Embed Size (px)
Citation preview

Review
Recent methodological advances in thediscovery of GPCR-associated proteincomplexesAvais M. Daulat*,1,2,3, Pascal Maurice*,1,2,4 and Ralf Jockers1,2
1 Institut Cochin, Universite Paris Descartes, Centre National de la Recherche Scientifique (UMR 8104),
Department of Cell Biology, F-75014 Paris, France2 Institut national de la sante et de la recherche medicale, U567, F-75014 Paris, France3 Present address: Leslie Dan Faculty of Pharmacy, University of Toronto, Toronto, ON M5S 3M2, Canada4 Present address: Centre de Recherche de l’Institut de Cardiologie de Montreal et Universite de Montreal, Montreal,
QC H1T 1C8, Canada
Protein-interaction networks have important roles incellular homeostasis and the generation of complexityin biological systems. G-protein-coupled receptors(GPCRs), the largest family of membrane receptorsand important drug targets, are integral parts of thesenetworks. Ligand stimulation and the dynamic inter-action with GPCR-associated protein complexes(GAPCs) constitute two important regulatory mechan-isms of GPCR function. Several genomic and proteomicapproaches have been developed to identify GAPCs inthe past. However, this task turned out to be particularlydemanding owing to difficulties in preserving the com-plex three-dimensional GPCR structure during receptorsolubilization and to inherent limitations in the use ofisolated receptor domains as bait. Newly emergingmethods have the potential to overcome these limita-tions and will certainly boost the identification of func-tionally relevant GAPCs to finally increase ourknowledge of the regulation of GPCRs and provide noveldrug targets. Here, we focus on the comparison of twocomplementary GAPC purification strategies, which arebased on soluble GPCR subdomains and entire GPCRs.
IntroductionThe superfamily of seven-transmembrane-spanning G-protein-coupled receptors (GPCRs) constitutes the largestfamily of membrane receptors with >800 members inhumans. GPCRs respond to a wide variety of extracellularstimuli and are the targets of 30–50% of available medi-cation for human diseases [1]. The initial view of the basicGPCR signaling module, which included a receptor, aheterotrimeric G protein and effectors, turned out to beoversimplified. Various recent studies show that GPCRsbind to a host of different proteins placing GPCRs at thecentre of large protein-interaction networks dedicated tothe export to the cell surface, signal transduction, desen-sitization, intracellular trafficking, fine-tuning and allo-steric regulation of these receptors. Depending on theextra- and intra-cellular environment and the physiologi-cal state of the cell, the composition and dynamics of these
Corresponding author: Jockers, R. ([email protected])* These authors contributed equally..
72 0165-6147/$ – see front matter � 2008 Elsevier L
complexes vary and, thus, participate in the dynamicregulation of receptor function.
The last ten years have seen an unprecedented revolu-tion in the field of proteomics, mainly owing to advances inprotein identification methods by mass spectrometry (MS)resulting in the description of interactomes of whole cells[2]. Despite these major advances, the characterization ofmembrane protein complexes is still in its infancy. Indeed,membrane proteins are usually under-represented innewly identified protein networks [2]. The analysis ofmembrane proteins such as GPCRs and associated proteincomplexes has always been a challenge because GPCRs aretypically expressed at low levels, are poorly immunogenicand are difficult to solubilize in a functionally active form.
In an attempt to avoid receptor solubilization, solublereceptor subdomains have been used as bait in yeast two-hybrid screens and affinity chromatography purifications.Several components of the GPCR-associated protein com-plexes (GAPCs) that interact directly and indirectly withthe receptor have been identified, with metabotropic glu-tamate (mGlu) and serotonin (5-HT) receptors being themost extensively investigated GPCRs, as summarized inrecent reviews [3–7]. Further methods such as the tandemaffinity purification (TAP) approach [8] and the split-ubi-quitin membrane two-hybrid system [9] that use the entirereceptor as bait have recently been added to the tool box.Here, we focus on the description and comparison of twocomplementary purification procedures, the peptide affi-nity chromatography purification and TAP, that are basedon soluble GPCR subdomains and entire GPCRs, respect-ively.
Identification of GAPCs using GPCR subdomainsSoluble GPCR subdomains, typically the C terminus, havebeen extensively used in the past to screen for GAPCs tocircumvent problems associated with the hydrophobicnature of full-length GPCRs. Several genomic- and proteo-mic-based approaches have been described, such as theyeast two-hybrid approach and protein micro-arrays,which will not be discussed here in detail. The yeasttwo-hybrid approach, which detects direct interacting part-ners from cDNA libraries, was applied to a great number of
td. All rights reserved. doi:10.1016/j.tips.2008.10.009 Available online 25 December 2008
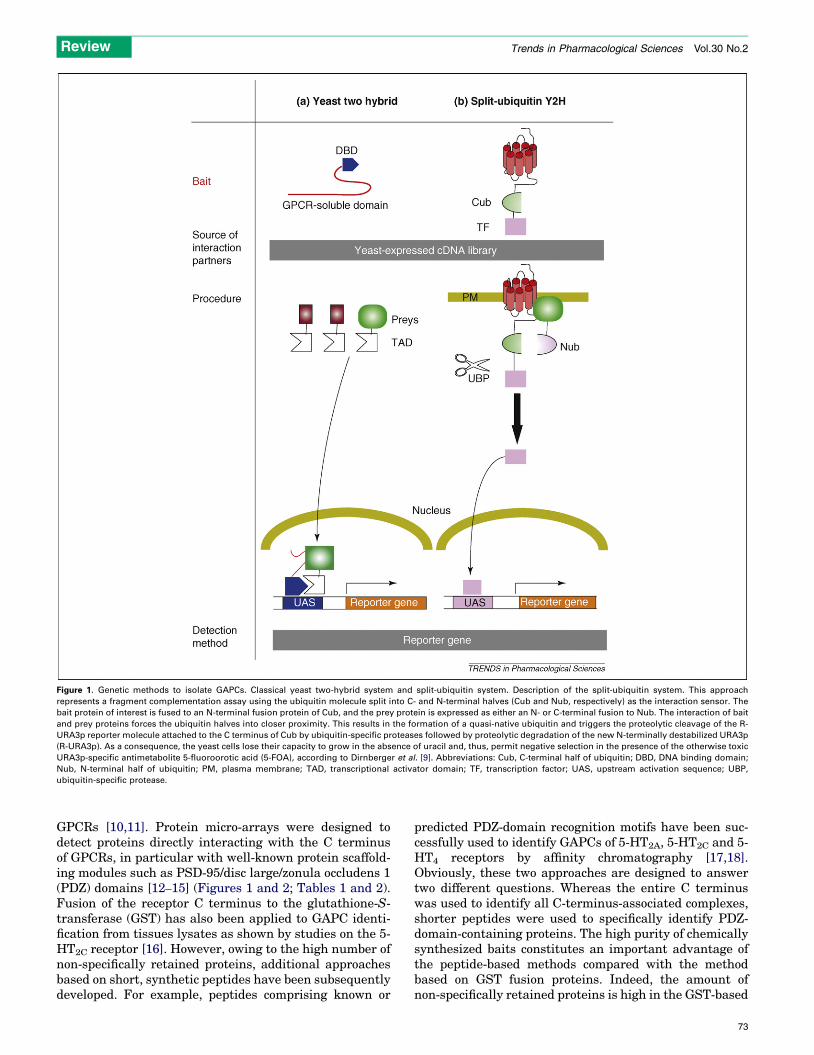
Figure 1. Genetic methods to isolate GAPCs. Classical yeast two-hybrid system and split-ubiquitin system. Description of the split-ubiquitin system. This approach
represents a fragment complementation assay using the ubiquitin molecule split into C- and N-terminal halves (Cub and Nub, respectively) as the interaction sensor. The
bait protein of interest is fused to an N-terminal fusion protein of Cub, and the prey protein is expressed as either an N- or C-terminal fusion to Nub. The interaction of bait
and prey proteins forces the ubiquitin halves into closer proximity. This results in the formation of a quasi-native ubiquitin and triggers the proteolytic cleavage of the R-
URA3p reporter molecule attached to the C terminus of Cub by ubiquitin-specific proteases followed by proteolytic degradation of the new N-terminally destabilized URA3p
(R-URA3p). As a consequence, the yeast cells lose their capacity to grow in the absence of uracil and, thus, permit negative selection in the presence of the otherwise toxic
URA3p-specific antimetabolite 5-fluoroorotic acid (5-FOA), according to Dirnberger et al. [9]. Abbreviations: Cub, C-terminal half of ubiquitin; DBD, DNA binding domain;
Nub, N-terminal half of ubiquitin; PM, plasma membrane; TAD, transcriptional activator domain; TF, transcription factor; UAS, upstream activation sequence; UBP,
ubiquitin-specific protease.
Review Trends in Pharmacological Sciences Vol.30 No.2
GPCRs [10,11]. Protein micro-arrays were designed todetect proteins directly interacting with the C terminusof GPCRs, in particular with well-known protein scaffold-ing modules such as PSD-95/disc large/zonula occludens 1(PDZ) domains [12–15] (Figures 1 and 2; Tables 1 and 2).Fusion of the receptor C terminus to the glutathione-S-transferase (GST) has also been applied to GAPC identi-fication from tissues lysates as shown by studies on the 5-HT2C receptor [16]. However, owing to the high number ofnon-specifically retained proteins, additional approachesbased on short, synthetic peptides have been subsequentlydeveloped. For example, peptides comprising known or
predicted PDZ-domain recognition motifs have been suc-cessfully used to identify GAPCs of 5-HT2A, 5-HT2C and 5-HT4 receptors by affinity chromatography [17,18].Obviously, these two approaches are designed to answertwo different questions. Whereas the entire C terminuswas used to identify all C-terminus-associated complexes,shorter peptides were used to specifically identify PDZ-domain-containing proteins. The high purity of chemicallysynthesized baits constitutes an important advantage ofthe peptide-based methods compared with the methodbased on GST fusion proteins. Indeed, the amount ofnon-specifically retained proteins is high in the GST-based
73
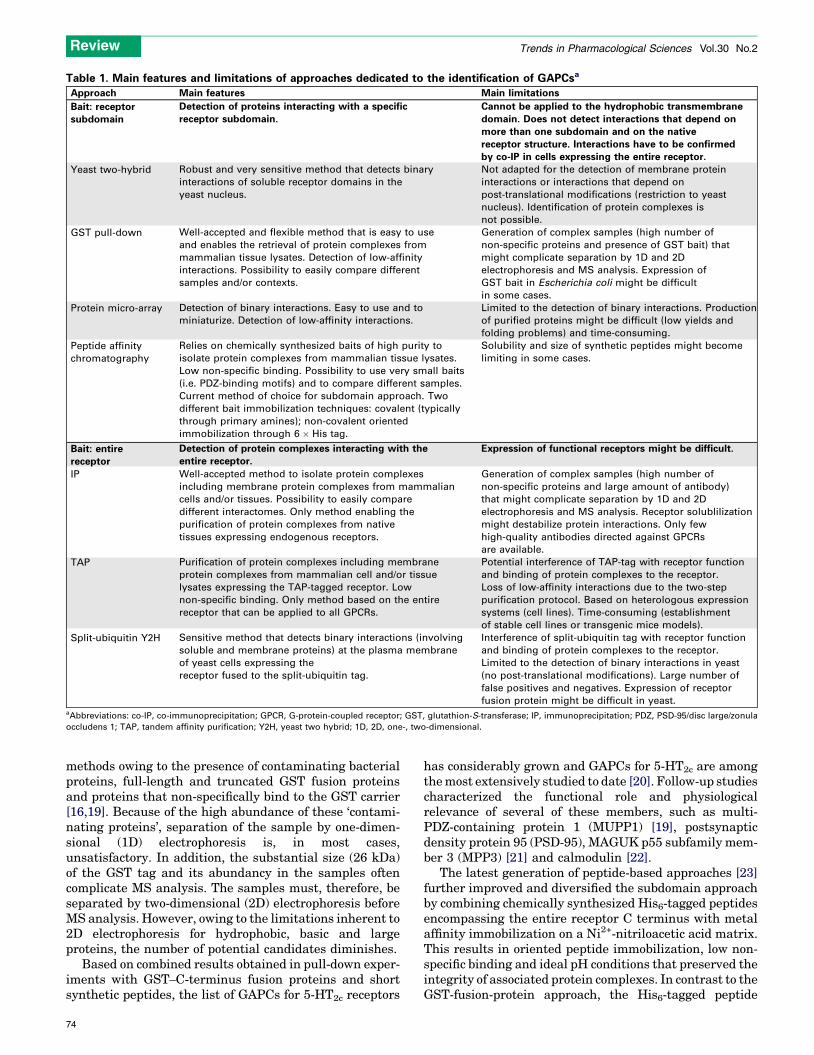
Table 1. Main features and limitations of approaches dedicated to the identification of GAPCsa
Approach Main features Main limitations
Bait: receptorsubdomain
Detection of proteins interacting with a specific
receptor subdomain.
Cannot be applied to the hydrophobic transmembrane
domain. Does not detect interactions that depend on
more than one subdomain and on the native
receptor structure. Interactions have to be confirmed
by co-IP in cells expressing the entire receptor.
Yeast two-hybrid Robust and very sensitive method that detects binary
interactions of soluble receptor domains in the
yeast nucleus.
Not adapted for the detection of membrane protein
interactions or interactions that depend on
post-translational modifications (restriction to yeast
nucleus). Identification of protein complexes is
not possible.
GST pull-down Well-accepted and flexible method that is easy to use
and enables the retrieval of protein complexes from
mammalian tissue lysates. Detection of low-affinity
interactions. Possibility to easily compare different
samples and/or contexts.
Generation of complex samples (high number of
non-specific proteins and presence of GST bait) that
might complicate separation by 1D and 2D
electrophoresis and MS analysis. Expression of
GST bait in Escherichia coli might be difficult
in some cases.
Protein micro-array Detection of binary interactions. Easy to use and to
miniaturize. Detection of low-affinity interactions.
Limited to the detection of binary interactions. Production
of purified proteins might be difficult (low yields and
folding problems) and time-consuming.
Peptide affinity
chromatography
Relies on chemically synthesized baits of high purity to
isolate protein complexes from mammalian tissue lysates.
Low non-specific binding. Possibility to use very small baits
(i.e. PDZ-binding motifs) and to compare different samples.
Current method of choice for subdomain approach. Two
different bait immobilization techniques: covalent (typically
through primary amines); non-covalent oriented
immobilization through 6 � His tag.
Solubility and size of synthetic peptides might become
limiting in some cases.
Bait: entire
receptor
Detection of protein complexes interacting with the
entire receptor.
Expression of functional receptors might be difficult.
IP Well-accepted method to isolate protein complexes
including membrane protein complexes from mammalian
cells and/or tissues. Possibility to easily compare
different interactomes. Only method enabling the
purification of protein complexes from native
tissues expressing endogenous receptors.
Generation of complex samples (high number of
non-specific proteins and large amount of antibody)
that might complicate separation by 1D and 2D
electrophoresis and MS analysis. Receptor solublilization
might destabilize protein interactions. Only few
high-quality antibodies directed against GPCRs
are available.
TAP Purification of protein complexes including membrane
protein complexes from mammalian cell and/or tissue
lysates expressing the TAP-tagged receptor. Low
non-specific binding. Only method based on the entire
receptor that can be applied to all GPCRs.
Potential interference of TAP-tag with receptor function
and binding of protein complexes to the receptor.
Loss of low-affinity interactions due to the two-step
purification protocol. Based on heterologous expression
systems (cell lines). Time-consuming (establishment
of stable cell lines or transgenic mice models).
Split-ubiquitin Y2H Sensitive method that detects binary interactions (involving
soluble and membrane proteins) at the plasma membrane
of yeast cells expressing the
receptor fused to the split-ubiquitin tag.
Interference of split-ubiquitin tag with receptor function
and binding of protein complexes to the receptor.
Limited to the detection of binary interactions in yeast
(no post-translational modifications). Large number of
false positives and negatives. Expression of receptor
fusion protein might be difficult in yeast.aAbbreviations: co-IP, co-immunoprecipitation; GPCR, G-protein-coupled receptor; GST, glutathion-S-transferase; IP, immunoprecipitation; PDZ, PSD-95/disc large/zonula
occludens 1; TAP, tandem affinity purification; Y2H, yeast two hybrid; 1D, 2D, one-, two-dimensional.
Review Trends in Pharmacological Sciences Vol.30 No.2
methods owing to the presence of contaminating bacterialproteins, full-length and truncated GST fusion proteinsand proteins that non-specifically bind to the GST carrier[16,19]. Because of the high abundance of these ‘contami-nating proteins’, separation of the sample by one-dimen-sional (1D) electrophoresis is, in most cases,unsatisfactory. In addition, the substantial size (26 kDa)of the GST tag and its abundancy in the samples oftencomplicate MS analysis. The samples must, therefore, beseparated by two-dimensional (2D) electrophoresis beforeMS analysis. However, owing to the limitations inherent to2D electrophoresis for hydrophobic, basic and largeproteins, the number of potential candidates diminishes.
Based on combined results obtained in pull-down exper-iments with GST–C-terminus fusion proteins and shortsynthetic peptides, the list of GAPCs for 5-HT2c receptors
74
has considerably grown and GAPCs for 5-HT2c are amongthemost extensively studied to date [20]. Follow-up studiescharacterized the functional role and physiologicalrelevance of several of these members, such as multi-PDZ-containing protein 1 (MUPP1) [19], postsynapticdensity protein 95 (PSD-95), MAGUK p55 subfamily mem-ber 3 (MPP3) [21] and calmodulin [22].
The latest generation of peptide-based approaches [23]further improved and diversified the subdomain approachby combining chemically synthesized His6-tagged peptidesencompassing the entire receptor C terminus with metalaffinity immobilization on a Ni2+-nitriloacetic acid matrix.This results in oriented peptide immobilization, low non-specific binding and ideal pH conditions that preserved theintegrity of associated protein complexes. In contrast to theGST-fusion-protein approach, the His6-tagged peptide

Figure 2. Biochemical methods to isolate GAPCs. Five different isolation procedures are compared on the level of the bait used, the source of interaction partners, the
purification protocol and the detection methods used. Abbreviations: GST, glutathione-S-transferase; His6, tag composed of six histidine amino acid residues; IP,
immunoprecipitation; MS, mass spectrometry; PM, plasma membrane; TAP, tandem affinity purification; TEV, Tobacco Etch Virus.
Review Trends in Pharmacological Sciences Vol.30 No.2
approach generates much less complex samples and can,therefore, easily be combined with 1D and 2D gel electro-phoresis to separate proteins, which considerablyincreases the number of identified interacting partners.
Table 2. Comparison of approaches dedicated to the identification
Y2H Split-ubiquitin
Y2H
IP
Bait type Soluble
subdomain
Entire tagged
receptor
Entire wild-
receptor
Detectable Interactions Binary Binary Protein
complexes
Cellular context Yeast cell Yeast cell Mammalian
and tissues
Post-translational
modifications
�b � +
Possibility to detect membrane
protein
� + +
Background (false positive and
negatives)
High High High
Confirmation by co-IP required + + �Identification of interactomes
upon agonist stimulation
� + +
Comparison of interactomes
from different cells and/or
tissues
� � +
aAbbreviations: co-IP, co-immunoprecipitation; GST, glutathion-S-transferase; IP, immub+, yes; �, no.cThe bait is not post-translationally modified but cell lysate might contain post-translatdSynthetic peptides could be chemically modified and serve as bait.
The advantage of combining 1D and 2D electrophoresis forsystematic identification of GAPCs is illustrated by ourrecent study using the C termini of themelatonin receptorsMT1 and MT2 as baits [23]. Among the 40 proteins ident-
of GAPCsa
TAP GST pull-down Peptide affinity
chromatography
Protein
micro-array
type Entire tagged
receptor
Soluble
subdomain
Soluble
subdomain
Soluble
subdomain
Protein
complexes
Protein
complexes
Protein
complexes
Binary
cells Mammalian
cells (tissues)
Mammalian
tissue lysate
Mammalian
tissue lysate
Purified
protein
+ +/�c +/�c,d �
+ � � �
Low High Low Low
� + + +
+ � � �
+ + + +
noprecipitation; TAP, tandem affinity purification; Y2H, yeast two hybrid.
ional modifications.
75

Review Trends in Pharmacological Sciences Vol.30 No.2
ified for the MT1 C terminus, 30 were obtained from 2Delectrophoresis and an additional 10 from 1D electropho-resis. For the MT2 C terminus, 11 proteins were identifiedfrom 2D electrophoresis and an additional 11 from 1Delectrophoresis. This clearly indicates that the use of only2D electrophoresis in previous studies led to an under-estimation ofmultiprotein complexes associatedwith theCterminus of GPCRs. Depending on the sample complexityand the presence of abundant proteins thatmightmask theidentification of less abundant proteins, trypsin digestionin solution (combined with liquid chromatography andmass spectrometry identification) can be an alternativeto separation by 1D and 2D electrophoresis. Identificationof ‘expected’ interaction partners such as Gi proteins andG-protein-coupled-receptor kinases 2 and 3 validatedinteraction partners identified with our approach anddemonstrated its sensitivity [23]. The functional relevanceof several newly identified interaction partners such astubulin [24], MUPP1 [11] and the regulator of G-proteinsignaling (RGS) 20, also called RGSZ1 [23], has also beshown.
In conclusion, substantial progress has been made indefining the GAPCs of several GPCRs using soluble re-ceptor subdomains as bait. Using only the soluble subdo-mains overcomes several technical difficulties associatedwith the hydrophobic transmembrane core domain ofGPCRs, and a specific subset of GAPCs interacting withthe selected subdomain or interaction motif can be rapidlyidentified. However, GAPCs identified by subdomainapproaches must be confirmed by other methods usingintact cells and entire receptors (Tables 1 and 2). Further-more, binding of GAPCs to receptors can depend on thepresence of multiple subdomains or conformational deter-minants that are not provided by a single subdomain. Toidentify theseGAPCs, complementary techniques based onthe use of entire receptors have been recently developed.
Identification of GAPCs using entire GPCRsIdeally, GAPCs are recovered and identified from nativetissue samples expressing the wild-type GPCR of interestin its natural environment and at physiological expressionlevels. Co-immunoprecipitation (IP) of GAPCs with recep-tor-specific antibodies currently represents the most suit-able approach to achieve this goal. Ideally, these antibodiesshould preferentially recognize epitopes located in theextracellular receptor domain to avoid interference withpotential GAPC binding sites in intracellular and trans-membrane receptor regions. Unfortunately, few high-affi-nity antibodies have been successfully developed forGPCRs, and successful isolation of GAPCs by co-IP hasbeen reported only in a few cases, such as the mGlu5
receptor [25]. Co-IPs are based on a one-step purification,which generates rich samples with high levels of non-specific proteins. However, the presence of large amountsof antibody in the eluate might interfere with MS analysisand the detection of low-abundant proteins.
The recently described TAP method overcomes many ofthese limitations while preserving the advantages of usingentire GPCRs as bait compared with isolated receptordomains. Moreover, the two-step purification greatlyreduces the amount of non-specific interacting proteins,
76
and no antibody is required for the purification of theGPCR and its GAPCs, thus decreasing interference withMS analysis. However, the wild-type receptors must bemodified with the TAP tag. The TAP method, initiallydesigned for large-scale purification of soluble proteincomplexes [2,26], was recently adapted to the specificneeds of GPCRs [8]. The TAP method relies on the expres-sion of the TAP-tagged target protein in the host cell. Theoriginal TAP tag consists of two immunoglobulin (IgG)binding units of protein A from Staphylococcus aureus, acleavage site for the Tobacco Etch Virus (TEV) proteaseand a calmodulin-binding peptide. The protein of interestand its associated protein complexes are recovered in atwo-step purification protocol that involves, first, bindingto IgG-coated sepharose beads and subsequent cleavage bythe TEV protease at the Glu-Xaa-Xaa-Tyr-Xaa-Gln/Serconsensus sequence and, second, the eluted complex isthen immobilized on calmodulin-coated beads in the pre-sence of calcium and finally eluted with ethylene glycoltetraacetic acid.
The TAP method has been successfully applied to MT1
and MT2 melatonin receptors to isolate GAPCs fromhuman embryonic kidney 293 cells stably expressing C-terminally TAP-tagged receptors [8]. After ensuring thatthe TAP tag did not affect subcellular localization, agonistaffinity and signaling capacity of these two TAP-taggedreceptors and optimizing the conditions of receptor solu-bilization and purification, an overall purification yield of15–30% was achieved for these two GPCRs. The co-puri-fication and MS identification of Gi proteins demonstratedthat the purification conditions were compatible with therecovery of ‘weakly’ associated GAPCs, as the receptor–G-protein interaction is usually considered to be fragile. Morerecently, GAPCs of theN-terminally TAP-tagged a1D-adre-noceptor was successfully identified [27]. Among theseproteins, the dystrophin proteins syntrophin, dystrobrevinand utrophin were of particular interest because binding ofthese proteins to the a1D-adrenoceptor was essential forexpression of functional receptors at the cell surface anda1D-adrenoceptor-associated regulation of blood pressure.Purification of TAP-tagged GPCRs has also been used tocompare GAPCs of the wild-type and Leu148Ser mutant(located within the intracellular loop 2) of the orphanGPR54 receptor, which is associated with idiopathic hypo-gonadotropic hypogonadism [28].
Taking together all currently available data on GAPCsidentified by the TAP approach, it is interesting to notethat recovered proteins are not only limited to thoseinvolved in signal transduction but also include proteincomplexes involved in receptor biosynthesis and traffick-ing, further strengthening the idea that GPCRs are accom-panied by GAPCs at each step of their life cycle.
Several future developments and applications of theTAP approach could be envisioned in the context of identi-fication and characterization of GAPCs. For example,improved versions of the TAPmethod have been developedto increase the purification yield in mammalian cells byintroducing tags of smaller size and higher avidity [29–31]and to stabilize the protein complexes by in vivo cross-linking [32,33]. Separation of subcellular compartmentsbefore the purification of complexes by the TAP approach

Review Trends in Pharmacological Sciences Vol.30 No.2
might help in the understanding of the spatial distributionof GAPCs. An interesting, but so far unexplored, feature isthe possibility to isolate complexes from unstimulated andstimulated cells. Comparison of different conditions alsoimplies the use of quantitative proteomics by labelingproteins or peptides with stable isotopes whose differencesin mass and peak intensities can be easily detected by MS(for reviews, see Refs [34,35]). Analysis of differentiallyrecruited GAPCs might give important insights into thefirst steps of GPCR-promoted signal transduction.
Furthermore, knock-in mice expressing TAP-taggedGPCRs are likely to become very interesting experimentalmodels for the isolation of GAPCs in vivo as shown forsoluble proteins [36,37]. Indeed, one of the advantages ofisolating GAPCs from their physiological context in livinganimals is the possibility of identifying specific inter-actions that might not be present in heterologous cell-culture preparations. In addition, the application of theTAP method to transgenic mice and to in vivo proteomicsmight offer the interesting possibility of studying tissue-specific GAPCs, GAPCs at different stages of developmentand GAPCs after in vivo pharmacological treatments.However, prior characterization of TAP-tagged GPCRsin cell lines is still required because it will provide valuableinformation on the solubilization and functionality of thesetagged receptors.
The flexibility of the TAP approach is further illustratedby the split-tag TAP version, in which the two tags arefused to two separate proteins [38], and was successfullyused for the identification of protein complexes associatedto the subunits UNC-29 and LEV-1 of the levamisole-sensitive nicotinic acetylcholine receptor ion channel inCaenorhabditis elegans [39]. This split-tag TAP versionmight be particularly useful in isolating GAPCs thatspecifically and differentially bind to GPCR homodimersversus heterodimers.
Further developments and new inputsThe first and crucial step for successful purification offunctional GAPCs is the solubilization of the receptor,which attempts to satisfy the diverging aims of solubilizingmaximal receptor amounts without destabilizing the re-ceptor structure and the integrity of GAPCs. Optimalconditions must be empirically determined in most casesfor each receptor. However, major progress in GPCR stabil-ization and solubilization, particularly due to recent effortsonGPCR crystallization, provides the scientific communitywith many different options [40].
Post-translational modifications such as phosphoryl-ation, sumoylation or ubiquitination are well-known trade-marks of GPCR activation. These reversible modificationscan occur either at the receptor level, its GAPCs or furtherdownstream in the signaling pathway, and analysis ofthese modifications by MS should become an importantcomponent of future research on GAPCs. The phosphoryl-ation state of proteins is usually determined after previousenrichment of phosphoproteins and/or phosphopeptides.The most commonly used enrichment approaches arebased onFe(III), Ga(III) or titaniumdioxide (TiO2) chroma-tography or IPs using anti-phosphotyrosine antibodies (forreview, see Ref. [35]). Such steps might be added at the end
of an IP or TAP protocol. Thus, instead of separating thefinal eluate by electrophoresis, the eluate might be directlysubmitted to trypsin digestion followed by the recovery ofphospho-peptides on TiO2 columns and identification ofretained peptides by MS [41].
The repertoire of available techniques might be furtherdiversified as shown by the recently described split-ubi-quitin membrane two-hybrid system, which was developedto overcome the limitations of the conventional yeast two-hybrid technology [42]. The system represents a fragmentcomplementation assay using the ubiquitin molecule splitinto C- and N-terminal halves as the interaction sensor,which was applied to monitor binary interactions betweenintegral membrane proteins and soluble proteins or be-tween two membrane proteins [9,43] (Figure 1). The split-ubiquitin system represents an important evolution of theclassical yeast two-hybrid system that relies on the assayrobustness and large-scale screening capacity of yeast-based assays. To date, this assay has only been appliedto the frizzled 1 receptor resulting in the identification of 16new interaction partners, which remain to be validated [9].
Concluding remarksThe classical view of GPCRs recruiting their cognate Gprotein upon receptor stimulation followed by the dis-sociation of the activated G protein into Ga and Gbg sub-units that diffuse to different effector systems has beenprogressively replaced by a dynamic model of preformedsignaling complexes with defined composition and stoichi-ometry [44]. In addition, these signaling complexes containfurther components such as adaptor proteins, proteinkinases and phosphatases and coexist with other com-plexes involved in receptor maturation, trafficking anddegradation. The most important challenge consists ofisolating and characterizing these receptor-proximal com-ponents of the cellular protein-interaction network.Clearly, a combination of different complementarymethods will be necessary to obtain a complete pictureof these networks. A careful analysis of the advantages anddrawbacks of the subdomain and the entire GPCRapproaches (TAPmethod) shows that both are complemen-tary rather than mutually exclusive. Whereas the entirereceptor approach enables the purification of protein com-plexes formed in intact cells, the subdomain approach ismore easily accessible for comparing the interactomes ofdifferent tissues and under different experimental con-ditions (different patho-physiological models or pharma-cological treatments) (Tables 1 and 2). Identification ofGAPCs might also provide valuable information on thefunction of the >100 orphan GPCRs, which remain notor-iously difficult to elucidate in the absence of a known ligand[45]. Interfering with the binding capacity of GAPCs toGPCRs by disruption or reinforcement might become a toolfor selective therapeutic intervention. Binding of GAPCs toGPCRs strongly depends on the cellular context andmight,thus, provide a new way for selective pharmacologicalintervention in one specific tissue without disturbing thereceptor function in another tissue. Intervention at thelevel of GAPC binding to the GPCR might also be aconvenient way to discriminate between closely relatedreceptor subtypes or splice variants of the same receptor
77

Review Trends in Pharmacological Sciences Vol.30 No.2
because differences in their intracellular domains areexpected to result in the binding of different GAPCs.
AcknowledgementsThis work was supported by grants from SERVIER, the FondationRecherche Medicale, Fondation pour la Recherche sur le CerveauNeurodon, Institut National de la Sante et de la Recherche Medicaleand Centre National de la Recherche Scientifique. A.M.D. holds an EGIDfellowship. We thank Patty Chen (Institut Cochin, Paris) for comments onthe manuscript.
References1 Lagerstrom, M.C. and Schioth, H.B. (2008) Structural diversity of G
protein-coupled receptors and significance for drug discovery.Nat. Rev.Drug Discov. 7, 339–357
2 Gavin, A.C. et al. (2002) Functional organization of the yeast proteomeby systematic analysis of protein complexes. Nature 415, 141–147
3 Bockaert, J. et al. (2004) GPCR interacting proteins (GIP). Pharmacol.Ther. 103, 203–221
4 Gavarini, S. et al. (2004) Molecular and functional characterization ofproteins interacting with the C-terminal domains of 5-HT2 receptors:emergence of 5-HT2 ‘receptosomes’. Biol. Cell 96, 373–381
5 Fagni, L. et al. (2004) Identification and functional roles ofmetabotropic glutamate receptor-interacting proteins. Semin. CellDev. Biol. 15, 289–298
6 Pluder, F. et al. (2006) Proteome analysis to study signal transductionof G protein-coupled receptors. Pharmacol. Ther. 112, 1–11
7 Enz, R. (2007) The trick of the tail: protein-protein interactions ofmetabotropic glutamate receptors. Bioessays 29, 60–73
8 Daulat, A.M. et al. (2007) Purification and identification of G protein-coupled receptor protein complexes under native conditions. Mol. Cell.Proteomics 6, 835–844
9 Dirnberger, D. et al. (2008) An optimized split-ubiquitin cDNA-libraryscreening system to identify novel interactors of the human Frizzled 1receptor. Nucleic Acids Res. 36, e37
10 Milojevic, T. et al. (2006) The ubiquitin-specific protease Usp4 regulatesthe cell surface level of the A2A receptor.Mol. Pharmacol. 69, 1083–1094
11 Guillaume, J.L. et al. (2008) The PDZ protein mupp1 promotes Gi
coupling and signaling of the Mt1 melatonin receptor. J. Biol. Chem.283, 16762–16771
12 Heydorn, A. et al. (2004) A library of 7TM receptor C-terminal tails.Interactions with the proposed post-endocytic sorting proteins ERM-binding phosphoprotein 50 (EBP50), N-ethylmaleimide-sensitive factor(NSF), sorting nexin 1 (SNX1), and G protein-coupled receptor-associated sorting protein (GASP). J. Biol. Chem. 279, 54291–54303
13 Fam, S.R. et al. (2005) P2Y1 receptor signaling is controlled byinteraction with the PDZ scaffold NHERF-2. Proc. Natl. Acad. Sci.U. S. A. 102, 8042–8047
14 Stiffler, M.A. et al. (2007) PDZ domain binding selectivity is optimizedacross the mouse proteome. Science 317, 364–369
15 Paquet, M. et al. (2006) The PDZ scaffold NHERF-2 interacts withmGluR5 and regulates receptor activity. J. Biol. Chem. 281, 29949–
2996116 Becamel, C. et al. (2002) Synaptic multiprotein complexes associated
with 5-HT2C receptors: a proteomic approach. EMBO J. 21, 2332–234217 Becamel, C. et al. (2004) The serotonin 5-HT2A and 5-HT2C receptors
interact with specific sets of PDZ proteins. J. Biol. Chem. 279, 20257–
2026618 Joubert, L. et al. (2004) New sorting nexin (SNX27) and NHERF
specifically interact with the 5-HT4a receptor splice variant: roles inreceptor targeting. J. Cell Sci. 117, 5367–5379
19 Becamel, C. et al. (2001) Interaction of serotonin 5-hydroxytryptaminetype 2C receptors with PDZ10 of the multi-PDZ domain proteinMUPP1. J. Biol. Chem. 276, 12974–12982
20 Bockaert, J. et al. (2004) GPCR-interacting proteins (GIPs): nature andfunctions. Biochem. Soc. Trans. 32, 851–855
21 Gavarini, S. et al. (2006) Opposite effects of PSD-95 and MPP3 PDZproteins on serotonin 5-hydroxytryptamine2C receptor desensitizationand membrane stability. Mol. Biol. Cell 17, 4619–4631
22 Labasque, M. et al. (2008) Physical interaction of calmodulin with the5-HT2C receptor C-terminus is essential for G Protein-independent,arrestin-dependent, receptor signaling. Mol. Biol. Cell 19, 4640–4650
78
23 Maurice, P. et al. (2008) A generic approach for the purification ofsignaling complexes that specifically interact with the carboxy-terminal domain of G protein-coupled receptors. Mol. Cell.Proteomics 7, 1556–1569
24 Bondi, C.D. et al. (2008) MT1 melatonin receptor internalizationunderlies melatonin-induced morphologic changes in Chinesehamster ovary cells and these processes are dependent on Gi
proteins, MEK 1/2 and microtubule modulation. J. Pineal Res. 44,288–298
25 Farr, C.D. et al. (2004) Proteomic analysis of native metabotropicglutamate receptor 5 protein complexes reveals novel molecularconstituents. J. Neurochem. 91, 438–450
26 Rigaut, G. et al. (1999) A generic protein purificationmethod for proteincomplex characterization and proteome exploration. Nat. Biotechnol.17, 1030–1032
27 Lyssand, J.S. et al. (2008) Blood pressure is regulated by an a1D-adrenergic receptor/dystrophin signalosome. J. Biol. Chem. 283,18792–18800
28 Wacker, J.L. et al. (2008) Disease causing mutation in GPR54 revealsimportance of second intracellular loop for class A GPCR function. J.Biol. Chem. 283, 31068–31078
29 Drakas, R. et al. (2004) Control of cell size through phosphorylation ofupstream binding factor 1 by nuclear phosphatidylinositol 3-kinase.Proc. Natl. Acad. Sci. U. S. A. 101, 9272–9276
30 Burckstummer, T. et al. (2006) An efficient tandem affinity purificationprocedure for interaction proteomics inmammalian cells.Nat.Methods3, 1013–1019
31 Gloeckner, C.J. et al. (2007) A novel tandem affinity purificationstrategy for the efficient isolation and characterisation of nativeprotein complexes. Proteomics 7, 4228–4234
32 Guerrero, C. et al. (2006) An integrated mass spectrometry-basedproteomic approach: quantitative analysis of tandem affinity-purifiedin vivo cross-linked protein complexes (QTAX) to decipher the 26 Sproteasome-interacting network. Mol. Cell. Proteomics 5, 366–378
33 Tagwerker, C. et al. (2006) A tandem affinity tag for two-steppurification under fully denaturing conditions: application inubiquitin profiling and protein complex identification combined within vivo cross-linking. Mol. Cell. Proteomics 5, 737–748
34 Gingras, A.C. et al. (2007) Analysis of protein complexes using massspectrometry. Nat. Rev. Mol. Cell Biol. 8, 645–654
35 Yang, W. et al. (2008) Proteomic approaches to the analysis ofmultiprotein signaling complexes. Proteomics 8, 832–851
36 Angrand, P.O. et al. (2006) Transgenicmouse proteomics identifies new14-3-3-associated proteins involved in cytoskeletal rearrangementsand cell signaling. Mol. Cell. Proteomics 5, 2211–2227
37 Mori, M. et al. (2008) Stable form of Jab1 enhances proliferation andmaintenance of hematopoietic progenitors. J. Biol. Chem. 283, 29011–
2902138 Dziembowski, A. and Seraphin, B. (2004) Recent developments in the
analysis of protein complexes. FEBS Lett. 556, 1–639 Gottschalk, A. et al. (2005) Identification and characterization of novel
nicotinic receptor-associated proteins in Caenorhabditis elegans.EMBO J. 24, 2566–2578
40 Pocanschi, C.L. et al. (2006) Amphipathic polymers: tools to foldintegral membrane proteins to their active form. Biochemistry 45,13954–13961
41 Collins, M.O. et al. (2005) Robust enrichment of phosphorylated speciesin complex mixtures by sequential protein and peptide metal-affinitychromatography and analysis by tandem mass spectrometry. Sci.STKE 2005, pl6
42 Stagljar, I. and Fields, S. (2002) Analysis of membrane proteininteractions using yeast-based technologies. Trends Biochem. Sci.27, 559–563
43 Gisler, S.M. et al. (2008) Monitoring protein-protein interactionsbetween the mammalian integral membrane transporters and PDZ-interacting partners using a modified split-ubiquitin membrane yeasttwo-hybrid system. Mol. Cell. Proteomics 7, 1362–1377
44 Dupre, D.J. and Hebert, T.E. (2006) Biosynthesis and trafficking ofseven transmembrane receptor signalling complexes. Cell. Signal. 18,1549–1559
45 Levoye, A. et al. (2006) Do orphan G-protein-coupled receptors haveligand-independent functions? New insights from receptorheterodimers. EMBO Rep. 7, 1094–1098





























