.
326 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Quinn JM, Steele GL, Hickey CW, Vickers ML. 1994. Upper thermal tolerances of twelve NewZealand stream invertebrate species. New Zealand Journal of Marine and Freshwater Research.28:391–397.
Reid DJ, Quinn JM, Wright-Stow AE. 2010. Responses of stream macroinvertebrate communitiesto progressive forest harvesting: influences of harvest intensity, stream size and riparian buffers.Forest Ecology and Management. 260:1804–1815.
Richardson JS, Danehy RJ. 2007. A synthesis of the ecology of headwater streams and their riparianzones in temperate forests. Forest Science. 53:131–147.
Rios SL, Bailey RC. 2006. Relationship between riparian vegetation and stream benthic communi-ties at three spatial scales. Hydrobiologia. 553:153–160.
Robinson CA, Ghaffarzadeh M, Cruse RM. 1996. Vegetative filter strip effects on sediment concen-tration in cropland runoff. Journal of Soil and Water Conservation. 51:227–230.
Rosenberg DM, Resh VH, editors. 1993. Freshwater biomonitoring and benthic macroinvertebrates.New York: Chapman and Hall. p. 504.
Roy AH, Rosemond AD, Paul MJ, Leigh DS, Wallace JB. 2003. Stream macroinvertebrate responseto catchment urbanisation (Georgia, U.S.A.). Freshwater Biology. 48:329–346.
Scarsbrook MR, Halliday J. 1999. Transition from pasture to native forest land-use along streamcontinua: effects on stream ecosystems and implications for restoration. New Zealand Journalof Marine and Freshwater Research. 33:293–310.
Schulz R, Liess M. 1999. A field study of the effects of agriculturally derived insecticide input onstream macroinvertebrate dynamics. Aquatic Toxicology. 46:155–176.
Shandas V, Alberti M. 2009. Exploring the role of vegetation fragmentation on aquatic conditions:linking upland with riparian areas in Puget Sound lowland streams. Landscape and Urban Plan-ning. 90:66–75.
Shieh SH, Ward JV, Kondratieff BC. 2003. Longitudinal changes in macroinvertebrate production ina stream affected by urban and agricultural activities. Archiv fur Hydrobiologie. 157:483–503.
Short RA, Ward JV. 1980. Leaf litter processing in a regulated Rocky Mountain stream. CanadianJournal of Fisheries and Aquatic Sciences. 37:123–127.
Sponseller RA, Benfield EF, Valett HM. 2001. Relationships between land use, spatial scale andstream macroinvertebrate communities. Freshwater Biology. 46:1409–1424.
Stevens MHH, Cummins KW. 1999. Effects of long-term disturbance on riparian vegetation and in-stream characteristics. Journal of Freshwater Ecology. 14:1–17.
Storey RG, Cowley DR. 1997. Recovery of three New Zealand rural streams as they pass throughnative forest remnants. Hydrobiologia. 353:63–76.
Sweeney BW. 1993. Effects of streamside vegetation on macroinvertebrate communities of WhiteClay Creek in eastern North America. Proceedings of the Academy of Natural Sciences ofPhiladelphia. 144:291–340.
Sweeney BW, Vannote RL, Dodds PJ. 1986. The relative importance of temperature and diet tolarval development and adult size of the winter stonefly, Soyedina carolinensis (Plecoptera:Nemouridae). Freshwater Biology. 16:39–48.
Tran CP, Bode RW, Smith AJ, Kleppel GS. 2010. Land-use proximity as a basis for assessingstream water quality in New York State (USA). Ecological Indicators. 10:727–733.
Utz RM, Hilderbrand RH, Boward DM. 2009. Identifying regional differences in thresholdresponses of aquatic invertebrates to land cover gradients. Ecological Indicators. 9:556–567.
Vallania A, Corigliano MC. 2007. The effect of regulation caused by a dam on the distribution ofthe functional feeding groups of the benthos in the subbasin of the Grande River (San Luis,Argentina). Environmental Monitoring and Assessment. 124:201–209.
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE. 1980. The river continuum con-cept. Canadian Journal of Fisheries and Aquatic Sciences. 37:130–137.
Vannote RL, Sweeney BW. 1980. Geographic analysis of thermal equilibria: a conceptual modelfor evaluating the effect of natural and modified thermal regimes on aquatic insect communi-ties. The American Naturalist. 115:667–695.
V€or€osmarty CJ, McIntyre PB, Gessner MO, Dudgeon D, Prusevich A, Green P, Glidden S, BunnSE, Sullivan CA, Reidy Liermann C, et al. 2010. Global threats to human water security andriver biodiversity. Nature. 467:555–561.
Ward JV, Stanford JA. 1983. The serial discontinuity concept of lotic ecosystems. In: Fontaine TD,Bartell SM, editors. Dynamics of lotic ecosystems. Ann Arbor (MI): Ann Arbor Sciences Pub-lishers Inc. p. 29–42.
Journal of Freshwater Ecology 327
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13

Weigel BM, Lyons J, Paine LK, Dodson SI, Undersander DJ. 2000. Using stream macroinverte-brates to compare riparian land use practices on cattle farms in southwestern Wisconsin. Journalof Freshwater Ecology. 15:93–106.
Weigel BM, Wang LZ, Rasmussen PW, Butcher JT, Stewart PM, Simon TP, Wiley MJ. 2003. Rel-ative influence of variables at multiple spatial scales on stream macroinvertebrates in the North-ern Lakes and Forest ecoregion, U.S.A. Freshwater Biology. 48:1440–1461.
Wipfli MS, Richardson JS, Naiman RJ. 2007. Ecological linkages between headwaters and down-stream ecosystems: transport of organic matter, invertebrates, and wood down headwater chan-nels. Journal of the American Water Resources Association. 43:72–85.
Wright JF. 1995. Development and use of a system for predicting the macroinvertebrate fauna inflowing waters. Australian Journal of Ecology. 20:181–197.
Yang LZ, Feng YF, Shi WM, Xue LH, Wang SQ, Song XF, Chang ZZ. 2013. Review of the advan-ces and development trends in agricultural non-point source pollution control in China. ChineseJournal Eco-Agriculture. 21:96–101. Chinese.
Zhang WL, Wu SX, Ji HJ, Kolbe H. 2004. Estimation of agricultural non-point source pollution inChina and the alleviating strategies. I. Estimation of agricultural non-point source pollution inChina in early 21 century. Scientia Agricultura Sinica. 37:1008–1017. Chinese.
Zimmerman GM, Goetz H, Mielke Jr PW. 1985. Use of an improved statistical method for groupcomparisons to study effects of prairie fire. Ecology. 66:606–611.
328 Y. Zhang et al.
Dow
nloa
ded
by [
Uni
vers
ity o
f Il
linoi
s at
Urb
ana-
Cha
mpa
ign]
at 1
0:35
28
Sept
embe
r 20
13





























