Embed Size (px)
Citation preview

Rapid Effects of Triiodothyronine onImmediate-Early Gene Expression in
Schwann CellsGILLES MERCIER,* NATHALIE TURQUE, AND MICHAEL SCHUMACHER
U488 INSERM, Kremlin-Bicetre, France
KEY WORDS triiodothyronine; Schwann cells; transcription factors; myelin;Krox-20
ABSTRACT In the peripheral nervous system, triiodothyronine (T3) plays an im-portant role in the development and regeneration of nerve fibers and in myelin forma-tion. However, the target genes of T3 in peripheral nerves remain to be identified. Weinvestigated whether T3 activated genes of transcription factors in Schwann cells.Expression of egr-1 (krox-24), egr-2 (krox-20), egr-3, c-jun, junB, c-fos, fosB, fra-1, fra-2,and CREB genes was analyzed by reverse transcription-polymerase chain reaction(RT-PCR) in Schwann cells isolated from neonatal rat sciatic nerves and in the cell linesMSC-80 (mouse Schwann cells), NIH-3T3 (mouse fibroblasts), and CHO (Chinese ham-ster ovary cells). Some of these transcription factors have been shown to be involved inSchwann cell differentiation. T3 triggered a rapid (15–30 min), transient (1–2-h) andstrong (6- to 15-fold) stimulation of Egr-1, Egr-2, Egr-3, Jun B, c-Fos, and Fos B mRNAexpression in Schwann cells. In contrast, expression of c-Jun, Fra-1, Fra-2, and CREBmRNA was not affected by T3. The stimulatory effects of T3 could be abolished by addingactinomycin D. T3 triggered the same pattern of gene stimulation in the mouse Schwanncell line MSC80, but not in the NIH-3T3 and CHO cell lines. Serum activated all thegenes that responded to T3 and in addition fra-1 and fra-2, but not c-jun and CREB.Immunoblotting showed that the increase in Egr-1 and c-Fos mRNA levels was accom-panied by an increase in the corresponding proteins. In addition, shifts of the proteinbands indicated a posttranslational modification of the two proteins. These effects of T3are likely to be mediated by the intracellular T3 receptor, as the D-isomer RT3 and T0,which do not bind to T3 receptors, proved ineffective. The present data suggested that T3may regulate Schwann cell functions and differentiation by transiently activating theexpression of specific transcription factors. GLIA 35:81–89, 2001. © 2001 Wiley-Liss, Inc.
INTRODUCTION
Thyroid hormone plays an important role in neuro-nal maturation and in myelination (Legrand, 1986;Pollard, 1993; Chan and Kilby, 2000). Myelinating glialcells of the central and peripheral nervous system haverecently been shown to be targets for triiodothyronine(T3) and to express nuclear thyroid hormone receptors(TR). TR are encoded by two distinct genes, designatedTRa and TRb. Alternative splicing from each genegenerates multiple TR isoforms: TRa1, TRa2, andTRa3 from the TRa gene and TRb1 and TRb2 from theTRb gene (Oppenheimer, 1999; Zhang and Lazar,2000). In the central nervous system, TRa1 and TRa2
are expressed at all stages of oligodendrocyte matura-tion, whereas TRb1 is only present in postmitotic oli-godendrocytes (Baas et al., 1994; Carre et al., 1998). Infact, TRb1 has been proposed to play an important rolein oligodendroglial differentiation by inhibiting theproliferation of precursor cells and by triggering theirmaturation (Barres et al., 1994; Baas et al., 1997).
Grant sponsor: Association Francaise Contre les Myopathies (A.F.M.); Grantsponsor: Myelin Project (USA); Grant sponsor: Projet Myeline (France).
*Correspondence to: Gilles Mercier, U488 INSERM, 80, rue du GeneralLeclerc, 94276 Kremlin-Bicetre, France. E-mail: [email protected]
Received 29 December 2000; Accepted 20 April 2001
Published online 00 Month 2001
GLIA 35:81–89 (2001)
© 2001 Wiley-Liss, Inc.

Thyroid hormone is most likely not a survival factor formature oligodendrocytes, but in excess it increases thenumber of apoptotic cells (Marta et al., 1998).
In cranial nerves, T3 has been shown to acceleratemyelin gene expression in a coordinated manner bothin the distal part, which is myelinated by oligodendro-cytes, and in the proximal part, which is myelinated bySchwann cells (Knipper et al., 1998). As shown by theuse of selective antisera, Schwann cells mainly expressTRa1 (Glauser and Barakat, 1997). In the rat sciaticnerve, TR are expressed only during early postnatallife. However, adult Schwann cells exhibit TR immu-noreactivity again after nerve injury, when they aredeprived of axonal contact (Barakat-Walter et al.,1993; Barakat-Walter, 1999). These observations areconsistent with the finding that during developmentand regeneration thyroid hormone promotes axonalgrowth and myelination in the peripheral nervous sys-tem. Thirty years ago, it was reported that the diame-ter of axons and the number of myelinated fibers aredecreased in the rat sciatic nerve as a consequence ofneonatal hypothyroidism (Clos and Legrand, 1970; Pol-lard, 1993). Subsequently, it has been demonstratedthat treatment of rats with thyroid hormone acceler-ates the regeneration of nerve fibers (McIsaac and Ki-ernan, 1975; Barakat-Walter, 1999). Local administra-tion of a high dose of thyroid hormone in siliconeregeneration chambers was shown to promote regener-ation of the transected sciatic nerve by increasing thediameter of axons and the thickness of the newlyformed myelin sheaths and by improving the recoveryof sensory and motor functions (Voinesco et al., 1998).Interestingly, hypothyroidism has been involved in thedevelopment of peripheral neuropathies in humans(Perkins and Morgenlander, 1997).
There are several mechanisms by which actions ofthyroid hormone on Schwann cells may acceleratethe development and regeneration of peripheralnerves. First, T3 may have a role in Schwann cellproliferation. Indeed, T3 increases the incorporationof [3H]thymidine in cultured rat Schwann cells, andthe proliferation of Schwann cells is reduced in thesciatic nerve of hypothyroid chick embryos (Ussonand Saxod, 1988; Barakat-Walter, 1999). Second, T3may stimulate the production of neurotrophic factorsby Schwann cells, resulting in accelerated axonalgrowth. That is, T3 stimulates neurite outgrowthfrom dorsal root ganglia (DRG) neurons, but only ifthey are co-cultured with Schwann cells (Barakat-Walter and Riederer, 1996). Third, T3 may have adirect role in myelination, although this has still notbeen demonstrated experimentally. A role for T3 inmyelination is strongly suggested by the observationthat the hormone increases the expression of myelingenes and that a functional thyroid hormone re-sponse element (TRE) is present in the promoterregion of the myelin basic protein (MBP) gene, whichappears to be preferentially transactivated via TRb(Farsetti et al., 1992).
The multiple effects of thyroid hormone on Schwanncells are most likely mediated by other factors. A mech-anism by which T3 may regulate and coordinate vari-ous Schwann cell functions is by activating or repress-ing specific transcription factors involved in earlysignaling events. The study of the role of transcriptionfactors in the nervous system has only very recentlybecome an intense field of research (Herdegen andLeah, 1998). Several transcription factors have beenshown to play an important role in Schwann cell devel-opment (Zorick and Lemke, 1996; Topilko et al., 1997).This is particularly well documented by the transcrip-tion factor Egr-2 (Krox-20). Mice lacking the egr-2 genedo not form peripheral myelin and do not express anyof the major peripheral myelin proteins (Topilko et al.,1994). Schwann cell expression of Egr-2, and of othertranscription factors, such as c-Jun, is regulated byneuronal signals (De Felipe and Hunt, 1994; Stewart,1995; Murphy et al. 1996).
In the present study, using reverse transcription-polymerase chain reaction (RT-PCR), we have ana-lyzed the effect of T3 on the expression of 10 immedi-ate-early genes in cultured rat Schwann cells, namely,egr-1, egr-2, egr-3, junB, fosB, c-jun, c-fos, fra-1, fra-2,and CREB. Results show that T3 rapidly and tran-siently induces the expression of egr-1, egr-2, egr-3,junB, c-fos, and fosB, but not of the other studied tran-scription factors. In contrast to T3, both RT3 and T0,which do not bind to nuclear TR, were inactive. Inaddition, the pattern of transcription factor activationby T3 showed Schwann cell specificity.
MATERIALS AND METHODSCulture Conditions
Cultures of rat Schwann cells
Cultures of rat Schwann cells were prepared frompostnatal 3–4-day-old rats according to the method ofBrockes et al. (1979). Sciatic nerves were dissected ofconnective tissue, chopped, and dissociated by incubat-ing them for 20 min at 37°C in Dulbecco’s modifiedEagle’s medium (DMEM, Gibco-BRL) containing 0.25%trypsin and 0.1% collagenase type A (Sigma ChemicalCo.). Cells were collected by centrifugation and seededonto tissue culture flasks precoated with poly-L-lysine(Sigma) and cultured in DMEM supplemented with 4mM glutamine (Biomedia), 50 mg/L penicillin/strepto-mycin (Biomedia), and 10% fetal calf serum (FCS)(Gibco-BRL). After 48 h, 1025 cytosine arabinosine wasadded for 2 days for partial elimination of contaminat-ing fibroblasts. Schwann cells were further purified byanti-Thy1.1 monoclonal antibody and Low-Tox-M rab-bit complement (both from Cederlane). To stimulateSchwann cell growth, the medium was supplementedwith 5 mM forskolin (Calbiochem) and 4.1 mg/ml insulin(Organon) (Schumacher et al., 1993).
82 MERCIER ET AL.

Cell line cultures
Chinese hamster ovary (CHO), NIH 3T3 (mouse fi-broblasts), and MSC80 (mouse Schwann cells) weregrown in DMEM (Gibco-BRL) supplemented with 4mM glutamine (Biomedia), 50 mg /L penicillin/strepto-mycin (Biomedia), and 10% FCS (Gibco-BRL). MSC80is a murine Schwann cell line established by seruminduction (Boutry et al., 1992).
Treatment of Cells
For kinetic experiments, 106 cells were plated in 10-cmpetri dishes. After 2 days, cells were washed with phos-phate-buffered saline (PBS) and grown for 24 h inDMEM/Ham F12 (V/V) medium supplemented with 1.3mM insulin, 0.12 mM selenium, and 0.04 mM transferrin,before hormones or FCS were added. At different times,RNA and proteins were extracted from the cells.
Semiquantitative RT-PCR Experiments
Total RNA was extracted from cells with the QIA-GEN RNeasy mini kit and precipitated with ethanol.From 1mg of total RNA, first-strand cDNA was gener-ated using Moloney reverse transcriptase and reagentssupplied with the SuperScript II kit (Life Technologies)and random hexamer primers (Roche) under conditionsindicated by the manufacturer. The polymerase chainreactions (PCR) were carried out in a final volume of 50ml containing 2 ml of reverse transcriptase, 20 mMTris-HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl
2, 0.125
mCi a32P dCTP (Amersham), 0.1 mM of each primer(0.02 mM for S 26), 1 mM dNTP mix and 1 U of TaqDNA Polymerase (Life Technologies). Amplifications ofcDNA were performed for 1 min at 94°C, followed by 27cycles for 1 min at 94°C, 1 min at 60°C and 1 min at72°C strand completion was maximized by extendingthe final incubation at 72°C for an additional 4 min.
The primers we designed were the following:
egr-1Sense 59-AGAAGGCGATGGGTGGAGACGA-39Antisense 59 TGTGGATCTCTCTGGCACGG-39Size of fragment: 831
egr-3Sense 59-CGACTCGGTAGCCCATTACAATCA-39Antisense 59-TTGTAGTCAGGAATCATGGGGAA-AA-39Size of fragment: 464
c-junSense 59-GCCTGCACAGTGAGCCTCCG-39Antisense 59 AGTTGGCACCCACTGTTAAC-39Size of fragment: 489
junBSense 59-CCGGATGTGCACGAAAATGGAACAG-39Antisense 59-ACCGTCCGCAAAGCCCTCCTG-39Size of fragment: 408
CREBSense 59-CCAGCCACAGATTGCCACATTGC-39Antisense 59-CCCCTGGTGCATCAGAAGATAAGTC-39Size of fragment: 355
c-fosSense 59-TCCCAGAGGAGATGTCTGTG)-39Antisense 59 GGCTCCAGCTCTGTGACCAT-39Size of fragment: 409
fosBSense 59-TCATCACCCTCCGCCGAGTCTCAGT-39Antisense 59-GCTTGCCCCGCCAGTGCTGTAG-39Size of fragment: 305
fra-1Sense 59-GCGCTCTTCACGAAATCCAGCAG-39Antisense 59-TGGCCAATTGGGTTCACTGTCTG-AAG-39Size of fragment: 493
fra-2Sense 59-CTGCGCACGCCGAGTCCTACT-39Antisense 59-GATTCGACGCTTCTCCTCCTCCTCA-39Size of fragment: 304
S26Sense 59-AGGAGAAACAACGGTCGTTGCCAAAA-39Antisense 59-GCGCCAGCAGGTCTGAATCGTG-39Size of fragment: 305
Aliquots from the amplification products were re-solved on agarose gel and transferred onto a nitrocel-lulose membrane (Amersham) in 0.4 M NaOH. Bandswere quantified on an Instant Imager (Packard) andvisualized by autoradiography. Quantifications weremade by calculating the ratio of counts per minute(cpm) at each time to cpm of the corresponding control.The results were normalized by comparison with theribosomal S26 signal.
Western Blotting Experiments
Cells were resuspended in a lysis buffer composed of2% sodium dodecyl sulfate (SDS), 66 mM Tris pH 6.8,and 0.7 M b-mercaptoethanol 10% glycerol. The lysateswere boiled for 5 min and centrifuged for 10 min atmaximal speed in a minifuge. The determinations ofprotein concentrations were performed after the color-imetric method described by McKnight (1977).
A total of 20 mg of proteins for c-jun and c-fos immu-noblottings and 40 mg of proteins for Egr1 immunob-lottings was loaded and run on a denaturing 10% poly-acrylamide gel. Proteins were transferred onto a PVDFmembrane (NEN). The membranes were blocked in 5%dry milk in PBS-0.1% Tween and then incubated withantibodies diluted 1:2,000. The three antibodies arerabbit polyclonal antibodies purchased from SantaCruz: anti-Egr-1 antibody (sc-189), anti-c-jun antibody(sc-45-G), and anti-c-Fos antibody (sc-52-G). The sec-ondary antibody used in 1:30,000 dilution, was pur-chased from Vector (PI-1000). Revelation was per-formed by chemoluminescence (Amersham). Blots were
83T3 EFFECTS ON SCHWANN CELLS
developed with an ECL kit (Amersham) and detectedon Kodak film.
RESULTS.Effects of T3 on Immediate-Early Genes in
Schwann Cells
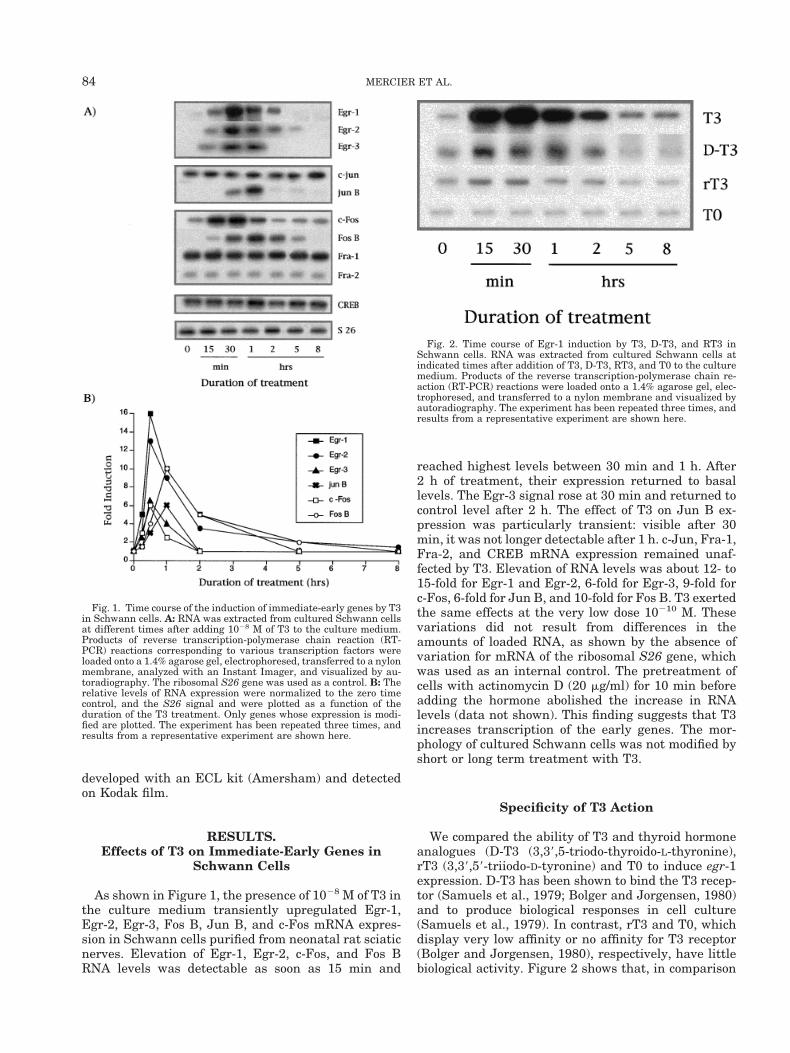
As shown in Figure 1, the presence of 1028 M of T3 inthe culture medium transiently upregulated Egr-1,Egr-2, Egr-3, Fos B, Jun B, and c-Fos mRNA expres-sion in Schwann cells purified from neonatal rat sciaticnerves. Elevation of Egr-1, Egr-2, c-Fos, and Fos BRNA levels was detectable as soon as 15 min and
reached highest levels between 30 min and 1 h. After2 h of treatment, their expression returned to basallevels. The Egr-3 signal rose at 30 min and returned tocontrol level after 2 h. The effect of T3 on Jun B ex-pression was particularly transient: visible after 30min, it was not longer detectable after 1 h. c-Jun, Fra-1,Fra-2, and CREB mRNA expression remained unaf-fected by T3. Elevation of RNA levels was about 12- to15-fold for Egr-1 and Egr-2, 6-fold for Egr-3, 9-fold forc-Fos, 6-fold for Jun B, and 10-fold for Fos B. T3 exertedthe same effects at the very low dose 10210 M. Thesevariations did not result from differences in theamounts of loaded RNA, as shown by the absence ofvariation for mRNA of the ribosomal S26 gene, whichwas used as an internal control. The pretreatment ofcells with actinomycin D (20 mg/ml) for 10 min beforeadding the hormone abolished the increase in RNAlevels (data not shown). This finding suggests that T3increases transcription of the early genes. The mor-phology of cultured Schwann cells was not modified byshort or long term treatment with T3.
Specificity of T3 Action
We compared the ability of T3 and thyroid hormoneanalogues (D-T3 (3,39,5-triodo-thyroido-L-thyronine),rT3 (3,39,59-triiodo-D-tyronine) and T0 to induce egr-1expression. D-T3 has been shown to bind the T3 recep-tor (Samuels et al., 1979; Bolger and Jorgensen, 1980)and to produce biological responses in cell culture(Samuels et al., 1979). In contrast, rT3 and T0, whichdisplay very low affinity or no affinity for T3 receptor(Bolger and Jorgensen, 1980), respectively, have littlebiological activity. Figure 2 shows that, in comparison
Fig. 2. Time course of Egr-1 induction by T3, D-T3, and RT3 inSchwann cells. RNA was extracted from cultured Schwann cells atindicated times after addition of T3, D-T3, RT3, and T0 to the culturemedium. Products of the reverse transcription-polymerase chain re-action (RT-PCR) reactions were loaded onto a 1.4% agarose gel, elec-trophoresed, and transferred to a nylon membrane and visualized byautoradiography. The experiment has been repeated three times, andresults from a representative experiment are shown here.
Fig. 1. Time course of the induction of immediate-early genes by T3in Schwann cells. A: RNA was extracted from cultured Schwann cellsat different times after adding 1028 M of T3 to the culture medium.Products of reverse transcription-polymerase chain reaction (RT-PCR) reactions corresponding to various transcription factors wereloaded onto a 1.4% agarose gel, electrophoresed, transferred to a nylonmembrane, analyzed with an Instant Imager, and visualized by au-toradiography. The ribosomal S26 gene was used as a control. B: Therelative levels of RNA expression were normalized to the zero timecontrol, and the S26 signal and were plotted as a function of theduration of the T3 treatment. Only genes whose expression is modi-fied are plotted. The experiment has been repeated three times, andresults from a representative experiment are shown here.
84 MERCIER ET AL.
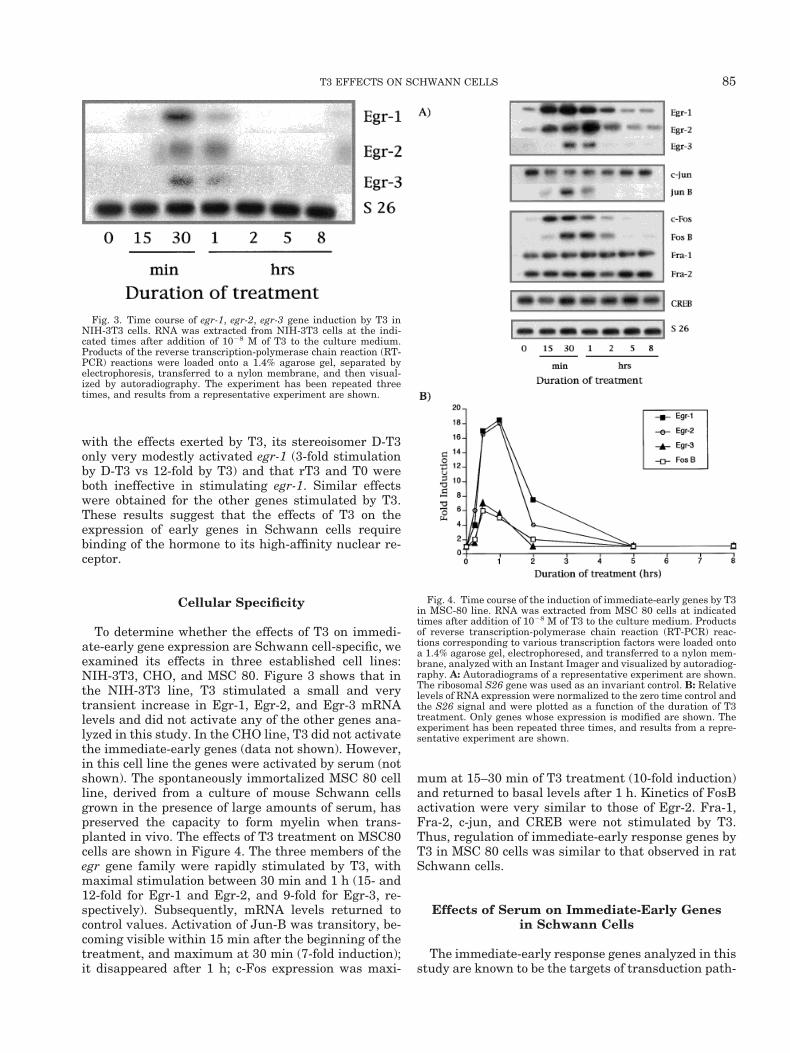
with the effects exerted by T3, its stereoisomer D-T3only very modestly activated egr-1 (3-fold stimulationby D-T3 vs 12-fold by T3) and that rT3 and T0 wereboth ineffective in stimulating egr-1. Similar effectswere obtained for the other genes stimulated by T3.These results suggest that the effects of T3 on theexpression of early genes in Schwann cells requirebinding of the hormone to its high-affinity nuclear re-ceptor.
Cellular Specificity
To determine whether the effects of T3 on immedi-ate-early gene expression are Schwann cell-specific, weexamined its effects in three established cell lines:NIH-3T3, CHO, and MSC 80. Figure 3 shows that inthe NIH-3T3 line, T3 stimulated a small and verytransient increase in Egr-1, Egr-2, and Egr-3 mRNAlevels and did not activate any of the other genes ana-lyzed in this study. In the CHO line, T3 did not activatethe immediate-early genes (data not shown). However,in this cell line the genes were activated by serum (notshown). The spontaneously immortalized MSC 80 cellline, derived from a culture of mouse Schwann cellsgrown in the presence of large amounts of serum, haspreserved the capacity to form myelin when trans-planted in vivo. The effects of T3 treatment on MSC80cells are shown in Figure 4. The three members of theegr gene family were rapidly stimulated by T3, withmaximal stimulation between 30 min and 1 h (15- and12-fold for Egr-1 and Egr-2, and 9-fold for Egr-3, re-spectively). Subsequently, mRNA levels returned tocontrol values. Activation of Jun-B was transitory, be-coming visible within 15 min after the beginning of thetreatment, and maximum at 30 min (7-fold induction);it disappeared after 1 h; c-Fos expression was maxi-
mum at 15–30 min of T3 treatment (10-fold induction)and returned to basal levels after 1 h. Kinetics of FosBactivation were very similar to those of Egr-2. Fra-1,Fra-2, c-jun, and CREB were not stimulated by T3.Thus, regulation of immediate-early response genes byT3 in MSC 80 cells was similar to that observed in ratSchwann cells.
Effects of Serum on Immediate-Early Genesin Schwann Cells
The immediate-early response genes analyzed in thisstudy are known to be the targets of transduction path-
Fig. 3. Time course of egr-1, egr-2, egr-3 gene induction by T3 inNIH-3T3 cells. RNA was extracted from NIH-3T3 cells at the indi-cated times after addition of 1028 M of T3 to the culture medium.Products of the reverse transcription-polymerase chain reaction (RT-PCR) reactions were loaded onto a 1.4% agarose gel, separated byelectrophoresis, transferred to a nylon membrane, and then visual-ized by autoradiography. The experiment has been repeated threetimes, and results from a representative experiment are shown.
Fig. 4. Time course of the induction of immediate-early genes by T3in MSC-80 line. RNA was extracted from MSC 80 cells at indicatedtimes after addition of 1028 M of T3 to the culture medium. Productsof reverse transcription-polymerase chain reaction (RT-PCR) reac-tions corresponding to various transcription factors were loaded ontoa 1.4% agarose gel, electrophoresed, and transferred to a nylon mem-brane, analyzed with an Instant Imager and visualized by autoradiog-raphy. A: Autoradiograms of a representative experiment are shown.The ribosomal S26 gene was used as an invariant control. B: Relativelevels of RNA expression were normalized to the zero time control andthe S26 signal and were plotted as a function of the duration of T3treatment. Only genes whose expression is modified are shown. Theexperiment has been repeated three times, and results from a repre-sentative experiment are shown.
85T3 EFFECTS ON SCHWANN CELLS

ways stimulated by serum and growth factors. We de-termined whether the pattern of gene activation byserum was different from that obtained with T3. Figure5 shows that the addition of FCS to quiescent cellstriggered the stimulation of all the genes studied, ex-cept for c-jun and CREB. The amplitude of the stimu-lation was higher with serum than with T3 (cf. Fig. 1):Fos B (40-fold by serum vs 10-fold by T3), Jun B (25-fold by serum vs 6-fold by T3), Egr-1 (37-fold by serumvs 16-fold by T3), Egr-2 (27-fold by serum vs 12-fold byT3), and Egr-3 (17-fold by serum vs 9-fold by T3). Fra-1and Fra-2, which were unresponsive to T3, were acti-vated by serum (7-fold for Fra-1 and 3-fold for Fra-2).
Unlike these genes, c-fos expression was only weaklystimulated (4-fold by serum versus 10-fold by T3).
egr1, c-jun, and c-fos Immunoblotting
In order to determine whether the variations in RNAlevels, observed in Schwann cells, were followed byequivalent variations at the protein level, we immuno-precipitated three proteins—Egr-1, c-Jun, and c-Fos—with specific antibodies and analyzed the immunopre-cipitated products by immunoblotting. In quiescentcells treated with serum, the anti-Egr-1 antiserum de-tected a very faint band (80 kD) after 30 min (Fig. 6).The signal increased very sharply, reaching a peak at1 h and then rapidly decreased, no signal being detect-able after 2 h. In T3 treated cells, the antibody detecteda strong band as soon as 30 min. The disappearance ofthe Egr-1 signal was accompanied by a shift in migra-tion. This shift was much more marked in cells treatedwith T3 after 1 h and 2 h than in cells treated withserum for 2 h. The origins of these differences in themigration profile remain to be elucidated. The anti-c-Jun antibody detected a high level of 42–44 kD proteinin untreated cells but did not detect any difference inthe c-jun protein level during T3 and serum treatment.The anti-c-Fos antibody showed that quiescent cellsexpress a relatively high level of c-Fos protein (60–62kD). The intensity of the signal increased within 30min after T3 or serum treatment, peaked at 1 h, andreturned to basal levels after 2 h. Unlike Egr-1, a shiftin migration was only observed with serum-treated
Fig. 5. Time course of the induction of immediate-early genes byserum in Schwann cells. RNA was extracted from cultured Schwanncells different times after treatment with serum. Products of thereverse transcription-polymerase chain reaction (RT-PCR) reactions,corresponding to different transcription factors, were loaded onto a1.4% agarose gel, electrophoresed and transferred to a nylon mem-brane, analyzed with an Instant Imager, and visualized by autora-diography. A: The autoradiogram of a representative experiment isshown. The ribosomal S26 gene was used as a control. B: The relativelevels of RNA expression normalized to the zero time control and theS26 signal were plotted as a function of the duration of serum treat-ment. Only genes whose expression is modified are shown. The ex-periment has been repeated three times, and results from a represen-tative experiment are shown.
Fig. 6. Immunoblot analysis of the time course of Egr-1, c-Jun, andc-Fos protein induction by T3 and serum in Schwann cells. Schwanncells were cultured in defined medium for 24 h and then treated withT3 (1028 M) or serum (10%) for different times. Total cellular proteinswere extracted and subjected to immunoblot analysis as described inMaterials and Methods. The samples were blotted with anti-Egr-1,anti-c-Jun, and anti-c-Fos polyclonal antibodies. The experiment hasbeen repeated three times, and results from a representative experi-ment are shown.
86 MERCIER ET AL.

cells. Similar patterns of Egr-1, c-Jun, and c-Fos pro-tein expression were obtained with MSC 80 cells (notshown). Thus, at least for these three genes, the vari-ation in RNA levels correlated with the variations ofprotein levels.
DISCUSSION
The systematic search for thyroid hormone-sensitivetranscription factors in Schwann cells represents afirst step in elucidating the mechanisms of action of thehormone and in identifying its target genes. The re-sults of the present study show that in cultured ratSchwann cells, T3 causes a marked rapid and transientincrease in mRNA levels of Egr-1, Egr-2, Egr-3, Jun B,c-Fos, and Fos B, but not of c-Jun, Fra-1, Fra-2, andCREB. T3 also does not affect the expression of SCIPand Sox-10 (data not shown), both of which are in-volved in Schwann cell differentiation (Mirsky and Jes-sen, 1999). The increase in mRNA levels is likely toresult from stimulation of gene transcription, as itcould be blocked by actinomycin D.
Early response transcription factors can be inducedby a wide range of extracellular stimuli, including hor-mones, neurotransmitters, neuropeptides, growth fac-tors, and stress factors (Herdegen and Leah, 1998). Theobserved effects of T3 are very likely to be biologicallymeaningful and relevant for the understanding of thy-roid hormone action in Schwann cells. First, T3 in-duced a strong increase in the expression of specifictranscription factors, in particular those belonging tothe EGR family that have been shown to play an im-portant role in Schwann cell development, such asEgr-1 (Krox-24) and Egr-2 (Krox-20) (Topilko et al.,1994, 1997). Second, the compounds that bind the nu-clear receptors (T3 and D-T3), but not rT3 and T0,which do not bind to TR, induced transcription factorexpression. The effects of T3 are thus likely to be me-diated by binding to its cognate receptor. Third, theinduction of transcription factors by T3 showed cellspecificity. Indeed, the same profile of transcriptionfactor induction was observed in rat Schwann cellspurified from neonatal sciatic nerves and in the mouseSchwann cell line MSC80, but not in the CHO andNIH-3T3 cell lines. With regard to the cell-specific ac-tion of T3, the hormone induced c-fos expression inSchwann cells but has been reported to repress c-fospromoter activity in pituitary cells (Perez et al., 1994).The difference in the sensitivity of IEG to T3 inSchwann cells, MSC80, NIH-3T3, and CHO lines raisesthe question of the expression of various TR in thesecells. The problem is now under investigation.
The expression of Egr-1 in the developing brain isaffected by the thyroid status of the animal (Mellstromet al., 1994). More recently, it has been shown that theegr-1 gene contains a thyroid hormone response ele-ment in the promoter region and that its transcriptionis increased by T3 in the cerebellum of newborn mice(Ghorbel et al., 1999). The observation that T3 induced
both Egr-1 and Egr-2 mRNA is interesting because ithas been reported that the expression of both tran-scription factors in Schwann cells is successive andmutually exclusive during embryonic development andduring regeneration, with Krox-24 restricted to undif-ferentiated and proliferating Schwann cells, whereasKrox-20 is present in mature Schwann cells. However,both factors are coexpressed at birth in about 30% ofSchwann cells before the onset of myelination (Topilkoet al., 1997).
Thyroid hormone may not only affect the expressionof transcription factors in Schwann cells, it may alsoinfluence their activity via the regulation of posttrans-lational modifications. This is strongly suggested bythe shift observed in the migration of T3-inducedEgr-1. The cause of this change in the migration profileof Egr-1 remains to be elucidated, but it may corre-spond to a change in the phosphorylation of the pro-tein. Multiple forms of phosphorylated Egr-1 have beenobserved and phosphorylation and dephosphorylationof the Egr-1 protein may be involved in the regulationof its activity (Cao et al., 1990).
The expression of four transcription factors was notaffected by T3: three members of the fos gene family(c-Jun, Fra-1, Fra-2) and the cAMP response elementbinding protein (CREB). In spite the fact that c-Jun isa member of the inducible transcription factors (ITF), itwas found to be expressed at high levels in culturedSchwann cells, and its expression was not modified byserum. However, c-Jun expression was previouslyshown to be inhibited by cAMP and by axonal signals(De Felipe and Hunt, 1994; Stewart, 1995). In contrast,the inducible transcription factors Fra-1 and Fra-2were induced by serum. As expected, the constitutivetranscription factor CREB was not induced by serum.The transcriptional activity of CREB is regulated byphosphorylation via different signaling cascades(Herdegen and Leah, 1998). Neuron–glial interactionsalso influence the phosphorylation of CREB, which hasbeen related to Schwann cell proliferation (Taberneroet al., 1998; Lee et al., 1999). It would thus be inter-esting to determine whether T3 regulates phosphory-lation of this and other transcription factors inSchwann cells.
Most transcription factors form dimers, such as theleucine zipper proteins of the jun and fos families (Ka-rin et al., 1997). These dimers differ in their DNA-binding properties, stability, and transactivation po-tential. In particular, in addition to homodimerization,c-Jun displays a wide range of dimerization with mostmembers of the ITF and CTF families, including tran-scription factors that were analyzed in the presentstudy: Jun B, c-Fos, Fos B, Fra-1, and Fra-2 (for review,see Herdegen and Leah, 1998). Although the ability oftranscription factors to form heterodimers is more orless restricted, the number of possible combinations islarge and depends on the relative concentration of eachfactor. Consequently, modifications in the expression oftranscription factors by T3 may result in changes in
87T3 EFFECTS ON SCHWANN CELLS

dimer formation and, consequently, in the regulation ofdownstream genes.
Our results were obtained with a purified populationof Schwann cells. Such population is not encounteredin the nervous system, where Schwann cells interactwith different cell types. Thus, Schwann cells are notable to differentiate fully in the absence of axons. In thecentral nervous system, neuronal contacts have beenshown to inhibit astroglial cell growth and to be animportant mechanism controlling gene expression inprimary cultures of astrocytes (Pennypacker et al,1996). Nevertheless, purified primary cells are a goodmodel for studying their responsiveness to any sub-stance. All the effects of T3 we have described are fastand transient. The characteristic of immediate-earlytranscription factors is their rapid on/off kinetics, evenwhen the stimulation is continuous. Transient activa-tion of IEG has been shown to be involved in the dif-ferentiation of numerous cellular systems (for review,see Herdegen and Leah, 1998). They are the first tar-gets of stimuli regulating differentiation or adaptiveresponse to environmental conditions. Knockout ofsome of these transcription factors results in loss ofinducibility of genes (Hu et al, 1994), in addition tosevere defects (Johnson et al, 1992). Our results sug-gest that acceleration of development and regenerationof peripheral nerves by T3 may be mediated by thestimulation of a set of transcription factors. But theeffects of T3 on gene expression in the peripheral nerveduring development and regeneration depend on otherfactors. This could explain why despite being stimu-lated by T3, Egr-1 and Egr-2 each display a specificexpression pattern during development and regenera-tion, as well as why the inducibility of IEG by externalstimuli changes during development (Herdegen andLeah, 1998).
The important role of hormones during developmentand regeneration of the nervous system is increasinglyrecognized and represents an emerging area of re-search. In addition to thyroid hormone, steroid hor-mones play an important role in the regeneration ofperipheral nerves. Originally considered only repro-ductive or stress hormones, it is now well establishedthat they have much broader activities in the nervoussystem, where they regulate vital glial and neuronalfunctions (Bock and Goode, 1995; Schumacher et al.,1996). For example, progesterone, well known as ahormone involved in pregnancy, is synthesized bySchwann cells (Guennoun et al., 1997; Robert et al.,2001) and plays an important role in myelination, asdemonstrated in vivo and in co-cultures of neurons andSchwann cells (Koenig et al., 1995; Chan et al., 1998;Notterpek et al., 1999; Melcangi et al., 1998). The pos-sible interactions of thyroid and steroid hormones inSchwann cell function and their distinct and eventu-ally common target genes require investigation.
In summary, thyroid hormone is an important sig-naling molecule in the peripheral nervous system,where it acts on Schwann cells and promotes axonalgrowth and myelin formation. However, its target
genes still need to be identified. The present studyrepresents the first systematic investigation of T3 onthe expression of immediate-early genes in Schwanncells. By regulating the expression of these immediate-early genes, thyroid hormone may influence, in a coor-dinated manner, the expression of Schwann cell genesinvolved in proliferation, growth factor synthesis, dif-ferentiation, and myelin formation. Indeed, a transientincrease in immediate-early response genes can havelong-lasting effects on cell growth, survival, and differ-entiation.
ACKNOWLEDGMENTS
This work was supported in part by the AssociationFrancaise Contre les Myopathies (A.F.M.), by a post-doctoral fellowship from the Myelin Project (USA), andby Projet Myeline (France) (to N.T).
REFERENCES
Baas D, Bourbeau D, Carre JL, Sarlieve LL, Dussault JH, PuymiratJ. 1994. Expression of alpha and beta thyroid receptors duringoligodendrocyte differentiation. NeuroReport 5:1805–1808.
Baas D, Bourbeau D, Sarlieve LL, Ittel ME, Dussault JH, Puymirat J.1997. Oligodendrocyte maturation and progenitor cell proliferationare independently regulated by thyroid hormone. Glia 19:324–332.
Barres BA, Lazar MA, Raff MC. 1994. A novel role for thyroid hor-mone, glucocorticoids and retinoic acid in timing oligodendrocytedevelopment. Development 120:1097–1108.
Barakat-Walter I. 1999. Role of thyroid hormones and their receptorsin peripheral nerve regeneration. J Neurobiol 40:541–559.
Barakat-Walter I, Duc C, Puymirat J. 1993. Changes in nuclear3,5,39-triiodothyronine receptor expression in rat dorsal root gan-glia and sciatic nerve during development: comparison with regen-eration. Eur J Neurosci 5:319–326.
Barakat-Walter I, Riederer BM. 1996. Triiodothyronine and nervegrowth factor are required to induce cytoplasmic dynein expressionin rat dorsal root ganglion cultures. Brain Res Dev Brain Res96:109–119.
Bock GR, Goode JA. 1995. Non-reproductive actions of sex steroids.Chichester: John Wiley & Sons.
Bolger MB, Jorgensen EC. 1980. Molecular interactions between thy-roid hormones analogs and the rat liver receptor. Partition of equi-librium binding free energy changes into substituent group inter-action. J Biol Chem 255:10271–10277.
Boutry JM, Hauw JJ, Gansmuller A, Di-Bert N, Pouchelet M, Baron-Van Evercoren A. 1992. Establishment and characterization of amouse Schwann cell line which produces myelin in vivo. J NeurosciRes 32:15–26.
Brockes JP, Fields KL, Raff MC. 1979. Studies on cultured ratSchwann cells. I. Establishment of purified populations from cul-tures of peripheral nerve. Brain Res 165:105–108.
Cao X, Koski RA, Gashler A, McKiernan M, Morris CF, Gaffney R,Hay R, Suskhatme VP. 1990. Identification and characterization ofthe Egr-1 gene product, a DNA-binding zinc finger protein inducedby differentiation and growth signals. Mol Cell Biol 10:1931–1939.
Carre JL, Demerens C, Rodriguez-Pena A, Floch HH, Vincendon G,Sarlieve LL.1998. Thyroid hormone receptor isoforms are sequen-tially expressed in oligodendrocyte lineage cells during rat cerebraldevelopment. J Neurosci Res 54:584–594.
Chan S, Kilby MD. 2000. Thyroid hormone and central nervous sys-tem development. J Endocrinol 165:1–8.
Chan JR, Philipps LJ, Glaser M. 1998. Glucocorticoids and progestinssignal the initiation and enhance the rate of myelin formation. ProcNatl Acad Sci USA 95:10459–10464.
Clos J, Legrand J .1970. Effects of thyroid deficiency and underfeed-ing on growth and myelination of the sciatic nerve fibers in theyoung white rat. An electron microscopic study. Brain Res 22:285–297.
88 MERCIER ET AL.

De Felipe C, Hunt SP. 1994. The differential control of c-Jun expres-sion in regenerating sensory neurons and their associated glialcells. J Neurosci 14:2911–2923.
Farsetti A, Desvergne B, Hallenbeck P, Robbins J, Nikodem V. 1992.Characterization of myelin basic protein thyroid hormone responseelement and its function in the context of native and heterologouspromoter. J Biol Chem 267:15784–15788.
Glauser L, Barakat-Walter I. 1997. Differential distribution of thyroidhormone receptor isoform in rat dorsal root ganglia and sciaticnerve in vivo and in vitro. J Neuroendocrinol 9:217–227.
Ghorbel MT, Seugnet I, Hadj-Sahraoui N, Topilko P, Levi G, Deme-neix B. 1999. Thyroid hormone effects on Krox-24 transcription inthe post-natal mouse brain developmentally regulated but are notcorrelated with mitosis. Oncogene 18:917–924.
Guennoun R, Schumacher M, Robert F, Delespierre B, Gouezou M,Eychenne B, Akwa Y, Robel P, Baulieu EE. 1997. Neurosteroids:expression of functional 3beta-hydroxysteroid dehydrogenase by ratsensory neurons and Schwann cells. Eur J Neurosci 9:2236–2247.
Herdegen T, Leah JD. 1998. Inducible and constitutive transcriptionfactors in the mammalian nervous system: control of gene expres-sion by Jun, Fos and Krox, and CREB/ATF proteins. Brain ResBrain Res Rev 28:370–490.
Hu E, Mueller E, Oliviero S, Papaioannou V, Johnson D, SpiegelmanBM. 1994. Targeted disruption of the c-fos gene demonstrate c-fosdependent and independent pathways for gene expression stimu-lated by growth factors or oncogenes. EMBO J 13:3094–3104.
Johnson RS, Spiegelman BM, Papaionnou V. 1992. Pleiotropic effectsof a null mutation in the c-fos proto-oncogene. Cell 71:577–586.
Karin M, Liu Z, Zandi E. 1997. AP-1 function and regulation. CurrOpin Cell Biol 9:240–246.
Knipper M, Bandtlow C, Gestwa L, Kopschall I, Rohbock K, WiechersB, Zenner HP, Zimermann U. 1998. Thyroid hormone affectSchwann cell and oligodendrocyte gene expression at the glial tran-sition zone of the VIIIth nerve prior to cochlea function. Develop-ment 125:3709–3718.
Koenig HL, Schumacher M, Ferzaz B, Do Thi AN, Ressouches A,Guennoun R, Jung-Testas I, Robel P, Akwa Y, Beaulieu EE. 1995.Progesterone synthesis and myelin formation by Schwann cells.Science 268:1500–1503.
Legrand J. 1986. Thyroid hormone effects on growth and develop-ment. In: Hennemann G, editor. Thyroid hormone metabolism. NewYork: Marcel Dekker. p 503–534.
Lee MM. Badache A, Devries GH. 1999. Phosphorylation of CREB inaxon-induced Schwann cell proliferation. J Neurosci Res 55:702–712.
Marta CB, Adamo AM, Soto EF, Pasquini JM. 1998. Sustained neo-natal hyperthyroidism in the rat affects myelination in the centralnervous system. J Neurosci Res 53:251–259.
McIsaac G, Kiernan JA. 1975. Accelerated recovery from peripheralnerve injury in experimental hyperthyroidism. Exp Neurol 48:88–94.
McKnight GS. 1977. A colorimetric method for the determination ofsubmicrogram quantities of protein. Anal Biochem 78:86–92.
Melcangi RC, Magnaghi V, Cavaretta L, Martini L, Piva F. 1998. Ageinduced decrease of glycoprotein P0 and myelin basic protein geneexpression in the rat sciatic nerve. Repair by steroid derivatives.Neurosciences 23:569–578.
Mellstrom B, Pipaon C, Naranjo JR, Perez-Castillo A, Santos A. 1994.Differential effect of thyroid hormone on NGFI-A gene expressionrat brain. Endocrinology 135:583–588.
Mirsky R, Jessen KR. 1999. The neurobiology of Schwann cells. BrainPathol 9:293–311.
Murphy P, Topilko P, Schneider-Mansoury S, Seitanidou T, CharnayP. 1996. The regulation of Krox-20 expression reveals importantsteps in the control of peripheral glial cell development. Develop-ment 122:2847–2857.
Notterpek L, Snipes GJ, Shooter EM 1999. Temporal expression pat-tern of peripheral myelin protein 22 during in vivo and in vitromyelination. Glia 25:358–369.
Oppenheimer JH. 1999. Evolving concepts of thyroid hormone action.Biochimie 81:539–543.
Pennypacker KR, Hong JS, Mullis SB. Hudson PM, McMillian MK.1996. Transcription factors in primary glial cultures: changes withneuronal interactions. 37:224–230.
Perez P, Palomino T, Schonthal A, Aranda A. 1994. Determination ofthe promoter elements that mediate repression of c-fos gene tran-scription by thyroid hormone and retinoic acid receptors. BiochemBiophys Res Commun 30:135–140.
Perkins AT, Morgenlander JC. 1999. Endocrinologic causes of periph-eral neuropathy. Pins and needles in a stocking-and-glove patternand other symptoms. Postgrad Med J 102:81–82.
Pollard JD. 1993. Neuropathy in diseases of the thyroid and pituitaryglands. In: Dyck PJ, Thomas PK, Griffin JW, Low PA, Poduslo JF,editors. Peripheral Neuropathy. Third edition. Philadelphia: WBSaunders. p 1266–1274.
Robert F, Guennoun R, Desarnaud F, Do Thi AN, Benmessahel Y,Baulieu EE, Schumacher M. 2001. Synthesis of progesterone inSchwann cells: regulation by neurons. Eur J Neurosci 13:916–924.
Samuels HH, Stanley F, Casanova J. 1979. Relation of receptor affin-ity to the modulation of thyroid hormone nuclear receptor levels andgrowth hormone synthesis by L-triidothyronine and iodothyronineanalogues in cultured GH 1 cells. J Clin Invest 63:1229–1240.
Schumacher M, Jung-Testas I, Robel P, Baulieu EE. 1993. Insulin-like growth factor I: a mitogen for rat Schwann cells in the presenceof elevated levels of cyclic AMP. Glia 4:232–240.
Schumacher M, Robel P, Baulieu EE. 1996. Development and regen-eration of the nervous system: a role for neurosteroids. Dev Neuro-sci 18:6–21.
Stewart HJ.1995. Expression of c-Jun, Jun B, Jun D and cAMPresponse element binding protein by Schwann cells and their pre-cursors in vivo and in vitro. Eur J Neurosci 7:1366–1375.
Tabernero A. Stewart HJS, Jessen K, Misky R. 1998. The neuron–gliasignal b neuregulin induces sustained CREB phosphorylation onser-133 in cultured rat Schwann cells. Mol Cell Neurol 10:309–322.
Topilko P, Schneider-Maunoury S, Levi G, Baron-Van Evercooren A,Chennoufi ABY, Seitanidou T, Babinet C, Charnay P. 1994. Krox-20controls myelination in the peripheral nervous system. Nature 371:796–799.
Topilko P, Levi G, Merlo G, Mantero S, Desmarquet C, Mancardi G,Charnay P. 1997. Differential regulation of the zinc finger genesKrox-20 and Krox-24 (Egr-1) suggests antagonistic roles inSchwann cells. J Neurosci Res 50:702–712.
Usson Y, Saxod R. 1988. Schwann cell proliferation in the sciaticnerve of hypothyroid chick embryos studied by autoradiographyand image analysis. J Neurocytol 17:641–649.
Voinesco F, Glauser L, Kraftsik R, Barakat-Walter I. 1998. Localadministration of thyroid hormones in silicone chamber increasesregeneration of rat transected sciatic nerve. Exp Neurol 150:69–81.
Zhang J, Lazar MA. 2000. The mechanism of action of thyroid hor-mones. Annu Rev Physiol 62:439–466.
Zorick TS, Lemke G. 1996. Schwann cell differentiation. Curr OpinCell Biol 8:870–876.
89T3 EFFECTS ON SCHWANN CELLS





























