Embed Size (px)
Citation preview

Photosynthesis Research 22: 101-113, 1989. © 1989 Kluwer Academic Publishers. Printed in the Netherlands.
Regular paper
101
Regeneration of the high-affinity manganese-binding site in the reaction center of an oxygen-evolution deficient mutant of Scenedesmus by protease action
Christopher Preston and Michael Seibert Photoconversion Research Branch, Solar Energy Research Institute,* Golden, CO 80401, U.S.A.
Received 16 February 1989; accepted in revised form 4 May 1989
Key words: manganese, oxygen evolution, D1 protein, protease, processing, Scenedesmus obliquus mutant (LF-I)
Abstract
The O2-evolution deficient mutant (LF-1) of Scenedesmus obliquus inserts an unprocessed D1 protein into the thylakoid membrane and binds less than half the wild type (WT) level of Mn. LF-1 photosystem II (PS II) membrane fragments lack that part of the high-affinity Mn 2÷ -binding site found in WT membranes which may be associated with histidine residues on the D1 protein (Seibert et al. 1989 Biochim Biophys Acta 974: 185-191). Hsu et al. (1987 Biochim Biophys Acta 890: 89-96) purport that the high-affinity site (characterized by competitive inhibition of DPC-supported DCIP photoreduction by/ IM concentrations of Mn 2+ ) in Mn-extracted PS II membranes is also the binding site for Mn functional in 02 evolution. Proteases (papain, subtilisin, and carboxypeptidase A) can be used to regenerate the high-affinity Mn z+ -binding site in LF-1 PS II membranes but not in thylakoids. Experiments with the histidine modifier, DEPC, suggest that the regenerated high-affinity Mn2÷-binding sites produced by either subtilisin or carboxypeptidase A treatments were the same sites observed in WT membranes. However, none of the protease treatments produced LF-1 PS II membranes that could be photoactivated. Reassessment of the processing studies of Taylor et al. (1988 FEBS Lett 237: 229-233) lead us to believe that their procedure also does not result in substantial photoactivation of LF-1 PS II membranes. We conclude that (1) the unprocessed carboxyl end of the DI protein in LF-I is located on the lumenal side of the PS II membrane, (2) the unprocessed fragment physically obstructs or perturbs that part of the high-affinity Mn 2÷ -binding site undetectable in LF-1, and (3) the D1 protein must be processed at the time of insertion into the membrane for normal O2-evolution function to result.
Abbreviations; Chl - chlorophyll, DCBQ - 2,6-dichloro-l,4-benzoquinone, DCIP - 2,6-dichlorophenol indophenol, DEPC - diethylpryocarbonate, DPC - 1,5-diphenylcarbazide, HEPES - 4-(2-hydroxyethyl)-l- piperazine-ethanesulfonic acid, LDS-PAGE - lithium dodecylsulfate polyacrylamide gel electrophoresis, LF-1 - a low-fluorescent mutant of Scenedesmus obliquus, MES - 4-morpholineethanesulfonic acid, PS II - photosystem II, PMSF - phenylmethylsulfonyl fluoride, RC - photosystem II reaction center, Tris - tris(hydroxymethyl)aminomethane, WT - wild type
* Operated by the Midwest Research Institute for the U.S. Department of Energy under contract DE-AC-02-83CH10093.
Introduction
In oxygenic photosynthesis, primary charge separation occurs with a z of about 3 ps (Wa-

102
sielewski et al. 1989) in isolated photosystem II (PS II) reaction center (RC) complex, composed of the D 1, D2 and cytochrome b-559 proteins (Nanba and Satoh 1987). Water-splitting function coupled to charge separation involves the sequential extrac- tion of four electrons from water, releasing 02 as a byproduct in thylakoid membranes (Kok et al. 1970). The so called 02-evolving complex is com- posed of several proteins and binds four Mn at its catalytic center (Amesz 1983, Dismukes 1986). Early evidence suggested that up to two of these Mn were bound to a 33-kDa extrinsic protein (Abramowicz and Dismukes 1984, Yamamoto et al. 1984). However, it was also shown that this protein could be removed without release of Mn (Ono and Inoue 1983, Miyao and Murata 1984) suggesting that all four Mn were bound to intrinsic proteins. Recently, several groups have suggested that the D1 protein may provide at least some ligands for binding functional Mn (Metz et al. 1986, Ikeuchi et al. 1988, Seibert et al. 1988, Tamura et al. 1989). All of these studies indicate that the reaction centre and the O2-evolving complex may consist of some of the same proteins.
In 1980, Metz and Bishop (1980) and Metz et al. (1980) reported a low fluorescent mutant (LF-1) of the green alga Scenedesmus obliguus that had an inactive O2-evolving apparatus and bound 40% of the Mn compared to wild type (WT). Moreover, LF-1 was undamaged on the reducing side of PS II, contained a functional reaction center, and display- ed active secondary donor function (Metz et al. 1985, Rutherford et al. 1988). Several groups have suggested that the mutant inserts a D 1 protein with an unprocessed carboxyl end (Metz et al. 1986, Diner et al. 1988, Taylor et al. 1988a). Apparently the unprocessed D1 protein renders the mutant unable to bind at least some functional Mn and hence inhibits its ability to evolve 02.
Recently Hsu et al. (1987) developed an essay for detecting a high-affinity, Mn 2- -binding site in PS II membranes which had been pre-treated with tris(hydroxymethyl)aminomethane (Tris)-HC1 to remove functional Mn. This assay depends on the ability of Mn 2+ , at micromolar concentrations, to inhibit light-induced electron donation from diph- enylcarbazide (DPC) to PS II competitively. Inhibi- tion occurs since Mn 2+ is not as good an electron donor as DPC. Substantial evidence indicates that the high-affinity site is the binding site for function-
al Mn (Hsu et al. 1987, Seibert et al. 1989, Tamura et al. 1989). Based on the above, Seibert et al. (1989) used the assay to investigate the M n 2+ -bind- ing site in PS II-enriched membranes from the LF- 1 mutant of Scenedesmus. They found that one com- ponent of the site, observed in WT PS II mem- branes, was missing in the mutant; that the missing component is associated with the D 1 protein; and that (a) histidine residue(s) also associated with this component provides ligand(s) for binding Mn. Fur- thermore the full complement of the high-affinity Mn 2 + -binding site was detected on isolated spinach D1, D2, cytochrome b-559 RC complex (Seibert et al. 1989). In the present study, we have treated PS II membranes from the LF-1 mutant with several proteases to cleave the unprocessed D I protein. Such treatments regenerate the high-affinity Mn 2÷ - binding site in this material as detected by the Hsu et al. (1987) assay.
Materials and methods
Materials
Scenedesmus obliquus WT and LF-1 mutant cells were grown heterotrophically in 200-ml batches as described in Bishop (1971). Cells were harvested and broken, and PS II-enriched membranes were prepared by the method of Metz and Seibert (1984). PS II membranes were stored at - 80°C in 20mM 4-morpholineethanesulfonic acid (MES)- NaOH (pH 6.5), 400mM sucrose, 20mM NaCI, and 5 mM MgC12 (Buffer A) until use.
Papain (E.C. 3.4.22.2) Type IV from papaya latex, subtilisin (E.C. 3.4.21.14) from Bacillus sub- tilis, and carboxypeptidase A (E.C. 3.4.17.1) Type II from bovine pancreas were obtained from Sigma (St. Louis, MO). Papain was activated by incubat- ing the enzyme at 1 mg protein, ml ~ for 30 rain on ice in 50mM Tris-HC1 (pH 7.5), 100mM NaC1, and 165#g.ml -~ of DL-cysteine. Carboxypep- tidase A and subtilisin were used as supplied.
Protease treatment of PS H membranes
PS II membranes at 200 #g chlorophyll (Chl). ml were incubated with papain for 20 rain at 20°C in the dark. The incubation buffer consisted of 40 mM

103
4-(2-hydroxyethyl)- 1 -piperazine-ethanesulfonic acid (HEPES)-NaOH (pH 6.8), 400mM sucrose, and 10mM NaC1 (Buffer B) with 0.1% w/v dith- iothreitol added. The reaction was stopped by ad- ding 20 volumes of ice cold Buffer B containing 1 mM phenylmethylsulfonyl fluoride (PMSF). The treated membranes were collected by centrifuga- tion (30,000 x g, l0 min) and washed twice in Buf- fer B before final resuspension in Buffer A.
Subtilisin and carboxypeptidase A treatments were performed by incubating PS II membranes (200 #g Chl. ml- 1 ) with enzyme for 30 rain at 20°C in the dark in 20mM MES-NaOH (pH 6.5), 400 mM sucrose, and 20 mM NaCl (Buffer C). The reactions were stopped by adding 20 volumes of ice cold Buffer C containing 1 mM PMSF (for sub- tilisin treatment) or 4mM 1,10-phenanthroline (for carboxypeptidase A treatment). The membranes were collected by centrifugation (30,000 x g, 10 rain) and washed once in Buffer C before resus- pension in Buffer A. Neither the PMSF nor 1,10- phenanthroline washes alone caused any dimin- ution of O2-evolution activity or DCIP photo- reduction with WT PS II membranes.
The protease concentrations used were deter- mined empirically, in such a way that the con- centration chosen resulted in a 50% inhibition of DPC-supported DCIP photoreduction by l0 #M MnC12 with LF-1 PS II membranes.
N H 2 O H treatment
PS II membranes at 500pg Chl-ml -l were in- cubated with 5 mM NH2 OH in Buffer A for 30 min at 4°C in the dark to remove functional Mn from the membrane (Tamara and Cheniae 1987). The membranes were collected by centrifugation (30,000 x g, 10 min) and washed once in Buffer A before resuspension in the same buffer. When both protease and NH2OH treatments were performed on the same preparation, the protease treatment always preceded NHEOH treatment. NH2OH treatment of LF-1 PS II membranes did not alter DPC-supported DCIP reduction, Mn 2+ inhibition of this reaction, or the photoactivation potential of these membranes (see also Seibert et al. 1989). For this reason NH2OH treatment of LF-1 PS II mem- branes was dispensed with.
D E P C treatment
Diethylpyrocarbonate (DEPC) was used to modify available histidine residues (Miles 1977). PS II membranes at 100/~gChl-m1-1 were incubated with 500/~M DEPC (from 100 mM fresh stock dis- solved in ethanol) in Buffer C for 1 h at 20°C in the dark (Tamura et al. 1989). The reaction was stopped by adding 5 volumes of ice cold Buffer C containing 10mM DL-histidine. After centrifuga- tion (30,000 x g, 10min) the membranes were washed once in Buffer C before resuspending in Buffer A. With WT PS II membranes, DEPC treat- ment was carried out subsequent to NHEOH treat- ment. Controls were treated in the same way except that DEPC was not added. Previous workers (Tamura et al. 1989) using the above treatment conditions have demonstrated that DEPC specifi- cally modified histidine residues in the D1 protein that provide ligands for Mn.
Photoactivation
Photoactivation was performed essentially as des- cribed by Tamura and Cheniae (1986, 1987). Briefly PS II membranes devoid of Mn were incubated at 200#gChl.ml -I in 50mM MES-NaOH (pH 6.5), 400mM sucrose, 20mM NaCl, 40mM CaC12, 2 mM MnCI2, and 100/~M DCIP for 45 min under illumination from a bank of cool white fluorescent lamps. The light intensity at the sample surface was 8W-m -2. The samples (2ml) were incubated at 20°C in flat 5-cm diameter petri dishes and shaken gently every 5 minutes. At the end of the illumina- tion period, the samples were centrifuged (30,000 x g, 10min) and washed once in Buffer A before resuspension in the same buffer.
02-evolution assays
O:-evolution activity was measured with a Clark- type 02 electrode. The assay buffer consisted of 50mM MES-NaOH (pH 6.5), 400mM sucrose, and 20 mM NaC1. Four ml of assay buffer contain- ing 6 #g Chl. ml- ~ were illuminated with saturating white light passed through a 2-cm water filter, and 200 #M DCBQ was used as an electron acceptor. PS II extrinsic proteins of 17-, 23- and 33-kDa

104
associated with O2 evolution were isolated from spinach PS II membranes by CaCI2 treatment (Ono and Inoue 1983). The crude protein extract was desalted by dialysis against 20mM MES-NaOH (pH 6.5) for 3h at 4°C. The material was then concentrated to about 2 mg protein-ml-~ on a bed of dry sucrose. For the photoactivation experi- ments, 10#l of the three extrinsic proteins were added to the O2-assay buffer and incubated with the PS II membranes for 1 min prior to illumina- tion.
DCIP photoreduction
Dichlorophenol indophenol (DCIP) photoreduc- tion was performed using an Aminco DW2a spec- trophotometer. The assay buffer was the same as for Oz-evolution measurements; however, 200 #M DPC was used as an electron donor and 30#M DCIP was added as an electron acceptor. Light was passed through a heat filter (Melles Griot 03MHG007) and a 630nm cut-off filter (Schott RG630) and focused on the sample cuvette. The photomultiplier was protected from the illuminat- ing light with a 600-rim narrow bandpass filter (Melles Griot 03FIV045), and DCIP reduction was monitored continuously at 600 nm. Samples con- tained 5 #g Chl. ml- l, the pathlength was I cm, and an extinction coefficient of 13 m M - ~ • cm- ~ (Arm- strong 1964) was used to calculate rates of DCIP reduction.
The DPC/Mn competition assay of Hsu et al. (1987) was used to determine the amount of high- affinity Mn 2+-binding site in the different materials. At the concentrations of DPC, MgC12, and NaC1 used in our studies, l0 #M Mn 2+ causes a 50% reduction in DPC-supported DCIP photoreduction in WT PS II membranes. MnC12 was added to the sample cuvette from a 100#M stock prepared in assay buffer. For each experi- mental point, two measurements were made on different samples and averaged. The two measure- ments were generally within 5% of each other. The data presented are representative of several experi- ments.
LDS-PAGE
Lithium dodecysulfate-polyacrylamide gel electro-
phoresis (LDS-PAGE) of Scenedesmus PS II was performed essentially as described in Metz et al. (1985) using a 22-cm 10-15% gradient acrylamide resolving gel. The long gel was necessary for resolv- ing bands in the 30-36 kDa range. Samples were dissociated by heating at 50°C for 5 min in 50 mM Tris-HCl (pH 7.8), 6% sucrose, 1% dithiothreitol, and 50:1 LDS:Chl. The papain-treated samples were dissociated in 50 mM Tris-HCl (pH 6.8), 10% glycerol, 1% dithiothreitol, 20mM PMSF (to in- hibit residual papain action during sample dissocia- tion), and 50:1 LDS:Chl.
Results
The addition of 10/~M MnC12 to a standard DPC to DCIP assay for PS II activity of Scenedesmus WT PS II membranes decreases the photoreduction rate of DCIP by about 50% under our conditions (Fig. 1A). Properties of the high-affinity Mn 2+- binding site equated with the binding site of fun- ctional Mn were originally determined by this technique (Hsu et al. 1987). Exposure of LF-1 membranes to the same conditions resulted in a smaller (about 25 %) decrease in DCIP photoreduc- tion (Fig. 1B). Treatment of WT PS II membranes with 1:150 (w/w) papain:Chl prior to assay did not alter substantially the properties of the MnC12 in- hibition (Fig. 1A). However, significant differences were observed when LF-1 PS II membranes were treated with papain (Fig. 1B). Not only did papain increase the effect of Mn 2+ on the DPC to DCIP assay but it also affected the concentration depen- dence (i.e., more MnC12 was required for full inhibi- tion). In all of these preparations, the rate of DCIP photoreduction was constant between 10 and 100/~M MnC12. Since papain treatment degrades many proteins in both WT and LF-1 PS II prepara- tions including the D1 protein (Fig. 2), it seems reasonable to suggest that treatment of LF- 1 PS II membranes removes enough of the unprocesses segment of the D1 protein to uncover (or perhaps reconstitute) the missing Mn 2+ -binding site.
Regeneration of the high-affinity Mn 2+ -binding site in LF-1 PS II membranes is significant because it might result in recovery of photoactivation potential. Unfortunately, photoactivation of pa- pain-treated LF-1 PS II membranes was little more successful than for the untreated LF-1 material (Table 1). Photoactivation of NH2OH-treated WT

105
100
80
~- 60 . 2 "5 -o 40
0_ o 20 D
I
A I I I I ~ I
~ B8
• ~ B I I I , , 1 1 ,
[3
0 I I I I I s j l I I I I I.t; i 0 2 4 6 8 1 0 1 0 0 0 2 4 6 8 1 0 1 0 0
MnCI 2 concentrat ion (ym)
• • 2 + " Fig. 1. The influence of papain treatment on the hlgh-aifinlty Mn -binding site in Scendesmus (A) WT and (B) LF-1 PS II membranes. Untreated PS II (o), PS II treated with 1:150 (w/w) papain:Chl (D). Maximum rates of DCIP photoreduction were 124, 118, 135 and ! 32 #moles DCIP mg Chl ~ h- 1 for WT, papain-treated WT, LF- I, and papain-treated LF- 1 PS II membranes, respectively.
PS II reconstituted about 80% of the control level of O2-evolution activity. Papain treatment of WT PS II membranes reduced O2-evolution activity by about 17%, but did not affect photoactivation potential of this material. Addition of CaCI 2 and the extrinsic proteins to photoactivated papain- treated LF-1 PS II membranes did not markedly increase 02 -evolution capacity of this material sug- gesting that loss of C a 2+ and/or of the extrinsic proteins was not the cause of inhibition. Instead Mn appeared not to ligate or more likely ligated incorrectly (see also Seibert et al. 1989) to the mem- brane during the photoactivation process.
While successful in providing access to the miss- ing Mn site(s), papain treatment did not allow photoactivation of LF-1 PS II membranes. Since this could be a consequence of papain cleavage of many proteins (see Fig. 2), other proteases were tried. Treatment of LF-1 PS II membranes with subtilisin at 1:200 (w/w) subtilisin:Chl reduced the apparent molecular weight of the LF-1 D1 protein by about 2.5-3kDa. (Fig. 2). Subtilisin also re- duced the apparent molecular weight of the WT D 1 protein, but only by about 0.5-1 kDa. Subtilisin treatment of LF-1 PS II increased MnC12 inhibition of DPC ~ DCIP activity (Fig. 3B). As with the papain treatment, the concentration dependence of the MnC12 inhibition was different for subtilisin- treated material compared to control membranes. This change was similar to that for the papain-
treated material where higher MnCI2 concentration was required for full inhibition of LF-1 than for the WT PS II membranes. However unlike papain treatment ofWT PS II membranes, subtilisin treat- ment did seem to affect MnC12 inhibition of DPC --, DCIP activity (Fig. 3A). In treated WT material, MnCI2 was not as effective at inhibiting DPC donation, and activity was only reduced by 42% in the presence of 10#M MnCI2. We have no explanation for this result as the WT D1 protein is being cut to about the same length as the LF-1 D1 protein (Fig. 2). Photoactivation of the subtilisin treated LF-1 PS II membranes was not successful (Table 2). Addition of CaCI2 and the extrinsic pro- teins to this material after photoactivation did not increase activity. Photoactivation of WT and sub- tilisin-treated WT PS II membranes was more than 80% successful and subtilisin treatment of WT PS II membranes reduced 02 evolution by 25%. In many ways the non-specific proteases, papain and subtilisin, produced similar results.
The D1 protein of LF-1 is unprocessed at the carboxyl end (Diner et al. 1988; Taylor et al. 1988a) and consequently carboxypeptidase A was used in an attempt to remove the unprocessed fragment. Treatment of LF-1 PS II membranes with 1:20 (w/w) carboxypeptidase A:Chl increased the MnCl2-induced inhibition of the DPC ~ DCIP activity from about 25% to about 50% (Fig. 4B). Thus, treatment with carboxypeptidase A, like

106
Fig. 2. LDS-PAGE of protease-treated PS II membranes. Lanes 1, 5 and 9: WT PS II; lane 2: papain-treated WT PS II; lanes 3, 7, and 1 h LF-1 PS II; lane 4: papain-treated LF-1 PS II; lane 6: subtilisin-treated WT PS II; lane 8: subtilisin-treated LF-1 PS II; lane I0: carboxypeptidase A-treated WT PS II; lane 12: carboxypeptidase A-treated LF-1 PS II; and lane 13: molecular weight standards of 97-, 66-, 43-, 21.5- and 14.4-kDa. The positions of the WT and LF-1 D1 proteins (O) and the 30-kDa (equivalent to 33 kDa in spinach) extrinsic protein ( - ) are indicated.
papa in and subtilisin, is able to uncover the masked M n - b i n d i n g site on the D1 prote in of LF-1. However, in contras t to the papa in and subtil isin t reatments , carboxypept idase A t rea tment did no t alter the concen t ra t ion dependence of the MnCI2
inhib i t ion of D P C ~ D C I P activity. Fur thermore , carboxypept idase A t rea tment of W T PS II mem- branes did no t alter MnC12 inhibi t ion of D P C ~ D C I P activity (Fig. 4A). On the other hand, in both W T and LF-1 PS II membranes ,

107
Table 1. Photoactivation of papain-treated PS II membranes from Scenedesmus. NH2OH treatment and photoactivation procedures are described in the Materials and Methods. The proteins added to the 02 assays were a mixture of the 17-, 23- and 33-kDa extrinsic proteins, and CaCI 2 was added at 2 raM.
Sample and treatment 02 Evolution rate (~mole mg Chl - i h - t )
WT PS II LF-1 PS II
Untreated PS H Control 231 14 NH2OH treated 14 3 Photoactivated 179 12 Photoactivated + proteins, CaCI: 184 15
Papain-treated PS H Control 191 11 NH2OH treated 11 11 Photoactivated 122 17 Photoactivated + proteins, CaCI 2 146 27
100,
80,
60
40
0 20 121
0 1
l o l o o
MnCI 2 concentration (p.m)
Fig. 3. The influence of subtilisin treatment on the high-atfinity Mn :+ -binding site in (A) WT and (B) LF-I PS II membranes. Untreated PS II (o), PS II treated with 1:200 (w/w) subtilisin:Chl (U). Max imum rates of DCIP photoreduction were 111, 113, 123 and 125 #mole DCIP mg C h l - 1 h - ~ for WT, subtilisin-treated WT, LF- 1, and subtilisin-treated LF- 1 PS II membranes, respectively.
Table 2. Photoactivation of subtilisin-treated PS II membranes. N H : O H treatment and photoactivation procedures are described in the Materials and Methods. The proteins added to the O: assays were a mixture of the 17-, 23- and 33-kDa extrinsic proteins, and CaC12 was added at 2 raM.
Sample and treatment O: Evolution rate ~ m o l e mg Chl -I h -~)
WT PS II LF-I PS II
Untreated PS H Control 256 NH2OH treated 9 Photoactivated 210 Photoactivated + proteins, CaCI 2 247
Subtilisin-treated PS H Control 193 NH2OH treated 17 Photoactivated 166 Photoactivated + proteins, CaCI: 187
16
19 26

108
100q
80 (D
c 60 O
-~ 4o
o 20 c~
0 0
I I I A
I I I 2 4 6
i I f f ~ i , i , , [ [ ,
0 0
°n@ o o
I I / , , , I I I I I f [ l 8 10100 0 2 4 6 8 10100
MnCI 2 concentration (#m)
Fig. 4. The influence ofcarboxypeptidase A treatment on the high-affinity Mn 2+ -binding site in (A) WT and (B) LF-1 PS II membranes. Untreated PS II (o), PS II treated with 1:20 (w/w) carboxypeptidase A:Chl (O). Maximum rates of DCIP photoreduction were 129, 167, 172 and 224 #mole DCIP mg Chl-~ h-~ for WT, carboxypeptidase A-treated WT, LF-1, and carboxypeptidase A-treated LF-1 PS II membranes, respectively.
c a r b o x y p e p t i d a s e A t r ea tmen t increased the ra te o f D P C - s u p p o r t e d D C I P reduc t ion in the absence o f M n 2+ by 4 0 - 7 0 % depend ing on the p repa ra t ion . As yet we have no conclusive exp lana t ion for this result. C a r b o x y p e p t i d a s e A t r ea tmen t resul ted in a reduc t ion o f 1 k D a in the a p p a r e n t molecu la r weight o f the LF-1 D1 p ro te in (Fig. 2), thus only pa r t o f the unprocessed segment o f this p ro te in needed to be r emoved to res tore the full high-affin- ity M n 2÷-binding site. Despi te regenera t ion o f a M n - b i n d i n g site which appea r s very s imilar to the W T site, p h o t o a c t i v a t i o n o f the ca rboxypep t ida se A- t r ea t ed LF-1 PS II m e m b r a n e s was also no t
successful (Table 3). F u r t h e r m o r e , ca rboxypep- t idase A - t r e a t m e n t o f W T PS II membranes resul ted in a 50% inhib i t ion o f 02 evolut ion, but despi te this loss in act ivi ty, pho toa c t i va t i on o f this mate r ia l was still possible. This loss o f O2-evolu- t ion act ivi ty o f W T PS II membranes is p r o b a b l y a result o f c a rboxypep t ida se A ac t ion on the 30-kDa (33 k D a in spinach) pro te in which is a t t acked by this p ro tease (Fig. 2).
Using a his t idine chemical modif ier (DEPC) , Seibert et al. (1989) showed tha t the M n 2+ -b inding site missing f rom the LF-1 PS II membranes is assoc ia ted with a his t idine residue(s) on the D1
Table 3. Photoactivation of carboxypeptidase A-treated PS II membranes. NH2OH treatment and photoactivation procedures are described in the Materials and Methods. The proteins added to the O 2 assays were a mixture of the 17-, 23- and 33-kDa extrinsic proteins, and CaC12 was added at 2 mM.
Sample and treatment 02 Evolution rate (~mole mg Chl- ' h ' )
WT PS II LF-1 PS II
Untreated PS H Control 226 NH2OH treated 10 Photoactivated 186 Photoactivated + proteins, CaC12 205
Carboxypeptidase A-treated P S H Control 110 NH2OH treated 4 Photoactivated 97 Photoactivated + proteins, CaC1 z 109
6
8 15

protein (Tamura et al. 1989). We used the same chemical modifier to determine if the high-affinity Mn 2+-binding site we regenerated through protease action was the same as the WT histidine site. Figure 5 shows that the site reconstituted after treatment of LF-1 PS II membranes with carboxy- peptidase A was sensitive to DEPC. Specifically, DEPC treatment reduced MnC12 (10#M) inhibi- tion of DPC --* D C I P activity in protease-treated membranes from about 50% to about 25%, but had no effect on Mn2+-inhibition properties of LF-1 PS II membranes that were not treated with protease. DEPC also reduced MnC12 inhibition of DPC --* DCIP activity in subtilisin-treated LF-1 PS II membranes f rom about 50% to about 25% (Fig. 5B). These results indicate that the recon- stituted high-affinity Mn2+-binding sites in both carboxypeptidase A- and subtilisin-treated LF-1 PS II membranes are probably the same sites present in W T PS II membranes and missing in untreated LF-1 PS II.
When LF-1 thylakoids were treated with either carboxypeptidase A or subtilisin and then frac- tionated into PS II membrane fragments, 10#M MnC% inhibited DPC-suppor ted D C I P photoreduction by only 25%, the same value observed for untreated LF-1 PS II membranes
109
(Table 4). This suggests that both proteases are acting only on the lumenal side of the PS II membrane, and hence the carboxyl terminus of the unprocessed LF-1 D 1 protein is located on this side of the membrane (see also Seibert et al. 1989). The site of subtilisin action is probably after the last membrane spanning segment of the D1 protein, since only 2.5-3 kDa of this protein is removed by subtilisin in LF-1 PS II membranes (Fig. 2).
Taylor et al. (1988b) claim to have achieved photoactivation of LF-1 PS II membranes after processing of the D1 protein using a Triton X-100 extract from WT thyakoids (which presumably contains the natural D1 processing enzyme). This treatment processed the D I protein to the correct size and purportedly gave 83% photoactivation. We repeated this experiment and the results are shown in Table 5 along with those of Taylor et al. (1988b). As can be seen, the O2-evolution activity of control WT samples reported by Taylor et al. (1988b) was only 23% of that reported here. Fur- thermore, the activity reported for photoactivated LF-1 PS II membranes represented only 32% of their WT activity and is in fact lower than the rates we have obtained for comparable material. The level of photoactivation we have obtained for Triton X-100 extract-treated LF-1 PS II mem-
, ~ r , / / , • B i , , , I # , ~ I
100' A
"I II ° ~ ~ .
,- 60 0
' ~ D u 0 0
-o 40
a_
0 20 a
0 I I I I I i / I I 1 I I I l i f t 0 2 4 6 8 10100 0 2 4 6 8 10 100
MnCI 2 concent ra t ion (urn)
Fig. 5. Modification of the regenerated high-affinity Mn2+-binding site on protease-treated LF-I PS II membranes by the histidine chemical modifier DEPC. (A) Carboxypeptidase A-treated membranes and (B) subtilisin-treated membranes. Control LF-1 PS II membranes (o), DEPC-treated membranes (o), protease-treated membranes (D), and membranes treated with both protease and DEPC (11). Maximum rates of DCIP photoreduction in (A) were 149, 121, 218 and 1871tmoleDCIPmgChl -t h -I for control, DEPC-treated, carboxypeptidase A-treated, and carboxypeptidase A-/DEPC-treated LF-I PS II membranes, respectively, and in (B) were 158, 168, 170 and 166/~mole DCIP mg Chl- ~ h- ~ for control, DEPC-treated, subtilisin-treated, and subtilisin-/DEPC-treated LF-1 PS II membranes, respectively.

110
Table 4. The effects of protease treatment of LF-1 PS II membrane fragments and thylakoids on the high-atfinity M n 2+ -binding site. Protease treatments were done as described in Materials and Methods. After treatment, thylakoids were fractionated into PS II membrane fragments for assay. DPC-supported DCIP photoreduction rates* were measured in the presence and absence of 10#M MnCI~.
Sample and treatment Rate (+ IO#M MnC12)
Rate ( - MnC12)
PS II membrane fragments Subtilisin-treated PS II Carboxypeptidase A-treated PS II Subtilisin-treated thylakoids/PS II Carboxypeptidase A-treated thylakoids/PS II
0.72 0.51 0.52 0.76 0.76
* Maximum rates of DCIP photoreduction were 126, 118, 184, 98, and 103 #mole DCIP mg Chl- l h- t for control PS II, subtilisin- treated PS II, carboxypeptidase A-treated PS II, subtilisin-treated thylakoid/PS II, and carboxypeptidase A-treated thylakoid/PS II preparations, respectively.
branes is comparable to that of other protease treatments (Tables 1-3). We therefore conclude that photoactivation occurs only to a very limited extent in LF-1.
Discussion
The O2-evolution deficient LF-1 mutant of Scenedesmus contains an unprocessed fragment on the carboxyl end of the D1 protein (Diner et al. 1988, Taylor et al. 1988a), which appears to inter- fere with Mn-binding to the O2-evolving complex (Metz et al. 1986). Recent studies (Seibert et al. 1989) confirmed in the present work indicate that half of the high-affinity Mn-binding site is 'missing' in LF-I . By missing we mean that the high-affinity Mn 2+-binding properties of the site are altered due to steric hindrance or masking of the site by the unprocessed component of D 1. We have attempted to regenerate the missing site by removing the un- processed segment using a variety of proteases, including (1) papain, a general protease, which has a preference for arginine, histidine and lysine re- sidues (Smyth 1967); (2) subtilisin, another general protease the specificity of which depends on the protein being cleaved (Ottensen and Svendsen 1970); (3) carboxypeptidase A, which sequentially removes the carboxyl residue from proteins (Ambler 1967). All three protease treatments used here appear to restore the missing high-affinity Mn 2+- binding site in LF-1 PS II membranes as assayed by the Hsu et al. (1987) procedure. However, the pa- pain- and subtilisin-regenerated sites exhibited a higher MnC12 concentration requirement (10 pM as
opposed to the 2-5 pM that was needed to obtain maximum inhibition of DPC ~ DCIP activity in WT PS II membranes). Carboxypeptidase A treat- ment ofLF-1 PS II, on the other hand, did not alter the concentration dependence of MnC12 inhibition. The change in concentration dependence of the Mn-binding site in papain- and subtilisin-treated material might be caused by: (1) creation of a new non-specific Mn-binding site after treatment with these proteases, (2) cleavage of the D1 at points other than the carboxyl end or cleavage of other PS II proteins, or (3) obstruction of the Mn-binding site by a small amount of protein even after pro- tease treatment. In light of the chemical modifier experiments, the first possibility seems unlikely; a newly created site would not be expected to show the same sensitivity to DEPC as the original site (see Fig. 5). The second possibility can also be discounted since WT PS II membranes also should be affected in the same way as the mutant. In actuality we see no evidence that papain or sub- tilisin treatment of WT PS II membranes altered the concentration dependence of MnCI2 inhibition of DPC ~ D C I P activity. Consequently, we be- lieve that papain and subtilisin are cutting the D 1 protein in LF-1 PS II membranes, probably at the carboxyl end, but the cut fragment(s) remain bound (perhaps through hydrogen bonding) to the remainder of the protein. This impedes access of Mn 2+ to the high-affinity site and results in an increase of the apparent dissociation constant. A similar effect is not seen with carboxypeptidase A-treated LF-1 PS II because this enzyme removes the carboxyl end of the D1 protein one residue at a time.

111
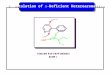
Despite our abil i ty to regenerate the high-affinity Mn2+-b ind ing site in LF-1 PS II membranes , we were unab le to photoact ivate any of the protease- treated LF-1 PS II preparat ions . This could be explained if, even after protease t reatment , one of the M n - b i n d i n g sites is still no t in the correct con- fo rmat ion to allow photoact iva t ion . This possibil- ity is i l lustrated conceptual ly in Fig. 6, where the W T PS II M n - b i n d i n g sites are in a well-structured conformat ion . F o u r M n - b i n d i n g sites have been shown since 4 M n are p robab ly required for 02- evolut ion (Amesz 1983, Dismukes 1986). In LF- I PS II, the unprocessed piece of prote in obstructs half of the M n - b i n d i n g site(s), bu t in addi t ion, the sites themselves may have been displaced from their correct posi t ions due to improper inser t ion of the D1 protein. Removal of the extra unprocessed f ragment by carboxypept idase A t rea tment or cut t ing it with subti l is in or papa in exposes the missing sites, bu t does no t re turn the sites to the correct conformat ion . Taylor et al. (1988b) reported 83% photoac t iva t ion of LF-1 PS II after t rea tment of LF-1 PS II membranes with a Tr i ton X-100 extract of W T thylakoids. This extract p resumably conta ins the na tu ra l Dl -process ing enzyme now k n o w n to cleave precursor D I between Ala-344 and Ala-345 (Takahashi et al. 1988). As we have shown in Table 5, the data of Taylor et al. (1988b) does no t indicate significant photoact iva t ion . In fact the a m o u n t of photoac- t ivat ion achieved after this t rea tment is comparable to the other protease t reatments we have used. We therefore conclude that the D1 prote in has to be processed to its correct length dur ing fo rmat ion of the PS II complex in order for the O2-evolving appara tus to be const ructed properly. This may
WT LF-1
CPASE LF-1 SUB LF-1
Fig. 6, A schematic model showing a conceptual organization of the Mn-binding sites in WT, LF-1, and protease-treated LF-1 PS II membranes. The circles represent Mn-binding sites and the wavy lines are stretches of protein, either D 1 or D2, which may provide ligands for binding Mn. Closed circles represent Mn sites altered in LF-1, Although we have evidence for only two types of high-affinity Mn2+-binding sites of approximately equal magnitude, we show four Mn sites because WT mem- branes contain four Mn per PS II reaction center. In the LF-1 PS II, not only are two of the sites obstructed, but the sites are oriented incorrectly due to improper insertion of DI into the membrane. Treatment of LF-1 PS II with proteases, either carbo×ypeptidase A (CPase, which sequentially removed car- boxyl-terminal amino acids) or subtilisin (Sub, which cleaves the carboxyl end of the protein), exposes the Mn-binding sites but does not return the sites to the proper conformation for 0 2 evolution. In the case of subtilisin-treatment, the excised car- boxyl end of D1 is shown as hydrogen-bonded onto PS II proteins to explain altered Mn 2+ -donation properties discussed in the Results section.
result f rom the fact that D1 protein inser t ion and assembly of PS II react ion center occurs in the s tromal membranes prior to t rans loca t ion to the
Table 5. 02-evolution activity of Scenedesmus PS II membranes before and after photoactivation. The Triton X-100 extraction of WT thylakoids and the treatment of LF-1 PS II membranes with this extract were done as described in Taylor et al. (1988b). LF-1 PS II membranes were treated with the Triton X-100 extract for 1 h. Photoactivation was done as described in the Materials and Methods. The data obtained here are compared to those reported by Taylor et al. (1988b). A mixture of the three extrinsic proteins (17, 23, and 33 kDa) and 2mM CaCI 2 were added to all assays.
PS II membranes 02 evolution rate (#mole mg Chl- ~ h - t )
This study Taylor et al. (1988b)
WT 256 60 Photoactivated WT 247 31 LF-I 14 0 Photoactivated LF- 1 15 0 LF-1 treated with Triton X-100 extract 10 - LF-1 treated with Triton X-100 extract then photoactivated 34 19

112
grana (Mat too and Edelman 1987). All o f our 'processing' experiments have, o f course, been done on appressed membrane fragments.
We are now able to place the unprocessed D1 fragment firmly on the lumenal side o f the LF- I membrane (see Table 4) and not on the stromal side (eliminating the stromal side D 1 processing model o f Diner et al. 1988). However , the assignment o f Mn-binding sites to part icular proteins is still ten- tative. T a m u r a et al. (1989) have indicated that histidine residue(s) on the D1 protein provide ligand(s) for functional Mn. Seibert et al. (1989) and the current work suggest that only half o f the detectable high-affinity Mn2+-binding site, that associated with histidine ligands, is located on this protein. None o f these studies rule out binding sites involving histidine residues on the D2 protein; however, we feel that this would be the less likely si tuation since carboxypept idase A only regenera- tes the high-affinity Mn 2+ -binding site associated with D1. Symmetry considerat ions within the reaction center might suggest that the remaining Mn-binding site(s) present in untreated LF-1 is located on the D2 protein. However , Virgin et al. (1988) have seen a parallel loss o f Mn and the D1 protein during extended photoinhibi t ion o f thyla- koid membranes where no loss o f the D2 protein occurred. This suggests that mos t if not all the functional Mn have at least one ligand associated with the D1 protein.
Apar t f rom the histidine residues ment ioned earlier, other amino acids, including lysine, the carboxyl residues, glutamate and aspartate, and their amines, glutamine and asparagine, may provide ligands for binding the Mn (Dismukes 1988). Future use o f the LF-1 mutan t o f S c e n e d e s -
m u s could assist in elucidating the location o f other residues that bind Mn based on our ability to restore the occluded half o f the high-affinity Mn 2+- binding site.
Acknowledgements
This work was supported by the Division o f Energy Biosciences, Office o f Basic Energy Sciences, U.S. Depar tment o f Energy Cont rac t No. 18-0006-89 (M.S.). The authors would like to thank Dr. N. T a m u r a for st imulating discussions and Dr. G. M.
Cheniae for helpful suggestions during the the in- ception o f this work. M.S. would like to use this oppor tun i ty to express sincere appreciat ion to his Ph.D. advisor Dr. D o n DeVaul t w h o m this issue honors for patience, guidance, and wisdom during four formative and enjoyable years.
References
Abramowicz DA and Dismukes GC (1984) Manganese proteins isolated from spinach thylakoid membranes and their role in 02 evolution. II. A binuclear manganese-containing 34 kil- odalton protein, a probable component of the water dehyd- rogenase enzyme. Biochim Biophys Acta 765:318-328
Ambler RP (1967) Enzymic hydrolysis with carboxypeptidases. Methods Enzymol 11:155-166
Amesz J (1983) The role of manganese in photosynthetic oxygen evolution. Biochim Biophys Acta 726:1-12
Armstrong JMcD (1964) The molar extinction coefficient of 2,6-dichlorophenol indophenol. Biochim Biophys Acta 86: 194-197
Bishop NI (1971) Preparation and properties of mutants: Scenedesmus. Methods Enzymol 23:130-143
Dismukes GC (1986) The metal centers of the photosynthetic oxygen-evolving complex. Photochem Photobiol 43:99-115
Dismukes GC (1988) The spectroscopically derived structure of the manganese site for photosynthetic water oxidatization and a proposal for the protein-binding sites for calcium and manganese. Chemica Scripta 28A: 99-104
Diner BA, Ries DF, Cohen BN and Metz JG (1988) COOH- terminal processing of polypeptide DI of the photosystem II reaction center of Scenedesmus obliquus is necessary for the assembly of the oxygen-evolving complex. J Biol Chem 263: 8972-8980
Hsu B-D, Lee J-Y and Pan R-L (1987) The high-affinity binding site for manganese on the oxidizing side of photosystem II. Biochim Biophys Acta 890:89-96
Ikeuchi M, Koike H and Inoue Y (1988) Iodonation of D1 (herbicide binding protein) is coupled with photooxidation of 1251- associated with CI- -binding site in photosystem-II wat- er-oxidation system. Biochim Biophys Acta 932:160-169
Kok B, Forbusch B and McGloin M (1970) Cooperation of charges in photosynthetic 02 evolution-- I. A linear four step mechanism. Photochem Photobiol 11:457475
Mattoo AK and Edelman M (1987) Intramembrane transloca- tion and posttranslational palmitoylation of the chloroplast 32-kDa herbicide-binding protein. Proc Natl Acad Sci USA 84: 1497-150l
Metz JG and Bishop NI (1980) Identification of a chloroplast membrane polypeptide associated with the oxidizing side of photosystem II by the use of select low-fluorescent mutants of Scenedesrnus. Biochem Biophys Res Commun. 94:560-566
Metz JG and Seibert M (1984) Presence in photosystem II core complexes of a 34-kilodalton polypeptide required for water photolysis. Plant Physiol 76:829-832

Metz JG, Wong J and Bishop NI (1980) Chances in electro- phoretic mobility of a chloroplast membrane polypeptide associated with loss of the oxidizing side of photosystem II in low fluorescent mutants of Scenedesmus. FEBS Lett 114: 61-66
Metz JG, Bricker TM and Seibert M (1985) The azido [~4C] atrazine photoaffinity technique labels a 34-kDa protein in Scenedesmus which functions on the oxidizing side of photosystem II. FEBS Lett 185:191-196
Metz JG, Pakrasi HB, Seibert M and Arntzen CJ (1986) Ev- idence for dual function of the herbicide-binding D I protein in photosystem II. FEBS Lett 205:269-274
Miles EW (I 977) Modification of histidyl residues in proteins by diethylpyrocarbonate. Methods Enzymol 47:431-442
Miyao M and Murata N (1984) Effect of urea on photosystem II particles. Evidence for an essential role of the 33 kilodalton polypeptide in photosynthetic oxygen evolution. Biochim Biophys Acta 765:253-257
Nanba O and Satoh K (1987) Isolation of a photosystem II reaction center consisting of D-1 and D-2 polypeptides and cytochrome b-559. Proc Natl Acad Sci USA 84:109-112
Ono T-A and Inoue Y (1983) Mn-preserving extraction of 33-, 24- and 16-kDa proteins from O2-evolving PS II particles by divalent salt-washing. FEBS Lett 164:255-260
Ottensen M and Svendsen I (1970) The subtilisins. Methods Enzymol 19:199-215
Rutherford AW, Seibert M and Metz JG (1988) Characterisa- tion of the low-fluorescent (LF-1) mutant of Scenedesmus by EPR. Biochim Biophys Acta 932:171-176
Seibert M, Cotton TM and Metz JG (1988) Surface-enhanced Raman scattering spectroscopy: probing the lumenal surface of photosystem II membranes for evidence of manganese. Biochirn Biophys Acta 934:235-246
Seibert M, Tamura N and Inoue Y (1989) Lack of photoactiva- tion capacity in Scenedesmus obliquus LF-1 results from loss of half the high-affinity manganese-binding site. Relationship to the unprocessed D1 protein. Biochim Biophys Acta 974: 185-191
113
Smyth DG (1967) Techniques in enzymic hydrolysis. Methods Enzymol 11:214-231
Takahashi M, Shiraishi T and Asada K (1988) COOH-terminal residues of D 1 and the 44 kDa CPa-2 at spinach photosystem II core complex. FEBS Lett 240:6-8
Tamura N and Cheniae GM (1986) Requirements for the photoligation of Mn 2+ in PS II membranes and the expression of water-oxidizing activity of the polynuclear Mn-catalyst. FEBS Lett 200:231-236
Tamura N and Cheniae GM (1987) Photoactivation of the water-oxidizing complex in photosystem II membranes de- pleted of Mn and extrinsic proteins. I. Biochemical and kinet- ic characterization. Biochim Biophys Acta 890:179-194
Tamura N, Ikeuchi M and Inoue Y (1989) Assignment of his- tidine residues in DI protein as possible ligands for functional manganese in photosynthetic water-oxidizing complex. Biochim Biophys Acta 973:281-289
Taylor MA, Nixon PJ, Todd CM, Barber J and Bowyer JR (1988a) Characterisation of the D I protein in a photosystem II mutant (LF-1) of Scenedesmus obliquus blocked on the oxidising side. Evidence supporting non-processing of D 1 as the cause of the lesion. FEBS Lett 235:109-116
Taylor MA, Packer JCL and Bowyer JR (1988b) Processing of the D1 polypeptide of the photosystem II reaction centre and photoactivation of a low fluorescence mutant (LF-I) of Scenedesmus obliquus. FEBS Lett 237:229-233
Virgin I, Styring S and Andersson B (1988) Photosystem II disorganization and manganese release after photoinhibition of isolated spinach thylakoid membranes. FEBS Lett 233: 408-412
Wasielewski MR, Johnson DG, Seibert M and Govindjee (1989) Determination of the primary charge separation rate in is- olated photosystem II reaction centres with 500-fs time re- solution. Proc Natl Acad Sci USA 86:524-528
Yamamoto Y, Shinkai H, Isogai Y, Matsuura K and Nishimura M (1984) Isolation of an Mn-carrying 33-kDa protein from an oxygen-evolving photosystem-II preparation by phase par- titioning with butanol. FEBS Lett 175:429-432