Embed Size (px)
Citation preview

Radiation therapy causes loss of dermal lymphatic vessels and interferes withlymphatic function by TGF-�1-mediated tissue fibrosis
Tomer Avraham,1 Alan Yan,1 Jamie C. Zampell,1 Sanjay V. Daluvoy,1 Adriana Haimovitz-Friedman,2
Andrew P. Cordeiro,1 and Babak J. Mehrara1
1Division of Plastic and Reconstructive Surgery, Department of Surgery, and 2Department of Radiation Oncology, MemorialSloan-Kettering Cancer Center, New York, New York
Submitted 8 December 2009; accepted in final form 1 June 2010
Avraham T, Yan A, Zampell JC, Daluvoy SV, Haimovitz-Friedman A, Cordeiro AP, Mehrara BJ. Radiation therapycauses loss of dermal lymphatic vessels and interferes with lym-phatic function by TGF-�1-mediated tissue fibrosis. Am J PhysiolCell Physiol 299: C589 –C605, 2010. First published June 2, 2010;doi:10.1152/ajpcell.00535.2009.—Although radiation therapy is amajor risk factor for the development of lymphedema followinglymphadenectomy, the mechanisms responsible for this effect remainunknown. The purpose of this study was therefore to determine theeffects of radiation on lymphatic endothelial cells (LECs) and lym-phatic function. The tails of wild-type or acid sphingomyelinase(ASM)-deficient mice were treated with 0, 15, or 30 Gy of radiationand then analyzed for LEC apoptosis and lymphatic function atvarious time points. To analyze the effects of radiation fibrosis onlymphatic function, we determined the effects of transforming growthfactor (TGF)-�1 blockade after radiation in vivo. Finally, we deter-mined the effects of radiation and exogenous TGF-�1 on LECs invitro. Radiation caused mild edema that resolved after 12–24 wk.Interestingly, despite resolution of tail edema, irradiated animalsdisplayed persistent lymphatic dysfunction. Radiation caused loss ofcapillary lymphatics and was associated with a dose-dependent in-crease in LEC apoptosis. ASM�/� mice had significantly less LECapoptosis; however, this finding did not translate to improved lym-phatic function at later time points. Short-term blockade of TGF-�1function after radiation markedly decreased tissue fibrosis and signif-icantly improved lymphatic function but did not alter LEC apoptosis.Radiation therapy decreases lymphatic reserve by causing depletion oflymphatic vessels and LECs as well as promoting soft tissue fibrosis.Short-term inhibition of TGF-�1 activity following radiation im-proves lymphatic function and is associated with decreased soft tissuefibrosis. ASM deficiency confers LEC protection from radiation-induced apoptosis but does not prevent lymphatic dysfunction.
lymphedema; endothelium; transforming growth factor-�1
LYMPHEDEMA IS a debilitating disorder that affects more than3 million Americans (2, 48). In the United States the mostcommon cause of lymphedema is lymph node dissection forthe treatment of a variety of malignancies (44, 60). Becauseof the relative prevalence of breast cancer and the fact thatlymphatic metastasis is the primary source of metastasis inthese patients, axillary lymph node dissection (ALND) is themost commonly encountered cause of lymphedema clini-cally in Western countries (23). Patients with lymphedemaare easily recognizable by their dependence on tight-fittinggarments needed to prevent the inevitable progression ofthis disorder. Because of the lack of effective treatmentoptions, these patients have significantly decreased quality
of life with frequent infections, decreased function, anddisfigurement (3, 60, 61).
A large number of clinical studies have identified risk factorsfor the development of lymphedema (22, 35, 42, 59, 61). Thevast majority of these studies have identified radiation therapyas an independent risk factor for the development of lymphed-ema (23). In fact, it is estimated that postoperative radiationtherapy increases the risk of developing lymphedema by asmuch as 10-fold (44). In addition, it appears that the combi-nation of surgery and radiation is necessary for the develop-ment of lymphedema since radiation alone is infrequentlyassociated with clinically apparent lymphedema. Nevertheless,the potential mechanisms by which radiation therapy increasesthe risk of lymphedema remain essentially unknown.
One previous study demonstrated that intestinal lymphaticendothelial cells (LECs) are relatively radioresistant comparedwith microvascular endothelial cells (57). This finding was notreproduced by a more recent study demonstrating intestinallymphatic endothelial apoptosis shortly after total body irradi-ation (1). The clinical relevance of studies on intestinal LECsis somewhat difficult to interpret since lymphatic dysfunctionand subsequent extremity lymphedema are related to cutaneouslymphatic channels, which may be structurally or functionallydifferent from intestinal lymphatics. In fact, Jackowski et al.(27) have shown that decreased numbers of cutaneous lym-phatics after radiation therapy in breast cancer patients areassociated with an increased rate of lymphedema. In addition,Mortimer et al. (40) have shown in limited studies that radia-tion therapy is associated with cutaneous lymphatic dysfunc-tion in pigs. Therefore, it is possible that radiation therapycontributes to the development of lymphedema by causingdepletion or dysfunction of cutaneous lymphatic channels.
Radiation therapy may also increase the risk of lymphedemaafter ALND by promoting tissue fibrosis. Radiation therapy iswell known to cause tissue fibrosis as a result of transforminggrowth factor (TGF)-�1-dependent mechanisms (36, 37). Werecently showed (8) that fibrosis is a critical regulator oflymphatic regeneration and that this effect is largely dependenton TGF-�1 expression. In addition, we (8) and others (41) haveshown that TGF-�1 is a direct inhibitor of LEC proliferationand function. Therefore, it is feasible that at least some of thenegative consequences of radiation therapy on the lymphaticsystem may be secondary to tissue fibrosis. This hypothesis issupported by the fact that clinical factors that increase the riskof fibrosis (e.g., extensive surgery, infections, or obesity) alsosignificantly increase the risk of lymphedema (34, 35).
The purpose of these studies was to determine the effects ofionizing radiation therapy on cutaneous lymphatic vessels.Using a mouse tail model, we show that radiation causes a
Address for reprint requests and other correspondence: B. J. Mehrara, 1275York Ave., Rm. MRI 1005, New York, NY 10065 (e-mail: [email protected]).
Am J Physiol Cell Physiol 299: C589–C605, 2010.First published June 2, 2010; doi:10.1152/ajpcell.00535.2009.
0363-6143/10 Copyright © 2010 the American Physiological Societyhttp://www.ajpcell.org C589
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

dose-dependent, long-term decrease in lymphatic function.Impaired lymphatic function resulting from radiation is asso-ciated with a combination of LEC apoptosis, long-term de-crease in the number of cutaneous lymphatic vessels, and softtissue fibrosis. We show that LECs in acid sphingomyelinase(ASM)-deficient animals, similar to microvascular endothelialcells, are significantly protected from radiation-induced apop-tosis. However, this protective effect does not inhibit develop-ment of tissue fibrosis or improve lymphatic function at latertime points. Short-term inhibition of TGF-�1 significantlydecreased tissue fibrosis and markedly improved lymphaticfunction but did not provide a protective effect for LECapoptosis or lymphatic depletion. Finally, radiated lymphaticsdemonstrated evidence of endothelial-mesenchymal transition(EMT) in vivo. In vitro exposure of LECs to recombinantTGF-�1 increased expression of collagen and markers offibroblast differentiation while simultaneously decreasing theexpression of lymphatic markers [lymphatic vessel endothelialreceptor-1 (LYVE-1)]. Together, these results indicate thatlymphatic dysfunction resulting from radiation therapy doesnot cause clinically apparent lymphedema but may predisposeto the development of lymphedema when combined with otherinsults by promoting tissue fibrosis (e.g., surgery). In addition,our results indicate that prevention of tissue fibrosis resultingfrom radiation may be a means of protecting against thispotentially devastating complication.
METHODS
Mouse tail irradiation. All animal experiments were approved bythe Research Animal Resource Center Institutional Animal Care andUse Committee (IACUC) at Memorial Sloan-Kettering Cancer Cen-ter. C57BL/6 mice were anesthetized with ketamine-xylazine and thencarefully shielded, and their tails were then irradiated with the X-RAD320 (Precision X-Ray, North Branford, CT) with doses of 15 or 30Gy. We chose the mouse tail for irradiation because it is an appendagethat has been previously used extensively in studies of lymphaticfunction and regeneration and is also easily shielded from the rest ofthe animal. Control mice received anesthetic and were placed in theirradiator but received no radiation. Mice were then euthanized for tailharvest after 4 h, 10 h, 1 wk, 4 wk, 12 wk, and 24 wk followingirradiation (n � 5 for each dose at each time point evaluated).
Irradiation of acid sphingomyelinase-deficient mice. Mice deficientin the ceramide ASM have been previously shown to be protectedfrom radiation-induced apoptosis in a variety of cell types includingmicrovascular endothelial cells (43). As such, we evaluated the impactof this deficiency on LEC apoptosis following irradiation. ASM�/�mice established on the SV129/C57BL/6 background along withwild-type (WT) littermates underwent tail irradiation as describedabove. Tissues were harvested at 10 h and 4 wk after radiation (n �3 for each time point). The 4 wk time point was chosen as the longestfollow-up since this represented our longest follow-up in other groupsfor assessment of apoptosis.
Short-term TGF-�1 blockade with LY-364947. To determine theeffectiveness of TGF-�1 blockade in alleviating the affects of radia-tion therapy on lymphatic function, we treated animals with LY-364947 (Sigma, St. Louis, MO), a well-described small-moleculeinhibitor. This molecule is an ATP-competitive inhibitor of TGF-�receptor (TGF-BR)I and blocks association with TGF-BRII and sub-sequent Smad3 phosphorylation (52). LY-364947 was dissolved at aconcentration of 5 mg/ml in DMSO. Experimental animals (n � 5)were treated with LY-364947 (1 mg/kg diluted in PBS) deliveredintraperitoneally starting 1 day before tail irradiation with 15 Gy.Animals were subsequently treated every other day for three addi-tional doses. Control mice were irradiated in an identical manner but
treated with DMSO diluted in PBS. While long-term inhibition ofTGF-�1 may be associated with immune dysfunction, short-terminhibition as performed in our study is well tolerated (71). No adverseside effects were noted in our animals treated with LY-364947.
Tail volume measurements. To evaluate the degree of acutelymphedema following irradiation, tail volumes were determined withthe truncated cone formula [V � 1/4�(C1C2 � C2C3 �� � C7C8); Cis circumference] as previously described (53). Briefly, tail circum-ference was measured by blinded reviewers at four points at 10-mmintervals starting 20 mm from the tail base with a digital caliper. Tailvolume changes from baseline were calculated for each animal inorder to minimize the potential effects of interanimal baseline differ-ences in tail volumes.
Lymphoscintigraphy. To quantify lymphatic transport after irradi-ation, 99Tc-sulfur colloid (100-nm particle size; 400–800 �Ci in �50�l) was injected intradermally �20 mm from the tail tip with ourpreviously reported methods (5). This colloid is taken up by thelymphatics and transported to the lymph nodes at the base of the tailand is a sensitive, easily quantifiable measure of lymphatic transport.Briefly, dynamic planar gamma camera images were acquired in 15-to 600-s frames for up to 130 min after injection with an X-SPECT(Gamma Medica, Northridge, CA) fitted with low-energy parallel-hole collimators. Resulting dynamic images were analyzed withASIPro (CTI Molecular Imaging, Knoxville, TN), and region ofinterest (ROI) analysis was performed to derive the decay-adjustedactivity at the lymph nodes at the base of the mouse tail. Time-activitydata were then fit to a function exponentially decreasing to anasymptotic value for the injection site or a function exponentiallyincreasing to an asymptotic value for the individual lymph nodes.Maximal 99Tc uptake was calculated and plotted.
Specimen preparation, histology, and immunostaining. After eu-thanasia, mouse tails were fixed in 4% paraformaldehyde at 4°C for 24h and then decalcified in Immunocal (Decal Chemical, Tallman, NY).A 1-cm portion of tail was harvested 2 cm from the base of tail andsectioned longitudinally. Additionally, 2-mm cross sections of tailwere obtained both immediately distal and proximal to this piece. Allthree pieces of tail were embedded in paraffin and sectioned at 5 �m.
Sections were histologically stained with standard protocols andevaluated with a Leica microscope (Wetzlar, Germany). Histochem-ical staining for scar tissue was performed with picro-Sirius red(Direct Red 80, Sigma) as previously described (16). Sirius red-stained specimen were examined by polarized light microscopy (LeicaTCS AOBSSP2), and the scar index was calculated as previouslydescribed with Metamorph Offline software (Molecular Devices,Sunnyvale, CA) in a minimum of three 70,000-�m sections peranimal (16). A minimum of three animals were evaluated per group/per time point. Sirius red staining quantification of scarring andfibrosis is based on the fact that normal skin is characterized by thin,randomly oriented collagen fibers demonstrating yellow-green bire-fringence while scarring is associated with deposition of thick parallelcollagen bundles with orange-red birefringence (15, 16). The scarindex is the ratio of orange-red to yellow-green pixels, with highervalues representing increased scarring.
Immunohistochemical and immunofluorescent staining were per-formed as previously described to localize LECs, growth factors, andmyofibroblast markers (39). Primary antibodies used were againstLYVE-1 (rabbit polyclonal, Abcam, Cambridge, MA), podopla-nin (Syrian hamster monoclonal, Abcam), phosphorylated Smad3(pSmad3, rabbit polyclonal, Abcam), and �-smooth muscle actin(�-SMA, Abcam). Immunofluorescent secondary antibodies used flu-orescein (R&D Systems, Minneapolis, MN) or Cy3 Zymed (Invitro-gen Molecular Probes, Carlsbad, CA). For immunohistochemistry,secondary antibody was from the VECTASTAIN ABC Kit (Vector,Burlingame, CA) and developed with 3,3=-diaminobenzidine (DAB).For all immunohistochemical or immunofluorescent studies, negativecontrol included tissue incubated with secondary antibody but notprimary antibody.
C590 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

Images were obtained by bright-field microscopy (Leica TCS) forimmunohistochemistry and with a confocal microscope for immuno-fluorescence (Leica). Lymphatic vessel and lymphatic endothelial(LEC) cell counts were performed by two blinded observers in threeto five random high-power fields (HPFs) in a minimum of threeanimals/group fields. There was minimal interobserver variation inthe analysis. For colocalization of podoplanin and �-SMA, slideswere scanned with the Mirax Slide Scanner (Zeiss). Captured bright-field and fluorescent images taken at the same location on the slidewere overlaid with Adobe Photoshop (Adobe, San Jose, CA).
In vivo detection of LEC apoptosis was performed on paraffin-embedded tissue sections. Apoptotic cells were detected with theApoptag Red In Situ Apoptosis Detection Kite per the manufacturer’sprotocol (Millipore, Billerica, MA). This assay labels apoptotic cellsby using terminal deoxynucleotidyltransferase-mediated dUTP nickend labeling (TUNEL) staining. To localize apoptotic LECs, sectionswere double stained with anti-LYVE-1 antibody as described above,labeled with a fluorescein secondary antibody, and imaged with aconfocal microscope (Leica). For quantification of LEC apoptosis thetotal number of LYVE-1� cells was counted by two blinded review-ers in three to five random HPFs (400 magnification) in a minimumof three animals/time point/group. We identified cells that expressedboth TUNEL and LYVE-1, and these were considered apoptoticLECs; their proportion was expressed as a percentage of the totalnumber of LECs.
Western blot analysis. Total cellular protein was isolated from tailsections inclusive of the skin and subcutaneous tissues 1 wk afterirradiation from animals treated with LY-364947 or vehicle control.Protein was harvested with the Qiagen DNA/RNA/Protein Mini Kitaccording to the manufacturer’s protocol (Qiagen, Valencia, CA) andquantified with the Bradford method. Western blotting was performedas previously described (8). Western blot analysis for TGF-�-induced(TGFBi), a protein that is upregulated by and represents a surrogatemarker of TGF-�1 activity, was performed with a rabbit polyclonalantibody (Abcam) and normalized to actin levels (24, 62). Antibodiesfor N-cadherin, E-cadherin, LYVE-1, fibroblast activation protein(FAP), and collagen I were all purchased from Abcam. Immunoreac-tivity was determined with the ECL chemiluminescence detectionsystem (Amersham, Arlington Heights, IL).
Cell culture and reagents. Human dermal lymphatic endothelialcells (HLECs) were obtained from PromoCell (Heidelberg, Ger-many), cultured in ECGM-MV containing 0.4% ECGS/H, 5% FCS,EGF (10 ng/ml), hydrocortisone (1 �g/ml), and penicillin-streptomy-cin (50 U/ml) (Invitrogen), and passaged every 48 h. LEC morphol-ogy was confirmed with immunofluorescent staining for LYVE-1 andprox-1 (not shown). Early-passage cells (10) were used for allexperiments.
For in vitro assay of LEC apoptosis, cells were grown to near-confluence in six-well plates or chamber slides. They were thenirradiated with varying doses of radiation and evaluated at varioustime points. Flow cytometric analysis for apoptosis by TUNEL assaywas carried out with the Apo-Direct TUNEL kit per the manufacturer’sprotocol (Millipore). Flow cytometry was performed on the FACS-caliber flow cytometer (BD Biosciences, San Jose, CA) and analyzedwith Flowjo software (Tree Star, Ashland, OR). Immunofluorescentconfirmation of LEC apoptosis was carried out with the Dual Apo-ptosis Kit per the manufacturer’s protocol (Biotium, Hayward, CA).This kit allows concomitant detection of annexin V and caspase-3 asdual markers of apoptotic cell death.
To assess LEC senescence after radiation therapy in vitro, cellswere grown to �60% confluence in six-well plates and then exposedto various doses of irradiation and grown for an additional 4 days.After 4 days, the medium was removed and the cells were briefly fixedin 4% paraformaldehyde followed by staining with the BiovisionSenescence Detection Kit (Biovision, Mountain View, CA). Thisassay is based on the fact that senescent cells stain for senescence-associated (SA)-�-galactosidase at a pH of 6.0 with X-gal and has
been used in a large number of studies to identify cellular senescencein vitro (25). The number of stained cells as a function of total numberof cells present was quantified by blinded reviewers in a minimum offour plates per group.
To evaluate the direct effects of TGF-�1 on LECs, LECs weregrown in vitro in fibronectin-coated flasks (Sigma). At the initiation ofexperiments medium was changed to DMEM with 5% FCS with orwithout 10 ng/ml recombinant human (rh)TGF-�1 (R&D Systems).Cells were then evaluated at specified time points.
Statistical analysis. Multigroup comparison was performed byone-way ANOVA with the Tukey-Kramer post hoc test. Student’st-test was used for analyzing differences between two groups. Data arepresented as means � SD or SE as noted, with P 0.05 consideredsignificant.
RESULTS
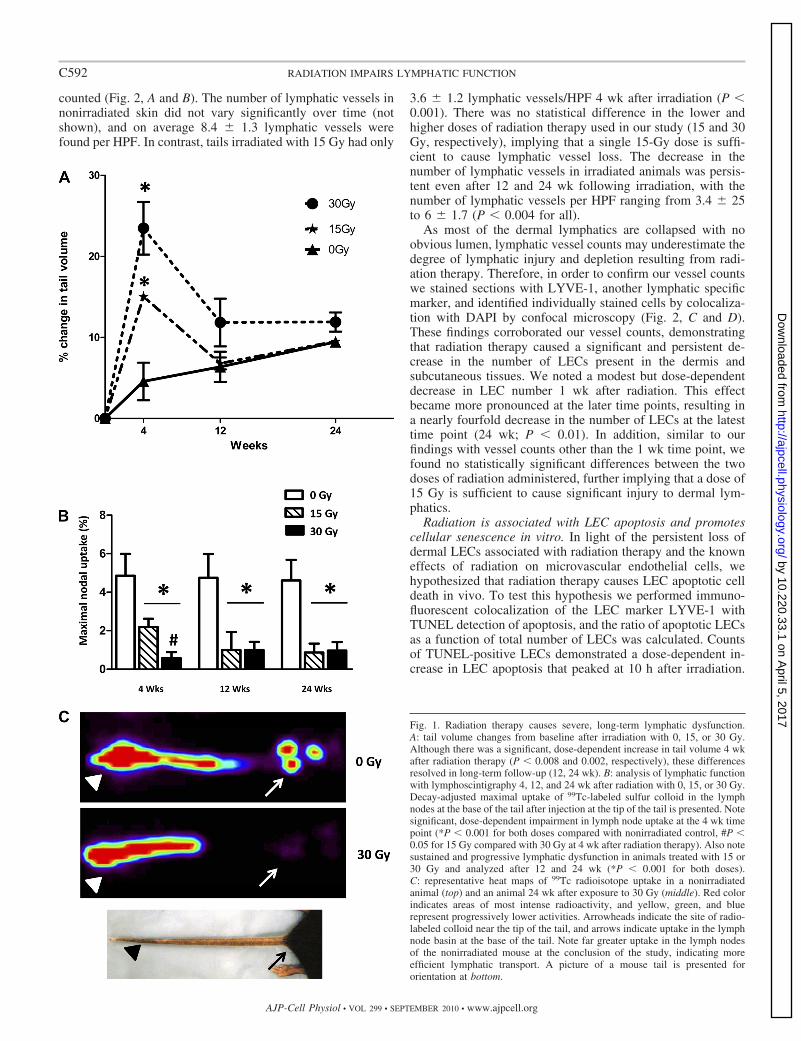
Radiation is associated with severe long-term lymphaticdysfunction. To assess the impact of radiation therapy on tailedema, mouse tails were radiated with a single dose of 0, 15,or 30 Gy and tail volume changes compared with baseline wereevaluated with the truncated cone formula (Fig. 1A). Fourweeks after radiation therapy with either 15 or 30 Gy, a modestalthough significant and dose-dependent increase in tail vol-umes was noted. Animals treated with a dose of 15 Gy had a�15% increase, while those treated with 30 Gy demonstratedan �25% increase in baseline tail volume (P 0.008 and0.002, respectively, compared with 0 Gy). These acute in-creases in tail edema were nearly resolved by 12 wk afterradiation therapy, demonstrating no significant differencesamong the various groups. In addition, no statistical changes intail volumes were noted in animals followed for as long as 24wk after irradiation. Thus, similar to the clinical scenario,radiation therapy alone does not appear to cause chronicswelling or lymphedema in our mouse tail model.
In an effort to evaluate the effect of radiation on dermallymphatic function, we performed lymphoscintigraphy with99Tc-labeled sulfur colloid and quantified the maximal uptakeof this material in the lymph node basin at the base of the tail.This analysis demonstrated that radiation therapy caused sig-nificant and dose-dependent decreases in lymphatic function 4wk after treatment compared with controls (P 0.0001 andP 0.003 for 30 Gy and 15 Gy, respectively, compared withcontrols; Fig. 1B). Interestingly, lymphatic dysfunction per-sisted at the later time points examined (12 and 24 wk),demonstrating severe lymphatic dysfunction despite resolutionof measurable tail lymphedema (Fig. 1B). In addition, bothradiation doses appeared to cause significant lymphatic dys-function, with an apparent progression of this phenomenon inanimals treated with 15 Gy such that lymph node uptake wasequivalent in severity with the 30-Gy-treated animals (Fig. 1B).Decay-adjusted lymph node uptake of radiolabeled sulfur col-loid was nearly fivefold lower in the treated animals comparedwith control (P 0.001). Image analysis of lymphoscinti-graphic data demonstrated gross impairment in nodal uptake ofradiolabeled colloid in irradiated animals compared with non-irradiated control animals (Fig. 1C).
Radiation therapy decreases number of dermal capillarylymphatic vessels and lymphatic endothelial cells. To deter-mine the impact of radiation therapy on the number of cuta-neous lymphatic vessels present tail sections were stained withthe lymphatic-specific marker podoplanin by immunohisto-chemistry and positively stained luminal structures were
C591RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from
counted (Fig. 2, A and B). The number of lymphatic vessels innonirradiated skin did not vary significantly over time (notshown), and on average 8.4 � 1.3 lymphatic vessels werefound per HPF. In contrast, tails irradiated with 15 Gy had only
3.6 � 1.2 lymphatic vessels/HPF 4 wk after irradiation (P 0.001). There was no statistical difference in the lower andhigher doses of radiation therapy used in our study (15 and 30Gy, respectively), implying that a single 15-Gy dose is suffi-cient to cause lymphatic vessel loss. The decrease in thenumber of lymphatic vessels in irradiated animals was persis-tent even after 12 and 24 wk following irradiation, with thenumber of lymphatic vessels per HPF ranging from 3.4 � 25to 6 � 1.7 (P 0.004 for all).
As most of the dermal lymphatics are collapsed with noobvious lumen, lymphatic vessel counts may underestimate thedegree of lymphatic injury and depletion resulting from radi-ation therapy. Therefore, in order to confirm our vessel countswe stained sections with LYVE-1, another lymphatic specificmarker, and identified individually stained cells by colocaliza-tion with DAPI by confocal microscopy (Fig. 2, C and D).These findings corroborated our vessel counts, demonstratingthat radiation therapy caused a significant and persistent de-crease in the number of LECs present in the dermis andsubcutaneous tissues. We noted a modest but dose-dependentdecrease in LEC number 1 wk after radiation. This effectbecame more pronounced at the later time points, resulting ina nearly fourfold decrease in the number of LECs at the latesttime point (24 wk; P 0.01). In addition, similar to ourfindings with vessel counts other than the 1 wk time point, wefound no statistically significant differences between the twodoses of radiation administered, further implying that a dose of15 Gy is sufficient to cause significant injury to dermal lym-phatics.
Radiation is associated with LEC apoptosis and promotescellular senescence in vitro. In light of the persistent loss ofdermal LECs associated with radiation therapy and the knowneffects of radiation on microvascular endothelial cells, wehypothesized that radiation therapy causes LEC apoptotic celldeath in vivo. To test this hypothesis we performed immuno-fluorescent colocalization of the LEC marker LYVE-1 withTUNEL detection of apoptosis, and the ratio of apoptotic LECsas a function of total number of LECs was calculated. Countsof TUNEL-positive LECs demonstrated a dose-dependent in-crease in LEC apoptosis that peaked at 10 h after irradiation.
Fig. 1. Radiation therapy causes severe, long-term lymphatic dysfunction.A: tail volume changes from baseline after irradiation with 0, 15, or 30 Gy.Although there was a significant, dose-dependent increase in tail volume 4 wkafter radiation therapy (P 0.008 and 0.002, respectively), these differencesresolved in long-term follow-up (12, 24 wk). B: analysis of lymphatic functionwith lymphoscintigraphy 4, 12, and 24 wk after radiation with 0, 15, or 30 Gy.Decay-adjusted maximal uptake of 99Tc-labeled sulfur colloid in the lymphnodes at the base of the tail after injection at the tip of the tail is presented. Notesignificant, dose-dependent impairment in lymph node uptake at the 4 wk timepoint (*P 0.001 for both doses compared with nonirradiated control, #P 0.05 for 15 Gy compared with 30 Gy at 4 wk after radiation therapy). Also notesustained and progressive lymphatic dysfunction in animals treated with 15 or30 Gy and analyzed after 12 and 24 wk (*P 0.001 for both doses).C: representative heat maps of 99Tc radioisotope uptake in a nonirradiatedanimal (top) and an animal 24 wk after exposure to 30 Gy (middle). Red colorindicates areas of most intense radioactivity, and yellow, green, and bluerepresent progressively lower activities. Arrowheads indicate the site of radio-labeled colloid near the tip of the tail, and arrows indicate uptake in the lymphnode basin at the base of the tail. Note far greater uptake in the lymph nodesof the nonirradiated mouse at the conclusion of the study, indicating moreefficient lymphatic transport. A picture of a mouse tail is presented fororientation at bottom.
C592 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

We identified very few apoptotic LECs in the nonirradiatedanimals (1.1%). In contrast, 10 h after irradiation, animalstreated with 15 Gy demonstrated apoptosis in 8.0% of LECs(P 0.005 compared with 0 Gy), while 30 Gy of radiationresulted in apoptosis in nearly 20% of LECs (P 0.006;Fig. 3, A and B).
To confirm our in vivo findings, we irradiated isolatedhuman dermal LECs in vitro with 0, 15, or 30 Gy of gamma
irradiation and identified apoptotic cells with flow cytometryand TUNEL assays 4 and 10 h later (Fig. 3C). LECs demon-strated a statistically significant increase in apoptosis comparedwith controls with a 30-Gy dose 10 h after irradiation (20.4%vs. 6.8%, P 0.001), but not with a 15-Gy dose. This could bea function of a higher background rate of apoptosis in this invitro study. It should be noted that the baseline rate of LECapoptosis in vivo was 1.1%, while in vitro it was 6.8%. This
Fig. 2. Radiation therapy decreases number of dermal capillary lymphatic vessels and lymphatic endothelial cells (LECs). A: analysis of capillary lymphaticvessels in the skin after irradiation with 0, 15, or 30 Gy. Tissues were stained with podoplanin antibody, and lymphatic vessel counts were performed in 3–5high-power field (HPF) sections in a minimum of 3 animals per group by 2 blinded reviewers. Vessel counts are presented as means � SD. Note significant andsustained decrease in the number of podoplanin-positive luminal structures in irradiated tissue sections compared with nonirradiated control sections (*P 0.05compared with nonirradiated control). B: representative 200 micrographs of tissue sections from a nonirradiated animal (0 Gy; top) and an animal 24 wk aftera 30-Gy dose of radiation (30 Gy; bottom) stained immunohistochemically for podoplanin (arrow demonstrates positively stained lymphatic). C: analysis of thenumber of LECs/HPF using immunofluorescent staining for lymphatic vessel endothelial receptor-1 (LYVE-1). Cell counts were performed in 3–5 HPFs/animalin a minimum of 3 animals per time point/group by blinded reviewers and are presented as means � SD. Again, note significant decrease in LEC counts inirradiated animals beginning 4 wk after irradiation (*P 0.05 for all doses and time points compared with baseline; P 0.001 at 24 wk) D: representative 100micrographs of sections stained for LYVE-1 (red) and DAPI nuclear counterstain (blue) in animal exposed to 0 Gy (top) or 30 Gy (bottom). Note grossly observeddecreased number of LECs (red cells).
C593RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

C594 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

phenomena is well recognized in endothelial cells, as theseoften have a significant rate of background apoptosis in vitroeven with meticulously maintained culture conditions (32).LEC apoptosis following exposure to radiation therapy was con-firmed by immunofluorescence for annexin V and caspase-3 (Fig.3D). This finding is interesting as it suggests that in vivo tissueinjury following radiation may sensitize LECs to apoptosis.Indeed, we recognize that a limitation of in vitro irradiationstudies is that these conditions fail to mimic changes in thespecific niche of individual cells and may therefore underesti-mate the effect of irradiation on isolated cells. However, invitro radiation experiments have been used in multiple previ-ous studies with the dose range described as a means ofvalidating and supporting in vivo observations (13, 51, 58, 63,70).
Irradiation has been previously shown to cause cellularsenescence by upregulating cell cycle regulators such as p21and p16 (63, 74). Senescent cells are metabolically active butdemonstrate significantly reduced potential for proliferationand differentiation (10). To determine the effects of sublethaldoses of irradiation on cellular senescence in isolated dermalLECs, cells were treated with 0, 4, 8, or 12 Gy. LECs were thengrown for an additional 4 days and evaluated for senescence by�-galactosidase staining at pH 6 (Fig. 3, E and F) (9). Inter-estingly, this analysis demonstrated that radiation therapy, evenat low doses, causes a statistically significant and dose-depen-dent increase in LEC senescence compared with controls at alldoses tested. This finding implies that radiation therapy mayimpair lymphatic function by causing premature senescence ofthese cells.
Protection of LECs from radiation-induced apoptosis doesnot prevent long-term lymphatic dysfunction. To study therelative impact of fibrosis and LEC apoptosis on the pathogen-esis of radiation-associated lymphatic dysfunction we irradi-ated the tails of ASM-deficient (ASM�/�) mice and WTlittermates. Evaluation of LEC apoptosis 10 h after treatmentwith 15 Gy demonstrated that ASM�/� mice had significantlyless apoptosis compared with WT control mice (4.5% vs. 8.1%,P 0.03; Fig. 4, A and B). Interestingly, however, thisprotection from apoptosis did not translate to significantlyimproved lymphatic function when assessed by lymphoscin-tigraphy 4 wk after radiation (Fig. 4C). In addition, ASMdeficiency did not significantly decrease radiation-induced fi-brosis as assessed by Sirius red staining (Fig. 4, D and E).
Radiation causes soft tissue fibrosis and increased TGF-�1activity. Radiation is known to cause soft tissue fibrosis (4, 68).In fact, clinically, many of the long-term wound healing andsurgical complications associated with radiation are directlyrelated to soft tissue fibrosis. Previous studies have shown thatpostradiation skin fibrosis is in large measure caused by acti-vation of TGF-�1 and its downstream mediator Smad3 (17).Furthermore, we (5, 8) and others (41) have previously dem-onstrated that TGF-�1 is an antilymphangiogenic cytokine byinhibiting LEC proliferation and function and that fibrosis is asignificant inhibitor of lymphatic function during wound heal-ing. Therefore, to determine the effects of fibrosis in general,and TGF-�1 function in particular, we evaluated fibrosismouse tails treated with 0, 15, or 30 Gy of irradiation.
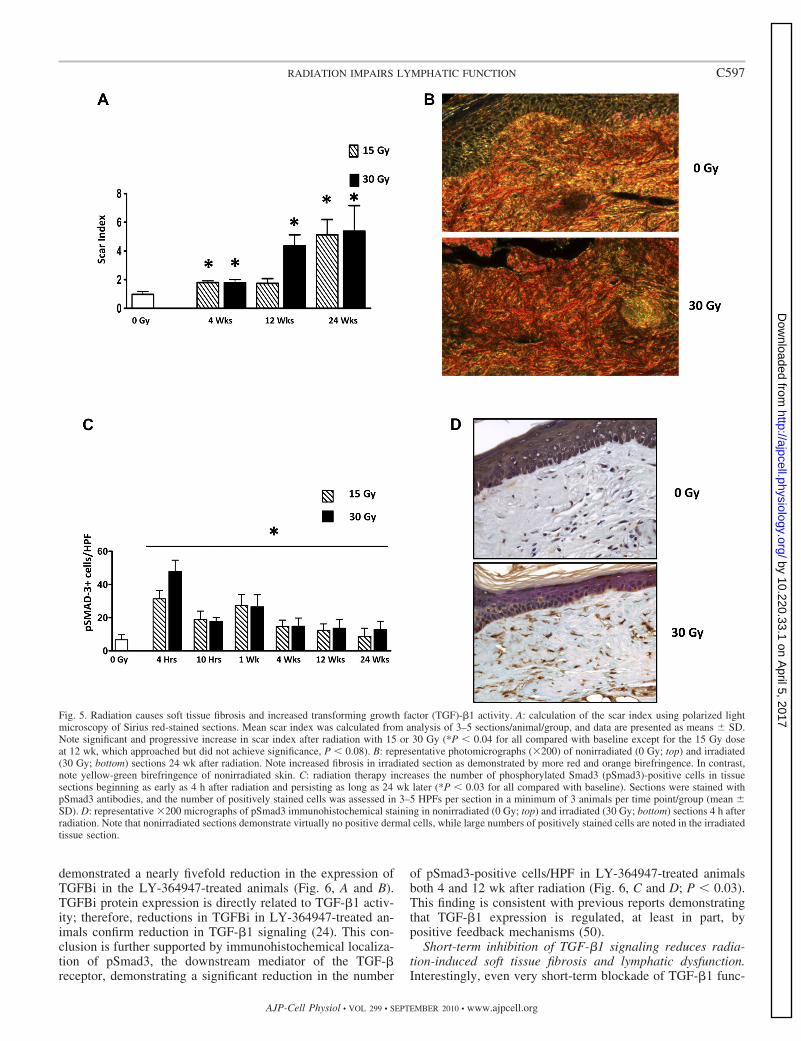
As expected, radiation therapy resulted in loss of hair folli-cles, epidermal thinning, dermal thickening, loss of fat, fibro-sis, and increased inflammation at all times evaluated afterradiation (not shown). Using Sirius red staining and birefrin-gence microscopy to calculate the scar index, we found thatradiation caused progressive fibrosis with a scar index that issignificantly higher than control starting at 4 wk after irradia-tion (P 0.03 for all doses at time points compared withcontrol) (Fig. 5, A and B). In addition, immunohistochemicallocalization of pSmad3, an important downstream mediator ofTGF-�1 signaling, demonstrated a significant increase in thenumber of pSmad3-positive cells as early as 4 wk after radia-tion therapy. Similar to previous reports, this increase inTGF-�1 activity persisted even 24 wk after radiation therapy inanimals treated with 30 Gy (P 0.03) (Fig. 5, C and D).
Treatment with LY-364947, a small-molecule inhibitor ofTGF-BRI, effectively blocks TGF-�1 signaling. Roberts andcolleagues (69) have previously shown that treatment of micewith small-molecule inhibitors of TGF-�1 for 3–5 wk candecrease the fibrosis associated with radiation therapy. How-ever, the effects of short-term TGF-�1 blockade on tissuefibrosis and lymphatic function remain unknown. This is im-portant since long-term inhibition of TGF-�1 function cancause significant morbidity due to the protean effects of thisgrowth factor. Therefore, we inhibited TGF-�1 function with asmall-molecule inhibitor of TGF-BRI (LY-364947) for just 1wk after radiation and evaluated the effects of this interventionon fibrosis, lymphatic function, and lymphatic vessel depletion.
Western blot analysis of tissues harvested 1 wk after radia-tion (15 Gy) from animals treated with or without LY-364947
Fig. 3. Radiation is associated with LEC apoptosis and promotes cellular senescence in vitro. A: identification of apoptotic LECs by colocalization of LYVE-1and terminal deoxynucleotidyltransferase-mediated dUTP nick end labeling (TUNEL). Individual LECs were identified by LYVE-1 staining and counted in 3–5HPFs in a minimum of 3 animals per time point/group by blinded reviewers. Percentage of apoptotic LECs was calculated by similarly counting the number ofLYVE-1�, TUNEL� cells and expressing this number as a function of total LEC number. Data are presented as means � SD. Increased LEC apoptosis wasnoted as early as 4 h after radiation and peaked at the 10 h time point in a dose-dependent manner (*P 0.01). B: representative immunofluorescent micrographs(200) of colocalization of the LEC marker (red) with TUNEL (green) and DAPI nuclear stain (blue) 10 h after irradiation with 0, 15, or 30 Gy. Note thedose-dependent increase in the presence of TUNEL-positive nuclei. C: flow cytometric TUNEL analysis of isolated LECs irradiated in vitro demonstrated thatLECs in this setting are relatively radioresistant, requiring a 30-Gy dose of radiation to induce significant apoptosis (*P 0.0001). Data are presented as means �SD of triplicate experiments. D: this finding was confirmed by immunofluorescence for annexin V and caspase-3. In the presented representative images annexinV is shown by red immunofluorescence, caspase-3 with green immunofluorescence, and DAPI nuclear counterstain in blue. Note the greater number of cellsstaining for both annexin V and caspase-3 in the 30-Gy-treated cells (bottom). E: radiation therapy induces LEC senescence 4 days after treatment, at doses thatare much lower than those required to induce apoptosis. Cells were radiated with 0, 4, 8, or 12 Gy of ionizing radiation, and senescence was analyzed with�-galactosidase staining at pH 6. Positively stained (blue) cells were identified with light microscopy and quantified in 3–5 HPFs/treatment group. Allexperiments were performed in triplicate, and data are presented as means � SD. Note a significant, dose-dependent increase in LEC senescence (*P 0.001for all compared with nonirradiated control). F: representative photomicrographs (100) of �-galactosidase staining of nonirradiated (0 Gy; top) and irradiated(30 Gy; bottom) LECs.
C595RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from
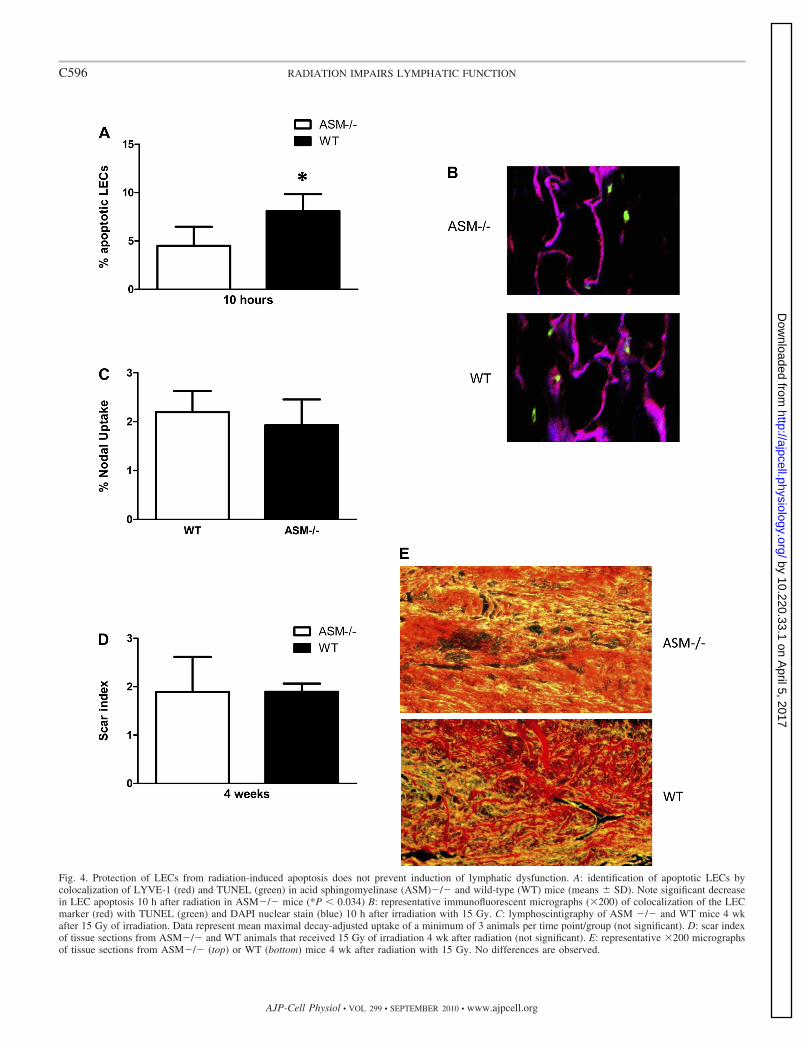
Fig. 4. Protection of LECs from radiation-induced apoptosis does not prevent induction of lymphatic dysfunction. A: identification of apoptotic LECs bycolocalization of LYVE-1 (red) and TUNEL (green) in acid sphingomyelinase (ASM)�/� and wild-type (WT) mice (means � SD). Note significant decreasein LEC apoptosis 10 h after radiation in ASM�/� mice (*P 0.034) B: representative immunofluorescent micrographs (200) of colocalization of the LECmarker (red) with TUNEL (green) and DAPI nuclear stain (blue) 10 h after irradiation with 15 Gy. C: lymphoscintigraphy of ASM �/� and WT mice 4 wkafter 15 Gy of irradiation. Data represent mean maximal decay-adjusted uptake of a minimum of 3 animals per time point/group (not significant). D: scar indexof tissue sections from ASM�/� and WT animals that received 15 Gy of irradiation 4 wk after radiation (not significant). E: representative 200 micrographsof tissue sections from ASM�/� (top) or WT (bottom) mice 4 wk after radiation with 15 Gy. No differences are observed.
C596 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from
demonstrated a nearly fivefold reduction in the expression ofTGFBi in the LY-364947-treated animals (Fig. 6, A and B).TGFBi protein expression is directly related to TGF-�1 activ-ity; therefore, reductions in TGFBi in LY-364947-treated an-imals confirm reduction in TGF-�1 signaling (24). This con-clusion is further supported by immunohistochemical localiza-tion of pSmad3, the downstream mediator of the TGF-�receptor, demonstrating a significant reduction in the number
of pSmad3-positive cells/HPF in LY-364947-treated animalsboth 4 and 12 wk after radiation (Fig. 6, C and D; P 0.03).This finding is consistent with previous reports demonstratingthat TGF-�1 expression is regulated, at least in part, bypositive feedback mechanisms (50).
Short-term inhibition of TGF-�1 signaling reduces radia-tion-induced soft tissue fibrosis and lymphatic dysfunction.Interestingly, even very short-term blockade of TGF-�1 func-
Fig. 5. Radiation causes soft tissue fibrosis and increased transforming growth factor (TGF)-�1 activity. A: calculation of the scar index using polarized lightmicroscopy of Sirius red-stained sections. Mean scar index was calculated from analysis of 3–5 sections/animal/group, and data are presented as means � SD.Note significant and progressive increase in scar index after radiation with 15 or 30 Gy (*P 0.04 for all compared with baseline except for the 15 Gy doseat 12 wk, which approached but did not achieve significance, P 0.08). B: representative photomicrographs (200) of nonirradiated (0 Gy; top) and irradiated(30 Gy; bottom) sections 24 wk after radiation. Note increased fibrosis in irradiated section as demonstrated by more red and orange birefringence. In contrast,note yellow-green birefringence of nonirradiated skin. C: radiation therapy increases the number of phosphorylated Smad3 (pSmad3)-positive cells in tissuesections beginning as early as 4 h after radiation and persisting as long as 24 wk later (*P 0.03 for all compared with baseline). Sections were stained withpSmad3 antibodies, and the number of positively stained cells was assessed in 3–5 HPFs per section in a minimum of 3 animals per time point/group (mean �SD). D: representative 200 micrographs of pSmad3 immunohistochemical staining in nonirradiated (0 Gy; top) and irradiated (30 Gy; bottom) sections 4 h afterradiation. Note that nonirradiated sections demonstrate virtually no positive dermal cells, while large numbers of positively stained cells are noted in the irradiatedtissue section.
C597RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

tion with LY-364947 after irradiation with 15 Gy significantlydecreased soft tissue fibrosis compared with control animals.The scar index of irradiated/LY-364947-treated animals wasmore than twofold less than vehicle treated/irradiated animalseven as long as 12 wk after radiation (Fig. 7, A and B; P 0.01). Short-term blockade of TGF-�1 function was associatedwith decreased tail volume 4 and 12 wk after radiation therapy,although this effect was not statistically different from vehiclecontrol-treated animals (Fig. 7C). This finding is not unexpectedsince in our earlier experiments we demonstrated that radiationonly caused mild, acute increases in tail volume measurementsthat resolved between 4 and 12 wk after treatment. Interestingly,however, we found that lymphatic function as assessed by lym-phoscintigraphy was significantly improved in animals treatedwith LY-364947 compared with control animals at both timepoints evaluated (Fig. 7D). On average, nodal uptake was nearly
4-fold and 2.5-fold higher in the LY-364947-treated animals 4 and12 wk after treatment, respectively, compared with control ani-mals (P 0.0001 and P 0.028, respectively). Image analysis oflymphoscintigraphic data demonstrated grossly increased uptakeof radiolabeled colloid in the lymph nodes of animals treated withLY-364947 compared with vehicle control-treated animals(Fig. 7E).
Improved lymphatic function in LY-364947-treated animalswas not due to a protective effect on LECs since quantificationof dermal LECs did not show significant differences in theLY-364947- or vehicle-treated animals (Fig. 7F). In addition,TGF-�1 blockade did not significantly alter LEC apoptosiswhen evaluated 10 h after treatment with 15 Gy (Fig. 7, G andH). Similarly, treatment of isolated LECs with recombinantTGF-�1 (10 ng/ml) did not result in increased rates of apopto-sis compared with controls.
Fig. 6. Treatment with LY-364947, a small-molecule inhibitor of TGF-�1 receptor (TGF-BR)I, effectively blocks TGF-�1 signaling. A and B: Western blot (A)and quantification (B) for TGF-�-induced (TGFBi) in vehicle- and LY-364947-treated animals 1 wk after irradiation with 15 Gy. Each lane represents proteinisolated from an individual animal. Note marked reduction (nearly 5-fold decrease) in TGFBi protein expression in LY-364947-treated animals, indicatinginhibition of TGF-�1 signaling (*P 0.02). C: calculation of the number of pSmad3-stained cells in 3–5 sections/animal in a minimum of 3 animals per group4 and 12 wk after irradiation with 15 Gy (mean � SD). Note significant and persistent decrease in the number of pSmad3-positive cells in animals treated withLY-364947 for 1 wk (*P 0.03). D: representative 200 micrographs of tissue sections from animals treated with vehicle (top) or LY-364947 (bottom) for1 wk after irradiation with 15 Gy. Tissue sections were harvested 12 wk after radiation therapy. Note marked decrease in the number of pSmad3-stained cellsin the LY-364947-treated animals.
C598 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

Fig. 7. Short-term inhibition of TGF-�1 signaling reduces radiation-induced soft tissue fibrosis and lymphatic dysfunction. A: scar index of tissue sections fromanimals that received 15 Gy of irradiation and were treated with vehicle or LY-364947 for 1 wk after irradiation (mean � SD scar index). Tissues were harvested4 or 12 wk after radiation. Note marked decrease in scar index in LY-364947-treated animals compared with control animals at both time points evaluated (*P 0.002, #P 0.02). B: representative 200 micrographs of tissue sections from animals treated with vehicle (top) or LY-364947 (bottom) 12 wk after radiationwith 15 Gy. Note decreased scarring as represented by decreased red-orange and increased yellow-green birefringence in the LY-364947-treated section. C: tailvolume measurements in animals treated with vehicle or LY-364947 4 or 12 wk after irradiation with 15 Gy (mean � SD). Treatment with LY-364947 resultedin mild but nonsignificant reduction in tail edema after irradiation. D: lymphoscintigraphy of animals treated with vehicle or LY-364947 4 or 12 wk after 15 Gyof irradiation. Data represent mean maximal decay-adjusted uptake of a minimum of 3 animals per time point/group. Note statistically significant increased lymphnode uptake in LY-364947-treated animals, particularly at the 4 wk time point (*P 0.0001). Statistically significant improved lymphatic function was also notedat the 12 wk time point; however, this difference was less marked than the 4 wk analysis (#P 0.03). E: representative heat maps of 99Tc radioisotope uptakein mice irradiated with 15 Gy and treated with either vehicle (left) or LY-364947 (right) at 4 and 12 wk after irradiation. Red color indicates areas of most intenseradioactivity, and yellow, green, and blue represent progressively lower activities. Arrowheads indicate the site of radiolabeled colloid near the tip of the tail,and arrows indicate uptake in the lymph node basin at the base of the tail. Note far greater uptake in the lymph nodes of animals treated with LY-364947,indicating more efficient lymphatic transport.
C599RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

Short-term inhibition of TGF-�1 signaling may reduceradiation-induced LEC EMT and lymphatic fibrosis.TGF-�1 is a known inducer of endothelial-mesenchymalcell transdifferentiation (EMT) resulting in vascular fibrosis(45– 47). In addition, we have more recently shown (8) thatLECs in animals treated with TGF-�1 or in an environmentcontaining high endogenous expression of TGF-�1 coex-press lymphatic and smooth muscle cell markers (LYVE-1and �-SMA, respectively) in dermal lymphatic capillaries.This is an abnormal phenotype, as dermal lymphatic capil-laries are not associated with pericytes. In addition, lym-phatic vessel fibrosis is a known clinical hallmark oflymphedema resulting in lymphatic vessel obliteration anddysfunction (56). Therefore, to determine whether radiationtherapy caused expression of fibroblast markers by lym-
phatic capillaries, we colocalized podoplanin and �-SMA innonirradiated animals and in animals irradiated with 15 Gyand treated with or without LY-364947 (Fig. 8A). As ex-pected we found no LECs that expressed �-SMA in nonir-radiated animals. Similarly, we found no lymphatic vesselsthat expressed �-SMA in irradiated animals treated withLY-364947. In contrast, we noted that in animals treatedwith vehicle and 15 Gy of irradiation, lymphatic capillariesthat coexpressed �-SMA could be seen in some sections,albeit these represented a minority of lymphatic capillariespresent.
To more directly study the effects of TGF-�1 on LECdedifferentiation and EMT, we treated isolated LECs withrecombinant TGF-�1 (10 ng/ml) and analyzed the expressionof a panel of fibroblast and LEC markers at various time points
Fig. 7—Continued F: quantification of the number of LECs in tissue sections from animals treated with vehicle or LY-364947 4 or 12 wk after 15 Gy ofirradiation (mean � SD). No significant differences were noted. G: identification of apoptotic LECs by colocalization of LYVE-1 and TUNEL (mean � SD;not significant). H: representative immunofluorescent micrographs (200) of colocalization of the LEC marker (red) with TUNEL (green) and DAPI nuclear stain(blue) 10 h after irradiation with 15 Gy. I: flow cytometric TUNEL analysis of isolated LECs demonstrating no induction of apoptosis in response to treatmentwith recombinant human (rh)TGF-�1 (10 ng/ml). Data are presented as means � SD of triplicate experiments.
C600 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from
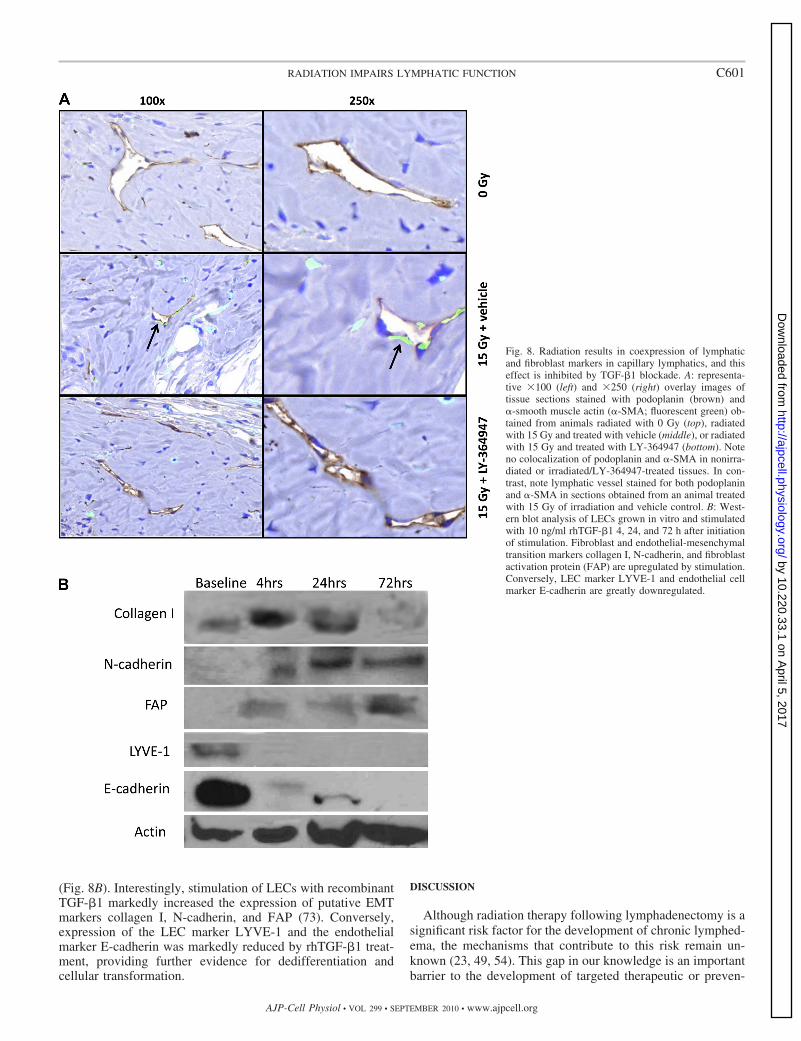
(Fig. 8B). Interestingly, stimulation of LECs with recombinantTGF-�1 markedly increased the expression of putative EMTmarkers collagen I, N-cadherin, and FAP (73). Conversely,expression of the LEC marker LYVE-1 and the endothelialmarker E-cadherin was markedly reduced by rhTGF-�1 treat-ment, providing further evidence for dedifferentiation andcellular transformation.
DISCUSSION
Although radiation therapy following lymphadenectomy is asignificant risk factor for the development of chronic lymphed-ema, the mechanisms that contribute to this risk remain un-known (23, 49, 54). This gap in our knowledge is an importantbarrier to the development of targeted therapeutic or preven-
Fig. 8. Radiation results in coexpression of lymphaticand fibroblast markers in capillary lymphatics, and thiseffect is inhibited by TGF-�1 blockade. A: representa-tive 100 (left) and 250 (right) overlay images oftissue sections stained with podoplanin (brown) and�-smooth muscle actin (�-SMA; fluorescent green) ob-tained from animals radiated with 0 Gy (top), radiatedwith 15 Gy and treated with vehicle (middle), or radiatedwith 15 Gy and treated with LY-364947 (bottom). Noteno colocalization of podoplanin and �-SMA in nonirra-diated or irradiated/LY-364947-treated tissues. In con-trast, note lymphatic vessel stained for both podoplaninand �-SMA in sections obtained from an animal treatedwith 15 Gy of irradiation and vehicle control. B: West-ern blot analysis of LECs grown in vitro and stimulatedwith 10 ng/ml rhTGF-�1 4, 24, and 72 h after initiationof stimulation. Fibroblast and endothelial-mesenchymaltransition markers collagen I, N-cadherin, and fibroblastactivation protein (FAP) are upregulated by stimulation.Conversely, LEC marker LYVE-1 and endothelial cellmarker E-cadherin are greatly downregulated.
C601RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

tative strategies for lymphedema. In the present study wedemonstrate that radiation therapy causes lymphatic dysfunc-tion due to depletion of LECs and soft tissue fibrosis. Similarto the clinical scenario in which lymphedema is rare followingradiation therapy without surgery, we found that radiation inour model resulted in only subtle increases in tail volume thatresolved over time. Interestingly, however, we found thatlymphatic function was markedly decreased after radiationtherapy, implying that radiation decreases the baseline clear-ance of lymphatic fluid without overwhelming the transportcapacity of existing vessels. This finding is supported by thefindings of Mortimer and colleagues (40) demonstrating im-paired dermal lymphatic function as measured by lymphoscin-tigraphy in a porcine model after 18 Gy of radiation. Our studyexpands the findings of Mortimer et al. by demonstrating thatlymphatic transport capacity is decreased for prolonged periodsof time (even 6 mo after radiation). These findings support thehypothesis that the long-term reduction in lymphatic functionresulting from radiation therapy is not usually sufficient tooverwhelm the transport capacity of extremity but when com-bined with additional injury from surgery goes on to causechronic lymphedema.
We hypothesized that one mechanism by which radiationtherapy may decrease lymphatic function is by directly injuringor depleting LECs. This hypothesis is supported by the fact thatmicrovascular endothelial cells are highly sensitive to radiationinjury and undergo apoptosis (43). In fact, depletion of thesecells is thought to be the underlying pathogenic mechanism forgastrointestinal toxicity resulting from radiation therapy (43).In the present study we show that dermal lymphatic vessels arealso susceptible to apoptosis after radiation injury and that thisresponse is, at least in part, due to LEC apoptosis in a processthat is mediated, at least in part, by ASM. Interestingly,however, we found that prevention of LEC apoptosis did nottranslate to improved lymphatic function as assessed by lym-phoscintigraphy, suggesting that other processes such as tissuefibrosis play an overall larger role in the regulation of lym-phatic function after radiation injury.
This proapoptotic effect of radiation was maximal 10 h afterradiation but was apparent even at the earliest time point weselected (4 h). Our findings are also supported by the work ofAbtahian et al. (1), who demonstrated intestinal LEC apoptosis4 h after 12 Gy of total body irradiation. In contrast, however,Sung et al. (57) have previously shown that intestinal andperitumoral lymphatics are resistant to radiation-induced apop-tosis. This difference from our findings may be related to anumber of factors. Most importantly, Sung and colleagues useda dose of 15 Gy and evaluated intestinal LECs. In the presentstudy, while we found a modest increase in LEC apoptosisafter 10 h with 15 Gy (�7% of LECs), 30 Gy caused a moresignificant increase in the number of apoptotic cells. Therelative doses of radiation to the skin are also likely to behigher since they are superficial compared with the intestinalvilli exposed to total body irradiation. It is also possible thatdermal lymphatics are phenotypically different than intestinallymphatics and therefore more susceptible to apoptosis afterionizing radiation injury. This idea is supported by the fact thatintestinal lymphatics, for example, are important in lipid trans-port, whereas dermal lymphatics are not. Furthermore, cellularresponses to radiation injury are known to be tissue dependentand influenced by the surrounding cellular niche. For example,
previous studies have shown that growth factors such asheparanase that are found in the extracellular matrix mayprovide a radioprotective effect for local endothelial cells (19).Thus dermal LECs may be exposed to a microenvironment thatis more inductive of apoptosis than that of intestinal LECs.
Interestingly, we demonstrated that LECs in culture arerelatively radioresistant and undergo apoptosis only when ex-posed to high doses such as 30 Gy. The observed differences inin vivo and in vitro radiation susceptibility may be due todifferences in the microenvironment of the cell in vivo. Thishypothesis is supported by the fact that impairment of lym-phatic vessel function in response to ultraviolet B radiation invivo is mediated by the release of VEGF-A, implying thatradiation therapy not only affects individual cells but alsochanges the cellular microenvironment (28). Furthermore, invivo radiation may cause generation of free oxygen radicals orother toxic substances that may increase the susceptibility toinjury. Kajiya and colleagues (29) have also demonstrated thatultraviolet B radiation therapy markedly downregulates theexpression of VEGF-C, leading to lymphatic dysfunction.Similar changes may contribute to decreased LEC and lym-phatic vessel numbers in our study since activation of VEGFreceptor (VEGFR)-3 is required for LEC survival and prolif-eration (18).
Our in vitro studies also demonstrated that human dermalLECs are sensitive to radiation therapy in terms of cellularsenescence. We found that even relatively low doses of radi-ation caused an increase in the number of senescent cells. Thisis an interesting finding as it suggests that chronic reduction inLEC and lymphatic vessel numbers may be related to prema-ture senescence of LECs. Similar findings have been demon-strated in vitro in a variety of cell types including fibroblasts,hematopoietic stem cells, and mesenchymal stem cells (7, 14,26, 64).
An important finding in our study that supports the conceptthat LECs are susceptible to radiation injury is the fact thatradiation therapy caused a significant depletion of LECs andlymphatic vessels in tissues even up to 6 mo after exposure.This is important as it demonstrates a direct mechanism bywhich radiation therapy decreases the transport capacity of thelymphatic system. Jackowski and colleagues (27) have evalu-ated the effects of radiation therapy on lymphatic vessels, usingbiopsies obtained at various times after treatment obtainedfrom women treated for breast cancer. Similar to our study, theauthors demonstrated that the number of small lymphaticvessels (10 �m) was decreased in the early periods afterradiation (0–5 mo), although this difference was not statisti-cally significant. However, in the patients evaluated between11 and 14 mo the authors found a significant increase in thenumber of small lymphatic vessels in the skin that they termedradiogenic lymphangiogenesis. These differences were nolonger apparent 17� mo after radiation therapy. These findingsimply that the timing of evaluation after radiation therapy isimportant and may be a reason for the differences observed inour study. Furthermore, the differences in the number oflymphatic vessels after radiation between our study and that ofJackowski et al. may be related to the radiation dose orfractionation protocols. Clinically patients were treated with atotal of 40 Gy of radiation delivered in a fractionated manner,whereas animals treated in our study were treated with a singledose. Thus fractionation may have a protective effect for
C602 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

lymphatic vessels. This may be reflected in the finding that thesubset of patients that developed lymphedema failed to mounta lymphangiogenic effect in response to radiation, implyingthat these patients had a more severe injury. Therefore, similarto our findings, patients with a reduction in the number ofcutaneous lymphatic vessels appeared to have decreased lym-phatic function and subsequent lymphedema.
The differences in the observations of Jackowski et al. fromour study may also stem from significant variability in thenumber of lymphatic vessels between patients. In fact, thesedifferences forced the authors to use a treated-to-control ratiofor their analysis that may have altered the findings in someway. This idea is further supported by studies demonstratingthat genetic polymorphisms play a role in an individual’s tissueresponse to radiation (6, 31). Interestingly, polymorphisms inthe gene coding for TGF-�1 have been directly implicated asmodulators of tissue response to radiation (72). The use ofgenetically inbred animals for the study of lymphatic functionobviates these differences and may simplify analysis. Theinherent variability in baseline lymphatic vessel numbers ob-served in clinical studies is interesting as it suggests thatgenetic or other differences may influence the baseline lym-phatic transport capacity and therefore contribute to the risk ofdeveloping lymphedema. This concept is supported by the factthat lymphatic imaging studies have demonstrated that the rateof lymphatic transport in patients is highly variable both aftersurgery and at baseline, and that patients with low baseline andpostsurgical clearance rates have a higher risk of developinglymphedema (55).
Another potential mechanism by which radiation therapycan decrease lymphatic function is soft tissue fibrosis. Fibrosedsoft tissues lose compliance and are characterized by loss ofnormal structures, which are replaced by scar tissue. In fact, theclinical hallmarks of lymphedema are soft tissue and lymphaticvessel fibrosis (56). Anatomic and cadaver studies have shownthat lymphatic vessels undergo fibrosis after lymphadenec-tomy, losing compliance and in many instances becomingcompletely stenosed and nonfunctional (56). We have previ-ously shown (5, 8) that lymphatic fluid stasis is associated withsoft tissue fibrosis and lymphatic dysfunction and that thiseffect is due, at least in part, to increased expression ofTGF-�1. In addition, we (8) and others (41) have shown thatTGF-�1 has antilymphangiogenic effects by inhibiting LECproliferation and tubule formation. These findings are impor-tant since radiation therapy is known to cause tissue fibrosis asa result of chronically increased TGF-�1 expression and maytherefore be additive to the effects of lymphatic stasis inpromoting soft tissue fibrosis and lymphatic dysfunction. Thishypothesis is supported by our findings in ASM�/� mice, inwhich LEC apoptosis was significantly decreased but lym-phatic dysfunction and tissue fibrosis persisted at later timepoints.
Several studies have shown that inhibition of TGF-�1 func-tion either in knockout animals or with small-molecule inhib-itors of TGF-�1 can decrease skin fibrosis resulting fromradiation (17, 36, 69). These findings have been difficult totranslate clinically, however, since long-term inhibition ofTGF-�1 function was required. In fact, the shortest effectivetime for treatment reported was 3–5 wk with the small-molecule inhibitor halofuginone (69). Prolonged TGF-�1 in-hibition can have significant negative side effects due to
immunosuppression and the protean effects of TGF-�1 in avariety of organ systems (33, 68). To circumvent these poten-tial difficulties, we evaluated the effects of short-term (1 wk)TGF-�1 inhibition on radiation fibrosis and lymphatic func-tion. We hypothesized that if the autoregulatory upregulationof TGF-�1 is avoided then long-term sequela of radiation canbe minimized. Indeed, we found that short-term blockade ofTGF-�1 with LY-364947 resulted in marked reduction of theprogressive fibrosis that is associated with radiation injury.This finding is consistent with the study by Wormstone andcolleagues (66), who showed that even a 2-day exposure toTGF-� results in long-term signaling changes in matrix con-traction, transdifferentiation, and fibrosis. In our study,TGF-�1 activity as measured by Smad3 phosphorylation wasattenuated far beyond the duration of therapy, suggesting thatwe had at least partially succeeded in breaking the positivefeedback loop of TGF-�1 activity that is seen with progressivefibrosis (68). These findings directly correlated with betterlymphatic function in the LY-364947-treated animals, consis-tent with the hypothesis that fibrosis is a mediator of lymphaticdysfunction. Interestingly, treatment with LY-364947 did notappear to confer any direct radioprotection to LECs in thepresent study, as evidenced by quantification of LEC depletion.While TGF-�1 has been shown to be inhibit LEC tubuleformation and proliferation, there are no data implicating thisgrowth factor in LEC death. Thus the present study indicatesthat radiation-induced LEC apoptosis is likely independent ofTGF-�1.
Previous studies have shown that TGF-�1 can cause EMTand microvascular vessel fibrosis (45, 46, 68). In addition, wehave previously shown (8) that increased TGF-�1 expression isassociated with coexpression of lymphatic and fibroblast mark-ers in capillary lymphatics. In the present study we found thatradiation therapy also induced the fibroblast marker �-SMA bycapillary LECs and that this effect could be abrogated byshort-term blockade of TGF-�1 function. These findings, to-gether with our in vitro experiments demonstrating thatTGF-�1 increases expression of EMT-associated markers andsimultaneous downregulation of lymphatic and endothelialmarkers, support the hypothesis that radiation therapy maydirectly contribute to lymphatic dysfunction by promotinglymphatic vessel fibrosis as a result of increased TGF-�1expression.
Radioprotective strategies are employed in a variety ofclinical scenarios to decrease the morbidity of radiation (11,20–21). For instance, these approaches have been used todecrease the early morbidity of head and neck cancers such asintraoral blistering and swelling. In addition, radioprotectionhas been effectively used to decrease the incidence of lateradiation-induced sequelae such as osteoradionecrosis andgrowth disturbances in children (12). These strategies are notcommonly used for breast cancer treatment, however, sinceradiation in this setting is thought to be relatively well toler-ated, causing little more than skin blistering and swelling. Ourfindings, however, suggest that radioprotective approachesmay have a role in the prevention of lymphedema by prevent-ing lymphatic dysfunction. This is critical since lymphedemaremains the most dreaded complication of breast cancer treat-ment and is a major source of morbidity for more than 500,000breast cancer survivors (22, 38). Future studies from ourlaboratory will explore these approaches in an effort to identify
C603RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

optimal methods of limiting radiation-induced injury to lym-phatic channels.
ACKNOWLEDGMENTS
The authors thank Dr. Zvi Fuks for his invaluable advice and guidance inthe planning and analysis of the presented studies.
GRANTS
This work was funded in part by grants from the Plastic Surgery Educa-tional Foundation to B. J. Mehrara and T. Avraham and the Breast CancerAlliance.
Technical services provided by the Memorial Sloan-Kettering CancerCenter Small-Animal Imaging Core Facility, supported in part by NationalInstitutes of Health (NIH) Small-Animal Imaging Research Program (SAIRP)Grant R24-CA-83084 and NIH Center Grant P30-CA-08748 are gratefullyacknowledged.
DISCLOSURES
None of the authors has any commercial associations or financial relation-ships that would create a conflict of interest with the work presented in thisarticle.
REFERENCES
1. Abtahian F, Guerriero A, Sebzda E, Lu MM, Zhou R, Mocsai A,Myers EE, Huang B, Jackson DG, Ferrari VA, Tybulewicz V, LowellCA, Lepore JJ, Koretzky GA, Kahn ML. Regulation of blood andlymphatic vascular separation by signaling proteins SLP-76 and Syk.Science 299: 247–251, 2003.
2. Abu-Rustum NR, Alektiar K, Iasonos A, Lev G, Sonoda Y, Aghaja-nian C, Chi DS, Barakat RR. The incidence of symptomatic lower-extremity lymphedema following treatment of uterine corpus malignan-cies: a 12-year experience at Memorial Sloan-Kettering Cancer Center.Gynecol Oncol 103: 714–718, 2006.
3. Ahmed RL, Prizment A, Lazovich D, Schmitz KH, Folsom AR.Lymphedema and quality of life in breast cancer survivors: the IowaWomen’s Health Study. J Clin Oncol 26: 5689–5696, 2008.
4. Arany PR, Flanders KC, DeGraff W, Cook J, Mitchell JB, RobertsAB. Absence of Smad3 confers radioprotection through modulation ofERK-MAPK in primary dermal fibroblasts. J Dermatol Sci 48: 35–42,2007.
5. Avraham T, Clavin NW, Daluvoy SV, Fernandez J, Soares MA,Cordeiro AP, Mehrara BJ. Fibrosis is a key inhibitor of lymphaticregeneration. Plast Reconstr Surg 124: 438–450, 2009.
6. Azria D, Ozsahin M, Kramar A, Peters S, Atencio DP, Crompton NE,Mornex F, Pelegrin A, Dubois JB, Mirimanoff RO, Rosenstein BS.Single nucleotide polymorphisms, apoptosis, and the development ofsevere late adverse effects after radiotherapy. Clin Cancer Res 14: 6284–6288, 2008.
7. Banito A, Rashid ST, Acosta JC, Li S, Pereira CF, Geti I, Pinho S,Silva JC, Azuara V, Walsh M, Vallier L, Gil J. Senescence impairssuccessful reprogramming to pluripotent stem cells. Genes Dev 23: 2134–2139, 2009.
8. Clavin NW, Avraham T, Fernandez J, Daluvoy SV, Soares MA,Chaudhry A, Mehrara BJ. TGF-beta1 is a negative regulator of lym-phatic regeneration during wound repair. Am J Physiol Heart Circ Physiol295: H2113–H2127, 2008.
9. Clavin NW, Fernandez J, Schonmeyr BH, Soares MA, Mehrara BJ.Fractionated doses of ionizing radiation confer protection to mesenchymalstem cell pluripotency. Plast Reconstr Surg 122: 739–748, 2008.
10. Crompton NE. Telomeres, senescence and cellular radiation response.Cell Mol Life Sci 53: 568–575, 1997.
11. Culy CR, Spencer CM. Amifostine: an update on its clinical status as acytoprotectant in patients with cancer receiving chemotherapy or radio-therapy and its potential therapeutic application in myelodysplastic syn-drome. Drugs 61: 641–684, 2001.
12. Damron TA, Spadaro JA, Horton JA, Margulies BS, Strauss JA,Farnum CE. Combinations of radioprotectants spare radiation-induceddamage to the physis. Clin Orthop Relat Res 426: 110–116, 2004.
13. de Magalhaes JP, Chainiaux F, Remacle J, Toussaint O. Stress-inducedpremature senescence in BJ and hTERT-BJ1 human foreskin fibroblasts.FEBS Lett 523: 157–162, 2002.
14. Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, MedranoEE, Linskens M, Rubelj I, Pereira-Smith O. A biomarker that identifiessenescent human cells in culture and in aging skin in vivo. Proc Natl AcadSci USA 92: 9363–9367, 1995.
15. Ehrlich HP, Desmouliere A, Diegelmann RF, Cohen IK, Compton CC,Garner WL, Kapanci Y, Gabbiani G. Morphological and immuno-chemical differences between keloid and hypertrophic scar. Am J Pathol145: 105–113, 1994.
16. Flanders KC, Major CD, Arabshahi A, Aburime EE, Okada MH,Fujii M, Blalock TD, Schultz GS, Sowers A, Anzano MA, Mitchell JB,Russo A, Roberts AB. Interference with transforming growth factor-beta/Smad3 signaling results in accelerated healing of wounds in previouslyirradiated skin. Am J Pathol 163: 2247–2257, 2003.
17. Flanders KC, Sullivan CD, Fujii M, Sowers A, Anzano MA, Arab-shahi A, Major C, Deng C, Russo A, Mitchell JB, Roberts AB. Micelacking Smad3 are protected against cutaneous injury induced by ionizingradiation. Am J Pathol 160: 1057–1068, 2002.
18. Flister MJ, Wilber A, Hall KL, Iwata C, Miyazono K, Nisato RE,Pepper MS, Zawieja DC, Ran S. Inflammation induces lymphangiogen-esis through upregulation of VEGFR-3 mediated by NF-kappaB andProx1. Blood 115: 418–429, 2009.
19. Godder K, Vlodavsky I, Eldor A, Weksler BB, Haimovitz-FreidmanA, Fuks Z. Heparanase activity in cultured endothelial cells. J CellPhysiol 148: 274–280, 1991.
20. Grdina DJ, Garnes BA, Nagy B. Protection by WR-2721 and WR-151327 against late effects of gamma rays and neutrons. Adv Space Res12: 257–263, 1992.
21. Grdina DJ, Murley JS, Kataoka Y. Radioprotectants: current status andnew directions. Oncology 63, Suppl 2: 2–10, 2002.
22. Hayes SC, Janda M, Cornish B, Battistutta D, Newman B. Lymphed-ema after breast cancer: incidence, risk factors, and effect on upper bodyfunction. J Clin Oncol 26: 3536–3542, 2008.
23. Hinrichs CS, Watroba NL, Rezaishiraz H, Giese W, Hurd T, FasslKA, Edge SB. Lymphedema secondary to postmastectomy radiation:incidence and risk factors. Ann Surg Oncol 11: 573–580, 2004.
24. Irigoyen M, Anso E, Salvo E, de las Herrerias JD, Martinez-Irujo JJ,Rouzaut A. TGFbeta-induced protein mediates lymphatic endothelial celladhesion to the extracellular matrix under low oxygen conditions. CellMol Life Sci 65: 2244–2255, 2008.
25. Itahana K, Campisi J, Dimri GP. Methods to detect biomarkers ofcellular senescence: the senescence-associated beta-galactosidase assay.Methods Mol Biol 371: 21–31, 2007.
26. Ito T, Sawada R, Fujiwara Y, Seyama Y, Tsuchiya T. FGF-2 sup-presses cellular senescence of human mesenchymal stem cells by down-regulation of TGF-beta2. Biochem Biophys Res Commun 359: 108–114,2007.
27. Jackowski S, Janusch M, Fiedler E, Marsch WC, Ulbrich EJ, Gais-bauer G, Dunst J, Kerjaschki D, Helmbold P. Radiogenic lymphangio-genesis in the skin. Am J Pathol 11: 338–348, 2007.
28. Kajiya K, Hirakawa S, Detmar M. Vascular endothelial growth factor-Amediates ultraviolet B-induced impairment of lymphatic vessel function.Am J Pathol 169: 1496–1503, 2006.
29. Kajiya K, Sawane M, Huggenberger R, Detmar M. Activation of theVEGFR-3 pathway by VEGF-C attenuates UVB-induced edema forma-tion and skin inflammation by promoting lymphangiogenesis. J InvestDermatol 129: 1292–1298, 2009.
30. Kokunai T, Urui S, Tomita H, Tamaki N. Overcoming of radioresis-tance in human gliomas by p21WAF1/CIP1 antisense oligonucleotide. JNeurooncol 51: 111–119, 2001.
31. Kong FM, Anscher MS, Sporn TA, Washington MK, Clough R,Barcellos-Hoff MH, Jirtle RL. Loss of heterozygosity at the mannose6-phosphate insulin-like growth factor 2 receptor (M6P/IGF2R) locuspredisposes patients to radiation-induced lung injury. Int J Radiat OncolBiol Phys 49: 35–41, 2001.
32. Langley RE, Bump EA, Quartuccio SG, Medeiros D, Braunhut SJ.Radiation-induced apoptosis in microvascular endothelial cells. Br JCancer 75: 666–672, 1997.
33. Leask A, Abraham DJ. TGF-beta signaling and the fibrotic response.FASEB J 18: 816–827, 2004.
34. Loudon L, Petrek J. Lymphedema in women treated for breast cancer.Cancer Pract 8: 65–71, 2000.
35. Mak SS, Yeo W, Lee YM, Mo KF, Tse KY, Tse SM, Ho FP, KwanWH. Predictors of lymphedema in patients with breast cancer undergoing
C604 RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from

axillary lymph node dissection in Hong Kong. Nurs Res 57: 416–425,2008.
36. Martin M, Lefaix J, Delanian S. TGF-beta1 and radiation fibrosis: amaster switch and a specific therapeutic target? Int J Radiat Oncol BiolPhys 47: 277–290, 2000.
37. Martin M, Lefaix JL, Pinton P, Crechet F, Daburon F. Temporalmodulation of TGF-beta1 and beta-actin gene expression in pig skin andmuscular fibrosis after ionizing radiation. Radiat Res 134: 63–70, 1993.
38. McLaughlin SA, Wright MJ, Morris KT, Sampson MR, BrockwayJP, Hurley KE, Riedel ER, Van Zee KJ. Prevalence of lymphedema inwomen with breast cancer 5 years after sentinel lymph node biopsy oraxillary dissection: patient perceptions and precautionary behaviors. J ClinOncol 26: 5220–5226, 2008.
39. Mehrara BJ, Mackool RJ, McCarthy JG, Gittes GK, Longaker MT.Immunolocalization of basic fibroblast growth factor and fibroblast growthfactor receptor-1 and receptor-2 in rat cranial sutures. Plast Reconstr Surg102: 1805–1817, 1998.
40. Mortimer PS, Simmonds RH, Rezvani M, Robbins ME, Ryan TJ,Hopewell JW. Time-related changes in lymphatic clearance in pig skinafter a single dose of 18 Gy of X rays. Br J Radiol 64: 1140–1146, 1991.
41. Oka M, Iwata C, Suzuki HI, Kiyono K, Morishita Y, Watabe T,Komuro A, Kano MR, Miyazono K. Inhibition of endogenous TGF-betasignaling enhances lymphangiogenesis. Blood 111: 4571–4579, 2008.
42. Pain SJ, Purushotham AD, Barber RW, Ballinger JR, Solanki CK,Mortimer PS, Peters AM. Variation in lymphatic function may predis-pose to development of breast cancer-related lymphoedema. Eur J SurgOncol 30: 508–514, 2004.
43. Paris F, Fuks Z, Kang A, Capodieci P, Juan G, Ehleiter D, Haimovitz-Friedman A, Cordon-Cardo C, Kolesnick R. Endothelial apoptosis asthe primary lesion initiating intestinal radiation damage in mice. Science293: 293–297, 2001.
44. Petrek JA, Senie RT, Peters M, Rosen PP. Lymphedema in a cohort ofbreast carcinoma survivors 20 years after diagnosis. Cancer 92: 1368–1377, 2001.
45. Roberts AB, Tian F, Byfield SD, Stuelten C, Ooshima A, Saika S,Flanders KC. Smad3 is key to TGF-beta-mediated epithelial-to-mesen-chymal transition, fibrosis, tumor suppression and metastasis. CytokineGrowth Factor Rev 17: 19–27, 2006.
46. Saika S, Kono-Saika S, Ohnishi Y, Sato M, Muragaki Y, Ooshima A,Flanders KC, Yoo J, Anzano M, Liu CY, Kao WW, Roberts AB.Smad3 signaling is required for epithelial-mesenchymal transition of lensepithelium after injury. Am J Pathol 164: 651–663, 2004.
47. Saika S, Kono-Saika S, Tanaka T, Yamanaka O, Ohnishi Y, Sato M,Muragaki Y, Ooshima A, Yoo J, Flanders KC, Roberts AB. Smad3 isrequired for dedifferentiation of retinal pigment epithelium followingretinal detachment in mice. Lab Invest 84: 1245–1258, 2004.
48. Sakorafas GH, Peros G, Cataliotti L. Sequelae following axillary lymphnode dissection for breast cancer. Expert Rev Anticancer Ther 6: 1629–1638, 2006.
49. Sakorafas GH, Peros G, Cataliotti L, Vlastos G. Lymphedema follow-ing axillary lymph node dissection for breast cancer. Surg Oncol 15:153–165, 2006.
50. Schmid P, Itin P, Cherry G, Bi C, Cox DA. Enhanced expression oftransforming growth factor-beta type I and type II receptors in woundgranulation tissue and hypertrophic scar. Am J Pathol 152: 485–493, 1998.
51. Schonmeyr BH, Wong AK, Soares M, Fernandez J, Clavin N, Me-hrara BJ. Ionizing radiation of mesenchymal stem cells results in dimi-nution of the precursor pool and limits potential for multilineage differ-entiation. Plast Reconstr Surg 122: 64–76, 2008.
52. Singh J, Chuaqui CE, Boriack-Sjodin PA, Lee WC, Pontz T, CorbleyMJ, Cheung HK, Arduini RM, Mead JN, Newman MN, PapadatosJL, Bowes S, Josiah S, Ling LE. Successful shape-based virtual screen-ing: the discovery of a potent inhibitor of the type I TGFbeta receptorkinase (TbetaRI). Bioorg Med Chem Lett 13: 4355–4359, 2003.
53. Sitzia J. Volume measurement in lymphoedema treatment: examination offormulae. Eur J Cancer Care (Engl) 4: 11–16, 1995.
54. Soran A, D’Angelo G, Begovic M, Ardic F, Harlak A, Samuel WieandH, Vogel VG, Johnson RR. Breast cancer-related lymphedema—whatare the significant predictors and how they affect the severity of lymphed-ema? Breast J 12: 536–543, 2006.
55. Stanton AW, Modi S, Bennett Britton TM, Purushotham AD, PetersAM, Levick JR, Mortimer PS. Lymphatic drainage in the muscle andsubcutis of the arm after breast cancer treatment. Breast Cancer Res Treat117: 549–557, 2009.
56. Suami H, Pan WR, Taylor GI. Changes in the lymph structure of theupper limb after axillary dissection: radiographic and anatomical study ina human cadaver. Plast Reconstr Surg 120: 982–991, 2007.
57. Sung HK, Morisada T, Cho CH, Oike Y, Lee J, Sung EK, Chung JH,Suda T, Koh GY. Intestinal and peri-tumoral lymphatic endothelial cellsare resistant to radiation-induced apoptosis. Biochem Biophys Res Com-mun 345: 545–551, 2006.
58. Suzuki K, Mori I, Nakayama Y, Miyakoda M, Kodama S, WatanabeM. Radiation-induced senescence-like growth arrest requires TP53 func-tion but not telomere shortening. Radiat Res 155: 248–253, 2001.
59. Tsai RJ, Dennis LK, Lynch CF, Snetselaar LG, Zamba GK, Scott-Conner C. The risk of developing arm lymphedema among breast cancersurvivors: a meta-analysis of treatment factors. Ann Surg Oncol 16:1959–1972, 2009.
60. Velanovich V, Szymanski W. Quality of life of breast cancer patientswith lymphedema. Am J Surg 177: 184–187, 1999.
61. Ververs JM, Roumen RM, Vingerhoets AJ, Vreugdenhil G, CoeberghJW, Crommelin MA, Luiten EJ, Repelaer van Driel OJ, Schijven M,Wissing JC, Voogd AC. Risk, severity and predictors of physical andpsychological morbidity after axillary lymph node dissection for breastcancer. Eur J Cancer 37: 991–999, 2001.
62. Wang M, Munier F, Araki-Saski K, Schorderet D. TGFBI genetranscript is transforming growth factor-beta1-responsive and cell density-dependent in a human corneal epithelial cell line. Ophthalmic Genet 23:237–245, 2002.
63. Wang Y, Blandino G, Givol D. Induced p21waf expression in H1299 cellline promotes cell senescence and protects against cytotoxic effect ofradiation and doxorubicin. Oncogene 18: 2643–2649, 1999.
64. Wang Y, Schulte BA, Zhou D. Hematopoietic stem cell senescence andlong-term bone marrow injury. Cell Cycle 5: 35–38, 2006.
65. Wendt J, Radetzki S, von Haefen C, Hemmati PG, Guner D, Schulze-Osthoff K, Dorken B, Daniel PT. Induction of p21CIP/WAF-1 and G2arrest by ionizing irradiation impedes caspase-3-mediated apoptosis inhuman carcinoma cells. Oncogene 25: 972–980, 2006.
66. Wormstone IM, Anderson IK, Eldred JA, Dawes LJ, Duncan G.Short-term exposure to transforming growth factor beta induces long-termfibrotic responses. Exp Eye Res 83: 1238–1245, 2006.
67. Wuerzberger-Davis SM, Chang PY, Berchtold C, Miyamoto S. En-hanced G2-M arrest by nuclear factor-kappaB-dependent p21waf1/cip1induction. Mol Cancer Res 3: 345–353, 2005.
68. Wynn T. Cellular and molecular mechanisms of fibrosis. J Pathol 214:199–210, 2008.
69. Xavier S, Piek E, Fujii M, Javelaud D, Mauviel A, Flanders KC,Samuni AM, Felici A, Reiss M, Yarkoni S, Sowers A, Mitchell JB,Roberts AB, Russo A. Amelioration of radiation-induced fibrosis: inhi-bition of transforming growth factor-beta signaling by halofuginone. JBiol Chem 279: 15167–15176, 2004.
70. Yan T, Seo Y, Schupp JE, Zeng X, Desai AB, Kinsella TJ. Methoxy-amine potentiates iododeoxyuridine-induced radiosensitization by alteringcell cycle kinetics and enhancing senescence. Mol Cancer Ther 5: 893–902, 2006.
71. Yang YA, Dukhanina O, Tang B, Mamura M, Letterio JJ, MacGre-gor J, Patel SC, Khozin S, Liu ZY, Green J, Anver MR, Merlino G,Wakefield LM. Lifetime exposure to a soluble TGF-beta antagonistprotects mice against metastasis without adverse side effects. J Clin Invest109: 1607–1615, 2002.
72. Yuan X, Liao Z, Liu Z, Wang LE, Tucker SL, Mao L, Wang XS,Martel M, Komaki R, Cox JD, Milas L, Wei Q. Single nucleotidepolymorphism at rs1982073:T869C of the TGFbeta 1 gene is associatedwith the risk of radiation pneumonitis in patients with non-small-cell lungcancer treated with definitive radiotherapy. J Clin Oncol 27: 3370–3378,2009.
73. Zeisberg M, Neilson EG. Biomarkers for epithelial-mesenchymal transi-tions. J Clin Invest 119: 1429–1437, 2009.
74. Zheng QH, Ma LW, Zhu WG, Zhang ZY, Tong TJ. p21Waf1/Cip1plays a critical role in modulating senescence through changes of DNAmethylation. J Cell Biochem 98: 1230–1248, 2006.
C605RADIATION IMPAIRS LYMPHATIC FUNCTION
AJP-Cell Physiol • VOL 299 • SEPTEMBER 2010 • www.ajpcell.org
by 10.220.33.1 on April 5, 2017
http://ajpcell.physiology.org/D
ownloaded from





























