Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol . 263, No. 33, Issue of November 25, pp. 17317-17321,1388 Printed in U. S. A .
Purification and Characterization of a Protein Associated with Peripheral-type Benzodiazepine Binding Sites*
(Received for publication, May 27, 1988)
Lucyna Antkiewicz-Michaluk, Alexey G. Mukhin, Alessandro Guidotti, and Karl E. KruegerS From the FIDIA-Georgetown Institute for the Neurosciences, Georgetown Uniuersity Medical School, Washington, D. C. 20007
The photoaffinity ligand [3H]PK 14106 was utilized to modify covalently peripheral-type benzodiazepine binding sites in rat adrenal mitochondrial prepara- tions. The photolabeled membrane preparations were then solubilized in 1% digitonin and the detergent- soluble extracts subjected to fractionation by ion-ex- change chromatography and reversed-phase high per- formance liquid chromatography. This scheme re- sulted in the purification of the putative binding site protein for PK 14106 which we have entitled PKBS. Purified preparations of PKBS exhibited a single band with a M, of approximately 17,000 when analyzed by sodium dodecyl sulfate-polyacrylamide gel electropho- resis followed by silver-staining or autoradiographic detection. Additional criteria examining the purity of PKBS preparations were provided by radioiodination with Bolton-Hunter reagent, amino acid analysis, gas- phase sequencing, and reversed-phase chromatogra- phy suggesting that this protein was purified to appar- ent homogeneity. These results demonstrate that a pro- tein associated with peripheral-type benzodiazepine recognition sites has been isolated thus facilitating more direct studies on the structure of this receptor and on the role of these binding sites in mediating responses elicited by benzodiazepines acting at these sites.
Benzodiazepines are one of the most widely used class of drugs by virtue of their anxiolytic and anticonvulsant effects. It is established that a subset of these drugs binds to at least two classes of binding sites termed central and peripheral- type benzodiazepine (PBZD)‘ binding sites, respectively (Braestrup and Squires, 1977; Mohler and Okada, 1977). Central-type benzodiazepine binding sites are almost exclu- sively present in the nervous system and are associated with GABA-regulated chloride channels (Schofield et al., 1987; Sigel et al., 1983, 1984). The therapeutic actions of benzodi- azepines are believed to be mediated through an action on GABA-ergic mechanisms (Costa et al,, 1975; Costa and Gui- dotti, 1979).
In contrast, PBZD binding sites are present in most tissues (De Souza et al., 1985; Anholt et al., 1985), and their levels
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed: FIDIA-George- town Institute for the Neurosciences, Georgetown University Medical School, 3900 Reservoir Rd., Washington, D. C. 20007.
The abbreviations used are: PBZD, peripheral-type benzodiaze- pine; GABA, y-aminobutyric acid; HPLC, high-performance liquid chromatography; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
vary quite substantially between different organs (Shoemaker et al., 1983; Anholt, 1986; Awad and Gavish, 1987). In periph- eral tissues, the primary subcellular localization of PBZD binding sites has been shown to be the mitochondrial com- partment (Anholt et al., 1986), and the tissue-dependent levels of these sites are also reflected in their abundance found in mitochondrial fractions from different tissues (Antkiewicz- Michaluk et al., 1988). The role of this class of receptors is not known, although they have been implicated in a number of cellular phenomena (Curran and Morgan, 1985; Wang et al., 1984; Mestre et al., 1985; Lueddens and Skolnick, 1987).
In addition to benzodiazepines, the PBZD binding domain also exhibits a high affinity for isoquinoline carboxamides (Le Fur et al., 1983a, 198313). The binding of both classes of compounds to PBZD recognition sites is mutually exclusive (Le Fur et al., 1983a, 1983b, 1983c), although they exhibit differential sensitivity to various forms of biochemical pertur- bation (Benavides et al., 1984; Skowronski et al., 1987). It has therefore been proposed that benzodiazepines and isoquino- line carboxamides may bind to different, but allosterically coupled, recognition sites located on PBZD receptors.
Recently, an isoquinoline carboxamide photoaffinity probe designated PK 14105 was synthesized, and its selectivity to insert covalently into PBZD binding sites was characterized (Doble et al., 1987). PK 14105 binds with high affinity to PBZD binding sites, and upon ultraviolet irradiation the association becomes irreversible. Photolabeling of cardiac membranes with [3H]PK 14105 is prevented by other isoqui- noline carboxamides and by benzodiazepines which show high affinities for PBZD binding sites. Photoincorporation is not blocked, however, by benzodiazepines which show high selec- tivity for central-type benzodiazepine binding sites over PBZD binding sites. Photoincorporation of PK 14105 results in an irreversible inhibition of isoquinoline carboxamide and benzodiazepine binding to PBZD binding sites. These obser- vations provide strong evidence that PK 14105 specifically binds to and covalently modifies PBZD binding sites at or near the recognition site for isoquinoline carboxamides.
The mode of covalent attachment of PK 14105 to PBZD binding sites is not known, although it is hypothesized that photoincorporation involves a nucleophilic aromatic substi- tution by an amino acid side chain functional group oriented in close proximity to the bound ligand (Doble et al., 1987). In cardiac membranes [3H]PK 14105 was found to label specifi- cally a single major polypeptide species with an apparent molecular mass of 17-18 kilodaltons. Subsequently, it has been verified that this polypeptide species is photolabeled in mitochondrial fractions from a wide variety of tissues con- taining PBZD binding sites (Antkiewicz-Michaluk et al., 1988). This 17-kDa protein is tentatively assumed to consti- tute a significant portion of the binding site domain for isoquinoline carboxamides, and therefore we have signified
17317

17318 Mitochondrial Benzodiazepine Receptor Protein Isolation
the acronym PKBS (PK binding site) in referring to this protein.
The number of so-called PBZD binding sites is quite sub- stantial in the central nervous system where their density can even exceed that of the central-type benzodiazepine binding sites (Villiger, 1985; Anholt, 1986). Recent electrophysiologi- cal and biochemical studies indicate that in some instances low affinity PBZD-like binding sites may be associated with one of the subtypes of GABAA receptor complexes in neurons (Skerritt et al., 1984; Gee, 1987; Gee et ai., 1988; Barbaccia et aL, 1988). This would imply that recognition sites having a binding profile similar to that of PBZD binding sites may be associated with GABA receptor complexes located on neu- ronal membranes.
Very recent studies employing recombinant DNA probes suggest that more than two subunit species may comprise GABAA receptors and also that multiple molecular forms of these receptors are possible (Barnard et al., 1988). In view of this fact, the possibility that the structure of PBZD binding sites may be found in different GABAA receptor subtypes needs to be considered. The purification of proteins which constitute PBZD binding sites would not only permit more direct studies on this class of recognition sites, but might also be of great value in elucidating related structures within various GABAA receptor subtypes.
The studies presented in this report demonstrate the use of the [3H]PK 14105 photoaffinity probe in monitoring the fractionation of the 17-kDa PKBS protein associated with mitochondrial PBZD binding sites. A simple purification scheme is presented resulting in the isolation of the PKBS protein to apparent homogeneity from rat adrenal mitochon- drial preparations.
EXPERIMENTAL PROCEDURES
Isolation of Adrenal Mitochondrial Fractions-Adrenal glands from Sprague-Dawley rats were decapsulated and placed in 20 volumes of 10 mM Tris-HC1 (pH 7.4), 0.32 M sucrose, containing a protease inhibitor mixture (0.1 mM phenylmethylsulfonyl fluoride, 0.1 p~ pepstatin A, 1 pg/ml aprotinin, 1 pg/ml leupeptin, 1 mM EDTA). The tissue was homogenized using 10 strokes in a Potter-Elvehjem tissue grinder, and the homogenate was centrifuged at 770 X g for 10 min. The supernatant was recovered and centrifuged at 5200 X g producing a pellet which was resuspended in the homogenization buffer and centrifuged again at 9000 X g. These mitochondrial-enriched pellets were stored at -70 "C until ready for use.
Photolabeling of Mitochondrial PBZD Binding Sites-Rat adrenal mitochondrial fractions were suspended at a concentration of 800 pg of protein/ml in 50 mM Tris-HC1 (pH 7.6) containing protease inhibitor mixture. Plastic culture dishes of 100-mm diameter con- tained 3 ml of suspension which was constantly stirred at 0 "C. To each dish [3H]PK 14105 was added to a concentration of 2 nM, and the samples were irradiated for 1 h from a distance of 3 cm using an ultraviolet light with maximum emission at 366 nm (Mineralight, UVGL-58). Following irradiation the membrane suspensions were diluted into 6 ml of 50 mM Tris-protease inhibitor mixture buffer containing 1 p~ PK 11195, centrifuged at 20,000 X g for 10 min, and the pellets washed once more in 6 ml of buffer containing 1 p M PK 11195.
Solubilization of Adrenal Mitochondrial Fractions and Purification of PKBS-Photolabeled mitochondrial preparations were combined with a 30-fold excess of unlabeled adrenal mitochondrial fractions, solubilized for 30 min at 0 "C with 10 mM Tris-HC1 (pH 7.4), 0.32 M sucrose, protease inhibitor mixture, 1% digitonin (w/v) at a concen- tration of 1 mg of protein/ml. The samples were then centrifuged for 30 min at 150,000 X g, and the supernatant fractions were recovered.
The digitonin-solubilized extracts were subjected to Q-Sepharose chromatography, and fractions containing PKBS were pooled as described under Fig. 1. These pooled fractions were concentrated with a Centriprep-30 membrane, diluted with 10 volumes of 0.1% trifluo- roacetic acid (w/v), and concentrated again to a volume of 1 ml. This additional washing procedure was found to improve the fractionation
of the sample by reversed-phase HPLC, which is described in the legend to Fig. 2.
Quantitative Protein Determination-Protein content was meas- ured by the procedure of Lowry et al. (1951), utilizing bovine serum albumin as a standard. For detergent-solubilized samples a modifi- cation of this method was employed as described (Bensadoun and Weinstein, 1976).
SDS-PAGE and Electroblotting of Protein Samples-Electropho- resis was performed by the method of Laemmli (1970) using 1.5-mm- thick 10-15% gradient polyacrylamide gels and a stacking gel of 3% polyacrylamide. Transfer of protein from gels to nitrocellulose mem- branes was accomplished with a Polyblot apparatus (American Bio- nuclear, Emeryville, CA) using the procedure suggested by the man- ufacturers. A discontinuous buffer system comprised of two anode buffers, 300 and 25 mM Tris (pH 10.4), each containing 20% metha- nol, and a cathode buffer of 25 mM Tris, 40 mM 6-aminohexanoic acid (pH 9.4), 20% methanol were utilized. Electroblotting was carried out for 1 h at 20 V.
Radioiodination of PKBS-Approximately 0.1 gg of purified PKBS recovered from reversed-phase HPLC was dried under vacuum, and 50 pl of 0.1 M NaB03 (pH 8.5) containing 20 pCi of [1Z51]diiodo- Bolton-Hunter reagent (4400 Ci/mmol) was introduced and allowed to react a t room temperature for 15 min. The reaction was terminated by adding 50 p1 of borate buffer containing 0.1 M glycine, and the sample was centrifuged at 12,000 x g for 3 min. The supernatant, which did not contain PKBS or other detectable labeled polypeptide species, was removed. The insoluble pellet fraction was washed once with 1 ml of borate buffer, solubilized in loading buffer for SDS- PAGE, and 1 X IO5 cpm was applied to the gel, electrophoresed, and transferred to nitrocellulose as described under Fig. 3.
RESULTS
Rat adrenal mitochondrial preparations possess a very high density of PBZD binding sites, approximately 140 pmol/mg protein, and contain a protein apparently common to all tissues expressing mitochondrial PBZD binding sites, of ap- proximately 17 kDa, which is covalently modified by PK 14105 during ultraviolet irradiation (Antkiewicz-Michaluk et al., 1988). Thus the 17-kDa protein should comprise at least 0.2% of the total protein from these membrane preparations. Because of this high abundance, rat adrenal mitochondrial fractions were used as an amenable source to purify the putative binding site protein for isoquinoline carboxamides. Adrenal mitochondrial membrane preparations photolabeled with [3H]PK 14105 were mixed with a 30-fold excess of unlabeled mitochondria and solubilized in a buffer containing 1% digitonin. The recoveries of membrane protein and radio- activity in the detergent-solubilized supernatant were at least 90% (Table I).
The solubilized extract was then applied to a Q-Sepharose column, and elution of the protein photolabeled by [3H]PK 14105 was performed with a linear gradient from 0 to 0.5 M NaCl. The majority of the labeled PKBS protein was eluted between 50-150 mM NaCl as evidenced by the radioactive profile of this separation method (Fig. 1). In addition to this peak two other radioactive peaks were observed. The first one
TABLE I Recoveries of protein and radioactivity from adrenal mitochondrial
fractions photolabeled with fH]PK 14105 Results from a single representative experiment are shown.
Fraction Total protein cpm (X 10-3)
mg Mitochondrial membrane fraction 90 1059 Digitonin-solubilized extract 82 903 Q-Sepharose 5.9 229 Reversed-Dhase HPLC 0.12" 40.0
"In addition to the procedure of Lowry et al. (1951), protein was also estimated by amino acid analysis assuming a molecular mass of 17 kDa (Table 11) and yielded the same value.
Mitochondrial Benzodiazepine Receptor Protein Isolation 17319
0.8 I I
A H 10
ML Eluant
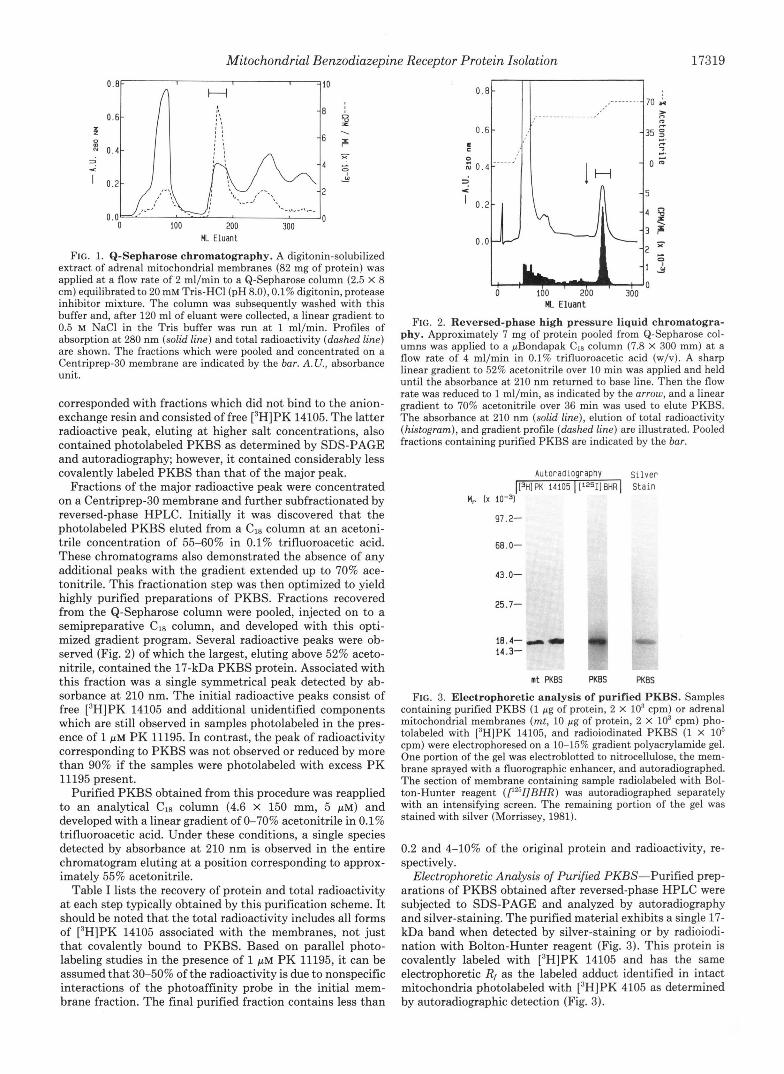
FIG. 1. Q-Sepharose chromatography. A digitonin-solubilized extract of adrenal mitochondrial membranes (82 mg of protein) was applied at a flow rate of 2 ml/min to a Q-Sepharose column (2.5 X 8 cm) equilibrated to 20 mM Tris-HC1 (pH 8.0), 0.1% digitonin, protease inhibitor mixture. The column was subsequently washed with this buffer and, after 120 ml of eluant were collected, a linear gradient to 0.5 M NaCl in the Tris buffer was run a t 1 ml/min. Profiles of absorption at 280 nm (solid line) and total radioactivity (dashed line) are shown. The fractions which were pooled and concentrated on a Centriprep-30 membrane are indicated by the bur. A. U., absorbance unit.
corresponded with fractions which did not bind to the anion- exchange resin and consisted of free [3H]PK 14105. The latter radioactive peak, eluting at higher salt concentrations, also contained photolabeled PKBS as determined by SDS-PAGE and autoradiography; however, it contained considerably less covalently labeled PKBS than that of the major peak.
Fractions of the major radioactive peak were concentrated on a Centriprep-30 membrane and further subfractionated by reversed-phase HPLC. Initially it was discovered that the photolabeled PKBS eluted from a C18 column at an acetoni- trile concentration of 55-60% in 0.1% trifluoroacetic acid. These chromatograms also demonstrated the absence of any additional peaks with the gradient extended up to 70% ace- tonitrile. This fractionation step was then optimized to yield highly purified preparations of PKBS. Fractions recovered from the Q-Sepharose column were pooled, injected on to a semipreparative Cla column, and developed with this opti- mized gradient program. Several radioactive peaks were ob- served (Fig. 2) of which the largest, eluting above 52% aceto- nitrile, contained the 17-kDa PKBS protein. Associated with this fraction was a single symmetrical peak detected by ab- sorbance at 210 nm. The initial radioactive peaks consist of free [3H]PK 14105 and additional unidentified components which are still observed in samples photolabeled in the pres- ence of 1 PM PK 11195. In contrast, the peak of radioactivity corresponding to PKBS was not observed or reduced by more than 90% if the samples were photolabeled with excess PK 11195 present.
Purified PKBS obtained from this procedure was reapplied to an analytical C18 column (4.6 X 150 mm, 5 pM) and developed with a linear gradient of 0-70% acetonitrile in 0.1% trifluoroacetic acid. Under these conditions, a single species detected by absorbance at 210 nm is observed in the entire chromatogram eluting at a position corresponding to approx- imately 55% acetonitrile.
Table I lists the recovery of protein and total radioactivity at each step typically obtained by this purification scheme. It should be noted that the total radioactivity includes all forms of [3H]PK 14105 associated with the membranes, not just that covalently bound to PKBS. Based on parallel photo- labeling studies in the presence of 1 p~ PK 11195, it can be assumed that 30-50% of the radioactivity is due to nonspecific interactions of the photoaffinity probe in the initial mem- brane fraction. The final purified fraction contains less than
E z 0.
?' ' 0.
0
<
0.
HL Eluant
FIG. 2. Reversed-phase high pressure liquid chromatogra- phy. Approximately 7 mg of protein pooled from Q-Sepharose col- umns was applied to a pBondapak CIS column (7.8 X 300 mm) at a flow rate of 4 ml/min in 0.1% trifluoroacetic acid (w/v). A sharp linear gradient to 52% acetonitrile over 10 min was applied and held until the absorbance at 210 nm returned to base line. Then the flow rate was reduced to 1 ml/min, as indicated by the arrow, and a linear gradient to 70% acetonitrile over 36 min was used to elute PKBS. The absorbance at 210 nm (solid line), elution of total radioactivity (histogram), and gradient profile (dashed line) are illustrated. Pooled fractions containing purified PKBS are indicated by the bur.
A u t o r a d l o g r a p n y S i l v e r [[3H] PK 14105 I [1251] BHR I S t a i n
H, [x lo-?
97.2-
68.0-
43.0-
25.7-
18.4- - - rw i4.3-
mt PKBS PKBS PKBS
FIG. 3. Electrophoretic analysis of purified PKBS. Samples containing purified PKBS (1 pg of protein, 2 X IO3 cpm) or adrenal
tolabeled with ['HIPK 14105, and radioiodinated PKBS (1 X lo6 mitochondrial membranes (mt, 10 pg of protein, 2 x lo3 cpm) pho-
cpm) were electrophoresed on a 10-15% gradient polyacrylamide gel. One portion of the gel was electroblotted to nitrocellulose, the mem- brane sprayed with a fluorographic enhancer, and autoradiographed. The section of membrane containing sample radiolabeled with Bol- ton-Hunter reagent (rz51]BHR) was autoradiographed separately with an intensifying screen. The remaining portion of the gel was stained with silver (Morrissey, 1981).
0.2 and 4-10% of the original protein and radioactivity, re- spectively.
Electrophoretic Analysis of Purified PKBS-Purified prep- arations of PKBS obtained after reversed-phase HPLC were subjected to SDS-PAGE and analyzed by autoradiography and silver-staining. The purified material exhibits a single 17- kDa band when detected by silver-staining or by radioiodi- nation with Bolton-Hunter reagent (Fig. 3). This protein is covalently labeled with [3H]PK 14105 and has the same electrophoretic R, as the labeled adduct identified in intact mitochondria photolabeled with 13H]PK 4105 as determined by autoradiographic detection (Fig. 3).

17320 Mitochondrial Benzodiazepine Receptor Protein Isolation
TABLE I1 Partial amino acid analysis of purified PKBS
hydrolysis in uacuo with 6 N HCI at 108" C for 24 h. Analysis of Approximately 20-30 pg of protein was subjected to gas-phase
amino acid residues was performed on a Beckman 121MB amino acid analyzer by the method of Spackman et al. (1958). Results are presented as the mean f S.D. of four separate analyses. All possible permutations of the observed amino acid composition uersus variable polypeptide size were checked using a computer program specifically designed for this purpose (L. Nadasdi and J. Kealey, personal com- munication). Using this algorithm, a polypeptide of 17,280 Da mani- fested the least error producing the theoretical molar contribution of each amino acid per mol of protein as indicated.
Amino acid Residue mol residue/ percent mol protein
Ala 10.3 f 0.55 15 Arg 6.1 & 0.53 10 Asx 5.8 f 0.47 9 Glx 8.8 f 0.33 13 GlY 10.6 k 0.65 16 His 1.8 f 0.17 3 Ile 3.4 k 0.56 6 Leu 12.8 f 0.41 20 LY s 4.7 * 0.96 8 Met 2.9 t 0.41 5 Phe 4.4 f 0.44 8 Pro 6.5 f 0.27 10 Ser 6.5 f 0.39 10 Thr 5.2 f 0.39 9 TY r 3.5 f 0.60 6 Val 6.7 f 0.60 11
Total 100 159
Partial Amino Acid Composition-Purified PKBS was sub- jected to amino acid analysis although no attempt was made to determine cysteine or tryptophan content. The relative abundance of amino acids in PKBS and its proposed compo- sition are listed in Table 11. Since corrections for incomplete hydrolysis or residue decomposition were not performed, some of the values obtained may be a slight underestimate of the true content. Four different analyses produced amino acid profiles which were most compatible with a protein of at least 15,560-17,560 Da. The mean of these four experiments yielded an amino acid composition which predicts a minimal molec- ular mass of 17,280 Da bearing the theoretical number of residues listed in Table 11. This theoretical calculation is in good agreement with the apparent molecular weight deter- mined by SDS-PAGE and provides significant criteria regard- ing the purity of the PKBS protein.
Added support in demonstrating the purity of PKBS prep- arations is given by the fact that attempts at obtaining partial amino acid sequence information from 200 pmol of purified PKBS failed to give a signal by gas-phase sequencing. Further studies by us in successfully obtaining partial amino acid sequences of peptide fragments from PKBS demonstrate that the amount of protein employed in this study greatly exceeded the limit of detection for sequence analysis. Therefore the ineffectiveness in obtaining a sequence from PKBS prepara- tions was not due to insufficient quantity of sample. This indicates that PKBS is blocked at its amino terminus, but moreover this also demonstrates the absence of contaminant polypeptides with accessible amino termini.
DISCUSSION
The results presented in this report demonstrate the puri- fication to apparent homogeneity of a protein associated with PBZD binding sites. This class of recognition sites had pre- viously been characterized as having binding specificity for at least two groups of organic compounds (Le Fur et al., 1983a,
1983b), benzodiazepines and isoquinoline carboxamides. PK 14105 was utilized in these studies to label covalently the putative binding sites for isoquinoline carboxamides and served as a powerful tool in monitoring the fractionation of the PKBS protein. SDS-PAGE of the purified protein exhib- ited a molecular mass of approximately 17 kDa in agreement with the [3H]PK 14105-photolabeled adduct identified from mitochondrial membrane fractions.
The yield of protein in the PKBS preparations accounts for approximately 0.1% of the total adrenal mitochondrial protein. Assuming that the density of PBZD binding sites in adrenal mitochondrial fractions is 100-150 pmol/mg protein (Antkiewicz-Michaluk et al., 1988), one might expect a yield in the range of 0.02-0.13% of the total protein in the final purified fraction if a 10-50% recovery of PKBS is achieved. The yield of protein obtained by this procedure is therefore not unreasonable, particularly when certain difficulty can be encountered when attempting to quantify accurately a rela- tively small amount of protein. The -fold purification of PKBS protein cannot be accurately determined in these stud- ies since technical errors arise when attempting to correlate the recovery of radioactivity with recovery of PKBS. The reasons for this are %fold. First, the proportion of [3H]PK 14105 in photolabeled membranes which is covalently bound to PKBS cannot be adequately determined. Secondly, it ap- pears as if the PK 14105-PKBS adducts may be susceptible to hydrolysis at low pH, thereby interfering in the determi- nation of PKBS yield during reversed-phase HPLC. These possibilities can account for the apparent low yield of total radioactivity listed in Table I, a value which is certainly a considerable underestimate of PKBS recovery.
Although the -fold purification of PKBS cannot be deter- mined, the degree of purity of these preparations was evalu- ated by five criteria. 1) Silver-staining of SDS-polyacrylamide gels on which PKBS was electrophoresed show only one heavily stained band. 2) SDS-PAGE analysis of PKBS prep- arations radioiodinated with Bolton-Hunter reagent revealed PKBS as the only labeled species migrating above the dye front. 3) Partial amino acid analysis of PKBS preparations predicts a minimal molecular mass of approximately 16-18 kDa for a protein possessing the determined amino acid composition, in good agreement with the value determined by SDS-PAGE. If other polypeptides were contaminating these preparations the estimations by both procedures would likely show a greater discrepancy. 4) PKBS subjected to gas-phase sequencing did not result in the release of phenylthiohydan- toin derivatives. Thus significant amounts of contaminating polypeptide species containing free amino termini were not present. 5) Only one absorbance peak at 210 nm is observed when PKBS is reapplied to a CI8 column and developed with a linear gradient of 0-70% acetonitrile. Based on these criteria PKBS can be considered to be essentially homogeneous.
At this point it is still not known whether the 17-kDa polypeptide is a fragment cleaved from a larger protein. The studies reported here employed a mixture of protease inhibi- tors suggesting that proteolytic activity does not produce this species from a larger protein, although this possibility can not be ruled out. Additional studies have shown that protease inhibitors are not essential in that the same 17-kDa protein is obtained from preparations lacking these inhibitors. These studies also establish that ultraviolet irradiation during pho- toactivation of PK 14105 does not generate the 17-kDa poly- peptide, since photolabeled mitochondrial fractions were com- bined with a 30-fold excess of unlabeled mitochondrial mem- branes. The purified PKBS from these preparations contain the 17-kDa species as observed by silver-staining indicating

Mitochondrial Benzodiazepine Receptor Protein Isolation 17321
that this polypeptide is present in nonirradiated membranes as well.
Radiation inactivation studies estimated a molecular mass of 34 kDa for the functional unit associated with PBZD binding sites (Paul et al., 1981). This might imply that the functional binding site may represent a dimer of the PKBS protein. It has been previously reported that the benzodiaze- pine flunitrazepam photolabels a 34-kDa protein believed to be the mitochondrial anion-channel porin (Snyder et al., 1987). PKBS is known to be a protein distinct from porin since additional studies by us indicate that amino acid se- quences obtained from peptide fragments of PKBS show no homology with yeast mitochondrial porin.' Biochemical stud- ies have suggested that benzodiazepine and isoquinoline car- boxamides may bind, in a mutually exclusive manner, to different sites which are allosterically coupled on PBZD rec- ognition site complexes (Benavides et al., 1984; Skowronski et al., 1987). This might account for the differences observed with the two photoaffinity probes; however, the relationships between PKBS, porin, and the PBZD binding site complex remain to be clarified.
The amino acid sequence information that we have eluci- dated by gas-phase sequencing of different peptide fragments derived from PKBS accounts for nearly one-half of the PKBS polypeptide length? The sequences obtained have been ana- lyzed with the protein sequence database of the National Biomedical Research Foundation (Washington, D. C.) , and no correspondence or significant homology with other known protein sequences has been found. There is also no apparent sequence homology to the polypeptide diazepam-binding in- hibitor (Mocchetti et al., 1986; Gray et al., 1986), the putative endogenous modulator of benzodiazepine receptors. PKBS is therefore probably a novel protein requiring more intensive characterization. The purification of PKBS will enable the molecular cloning and production of antibodies to this protein. This should undoubtedly prove to be an important step in understanding the molecular construction and cellular roles of PBZD receptors.
Acknowledgments-We wish to express our sincere gratitude to Drs. H. Van Hove (Commissariat a 1'Energie Atomique) and C. Gueremy (Pharmuka Laboratories) for their generous gifts of [3H] PK 14105 and PK 11195 used in these studies and to Dr. James Kealey (Biomolecular Resource Center, University of California, San Francisco) for his expertise in amino acid analysis and peptide sequencing.
REFERENCES Anholt, R. R. H. (1986) Trends Phurmncol. Sci. 7, 506-511 Anholt, R. R. H., De Souza, E. B., Oster-Granite, M. L., and Snyder,
Anholt, R. R. H., Pedersen, P. L., De Souza, E. B., and Snyder, S. H.
Antkiewicz-Michaluk, L., Guidotti, A., and Krueger, K. E. (1988)
Awad, M., and Gavish, M. (1987) J. Neurochem. 49,1407-1414 Barbaccia, M. L., Costa, E., and Guidotti, A. (1988) Annu. Rev.
Barnard, E. A., Burt, D. R., Darlison, M. G., Fujita, N., Levitan, E.
S. H. (1985) J. Phurmacol. Exp. Ther. 233,517-526
(1986) J. Biol. Chem. 261,576-583
Mol. Phurmacol. 34,272-278
Phurmmol. Toxicol. 28, 471-476
A. G. Mukhin, J. Kealey, and K. E. Krueger, unpublished data.
J., Schofield, E. J., Seeburg, P. H., Squire, M. D., and Stephenson, F. A. (1988) in The Allosteric Modulation of Amino Acid Receptors and Its Therapeutic Implications (Costa, E., and Barnard, E. A., eds) pp. 19-32, Raven Press, New York
Benavides, J., Begassat, F., Phan, T., Tur, C., Uzan, A., Renault, C., Dubroeucq, M. C., Gueremy, C., and Le Fur, G. (1984) Life Sci. 35,
Bensadoun, A., and Weinstein, D. (1976) Anal. Biochem. 70, 241-
Braestrup, C., and Squires, R. F. (1977) Proc. Natl. Acad. Sci. U. S.
Costa, E., and Guidotti, A. (1979) Annu. Rev. Phurmacol. Toxicol. 19,
Costa, E., Guidotti, A., Mao, C. C., and Suria, A. (1975) Life Sci. 17,
Curran, T., and Morgan, J. I. (1985) Science 229, 1265-1268 De Souza, E. B., Anholt, R. R. H., Murphy, K. M. M., Synder, S. H.,
and Kuhar, M. J . (1985) Endocrinology 116,567-573 Doble, A., Ferris, O., Burgevin, M. C., Menager, J., Uzan, A., Dub-
roeucq, M. C., Renault, C., Gueremy, C., and Le Fur, G. (1987) Mol. Phurmacol. 3 1,42-49
1249-1256
250
A. 74,3805-3809
531-545
167-186
Gee, K. W. (1987) J. Phurmncol. Exp. Ther. 240, 747-753 Gee. K. W., Brinton. R. E., and McEwen, B. S. (1988) J. Phurmacol.
Exp. Ther. 244,379-383 Grav. P. W.. Glaister. D.. Seebure. P. H.. Guidotti. A.. and Costa, E.
(l"986) Proc. Natl. Acad. Sci. UrS. A. 83, 7547-7551 Laemmli, U. K. (1970) Nature 227,680-685 Le Fur, G., Guilloux, F., Rufat, P., Uzan, A,, Renault, C., Dubroeucq,
M. C., and Gueremy, C. (1983a) Life Sci. 32, 1849-1856 Le Fur, G., Perrier, M. L., Vaucher, N., Imbault, F., Flamier, A.,
Uzan, A., Renault, C., Dubroeucq, M. C., and Gueremy, C. (1983b) Life Sci. 32, 1839-1847
Le Fur, G., Vaucher, N., Perrier, M. L., Flamier, A., Benavides, J., Renault. C.. Dubroeucq, M. C., Gueremy, C., and Uzan, A. (1983~) Life Sci.'33,449-457 "
Lowrv. 0. H.. Rosebroueh. N. J.. Farr. A. L.. and Randall. R. J. (1951) J. B h . Chkm. 193,265-275
. ,
Lueddens, H. W. M., and Skolnick, P. (1987) Eur. J. Phurmacol.
Mestre, M., Carriot, T., Belin, C., Uzan, A., Renault, C., Dubroeucq, M. C., Gueremy, C., Doble, A., and Le Fur, G. (1985) Life Sci. 36,
Mocchetti, I., Einstein, R., and Brosius, J. (1986) Proc. Natl. Acad.
Mohler, H., and Okada, T. (1977) Science 198, 849-851 Morrissey, J. H. (1981) Anal. Biochem. 117, 307-310 Paul, S. M., Kempner, E. S., and Skolnick, P. (1981) Eur. J. Phur-
macol. 76,465-466 Schofield, P. R., Darlison, M. G., Fujita, N., Burt, D. R., Stephenson,
F. A., Rodriguez, H., Rhee, L. M., Ramachandran, J., Reale, V., Glencorse, T., Seeburg, P. H., and Barnard, E. A. (1987) Nature
Shoemaker, H., Boles, R. G., Horst, W. D., and Yamamura, H. I.
Sigel, E., and Barnard, E. A. (1984) J. Biol. Chem. 259, 7219-7223 Sigel, E., Stephenson, F. A., Mamalaki, C., and Barnard, E. (1983) J.
Skerritt, J. H., Werz, M. A., McLean, M. J., and MacDonald, R. L.
Skowronski, R., Beaumont, K., and Fanestil, D. D. (1987) Eur. J.
Snyder, S. H., Verma, A., and Trifiletti, R. R. (1987) FASEB J. 1 ,
Spackman, D. H., Stein, W. H., and Moore, S. (1958) Anal. Chem.
Villiger, J. W. (1985) Neuro~hnrmncology 24,95-98
133,205-214
391-400
Sci. U. S. A. 83, 7221-7225
328,221-227
(1983) J. Pharmacol. Exp. Ther. 225,61-69
Biol. Chem. 268,6965-6971
(1984) Bruin Res. 310,99-105
Phurmacol. 143,305-314
282-288
30, 1190-1206
Wang, J. K. T., Morgan, J. I , and Spector, S. (1984) Proc. Natl. Acad. Sci. U. S. A. 81,3770-3772










![--Chapter 5 - Protein Purification and Characterization Techniques (1) [Compatibility Mode]](https://img.dokumen.tips/doc/110x75/552e34b34a7959b7628b4879/-chapter-5-protein-purification-and-characterization-techniques-1-compatibility-mode.jpg)


















