Embed Size (px)
Citation preview

Reevaluation of Oxidative Phosphorylationin Cardiac Mitochondria from NormalAnimals and Animals in Heart FailureBy George E. Lindenmayer, M.D., Louis A. Sordahl, Ph.D.,
and Arnold Schwartz, Ph.D.
ABSTRACTFor an adequate evaluation of mitochondria from diseased hearts, basic
characteristics of isolation, storage, media, ultrastructure and type of assaywere first determined using mitochondria from normal animals. A proteinaseprocedure yielded mitochondria from small laboratory animals, with low respira-tory control and marked permeability changes. The isolation medium yieldingthe most stable mitochondria with the highest respiratory control contained0 .18M KC1, 10mM EDTA, and 0.5% to 1% bovine serum albumin at pH 7.2.Heart failure in guinea pigs and rabbits was produced by varying degrees ofstenosis of the ascending aorta. An aberration in respiratory control was foundin mitochondria from hearts in severe failure. The quantitative differencesbetween normal and experimental respiratory control values were greatest whenthe highest possible normal respiratory control levels were obtained. The dif-ference between mitochondria prepared by a proteinase method from controland failing hearts was minimal. No changes in oxidative phosphorylation werenoted in mitochondria from hearts arrested by nitrogen, suggesting that acutehypoxia does not irreversibly damage energy-liberating reactions. It is con-cluded that severe heart failure is characterized by defects in mitochondria]oxidative phosphorylation, and that techniques of isolation or assay or bothare probably not causing the abnormalities.
ADDITIONAL KEY WORDShypoxia energy production
cardiac mitochondria respiratory controlcardiac arrest guinea pig rabbit
B The cellular and molecular events char-acteristic of congestive heart failure are stilllargely unknown. During the past 10 years,two general and opposing concepts of ener-getics in heart failure have evolved, oneimplicating an aberration in energy-liberating
From the Department of Pharmacology, BaylorUniversity College of Medicine, Houston, Texas77025.
These studies were supported by U. S. PublicHealth Service Grants HE 07906-06 and HE 05435-08P8 from the NationalHeart Institute and by grantsfrom the Texas and Houston Heart Associations.
Dr. Lindenmayer is a postdoctoral trainee of theU. S. Public Health Service (GM 00670-07). Dr.Schwartz is the recipient of a Research Career andDevelopment Award from the U. S. Public HealthService (K,HE 11,875-04).
A partial report of these studies was presented atthe 52nd Annual Meeting of the Federation of Ameri-can Societies for Experimental Biology.
Accepted for publication July 21, 1968.
reactions catalyzed by mitochondria (1-8)and the other describing normal energy pro-duction (9-11). The importance of mito-chondria in normal cardiac function isexemplified by the fact that the cardiacmuscle cell consists of at least 20% by dryweight of these organelles (12). The con-flicting data may be explained in one or bothof two ways: (a) The experimental modelsor human tissue employed were in differentdegrees or states of failure, ( b ) Isolation andassay conditions of subcellular componentsvaried significantly in different laboratories.In a recent assessment of the problem, Sobelet al. (13) reported no differences from con-trols in oxidative phosphorylation of mito-chondria from guinea pig hearts that failedafter partial stenosis of the ascending aorta.Their experimental model was the same asthat previously employed in two laboratories
Circulation Research, Vol. XXIII, September 19 439
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

440 LINDENMAYER, SORDAHL, SCHWARTZ
(2, 3) where significant alterations in mito-chondria were observed. Sobel et al. sug-gested that the Warburg manometric tech-nique, used for the above studies (2, 3)results in mitochondrial alterations because ofdamage during the relatively prolongedincubation period in fortified media. Theiruse of the more rapid oxygen electrodemethod revealed no differences betweenmitochondria from failing hearts and thosefrom controls (13). Since previous mano-metric studies were carried out under con-trolled conditions (2, 3, 5) the explanationby these authors appears doubtful. Whenproperly conducted and using fortified media,manometry is an adequate and useful methodfor estimating several important aspects ofmitochondrial activity (14).
Some of the difficulties involved in assayingactivities of heart mitochondria (e.g., reten-tion of endogenous substrate, ion content)have been discussed by others (15-19). Toresolve the problem of whether isolation orassay techniques or conditions are responsiblefor the reported mitochondrial defects inheart failure, we have investigated, in depth,basic characteristics of isolation, storage(aging), media, and type of assay of cardiacmitochondria from normal animals usuallyemployed in laboratory experiments. Allknown biochemical and ultrastructural testsfor mitochondrial intactness and activity wereused. Considering this information, a reex-amination of mitochondria derived from thepreviously employed "failing" model was thenundertaken. To avoid complex assay condi-tions, a polarographic procedure using anoxygen electrode with relatively simple in-cubation media was used. The results of thisinvestigation clearly support previous con-clusions (1-8) and reemphasize the impor-tance of mitochondria in severe heart failure.
Methods
Preparation of Mitochondria.— After removal ofthe hearts, the pericardium and vessels were re-moved by trimming. The whole heart was chilledin the particular isolation medium (indicated inthe tables), weighed, minced, and homogenizedwith a Teflon pestle and glass homogenizing ves-
sel or Polytron tissue processor (3 to 4 seconds ata rheostat setting of 2) as previously described(20, 21). Five isolation media were used: (a)0 . 2 5 M sucrose, lmM or 10mM ethylenediaminetetraacetate (EDTA), and 10mM tris-HCl; (b)0 . 1 8 M KC1, 10mM EDTA, and 0.5% bovine serumalbumin (Fraction V, Sigma); (c) 0 . 18M KC1and lOmM EDTA; (d) 0 . 1 8 M choline Cl andlOmMEDTA; (e) aproteinase-containingmedium(22, 23) (Nagarse, Enzyme Development Corp.,New York). The sodium salt of EDTA was usedthroughout, except for the choline medium, whichemployed the tris salt. All media were adjustedto pH 7.0 to 7.2 with tris base,
The homogenate was centrifuged at 600 X gfor 10 minutes; the supernatant fluid was strainedthrough several layers of cheesecloth and centri-fuged at 8,500 X g for 15 minutes; the pellet waswashed twice (20, 21).
Measurement of Respiratory Control—Respira-tory control and ADP-O ratios were determinedwith an oxygen electrode apparatus (Oxygraph,Gilson Medical Electronics) using either a vibrat-ing platinum electrode or a Clark oxygen elec-trode (Yellow Springs Instruments), in an assaymedium containing 0 . 2 5 M sucrose, 10mM tris-HClat pH 7.4, 8.5mM K2HPO4, 5mM tris-glutamateor tris-succinate and mitochondria (1.0 to 1.5mg/ml). The procedure and definitions have beencompletely described (21). Calculations of res-piratory control were made from data derivedafter the first addition of ADP. The fortified assaymedium in the manometric experiments was thesame as previously used (3).
Protein determinations were done by a biuretmethod (24). Assays using equivalent (±10%)amounts of mitochondrial protein were compared.
Congestive Heart Failure.—Albino guinea pigs(400 to 700 g) were anesthetized with 25 mg/kgpentobarbital ip and were ventilated by an intra-tracheal cannula and a Phipps and Bird respirator.The gas used was 955S O,-5« CO2 mixture. Theascending aorta was narrowed as previously de-scribed (3). After recovery, the animals wereclosely observed until the previously describedsigns (3) were manifest. At this time the guineapig with heart failure and a control or sham-operated guinea pig were anesthetized with 25mg/kg (control) or 15 mg/kg (experimental)pentobarbital, the trachea was intubated and thelungs were ventilated. The thoracic and peri-toneal cavities were examined prior to removalof the heart.
Acute Heart Failure.—This was induced inanesthetized, open-chest guinea pigs by narrowingthe ascending aorta 50% to 60% with 00 silk (3).The animal with partial aortic occlusion and acontrol animal were simultaneously ventilatedwith 95% O2-5% CO2. The hearts were removed
Circulation Research, Vol. XXIII, September 1968
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from
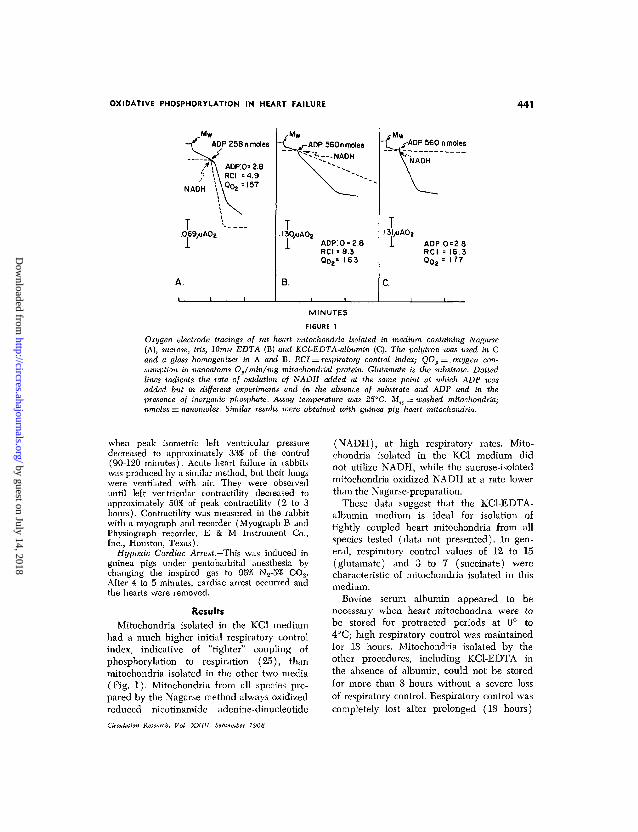
OXIDATIVE PHOSPHORYLATION IN HEART FAILURE 441
- f ADP 258 n molesV /"^ <
/ V \ ADP:O=2.8/ \ \ RCI =4.9
NADH \ \ ° O 2 = I 5 7
T I . . . .069/JAOZ
A.
_yM w
-ADP 560nmoles
^ . . - " N A D H
T. 130/iAOa
1 ADP:O = 2.8RCI = 8.3Qo2
s 1 63
B.
i-Mw"T r-ADP 560 n moles
VNADH
^ —
I3I /JAO 2
1 ADP:O=2.8RCI = 16.3Qo2 = 177
c.
FIGURE 1
Oxygen electrode tracings of rat heart mitochondria isolated in medium containing Nagarse(A), sucrose, tris, IOVIM EDTA (B) and KCl-EDTA-albumin (C). The polytron was used in Cand a glass homogenizer in A and B. RCI = respiratory control index; QOS = . oxygen con-sumption in nanoatoms Os/min/mg mitochondrial protein. Glutamate is the substrate. Dottedlines indicate the rate of oxidation of NADH added at the same point at which ADP wasadded but in different experiments and in the absence of substrate and ADP and in thepresence of inorganic phosphate. Assay temperature was 25°C. Mw = washed mitochondria;nmoles = nanomoles. Similar results were obtained with guinea pig heart mitochondria.
when peak isometric left ventricular pressuredecreased to approximately 33% of the control(90-120 minutes). Acute heart failure in rabbitswas produced by a similar method, but their lungswere ventilated with air. They were observeduntil left ventricular contractility decreased toapproximately 50% of peak contractility (2 to 3hours). Contractility was measured in the rabbitwith a myograph and recorder (Myograph B andPhysiograph recorder, E & M Instrument Co.,Inc., Houston, Texas).
Hypoxic Cardiac Arrest.—This was induced inguinea pigs under pentobarbital anesthesia bychanging the inspired gas to 95% N2-5% CO2.After 4 to 5 minutes, cardiac arrest occurred andthe hearts were removed.
Results
Mitochondria isolated in the KC1 mediumhad a much higher initial respiratory controlindex, indicative of "tighter" coupling ofphosphorylation to respiration (25), thanmitochondria isolated in the other two media(Fig. 1). Mitochondria from all species pre-pared by the Nagarse method always oxidizedreduced nicotinamide adenine-dinucleotide
Circulation Research, Vol. XXIII, September 196S
(NADH), at high respiratory rates. Mito-chondria isolated in the KC1 medium didnot utilize NADH, while the sucrose-isolatedmitochondria oxidized NADH at a rate lowerthan the Nagarse-preparation.
These data suggest that the KCl-EDTA-albumin medium is ideal for isolation oftightly coupled heart mitochondria from allspecies tested (data not presented). In gen-eral, respiratory control values of 12 to 15(glutamate) and 3 to 7 (succinate) werecharacteristic of mitochondria isolated in thismedium.
Bovine serum albumin appeared to benecessary when heart mitochondria were tobe stored for protracted periods at 0° to4°C; high respiratory control was maintainedfor 18 hours. Mitochondria isolated by theother procedures, including KC1-EDTA inthe absence of albumin, could not be storedfor more than 8 hours without a severe lossof respiratory control. Respiratory control wascompletely lost after prolonged (18 hours)
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

442 LINDENMAYER, SORDAHL, SCHWARTZ
10 15MINUTES
Oxygen electrode tracings of rat heart mitochondriaprepared in KCl-EDTA-albumin as described inMethods. Tracings A are mitochondria assayed for 20minutes at 25° C and tracings B the same preparationassayed at 37°C. Arrows indicate additions of 268nanomoles of ADP. Numbers to left of state 3 "bursts"are QOS values expressed in nanoatoms OJmin/mgmitochondrial protein. Glutamate is the substrate.Mm — washed mitochondria; RCI = respiratory con-trol index.
storage, when mitochondria were isolated in0.36M sucrose, lOmM EDTA, 0.5% albuminat pH 7.2, indicating the importance of KC1.
Figure 2 illustrates the stability of appro-priately isolated mitochondria from variousspecies at an elevated temperature and afterrepeated additions of ADP. When a fortifiedmedium of the Warburg type was used in amanometric device during prolonged incuba-tion (50 minutes at 37°C) no decrease inoxygen consumption (state 3) occurred. Mag-nesium chloride produced a stimulation ofoxygen consumption (state 4) in the oxygenelectrode system. Subsequent addition ofoligomycin totally inhibited the Mg2 +-stim-ulated respiration. High respiration due to anuncoupled state of oxidative phosphorylation
is not inhibited by these concentrations ofoligomycin (26).
The guinea pigs, 3 to 10 days after partialconstriction of the ascending aorta, devel-oped tachycardia, dyspnea, rales, hypotherm-ia, lethargy, and congested "nutmeg" liver(3). Gross pathological examination revealedan increase in the (heart weight/bodyweight) X 103 from 2.57 ±0.09 (SEM) to 3.74±0.25; (lung weight/body weight) X 103
from 5.57±0.20 to 8.34±0.48; and (liverweight/body weight) X 102 from 3.27 ±0.13to 4.19 ± 0.25. Many animals had pleuraleffusion and ascites. Three out of ten animals,however, had significant cardiac hypertrophywithout obvious failure.
Figure 3 shows representative traces ofoxygen consumption from mitochondria iso-lated from normal and experimental guinea
TABLE 1Oxidative Phosphorylation in Congestive Heart Failure(Guinea Pigs): Sucrose-Tris-EDTA Isolation Medium
Expt
12345
Mean± SE
12345
Mean± SE
RCI
5.46.94.25.13.6
5.00.6
1.72.41.42.51.8
2.00.3
GlutamateADP:O QO2
Control2.83.32.82.62.9
2.90.1
110103202126162
14119
Experimental
1.12.42.81.62.9
2.20.1
3853455162
505
RCI
1.82.52.51.92.2
2.20.1
1.41.61.41.01.7
1.40.1
SuccinateADP:O
1.41.82.01.91.4
1.90.1
1.01.62.51.21.2
1.50.2
QO."
116123214
99190
14820
6192
12541
162
9619
The mitochondria were isolated in 0.25M sucrose,lOmM tris buffer and lmM EDTA and assayed in thesucrose-tris medium (see Methods) at 30"C. Controlrefers to cardiac mitochondria of normal guinea pigsand experimental refers to mitochondria from heartsof guinea pigs in congestive heart failure. Paired ex-periments (control and experimental) were carriedout on the same day. RCI = respiratory control index;QO2 = oxygen consumption in nanoatoms O2/min/mgmitochondrial protein.
Circulation Research, Vol. XXlll, September 1968
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

OXIDATIVE PHOSPHORYLATION IN HEART FAILURE 443
GLUTAMATE-SUBSTRATE
T0.103 JJAO2
SUCCINATE-SUBSTRATE
MINUTESFIGURE 3
Representative oxygen electrode tracings of mitochondria from a normal guinea pig heart andone in congestive heart failure. The mitochondria were isolated in 0.25M sucrose, lOmiu tris,lmM EDTA. Glutamate or succinate was used as the substrate (5niM-final concentration);assays were carried out in the sucrose-tris medium at 30° C. ADP, 550 nanomoles, was addedwhere indicated. RCI = respiratory control index; QO2 = oxygen consumption in nanoatomsOJmin/mg mitochondrial protein.
QU2- 163
ADP:O-3.24
RCI -13.3
Q 0 2 - 121
RCI-3.1
QO,- 108
T0 . 0 9 7 / J A O 2
AOP:O-2.08
EXPERIMENTAL
MINUTES
FIGURE 4
Representative oxygen electrode tracings of mitochondria from a normal rabbit heart and onein acute heart failure. The mitochondria were isolated in 0.18M KCl-10mM EDTA. Rotenone,4 fig, was added to inhibit NADH-linked oxidation, after which ADP was not utilized untilsuccinate (5miu final concentration) was added. Assay conditions were as described in Figure 1.RCI — respiratory control index; QOS = nanoatoms OJmin/mg mitochondrial protein.
Circulation Research, Vol. XXIII, September 1968
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from
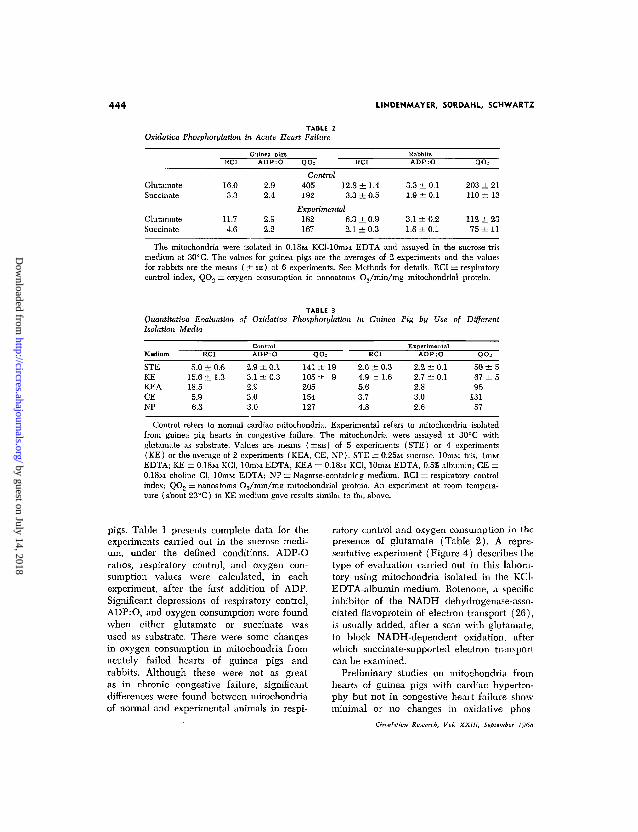
444 LINDENMAYER, SORDAHL, SCHWARTZ
TABLE 2Oxidative Phosphorylation in Acute Heart Failure
QO= RCI QO=
GlutamateSuccinate
GlutamateSuccinate
16.03.3
11.74.6
2.92.4
2.92.2
Control405 12.8 ± 1.4192 3.3 ± 0.5
Experimental182 6.3 ± 0.9167 2.1 ± 0.3
3.3 ±0.11.9 ±0.1
3.1 ± 0.21.8 ± 0.1
203 ± 21110 ± 13
112 ± 2375 ± 1 1
The mitochondria were isolated in 0.18M KCl-lOniM EDTA and assayed in the sucrose-trismedium at 30° C. The values for guinea pigs are the averages of 2 experiments and the valuesfor rabbits are the means ( ± S E ) of 6 experiments. See Methods for details. RCI = respiratorycontrol index; QO.-> = oxygen consumption in nanoatoms O.j/min/mg niitochondrial protein.
TABLE 3Quantitative Evaluation of Oxidative Phosphorylation in Guinea Pig by Use of DifferentIsolation Media
Medium
STEKEKEACE
NP
RCI
5.0 ±15.6 ±18.5
5.96.3
0.61.3
ContiADP
2.9 ±3.1 ±2.93.03.0
rol:O
0.10.3
QO=
141 ±105 ±205154127
199
RCI
2.0 ± 0.34.9 ± 1.65.63.74.8
ExperimA D P
2.2 ±2.7 ±2.83.02.6
ental:O
0.10.1
QOs
50 ±567 ±596
13157
Control refers to normal cardiac mitochondria. Experimental refers to mitochondria isolatedfrom guinea pig hearts in congestive failure. The mitochondria were assayed at 30°C withglutamate as substrate. Values are means ( ±SE ) of 5 experiments (STE) or 4 experiments(KE) or the average of 2 experiments (KEA, CE, NP). STE = 0.25M sucrose, lOmM tris, lmMEDTA; KE = 0.18M KC1, 10mM EDTA; KEA = 0.18M KC1, lOniM EDTA, 0.5% albumin; CE =0.18M choline Cl, lOmM EDTA; NP = Nagarse-containing medium. RCI = respiratory controlindex; QOO = nanoatoms O2/min/mg niitochondrial protein. An experiment at room tempera-ture (about 23°C) in KE medium gave results similar to the above.
pigs. Table 1 presents complete data for theexperiments carried out in the sucrose medi-um, under the defined conditions. ADP-0ratios, respiratory control, and oxygen con-sumption values were calculated, in eachexperiment, after the first addition of ADP.Significant depressions of respiratory control,ADP:O, and oxygen consumption were foundwhen either glutamate or succinate wasused as substrate. There were some changesin oxygen consumption in mitochondria fromacutely failed hearts of guinea pigs andrabbits. Although these were not as greatas in chronic congestive failure, significantdifferences were found between mitochondriaof normal and experimental animals in respi-
ratory control and oxygen consumption in thepresence of glutamate (Table 2). A repre-sentative experiment (Figure 4) describes thetype of evaluation carried out in this labora-tory using mitochondria isolated in the KC1-EDTA-albumin medium. Rotenone, a specificinhibitor of the NADH dehydrogenase-asso-ciated flavoprotein of electron transport (26),is usually added, after a scan with glutamate,to block NADH-dependent oxidation, afterwhich succinate-supported electron transportcan be examined.
Preliminary studies on mitochondria fromhearts of guinea pigs with cardiac hypertro-phy but not in congestive heart failure showminimal or no changes in oxidative phos-
Circulalion Research, Vol. XXI11, September 1968
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from
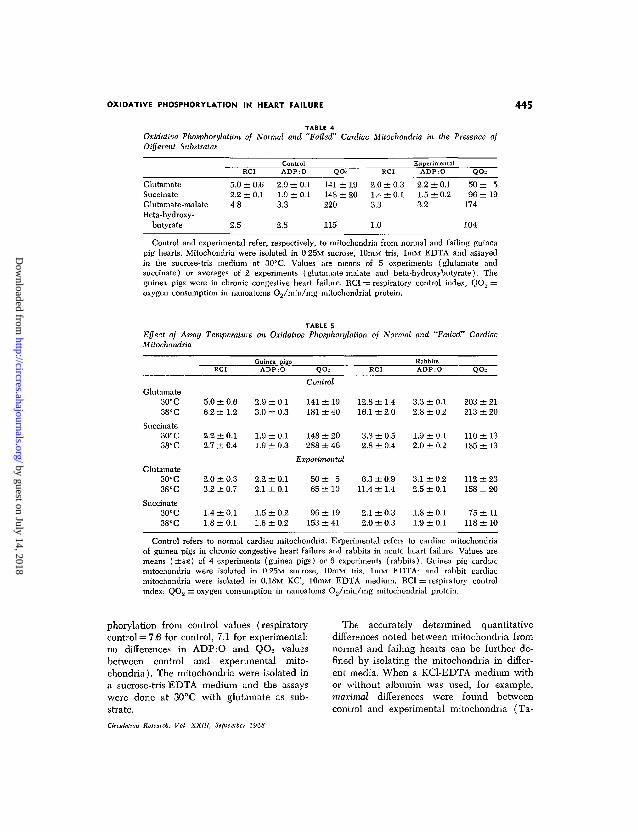
OXIDATIVE PHOSPHORYLATION IN HEART FAILURE 445
TABLE 4Oxidative Phosphorylation of Normal and "Failed" Cardiac Mitochondria in the Presence ofDifferent Substrates
GlutamateSuccinateGlutamate-malateBeta-hydroxy-
butyrate
5.02.24.8
2.5
RCI
±0.6±0.1
ContiA D P
2.9 ±1.9 ±3.3
2.8
:ol
:O
0.10.1
Q
141148220
115
± 1 9± 2 0
2.01.43.3
1.0
RCI
±0.3±0.1
ExperimentalADP:O
2.2 ±0.11.5 ±0.23.2
QO-
50 ±96 ±
174
104
519
Control and experimental refer, respectively, to mitochondria from normal and failing guineapig hearts. Mitochondria were isolated in 0,25M sucrose, 10mM tris, lmM EDTA and assayedin the sucrose-tris medium at 30°C. Values are means of 5 experiments (glutamate andsuccinate) or averages of 2 experiments (glutamate-malate and beta-hydroxybutyrate). Theguinea pigs were in chronic congestive heart failure. RCI == respiratory control index; QO2 =oxygen consumption in nanoatonis 02/min/mg mitochondrial protein.
TABLE SEffect of Assay Temperature on Oxidative Phosphorylation of Normal and "Failed" CardiacMitochondria
Glutamate30°C38°C
Succinate30° C38°C
Glutamate30° C38°C
Succinate30°C38°C
RCI
5.0 ± 0.66.2 ± 1.2
2.2 ± 0.12.7 ± 0.4
2.0 ± 0.33.2 ± 0.7
1.4 ±0.11.8 ±0.1
Guinea pigsADP:O
2.9 ± 0.13.0 ± 0.3
1.9 ±0.11.9 ±0.3
2.2 ± 0.12.1 ±0.1
1.5 ±0.21.6 ± 0.2
QO2
Control
141 ± 19181 ± 40
148 ± 20288 ± 46
Experimental
50 ± 565 ±10
96 ±19153 ±41
RCI
12.8 ±1.416.1 ±2.0
3.3 ± 0.52.8 ± 0.4
6.3 ± 0.911.4 ±1.4
2.1 ±0.32.0 ± 0.3
RabbitsADP:O
3.3 ± 0.12.8 ± 0.2
1.9 ± 0.12.0 ± 0.2
3.1 ±0.22.5 ± 0.1
1.8 ±0 .11.9 ± 0.1
QOs
203 ± 21213 ± 20
110 ± 13185 ± 13
112 ± 23158 ± 20
75 ± 1 1118 ± 10
Control refers to normal cardiac mitochondria. Experimental refers to cardiac mitochondriaof guinea pigs in chronic congestive heart failure and rabbits in acute heart failure. Values aremeans ( ± S E ) of 4 experiments (guinea pigs) or 6 experiments (rabbits). Guinea pig cardiacmitochondria were isolated in 0.25M sucrose, lOmM tris, lmM EDTA; and rabbit cardiacmitochondria were isolated in 0.18M KC1, 10mM EDTA medium. RCI = respiratory controlindex; QO2 = oxygen consumption in nanoatoms O2/min/mg mitochondrial protein.
phorylation from control values (respiratorycontrol = 7.6 for control, 7.1 for experimental;no differences in ADP:O and QO2 valuesbetween control and experimental mito-chondria). The mitochondria were isolated ina sucrose-tris-EDTA medium and the assayswere done at 30°C with glutamate as sub-strate.
Circulation Research, Vol. XXIII, September 1968
The accurately determined quantitativedifferences noted between mitochondria fromnormal and failing hearts can be further de-fined by isolating the mitochondria in differ-ent media. When a KC1-EDTA medium withor without albumin was used, for example,maxinuil differences were found betweencontrol and experimental mitochondria (Ta-
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

446 LINDENMAYER, SORDAHL, SCHWARTZ
ble 3). The choline or Nagarse-containingmedia were the poorest for detecting differ-ences between control and experimentalmitochondria. Interestingly, no difference inNADH utilization was found between mito-chondria from normal and failing hearts.
The ability of mitochondria from normaland failing hearts to use different substratesfor oxidative phosphorylation is shown inTable 4. Glutamate, glutamate-malate, andbeta-hydroxybutyrate are all NADH-linkedsubstrates. Glutamate appears to be the bestsubstrate in this group for detecting possibledifferences in oxidative phosphorylation. Thedata also show that although significant differ-ences were observed in succinate-supportedoxidative phosphorylation, the magnitudewas not as great as that found with glutamate.
In an effort to exaggerate differences be-tween mitochondria from normal and failinghearts, comparative assays were carried outat 30° and at 38°C. The data do not indicateany significant differences induced by tem-perature (Table 5), nor do they implicateany damage to mitochondria by the higherincubation temperatures. As would be ex-pected, increasing the temperature from 30°
TABLE 6Effect of ADP-Induced Repetitive Bursts on Respira-tory Control and Oxygen Consumption
Burst
1234
1234
Control*
5.3 ± 0.84.3 ± 0.63.9 ±0.8
141 ± 19110 ±3184 ± 32
111
CHF*
RCI
2.7 ±2.8 ±2.9 ±
50 ±58 ±39 ±60
0.90.81.0
52812
Controlf
16.018.0
6.86.2
405400254236
AHFf
11.78.55.06.7
182133104104
'Mitochondria from guinea pig hearts isolated inthe 0.25M sucrose, lOmM tris, lniM EDTA medium(4 experiments, means ± S E ) ; tin KC1-EDTA medi-um (averages of 2 experiments). Glutamate wassubstrate and the assays were carried out at 30°C.
CHF = congestive heart failure; AHF = acute heartfailure; RCI = respiratory control index; QO, =nanoatoms 02/min/mg mitochondrial protein.
to 3S°C caused an increase in oxygen con-sumption.
Exaggerated differences between mito-chondria from normal and failing hearts werefound, however, when the mitochondria wereincubated for 60 to 180 minutes at 0°C priorto assay (data not shown).
The data in Table 6 indicate the possibleerror of averaging repetitive bursts in estima-tion of respiratory control and ADP:O values.Respiratory control and other indices of mito-chondria isolated in a sucrose medium tendto decrease with repetitive additions of ADPand the normals tend to reach that of themitochondria from failing hearts. ADP:O val-ues remain essentially constant after repetitiveadditions of ADP for both control andexperimental mitochondria. However, whenmitochondria are isolated in the KC1-EDTA-albumin medium, multiple additions of ADPmay be made without significant respiratorycontrol changes (Fig. 2).
Mitochondria from guinea-pig hearts arenot so rigorously dependent upon adequatein vivo oxygenation for the maintenance offunctional integrity as one might suspect. Fiveto 6 minutes of hypoxia in guinea pigs (severeenough to cause cardiac arrest) had no sig-nificant effect on oxidative phosphorylation(respiratory control = 12.0 for control, 10.7for hypoxia; ADP:O-2.8 for control, 2.8 forhypoxia; oxygen consumption [nanoatomsOo/min/mg mitochondrial protein] =214 forcontrol, 218 for hypoxia; these values wereobtained with glutamate as substrate at 30°C;similar results were noted when succinate wasemployed).
DiscussionIn an appraisal of molecular events existing
in or leading to a pathologic state, one mustusually disrupt the cell and study subcellularsystems. In doing this, the organelles areplaced in at least two artificial environments,the isolation and assay media. If any estimateof in vivo function is to be made, the isolationand assay media at least, should be as non-destructive and as "physiological" as possible.
Circulation Research, Vol. XXI11, September 1968
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

OXIDATIVE PHOSPHORYLATION IN HEART FAILURE 447
The obvious conclusion is that the probabili-ty of finding a difference between a normaland diseased state is decreased if the sub-cellular systems are damaged or altered dur-ing isolation and assay. The present studysupports this reasoning as applied to study ofmitochondria from normal and failing hearts.
Initial respiratory control of mitochondriafrom normal hearts prepared by the KC1-EDTA medium with or without albumin (orwith choline-EDTA-albumin medium) washigher than that obtained using other pub-lished methods (9, 22, 23, 27). Mitochondriaprepared by this method also maintain theirbiochemical integrity (as determined byrespiratory control) during protracted peri-ods of storage. A relatively high level ofEDTA (lOmivi) should be used when isolat-ing cardiac mitochondria because of the largeamounts of calcium present in cardiac tissueand because Ca2+ can cause uncoupling ofmitochondria (26). The role of albumin inmaintaining the biochemical integrity of car-diac mitochondria is unknown, although theevidence is in favor of mechanisms morecomplex than free fatty acid binding (28-31).
Using the conditions defined for mito-chondrial isolation and assay, it is clear thatdefects in oxidative phosphorylation do existin both chronically and acutely failing, butnot hypoxic, hearts of the experimental mod-els employed. How can this conclusion beresolved with those studies indicating nochanges, using the same experimental model(9, 13)? The simplest explanation is thatthere is variance in the degree of failure inanimals used in different laboratories. Thepresence of normal mitochondria in differenttypes of experimental models has been pre-viously discussed (3, 32).
More important, however, is the possibilitythat varying aspects of technique are relevantin explaining the divergent data. The presentstudy directs attention to the followingpoints:
1. Averaging of values for respiratory con-trol or selection of values for Qo2 afterrepetitive additions of ADP to mitochondria
Circulation Research, Vol. XXIII, September 1968
that are isolated in certain media (13) maylead to erroneous conclusions.
2. In terms of isolation, the proteinaseprocedure for small laboratory animals oftenyields mitochondria with significant perme-ability changes which might make compari-sons difficult. Thus Nagarse-prepared mito-chondria (both normal and experimental)rapidly utilized NADH, which implies "leakymembranes," i.e., an increase in permeabilityof the external mitochondrial membrane (33).Differences between control and experimentalmitochondria in the present study were muchless apparent when the Nagarse procedurewas used than when other isolation methodswere used.
3. Properly prepared cardiac mitochondriafunction adequately at 25°, 30°, and 37°Cwith minimal or no deterioration of activityor structure for at least 20 to 30 minutes. Noinformation on possible NADH oxidation inprevious studies was reported (9-11, 13). Ifa significant loss of respiratory control doesoccur when mitochondria are incubated at37°C in the presence or absence of fortifiedmedia, the isolation technique should bequestioned (13).
Sobel et al. (13) have presented threesuggestions to explain how differences inoxidative phosphorylation between controland experimental animals might be artifacts,(a) The release of free fatty acids (FFA)during prolonged incubation at 37°C mayhave deleterious effects on mitochondrialfunction. This is improbable since the presentdata clearly show that inclusion of albuminand EDTA, both inhibitors of FFA-induceduncoupling of mitochondria (31, 34), resultedin maximal differences in respiratory controlbetween mitochondria from normal and fail-ing hearts, (b) The use of Warburg manom-etry, which requires relatively long incubationperiods, may result in deterioration of mito-chondria. There is no evidence to supportthis contention. Magnesium does not un-couple or damage oxidative phosphorylationbut, rather, maintains a high rate of ADPproduction. Manometry, as an adequate and
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

448 LINDENMAYER, SORDAHL, SCHWARTZ
important technique, has recently been dis-cussed by Slater (14). Furthermore, sincecontrol and experimental mitochondria werestudied concomitantly (2, 3, 5), it is doubtfulthat prolonged incubation alone would se-lectively damage only the mitochondria fromfailing hearts. If this were in fact true, itwould reflect a significant difference (perhapsfragility) between the control and experimen-tal preparations. In any case the presentreport vitiates the incubation argument byemploying an oxygen electrode system anda relatively simple assay medium, and theresults obtained were similar to those pre-viously reported (1, 3, 5). The results ofprolonged storage at 0°C of control andexperimental mitochondria, presented in thiscommunication, suggest that the experimentalmitochondria are perhaps more fragile thannormal, (c) The reported changes might bedue to acute hypoxia resulting from pro-longed time between death of the animaland removal of the heart. The present datamake this possibility remote, since acute invivo hypoxia does not effect significantchanges in respiratory control of the mito-chondria. These data are consistent withprevious studies showing recovery of the con-tractility of the acutely anoxic heart concomi-tant with a partial restoration of energy-richphosphate compounds and oxidative phos-phorylation (35). Currently, this informationis of primary importance in evaluation of cardi-ac transplantation procedures. However, thesedata do not preclude the interesting possibilitythat the observed mitochondrial differencesin failure may be secondary to a chronic invivo hypoxia.
Summarizing our findings, the isolated car-diac mitochondria used in the present study,to our knowledge, are in the best currentlyknown in vitro biochemical state. This wasdetermined by several established criteria:very high respiratory control, high oxygenconsumption, stability at various temperaturesand after repeated additions of ADP, limitedNADH utilization indicating intact outermembranes, and maintenance of activity af-ter prolonged storage. These observations are
further substantiated by electron microscopicstudies showing no significant ultrastmcturalchanges of isolated normal mitochondria evenafter 1 hour of incubation (state 4) at 30°Cor 20 minutes of incubation at 37°C (datanot presented). The biochemical integrity ofcardiac mitochondria has not been so docu-mented in all the previously reported studies(1-11, 13). Considering this information onnormal mitochondria, the changes reportedin this study have added meaning. It is highlydoubtful that the observations are artifactsof either isolation or assay (13). This studywas specifically undertaken to eliminate suchan argument by using as wide a range ofisolation and assay techniques as possible.
Chidsey et al. have presented an interest-ing discussion on divergencies in data (11).They found no mitochondrial changes inhuman papillary muscle from failing hearts.These authors suggest that the followingreasons could apply. No changes exist inhuman heart failure; their patients were onmaintenance doses of digoxin and may nothave been in the same severe state of failureas that in which changes have been reported.Humans typically develop hypertrophy andfailure much more slowly than most experi-mental models. Papillary muscle might notbe representative of the ventricular wall. Wewould like to add that data obtained fromisolated ventricular muscle obtained by biop-sy may not represent the average state ofenergy-liberating mechanisms either in thetotal ventricle or in the whole heart. Theheterogeneity of heart muscle may precludethe validity of extrapolating data obtainedfrom small segments to total tissue. For thisreason we studied energy liberation in theentire heart. The possibility that defectivemitochondria were diluted with normal mito-chondria only serves to make the differencesbetween the average values obtained fromnormal and diseased hearts that much moremeaningful. Obviously, an examination ofthis type yields only minimal informationconcerning the development of a pathologicalstate in individual areas of the heart.
Circulation Research, Vol. XXI11, September 1968
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

OXIDATIVE PHOSPHORYLATION IN HEART FAILURE 449
We wish to stress the possible importanceof the stage of heart failure. Meerson (36)has defined three significant developmentalstages in the failing heart: (1) transientbreakdown stage, characterized by contractileinsufficiency with a deficiency of certainenzymes; (2) protracted stage of relativelystable hyperfunction, characterized by the ab-sence of cardiac insufficiency, by hypertrophy,and by adequate oxidative phosphorylation;and (3) protracted stage of progressingcardiosclerosis and gradual exhaustion ofthe hypertrophied heart, characterized bydisturbances in protein synthesis and sus-tained hypoxia. We suggest that stage 3might be characterized by or caused by anaberration of mitochondrial function(s). Itis of interest that Syrian hamsters in severecongestive heart failure, secondary to agenetically linked cardiomyopathic process,were not in a preterminal condition but didshow significant differences between controland experimental mitochondria (37, 38).Hamsters that were in the hypertrophic phase(2, above) but did not exhibit congestiveheart failure, however, showed normal mito-chondrial activity (39). Mitochondria iso-lated from the hearts of animals or humanswith hypertrophy in the absence of severefailure might be expected, therefore, to showlittle or no abnormality.
References1. SZEKERES, L., AND SCHEIN, M.: Cell metabolism
of the overloaded mammalian heart in situ.Cardiologia 34: 18, 1959.
2. GEHTLEB, M. M.: Differences in efficiency ofenergy transfer in mitochondrial systems de-rived from normal and failing hearts. Proc.Soc. Exptl. Biol. Med. 106: 109, 1961.
3. SCHWARTZ, A., AND LEE, K. S.: Study of heartmitochondria and glycolytic metabolism in ex-perimentally induced cardiac failure. Circula-tion Res. 10: 321, 1962.
4. MEERSON, F. Z., ZALETAYEVA, T. A., LACUTCHEV,
S. S., AND PSHENNIKOVA, M. D.: Structure and
mass of mitochondria in the process of com-pensatory hyperfunction and hypertrophy ofthe heart. Exptl. Cell Res. 36: 568, 1964.
5. ARGUS, M. F., AHCOS, J. C , SARDESAI, V. M.,
AND OVEHUY, J. L.: Oxidative rates and phos-phorylation in sarcosomes from experimentally-
Circiitation Research, Vol. XXIII, September 1968
induced failing rat heart. Proc. Soc. Exptl. Biol.Med. 117: 380, 1964.
6. BlNC, R. J., WU, C, AND GuDBJAHNASON, S.:Mechanism of heart failure. Circulation Res.15 (supp. II): 11-64, 1964.
7. WOLLENBERGER, A., KXEITKE, B., AND RAABE, G.:Some metabolic characteristics of mitochondriafrom chronically overloaded, hypertrophiedhearts. Exptl. Mol. Path. 2: 251, 1965.
8. Fox, A. C , AND REED, G. E.: Exchanges of
nucleotide phosphates in normal and in failingcanine hearts. Am. J. Physiol. 110: 1383, 1966.
9. PLAUT, G. W. E., AND GERTLER, M. M.: Oxida-
tive phosphorylation studies in normal andexperimentally produced congestive heart fail-ure in guinea pigs: A comparison. Ann. N. Y.Acad. Sci. 72: 515, 1959.
10. OLSON, R. E.: Abnormalities of myocardial me-tabolism. Circulation Res. 15 (suppl. I I ) :11-109, 1964.
11. CHIDSEY, C. A., WEINBACH, E. C , POOL, P. E.,
AND MORROW, A. G.: Biochemical studies ofenergy production in the failing human heart.J. Clin. Invest. 45: 40, 1966.
12. PAGE, E., AND PACE, E. G.: Distribution of ions
and water between tissue compartments in theperfused left ventricle of the rat heart. Circu-lation Res. 22: 435, 1968.
13. SOBEL, B. E., SPANN, J. F., POOL, P. E. SONNEN-
BLICK, E. H., AND BRAUNWALD, E,: Normal
oxidative phosphorylation in mitochondriafrom the failing heart. Circulation Res. 21:355, 1967.
14. SLATEH, E. C : Manometric methods and phos-phate determination. In Methods of Enzymol-ogy, vol. 10, edited by R. W. Estabrook andM. E. Pullman. New York, Academic Press,1967, p. 19.
15. TARJAN, E. M., AND VON KORFF, R. W.: Factors
affecting the respiratory control ratio of rabbitheart mitochondria. J. Biol. Chem. 242: 318,1967.
16. BAUER, C , AND VON KORFF, R. W.: Variation of
endogenous substrates and pyruvate metab-olism in isolated heart mitochondria of sev-eral species. Biochim. Biophys. Acta 131: 280,1967.
17. OLSON, M. S., AND VON KORFF, R. W.: Changes
in endogenous substrates of isolated rabbitheart mitochondria during storage. J. Biol.Chem. 242: 325, 1967.
18. OZAWA, K., SETA, K., ARAKT, H., AND HANDA, H.:
Dependence of brain mitochondrial respirationon potassium ion. Biochem. J. 61: 352, 1967.
19. PACKER, L.: Coupled phosphorylation in rat heartmuscle sarcosomes. Arch. Biochem. Biophys.70: 290, 1957.
20. SAFER, B., AND SCHWARTZ, A.: Active transport
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

450 LINDENMAYER, SORDAHL, SCHWARTZ
of potassium ion in heart mitochondria. Circu-lation Res. 21: 25, 1967.
21. SOHDAHL, L. A., AND SCHWARTZ, A.: Effects ofdipyridamole on heart muscle mitochondria.Mol. Pharm. 3: 509, 1967.
22. CHANCE, B., AND HACMARA, C : Initiation ofsuccinate oxidation in aged pigeon heart mito-chondria. Biochem. Biophys. Res. Commun.3: 1, 1960.
23. SOBEL, B., JEQUIER, E., SJOERDSMA, A., ANDLOVENBERG, W.: Effect of catecholamines andadrenergic blocking agents on oxidative phos-phorylation in rat heart mitochondria. Circu-lation Res. 19: 1050, 1966.
24. JACOBS, E. A., JACOBS, M., SANADI, D. R., ANDBRADLEY, L. B.: Uncoupling of oxidative phos-phorylation by cadmium ion. J. Biol. Chem.223: 147, 1956.
25. CHANCE, B., AND WILLIAMS, G. R.: The respira-tory chain and oxidative phosphorylation. Ad-van. Enzymol. 17: 65, 1956.
26. LEHNINGER, A. L.: The Mitochondrion. NewYork, W. A. Benjamin, Inc., 1965.
27. SCHNEIDER, W. C : Intracellular distribution ofenzymes: III. The oxidation of octanoic acidby rat liver fractions. J. Biol. Chem. 176: 259,1948.
28. WEINBACH, E. C, GAHBUS, J., AND SCHEFFIELD,H. G.: Morphology of mitochondria in thecoupled, uncoupled and recoupled states. Exptl.Cell. Res. 46: 129, 1967.
29. DAVIS, J. T.: Technique for the isolation ofmitochondria from bovine lymphocytes. InMethods in Enzymology, vol. 10, edited byR. W. Estabrook and M. E. Pullman, NewYork, Academic Press, 1967, p. 115.
30. WEINBACH, E. C, AND GARBUS, J.: Structuralchanges in mitochondria induced by un-
coupling reagents, the response to proteolyticenzymes. Biochem. J. 106: 711, 1968.
31. VAN DEN BERGH, S. G.: Fatty acid activationand oxidation by mitochondria. In Methods inEnzymology, vol. 10, edited by R. W. Esta-brook and M. E. Pullman, New York, Aca-demic Press, 1967, p. 749.
32. PROCITA, L., SCHWARTZ, A., AND LEE, K. S.:Oxidative phosphorylation in the failing dogheart-lung preparation. Circulation Res. 16:391, 1965.
33. LEHNINCER, A. L.: Phosphorylation coupled tooxidation of DPNH, J. Biol. Chem. 190: 359,1951.
34. KENDAHL, F. E.: Studies on human serum pro-tein: II. Crystallization of human serum al-bumin. J. Biol. Chem. 138: 97, 1941.
35. FEINSTEIN, M. B.: Effects of experimental con-gestive heart failure, ouabain, and asphyxia onthe high-energy phosphate and creatine con-tent of the guinea pig heart. Circulation Res.10: 333, 1962.
36. MEERSON, F. Z.: Compensatory hyperfunction ofthe heart. Circulation Res. 10: 250, 1962.
37. SCHWARTZ, A., LINDENMAYER, G., AND HARIGAYA,A.: Respiratory control and calcium transportin heart mitochondria from the cardiomyo-pathic Syrian hamster. Ann. N. Y. Acad. Sci.(in press).
38. OPIE, L. H., LOCKNER, A., BRINK, A. J., HOM-BURGEH, G., AND NIXON, C. W.: Oxidativephosphorylation in hereditary myocardiopathyin the Syrian hamster. Lancet 2: 1213, 1964.
39. WROGEMANN, K., AND BLANCHAEB, M. C: Res-piration and oxidative phosphorylation bymuscle and heart mitochondria of hamsterswith hereditary myocardiopathy and polymyo-pathy. Can. J. Biochem. 46: 323, 1968.
cb, Vol. XX11I, September 1968
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

SCHWARTZGEORGE E. LINDENMAYER, LOUIS A. SORDAHL and ARNOLD
from Normal Animals and Animals in Heart FailureReevaluation of Oxidative Phosphorylation in Cardiac Mitochondria
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 1968 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleCirculation Research
doi: 10.1161/01.RES.23.3.4391968;23:439-450Circ Res.
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer
page under Services. Further information about this process is available in therequested is located, click Request Permissions in the middle column of the Web Once the online version of the published article for which permission is beingRightsLink, a service of the Copyright Clearance Center, not the Editorial Office.
can be obtained viaCirculation Researcharticles originally published in Requests for permissions to reproduce figures, tables, or portions ofPermissions:
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from

http://circres.ahajournals.org/content/23/3/439services, is located on the World Wide Web at:
The online version of this article, along with updated information and
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer
page under Services. Further information about this process is available in therequested is located, click Request Permissions in the middle column of the Web Once the online version of the published article for which permission is beingRightsLink, a service of the Copyright Clearance Center, not the Editorial Office.
can be obtained viaCirculation Researcharticles originally published in Requests for permissions to reproduce figures, tables, or portions ofPermissions:
by guest on July 14, 2018http://circres.ahajournals.org/
Dow
nloaded from





























