Embed Size (px)
Citation preview

Plant and Soil 243: 31–42, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
31
Quantitative effects of soil nitrate, growth potential and phenology onsymbiotic nitrogen fixation of pea (Pisum sativum L.)
Anne-Sophie Voisin1,3, Christophe Salon1, Nathalie G. Munier-Jolain1 & Bertrand Ney2
1INRA, Unite de Malherbologie et d’Agronomie, BP 86510, Dijon 21065 cedex, France. 2INRA-INA-PG, UniteEnvironnement et grandes cultures, BP 01, 78850 Thiverval Grignon, France. 3Corresponding author∗
Received 4 July 2001. Accepted in revised form 13 March 2002
Key words: 15N isotope dilution technique, nitrate, pea, Pisum sativum, symbiotic nitrogen fixation
Abstract
The influence of soil nitrate availability, crop growth rate and phenology on the activity of symbiotic nitrogenfixation (SNF) during the growth cycle of pea (Pisum sativum cv. Baccara) was investigated in the field underadequate water availability, applying various levels of fertiliser N at the time of sowing. Nitrate availability inthe ploughed layer of the soil was shown to inhibit both SNF initiation and activity. Contribution of SNF to totalnitrogen uptake (%Ndfa) over the growth cycle could be predicted as a linear function of mineral N content of theploughed layer at sowing. Nitrate inhibition of SNF was absolute when mineral N at sowing was over 380 kg Nha−1. Symbiotic nitrogen fixation was not initiated unless nitrate availability in the soil dropped below 56 kg Nha−1. However, SNF could no longer be initiated after the beginning of seed filling (BSF). Other linear relationshipswere established between instantaneous %Ndfa and instantaneous nitrate availability in the ploughed layer of thesoil until BSF. Instantaneous %Ndfa decreased linearly with soil nitrate availability and was nil above 48 and 34kg N ha−1 for the vegetative and reproductive stages, respectively, levels after which no SNF occurred. Moreover,SNF rate was shown to be closely related to the crop growth rate until BSF. The ratio of SNF rate over crop growthrate decreased linearly with thermal time. Maximum SNF rate was about 40 mg N m−2 degree-day−1, equivalentto 7 kg N ha−1, regardless of the N treatment. From BSF to the end of the growth cycle, the high N requirementsof the crop were supported by both SNF and nitrate root absorption but, of the two sources, nitrate root absorptionseemed to be less affected by the presence of reproductive organs. However, since soil nitrate availability was lowat the end of the growth cycle, SNF was the main source of nitrogen acquisition. The onset of SNF decrease at theend of the growth cycle seemed to be first due to nodule age and then associated to the slowing of the crop growthrate.
Abbreviations: BSF – beginning of seed filling; SNF – symbiotic nitrogen fixation; %Ndfa – percentage of nitrogenderived from air
Introduction
The contribution of symbiotic nitrogen fixation (SNF)to overall nitrogen acquisition by legumes (%Ndfa)is known to be affected by mineral N availability inthe soil. Indeed, the negative effect of nitrate on SNF,even at the smallest concentrations, has been reportedby several authors (Mac Duff et al., 1996 for whiteclover; Waterer and Vessey, 1993 for pea). Although
∗ FAX No: +33 (0)3-80-69-32-62; E-mail: [email protected]
the mechanisms involved are still being debated, soilnitrate availability has been shown to inhibit bothnodule formation and nitrogenase activity (Sprent etal., 1988). However, few studies have attemptedto establish a quantitative relationship between SNFactivity and mineral N availability in the soil (exceptfor Jensen, 1997 for pea) and the critical levels forinhibition have not been yet determined.
Symbiotic nitrogen fixation is known to vary alongthe growth cycle of legumes. The early growth ofpea is sustained by seed N and for some legumes a

32
period of hunger can appear after exhaustion of seedN reserves and before SNF is initiated (Mahon andChild, 1979). However this is quite unlikely to happenfor pea because translocation of fixed N compoundsis simultaneous to nodulation (Sprent and Thomas,1984). Symbiotic nitrogen fixation increases until theearly stages of reproductive development and declinesthereafter as a result of competition for carbon with re-productive organs (Bethlenfalvay and Phillips, 1997b;Jensen, 1987; Pate, 1985). At the end of the growthcycle, the exogenous nitrogen provided by both SNFand nitrate absorption is preferentially diverted to thefilling seeds. However, the high demand for nitro-gen by the filling seeds (Pate, 1985; Sinclair and deWitt, 1975) may not be totally satisfied by exogenousnitrogen supply, which results in remobilisation of ni-trogen from the vegetative organs to the seeds (Pateand Flinn, 1973). Although SNF has often been shownas complementary to root nitrate absorption even tooptimum nitrogen nutrition (Sagan et al., 1993; Voisinet al., 2001), very little is known regarding the relativetiming between SNF activity and root nitrate absorp-tion during the growth cycle. Hypotheses regardingthe ability to take up mineral N during reproductivegrowth are controversial, with some authors statingthat legumes would not effectively assimilate nitrate(Imsande, 1989). Still others observed high fertiliseruse efficiency during the late period of seed filling(Jensen, 1986).
Despite much attention, processes regulating SNFhave not been completely elucidated but they seem toresult from the effect of C and N fluxes in phloem(Baccanamwo and Harper, 1996). Accordingly, thenumber of nodules initiated and fully developed wouldbe regulated by a systematic regulatory mechanism(Caetano-Anolles and Gresshoff, 1991; Markwei andLarue, 1997). Nodule growth would be under thecontrol of assimilate availability (Hacin et al., 1997).Nitrogenase activity would be controlled by adjust-ments in the permeability of the O2 diffusion barrier,which is affected by the combined effects of directnitrate inhibition in the nodules (Faurie and Sous-sana, 1993; Minchin et al., 1986; Vessey and Waterer,1992), feed back of amino compounds circulating inthe phloem (Hartwig et al., 1994; Neo and Layzell,1997; Oti-Boateng, 1994; Parsons et al., 1993) andcarbon status of the plant (Finn and Brun, 1982; Sous-sana and Hartwig, 1996). Symbiotic nitrogen fixationwas also shown to be closely linked to current photo-synthesis (Bethlenfalvay and Phillips, 1977a; Kouchi,1986; Sinclair and de Witt, 1975) and thus correl-
ated to crop growth rate, at least in the early stagesof growth (Jensen, 1987). Moreover the indeterminatefeature of pea results in competition for assimilatesbetween reproductive and vegetative sinks that aresimultaneously present on the plant, through eitherdirect or amino compound-medicated carbon depriva-tion of the nodules, as previously stated. In particular,filling seeds have priority for assimilates at the end ofthe growth cycle (Jeuffroy and Warembourg, 1991).
In this study, variations of nodule activity wereanalysed in relation to assimilate availability to nod-ules that can be modulated both by environmentalfactors (through crop growth rate) and internal plantfactors varying with the phenological stage. The aimwas: (i) to quantify the effect of nitrate availabilityin the soil on SNF of adequately watered field grownpea (Pisum sativum L.), and (ii) to determine the re-spective effects of nitrate availability, growth potentialand phenological stage on changes of SNF along thegrowth cycle.
Material and methods
Site
The experiments were carried out in 1999 and 2000in Dijon (France) on a clayey calcic brown soil. Theclay content and the CACO3 content were, respect-ively, 41.6% and 0.1 ‰in the ploughed layer (0–35cm). At sowing, it contained 27 and 10 kg N ha−1
mineral N in 1999 and 2000, respectively, and the or-ganic nitrogen content was 1.5 ‰. Adequate P and Kfertilisation was applied during the autumn precedingthe experiment. No inoculation was needed as efficientindigenous strains of Rhizobium leguminosarum by.viciae were present in the soil. The field was irrigatedin order to avoid water stress.
Treatments and experimental design
Different levels of mineral N (0, 100, 200 and 400kg N ha−1 in 1999 and 0, 50 and 100 kg N ha−1 in2000) were applied in the form of NH4NO3 at sowingon field pea (Pisum sativum L.) cv. Baccara. The 15Nisotope dilution technique was used to estimate SNF(Duc et al., 1988). For this purpose, 5 kg N ha−1 ofNH4NO3 with 1% 15N atom excess were dissolved inwater and sprayed at a rate of 300 1.ha−1 at sowing onthe whole experimental area. Spring barley (Hordeumvulgare L. cv. Scarlett) was chosen as the non-fixingreference crop in order to provide an estimate of the

33
15N enrichment of soil mineral N. Pea and barley weresupplied with the same amounts of fertiliser and weregrown exactly under the same conditions. The cropswere sown on March 15th, 1999 and March 7th, 2000in randomised split-plot designs with crops as mainplots, N treatments as sub-plots and four replications.Each sub-plot consisted of 10 (pea) or 5 (barley) microplots of six rows 12 m long and spaced 0.20 m. Dens-ities were, respectively, 80 and 340 plants m−2 for peaand barley.
Sampling procedures and measurements
Aerial parts of pea were harvested twice a week, start-ing with the development of the fifth leaf and endingat physiological maturity. Each harvest consisted ofcollecting 1 m of the four central rows. Ten plantswere randomly taken aside for developmental observa-tions (Voisin et al., 2001). Barley was harvested oncea week.
The amount of residual mineral N in the soil wasmeasured for each 20 cm soil layer at sowing for bothyears and at six other dates during the growth cycle in1999 (about once every 2 weeks), for each treatmentand each repetition. Soil samples were cored on thesame plots that were used for shoot harvest by using a58 mm diameter auger.
Biomass, nitrogen concentration and 15N enrich-ment measurements were performed on the shoot ofboth crops. Dry matter was determined after oven dry-ing at 80 ◦C for 48 h. Samples were ground and Nconcentration was determined by the Dumas proced-ure (Allen, 1974). The 15N enrichment was measuredusing a dual inlet mass spectrometer (Fison Isochrom,Micromass, Lyon France). Soil mineral N contentwas determined in a K2SO4 extract (100 ml 0.05NK2SO4 per 20 g of fresh soil, shaken for half anhour), with an autoanalyser (Perstop analytical Inc,Silver Spring, USA) using copper reduction and theGriess Ilosvay reaction for nitrate and the indophenolmethod for ammonium (Keeney and Wilson, 1982).Both measurements were made colorimetrically.
Calculations and statistical analysis
Developmental stages were recorded to identify peri-ods characterised by the different sources and sinksthat are simultaneously present on the plant (Ney andTurc, 1993). Progress through the growing season wasexpressed as cumulative degree-days from emergencewith a 0 ◦C base temperature (Etévé and Derieux,1982; Ney and Turc, 1993).
The percentage of nitrogen that was derived fromfixation (%Ndfa) was calculated using the isotope di-lution principle (Amarger et al., 1977, 1979; Mariottiet al., 1983), specifically: %Ndfa = 100. (δ15N legume– δ15N reference plant) / (εfix δ15N reference plant),where εfix (−1 for pea) is the isotopic fractionationfactor associated with N2 fixation processes (Mariottiet al., 1980, 1983).
Values of 15N enrichments in crops (pea and bar-ley) were corrected considering seed-borne N as an-other source of nitrogen for the plant, as described byJensen et al. (1985):
δ15N corrected = (δ15N crop – (QN seed borne/ QN crop). �15N seed borne))/(1-(QN seed borne /QN crop)), where it is assumed that 50% of the seedborne N was located above ground (QN seed borne)and that QN crop is the quantity of nitrogen measuredin shoots.
Crop growth rate and SNF rate at date dn werecalculated over one week period as follows: (qn−1)-qn+1)/(dn−1-dn+1) where qn is either dry matter or thequantity of nitrogen that comes from SNF as measuredat date dn, expressed in cumulative degree-days fromemergence.
Analysis of variance was performed with GLMprocedure of SAS and means were classified usingthe least significative difference test (LSD) at the 0.05probability level (SAS institute, 1987).
Results and discussion
Changes in soil nitrate availability during the growthcycle
Symbiotic nitrogen fixation is known to be mainly af-fected by soil nitrate, ammonium effects being lessevident or antagonist (Streeter, 1988; Waterer and Ves-sey, 1993). In the field, nodules are held by the primaryroot and by the first order laterals in the upper 30 cm ofthe soil (Tricot, 1993). Even though nitrate inhibitionof SNF probably occurs through systemic regulationthrough the whole root system (Silsbury et al., 1986),more than 70% of pea roots are located in the upper 30cm of the soil (Voisin et al., 2001). Thus, in our study,mineral N effects on SNF were studied consideringspecifically nitrate availability in the upper soil layer.
The change in nitrate content of the 0–40 cm soillayer was plotted against cumulative degree-days fromemergence (Figure 1). The first measurement wasat sowing, at the same time N fertilisation occurred

34
Figure 1. Changes of nitrate content of the 0–40 cm upper soil layeracross time expressed in cumulative degree-days from emergencefor the four fertiliser N treatments at sowing (0 N (�) 100 N (�)200 N (�) 400 N(♦). Each point is the mean value of four replicates.Vertical bars represent LSD (p<0.05).
through the addition of NH4NO3. For every treat-ment, nitrate availability in the soil first increased until300 degree-days and decreased afterwards. The earlyincrease was assumed to be the result of: (i) miner-alization processes of organic matter in the ploughedlayer, as shown by the 0 N treatment, and (ii) transfon-nation of the ammonium added through N fertilisationinto nitrate. After emergence, soil nitrate content de-creased with nitrate uptake by the crop until harvest.At harvest, the quantity of nitrate left in the soil wassimilar for the 0 N, 100 N and 200 N treatments(around 8 to 10 kg N ha−1), while it was slightlyhigher for the 400 N treatment (30 kg N ha−1.
Overall effect of soil nitrate availability on symbioticnitrogen fixation
Total N accumulation in the crop over the growthcycle was not significantly different among treatmentswithin each year, except for the 400 N treatment (Table1). The contribution of SNF to total N uptake (%Ndfa)decreased with the N fertilisation level, from 84% forthe 0 N treatment down to nil for the 400 N treat-ment. Therefore, the amount of N derived from SNFdecreased with the N treatment, from a maximum of238 kg N ha−1 for the ON treatment (Table 1).
To establish a relationship between %Ndfa over thegrowth cycle and the total amount of nitrate availableto the crop along the growth cycle, mineral N (ni-trate and ammonium) measured in the ploughed soillayer at sowing was chosen as an indicator of cumu-lated nitrate availability over the growth cycle. Bothnitrate and ammonium were considered because am-monium is normally transformed into nitrate with time
Figure 2. Effect of mineral N availability in the ploughed layerat sowing (N) on the percentage of nitrogen derived from SNF(%Ndfa) measured at harvest (�) for 1999 and 2000 experiments.Each point is the mean value of four replicates. Vertical bars repres-ent two standard error deviations.— %Ndfa = –0.230 (N) ± 87.3 18(r2 = 0.976). (�) Points from Jensen, 1987 for comparison.
(Nicolardot et al., 1996) and thus accounts for a com-ponent of nitrate availability at the growth cycle level.Moreover, the amounts of ammonium measured atharvest were negligible suggesting that it was entirelytransformed into nitrate across the growth cycle (datanot shown). As nitrate leaching might occur becauseof the high doses of fertiliser added at sowing (200 Nand 400 N treatments), the amount of nitrate being lostbetween sowing and the first soil measurement was re-trieved for those treatments, using N budget at the croplevel. A linear relationship was established between%Ndfa over the growth cycle and soil mineral N in theploughed layer at sowing (Figure 2), demonstrating aquantitive limitation of SNF by soil mineral N avail-ability. Inhibition was absolute when soil mineral Navailability at sowing was over 380 kg N ha−1. Theestablished relationship (Figure 2) is a way of quanti-fying globally the effect of soil nitrate availability onthe potential of SNF, i.e. throughout the growth cycleof the crop. In cases where limitation of SNF by nitratehappens to be higher than model predictions, addi-tional sources of inhibition, such as water limitationor soil composition, must be investigated.
However, it has to be emphasised that this relation-ship probably depends on soil type (especially claycontent), in relation with the climate, that togetherdetermine the quantity of N that can potentially bemineralised from the soil organic matter during thegrowing season. As this relationship was establishedin Dijon (temperate climate) on a clayey calcic brownsoil, its applicability to other systems may be restric-

35
Table 1. Contribution of symbiotic nitrogen fixation to total crop nitrogenacquisition over the growth cycle as influenced by N-fertiliser supply at sow-ing (Values represent mean ± SE. Mean comparison were performed insideeach phase with LSD at p <0.05; values followed by the same letter are notsignificantly different)
N treatment N derived from fixation N accumulated by the crop
kg N ha−1 % kg N ha−1
1999 experiment
0 N 84 ± 2 (A) 284 ± 28 (A)
100 N 60 ± 2 (B) 310 ± 17 (A)
200 N 42± 8 (C) 282 ± 25 (A)
400 N 0 ± 0 (D) 264 ± 29 (B)
2000 experiment
0 N 83 ± 1 (A) 262 ± 30 (A)
50 N 70 ± 3 (B) 246 ± 24 (A)
100 N 55 ± 5 (C) 248 ± 6 (A)
ted. Nevertheless, these results are consistent withthose of Jensen (1987) in Denmark on a loamy sandsoil (Figure 2).
Changes in symbiotic nitrogen fixation activity duringthe growth cycle
The short time interval between measurements al-lowed a precise analysis of the changes in SNF alongthe growth cycle (Figure 3). Symbiotic nitrogen fix-ation started early in the growth cycle for the 0 Ntreatment (240 degree-days) whereas the onset of SNFwas delayed for plants that had received fertiliser Nat sowing: it started around 460 and 760 degree-daysfor the 100 N, 200 N, respectively while SNF wasalways nil for the 400 N treatment. The maximumSNF rate (40 mg N m−2 degree-days−1) was ratherunchanged by the fertiliser N treatment, as found byJensen (1987). Usually, SNF increases up to a plateauduring flowering and sharply drops to almost nil dur-ing seed maturation (Armstrong et al., 1994; Herdinaand Silbury, 1990; Bethlenfalvay and Phillips, 1977b).In our experiment (Figure 3), for the 0 N treatment,the plateau was not reached until the beginning ofseed filling and SNF activity occurred during the seedfilling period (Figure 3C), suggesting that it can bemaintained late in the growth cycle. Several authorshave also observed that SNF was still active duringreproductive stages and only ceased at physiologicalmaturity (Bergerson, 1992 for soybean, Jensen, 1987;Sparrow et al., 1995; Vikman and Vessey, 1992)
Moreover, a second peak of SNF was sometimesreported during reproductive stages if optimum tem-perature and water conditions occurred: Sparrow et al.(1995) and Jensen (1987) for pea, Rennie and Kemp(1980) and Franco et al (1979) for Phaseolus vulgaris,Dean and Clark (1980) for fababean. This happenedeven after a decline in SNF during seed filling, whichwas attributed to a carbon shortage for nodules arisingpresumably from the combined effects of a decreasedgrowth rate and competition with reproductive organs.Thus even if SNF always decreased during reproduct-ive growth, it could have been maintained as longas photosynthesis was not limited by environmentalconditions, and provided there were non-stressing soilconditions.
However, origins of SNF variations across thegrowth cycle remain unclear. Symbiotic nitrogen fix-ation may be affected by both environmental andinternal factors: high levels of nitrate inhibit SNF andit is also widely accepted that SNF activity is directlyrelated to current photosynthesis (Bethlenfalvay andPhillips, 1977a; Kouchi, 1986) and N demand by theplant as determined by plant growth (Hartwig et al.,1994). In addition, carbon allocation to nodules canbe modulated along the growth cycle by the presenceof other organs (Jeuffroy and Warembourg, 1991).In this adequately watered experiment, we thereforemade the hypothesis that three main factors could po-tentially limit the level of SNF: nitrate availability,growth potential, and presence of other competing or-gans for carbon, as indicated by phenology. Moreover,

36
Table 2. Values of the slopes of the linear regressions of symbiotic nitro-gen fixation rates against time (mg N m−2 cumulative degree-days fromemergence−2). (Mean comparison were performed inside each phase withLSD at p <0.05; values followed by the same letter are not significantlydifferent)
N treatment Phase I Phase 11a Phase 11b Phase III
kg N ha−1
0 N + 0.092 (B) – 0.195 (B) + 0.127 (A) – 0.079 (A)
100 N + 0.160 (A) – 0.099 (A) + 0.101 (A) – 0.064 (A)
200 N + 0.102 (B) – – – 0.154 (B)
Figure 3. Changes in SNF rates ( — ) and nitrate content in the 0–40upper soil layer ( ...... ) along the growth cycle for ON treatment (A),100 N treatment (B) and 200 N treatment (C). F: Flowering; BSF:Beginning of Seed Filling; ESF: End of Seed Filling (physiologicalmaturity). Each point is the mean value of four replicates. Lettersabove the solid lines denote the different periods of changes insymbiotic nitrogen fixation rates.
the level SNF activity obviously also depends uponnodule developmental stage.
Time course of SNF of the 0 N, 100 N and 200N treatments were compared using SNF rates cal-culated over one week period (Figure 3A, 3B and3C, respectively). Linear regressions were performedbetween SNF rates and cumulative degree-days fromemergence and slopes were compared using varianceanalysis (Table 2). Even if data concerning nodule de-velopment were not available, changes in SNF activitywere considered as indicators of the different periodsof the nodule lifecycle:
Period I: the first period of increase of SNF ratespanned nodule morphogenesis (I, Figure 3) and rep-resented a nodule formation period.
Period II: for the 0 N and 100 N treatments only,the first phase associated with a decrease (IIa) and thesecond phase associated with an increase (IIb, Figure3) were analysed and compared.
Period III: the final phase, associated with a finalSNF decrease, was attributed to nodule senescence(III, Figure 3).
Obviously, nitrogenase activity presumably startedas soon as nodules were established, and ended atphysiological maturity, but nodule activity was con-sidered to be independent from nodule morphogenesisand from nodule senescence during the period thatstarted at the end of phase I and ended at the beginningof phase III. These stages were simultaneous for the200 N treatment (Figure 3C).
The maximum potential of SNF during a givenperiod of the growth cycle was assumed to correspondto the total amount of N that was accumulated by thecrop during this period, following the hypothesis thatSNF could potentially sustain total N supply of theplant. The instantaneous %Ndfa was then defined asthe ratio of the instantaneous SNF rate over instantan-eous total N uptake rate. As such, the instantaneous

37
Figure 4. Symbiotic nitrogen fixation rate (A), crop growth rate (B)and instantaneous %Ndfa (C) as affected by mineral N availability inthe soil along the growth cycle. (0 N (�) 100 N (�) 200 N (� 4000N (♦). F: Flowering; BSF: Beginning of Seed Filling; ESF: End ofSeed Filling (physiological maturity). Each point is the mean valueof four replicates.
%Ndfa reflected the balance between SNF and min-eral N absorption. Since N accumulation is known tobe closely linked to biomass accumulation (Lemaireet al., 1997), this expression of SNF was consideredto be independent from crop growth rate. Values be-low 100% were interpreted as a limitation of SNFdue either to soil nitrate availability or to competi-tion for carbon allocation between nodules and theremaining sinks or to nodule internal factors. Instant-aneous %Ndfa was plotted against time (Figure 4C)and compared with the SNF rates and crop growthrates (Figure 4A and 4B). Three characteristic phasescould be distinguished:
Phase 1 (0–600 degree-days): instantaneous%Ndfa increased with thermal time for the 0 N treat-
ment, while it started to increase from 450 degree-daysfor the 100 N treatment, after a lag of full inhibi-tion. SNF was fully inhibited fro the 200 N treat-ment throughout this phase. During this vegetativeperiod, the differences of inhibition of SNF betweentreatments could be incurred to nitrate.
Phase 2 (600–1100 degree-days): this periodspanned the end of the vegetative stage, floweringand the BSF phase. The inhibition of SNF of the 200N treatment was alleviated during this period (Figure4C), as was observed earlier in the growth cycle forthe other treatments, probably in relation with the re-duction of soil nitrate availability (Figure 3). Duringthis phase, limitation was noted even though SNF ofthe 0 N and the 100 N treatments should no longer belimited by soil nitrate availability since nitrate limit-ation has been shown to decrease down to nil duringthe preceding period and since soil nitrate availabilitywent on decreasing since then (Figure 1). However,as the expression of instantaneous %Ndfa discardedany effect of the growth potential, it was probablythe consequence of an increase of nitrate assimilation,which replaced SNF that was limited by the simultan-eous low crop growth rate (Figure 4B) between 600and 900 degree-days. Probably because soil nitrateavailability was higher for the 100 N treatment (Figure1), the contribution of root mineral N absorption washigher for this treatment than for the 0 N treatment(Figure 4C). The end of the period included the endof flowering and the very beginning of seed filling,when competition for assimilates might have occurredbetween developing seeds and SNF activity. As a con-sequence, occurrence of nitrate root absorption mayhave resulted from the high N demand.
Phase 3 (1100–1400 degree-days): this periodwas the time when all seeds were filling and endedwith physiological maturity. Instantaneous %Ndfa de-creased strongly for all the treatments (Figure 4C)and was attributed to senescence of nodule tissuesas the SNF rates roughly linearly decreased down to0 with thermal time (Figure 4A). although relativelyhigh contributions of nitrate absorption to total N up-take were observed, they actually corresponded to verysmall accumulations of N since crop growth rates werevery low at this stage (Figure 4B).
Hence, SNF appeared to be mainly limited bysoil nitrate availability at the beginning of the growthcycle whereas it seemed to be more related to cropgrowth rate and to the presence of other competingsinks for carbon afterwards, and eventually to nodulesenescence.

38
Onset of the potential of symbiotic nitrogen fixationactivity
Nitrate inhibition of SNF resulted in a delayed onset ofSNF (Figure 3); it started late in the vegetative phasefor the 100 N treatment and at flowering for the 200N treatment. The rate of nodule establishment, as in-dicated by the slope of the first increase in SNF rate(Figure 3), differed with the N treatments, with that ofthe 100 N treatment being the highest, in comparisonwith the 0 N and 200 N treatments which were notsignificantly different from one another (Table 2). Inthe case of the 100 N treatment, SNF set up took placeduring the end of the vegetative phase (Figure 3B),which is a period of high crop growth rate potential(Figure 4B), whereas there must have been competi-tion between shoot and nodulated root settlement forthe 0 N treatment (Figure 3A). Considering the 200N treatment, onset of SNF happened during the endof flowering and beginning of seed filling phase (Fig-ure 3C), resulting in competition for carbon betweenreproductive organs and nodules.
The onset of SNF was related to soil nitrate avail-ability (Figure 3) in order to determine the thresholdunder which the inhibition of SNF could be allevi-ated. Symbiotic nitrogen fixation started when nitrateavailability dropped below 30, 56 and 33 kg N ha−1
for the ON, 100 N and 200 N treatments, respect-ively. For the 100 N treatment, SNF set up took placeduring a period when nitrate availability was prob-ably the only limiting factor to nodule growth, as hasbeen previously suggested above. Hence, the 100 Ntreatment value could be assumed to be the actualthreshold under which SNF set up was possible. Forthe 0 N treatment, residual mineral N at sowing aver-aged 27 kg N ha−1 so that SNF set up was assumedto have never been completely inhibited by solid ni-trate. Moreover, there obviously must be a lag phase atthe beginning of the growth cycle when root morpho-genesis and crop establishment has priority towardsnodule set up (Sprent and Thomas, 1984). In ourstudy, using the 0 N treatment, the duration of this lagphase could be estimated to last about 235 cumulat-ive degree-day starting from emergence. SNF of the200 N treatment started only when nitrate availabilityin the soil was under 33 kg ha−1 (Figure 3C). Com-pared to the 100 N treatment, there may have beenan additional delay (about 130 degree-days) resultingfrom low carbon availability for nodules associatedto the low crop growth rate between 600 and 700degree-days and/or to lower sink strength of nodules
Figure 5. Instantaneous %Ndfa as a function of instantaneous ni-trate availability in the upper 0–40 cm soil layer (N) for thebegetative (closed symbols) and reproductive stages (open symbols)until BSF (0 N (�) 100 N (�) 200 N (�) 400 N (♦)) Each pointis the mean value of four replicates. — % inhibition –2.96 (N) ±142.77 (r2 = 0.54). ...... % inhibition –3.27 (N) + 110.19 (r2 = 0.92).
at this stage (Figure 4B). In contrast, SNF was neverobserved for the 400 N treatment, even when nitrateavailability in the soil decreased down to 30 kg N h−1
around the middle of BSF (Figure 1). Hence, priorityof filling seeds for carbon (Jeuffroy and Warembourg,1991) may have been the reason that prevented theonset of nodulation at that time.
Therefore, we suggest that, the onset of SNF wasmade possible when soil nitrate was sufficiently low.The rate of establishment of SNF potential could bemodulated by carbon availability to the nodules, asdetermined by crop growth rate and phenology.
Modulation of symbiotic nitrogen fixation activity
As nitrate was identified as the major limiting factor ofSNF until the beginning of seed filling, instantaneous%Ndfa was plotted against instantaneous nitrate avail-ability in the ploughed layer, before this stage and forall the treatments of 1999 experiment (Figure 5). Twolinear relationships were established, distinguishingpoints belonging to the vegetative stage (V) and pointsbelonging to the flowering and BSF stage (R). Instant-aneous %Ndfa decreased linearly from 100% to 0 asnitrate availability varied from 0 up to about 48 and 34kg N ha−2 for the V and R phases, respectively (Fig-ure 5) and remained full thereafter. These relationshipsheld regardless of the N treatment, nitrate limitation ofSNF being highest in presence of reproductive organ(R). Nitrate levels of total SNF inhibition are con-sistent with the previously discussed results about theonset of fixation (Figure 3). Even if the effects ofsoil nitrate availability on either SNF set up or ni-
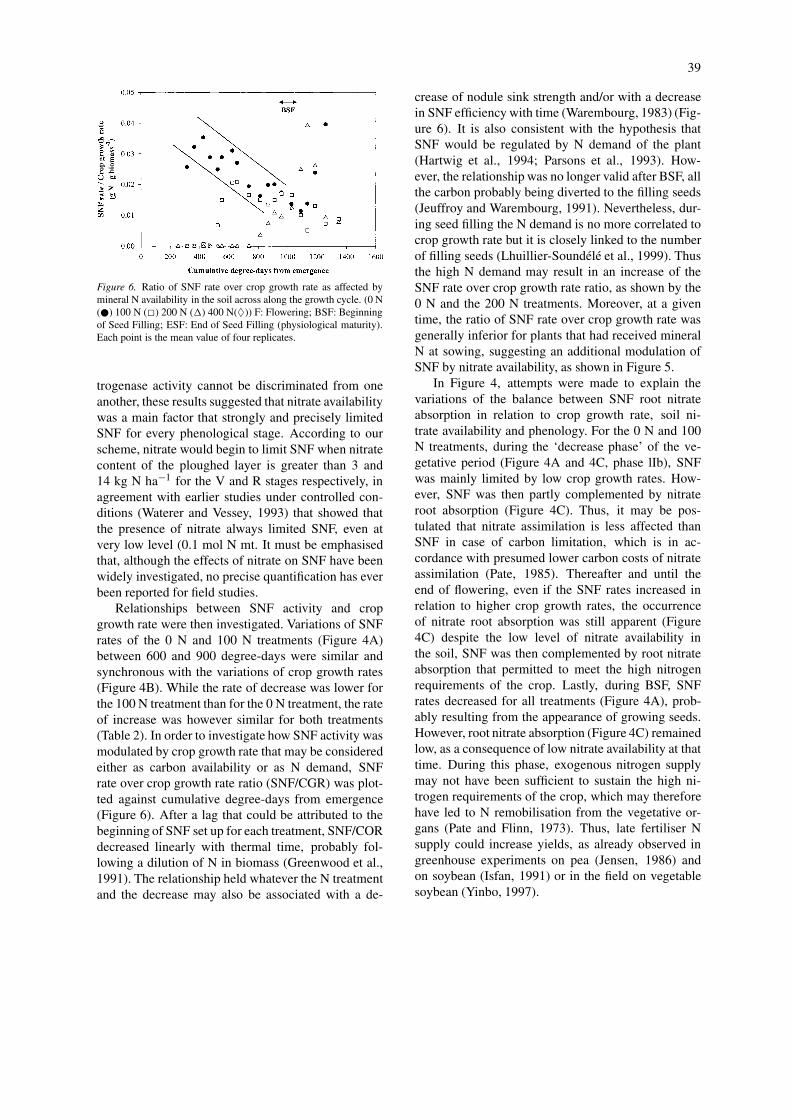
39
Figure 6. Ratio of SNF rate over crop growth rate as affected bymineral N availability in the soil across along the growth cycle. (0 N(�) 100 N (�) 200 N (�) 400 N(♦)) F: Flowering; BSF: Beginningof Seed Filling; ESF: End of Seed Filling (physiological maturity).Each point is the mean value of four replicates.
trogenase activity cannot be discriminated from oneanother, these results suggested that nitrate availabilitywas a main factor that strongly and precisely limitedSNF for every phenological stage. According to ourscheme, nitrate would begin to limit SNF when nitratecontent of the ploughed layer is greater than 3 and14 kg N ha−1 for the V and R stages respectively, inagreement with earlier studies under controlled con-ditions (Waterer and Vessey, 1993) that showed thatthe presence of nitrate always limited SNF, even atvery low level (0.1 mol N mt. It must be emphasisedthat, although the effects of nitrate on SNF have beenwidely investigated, no precise quantification has everbeen reported for field studies.
Relationships between SNF activity and cropgrowth rate were then investigated. Variations of SNFrates of the 0 N and 100 N treatments (Figure 4A)between 600 and 900 degree-days were similar andsynchronous with the variations of crop growth rates(Figure 4B). While the rate of decrease was lower forthe 100 N treatment than for the 0 N treatment, the rateof increase was however similar for both treatments(Table 2). In order to investigate how SNF activity wasmodulated by crop growth rate that may be consideredeither as carbon availability or as N demand, SNFrate over crop growth rate ratio (SNF/CGR) was plot-ted against cumulative degree-days from emergence(Figure 6). After a lag that could be attributed to thebeginning of SNF set up for each treatment, SNF/CORdecreased linearly with thermal time, probably fol-lowing a dilution of N in biomass (Greenwood et al.,1991). The relationship held whatever the N treatmentand the decrease may also be associated with a de-
crease of nodule sink strength and/or with a decreasein SNF efficiency with time (Warembourg, 1983) (Fig-ure 6). It is also consistent with the hypothesis thatSNF would be regulated by N demand of the plant(Hartwig et al., 1994; Parsons et al., 1993). How-ever, the relationship was no longer valid after BSF, allthe carbon probably being diverted to the filling seeds(Jeuffroy and Warembourg, 1991). Nevertheless, dur-ing seed filling the N demand is no more correlated tocrop growth rate but it is closely linked to the numberof filling seeds (Lhuillier-Soundélé et al., 1999). Thusthe high N demand may result in an increase of theSNF rate over crop growth rate ratio, as shown by the0 N and the 200 N treatments. Moreover, at a giventime, the ratio of SNF rate over crop growth rate wasgenerally inferior for plants that had received mineralN at sowing, suggesting an additional modulation ofSNF by nitrate availability, as shown in Figure 5.
In Figure 4, attempts were made to explain thevariations of the balance between SNF root nitrateabsorption in relation to crop growth rate, soil ni-trate availability and phenology. For the 0 N and 100N treatments, during the ‘decrease phase’ of the ve-getative period (Figure 4A and 4C, phase lIb), SNFwas mainly limited by low crop growth rates. How-ever, SNF was then partly complemented by nitrateroot absorption (Figure 4C). Thus, it may be pos-tulated that nitrate assimilation is less affected thanSNF in case of carbon limitation, which is in ac-cordance with presumed lower carbon costs of nitrateassimilation (Pate, 1985). Thereafter and until theend of flowering, even if the SNF rates increased inrelation to higher crop growth rates, the occurrenceof nitrate root absorption was still apparent (Figure4C) despite the low level of nitrate availability inthe soil, SNF was then complemented by root nitrateabsorption that permitted to meet the high nitrogenrequirements of the crop. Lastly, during BSF, SNFrates decreased for all treatments (Figure 4A), prob-ably resulting from the appearance of growing seeds.However, root nitrate absorption (Figure 4C) remainedlow, as a consequence of low nitrate availability at thattime. During this phase, exogenous nitrogen supplymay not have been sufficient to sustain the high ni-trogen requirements of the crop, which may thereforehave led to N remobilisation from the vegetative or-gans (Pate and Flinn, 1973). Thus, late fertiliser Nsupply could increase yields, as already observed ingreenhouse experiments on pea (Jensen, 1986) andon soybean (Isfan, 1991) or in the field on vegetablesoybean (Yinbo, 1997).

40
Decrease of symbiotic nitrogen fixation activity at theend of the growth cycle
Nodule senescence was characterised by the last de-crease in SNF rate at the end of the growth cycle forall treatments (Figure 3, Phase III). While SNF de-crease was initiated at the beginning of seed fillingfor the 0 N and 100 N treatments, which had sim-ilar decreasing slopes (Table 2), the onset of SNFdecrease of the 200 N treatment was delayed untilthe end of seed filling (Figure 3C) and the rate ofSNF decrease of the 200 N treatment was the highest(Table 2). Nodule activity ceased simultaneously forall the treatments at physiological maturity (around1400 cumulative degree-days from emergence). Al-though SNF rates continued to decrease during theseed filling period for the 0 N and 100 N treatments,a peak of SNF was observed during this period for the200 N treatment and attributed to newly formed, andthus more effective nodules whereas those of the 0 Nand the 100 N treatments were probably older, imply-ing larger carbon costs of SNF (Warembourg, 1983).This suggested that the onset of nodule senescencewould rather be due to tissue-initiated processes ratherthan to carbon deprivation of the nodules. However,the decrease in SNF rates for all the treatments downto nil at the end of the growth cycle was probablyaccelerated by the sharp decrease of the crop growthrate, resulting in carbon shortage for nodules.
Conclusion
According to the growth cycle of pea, two main peri-ods may be distinguished: until the beginning of seedfilling, SNF seemed to be affected mainly by soil ni-trate availability and variations of the crop growth po-tential while afterwards, the increase of sink strengthof reproductive organs and the onset of nodule tissuesenescence appeared to be the major limiting factors toSNF. This study also provided precise quantificationof the relationships between soil nitrate availability,crop growth rate and SNF under field conditions. Ourresults demonstrated that, under adequately wateredconditions, the contribution of SNF to overall nitro-gen acquisition can be predicted either, at the growingseason level, as a function of the amount of mineral Npresent at sowing, or at any given later time, as a func-tion of the instantaneous mineral N availability. Thetiming of onset of SNF moreover depends on the levelof soil nitrate availability. Our results also showed that
the level of SNF fixation rate is likely to vary accord-ing to the carbon that is allocated to the nodules, inrelation to endogenous factors, current photosynthesis,crop growth rate and other competing sinks for carbon.
Acknowledgements
Our grateful thanks are due to M. Donguy, V. Durey,J. Gonthier, C. Jeudy, and P. Mathey for their excellenttechnical assistance. We also thank J. Lévêque andMJ. Milloux for 15N isotopic analysis (Geosol, Uni-versité de Bourgogne). This work was partly foundedby INIRA, UNIP and Conseil Régional de Bourgogne.
References
Allen S E 1974 Chemical Analysis of Ecological Materials. JohnWiley and Sons, New York.
Amarger N, Mariotti A and Mariotti F 1977 essai d’estimation dutaux d’azote fixé symbiotiquement chez le lupin par le traçageisotopique naturel. C. R. Acad. Sc. 284, 2179–2182.
Amarger N, Mariotti A, Mariotti F, Durr J C, Bourguignon C andLagacherie B 1979 Estimate of symbiotically fixed nitrogen infield grown soybean using variations in 15N natural abundance.Plant and Soil 52, 269–280.
Armstrong E L, Pate J S and Unkovich m J 1994 Nitrogen balanceof field pea crops in South West Australia, studied using the 15Nnatural abundance technique. Aust. J. Plant Physiol. 21, 533–549.
Bacanamwo m and Harper J E 1996 Regulation of nitrogenase activ-ity in Bradyrhizobium Japonicum, soybean symbiosis by plantN status as determined by shoot C/N ratio. Physiol. Plant. 98,529–538.
Bergersen F J, Turner C, Peoples m B, Gault R R, Morthorpe L Jand Brockwell J 1992 Nitrogen fixation during vegetative andreproductive growth of irrigated soybean in the field. Aust. J.Agric. Res. 43, 145–153.
Bethienfalvay g J and Phillips D A 1977a Effect of light intensityon efficiency of carbon dioxide and nitrogen reduction in Pisumsativum L. Plant Physiol. 60, 868–871.
Bethlenfalvay G J and Phillips D A 1977b Ontogenic interac-tions between photosynthesis and symbiotic nitrogen fixation inlegumes. Plant physiol. 60, 419–421.
Caetano-Anolles G and Gresshoff P M 1991 Plant genetic controlof nodulation. Annu. Rev. Microbiol. 45, 345–382.
Dean JR and Clark K W 1980 Effect of low level nitrogen fertil-ization on nodultation, acetylene reduction and dry matter infaba bean (broadbeans) and three other legumes (peas, beans,soybeans). Can. J. Plant Sci. 60, 121–130.
Duc g, Mariotti A and Amarger N 1988 Measurements of geneticvariability for symbiotic dinitrogen fixation in field-grown fababean (Vicia faba L.) using a low level 15N-tracer technique. Plantand Soil 106, 269–276.
Etévé U and Derieux M 1982 Variabilitié de la durée de la phasevégétative chez le pois (Pisum sativum L.). Application à Iasélection de types resistants à l’hiver et à la détermination de Iadate de semis. Agronomie 2, 813–817.

41
Faurie 0 and Soussana J F 1993 Oxygen-induced recovery fromshort-term nitrate inhibition of N2 fixation in white clover plantsfrom space and dense stands. Physiol. Plant. 89, 467–475.
Finn G A and Brun W A 1982 Effect of atmospheric CO2 enrich-ment on growth, non- structural carbohydrate content and rootnodule activity in soybean. Plant Physiol. 69, 327–331.
Franco A A, Pereira J C and Neyra C A 1979 Seasonal patterns ofnitrate reductase and nitrogenase activities in Phaseolus vulgaris.Plant Physiol. 63, 421–424.
Greenwood D J, Gastal F, Lemaire G, Draycott A, Millard P andNeeteson J J 1991 Growth rate of %N of field grown crops:theory and experiments. Ann. Bot. 67, 77–151.
Hacin J I, Ben Bohlool B and Singleton PW 1997 Partitioning of14C-labelled photosynthate to developing nodules and roots ofsoybean (Glycine max.). New Phytol. 137, 257–265.
Hartwig U A, Heim I, Lfischer A and Nosberger J 1994 Thenitrogen-sink is involved in the regulation of nitrogenase activityin white clover after defoliation. Physiol. Plant. 92, 375–382.
Herdina and Silsbury J H 1990 Growth, nitrogen accumulation andpartitioning, and N2 fixation in faba bean (Vicia faba cv. Fiord)and pea (Pisum sativum cv. Early Dun). Field Crop Res. 24, 173–188.
Imsande J 1989 Rapid dinitrogen fixation during soybean podfill enhances net photosynthetic output and seed yield: a newperspective. Agron. J. 81, 49–556.
Isfan D 1991 Fertilizer nitrogen uptake by soybean as related tocultivars and time of application using 15N dilution technique.J. Plant Nutr. 14 (12), 1369–1380
Jensen E S 1987 Seasonnal patterns of growth and nitrogen fixationin field-grown pea. Plant and Soil 101, 29–37.
Jensen E S 1986 The influence of rate and time of nitrate supply onnitrogen fixation and yield in pea (Pisum sativum L.) Fertil. Res.10, 193–202.
Jensen E S 1997 The role of grain legume N2 fixation in the nitrogencycling of temperate cropping systems. Ed. Riso-R-885 (EN),Holback Center-Tryk, DK. 202 p.
Jensen E S, Andersen A J and Thomsen J D 1985 The influence ofseed-borne N in 15N isotope dilution studies with legumes. ActaAgric. Scand. 35, 438–443.
Jeuffroy m H and Warembourg F R 1991 Carbon transfer and par-titioning between vegetative and reproductive organs in Pisumstivum L. Plant Physiol. 97, 440–448.
Keeney D R and Wilson D W 1989 Nitrogen-inorganic forms. InMethods of Soil Analysis. Part II Chemical and MicrobiologicalProperties, 2nd ed., Eds. A L Page, R H Miller and D R Keeney.American Society of Agronomy, Madsion, WI, USA.
Kouchi H, Akao S and Yoneyama T 1986 Respiratory utilization of13C-labelled photosynthate in nodulated root systems of soybeanplants. J. Exp. Bot. 37 (180), 985–993.
Lemaire G, Gastal F, Plenet D and Le Bot J 1997 Le prélèvementde l’azote par les peuplements végétaux et la production descultures. In Maìtrise de l’azote dans les agrosystèmes, Eds. GLemaire and B Nicolardot. pp 121–139. Série les colloques del’INRA, INRA-Editions, Paris.
Lhuillier-Soundélé A, Munier-Jolain N and Ney B 1999 Depend-ence of seed nitrogen concentration on plant nitrogen availabilityduring the seed filling in pea. Eue J. Agron. 11, 157–166.
MacDuff J H, David S C and Divisdon I A 1996 Inhibition of N2 fix-ation by white clover (Trifolium repens L.) at low concentrationsin NO−
3 in flowing solution cultue. Plant and soil 180, 287–295.Mahon J D and Child J J 1979 Growth response of inoculated peas
(Pisum sativwn) to combined nitrogen. Can. J. Bot. 57, 1687–1693.
Mariotti A, Mariotti F and Amarger N 1983 Utilisation du traçageisotopique nature] par 15N pour la mesure du taux d’azote fixésymbiotiquement par les légumineuses. Physiol. Vég. 21 (2),279–291.
Mariotti A, Mariotti F and Amarger N 1980 Fractionnements iso-topiques de l’azote lors des processus d’absorption des nitrateset de fixation de l’azote atmosphérique par les plantes. Physiol.Vég. 18 (1), 163–181.
Markwei C M and Lame T A 1997 Phenotypic characterizationof sym 21, a gene conditioning shoot-controlled inhibition ofnodulation in Pisum sativum cv. Sparkle. Physiol. Plant. 100,927–932.
Minchin F R, Minguez I, Sheehy J E, Witty J F and Skot L 1986Relationships between nitrate and oxygen sypply in symbioticnitrogen fixation by white clover. J. Exp. Bot. 37, 1103–1113.
Neo H H and Layzell D B 1997 Phleom glutamine and the regulationof O2 diffusion in legume nodules. Plant Physiol. 113, 259–267.
Ney B and Turc O 1993 Heat-Unit Based description of thereproductive developpment of pea. Crop Sci. 33, 510–514.
Nicolardot B, Mary B, Houot S and Recous S 1996. La dynamiquede l’azote dans les sols cultivés. In: Maitrise de l’azote dans lesagrosyst mes, Eds. G Lemaire and B Nicolardot. pp 87–103.Série les colloques de INRA, INRA-Editions, Paris.
Parsons R, Stanforth A, Raven J A and Sprent J I 1993 Nodulegrowth and activity may be regulated by a feed-back mechanisminvolving phloem nitrogen. Plant Cell Environ. 16, 1 125–1136.
Pate J 5 1985 Physiology of pea – a comparison with other legumesin terms of economy of carbon and nitrogen in whole-plant andorgan functioning. In The Pea Crop. Eds. P D Hebblethwaite,M C Heath and T C K Dawkins. pp 279–296. Butterworths,London.
Pate J S and Flinn A m 1973 Carbon and nitrogen transfer fromvegetative organs to ripening seed of field pea (Pisum arvenseL.). J. Exp Bot. 24, 1090–1099.
Rennie R J and Kemp G A 1980 Dinitrogen fixation in pea beans(Phaseolus vulgaris) as affected by growth stage and temperatureregime. Can. J. Bot. 59, 1181–1188.
Sagan M, Ney B and Duc G 1993 Plant symbiotic mutants as atool to analyse nitrogen nutrition and yield relationships in fieldgrown peas (Pisum sativum L.). Plant and Soil 153, 33–45.
SAS Institute, 1987 SAS/STAT Guide for Personal Computer. 6thedition. SAS institute, Cary, NC.
Silsbury J H, Catchpoole D W and Wallace W 1986 Effects of ni-trate and ammonium on nitrogenase (C2H2 reduction) activity ofawards of subterranean clover, Trifolium subterraneum L. Aust.J. Plant Physiol. 13, 257–273.
Sinclair T R and de Witt C T 1975 Photosynthate and nitrogen re-quirements for seed production by various crops. Science 189,565–567.
Sinclair T R and de Wit C T 1976 Analysis of the carbon andnitrogen limitations to soybean yield. Agron. J. 68, 319–324.
Soussana J F and Hartwig U A 1996 The effect of elevated CO2on symbiotic N2 fixation: a link between the carbon and nitrogencycles in grass land ecosystems. Plant and Soil 187 (2), 321–332.
Sparrow S D, Cochran V L and Sparrow E B 1995 Crops: dinitrogenfixation by several legume crops in Alaska. Agron. J. 87, 34–41.
Sprent J I, Stephens J H and Rupela OP 1988 Environmental effectson nitrogen fixation. In Worldcrops: Cool Season Food LegumesEd. R J Sumerfield. pp 801–810. Kluwer Academic publishers,Dordrecht.
Sprent J I and Thomas R J 1984 Nitrogen nutrition of seedling grainlegumes. Some taxonomic, morphological and physiologicalconstrainsts. Plant Cell Environ. 7, 637–645.

42
Streeter J 1988 Inhibition of legume nodule formation and N2fixation by nitrate. CRC Cdit. Rev. Plant Sci. 7, 1–23.
Tricot F 1993 Mise en place des nodosités du pois protéagineuxde printemps (Pisuin sativum L.) Influence de la nutrition car-bonée. Thèse de Doctorat, Université Paris-Sud, centre d’Orsay(France), 78 p.
Vessey J K and Waterer J 1992 In search of the mechanism of nitrateinhibition of nitrogenase activity in legume nodules. Physiol.Plant. 84, 171–176.
Vikman P and Vessey J K 1992 The decline in N2 fixation rate incommon bean with the onset of pod-filling: fact or artifact. Plantand Soil 147, 95–105.
Voisin A-S, Salon C, Munier-Jolain N U and Ney B 2001 Effect ofmineral nitrogen on nitrogen nutrition and biomass partitioning
between the shoot and roots of pea (Pisum satium L.), Plant andSoil 242, 251–262.
Warembourg F R 1983 Estimating the true cost of dinitrogen fix-ation by nodulated plants in undisturbed conditions. Can. J.Microbiol. 29, 930–937.
Waterer J G and Vessey J K 1993 Effect of low static nitrate con-centrations on mineral nitrogen uptake nodulation, and nitrogenfixation in field pea. J. Plant Nutr. 16 (9), 1775–1789.
Yinbo G, Peoples MB and Rerkasem B 1997 The effect of N fertil-izer on N2 fixation, growth and yield of vegetable soybean. FieldCrop Res. 51: 221–229.
Section editor: H. Lambers





























