Embed Size (px)
Citation preview

REGULATION OF VERTEBRATE PLANAR CELL POLARITY
by
Jason Trinh
A thesis submitted in conformity with the requirements
for the degree of Master of Science
Graduate Department of Molecular Genetics
University of Toronto
© Copyright by Jason Trinh 2009

ii
REGULATION OF VERTEBRATE PLANAR CELL POLARITY Jason Trinh Master of Science 2009 Graduate Department of Molecular Genetics, University of Toronto Abstract
Planar cell polarity (PCP) provides positional information to a field of cells, coordinating
the orientation of polarized structures or the direction of polarized cell movements. An
evolutionarily conserved signalling pathway regulates PCP, however, the cue that
establishes PCP is unknown. There is a strong precedent for Wnt signalling to act as the
cue to establish PCP. Here I perform in vivo assays of cell polarity to examine the role of
non-canonical Wnt signalling in regulating PCP, using zebrafish neural progenitor cells
and asymmetric membrane localization of GFP-Prickle (a PCP cytoplasmic effector
molecule) as a model system. My preliminary evidence suggests Wnt4a provides
positional information to cells in the neural tube. In addition, using a membrane-yeast-
two-hybrid approach to discover novel regulators of PCP, I identified Ring Finger 41 as a
new binding partner to Van-gogh-like-2 (an essential PCP signalling molecule) and a
novel regulator of vertebrate PCP.

iii
Acknowledgements I would like to thank Dr. Brian Ciruna for his supervision, guidance and support
throughout my graduate school studies. I would like to thank my committee members,
Dr. Julie Brill and Dr. Helen McNeill for their guidance and expertise. Thank you to the
Ontario Graduate Scholarship Program and SickKids Research Training Centre for
providing funding. In addition, thank you to: Dr. Dan Voskas for being a mentor and
friend, Dr. Dani Gelinas for her continuous encouragementand support, Sasha Fernando
for his expert care of the zebrafish as well as all past and present members of the Ciruna
Lab. I would like to thank my family for supporting me without really understanding
what I do. Finally, I would like to thank Andrea for being my fellow passenger on this
adventure. You are my light.

iv
Table of Contents CHAPTER 1: BACKGROUND 1 1.0: Introduction 1 1.1: Planar Cell Polarity Signalling 2 1.2: Regulation of Planar Cell Polarity 5 1.3: Vertebrate Planar Cell Polarity Regulates Morphogenesis 8 1.4: Vertebrate Planar Cell Polarity Signalling 12 1.5: Regulation of Vertebrate Planar Cell Polarity 16 1.5.1: Non-canonical Wnt signalling regulates planar cell polarity 16 1.5.2: Vertebrate specific regulators of planar cell polarity are associated with Wnt signalling 17 1.5.3: The search for a global cue that establishes planar cell polarity 19 1.6: References 20 CHAPTER 2: INVESTIGATING THE ROLE OF WNT SIGNALLING IN VERTEBRATE PLANAR CELL POLARITY 24 2.0: Introduction 24 2.1: Results 30
2.1.1: Heterochronic cell transplantation is possible 30 2.1.2: Characterization of Wnt clones 33 2.1.3: Localized ectopic Wnt4a expression is able to alter neural
progenitor cell behaviour 38 2.2: Discussion 43
2.2.1: Anterior-posterior Wnt4a gradient provides positional information to neural progenitor cells 43
2.3: Methods 46 2.3.1: Materials 46 2.3.2: Zebrafish embryo microinjection 47 2.3.3: Cell transplants 48 2.3.4: Microscopy 48 2.3.5: Transgenesis 48
2.4: References 49 CHAPTER 3: DISCOVERY OF NOVEL REGULAORS OF PLANAR CELL POLARITY 3.0: Introduction 51 3.1: Results 54 3.1.1: Characterization of baits 54 3.1.2: Analysis of screen hits 56 3.1.3: Validation of MYTH screen 62 3.1.4: Ectopic expression of RNF41 disrupts PCP signalling 65 3.1.5: RNF41 subcellular localization suggests a new link between PCP and cilia 71

v
3.2: Discussion 72 3.2.1: RNF41 may regulate planar cell polarity through Vangl2 72 3.2.2: RNF41 localizes to the basal body and may regulate non-canonical signalling 74 3.3: Methods 74 3.3.1: Materials 74 3.3.2: Bait/Prey vector construction by homologous recombination gap repair 75 3.3.3: NubG/I Test 76 3.3.4: MYTH 76 3.3.5: Zebrafish microinjection 77 3.3.6: Immunoprecipitation and western blotting 78 3.2.7: in situ hybridization 79 3.2.8: Microscopy 79 3.4: References 80
CHAPTER 4: FUTURE DIRECTIONS 4.0: Preliminary results suggest Wnt4a provides positional information to neural progenitor cells 81 4.1: Addressing the instructive/permissive role of non-canonical Wnts 82 4.2: The role of RNF41 in regulating planar cell polarity 85 4.3: The cue that establishes planar cell polarity remains elusive 86 4.4: References 88

vi
List of Figures and Tables Figure 1 Planar cell polarity generates positional information 3 Figure 2 Models of PCP establishment 6 Figure 3 Convergent extension movements 10 Figure 4 Wnt signalling 13 Figure 5 Instructive versus permissive functions of Wnt 25 Figure 6 GFP-Pk as a maker of PCP 28 Figure 7 Heterochronic cell transplants are possible and do not disrupt
PCP 32 Figure 8 Injected Wnt mRNA is expressed and secreted 34 Figure 9 Ectopic non-canonical Wnt expression induces morphological defects 36 Figure 10 Ectopic Wnt4a expression induces neural tube defects with GFP-Pk puncta localized to the membrane 37 Figure 11 Neural progenitor cells respond differently to a Wnt4a clone with respect to its position 40 Figure 12 Transgenic approach to generate localized Wnt source in the neural tube 42 Figure 13 Wnt4a gradients may provide positional information to neural progenitor cells 44 Figure 14 Membrane yeast two hybrid (MYTH) 53 Table 1 Summary of bait construction 57 Figure 15 Characterization of baits 57-58 Table 2 MYTH screen hits 59 Figure 16 Analysis of hits 63 Figure 17 Validation of MYTH screen with known protein interactions 64 Figure 18 Co-immunoprecipitation shows that Vangl2 and RNF41 physically interact 66 Figure 19 Endogenous RNF41 expression 68 Figure 20 Ectopic RNF41 expression induces convergent extension defets 69 Figure 21 RNF41 regulates GFP-Pk localization 70 Figure 22 RNF41 localization suggests a role in regulating cilia 73 Figure 23 Temperature versus voltage standard curve 84 Figure 24 Local heat shock with a modified soldering iron is able to induce transgene expression in cells in the neural tube 84

1
CHAPTER 1: Background 1.0: Introduction
Cell polarity exists when cellular components have asymmetric characteristics at
different points within the cell. Potentially, proteins are localized or cellular structures are
formed in one region of the cell and not another. Many different cell types exhibit
individual or cellular polarization. For example, epithelial cells are polarized along the
apical-basal axis and mesenchymal cells are polarized in the direction of migration. During
animal development, cells are individually polarized, but often groups of cells within a
tissue are coordinated with the polarity of the tissue [1]. Tissue or planar polarity is
required to give cells positional information in the plane in order to position polarized
structures or undergo directed cell movement. [2]. Coordinated polarization across a field
of cells is called planar cell polarity (PCP). There are many examples of PCP in numerous
model systems. PCP has been shown to be required for proper alignment of hairs on
mammalian skin and on the Drosophila wing as well as coordinated mass movements of
cells during vertebrate embryonic development [1][3]. Early studies of PCP were
conducted in Drosophila, as it was one of the first animals where PCP was recognized
and can be found in adult tissues [2][4]. In Drosophila, mutations of PCP genes caused
disorganization of cuticle structures and the compound eye [2]. More specifically, wing
cells which are normally polarized along the proximal-distal axis, display swirly hair
patterns [2]. In the Drosophila eye, PCP mutants have defects in the organization of
photoreceptors [2]. Based on these phenotypes, mutants were identified and a group of
genes (core PCP genes) that are required in all polarized tissues was identified [4].

2
1.1: Planar Cell Polarity Signalling
The genetic control of PCP was first studied in the cuticle structures of
Drosophila, namely the hairs of the wing blade where loss-of function mutants displayed
swirly hair patterns [2]. A set of six genes were discovered, the core PCP genes that are
required in all tissues: frizzled (fz), which encodes a seven-pass transmembrane protein
with an extracellular cysteine-rich domain (CRD) which binds Wingless/Wnt protein;
dishevelled (dsh), which encodes a cytoplasmic adaptor protein that interacts with small
GTPases; van gogh (vang), which encodes a four-pass transmembrane protein that
interacts with Prickle (Pk), a cytoplasmic effector protein; flamingo (fmi), which encodes
a seven-pass transmembrane atypical cadherin; and diego (dgo) which encodes a
cytoplasmic effector ankryin-repeat protein. In the wing, the core PCP genes are required
for placement of hairs that are produced by each cell [1]. Normally, the hairs grow on the
apical surface of the distal side of the wing cell, growing out distally (Figure 1A). Loss-
of-function mutations in the core PCP genes disrupt the organization of hairs resulting in
growth of hairs from the centre of the apical surface of the wing cell [1].
Cells within a plane require PCP for positional information to orient polarized
structures like hairs. The generation of positional information is possibly the result of
asymmetric sub-cellular localization of the core PCP gene products (Figure 1B). In the
Drosophila wing, before the onset of PCP signalling, all core PCP proteins are localized
uniformly around the apical-lateral membrane, partially overlapping with cellular
junctions [2]. PCP signalling induces asymmetric localization of the PCP components
into two complexes across the proximal-distal axis. At the distal side, Fz recruits Dsh

3
Figure 1: Planar Cell Polarity Generates Positional Information (A) PCP genes give positional information to a field of cells required to orient a polarized structure. In the Drosophila wing, hairs are formed on the distal side of the cell, where in core PCP mutants, cells are not polarized in the plane and hairs are formed in the centre of the cell. (B) The core PCP components generate positional information from asymmetric localization of the core PCP components inside the cell. Fz, Dsh and Dgo act positively to promote PCP are localized to the distal side of the wing cell, while negative factors are localized to the proximal side of the wing cell.

4
and Dgo and acts positively, promoting downstream activation of Dsh effectors and
ultimately generation of a polarized structure. At the proximal side, Vang and Pk are
localized and act negatively to limit Dsh activation [5]. The only core PCP protein that is
not asymmetrically localized is Fmi, as it localizes to both Fz and Vang. The localization
of the core PCP proteins appears to be dependent on the transmembrane core PCP
proteins. Absence of Fz, Vang or Fmi results in loss of or strongly reduced apical
localization of the other core PCP components. Loss of Dsh, Pk or Dgo alone does not
affect apical localization of the other core PCP proteins [4].
Fz and Vang are able to induce polarization of PCP components in both cell-
autonomous and non-autonomous fashion. Fz and Vang are required for apical
localization of the core PCP components and recruitment of downstream effectors to
distal and proximal sides. In addition, Fz and Vang can also alter polarization and
localization of PCP components in neighbouring cells, suggesting that Fz/Vang are
involved in cell-cell communication and propagation of PCP establishment. Clonal
analysis of fz and vang in Drosophila has clearly demonstrated domineering non-
autonomy of fz and vang, which means that loss-of-function fz clones in the Drosophila
wing are able to affect the polarity of adjacent WT cells, resulting in hairs pointing
towards the fz clone. Similarly, loss-of-function of vang clones are able to change
polarity of neighbouring wing cells, however the hairs point away from the vang clone.
When examining other PCP genes like dsh or fmi, loss-of-function clones of dsh or fmi do
not act cell non-autonomously as indicated by normal polarization of hairs in adjacent
wildtype (WT) cells [4]. A possible mechanism for non-autonomy may be through direct
interaction of the Fz CRD and Vang, which have been shown to interact physically [6].

5
Alternatively, PCP propagation may be mediated through Fmi. Fmi is required for apical
localization of PCP components, but does not act non-autonomously to polarize
neighbouring cells. However, Fmi interacts homophilically in trans with Fmi on adjacent
cells to stabilize Fz and Vang complexes [4]. It has been proposed that Fmi-Fmi
interaction between cells activates Vang and represses Fz in the adjacent cell. Another
possibility is that Fmi is required to localize Fz and Vang in close proximity such that Fz
can directly interact with Vang. How PCP is propagated across a field of cells is still
unknown. One model suggests that complexes with the core PCP proteins form in trans
and propagate PCP in a ‘domino effect’ where polarization is initiated in a subset of cells
and is propagated across the plane through neighbouring cell-cell communication (Figure
2A) [7].
1.2: Regulation of Planar Cell Polarity
Polarization across a field of cells may occur by domineering non-autonomy of Fz
and Vang; however, another models suggest that a global cue may polarize the entire
plane (Figure 2B) [7]. Common to both models is an initial polarization event that either
begins the cascade of polarization mediated through cell-cell communication or acts to
polarize the entire plane. The identity of this initial polarization cue remains elusive. A
likely candidate is a diffusible cue that establishes PCP by acting on the core PCP
proteins. This cue most likely acts on Fz, Vang or Fmi since Pk, Dsh, Dgo act
downstream of Fz, Vang and Fmi [4]. There are no known soluble ligands of Vang or
Fmi; however, Fz is the receptor for the Wnt family [4]. Potentially Wnt proteins may
act as cue to establish

6
Figure 2: Models of PCP Establishment (A) Cell-Cell relay model: PCP is established and is propagated through a field of cells by domineering non-autonomy, or responding cells generate a polarizing cue that is secreted from one cell to the next. (B) Gradient model: PCP is established by a diffusible cue, and cells are polarized in the direction of the gradient.

7
PCP. However, clonal analysis of wingless (wg)/wnt mutants in the Drosophila abdomen,
revealed no changes in polarity of adjacent cells with all seven Wg/Wnt proteins tested
individually [8]. Unlike in the wing and compound eye, hedgehog signalling influences
PCP in the abdomen and may mask affects of mutant wg/wnt clones or possibly Wg/Wnt
proteins are functionally redundant [4]. Gain-of-function experiments with dWnt4 can
alter cellular orientation of PCP in the wing, however Drosophila Wnt proteins activate
other downstream pathways and it is difficult to distinguish a direct from indirect effects
on PCP signalling. Possibly there is an undiscovered Fz ligand that acts to polarize cells
[4]. Models for PCP establishment often include a diffusible cue termed Factor X. Factor
X could act in a long-ranged gradient produced by a few cells and spread across the
plane, or as a short-range cue generated throughout the field of cells, but in varying
amount based on the position of the cells [5].
In addition to the core PCP proteins, three additional genes fat (ft), dachsous (ds)
and four-jointed (fj), generate similar mutant phenotypes such as swirly hair and
disorganized photoreceptors. ft and ds are atypical cadherins that interact in trans, while fj
encodes a type II membrane protein localized to the Golgi. The role of the Ft-Ds group is
unclear, but there is evidence to support an upstream and parallel role to the core PCP
group. Ds and Fj are expressed in a gradient in the eye, wing and abdomen, creating a Ft
activity gradient. The only identified downstream effector of the Ft-Ds signalling is
atrophin, a transcriptional co-repressor downstream of Ft [9]. In the Drosophila wing,
ft/ds mutant clones alter Dsh, Pk and Fmi localization, whereas the localization of Ft and
Ds is unaltered in fz mutant clones [10]. Based on the Ft-Ds expression and epistasis data,
the Ft-Ds group could act upstream of the core PCP components and establish polarity.

8
However, it is unclear if Ft or Atrophin acts directly on the core PCP components. There
is also evidence to support a parallel role between the core PCP and Ft-Ds group. Clonal
analysis in the Drosophila abdomen shows that gain- and loss-of-function ft or ds clones
are able to repolarize neighbouring mutant fz and fmi cells [11]. In the larvae cuticle, PCP
defects are only observed when both the core PCP and Ft-Ds groups are affected. In
addition, fmi and ft double mutants have a more severe phenotype than the single
mutants, suggesting the core PCP and Ft-Ds group act in parallel [11].
1.3: Vertebrate Planar Cell Polarity Regulates Morphogenesis
The study of PCP in vertebrates is complex relative to the fly, as most PCP genes
have numerous homologues in vertebrates as compared to only one homologue in
Drosophila. For example, although there are five Drosophila frizzled genes, only one (fz)
regulates tissue polarity, whereas the other four show similar phenotypes to wingless (wg)
mutants [12]. In mice, there are ten frizzled genes and fz3 and fz6 have been implicated
in, but not dedicated to, regulation of PCP [4][13]. In addition, some homologues have
non-overlapping expression patterns, while others have overlapping expression patterns
and are functionally redundant. As a result, creating loss-of-function mutants is difficult,
especially if numerous genes are required to achieve a full PCP defect [4].
The same genes that control PCP in Drosophila are conserved in vertebrates,
where they give positional information to a field of cells to coordinate polarized
structures. For example, PCP is required for proper orientation of hairs on the mouse coat
[2]. In addition to the planar polarization of epithelia cells, vertebrate PCP can also be
extended to the polarization of mesenchymal cells. During vertebrate embryogenesis,

9
hundreds of individual cells move together in a highly organized pattern [3]. PCP has
been shown to regulate polarized cell movement that shapes the vertebrate body plan
[14]. One type of mass movement controlled by PCP is convergent extension (CE),
which results in narrowing of one axis and elongation of an orthogonal axis (Figure 3).
CE was initially studied by Xenopus researchers examining morphogenesis of the
posterior mesoderm and neuroectoderm. It was observed that the mesoderm cells
underwent radial and mediolateral (ML) intercalation, while the neuroectoderm cells
predominately performed ML intercalation [15]. Similar observations have been made in
zebrafish with the movements of mesoderm and neuroectoderm during gastrulation and
neurulation respectively [16][17].
The study of vertebrate PCP in zebrafish embryos combines several features that
are ideal for cellular and molecular analysis of PCP. Zebrafish embryos develop rapidly
externally and are easily accessible for embryonic manipulations such as cell
transplantations and microinjections. Simultaneous knock-down of many genes can be
performed with injection of anti-sense morpholino oligonucleotides. Zebrafish embryos
are optically clear making these embryos ideal for real-time in vivo imaging at the
morphological and cellular levels. Although zebrafish is hindered likewise with other
vertebrate models with redundancy and numerous homolouges of PCP genes, the distinct
advantages make zebrafish an amenable vertebrate model to study PCP.
During early zebrafish gastrulation, lateral mesoderm cells converge to the dorsal
side by directed migration. On the dorsal side, the axial mesoderm undergoes ML
intercalations that narrows the ML axis extends the anterior-posterior (AP) axis (Figure
3A). The presomitic mesoderm undergoes extension of the AP axis as a result of radial

10
B. Coordinated radial/medial Intercalation
Figure 3: Convergent Extension Movements Convergent extension movements result in anisotropic tissue movements where narrowing of one axis and elongation of an orthogonal axis occurs. A: Mediolateral (ML) Intercalation occurs within the same plane, where cells converge along one axis, resulting in extension in an orthogonal-planar axis. B: Coordinated radial/medial intercalation occurs where cells enter the plane and immediately medially intercalate. Convergence of two planes occurs, with extension of an orthogonal axis. C: Radial intercalation occurs where cells enter the plane and directly intercalate, extending the orthogonal axis. This results in the same mass movement as coordinate radial.medial intercalation, where there is convergence of two cell planes and extension of the orthogonal axis. D: Radial/ML intercalation occurs when cells enter the plane and separate medial cells, expanding the mediolateral axis. Radial/ML intercalations extend the orthogonal axis to coordinate radial/medial and radial intercalations. Adapted from Yin et al. JCB, 2008.
A. Mediolateral Intercalation
C. Radial Intercalation
D. Radial/Mediolateral Intercalation

11
intercalation, more specifically three types of polarized movements that result in
anisotropic tissue expansion [16]. The first type of anisotropic radial intercalation is
coordinated radial/medial intercalation, where a cell enters the plane and immediately
intercalates medially, extending the AP axis (Figure 3B) [16]. The second type is
radial/AP intercalation, where a cell intercalates directly into the plane and extends the
AP axis (Figure 3C) [16]. Finally, the third is radial/ML intercalation, where a cell enters
the plane and separates the medial cells, expanding the ML axis, counteracting CE
movements (Figure 3D) [16]. The majority of presomitic mesoderm during gastrulation
has been observed to undertake radial intercalations that result in extension of the AP
axis, where the majority of cells appear to undergo radial/AP intercalation [16]. Defects
in PCP signalling result in an embryo with shortened AP axis and broaden ML axis. More
specifically, the frequencies of cell intercalations are altered in PCP mutants, resulting
from impaired ML intercalation of axial mesoderm, and the anteroposterior bias of radial
intercalations is lost in the presomitic mesoderm [16][18].
During zebrafish neurulation, a single-cell layer of neuroepithelium called the
neural plate folds inwards forming a solid structure termed the neural keel [19]. The
neural keel eventually rounds up and detaches from the adjacent epidermis to form the
neural rod [19]. The neural tube is formed by retraction of the apical surfaces of cells at
the midline to form a lumen [19]. During neural keel stage, neural progenitor cells
(NPCs) undergo stereotypic cell divisions where dividing NPCs equally contribute
daughter cells to both sides of the developing neural tube [17]. More specifically, as a
NPC begins to divide, it rounds up its cell body and approaches the midline, while
maintaining contact with the basal side of the neural keel [17]. At the midline, the NPC

12
divides and the apical daughter cell directs protrusions towards the contralateral side
along the mediolateral axis, while the basal daughter cell reinserts into the ipsilateral side
[17]. The apical daughter NPC intercalates between neighbouring cells, resulting in
extension along the AP axis and convergence of the ML axis. In zebrafish vang (vangl2)
mutants, loss of PCP results in decreased CE and a neural tube defect, where NPCs
accumulate ectopically at the midline. PCP signalling is required during neurulation to
accommodate the act of cell division. When a NPC divides, the daughter loses contact to
the neuroepithelium; it requires PCP to provide positional information to direct cellular
protrusions and intercalate to the contralateral side. In vangl2 mutants where PCP is
absent, apical daughter cells have no directional information. They fail to form stable
protrusions and cannot intercalate, instead the daughter cell remains in the position where
it was born. The result is an accumulation of daughter cells at the midline [17].
1.4: Vertebrate Planar Cell Polarity Signalling
In vertebrates, there are approximately 10 frizzled genes, where fz7 is best
characterized in zebrafish and acts in the PCP pathway [20][21][22]. There are three
pathways that have been characterized downstream of Fz: canonical Wnt, Wnt-Ca++ and
non-canonical Wnt/PCP signalling (Figure 4). The canonical Wnt signalling pathway
regulates patterning by controlling transcription. In the absence of Wnt, ß-catenin (ß-
cat), a transcriptional activator, is degraded by the destruction complex. The destruction
complex consists of two scaffold proteins, Axin and Adenomatous polyposis coli (APC),
which bind ß-cat, and two kinases, Glycogen synthase kinase 3 (GSK3) and Casein
kinase 1α (CK1α). The two kinases phosphorylate ß-cat, which is then recognized by a

13
Figure 4: Wnt Signalling (A) Canonical Wnt signalling is ß-catenin dependent and induces a transcriptional response. (B) Wnt/Ca++ signalling is vertebrate specific and results in an increase in intracellular calcium. (C) Non-canonical Wnt/PCP signalling is ß-catenin independent and affects cell polarity.

14
Cullin E3 ubiquitin ligase that targets ß-cat for degradation. In the absence of canonical
Wnt signalling, there are low levels of ß-cat inside the cell and as a result target genes
remain repressed by Groucho, a transcriptional repressor that binds to Lymphoid
enhancer factor (LEF) and T cell factor (TCF), co-transcription factors. In the presence of
canonical Wnt signalling, Wnt binds Fz and its co-receptor Low-density-lipoprotein
receptor-related protein (LRP) 5/6 forming a complex at the cell surface. Inside the cell,
the Fz-LRP complex activates Dsh and recruits Axin to the LRP co-receptor. This
prevents formation of the destruction complex and ß-cat is not degraded. Stabilized ß-cat
accumulates in the cytoplasm and begins to translocate into the nucleus, where it
displaces Groucho from LEF/TCF and promotes transcription of LEF/TCF targeted genes
[23]. In zebrafish, canonical Wnt signalling is responsible for formation of the dorsal-
ventral (DV) axis, where it plays two distinct roles to specify dorsal and ventral cell fates
[24]. Before the onset of gastrulation, maternal ß-cat is asymmetrically localized to the
future dorsal side of the embryo [24]. Canonical Wnt signalling is required for formation
of the dorsal organizer, where an increase or decrease of ß-cat causes dorsalization or
ventralization respectively [24]. Later, during gastrulation, canonical Wnt signalling is
also important for specifying ventral cell fates [24]. Wnt8 is expressed in a decreasing
ventral to dorsal gradient, where increases or decreases in Wnt8 can cause ventralization
or dorsalization respectively [24].
The Wnt/Ca++ signalling pathway is the least characterized of the three pathways
and appears to be vertebrate specific [4]. Fz is classified as a novel type of G-protein
coupled receptor, where signalling downstream of the receptor may occur through

15
trimeric G-proteins. Upon Wnt stimulation, phospholipase C mediated increase in
intracellular Ca++ levels lead to activation of Ca++/calmodulin dependent effectors [25].
Unlike the canonical Wnt signalling pathways, the non-canonical Wnt/PCP
signalling does not require transcriptional activity of ß-cat. In vertebrates, non-canonical
Wnt/PCP signalling controls CE during gastrulation and neurulation. Similar to
Drosophila PCP, Fz recruits Dsh and Diversin (homologue of Drosophila Diego) and
acts positively, promoting downstream activation of Dsh effectors. One effector is the
Rho family of small GTPases that affects cytoskeleton actin-dynamics [26]. Vangl2 and
Pk are asymmetrically localized opposite to Fz and Dsh and are thought to act negatively
to limit Dsh activation. Similar to Drosophila, positional information is generated
through asymmetric localization of the core PCP components in vertebrates. In zebrafish,
Dsh and Pk have been shown to localize asymmetrically in cells undergoing CE
movements. Fluorescent protein tagged-Dsh localizes to the posterior apical membrane in
mesoderm cells, whereas GFP-Pk localizes to the anterior apical membrane in NPC
[16][17]. Together, these data suggest that the vertebrate core PCP proteins are localized
across the AP axis. In Drosophila, the asymmetric localization of core PCP components
lies along the same axis as planar polarization. In the Drosophila wing, the core PCP
components are found along the proximal-distal axis, where Fz localization coincides
with hair formation [5]. Although in zebrafish, subcellular asymmetries exist within the
cell, the localization of PCP components is orthogonal to the axis of polarized movement,
more specifically, protrusion formation and intercalation occurs along the ML axis. There
may be a difference how PCP is interpreted between epithelial and mesenchymal cells. It
appears that asymmetries of the core PCP components are a universal hallmark of PCP

16
signalling; however, the positional information that is generated appears to be interpreted
differently in mesenchymal cells.
1.5: Regulation of Vertebrate Planar Cell Polarity
1.5.1: Non-canonical Wnt signalling regulates planar cell polarity
Although there is no clear Drosophilia Wg/Wnt implicated in PCP signalling,
there is strong evidence in zebrafish to support a role for a non-canonical Wnt protein to
regulate PCP. First, several zebrafish mutants exhibit reduced CE movements without
major patterning defects [15]. The trilobite (tri), silberblick (slb), pipetail (ppt) and
knypek (kny) zebrafish mutants all exhibit CE defects, suggesting that these genes lie in
the same pathway. The tri mutant encodes for a core PCP component, Vangl2. [27]. Both
slb and ppt are Wnt proteins, Wnt11 and Wnt5b respectively, which have been classified
as non-canonical Wnt ligands [15]. kny encodes a member of the glypican family of
heparin sulphate proteoglycans, which potentate non-canonical Wnt signalling [28].
Based on the classification of Wnt from ectopic expression studies in Xenopus, there are
three non-canonical Wnt genes: wnt4, wnt5, wnt11. More specifically, the zebrafish non-
canonical wnt homologues that have been identified are: wnt4a, wnt4b, wnt5b, wnt11 and
wnt11-related. wnt4b and wnt11-related are most likely not involved in establishing PCP
since they are first expressed at later stages of development, approximately at 18- and
14-somite stages respectively [29][30], whereas wnt4a, wnt5b and wnt11 are expressed
when the embryo is undergoing CE movements [31][32][33]. There is no known mutant
for wnt4a; however, morpholino oligonucleotides (MO) that block translation of wnt4a
mRNA were shown to exacerbated the CE defect in slb;ppt mutants as compared with the

17
double mutant alone [17]. In addition, the absence of wnt4a/5b/11 leads to the loss of
polarized Pk localization in zebrafish embryos. Taken together, non-canonical Wnt
proteins are able to regulate PCP, as seen in the wnt5 and wnt11 mutants with CE defects,
as well as loss of GFP-Pk localization, an indicator of disrupted PCP signalling.
1.5.2: Vertebrate specific regulators of planar cell polarity are associated with Wnt
signalling
In vertebrates, Wnt protein has been classified as either canonical or non-
canonical depending on which signalling pathway is activated; however, another model
suggests that receptor availability may also regulate which pathway is activated [34]. For
example, it has been shown that LRP5/6 is necessary for canonical Wnt signalling. With
non-canonical Wnt signalling, a few co-receptors have been implicated in vertebrate
PCP: Knypek (Kny), Protein Tyrosine Kinase 7 (PTK7), RYK and ROR2. In vertebrates,
other regulators in addition to the core PCP group that appear to be able to regulate PCP;
however, it is unclear whether these regulators aid in interpreting the global PCP cue.
Kny is thought to be a positive regulator of PCP, where it may act to bind and promote
transmission of the non-canonical Wnt signal [35]. Kny has been shown to physically
interact with Dickkopf-1 (Dkk1), a secreted protein that negatively modulates canonical
Wnt signalling by inhibiting the LRP5/6-Fz interaction. Possibly Kny may promote PCP
signalling by down-regulating canonical signalling, for example, by increasing Fz
receptor occupancy for non-canonical Wnt protein [36]. The requirement for Kny in PCP
signalling appears to be permissive, since kny mutants are rescued with global expression
with kny mRNA [35].

18
PTK7 encodes a single-pass transmembrane protein with seven extracellular
immunoglobulin (Ig) domains and an intracellular tyrosine kinase homology domain.
PTK7 has been shown to genetically interact with Vangl2 in mice and loss-of-function
studies in the mouse reveal defects in CE movements and neural tube closure [37].
Studies in Xenopus show physical interaction between PTK7 and Fz and ability of PTK7
to recruit Dsh to the membrane; however, PTK7 mouse mutants show no difference in
Dsh localization [38][39]. PTK7 appears to play a conserved role in regulating CE
movements in vertebrates, yet whether it has a direct effect on PCP signalling is unclear.
RYK is a single-pass transmembrane atypical receptor tyrosine kinase identified
initally to play a role in axon guidance and neurite outgrowth in flies and mammals [40].
RYK consists of an extracellular WIF (Wnt inhibitory factor) domain, an intracellular
atypical kinase domain and PDZ binding motif [40]. Studies in Xenopus suggest a role in
regulating PCP, since over-expression or knock-down results in CE defects [41]. RYK
has been characterized as a Wnt receptor since it is able to bind Wnt1/3/5a independent
of Fz, but it also appears to form complexes with Wnt1/Fz8 and Wnt11/Fz7 [41]. Tertiary
complex formation of Wnt, Fz and RYK appears to regulate PCP signalling by
controlling Fz-Dsh endocytosis and receptor trafficking [41].
ROR2 is a single-pass transmembrane orphan receptor tyrosine kinase consisting of
extracellular CRD and Kringle domains and an intracellular tyrosine kinase domain [42].
Although ror2-/- mutant mice do not exhibit CE or neural tube defects, ROR2 has been
implicated in PCP since the mutant phenotype resembles the wnt5a-/- mutant and in vitro
studies show a physical interaction between Wnt5a and ROR2 [42].

19
1.5.3: The search for a global cue that establishes vertebrate planar cell polarity
Based on Drosophila PCP studies, the global cue that establishes vertebrate PCP
most likely affects the core membrane PCP components, Fmi, Vangl2 and Fz. Fmi is
believed to play an adhesion role since it forms homodimers with adjacent cells with its
extracellular cadherin domains. Fmi may regulate PCP signalling by stabilizing PCP
complexes rather than responding to cues that establish PCP, since it appears that Fmi
regulates PCP only in complex with Fz. Over-expression of Fmi in zebrafish embryos
does not disrupt Dsh localization, but is able to block Fz induced membrane localization
of Dsh [21].
Given that there are no known ligands for Vangl2, its role in PCP may be to
propagate PCP signalling to adjacent cells. In zebrafish, Vangl2 acts cell non-
autonomously, as wildtype NPC loose polarity when transplanted into a vangl2 mutant
host [17]. Vertebrate PCP may also be propagated as in Drosophila by domineering non-
autonomy through interaction of Vangl2 with the Fz CRD [6]. However there is a
possibility that there is an undiscovered Vangl2 ligand or co-receptor that responds to a
global PCP cue to establish polarity. Investigation into new binding partners of Vangl2
may help discover other cues that may globally regulate PCP.
In vertebrates, Wnt proteins are able to regulate PCP where loss-of-function of
wnt4/5/11 zebrafish embryos display CE and neural tube defects. The vertebrate
extracellular regulators of PCP further support a role for non-canonical Wnt protein to
regulate PCP, since RYK and ROR2 are able to bind non-canonical Wnts and Kny and
PTK7 physically interacts with receptors of Wnt signalling. Fz is a core PCP protein and
may act to receive polarity information from a non-canonical Wnt protein. Non-canonical

20
Wnt(s) may act as the initial polarization cue, which is propagated through the plane by
cell-cell communication or possibly acting in a global Wnt gradient that establishes
polarity across the entire plane. In this thesis, I examined the regulation of vertebrate
PCP. More specifically, I investigated the role of non-canonical Wnt signalling in
regulating vertebrate PCP (Chapter 2). To identify novel regulators of PCP, I performed
a screen for new protein interactions with Vang (Chapter 3). Finally, I discuss the
preliminary results obtained and possible future directions (Chapter 4).
1.6: References
[1] D. Strutt, “Frizzled signalling and cell polarisation in Drosophila and vertebrates,” Development (Cambridge, England), vol. 130, Oct. 2003, pp. 4501-4513.
[2] J.R.K. Seifert and M. Mlodzik, “Frizzled/PCP signalling: a conserved mechanism regulating cell polarity and directed motility,” Nature Reviews. Genetics, vol. 8, Feb. 2007, pp. 126-138.
[3] R. Keller, D. Shook, and P. Skoglund, “The forces that shape embryos: physical aspects of convergent extension by cell intercalation,” Physical Biology, vol. 5, 2008, p. 15007.
[4] J. Wu and M. Mlodzik, “A quest for the mechanism regulating global planar cell polarity of tissues,” Trends in Cell Biology, vol. 19, Jul. 2009, pp. 295-305.
[5] M. Fanto and H. McNeill, “Planar polarity from flies to vertebrates,” Journal of Cell Science, vol. 117, Feb. 2004, pp. 527-533.
[6] J. Wu and M. Mlodzik, “The frizzled extracellular domain is a ligand for Van Gogh/Stbm during nonautonomous planar cell polarity signaling,” Developmental Cell, vol. 15, Sep. 2008, pp. 462-469.
[7] H. Strutt and D. Strutt, “Long-range coordination of planar polarity in Drosophila,” BioEssays: News and Reviews in Molecular, Cellular and Developmental Biology, vol. 27, Dec. 2005, pp. 1218-1227.
[8] P.A. Lawrence, J. Casal, and G. Struhl, “Towards a model of the organisation of planar polarity and pattern in the Drosophila abdomen,” Development (Cambridge, England), vol. 129, Jun. 2002, pp. 2749-2760.
[9] M. Fanto, L. Clayton, J. Meredith, K. Hardiman, B. Charroux, S. Kerridge, and H. McNeill, “The tumor-suppressor and cell adhesion molecule Fat controls planar polarity via physical interactions with Atrophin, a transcriptional co-repressor,” Development (Cambridge, England), vol. 130, Feb. 2003, pp. 763-774.
[10] D. Ma, C. Yang, H. McNeill, M.A. Simon, and J.D. Axelrod, “Fidelity in planar cell polarity signalling,” Nature, vol. 421, Jan. 2003, pp. 543-547.
[11] J. Casal, P.A. Lawrence, and G. Struhl, “Two separate molecular systems,

21
Dachsous/Fat and Starry night/Frizzled, act independently to confer planar cell polarity,” Development (Cambridge, England), vol. 133, Nov. 2006, pp. 4561-4572.
[12] C. Wu and R. Nusse, “Ligand receptor interactions in the Wnt signaling pathway in Drosophila,” The Journal of Biological Chemistry, vol. 277, Nov. 2002, pp. 41762-41769.
[13] Y. Wang, N. Guo, and J. Nathans, “The role of Frizzled3 and Frizzled6 in neural tube closure and in the planar polarity of inner-ear sensory hair cells,” The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, vol. 26, Feb. 2006, pp. 2147-2156.
[14] R. Keller, “Shaping the vertebrate body plan by polarized embryonic cell movements,” Science (New York, N.Y.), vol. 298, Dec. 2002, pp. 1950-1954.
[15] M. Tada, M.L. Concha, and C.P. Heisenberg, “Non-canonical Wnt signalling and regulation of gastrulation movements,” Seminars in Cell & Developmental Biology, vol. 13, Jun. 2002, pp. 251-260.
[16] C. Yin, M. Kiskowski, P. Pouille, E. Farge, and L. Solnica-Krezel, “Cooperation of polarized cell intercalations drives convergence and extension of presomitic mesoderm during zebrafish gastrulation,” The Journal of Cell Biology, vol. 180, Jan. 2008, pp. 221-232.
[17] B. Ciruna, A. Jenny, D. Lee, M. Mlodzik, and A.F. Schier, “Planar cell polarity signalling couples cell division and morphogenesis during neurulation,” Nature, vol. 439, Jan. 2006, pp. 220-224.
[18] N.S. Glickman, C.B. Kimmel, M.A. Jones, and R.J. Adams, “Shaping the zebrafish notochord,” Development (Cambridge, England), vol. 130, Mar. 2003, pp. 873-887.
[19] E. Hong and R. Brewster, “N-cadherin is required for the polarized cell behaviors that drive neurulation in the zebrafish,” Development (Cambridge, England), vol. 133, Oct. 2006, pp. 3895-3905.
[20] S. Witzel, V. Zimyanin, F. Carreira-Barbosa, M. Tada, and C. Heisenberg, “Wnt11 controls cell contact persistence by local accumulation of Frizzled 7 at the plasma membrane,” The Journal of Cell Biology, vol. 175, Dec. 2006, pp. 791-802.
[21] F. Carreira-Barbosa, M. Kajita, V. Morel, H. Wada, H. Okamoto, A. Martinez Arias, Y. Fujita, S.W. Wilson, and M. Tada, “Flamingo regulates epiboly and convergence/extension movements through cell cohesive and signalling functions during zebrafish gastrulation,” Development (Cambridge, England), vol. 136, Feb. 2009, pp. 383-392.
[22] S. El-Messaoudi and A. Renucci, “Expression pattern of the frizzled 7 gene during zebrafish embryonic development,” Mechanisms of Development, vol. 102, Apr. 2001, pp. 231-234.
[23] S. Angers and R.T. Moon, “Proximal events in Wnt signal transduction,” Nature Reviews. Molecular Cell Biology, vol. 10, Jul. 2009, pp. 468-477.
[24] A.F. Schier and W.S. Talbot, “Molecular genetics of axis formation in zebrafish,” Annual Review of Genetics, vol. 39, 2005, pp. 561-613.
[25] G. Schulte and V. Bryja, “The Frizzled family of unconventional G-protein-coupled receptors,” Trends in Pharmacological Sciences, vol. 28, Oct. 2007, pp. 518-525.
[26] H. Moeller, A. Jenny, H. Schaeffer, T. Schwarz-Romond, M. Mlodzik, M. Hammerschmidt, and W. Birchmeier, “Diversin regulates heart formation and

22
gastrulation movements in development,” Proceedings of the National Academy of Sciences of the United States of America, vol. 103, Oct. 2006, pp. 15900-15905.
[27] J.R. Jessen, J. Topczewski, S. Bingham, D.S. Sepich, F. Marlow, A. Chandrasekhar, and L. Solnica-Krezel, “Zebrafish trilobite identifies new roles for Strabismus in gastrulation and neuronal movements,” Nature Cell Biology, vol. 4, Aug. 2002, pp. 610-615.
[28] J. Topczewski, D.S. Sepich, D.C. Myers, C. Walker, A. Amores, Z. Lele, M. Hammerschmidt, J. Postlethwait, and L. Solnica-Krezel, “The zebrafish glypican knypek controls cell polarity during gastrulation movements of convergent extension,” Developmental Cell, vol. 1, Aug. 2001, pp. 251-264.
[29] A. Liu, A. Majumdar, H.E. Schauerte, P. Haffter, and I.A. Drummond, “Zebrafish wnt4b expression in the floor plate is altered in sonic hedgehog and gli-2 mutants,” Mechanisms of Development, vol. 91, Mar. 2000, pp. 409-413.
[30] T. Matsui, A. Raya, Y. Kawakami, C. Callol-Massot, J. Capdevila, C. Rodríguez-Esteban, and J.C. Izpisúa Belmonte, “Noncanonical Wnt signaling regulates midline convergence of organ primordia during zebrafish development,” Genes & Development, vol. 19, Jan. 2005, pp. 164-175.
[31] C.P. Heisenberg, M. Tada, G.J. Rauch, L. Saúde, M.L. Concha, R. Geisler, D.L. Stemple, J.C. Smith, and S.W. Wilson, “Silberblick/Wnt11 mediates convergent extension movements during zebrafish gastrulation,” Nature, vol. 405, May. 2000, pp. 76-81.
[32] B. Kilian, H. Mansukoski, F.C. Barbosa, F. Ulrich, M. Tada, and C.P. Heisenberg, “The role of Ppt/Wnt5 in regulating cell shape and movement during zebrafish gastrulation,” Mechanisms of Development, vol. 120, Apr. 2003, pp. 467-476.
[33] A.R. Ungar, G.M. Kelly, and R.T. Moon, “Wnt4 affects morphogenesis when misexpressed in the zebrafish embryo,” Mechanisms of Development, vol. 52, Aug. 1995, pp. 153-164.
[34] A.J. Mikels and R. Nusse, “Purified Wnt5a protein activates or inhibits beta-catenin-TCF signaling depending on receptor context,” PLoS Biology, vol. 4, Apr. 2006, p. e115.
[35] J. Topczewski, D.S. Sepich, D.C. Myers, C. Walker, A. Amores, Z. Lele, M. Hammerschmidt, J. Postlethwait, and L. Solnica-Krezel, “The zebrafish glypican knypek controls cell polarity during gastrulation movements of convergent extension,” Developmental Cell, vol. 1, Aug. 2001, pp. 251-264.
[36] L. Caneparo, Y. Huang, N. Staudt, M. Tada, R. Ahrendt, O. Kazanskaya, C. Niehrs, and C. Houart, “Dickkopf-1 regulates gastrulation movements by coordinated modulation of Wnt/beta catenin and Wnt/PCP activities, through interaction with the Dally-like homolog Knypek,” Genes & Development, vol. 21, Feb. 2007, pp. 465-480.
[37] X. Lu, A.G.M. Borchers, C. Jolicoeur, H. Rayburn, J.C. Baker, and M. Tessier-Lavigne, “PTK7/CCK-4 is a novel regulator of planar cell polarity in vertebrates,” Nature, vol. 430, Jul. 2004, pp. 93-98.
[38] I. Shnitsar and A. Borchers, “PTK7 recruits dsh to regulate neural crest migration,” Development (Cambridge, England), vol. 135, Dec. 2008, pp. 4015-4024.
[39] W.W. Yen, M. Williams, A. Periasamy, M. Conaway, C. Burdsal, R. Keller, X. Lu, and A. Sutherland, “PTK7 is essential for polarized cell motility and convergent

23
extension during mouse gastrulation,” Development (Cambridge, England), vol. 136, Jun. 2009, pp. 2039-2048.
[40] W. Lu, V. Yamamoto, B. Ortega, and D. Baltimore, “Mammalian Ryk is a Wnt coreceptor required for stimulation of neurite outgrowth,” Cell, vol. 119, Oct. 2004, pp. 97-108.
[41] G. Kim, J. Her, and J. Han, “Ryk cooperates with Frizzled 7 to promote Wnt11-mediated endocytosis and is essential for Xenopus laevis convergent extension movements,” The Journal of Cell Biology, vol. 182, Sep. 2008, pp. 1073-1082.
[42] I. Oishi, H. Suzuki, N. Onishi, R. Takada, S. Kani, B. Ohkawara, I. Koshida, K. Suzuki, G. Yamada, G.C. Schwabe, S. Mundlos, H. Shibuya, S. Takada, and Y. Minami, “The receptor tyrosine kinase Ror2 is involved in non-canonical Wnt5a/JNK signalling pathway,” Genes to Cells: Devoted to Molecular & Cellular Mechanisms, vol. 8, Jul. 2003, pp. 645-654.

24
CHAPTER 2: Investigating the Role of Wnt Signalling in Vertebrate Planar Cell Polarity 2.0: Introduction
There are two models for planar cell polarity (PCP) establishment, the cell-cell
relay model and gradient model (Figure 2). The cell-cell relay model results in sequential
polarization of a field of cells, where Factor X initiates polarization and adjacent cells are
polarized either by production of a secreted cue acting in small gradients or by
domineering non-autonomy. The gradient model suggests that PCP is established by a
long-ranged gradient of Factor X, where cells sense an increasing/decreasing gradient and
establish polarity accordingly. The identity of Factor X is unknown, however there is a
strong precedent for non-canonical Wnt protein to establish PCP in vertebrates. One of
the core PCP components is Frizzled (Fz), a transmembrane protein that contains an
extracellular cysteine-rich domain (CRD), which binds Wnt protein. Zebrafish non-
canonical Wnt mutants (wnt5b/11) display convergent extension (CE) defects that
phenocopy other core PCP mutants. Finally, other vertebrate regulators of PCP, knypek,
protein tyrosine kinase 7, RYK and ROR2 are all associated with Wnt signalling.
In both models of PCP establishment, Factor X needs to act instructively to
establish PCP across the plane. To generate polarity across a field of cells, the initial cue
needs to create directionality. An instructive cue can elicit different responses within a
plane, whereas the response a cell has to the cue will vary with its relative position
(Figure 5). In the gradient model, an instructive cue would polarize the entire plane with
each cell responding to a gradient of Factor X. In the cell relay model, the initial factor

25
Figure 5: Instructive Versus Permissive Functions of Wnt The role of Wnt signalling in regulating PCP is unclear. Wnt signals could possibly act in two ways to regulate PCP either as an instructive or permissive signal. Wnt as an instructive signal: Wnt signals are required for PCP and establish the direction of cell polarization (arrows). The polarity of cells surrounding a Wnt source will reflect their relative position to the localized polarizing cue. Example: Wnt PCP. Wnt as a permissive cue: Wnt signals are required for PCP, however they do not dictate the direction of cell polarization. The polarity of cells is established irrespective of the Wnt source. Potentially another factor instructs PCP. Example: Wnt + FactorX PCP.
Wnt
Wnt
X
Permissive
Instructive

26
dictates the directionality, where positional information the first cell receives from the
cue will be propagated to the next cell and eventually across the entire plane. Conversely,
a signal that is required by the cell, where the cell’s response is the same irrespective of
its position, to the cue is considered a permissive cue (Figure 5). To gauge whether non-
canonical Wnt proteins act as Factor X in establishing PCP, non-canonical Wnt protein
needs to be assayed for either a permissive or instructive role in regulating PCP. There is
some evidence from zebrafish studies suggesting that Wnt11 acts permissively and wnt5b
acts instructively to regulate PCP. wnt11 mutants can be rescued with global over-
expression of wnt11 mRNA, demonstrating that graded/localized Wnt11 expression is not
required to establish PCP [1]. However, rescue of the wnt5b mutant cannot be achieved
with ubiquitous expression of wnt5b mRNA, possibly because a specific Wnt5b
expression pattern is required in the embryo to establish PCP [2]. There is evidence from
other model organisms showing the Wnt protein can act as instructive cues. In the
developing chick embryo, myocytes normally elongate parallel to the neural tube. When
a cell expressing Wnt11 is transplanted in between two somites, elongating myocytes are
found swirling around the local Wnt11 source. When Wnt11 is globally over-expressed,
myocyte elongation is disorganized. Taken together, it suggests that Wnt11 acts
instructively to regulate myotcye elongation in the chick embryo [3]. During C. elegans
development at the four-cell stage, the P2 cell (signalling cell) determines the division
plane in the EMS (responding cell). Positional MOM-2 (C. elegans Wnt) signal secreted
from the P2 cell acts instructively on the EMS cell directing spindle orientation [4].
Although Wnt Protein can act instructively, PCP has not been implicated in directing
myocyte elongation or spindle orientation in the EMS cell. To assay if Wnt acts

27
instructively to establish PCP, similar assays with a local Wnt source need to be
performed, however in a system/process where PCP signalling is required.
Zebrafish neurulation is a model system that can be used to assay the affects of
local Wnt protein expression on PCP. It is amenable to embryonic manipulation, in which
there are various means to induce localized ectopic Wnt expression. To gauge if Wnt
protein is acting instructively or permissively, differences in Prickle (Pk) localization can
be used. Asymmetric localization of the PCP components is a hallmark of PCP
signalling. In the developing zebrafish neural tube, it has been shown that a green
fluorescent protein (GFP)-Pk fusion localizes to the anterior side of the membrane in
discrete puncta in neural progenitor cells (NPCs) (Figure 6A). Disruption of PCP such as
in vangl2 mutant and wnt5b/11 double mutant embryos injected with wnt4 morpholino
oligonucleotide (MO) show loss of puncta on the anterior membrane (Figure 6B) [5]. In
response to a local Wnt source, the polarity of NPCs can be examined. If Wnt acts
permissively to establish PCP, a local Wnt source should not change the directionality of
polarization. Accordingly, GFP-Pk localization should not be altered around the local
Wnt clone. If Wnt protein acts instructively to establish PCP, GFP-Pk localization should
change with respect to the local Wnt source. Potentially, normal polarity (GFP-Pk
anterior localization) would be observed in one direction, while polarity may be reversed
in the opposite direction.
There are three non-canonical Wnts, wnt4a, wnt5b and wnt11, that are expressed
during zebrafish gastrulation/neurulation. wnt11 expression in WT zebrafish embryos is
first detected in the germ ring at shield stage and is later detected in the anterior paraxial
mesoderm and neuroectoderm [1]. By late gastrulation, the neuroectoderm domain lies

28
Figure 6: GFP-PK as a marker of PCP (A) In WT zebrafish neural progenitor cells (NPCs), GFP-Prickle (Pk) puncta localized to the anterior membrane. (B) When PCP signalling is disrupted like in materal-zygotic vangl2 mutants, GFP-Pk puncta at the anterior membrane is lost. GFP-Pk remains cytoplasmic. Ciruna et al. Nature (2006) 439, 220-224.

29
posterior to the presumptive forebrain [1]. wnt5b is maternally provided and zygotic
expression is first detected in the germ ring at shield stage and later in the posterior
paraxial and axial mesoderm, adjacent to the wnt11 expression, becoming restricted to the
posterior mesendoderm and tailbud [2]. wnt4 is first detected at the 3-somite stage in the
forebrain and spreads posteriorly through the hindbrain, along the lateral dorsal neural
keel stopping short of the tailbud at approximately the 8-10 somite stage [7]. At later
stages, wnt4a is found in the anterior lateral plate mesoderm and mainly in the floor plate
of the neural tube [6]. The non-canonical Wnts have over-lapping expression patterns and
appear to act redundantly to regulate PCP. wnt11 mutants can be rescued with injection
of wnt5b mRNA. Loss-of-function of all three non-canonical Wnts is required to achieve
the most severe phenotype [17]. As a result, creating single mutant loss-of-function Wnt
clones and assaying polarity on neighbouring cells may not be informative. Using gain-
of-function clones, each non-canonical Wnt protein can be assayed for its instructive or
permissive affect on PCP.
To assay the direct affect of Wnt on PCP, a clone expressing Wnt protein was
introduced into the neural keel. This requires spatial and temporal control of the clone.
One method is to transplant beads coated with growth factors, which can be transplanted
into any location at any point in development. The major limitation to this method is the
isolation and purification of the growth factor of interest. Although Wnt proteins are
secreted, active Wnt molecules are difficult to solubilize and consequently difficult to
purify [8]. Their inability to solubilize is a result of lipid-modifications resulting in a
more hydrophobic protein than predicted by the amino acid sequence [8]. To generate
Wnt clones, an in vivo approach was taken where cells that ectopically express high level

30
of Wnt protein were transplanted into the neural keel labelled with GFP-Pk, the marker of
cell polarity. One method to generate chimeras in zebrafish is by homochronic cell
transplantation, where donor cells from a mid-blastua stage embryo (donor) are
transplanted into a staged-matched embryo (host). The donor embryo is injected with wnt
mRNA and cells are transplanted into the presumptive neural tube of a GFP-Pk labelled
host embryo. Although homochronic cell transplantation would generate Wnt clones in
the neural tube, there are two concerns that may complicate the analysis. First, the Wnt
clone will be active throughout development and it will be unclear whether observed
changes in polarity are a direct result of Wnt on NPC or a secondary result from altered
PCP during gastrulation. Second, many Wnt clones will be generated, possibly making
the analysis difficult when examining NPC surrounded by multiple Wnt clones and
attempting to determine the direction of the Wnt gradient. To clearly demonstrate an
instructive or permissive role for Wnt on PCP, one Wnt clone that is active only during
neurulation needs to be generated. Accordingly, any observed differences in the polarity
of NPC can be attributed to the Wnt clone. Described herein is the first demonstration
that transplantation of mid-blastula stage cells directly into the neural keel is possible and
can be used to assay the role of Wnt in regulating PCP. My preliminary results suggest
that Wnt4a is able to alter NPC behaviour in the developing neural tube.
2.1: Results
2.1.1: Heterochronic cell transplantation is possible
Transplantation is usually performed with mid-blastula staged embryos since the
cells are easily manipulated and have less adhesion between them. Cells can be

31
withdrawn or deposited into the mid-blastula staged embryo without altering
development. For temporal and spatial control of the Wnt clone, heterochronic cell
transplants (HCT) were performed, where cells from a donor mid-blastula staged embryo
injected with wnt mRNA were transplanted into the neural keel of host embryo labelled
with GFP-Pk. To test whether HCT were possible, donor cells labelled with membrane-
GFP (mGFP) were transplanted into a membrane red fluorescent protein (memRFP)
labelled host. Using confocal microscopy, host embryos were imaged two hours after
transplantation for the presence of transplanted cells (mGFP labelled) in the neural tube
(Figure 7A). The donor cells are much larger than the NPC since the donor cells are taken
from an earlier staged embryo and transplanted into a later staged host. There is no non-
specific or absent fluorescence indicating donor cells are alive and transplantation does
not cause cellular damage. The donor cells appear to be well integrated into the host
suggesting that HCT are possible.
The affect of HCT on the endogenous polarity was investigated to ensure that
HCT did not disrupt PCP signalling. GFP-Pk was used as a marker of polarity, as GFP-
Pk forms membrane associated puncta localized to the anterior membrane in wildtype
(WT) NPCs. If PCP is disrupted, as with loss-of-function of non-canonical Wnts, GFP-Pk
puncta are lost from the membrane. To test whether HCT disrupts PCP in the host, WT
mid-blastula staged cells labelled with mGFP were transplanted into the neural keel of a
memRFP and GFP-Pk scatter labelled host. GFP-Pk was not globally expressed since it is
difficult to discern anterior localization of puncta in one cell from posterior localization
of puncta in an adjacent cell when all cells are labelled. Instead, GFP-Pk was scatter-
labelled, where one blastomere of an 8-cell staged embryo was injected resulting in

32
Figure 7: Heterochronic cell transplants are possible and do not disrupt PCP A. Cells from sphere stage embryo labelled with mGFP transplanted into host 4-6 somite staged embryo labelled with memRFP. Confocal section, dorsal view. B. Cells from sphere stage embryo injected with mGFP transplanted into host 4-6 somite stage embryo labelled with memRFP and scatter-labeled with GFP-Pk. Puncta are found on the anterior side of the membrane. Confocal sections, dorsal view.
Ant

33
patches of GFP-Pk labelled cells. Examining the host embryos after HCT showed
integrated donor cells (mGFP) in the neural keel (memRFP). The localization of GFP-Pk
puncta was along the anterior membrane in cells near the donor cells, suggesting that
HCT does not disrupt PCP in the host embryo (Figure 7B).
2.1.2: Characterization of Wnt clones
HCT is a method that allows for mid-blastula cells to be transplanted into the host
neural keel; however, it is unclear if the transplanted donor cells can act as a source of
Wnt signal. For the Wnt clone to act as a local source of Wnt protein, secretion of
functional Wnt protein to neighbouring NPCs is required. To characterize the donor cells,
several tests were performed to show that cells transplanted from embryos injected with
wnt mRNA could express, secrete and generate functional Wnt protein. First, to show that
wnt mRNA injected into embryos is translated into protein, mRNA encoding wnt11 fused
to yellow fluorescent protein (YFP) mRNA was injected into 1-cell stage embryos and
observed at mid-blastula stage for YFP fluorescence. Injected embryos at mid-blastula
showed YFP fluorescence indicating the injected wnt mRNA is translated into protein
(Figure 8A). Next, to test if Wnt protein is secreted, cells from embryos injected with
wnt11YFP mRNA at mid-blastula stage were transplanted into the animal cap of a
memRFP labelled stage-matched host. Using confocal microscopy, host embryos were
imaged and donor cells expressing Wnt11YFP were detected. YFP fluorescence was
observed as cytoplasmic puncta inside the donor cells as well as discrete puncta among
the host cells suggesting that Wnt protein is secreted from transplanted mid-blastula stage
cells (Figure 8B).
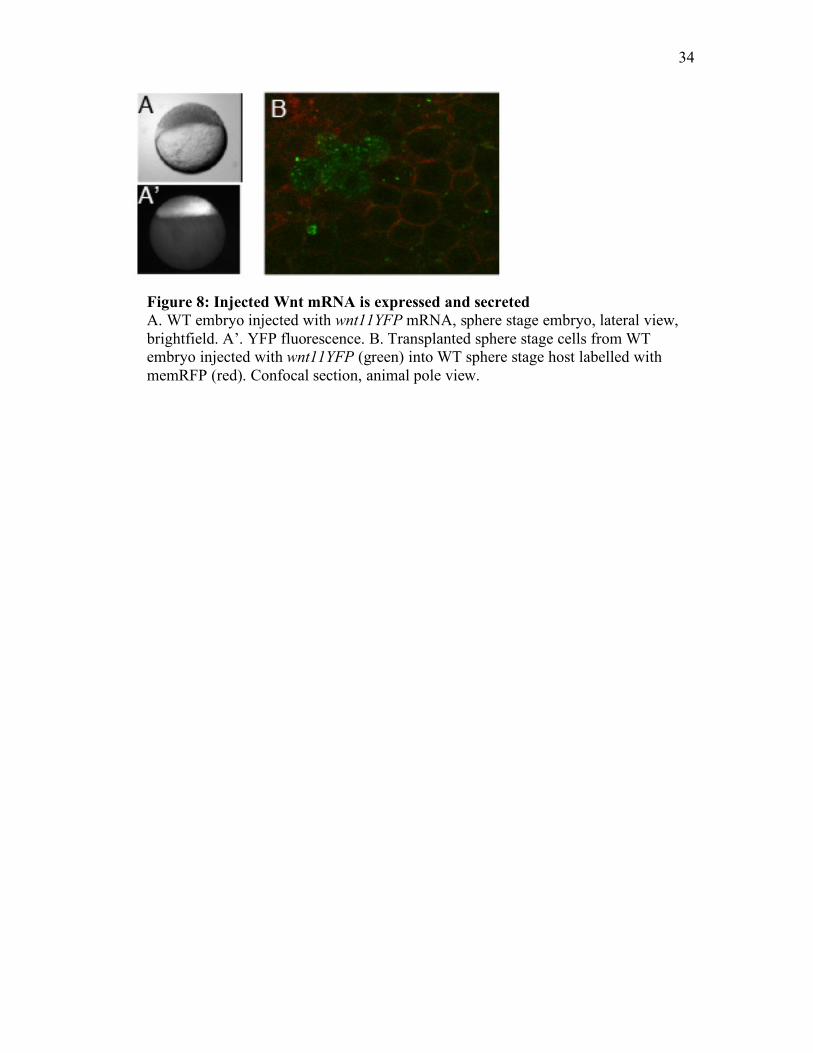
34
Figure 8: Injected Wnt mRNA is expressed and secreted A. WT embryo injected with wnt11YFP mRNA, sphere stage embryo, lateral view, brightfield. A’. YFP fluorescence. B. Transplanted sphere stage cells from WT embryo injected with wnt11YFP (green) into WT sphere stage host labelled with memRFP (red). Confocal section, animal pole view.

35
To test whether injected wnt mRNA results in functional Wnt protein, non-
canonical wnt mRNA was globally over-expressed in zebrafish embryos and scored for
an extension defect at a mid-somite stage (Figure 9). Wnt misexpression has been shown
to induce morphogenesis defects such as decreased extension of axial tissues and anterior
migration of cells [1][2][7]. Comparing phenotypes to uninjected controls, injection of
200 pg of wnt11YFP and wnt5b consistently generated an over-expression phenotype, as
seen with decreased anterior-posterior extension of the embryo. Surprisingly, injection of
20 pg of wnt4a was able to generate a similar phenotype. Although the non-canonical
Wnts may be functionally redundant as demonstrated with the rescue of the wnt11 mutant
with wnt5b expression, they appear to have distinct activities where rescue of wnt11
mutants is achieved with ten times less wnt11 than wnt5b mRNA. Wnt4a was the first
Wnt chosen to assay using HCT since a low amount of mRNA was required to generate a
phenotype, and further increases in mRNA concentration would be possible without
inducing non-specific mRNA toxicity indicated by necrosis or the embryo.
To further characterize the effect of Wnt4a ectopic expression on NPCs, wnt4a
mRNA was injected into WT 1-cell stage embryos and the neural tube was examined for
neural tube defects and the presence of GFP-Pk puncta at mid-somite stage. As seen with
vangl2 mutants, disruption of PCP leads to defects in neural tube formation, where an
ectopic accumulation of apical daughter cells occurs at the midline. Embryos were
injected with 20 pg of wnt4a mRNA and labelled with memRFP. Using confocal
microscopy, injected embryos displayed an accumulation of cells in the neural tube
suggesting Wnt4a is capable of disrupting PCP (Figure 10A). To gauge if Wnt protein

36
Figure 9: Ectopic non-canonical Wnt expression induces morphogenesis defects A-D: Lateral views, anterior is to the right, posterior is to the left. A. Non-injected WT embryo during segmentation period (12-16 hpf). B. WT embryos injected with 20 pg of wnt4 mRNA. C. WT embryos injected with 200 pg of wnt11YFP mRNA. D. WT embryo injected with 200 pg of wnt5 mRNA. E. WT embryos injected with 200 pg of wnt11r mRNA.
37
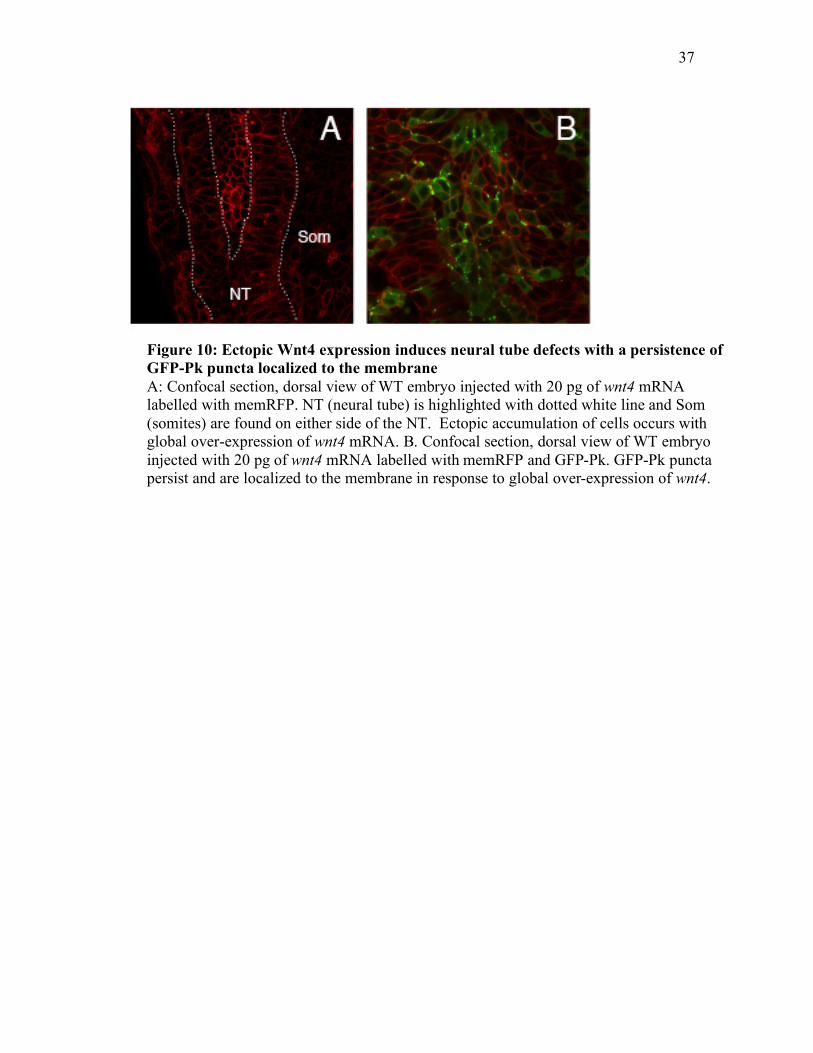
Figure 10: Ectopic Wnt4 expression induces neural tube defects with a persistence of GFP-Pk puncta localized to the membrane A: Confocal section, dorsal view of WT embryo injected with 20 pg of wnt4 mRNA labelled with memRFP. NT (neural tube) is highlighted with dotted white line and Som (somites) are found on either side of the NT. Ectopic accumulation of cells occurs with global over-expression of wnt4 mRNA. B. Confocal section, dorsal view of WT embryo injected with 20 pg of wnt4 mRNA labelled with memRFP and GFP-Pk. GFP-Pk puncta persist and are localized to the membrane in response to global over-expression of wnt4.

38
acts instructively (changes in GFP-Pk puncta localization) or permissively (no changes
in GFP-Pk puncta localization), GFP-Pk puncta need to be present in response to a Wnt
clone. To test if GFP-Pk puncta persist in NPC in response to ectopic Wnt4a expression,
embryos were injected with wnt4a mRNA and labelled globally with memRFP and GFP-
Pk. Using confocal microscopy, NPCs in injected embryos had GFP-Pk puncta present at
the membrane, demonstrating in response to ectopic Wnt4a, GFP-Pk puncta persist and
can be used to as a readout for the Wnt instructive/permissive assay (Figure 10B). Since
the GFP-Pk are present, differences in localization or quantity of puncta can be examined
in response to a localized Wnt4a source.
2.1.3: Localized ectopic Wnt4a expression is able to alter neural progenitor cell
behaviour
After ensuring HCT were possible and did not disrupt endogenous PCP as well as
characterizing the donor cells as a Wnt clone capable of secreting functional Wnt, HCT
were performed generating a Wnt4a clone inside the neural keel. If Wnt4a plays an
instructive role, it would be expected that the polarity of NPCs would be established with
respect to the location of the Wnt clone. Based on wnt4a in situ hybridization data, there
is an endogenous decreasing gradient along the anterior-posterior axis. Possibly, normal
polarity (anterior localized GFP-Pk puncta) would be established posterior to the Wnt
clone since NPCs would be exposed to the same decreasing gradient. In addition, polarity
would be reversed anterior to the Wnt clone, as the NPCs would be exposed to an
increasing Wnt4a gradient, opposite to the endogenous gradient. Thus, if Wnt4a acts

39
permissively, polarity should be affected equally anterior and posterior to the Wnt clone,
and GFP-Pk localization should be the same in all NPCs surrounding the Wnt clone.
Unfortunately, the majority of attempts to perform the assay were unsuccessful,
generating uninformative data. The main issue was that variability within each step of the
procedure decreased the overall success of the experiment. The position of the Wnt clone
is not fixed, when imaging the transplant location (near the 6th somite) donor cells were
often not detected suggesting the donor cells shifted position or died. GFP-Pk labeling
was clustered and uneven, making it difficult to determine on which membrane the
puncta were localized. In addition, the entire anterior-posterior axis was not labelled,
making it difficult to compare anterior and posterior regions relative to the Wnt clone.
Finally, due to the length of the procedure, there are a finite number of HCTs that can be
performed and examined by confocal microscopy, making it difficult to achieve a high
number of replicates. Taken together, it is unclear whether Wnt4a acts instructively or
permissively to establish PCP.
Despite the majority of data being uninformative, a small proportion (n=2) of
HCT resulted in a surprising phenotype. NPCs appear to respond differently to a local
Wnt4a source with respect to its relative position. Although the GFP-Pk labeling could
not be used to gauge if PCP was affected, GFP-Pk labeling was used as a cell tracer.
Scatter-labeling by injection of one blastomere at the 8-cell stage often results in labeling
cells on one half of the embryo along the anterior-posterior axis. During neurulation,
NPC from one side of the neural tube will divide generating a daughter cell that will cross
the midline and intercalate into the contralateral side. As a result, there is equal
contribution

40
Figure 11: Neural progenitor cells respond differently to a Wnt4a clone with respect to its position Heterochronic cell transplants generating a local Wnt4a clone. A-C confocal sections, dorsal views, anterior top, posterior bottom. Wnt source (*), donor cells labelled with mGFP and injected with 20 pg of Wnt4. A. Majority of neural progenitor cells anterior to the Wnt source do not cross the midline. B. Neural progenitor cells adjacent to the Wnt source are disordered. No clear midline is found. C. Posterior to the Wnt source, neural progenitor cells can be found on both sides of the neural tube.
Anterior
Posterior
*
*
*

41
of cells from both sides of the neural tube. In the HCT assay, NPCs posterior to the
Wnt4a clone appear to behave normally, with a distribution of GFP-Pk labelled cells on
both sides of the neural tube (Figure 11C). NPCs adjacent to the Wnt4a clone are
disordered, and there is no clear midline (Figure 11B). Anterior to the Wnt4a clone, GFP-
Pk labelled cells appear to have not crossed the midline to the contralateral side, but have
not accumulated at the midline (Figure 11A). The HCT data suggest that Wnt4a affects
NPC behaviour along the anterior-posterior axis.
To support the results obtained from HCT, a transposon-mediated transient
transgenic approach was used. The transient transgenic approach permits temporal and
some spatial control in generating Wnt clones. The temporal control results from using a
heat-shock promoter to drive gene expression at a desired time in development. Spatial
control of Wnt clones results from chimeric integration of the transgene, such that only a
few cells will express the transgene (Figure 12B). The transgene used to generate Wnt4a
clones consisted of a heat-shock promoter driving expression of wnt4a fused to an
internal ribosomal entry site (IRES) and mGFP reporter (Figure 12A). Successful
transgene activation was indicated by mGFP expression. To track cell movement, cells
were scatter labelled with memRFP by injecting one blastomere at the 8-cell stage. Such
cells will often mark only half the embryo along the anterior-posterior axis. As NPCs
begin to divide, apical daughter cells will intercalate into the contralateral membrane,
where cells labelled with memRFP should be found on sides of the developing neural
tube (Figure 12C). Embryos were heat-shocked at 6-somite stage and imaged using
confocal microscopy. The cells surrounding the Wnt4a clone were examined for

42
Figure 12: Transgenic approach to generate localized Wnt source in the neural tube Transgenesis is mediated by injection of tol2 transposase mRNA and transgene vector (A) is chimeric. Confocal sections, dorsal view. B. Heatshock induced transgenic with low level of transgene integration C. Control embryo scatter labelled with memRFP demonstrating that as cells divide, apical daughter cells are deposited on the contralateral side of the developing neural tube. D. Heatshock transgenic embryo scatter labelled with memRFP. Cells expressing transgene are labelled with mGFP. Cells anterior to Wnt4 source are found on one side of the neural tube, while cells posterior to the Wnt4 source are found on both sides of the embryo.
HSP70 Wnt IRES mGFP
C
A

43
differences in cell movement across the midline anterior and posterior to the local Wnt4a
source. Similar to the HCT experiment, NPCs posterior to the Wnt4a clone were found
on both sides of the neural tube. NPCs anterior to the Wnt4a clone were only found one
side of the neural tube (Figure 12D). Taken together, Wnt4a appears to affect the
behaviour of NPC across the midline anterior to the Wnt4a clone.
2.2: Discussion
2.2.1: Anterior-posterior Wnt4a gradient provides positional information to neural
progenitor cells
There is a striking difference to the response of NPC anterior and posterior to a
local Wnt4a source. Comparing the anterior and posterior regions, the main difference is
the direction of the Wnt4a gradient that is formed. Posterior to the Wnt4a clone, a
decreasing gradient along the anterior-posterior axis is generated, which is the same
direction as the endogenous wnt4a gradient. The NPCs anterior to the Wnt4a clone are
exposed to an increasing gradient along the anterior-posterior axis, opposite to the
endogenous wnt4a gradient. The Wnt4a gradient along the anterior-posterior possibly
provides some kind of information, where a ‘reverse’ gradient may alter NPC behaviour
(Figure 13). Considering the expression pattern of wnt5b and wnt11 at early somite
stages, wnt5b expression is found in the tailbud and the posterior mesendoderm, while
wnt11 expression is found in the forebrain and anterior mesoderm. Possibly, Wnt5b
and/or Wnt11 act to establish the anterior-posterior identity by PCP signalling and Wnt4a
either acts redundantly or has a distinct role in regulating neural tube morphogenesis.
This is based on the assumption that Wnt4a is being secreted and Wnt4a is able to

44
Figure 13: Wnt4a gradients may provide positional information to neural progenitor cells Generation of a local Wnt4a source and resulting Wnt4a gradients may provide positional information to neural progenitor cells. Cells posterior to the Wnt4a source are exposed to a ‘normal’ anterior-posterior decreasing gradient and labelled cells are found on both sides of the neural tube. Cells anterior to the Wnt4a source are exposed to a ‘reverse’ anterior-posterior increasing gradient and labelled cells are found on one side of the neural tube. Cells adjacent to the Wnt4a are exposed to no gradient and labelled cells accumulate ectopically at the midline.

45
mediate a respond in NPCs in both directions. Potentially, Wnt4a is not secreted in the
anterior direction, which can be confirmed by generating a local Wnt source expressing
fluorescently tagged Wnt protein. If fluorescent Wnt puncta are found anterior to the
clone, it suggests that Wnt protein is secreted in the anterior direction. To test whether
NPC anterior to the Wnt clone are able to transduce Wnt4a signal, a downstream effector
such as GFP-Pk localization can be used. If anterior NPCs are able to respond to Wnt4a,
potentially a change in GFP-Pk localization will be observed.
If Wnt4a is secreted in all directions and all NPCs can respond to Wnt4a, there are
three possibilities for how a reversed Wnt4a gradient may affect NPCs. First, Wnt4a
could regulate the direction of intercalation. PCP has been shown to couple cell division
with mediolateral intercalation; however, the mechanism by which the direction of
intercalation is decided is unknown. In Drosophila studies, Fz co-localizes with the
position of the polarized structure. Conversely, in vertebrates, Fz has not been shown to
co-localize with stable mediolateral cellular protrusions. The localization of GFP-Pk
suggests that the PCP components in vertebrates are localized along the anterior-posterior
axis. It is unclear how Wnt4a would regulate the position of protrusion formation.
Possibly in response to the endogenous decreasing anterior-posterior gradient, apical
daughter cells intercalate into the contralateral side. When apical daughter cells are
exposed to a reverse gradient, an increasing anterior-posterior gradient, the apical
daughter cell intercalates into the ipsilateral side. As seen with the HCT data (Figure
12B) and Wnt4a global over-expression (Figure 10A), possibly when no Wnt4a gradient
is present, the ability to direct protrusions to either side is disrupted, resulting in NPCs
accumulating ectopically The loss of the Wnt4a gradient such as with wnt4a morphants,

46
no phenotype observed, suggesting that the Wnt4a gradient does not inhibit formation of
protrusions. Possibly, loss of a Wnt4a gradient randomizes the direction of protrusions,
where in wnt4a morphants, daughter cells are deposited randomly on either side, but
equally from both sides, which would possibly result in no morphological phenotype.
The second and third possibilities are potential affects of Wnt4a on cell division.
Possibly, Wnt4a may affect the direction of cell division, where the absence of NPCs
crossing the midline is a result of NPC divisions along the anterior-posterior axis. The
third possibility is that Wnt4a regulates the decision for NPCs to divide. NPCs found
anterior to the Wnt4a source may not divide, resulting no daughter NPCs crossing the
midline. It is unclear how a reversed gradient would affect cell division; however both
scenarios are possible since either could result in NPCs not crossing the midline.
To determine the specific role of Wnt4a, NPC division needs to be observed in
response to a local Wnt4a source. Using confocal microscopy and time-lapse imaging,
NPCs can be tracked to determine if cells are dividing and in which direction. The
evidence from the HCT experiments, supported by the transgenic data, suggests that
Wnt4a provides positional data to NPCs. It is unclear if Wnt4a acts through PCP
signalling. To determine if the differential NPC behaviour is a result of changes in PCP
signalling, differences in GFP-Pk localization can compared with NPC movement in
response to a local Wnt4a source.
2.3: Methods
2.3.1: Materials
An in vitro RNA synthesis kit to generate capped mRNA for microinjection and DEPC-

47
treated water used to dilute mRNA was obtained from Ambion. Restriction enzymes,
bovine serum albinum and alkaline phosphatase were obtained from New England
Biosciences. High-fidelity Taq Phusion PCR kit was obtained from Finzymes and
primers used in PCR were obtained from Sigma-Aldrich. Cloning was performed using
DH5α, TOP10 cells and all gateway cloning reagents (LR clonase, BP clonase) were
obtained from Invitrogen. DNA ligation kit was obtained from Roche Biosciences.
Plasmid DNA isolation, gel extraction and PCR purification kits were obtained from
Qiagen. Glass capillary tubes with filaments and without filaments were obtained from
World precision instruments and were pulled on a Sutter Instruments Co. glass puller to
generate needles for microinjections and cell transplantation. Phenol red, low melt agarose,
pronase and other biological buffers/reagents were obtained from Sigma-Aldrich.
Embryos were mounted in glass bottom Petri dishes obtained from MatTek Corporation
and imaged using LSM710 confocal microscope from Zeiss equipped with ZEN software.
Figures were processed using ZEN and Adobe Creative Suite 4.
2.3.2: Zebrafish embryo microinjection
Zebrafish embryos were obtained from natural mating of wildtype (WT) TL and AB
hybrid backgrounds. Plasmids containing membrane-localized red fluorescent protein
(memRFP), membrane-localized green fluorescent protein (mGFP), EGFP-Prickle (GFP-
Pk), Wnt4a, Wnt5b, Wnt11, Tol2, were linearized and sense-strand-capped mRNA was
synthesized with the mMESSAGE mMACHINE system. Zebrafish embryos were
dechorionated by treatment of pronase and injected at the one-cell stage. Scatter labeling

48
was obtained by injecting two blastomeres at the eight-cell stage.
2.3.3: Cell transplants
Cell transplants were performed with mid-blastula stage embryos. For Wnt secretion
assay, embryos were dechorionated with Pronase treatment. Donor and host embryos
were injected at the 1-cell stage with capped wnt11-yfp and memRFP mRNA
respectively. Approximately 20 cells were transplanted into the animal cap of the host
embryo. Heterochronic cell transplants were performed with mid-blastua stage and 6-
somite staged embryos. With initial transplants, donor cells were labeled with rhodhamin-
dextran. Host embryos were injected with memRFP mRNA and scattered labeled with
GFP-Pk mRNA to the 6-stage. Donor embryos were injected with mGFP and Wnt
mRNA to the mid-blastua stage. 10-20 cells from the donor embryo were transplanted
into a single location at the 6-somite in the host embryo.
2.3.4: Microscopy
Live embryos were mounted in 0.8% low melt agarose before imaging. Fluorscent images
of embryos injected with memRFP, mGFP, GFP-Pk or transgenics were obtained with a
Zeiss 510/710 confocal microscope. Images were captured along the dorsal-ventral axis
through the neuroepithelium to the notocord at the 6-12 somite stage.
2.3.5: Transgenesis
To generate transgenic zebrafish, tol2 mediated transgenesis was used. Using the Gateway

49
cloning system from Invitrogen following the manufacture’s instructions, middle entry
clones containing Wnt open reading frames were amplified with recombination sites from
plasmids by Phusion polymerase chain reaction (PCR) from Finnzymes following the
manufacteur’s instructions. Primers used to generate inserts for middle entry vectors:
Heat-shock protein 70 (Hsp70) promoter in 5’entry plasmid, and IRES:mGFP 3’entry
plasmids were obtained from the Lawson Tol2 Kit. Entry plasmids were recombined into
a destination vector to generate Hsp70:wnt:IRESmGFP constructs. Chorinated zebrafish
embryos obtained from a natural mating were injected with 25 pg of Tol2 mRNA and 25
pg of transgene plasmid DNA. Embryos at early somite stages were subject to heat-
shock at 37°C for 30 minutes to induce expression of Wnt protein and drive mGFP
expression.
2.4: References
[1] C.P. Heisenberg, M. Tada, G.J. Rauch, L. Saúde, M.L. Concha, R. Geisler, D.L. Stemple, J.C. Smith, and S.W. Wilson, “Silberblick/Wnt11 mediates convergent extension movements during zebrafish gastrulation,” Nature, vol. 405, May. 2000, pp. 76-81.
[2] B. Kilian, H. Mansukoski, F.C. Barbosa, F. Ulrich, M. Tada, and C.P. Heisenberg, “The role of Ppt/Wnt5 in regulating cell shape and movement during zebrafish gastrulation,” Mechanisms of Development, vol. 120, Apr. 2003, pp. 467-476.
[3] J. Gros, O. Serralbo, and C. Marcelle, “WNT11 acts as a directional cue to organize the elongation of early muscle fibres,” Nature, vol. 457, Jan. 2009, pp. 589-593.
[4] B. Goldstein, H. Takeshita, K. Mizumoto, and H. Sawa, “Wnt signals can function as positional cues in establishing cell polarity,” Developmental Cell, vol. 10, Mar. 2006, pp. 391-396.
[5] B. Ciruna, A. Jenny, D. Lee, M. Mlodzik, and A.F. Schier, “Planar cell polarity signalling couples cell division and morphogenesis during neurulation,” Nature, vol. 439, Jan. 2006, pp. 220-224.
[6] T. Matsui, A. Raya, Y. Kawakami, C. Callol-Massot, J. Capdevila, C. Rodríguez-Esteban, and J.C. Izpisúa Belmonte, “Noncanonical Wnt signaling regulates midline convergence of organ primordia during zebrafish development,” Genes &

50
Development, vol. 19, Jan. 2005, pp. 164-175. [7] A.R. Ungar, G.M. Kelly, and R.T. Moon, “Wnt4 affects morphogenesis when
misexpressed in the zebrafish embryo,” Mechanisms of Development, vol. 52, Aug. 1995, pp. 153-164.
[8] M. Kurayoshi, H. Yamamoto, S. Izumi, and A. Kikuchi, “Post-translational palmitoylation and glycosylation of Wnt-5a are necessary for its signalling,” The Biochemical Journal, vol. 402, Mar. 2007, pp. 515-523.
[9] B. Geldmacher-Voss, A.M. Reugels, S. Pauls, and J.A. Campos-Ortega, “A 90-degree rotation of the mitotic spindle changes the orientation of mitoses of zebrafish neuroepithelial cells,” Development (Cambridge, England), vol. 130, Aug. 2003, pp. 3767-3780.

51
CHAPTER 3: Discovery of Novel Regulators of Planar Cell Polarity
3.0: Introduction
Drosophila planar cell polarity (PCP) studies suggest the global cue that
establishes PCP most likely acts through the membrane proteins of the core PCP group,
namely Flamingo (Fmi), Frizzled (Fz) or Van gogh like 2 (Vangl2). Fmi is a seven-pass
atypical cadherin, thought to regulate adhesion between cells and support formation of
complexes with Fz and Vangl2. The global cue that establishes PCP is not likely
transduced through Fmi since Fmi acts cell autonomously and is not asymmetrically
localized across the cell. Fz is characterized as a Wnt receptor, suggesting that Wnt
protein may regulate PCP. The role of Wnt signalling in regulating PCP is the topic of
investigation in Chapter 2. Vangl2 is a four pass transmembrane protein that is
asymmetrically localized and acts cell non-autonomously. There are no known soluble
ligands for Vangl2 and very few membrane proteins interact with Vangl2. Scribble and
Discs Large, two proteins involved in apical-basal polarity and Fmi have been identified
to physically interact in cis [1][2]. To further characterize Vangl2, an essential PCP
component, a screen for novel protein interactions was performed to potentially discover
a novel co-receptor that may respond to the global cue to establish PCP.
A common approach to search for novel protein-protein interactions is to perform
a yeast-two-hybrid (YTH), where a bait protein is screened against a library of prey
proteins. The bait consists of the protein of interest fused to a portion of a transcription
factor. The prey consists of a protein from a library fused to the complementary portion
of the transcription factor. When the bait and prey proteins interact, the complementary
portions of the transcription factor interact and form an intact functional transcription

52
factor. Subsequently, the bait-prey complex can enter the nucleus and activate target
reporter genes. Although the conventional YTH is a useful tool to assess protein-protein
interactions, there are some limitations. Integral membrane proteins, due to their highly
hydrophobic nature cannot be analyzed by the conventional method since they are unable
to translocate to the nucleus to activate the reporter genes.
To detect protein-protein interactions with membrane proteins, a modified YTH
method has been employed in collaboration with Igor Stagjlar. Membrane-yeast-two-
hybrid (MYTH) allows for membrane protein-protein interactions to be detected. MYTH
uses a split-ubiquitin approach, where the transcription factor translocates to the nucleus
independent of the bait and prey proteins, allowing membrane proteins to be used in the
screen (Figure 14) [3]. MYTH is based on observation that ubiquitin can be separated
into two components (termed Nub and Cub) that functionally reconstitute when in close
proximity to one another [3]. The bait is tagged with the Cub (C-terminal half of
ubiquitin) and an artificial transcription factor [3]. The prey is tagged with the Nub (N-
terminal half of ubiquitin). The bait-prey interaction brings the Cub and Nub moieties in
close proximity to each other and facilitates reconstitution of ubiquitin [3]. The presence
of ubiquitin is recognized by ubiquitin-specific proteases found in all eukaryotic cells,
resulting in cleavage of the transcription factor attached to the Cub domain [3]. The
released transcription factor enters the nucleus and activates the reporter genes [3].

53
Figure 14: Membrane Yeast Two Hybrid (MYTH) A. Membrane protein of interest is fused to Cub domain followed by an artificial transcription factor LexA-VP16, while another prey protein (cytosolic or membrane) is fused to the NubG domain. If bait and prey do not interact, the Cub and Nub domains will not reconstitute to form ubiquitin, no cleavage of the transcription factor occurs, resulting in no activation of the reporter genes. B. If bait and prey interact, Nub and Cub domains are brought into close proximity and ubiquitin reconstitution is able to occur. Ubiquitin is recognized by proteases inside the cell and cleaves the artificial transcription factor and enters the nucleus. Inside the nucleus, the transcription factor binds the LexA operator activating reporter genes.

54
In zebrafish, there are two Vang homologues, vangl1 and vangl2. The expression
of zebrafish vangl2 is widely expressed throughout zebrafish development and is
maternally provided. vangl1 is not expressed maternally and is first detected at the 15-
somite stage in the developing nervous system [4]. The expression of vangl1 appears to
not overlap with vangl2, since by 1 day post fertilization, vangl1 expression is restricted
to the hindbrain whereas vangl2 expression is found in the neuroectoderm, neural tube
and hatching gland [4]. Despite the non-overlapping expression patterns, injection of
vangl1 mRNA into vangl2 mutant zebrafish embryos is able to partial rescue convergent
extension defects, suggesting a functional similarity between Vangl1 and Vangl2 [4]. The
distinct roles of Vangl1 and Vangl2 is unclear. The objective of the MYTH screen is to
discover novel regulators of PCP by identifying new protein-protein interactions with
Vangl1 and Vangl2. Possibly, other membrane proteins that may act as PCP co-receptors
will be identified as well as downstream proteins that will show similar or distinct roles
for Vangl1 and Vangl2.
3.1: Results
3.1.1: Characterization of Baits
Bait proteins were constructed using an in vivo recombination approach in yeast.
Using primers, the open reading frames of Vangl1 and Vangl2 were amplified by
polymerase chain reaction (PCR) with flanking regions homologous to the bait vector.
Bait vectors were linearized and transformed into competent yeast cells with purified
PCR products. Bait vectors encode the leu2 gene product, which permits yeast to grow on
selective plates without leucine. Yeast growing on plates without leucine were

55
individually picked and plasmids were isolated and sequenced. Vangl1 and Vangl2 baits
with N- and C-terminal tags were constructed using this method.
To characterize the constructed bait proteins, tests for activation of the reporter,
localization of the bait and stringency of interaction were performed. The NubG/I test
was used, inwhich the N-terminal portion of ubiquitin (NubI) or N-terminal portion of
ubiquitin with a point mutation (NubG) is transformed with the bait vector into
competent yeast cells. NubI acts as a positive control since it has a high affinity for the
Cub component and will spontaneously reconstitute in vivo to form ubiquitin. The
transcription factor is cleaved and able to translocate into the nucleus to activate the three
reporter genes, HIS3, ADE2 and lacZ. The gene products of HIS3 and ADE2 allow yeast
to grow on plates without histidine and adenine, which normally inhibits growth. Growth
on plates (without histidine and adenine) suggests that the bait constructs are functional
and capable of activating the reporter.
NubG acts as a negative control since it contains a point mutation that prevents
spontaneous reconstitution with the Cub domain. Instead, NubG needs to be brought into
close proximity, such as with bait-prey interaction. However since only NubG will be
transformed with the bait, no interaction should occur, resulting in no activation of the
reporter. Any growth on the selective plates is indicative of background that will be
obtained in the MYTH screen. To determine the stringency of the screen, yeast cells were
transformed with the bait and NubG and assayed on selective plates (minus histidine and
adenine) with varying concentrations of 3-amino-1,2,4-triazole (3-AT). 3-AT is a
competitive inhibitor of the HIS3 gene product and can be added to the plate media to
remove background due to the leakiness of the reporter. Finally to test for localization of

56
the bait construct, the Nub proteins were targeted to the plasma membrane and assayed
for presence in the endoplasmic reticulum (ER). The localization of the bait can be
gauged by localization of the NubI, activation of the reporter and growth on the selective
plates.
Four bait constructs consisting of N- and C-terminal Cub tagged Vangl-1 and
Vangl-2 were generated (Table 1). All constructs appeared to be capable of activating
reporter since there was growth when the baits were transformed with NubI. The N-
terminal Cub-Vangl1 appeared to localize to the ER and plasma membrane. However, the
N-terminal Cub-Vangl2 bait appeared to localize only to the plasma membrane (Figure
15C). The C-terminal Vangl1/2-Cub fusion appeared to localize to both the ER and the
plasma membrane. Examining the leakiness of the reporter, there appeared to be some
background with the C-terminal Vangl2-Cub bait, which can be decreased by addition of
10 mM 3-AT. Since the C- terminus of Vangl2 is better characterized and was shown to
consist of a PDZ binding domain, the MYTH screen was conducted with the C-terminal
Cub baits.
3.1.2: Analysis of MYTH screen hits
Three screens were performed with the C-terminal baits for Vangl-1 and -2.
Collectively, the Vangl-1 screens generated 90 hits of which 39 were unique hits. The
Vangl-2 screen generated 67 hits where 56 hits were unique (Table 2). Expected hits such
as other members of the PCP signalling pathway, namely Dishevelled and Prickle, were
not detected in the screen. However, proteins affiliated with the pathway, such as
Receptor Tyrosine Kinase-like Orphan receptor 1 (ROR1) and Vangl-1 were found to

57
Table1: Summary of bait construction Bait Plasma ER Stringency
V1-Cub Y Y 0 mM V2-Cub Y Y 10 mM Cub-V1 Y Y 0 mM Cub-V2 N Y 0 mM
A
B
AS

58
C
D
Figure 15: Characterization of Baits NubG/I test to check for activation of reporter, localication and background of bait proteins NubI has a high affinity for the Cub domain, acts as a positive control. NubG contains a point mutation and needs to be brought into close proximity with protein interaction. NubG should act as a negative control since no prey protein is present. Selective conditions are indicated as follows: W - tryptophan, L - Leucine, A – Adenine, H – Histidine, 10/25/50/100 mM amounts of 3AT. –W selects for presence of prey vector, -L selects for presence of bait vector. –AH selects for activation of reporter genes. A. Cub-Vangl1 is localized to the ER and plasma membrane and is able to activate the reporter without any background B. Vangl1-Cub is localized to both the ER and plasma membrane and is able to activate the reporter without any background. C. Cub-Vangl2 is localized only to the plasma membrane and is able to activate the reporter without any background. D. Vangl2-Cub localizes to both the ER and plasma membrane and is able to activate the reporter with little background. Addition of 3AT diminishes the background.

59
Table2a: Vangl1 MYTH screen hits Accession # Gene Frequency NM_001001995 glycoprotein M6B (GPM6B) 2 NM_001044 solute carrier family 6 member 3 (SLC6A3) 2 NM_001083592 receptor tyrosine kinase-like orphan receptor 1 (ROR1) 8 NM_002155 heat shock 70kDa protein 6 1 NM_002333 low density lipoprotein receptor-related protein 3 1 NM_002501 nuclear factor I/X (CCAAT-binding TF) (NFIX) 1 NM_003257 tight junction protein 1 (zona occludens 1) (TJP1) 1 NM_003277 claudin 5 1 NM_004607 tubulin folding cofactor A (TBCA) 1 NM_005669 receptor accessory protein 5 (REEP5) 1 NM_005745 B-cell receptor-associated protein 31 (BCAP31) 10 NM_005909 microtubule-associated protein 1B (MAP1B) 1 NM_006044 histone deacetylase 6 (HDAC6) 1 NM_006292 tumor susceptibility gene 101 (TSG101) 1 NM_014041 signal peptidase complex subunit 1 homolog 1 NM_014051 transmembrane protein 14A 12 NM_014145 chromosome 20 open reading frame 30 (C20orf30) 1 NM_014445 stress-associated endoplasmic reticulum protein 1 1 NM_014713 lysosomal-associated protein transmembrane 4 alpha 1 NM_015407 abhydrolase domain containing 14A (ABHD14A) 1 NM_016145 chromosome 19 open reading frame 56 (C19orf56) 1 NM_016224 sorting nexin 9 (SNX9) 1 NM_017655 GIPC PDZ domain containing family, member 2 (GIPC2) 1 NM_018930 protocadherin beta 10 (PCDHB10) 2 NM_018936 protocadherin beta 2 (PCDHB2) 1 NM_020762 SLIT-ROBO Rho GTPase activating protein 1 1 NM_021009 ubiquitin C 2 NM_024532 sperm associated antigen 16 (SPAG16) 1 NM_032927 transmembrane protein 128 (TMEM128) 1 NM_138959 vang-like 1 (van gogh, Drosophila) (VANGL1) 1 NM_152464 transmembrane protein 199 (TMEM199) 1 NM_153757 nucleosome assembly protein 1-like 5 (NAP1L5) 1 NM_173834 Yip1 domain family, member 6 (YIPF6) 2 NM_173852 keratinocyte associated protein 2 (KRTCAP2) 1 NM_177424 syntaxin 12 (STX12) 1 NM_194359 ring finger protein 41 (RNF41) 20 NM_207521 reticulon 4 (RTN4) 1 XM_001717663 PREDICTED: similar to hCG23722 (LOC100128731) 1 XM_002344544 PREDICTED: similar to Cytochrome c oxidase subunit 2 1

60
Table2b: Vangl2 MYTH screen hits Accession # Gene Frequency NM_175607 contactin 4 (CNTN4) 1 NM_178014 tubulin, beta (TUBB) 1 ABG29260 NADH dehydrogenase subunit 2 1 NM_001002021 phosphofructokinase, liver (PFKL) 1 NM_001003 ribosomal protein, large, P1 (RPLP1) 1 NM_001006623 WD repeat domain 33 (WDR33) 1 NM_001008390 CGG triplet repeat binding protein 1 (CGGBP1) 1 NM_001024660 kalirin, RhoGEF kinase (KALRN) 1 NM_001040058 secreted phosphoprotein 1 (SPP1) 1 NM_001204 bone morphogenetic protein receptor, type II 2 NM_001614 actin, gamma 1 (ACTG1) 3 NM_001806 CCAAT/enhancer binding protein (C/EBP), gamma (CEBPG) 1 NM_002045 growth associated protein 43 (GAP43) 1 NM_002055 glial fibrillary acidic protein (GFAP) 2 NM_002585 pre-B-cell leukemia homeobox 1 (PBX1) 1 NM_002601 phosphodiesterase 6D, cGMP-specific, rod, delta (PDE6D) 1 NM_002792 proteasome subunit, alpha type, 7 (PSMA7) 1 NM_002847 protein tyrosine phosphatase receptor N polypeptide2(PTPRN2) 1 NM_003127 spectrin, alpha, non-erythrocytic 1 (alpha-fodrin) 2 NM_003277 claudin 5 1 NM_003388 CAP-GLY domain containing linker protein 2 (CLIP2) 1 NM_004537 nucleosome assembly protein 1-like 1 (NAP1L1) 1 NM_004615 tetraspanin 7 (TSPAN7) 1 NM_005345 heat shock 70kDa protein 1 NM_005917 malate dehydrogenase 1, NAD (soluble) (MDH1) 1 NM_007369 G protein-coupled receptor 161 (GPR161) 1 NM_013442 stomatin (EPB72)-like 2 (STOML2) 1 NM_014717 zinc finger protein 536 (ZNF536) 1 NM_014787 DnaJ (Hsp40) homolog, subfamily C, member 6 (DNAJC6) 1 NM_015123 FERM domain containing 4B (FRMD4B) 1 NM_015570 autism susceptibility candidate 2 (AUTS2) 1 NM_017811 ubiquitin-conjugating enzyme E2R 2 (UBE2R2) 1 NM_017812 coiled-coil-helix-coiled-coil-helix domain containing 3 (CHCHD3) 1 NM_017921 nuclear protein localization 4 homolog 1 NM_018273 transmembrane protein 143 (TMEM143) 1 NM_020240 CDC42 small effector 2 (CDC42SE2) 1 NM_021948 brevican (BCAN) 1 NM_022777 RAB, member RAS oncogene family-like 5 (RABL5) 1 NM_022895 chromosome 12 open reading frame 43 (C12orf43) 1 NM_030939 chromosome 6 open reading frame 62 (C6orf62) 1 NM_033119 naked cuticle homolog 1 (Drosophila) (NKD1) 1 NM_170692 RAS protein activator like 2 1 NM_170774 Ras association domain family member 2(RASSF2) 1 NM_182472 EPH receptor A5 (EPHA5) 1 NM_182480 coenzyme Q6 homolog, monooxygenase 1

61
Table 2b: Vangl2 MYTH screen hits (continued) Accession # Gene Frequency NM_194317 LY6/PLAUR domain containing 6 (LYPD6) 1 NM_194359 ring finger protein 41 (RNF41) 7 NM_203401 stathmin 1/oncoprotein 18 (STMN1) 1 NM_206962 protein arginine methyltransferase 2 (PRMT2) 1 XM_001716421 R3H domain and coiled-coil containing 1 (R3HCC1) 1 XM_001721542 hypothetical protein 1 XM_035299 zinc finger, SWIM-type containing 6 (ZSWIM6) 1 XR_017149 PREDICTED: Homo sapiens misc_RNA (LOC392437) 1 XR_017548 PREDICTED: Homo sapiens misc_RNA (LOC399942) 1 XR_040065 PREDICTED: Homo sapiens misc_RNA (OTUD1) 1 XR_041470 PREDICTED: Homo sapiens misc_RNA (LOC389634) 1

62
interact with Vangl-1 [5]. The Vangl-2 screen showed interaction with Naked Cuticle
and Bone Morphogenetic Protein Receptor, both of which have been implicated in PCP
signalling [6][7]. MYTH data were analyzed through the use of Ingenuity Pathways
Analysis, where the hits were clustered into known signalling networks and signalling
pathways (Ingenuity® Systems, www.ingenuity.com). The screens resulted in being
clustered to the molecular/cellular network, more specifically cellular
assembly/organization (Figure 16A). The top signalling pathway associated with the hits
was tight-junction signalling for both Vang homologues (Figure 16B). In Drosophila
studies, the core PCP proteins have been found to localize and interact with many of the
junction components, suggesting the identified interactions in the MYTH screen are valid
[8].
3.1.3: Validation of MYTH screen
The MYTH screens conducted did not yield any known protein-protein
interactions that have been shown to interact physically with Vangl1/2. To ensure the hits
obtained from the MYTH screen represented valid protein-protein interactions, positive
control prey constructs were made. Scribble and Prickle have been shown to interact
physically with Vangl-2 and by transforming the Vangl-2-Cub bait with NubG-Scribble
or NubG-Prickle prey vectors, revealed a direct protein interaction (Figure 17) [1][9].
Both NubG-Scribble and NubG-Prickle1 activated the reporter, where growth was
observed on selective plates. Another method to validate the MYTH screen was to choose
one of the hits and check for physical interaction with Vangl1/2. The hit selected was

63
Figure 16: Analysis of Hits Using Ingenuity Pathway Analysis software, hits from the MYTH screen were clustered into (A) Network Functions and (B) Signalling Pathways.
A
B

64
Figure 17: Validation of MYTH Screen with Known Protein Interactions To validate the screen, prey constructs of proteins known to interact with Vangl2 were chosen. –W (tryptophan) selects for presence of prey vector. –L (Leucine) selects for presence of bait. –AH (adenine, histidine) selects for interaction. 3AT, 25 mM is added to remove possible background. Xgal, substrate for ß-galacosidase reporter gene, results in blue colonies. Scribble and Prickle have been shown to interact physically with Vangl2, which is shown by growth on selective plates with stringency and blue colonies on the selective Xgal plate. PTK7 although implicated in PCP signalling has not been shown to interact with Vangl2. No growth is seen on selective plates with 3AT and no blue colony is formed.

65
RING Finger 41 (RNF41), since it was found to interact with both Vangl-1 and Vangl-2
with the highest frequency. To confirm the interaction, an in vivo approach was taken
utilizing zebrafish embryos. Zebrafish RNF41 was cloned from cDNA and subcloned
into an in vitro mRNA expression vector. mRNA encoding myc-tagged zebrafish Vangl-
2 and flag-tagged zebrafish RNF41 were injected into zebrafish embryos and protein
lysate was collected. To test for a physical interaction, co-immunoprecipitation was
performed which showed that Vangl-2 and RNF41 physically interact (Figure 18).
RNF41 encodes an E3 ubiquitin ligase that contains a ring finger motif and USP8
interaction domain. E3 ubiquitin ligases are involved in transferring ubiquitin from the
E2 ubiquitin ligase to the substrate to be ubiquitinated. The ring finger motif is
responsible for substrate specificity. USP8 is a deubiquitinating enzyme, which has been
shown to physically interact with RNF41 in the USP8 interaction domain. RFN41 has
been shown to interact and regulate steady-state levels of the ErbB3 receptor,
ubiquitinates Parkin as well as regulating its own stability [10]. Expression of USP8
appears to stabilize RNF41, further suggesting that RNF41 is ubiquitinated. From the
MYTH screen, possibly RNF41 may regulate the trafficking or stability of Vangl-1 and -
2 and may act as a novel regulator of PCP signalling. Ubiquitination has recently been
shown to regulate PCP signalling and potentially RNF41 may act in the pathway to
regulate PCP protein stability[11].
3.1.4: Ectopic expression of RNF41 disrupts PCP signalling
The protein interaction between RNF41 and Vangl2 suggests RNF41 may
regulate PCP. First, the endogenous expression was examined to determine if RNF41 is

66
Figure 18: Co-immunoprecipitation shows that Vangl2 and RNF41 physically interact IB: Immunoblotted, IP: Immunoprecipitated, IgGH: Immunoglobulin Heavy-chain, IgGL: Immunoglobulin Light-chain, + presence, - absence. A light-chain specific antibody was used since Vangl2-Myc separated to approximately the same molecular weight as the heavy-chain. Two murine antibodies were used to IP and IB and even with the IgGL-specific secondary antibody, a faint band was detected in all sample lanes, most likely corresponding to the heavy chain. * indicates a higher molecular weight band detected in blots probed with myc antibody when Vangl2-myc is expressed. * appears in the lysate control (not shown) and IP, suggesting Vangl2-Myc was immunoprecipitated. Protein concentrations were not equalized between samples.
Flag-RNF41 - + + - Vangl2-Myc + - + -
IP: Myc
IB: Flag
IgG L
IgG H
Flag-RNF41 - + + -
Vangl2-Myc + - + -
Lysates
IB: Myc
IB: Flag
IB: Myc
* *

67
expressed at time when the embryo is undergoing convergent extension (CE) movements
(Figure 19). An anti-sense RNA probe was generated using the full length RNF41.
Zebrafish RNF41 appears to be expressed maternally and persists ubiquitously after the
onset of zygotic transcription and at early stages of neurulation. Since RNF41 mRNA is
expressed when the embryo is undergoing convergent extension movements, the next
assay was to determine if RNF41 could regulate PCP signalling. To test whether PCP
signalling was disrupted, RNF41 was ectopically expressed by injecting RNF41 mRNA
into the 1-cell stage embryo. A concentration dependent defect in the extension of the
embryo was observed as compared to un-injected embryos, suggesting that RNF41 may
regulate PCP (Figure 20A-E). To ensure a defect in CE was the cause of the phenotype,
embryos injected with 200 pg of RNF41 mRNA were probed for krox20 and myoD which
marks the midbrain-hindbrain boundary and somites respectively to clearly show the
presence of a CE defect (Figure 20F,G). Embryos were time-matched and flat mounted
and comparing a non-injected embryo to a RNF41 mRNA injected embryo, the embryo
with RNF41 over-expression shows a broader and shorter body axis as compared to the
non-injected control.
To determine if RNF41 has a direct role in regulating PCP signalling, GFP-Pk
localization was examined in response to RNF41 over-expression. Embryos were
injected with GFP-Pk at the 1-cell stage and subsets of those embryos were injected again
with varying concentrations of RNF41 mRNA (Figure 21). WT embryos injected with
GFP-Pk possess puncta localized to the membrane. With increasing concentrations of
RNF41, the number of puncta decreased, where with injection 400 pg of RNF41 mRNA,

68
Figure 19: Endogenous RNF41 expression RNF41 RNA in situ hybridization of zebrafish embryos at different stages. A-C: RNF41 anti-sense RNA probe. D-F: RNF41 sense probe A,D: 2-cell stage. B,D: Sphere stage. C,E:. 6-somite stage.

69
Figure 20: Ectopic RNF41 expression induces convergent extension defects A-E: Lateral views of zebrafish embryos at 14-16 hpf. A: WT. B. 100 pg RNF41 mRNA injection. C: 200 pg RNF41 mRNA injection. D: 200 pg GFP-RNF41 mRNA injection. E: 400 pg GFP-RNF41 mRNA injection. F,G: Flat mount MyoD and Krox20 anti-sense RNA in situ hybridization of time-matched zebrafish embryos at 14-16 hpf. * indicates Krox20 staining. Dash line indicates MyoD staining. F: WT embryo. G: 200 pg RNF41 mRNA injection.
* *

70
Figure 21: RNF41 regulates GFP-Pk localization Confocal microscopic sections along the dorsal-ventral axis of 14-16 hpf zebrafish embryos injected with memRFP (red) and GFP-Pk (green). All WT embryos were injected with GFP-Pk and memRFP. A: Subset of embryo with no second injection. B: Subset of embryos with second injection of 100 pg of RNF41 mRNA. C: Subset of embryos with second injection of 200 pg of RNF41 mRNA. D: Subset of embryos with second injection of 400 pg of RNF41 mRNA.
A B
C D

71
GFP-Pk puncta localization was lost. The cytoplasmic levels of GFP-Pk also appeared to
be reduced in a concentration dependent manner, with the levels of memRFP remaining
constant at all concentrations. This suggests that RNF41 may directly regulate PCP
signalling by interacting with GFP-Pk and potentially regulating its stability.
3.1.5: RNF41 subcellular localization suggests a new link between PCP and cilia
To further characterize the role of RNF41, GFP-RNF41 fusion protein was
constructed and injected into zebrafish embryos. GFP fluorescence was found throughout
the neural keel with weak cytoplasmic staining. At neural rod stages, GFP puncta were
observed just below the apical midline, near the ventral floor plate. The punctate pattern
observed appeared similar to the organization of cilia at the floor plate of the neural tube
(Figure 22A). To determine if GFP-RNF41 puncta were related to cilia, a marker of cilia
available in the lab was used. GFP-RNF41 mRNA was injected into Cilia-RFP marker
transgenic embryos and imaged using confocal microscopy (Figure22B). Examining the
ventral floor plate at neural rod stages showed similar expression pattern, where RNF41
puncta localized to the base of the cilium, possibly at the basal body.
3.2: Discussion
3.2.1: RNF41 may regulate planar cell polarity through Vangl2
RNF41 is an E3 ubiquitin ligase that directly interacts with Vangl2 and may
regulate PCP by affecting Vangl2 stability, Vangl2 localization or Pk stability.
Ubiquitination of membrane proteins causes either the recycling of the membrane protein

72
Figure 22: RNF41 localization suggests a role in regulating cilia Confocal sections along the dorsal-ventral axis of zebrafish neural rod. A: Embryos injected with memRFP (red) and 200 pg of GFP-RNF41 (green) mRNA. GFP-RNF41 is cytoplasmic and found in all cells the neural keel/rod. When the midline forms, puncta can be found on either side of the midline, close to the membrane. B: Embryos derived from a cross of cilia-RFP/- transgenic zebrafish (red) injected with 200 pg of GFP-RNF41 (green) mRNA. RNF41 appear to localize with each cilium.
A B

73
through the endosomal pathway or it is degraded by the lysosome. It is unclear whether
Vangl2 is ubiquitinated, since no other E3 ubiquitin ligase has been found to interact with
Vangl2. To begin to assess whether RNF41 regulate Vangl2 stability, injection of
zebrafish embryos with Vangl2-GFP and RNF41 can be performed. If RNF41 regulates
Vangl2 stability, loss of Vangl2-GFP at the membrane would be expected with co-
expression of RNF41. If RNF41 does not regulate Vangl2 stability, Vangl2-GFP
membrane fluorescence would expected to be comparable to WT expression.
Asymmetric localization of PCP components is a hallmark of PCP signalling. Studies in
cell culture have shown that Smurf E3 ubiquitin ligases regulate asymmetric localization
of Pk [11]. Upon complex formation of Pk, Dsh and Par6, Smurf is recruited to the
complex and targets Pk for ubiquitin-dependent degradation [11]. Possibly, Vangl2 is
degraded by RNF41 and if RNF41 is itself asymmetrically localized, it may promote the
asymmetric distribution of Vangl2. The endogenous localization of RNF41 needs to be
examined. It is unclear whether RNF41 acts on Vangl2 or Pk to regulate PCP. The loss of
GFP-Pk puncta could result from RNF41 directly regulating Pk stability. Vangl2 could
act to recruit Pk and RNF41, allowing RNF41 to interact and target Pk for ubiquitin-
mediated degradation. This additional method to regulate Pk protein levels could act with
Smurf to provide a means to regulate Pk stability across the entire cell. The model that I
propose is that Vangl2 acts as a scaffold to localize Pk and RNF41. When in close
proximaty, RNF41 binds and ubiquitinates Pk, targeting it for degradation.

74
3.2.2: RNF41 localizes to the basal body and may regulate non-canonical signalling
Disruption of basal body by loss-of-function of basal body proteins such as
bardet-biedl syndrome 4 (bbs4) in zebrafish results in embryos with CE defects [12].
Furthermore, epistasis analysis indicates that bbs4 functions downstream of wnt5b and
wnt11 [12]. In addition to the PCP defects, bbs4 morphants also possess canonical Wnt
defects, indicated by the expanded axin2 domain relative to wild-type [12]. The affect on
canonical and non-canonical Wnt signalling was further characterized with in vitro
experiments. In mono-ciliated mammalian cells, stimulation of Wnt5a has been shown to
inhibit canonical signalling by inhibiting up-regulation of the canonical Wnt signalling co-
transcription factor (TCF/LEF). With disruption of the basal body, the ability of Wnt5a
to suppress TCF/LEF up-regulation is lost [12]. Possibly disruption of the basal body
alters the behaviour of a common effector molecule to both pathways, desensitizing the
cell to Wnt stimulation [12]. A possible candidate is Dsh as it mediates both canonical
and non-canonical Wnt signalling as well as mis-expression of Dsh is able to partially
rescue bbs4 morphants [12]. Potentially, RNF41 localizes to the basal body and is able to
mediate some aspect downstream of non-canonical Wnt signalling.
3.3: Methods
3.3.1: Materials
Amino acids, agar and components for yeast media were obtained from BioShop.
Bait/prey vectors and Human embryonic cDNA library were obtained from Igor
Stagljar/Dualsystems Biotech. Saccharomyces cercisiae THY.AP4 [MATa leu2-3, 112

75
ura3-52 trp1-289 lexA::HIS3 lexA::ADE2 lexA::lacZ] yeast strain was obtained from
Igor Stagljar Lab. Plasmid isolation, gel extraction and PCR puficiation kits were
obtained from Qiagen. Other biological reagents and buffers were obtained from Sigma-
Aldrich. High-fidelity Taq Phusion PCR kit was obtained from Finzymes and primers
used in PCR were obtained from Sigma-Aldrich. Human Vangl-1, Vangl-2 and Prickle-1
clones were obtained from Open Biosystems and the mouse Scribble plasmid was
obtained from Jane McGlade’s Laboratory.
3.3.2: Bait/Prey vector construction by homologous recombination/gap repair
PCR primers containing the homologous region of vector backbone and 5’ or 3’ region of
the open-reading frame of the gene of interest (Vangl-1, Vangl-2, Prickle-1, Scribble)
were used to obtain a PCR product with homologous regions flanking both ends of the
open-reading frame. Primers used to generate bait/prey inserts: Vangl2-Cub Forward:
ATGTCTGATGCGGCTCCTTCATTGAGCAATCTATTTTATATGGACACCGAGTC
CCAGTAC, Reverse: GTTGATCTGGAGGGATCCCCCCCGACCAATCTATTTTATA
TGGATACCGAATCCACTTATTC. Vangl1-Cub Forward: ATGTCTGATGCGGCTCC
TTCATTGAGCAATCTATTTTATATGGATACCGAATCCACTTATTC, Reverse:
GTTGATCTGGAGGGATCCCCCCCGAC ATGGTCGACGGTATAACGGATGTCT
CAGACTGTAAGC. Cub-Vangl2 Forward: GCCA GGCCTTTAATTAAGGCCCCAAT
GGACACCGAGTCCCAGTAC, Reverse: CATGACCT ATTAAGATCTGACGTCAGC
GCTCCGCGTCCGCGTCACACTGAGGTCTCAGACTG. Cub-Vangl1 Forward:
GCCAGGCCTTTAATTAAGGCCGCCTCGGCCCCAATGGATAC CGAATCCACTT
ATTC, Reverse: CATGACCTATTAAGATCTGACGTCAGCGCTCCGC GTTAAACG

76
GATGTCCAGACTG. Nub-Scrib Forward: CCAAGCAGTGGTATCAACGCAGAGTG
GCCATTACGGCCCATGCTGAAGTGCATCCCGC. Reverse: CGAATTCTCGAG
AGGCCGAGGCGGGCCGACATGTTTTTTCCCCTAGGAGGGCACAGGGCCC.
Nub-Prickle1 Forward: CCAAGCAGTGGTATCAACGCAGAGTGGCCATTACG
GCCCATGCC TTTGGAGATGGAGCCC , Reverse: CGAATTCTCGAGAGGCCGA
GGCGGGCCGACAT GTTTTTTCCCTTAAGAAATAATACAATTTTTGCCC. PTK7-
Nub Forward: CCAAGCAG TGGTATCAACGCAGAGTGGCCATTACGGCCCATGG
GAGCTGCGCGGGGATCC, Reverse: CGAATTCTCGAGAGGCCGAGGCGGCCGA
CATGTTTTTTCCCTCACGGCT TGCTGTCCACGG. High-fidelity Taq Phusion PCR
kit was used to amplify the open-reading frame following the manufacture’s protocol
without any deviations. PCR products were purified using Qiagen PCR purification kits
following the manufacture’s directions without any changes. Compentent yeast cells were
transformed with linearized bait (pCCWSTE linearized with HindIII and PstI, pBTN3
linearized with NcoI and SacII) or prey (SmaI) and PCR products containing open-reading
frames of Vangl-1 and -2. Bait and prey plasmids containing insert were selected for by
growing transformed yeast on media plates without leucine or tryptophan respectively.
3.3.3: NubG/I Test
Bait vectors are transformed into competent yeast cells and plated on selective plates (-
leucine) for successful transformation. Yeast containing bait vectors are grown to
competency and transformed with empty prey vectors (Nub domain only) targeted either
to the plasma membrane or endoplasmic reticulum (ER). Two different Nub domains are
used, NubI and NubG to test for ability to activate the reporter (positive control) and the

77
amount of background (negative control) respectively. To test for the level of stringency,
the transformed yeast containing both the bait and empty prey vectors are plated on
media with different concentrations of 3-aminotriazole, a competitive inhibitor of the his3
reporter gene.
3.3.4: MYTH
Bait vectors are transformed into competent yeast cells and plated on selective plates (-
leucine) for successful transformation. Yeast containing bait vectors are grown to
competency and transformed with a cDNA prey library (human fetal brain obtained from
the Igor Stagljar Lab). Transformed yeast was selected for interaction on plates without
leucine, tryptophan, adenine, histidine and corresponding amount of 3-AT (10
mMVangl2-Cub and 0 mM Vangl1-Cub) as determined by the NubG/I test. Colonies on
selection plates were replated on plates containing x-gal, where positive blue colonies
were picked and plasmid DNA was isolated using QIAGEN plasmid DNA isolation kits.
Prey cDNA was amplified using High-fidelity Taq Phusion PCR kit with primers:
Forward: GTCGAAATTCAAGACAA GG, Reverse: CGTGAATGTAAGCGTGAC.
PCR products were purified using QIAGEN PCR purification kits and sequenced using
the forward PCR primer for sequencing. Sequenced prey cDNA was identified using
BLAST against human genome.
3.3.5: Zebrafish microinjection
Zebrafish embryos were obtained from natural mating of wildtype (WT) TL and AB
hybrid backgrounds. Plasmids containing membrane-localized red fluorescent protein

78
(memRFP), EGFP-Prickle (GFP-Pk), Myc-Vangl2, RNF41, GFP-RNF41, Flag-
RNF41were linearized and sense-strand-capped mRNA was synthesized with the
mMESSAGE mMACHINE system. Zebrafish embryos were dechorionated by treatment
of pronase and injected at the one-cell stage. Scatter labeling was obtained by injecting
two blastomeres at the eight-cell stage.
3.3.6: Immunoprecipitation and Western Blotting
WT embryos at the 1-cell stage were injected with Myc-Vangl2 and/or Flag-RNF41
mRNA. Embryos were developed to sphere stage and cells were mechanically dissociated
using calcium-free conditions following the procedure outline in the Zebrafish Book
(Westerfield, M. (2000). The zebrafish book. A guide for the laboratory use of zebrafish
(Danio rerio). 4th ed., Univ. of Oregon Press, Eugene.). Sphere stage embryos were
rinsed in calcium-free Ringer’s solution (116 mM NaCl, 2.9 mM KCl, 5 mM HEPES pH
7.2). Approximately 100 embryos were placed in a single drop of Ringer’s solution on a
tissue culture plate. A glass cover-slip was placed onto the embryos and smashed. Petri
dish was washed with Ringer’s solution and using a narrow glass pipette, the dissociated
cells were collected into eppendorf tubes. Dissociated cells were pelleted for 5 min at 300
g and resuspended with a narrow bore glass pipette with Ringer’s solution to remove
excess yolk and centrifuged again. Cell pellets were frozen at -80ºC for future use. To
prepare protein for western blotting, cell pellets were resuspended in PLC lysis buffer,
run through needle syringe and centrifuged 13000 rpm to precipitate protein and remove
cellular debris. For immunoprecipitation, protein pellet was resuspended in PLC lysis
buffer and 40 µl of Protein G beads were added with 1 µg of Myc-antibody and PLC

79
lysis buffer was added to a total volume of 1 ml. Mixture was incubated at 4ºC for 2-3
hours with nutation. Beads were pelleted at 4ºC and supernatant was removed. Beads
were washed with PLC lysis buffer and centrifuged and repeated three times. 2x sample
buffer was added to the beads and boiled for 5 minutes. An acrylamide gel was prepared
and boiled samples were loaded in the gel. SDS-PAGE was performed according to
manufacture’s instructions. Separated proteins in the gel were transferred to PVDF
membrane and blotted with anti-flag antibody (Sigma, 1:2500) and detected with goat-
anti-mouse-HRP (BioRad, 1:10000). Protein lysate controls were blotted with anti-myc
antibody (Abcam, 1:5000) and anti-flag antibody and detected with goat-anti-mouse-
HRP. Protein bands were detected by chemilummenescence using ECL plus (GE Health
Care).
3.3.7: in situ hybridization
Embryos for in situ hybridization were fixed overnight in 4% paraformaldehyde in PBS.
Standard whole-mount in situ hybridization protocol was performed using digoxygenin-
labeled anti-sense RNA probes synthesized by in vitro transcription (Roche).
myoD/krox20, RNF41 riboprobes at 0.5-1.0 ng/µl were used and stained embryos were
imaged using Leica dissecting scope and OpenLab software.
3.3.8: Microscopy
Live embryos were mounted in 0.8% low melt agarose before imaging. Fluorescent images
of embryos injected with memRFP, GFP-Pk or transgenics were obtained with a Zeiss
510/710 confocal microscope. Images were captured along the dorsal-ventral axis through

80
the neuroepithelium to the notocord at the 6-12 somite stage.
3.4: References
[1] L.M. Kallay, A. McNickle, P.J. Brennwald, A.L. Hubbard, and L.T. Braiterman, “Scribble associates with two polarity proteins, Lgl2 and Vangl2, via distinct molecular domains,” Journal of Cellular Biochemistry, vol. 99, Oct. 2006, pp. 647-664.
[2] O. Lee, K.K. Frese, J.S. James, D. Chadda, Z. Chen, R.T. Javier, and K. Cho, “Discs-Large and Strabismus are functionally linked to plasma membrane formation,” Nature Cell Biology, vol. 5, Nov. 2003, pp. 987-993.
[3] K. Iyer, L. Bürkle, D. Auerbach, S. Thaminy, M. Dinkel, K. Engels, and I. Stagljar, “Utilizing the split-ubiquitin membrane yeast two-hybrid system to identify protein-protein interactions of integral membrane proteins,” Science's STKE: Signal Transduction Knowledge Environment, vol. 2005, Mar. 2005, p. pl3.
[4] J.R. Jessen and L. Solnica-Krezel, “Identification and developmental expression pattern of van gogh-like 1, a second zebrafish strabismus homologue,” Gene Expression Patterns: GEP, vol. 4, May. 2004, pp. 339-344.
[5] J.L. Green, S.G. Kuntz, and P.W. Sternberg, “Ror receptor tyrosine kinases: orphans no more,” Trends in Cell Biology, vol. 18, Nov. 2008, pp. 536-544.
[6] T.J. Van Raay, R.J. Coffey, and L. Solnica-Krezel, “Zebrafish Naked1 and Naked2 antagonize both canonical and non-canonical Wnt signaling,” Developmental Biology, vol. 309, Sep. 2007, pp. 151-168.
[7] D.C. Myers, D.S. Sepich, and L. Solnica-Krezel, “Bmp activity gradient regulates convergent extension during zebrafish gastrulation,” Developmental Biology, vol. 243, Mar. 2002, pp. 81-98.
[8] J. Wu and M. Mlodzik, “A quest for the mechanism regulating global planar cell polarity of tissues,” Trends in Cell Biology, vol. 19, Jul. 2009, pp. 295-305.
[9] A. Jenny, R.S. Darken, P.A. Wilson, and M. Mlodzik, “Prickle and Strabismus form a functional complex to generate a correct axis during planar cell polarity signaling,” The EMBO Journal, vol. 22, Sep. 2003, pp. 4409-4420.
[10] X. Wu, L. Yen, L. Irwin, C. Sweeney, and K.L. Carraway, “Stabilization of the E3 ubiquitin ligase Nrdp1 by the deubiquitinating enzyme USP8,” Molecular and Cellular Biology, vol. 24, Sep. 2004, pp. 7748-7757.
[11] M. Narimatsu, R. Bose, M. Pye, L. Zhang, B. Miller, P. Ching, R. Sakuma, V. Luga, L. Roncari, L. Attisano, and J.L. Wrana, “Regulation of planar cell polarity by Smurf ubiquitin ligases,” Cell, vol. 137, Apr. 2009, pp. 295-307.
[12] J.M. Gerdes, Y. Liu, N.A. Zaghloul, C.C. Leitch, S.S. Lawson, M. Kato, P.A. Beachy, P.L. Beales, G.N. DeMartino, S. Fisher, J.L. Badano, and N. Katsanis, “Disruption of the basal body compromises proteasomal function and perturbs intracellular Wnt response,” Nature Genetics, vol. 39, Nov. 2007, pp. 1350-1360.

81
CHAPTER 4: Future Directions
4.0: Preliminary results suggest Wnt4a provides positional information to neural
progenitor cells.
Non-canonical Wnt molecules have been shown in vertebrates to regulate planar
cell polarity (PCP). There is some evidence to suggest that Wnts act redundantly;
however, the specific role of each non-canonical Wnt has not been fully characterized.
There is no known mutant for wnt4a and knock-down by morpholino oligonucleotides
(MO) does not show a phenotype. However, Wnt4a has been implicated in PCP
signalling since further knock-down of wnt4a in wnt5b/11 double mutants generates a
more severe phenotype. In addition, Prickle (Pk) tagged with green fluorescent protein
(GFP), a marker of polarity is lost compared to the double mutant. To examine the
specific role of Wnt4a in regulating PCP, a local source of Wnt4a was generated in the
developing neural tube and the affect of Wnt4a on neighbouring neural progenitor cells
(NPCs) was examined. Although the initial objective was to determine differences in
PCP using GFP-Pk localization, technical difficulties with the heterochronic cell
transplantation procedure limited the depth of analysis. However, an interesting
observation did arise from a subset of the experiments. Generation of a local Wnt4a
source appeared to alter the behaviour of NPCs along the anterior-posterior axis. Based
on in situ hybridization data, wnt4a is normally expressed in a decreasing gradient along
the anterior-posterior axis, and I propose that this gradient provides positional
information to NPCs during neurulation. When this gradient is disrupted, differences in
NPC cell behaviour will be observed. When NPCs are exposed to a ‘reversed’ gradient,
an increasing anterior-posterior gradient, NPCs appear to not cross the midline. It is

82
unclear what the mechanism is as it could possibly result from changes in the direction of
intercalation or direction of cell division; however, this phenotype has been observed by
two different methods. To begin to understand how alterations in Wnt4a affect NPC
behaviour, real-time in vivo imaging needs to be performed to address how NPC
division/movement is affected when exposed to different Wnt4a gradients. Although
loss-of-function of wnt4a does not result in a morphological phenotype, there may be a
cell movement phenotype. My preliminary results suggest that loss of Wnt4a may alter
the direction of protrusions, where loss of wnt4a results in daughter NPCs to be deposited
randomly but equally on both sides, which would not result in a phenotype. Potentially,
by observing NPC movement in a wnt4a morphant, a cellular phenotype may be
observed.
First to address the assumption that Wnt4a is not being secreted in all directions
from the Wnt clone, a local source of Wnt needs to be generated expressing GFP tagged
Wnt and the secretion pattern examined. To assay whether NPCs anterior/posterior to the
Wnt clone are able to sense the Wnt signal, GFP-Pk or GFP-Dsh localization can be used.
Enriched fluorescence at the membrane will indicate NPCs are able to respond to Wnt4a.
Second, to specifically characterize the affect of Wnt4a on NPCs, individual cell tracking
needs to be performed. After generation of a local Wnt4a source in an embryo with single
cells labelled (homochronic cell transplants/scatter labeling with a membrane marker),
individual cells can be observed for ability to divide, the direction of cell division and
direction of intercalation after cell division. By capturing the behaviour of NPCs in real-
time, the affect of Wnt4a on NPC behaviour can be determined.

83
4.1: Addressing the instructive/permissive role of non-canonical Wnt
The instructive/permissive role of non-canonical Wnts in regulating PCP is still
an outstanding question that needs to be addressed. The heterochronic cell transplant
procedure requires more optimization to increase the overall success rate of the
technique. For example, to improve the GFP-Pk scatter labeling, transplantation of GFP-
Pk labeled cells into the presumptive neural tube may generate more even labeling along
the anterior-posterior axis. Possibly performing transplants into hosts at earlier/later
stages will improve the likelihood of finding the donor cells in the transplant location
during confocal examination. Despite its shortcomings, heterochronic cell transplantation
remains one method to generate a local Wnt source in the developing neural tube.
Another method to generate one Wnt clone with spatial temporal control is with the use
of transgenics. Transient Wnt transgenics were used in this study (Figure 12), where
embryos were heat shocked to generate Wnt clones. However the number of Wnt clones
was variable since the transgene was not stably expressed and global heat shock was
performed. To generate a single Wnt clone in the developing neural tube, stable
transgenic lines with ubiquitous transgene expression in the neural tube are required.
Spatial and temporal control can be achieved by heat shocking a local area at a chosen
time point, driving expression of the transgene consisting of a heat-shock promoter
inducing Wnt and GFP expression. A similar technique has previously been used to
activate transgene expression in the zebrafish eye and somites and preliminary work has
been successful at adapting this procedure for analysis in the neural tube [1]. To heat-
shock a small region in the neural tube, a modified soldering iron has been constructed
and attached to a power supply where the voltage can be adjusted. Changes in voltage

84
correspond to different solder iron tip temperatures, where 28 V results in solder iron tip
temperature of 60°C measured in air (Figure 23). To optimize the local heat-shock, a
strong GFP (FGFR-GFP) line was used, where embryos were heat-shocked and GFP
expression was scored 4-6 hours after induction. Heat-shock between 3-5 minutes was
able to induce varying degrees of GFP expression. Examination of GFP fluorescence
showed a range of expression where individual cells to small clusters were induced
(Figure 24). The local heat-shock method allows targeting at a specific time and place for
a Wnt clone to be produced. In comparison to the heterochronic cell transplantation
procedure, the local heat-shock method can generate more replicates with better accuracy
in a less time-consuming manner. The only drawback is the generation of stable
transgenic lines and screening for founders with ubquitious transgene expression. Once
the stable transgenic lines have been established, transgenic embryos can be scatter-
labelled with GFP-Pk. At early somite stages, these embryos can be locally heat-shocked
and screened 4-6 hours later for GFP expression under a dissecting scope. Embryos
positive for a Wnt clone can be analyzed by confocal microscopy for differences in GFP-
Pk localization with respect to the Wnt clone. Hopefully by using this approach, the
instructive/permissive role of non-canonical Wnts can be found.
4.2: The role of RNF41 in regulating planar cell polarity
Preliminary evidence suggests that RNF41 regulates PCP; however, the specific
mechanism is unclear. RNF41 is an E3 ubiquitin ligase that physically interacts with
Vangl2, which suggests that Vangl2 may be a substrate for RNF41. To determine if
RNF41 directly regulates Vangl2, co-expression studies with GFP tagged Vangl2 and

85
Figure 23: Temperature Voltage Standard Curve By adjusting the voltage supply to the modified soldering iron, the temperature of the tip proportionally changes
A Aʼ
Figure 23: Local heat shock with a modified soldering iron is able to induce transgene expression in cells in the neural tube (A) Transgenic embryo 4 hours post-heat-shock, dorsal view. (A’) Enlargement of heat-shock area showing individual cells expressing transgene

86
RNF41 can be performed. It would be expected if RNF41 regulates Vangl2, membrane
fluorescence of Vangl2-GFP would decrease with co-expression of RNF41 in a
concentration dependent manner. To further suggest that whether loss of Vangl-2 is a
result of ubiquitin-dependent degradation, embryos can be treated with proteasome
blockers, which should block the loss of Vangl2 membrane fluorescence. To show that
Vangl-2 is ubiquitinated, immunoblots for ubiquitinated Vangl2 can be performed. To
test and screen for substrates of RNF41, a mass-spectrometry approach can be taken,
where cells over-expressing RNF41 and treated with proteasome blockers can be
analyzed for ubiquitinated peptides. As a positive control, the known targets (ErbB3,
Parkin, RNF41) should be detected. Potentially either Vangl2 or Pk will be detected,
indicating the substrate of RNF41. Finally, In zebrafish, knock-down of gene function is
easily accomplished by injection of MO. To further characterize the role of RNF41 in
regulating PCP, loss-of-function studies can be performed.
4.3: The cue that establishes planar cell polarity remains elusive
The global cue that establishes PCP remains elusive; however, non-canonical Wnt
remain a likely candidate. To investigate the role of non-canonical Wnts in regulating
PCP, the various techniques to generate a local Wnt source in the neural tube can be
further developed and eventually applied. If non-canonical Wnts do not play instructive
roles in regulating PCP, there are many other possible leads generated by the membrane-
yeast-two-hybrid screen, which identified 95 protein-protein interactions with Vangl1/2.
The MYTH screen will not directly determine the global PCP cue; however, other
membrane proteins that may act as co-receptors may be found or other signalling

87
pathways that interact with Vangl1/2 can be tested where the upstream components of the
interacting protein might included a ligand that may regulate PCP as well.
From the screen, there are some interesting hits that should be validated by
immunoprecipitation and further characterized. Receptor-tyrosine kinase-like orphan
receptor 1 (ROR1) is a functional homologue to ROR2, which has been shown in
mammalian cell culture to bind Wnt5a. ROR1/2 contain an intracellular tyrosine kinase
domain, which may possibly phosphorylate Vangl2 or other PCP components. Another
possibility is that Bone morphogenetic protein (BMP) may act as a global cue since
BMP receptor type II was found to interact with Vangl2. BMP has been previously
implicated in regulating CE during zebrafish gastrulation [41]. During gastrulation, BMP
is expressed along the dorsal-ventral axis in a gradient with highest Bmp expression on
the ventral side. Examining cell movements in regions of different Bmp concentration,
different cell movement behaviours were observed [41]. Bmp appears to negatively
regulate non-canonical wnt signalling (wnt 5b/11), which controls CE during gastrulation
[41]. Potentially, the Bmp gradient found across the dorsal-ventral axis may indirectly
regulate PCP signalling by regulating non-canonical Wnt signalling which in turn
maintains PCP. Recent evidence indicates that Bmp signalling also directly regulates
PCP signalling [42]. Smurf1/2, an E3 ubiquitin ligase and regulator of Bmp signalling
has been shown to regulate Pk stability in complex with Dsh and Par6 [42]. Asymmetric
localization of PCP components appears to be generated by targeted degradation. In lieu
of the RNF41 data, other proteins from the MYTH screen suggest that Vangl1/2 is
potentially ubiquitinated. Membrane proteins that are ubiquitinated are normally
endocytosed and delievered to the lysosome for degradation. Proteins such as Sorting

88
Nexin 9, Low-density-lipoprotein Receptor Related Protein 3 and lysosomal-associated
protein transmembrane 4α were all found to interact with Vangl1 suggesting
ubiquitination as a means to generate asymmetric localization of Vangl1/2. In summary,
there are many possible candidates that could act to establish PCP. Hopefully, with the
preliminary findings described herein, a contribution has been made to aid in the search.
4.4: References
[1] M.E. Hardy, L.V. Ross, and C. Chien, “Focal gene misexpression in zebrafish embryos induced by local heat shock using a modified soldering iron,” Developmental Dynamics: An Official Publication of the American Association of Anatomists, vol. 236, Nov. 2007, pp. 3071-3076.
[2] J.R. Jessen, J. Topczewski, S. Bingham, D.S. Sepich, F. Marlow, A. Chandrasekhar, and L. Solnica-Krezel, “Zebrafish trilobite identifies new roles for Strabismus in gastrulation and neuronal movements,” Nature Cell Biology, vol. 4, Aug. 2002, pp. 610-615.
[3] B. Ciruna, A. Jenny, D. Lee, M. Mlodzik, and A.F. Schier, “Planar cell polarity signalling couples cell division and morphogenesis during neurulation,” Nature, vol. 439, Jan. 2006, pp. 220-224.










![Enhanced Kat3A/Catenin transcription: a common mechanism of … · 2019. 9. 28. · mediated transcription) or non-canonical (planar cell polarity, Ca2+/PKC activation)[42,43]. Canonical](https://img.dokumen.tips/doc/110x75/60c2e324f6a8a620c25ac30f/enhanced-kat3acatenin-transcription-a-common-mechanism-of-2019-9-28-mediated.jpg)



![Retromer Controls Planar Polarity Protein Levels and ......they form intercellular complexes that link neigh-boring cells [1–3]. Specifically, the seven-pass transmembrane protein](https://img.dokumen.tips/doc/110x75/60da5598397da01d004943d0/retromer-controls-planar-polarity-protein-levels-and-they-form-intercellular.jpg)














