Embed Size (px)
Citation preview

Tumor Biology and Immunology
PTBP3-Mediated Regulation of ZEB1 mRNAStability Promotes Epithelial–MesenchymalTransition in Breast CancerPingfu Hou1, Lin Li1, Fang Chen1, Yansu Chen3, Hui Liu4, Jingjing Li5, Jin Bai1, andJunnian Zheng1,2,6
Abstract
The RNA polypyrimidine tract-binding protein PTBP3 is a littlestudied paralog of PTBP1, which has oncogenic properties. In thisstudy,wedemonstrate thatPTBP3 induces epithelial–mesenchymaltransition (EMT) in breast tumor cells and promotes their invasivegrowth and metastasis. Elevated expression of PTBP3 associatedsignificantly with lymph node metastasis, advanced histologygrade, TNM stage, and poor 5-year overall survival of patients. Inhuman mammary epithelial cells, PTBP3 overexpression was suf-ficient to induce EMT and to enhance cell migration, invasion, andcancer stem-like cell properties. PTBP3 regulated expression of the
EMT regulatory transcription factor ZEB1 by binding the 30UTR ofits mRNA, thereby preventing its degradation. Conversely, ZEB1ablation blocked the ability of PTBP3 to induce EMT. Overall, ourfindings define PTBP3 as a regulator of EMT that acts by governingexpression of ZEB1, and they establish an oncogenic function ofPTBP3, suggesting its candidacy as a theranostic target.
Significance: Thesefindings define PTBP3 as a regulator of EMTthat acts by governing expression of ZEB1, and they establishan oncogenic function of PTBP3, suggesting its candidacy as atheranostic target. Cancer Res; 78(2); 387–98. �2017 AACR.
IntroductionBreast cancer is the most common cancer among women
worldwide. Distant metastases are the cause of about 90% ofdeaths in breast cancer (1). The epithelial-to-mesenchymal tran-sition (EMT) process, first described in embryogenesis, is charac-terized by changes in cell morphology, behavior, and plasticity(2, 3). The activation of EMT program depends on a diverse arrayof proteins, including transcription factors, cell signaling regula-tors, and secretory factors, as well as regulation by long noncodingRNAs (lncRNA). EMT is widely accepted as an essential step forfueling tumor initiation and metastatic spread (4).
EMT is associated with decreased expression of epithelialmarkers, such as E-cadherin, and increased expression of mesen-chymal markers (5). E-cadherin is repressed at the transcriptionlevel bymultiple EMT inducers, including Snail, Slug, Twist, ZEB1,
and ZEB2 (6). ZEB1 can suppress E-cadherin expression bybinding to the E-box region of CDH1 gene promoter (7, 8),thereby acting as a master inhibitor of EMT (9). Recent studieshave shown that Slug can directly activate ZEB1 at the transcrip-tional level (10). However, ZEB1 expression is posttranscription-ally repressed by several miRNAs, including mir-200 family, mir-205 (11), and mir-216a (12). It is therefore generally agreed thatZEB1 is an important activator of EMT and may serve as amoderator of breast cancer stem cells (BCSC; ref. 13).
Recently, increasing evidence suggests that diverse RNA-binding proteins (RBP) are involved in cancer process. PTBP1is a member of RBPs that interacts with regulatory RNAs toregulate mRNA splicing (14), stability (15), localization (16),and translation (17). PTBP1 has two paralogs, PTBP2 (nPTB;neural PTB) and PTBP3 (ROD1; regulator of differentiation 1),with a similar protein architecture and containing four RNArecognition motifs (18). The functions of PTBP1 and PTBP2have been well studied, whereas the role of PTBP3 has beenrelatively neglected in a long time. Recent studies have shownthat PTBP3 plays a role in nonsense-mediated mRNA decay andfunctions as a splicing repressor (19). Considering the multiplefunctions of PTBP1 and PTBP2 in various biological processesand the similar structure of PTBP3 with PTBP1/2, we expectedthat PTBP3 would have an important nonredundant role incellular processes.
In this study, we have extensively analyzed the function ofPTBP3 using cell-based approaches, mouse xenograft models,and IHC-based human breast cancer correlation studies. Theresults have uncovered previously unrecognized roles forPTBP3 in EMT and breast cancer progression. We demonstratedthat high expression of PTBP3 in cancers was confirmed to be apredictor of poorer survival of breast cancer patients. PTBP3acted as an inducer of EMT program and promoted cell migra-tion, invasion, proliferation, and breast cancer stemness
1Cancer Institute, Xuzhou Medical University, Xuzhou, Jiangsu, China. 2Center ofClinical Oncology, Affiliated Hospital of Xuzhou Medical University, Xuzhou,Jiangsu, China. 3School of Public Health, Xuzhou Medical University, Xuzhou,Jiangsu, China. 4School of Pathology, Xuzhou Medical University, Xuzhou,Jiangsu, China. 5School of Medical Imaging, Xuzhou Medical University, Xuzhou,Jiangsu, China. 6Jiangsu Center for the Collaboration and Innovation of CancerBiotherapy, Cancer Institute, XuzhouMedical University, Xuzhou, Jiangsu, China.
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
P. Hou and L. Li contributed equally to this article.
Correspondence Authors: Junnian Zheng, Jiangsu Center for the Collaborationand Innovation of Cancer Biotherapy, Cancer Institute, Xuzhou Medical Univer-sity, TheAffiliated Hospital of XuzhouMedical University, 84West Huaihai Road,Xuzhou 221002, Jiangsu Province, China. Phone: 86-516-85582530; Fax: 86-516-85582530; E-mail: [email protected]; and Jin Bai, [email protected]
doi: 10.1158/0008-5472.CAN-17-0883
�2017 American Association for Cancer Research.
CancerResearch
www.aacrjournals.org 387
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

acquisition. In addition, knockdown of PTBP3 was testified torepress tumor growth and metastasis in vivo. Finally, the impor-tant role of ZEB1 for PTBP3-induced EMT was revealed. Inmechanism, PTBP3 regulated ZEB1 expression by preventingZEB1 mRNA degradation through binding to the 30UTR regionof ZEB1 mRNA.
Materials and MethodsCell line culture
MCF10A, MCF7, MDA-MB-231, BT474, and HEK293T celllines were obtained from ATCC and immediately expanded andfrozen down as master stocks. A vial of working stock wasalways used to start a new experiment, and the cells werereplenished with another working stock after being culturedfor no more than 6 weeks. In addition, these cell lines wereauthenticated by short tandem repeat profiling by ATCC orGenewiz, CN. MCF10A cells were cultured as described previ-ously (20). MCF7 and BT474 cells were cultured in RPMI1640medium supplemented with 10% FBS. MDA-MB-231 cells werecultured in L15 medium with 10% FBS at 37�C without CO2.HEK293T cells were cultured in DMEM medium containing10% FBS.
Gene transfection and silencingFor siRNA transfections, cells were transfected with 20 nmol/L
siRNA (Shanghai GenePharma) by siLentFect (Bio-Rad Labora-tories, Inc.). For plasmid transfections, cells were transfectedusing Lipofectamine 2000 (Life Technologies), according tothe manufacturer's instructions. The siRNAs and shRNAsequences are described as follows: siPTBP3#1:CCCAGUAAAUG-CACAUUAUTT, siPTBP3#2: GCCCUGUGCUUCGAAUAAUTT,siZEB1#1:GGAUCAGAAUGACUCUGAUTT, siZEB1#2: GGU-CUUAUUCUCAACACAUTT, siPTBP1: GCCUCAACGUCAA-GUACAATT, shCtrl-For: CCGGCCTAAGGTTAAGTCGCCCTCTC-GAGAGCGAGGGCGACTTAACCTTAGGTTTTTG, shPTBP3#1-for: CCGGACCAGGAAATTCTGTTCTACTCTCGAGAGTAGAA-CAGAATTTCCTGGTTTTTTG, shPTBP3#2-for: CCGGCAGA-GACTTCACTCGCTTAGACTCGAGTCTAAGCGAGTGAAGTCTC-TGTTTTTG.
Plasmids and virus infectionThe vector pCMV6-XL5-PTBP3 was a kind gift from Dr. Luis F.
Congote, McGill University Health Centre (Montreal, Canada;ref. 21). PTBP3 cDNA was cloned by PCR from pCMV6-XL5-PTBP3 using TransStart FastPfu Fly DNA Polymerase Kit (Trans-Gen Biotech) and inserted into pCDH1-CMV-MSC-EF1-GFP-Puro lentivirus vector (System Biosciences) at ECOR1/BamH1sites. PTBP3 shRNA sequences were cloned to the vector pLko.1 atAge1/ECOR1 sites. PTBP3 overexpression and knockdown lenti-viruses were generated by cotransfecting 293T cells with the othertwo packing vectors pMD2G and psPAX and concentrated asdescribed previously (20). Stable cell lines overexpressing orlacking PTBP3 were generated by being infected with lentivirusand selected with 2 mg/mL puromycin for about 2 weeks. Thevectors pCDNA3-Flag-MS2bp and pCDNA3-12 � MS2bs wereprovided by Dr. Xiaofei Zheng (Academy of Military MedicalSciences, Beijing, China). ZEB1 30UTR was cloned by PCR andinserted into pCDNA3-12 � MS2bs between ECOR1 and Not1sites. Primer sequences for vector cloning are available inSupplementary Experimental Procedures.
Immunofluorescence and Western blottingImmunofluorescence and Western blotting were done as
described previously (20). Primary antibodies against the follow-ing proteins were used: E-cadherin (BD Biosciences, 610181),N-cadherin (BD Biosciences, 610920), vimentin (BD Bio-sciences, 550513), fibronectin (BD Biosciences, 610077), ZEB1(Santa Cruz Biotechnology, sc-25388), ZEB2 (Santa Cruz Bio-technology, sc-271984), AGO2 (Santa Cruz Biotechnology,sc-53521), PTBP3 (Santa Cruz Biotechnology, sc-100845),PTBP1 (Santa Cruz Biotechnology, sc-16547), GAPDH (SantaCruz Biotechnology, sc-271984), PKM1 (SAB, #21577), PKM2(SAB, #21578), C-myc (Genescript, A00704), and Flag (Sigma,F1804). Secondary antibodies included horseradish peroxidase(HRP)-goat anti-mouse, HRP-goat anti-rabbit, and HRP-rabbitanti-goat (ZSGB-BIO). Protein bands were detected by Tanon5200 automatic chemiluminescence imaging analysis systemusing ECL reagent (Tanon).
Cellular proliferation, colony formation, invasion, andmigration assays
CCK-8 assay was applied to measure the cell proliferationaccording to the Cell Counting Kit-8 manufacturer's protocol(Dojindo). Colony formation assay was performed in 60-mmdishes in which 1,000 cells were seeded and cultured for 2 weeks.Colonies were counted manually after staining with 0.1% crystalviolet. Migration assays and Matrigel invasion assays in breastcancer cell lineswere conducted as describedpreviously (20),withminor modification. Cells were treated with serum starvationovernight, and then migration and invasion assays were per-formed as described previously.
RNA extraction, RT-PCR, and real-time PCR analysisTRIzol reagent was used to extract RNA from cells and tissues,
and the cDNA was generated with random primers (Promega)using theReverse Transcription System (Promega). Real-time PCRwas carried out on ABI-7500 using SYBR Green Real-Time PCRMaster Mix (Vazyme Biotech). GAPDH was used for normaliza-tion of qRT-PCR data. Primer sequences are available in Supple-mentary Experimental Procedures.
Immunoprecipitation and RNA pull-down assayImmunoprecipitation was carried out as described previously
(22). Briefly, MCF7 lysates were prepared in immunoprecipita-tion lysis buffer (20 mmol/L Tris-Cl, pH 8.0, 10 mmol/L NaCl, 1mmol/L EDTA, 0.5% NP-40) containing a protease inhibitorcocktail (Sigma). Two micrograms of cell extracts was preclearedwith 50 mL protein A/G-agarose (Santa Cruz Biotechnology) at4�C for 2 hours, and the supernatant was incubated with corre-sponding antibodies with gently shacking at 4�C overnight,followed by the addition of 50 mL of protein A/G-agarose foranother 2 hours. The beads were washed and then resuspended in60mL of 1� loading buffer andboiled for 5minutes, and followedwithWestern blot detection. RNApull-down assaywas carried outusing Flag-MS2bp-MS2bs–based pull-down assay as describedpreviously (20), with minor modification. Specifically, pcDNA3-FlagMS2bp and pcDNA3-ZEB1-30UTR-MS2bs (or mock vector)were cotransfected with pCMV6-XL5-PTBP3 to 293T cells, and thecells were harvested after 48 hours. About 2� 107 cells were lysedin lysis buffer (20 mmol/L Tris-Cl, pH 8.0, 10 mmol/L NaCl,1 mmol/L EDTA, 0.5% NP-40) supplemented with RNasin(80 U/mL, NEB) and a protease inhibitor cocktail (Sigma).
Hou et al.
Cancer Res; 78(2) January 15, 2018 Cancer Research388
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

A total of 2.5 mg monoclonal anti-Flag(R) M2 antibody (sigma)was added to each binding reaction tube and incubated at 4�C for4 hours, followed by addition of 50 mL of protein A/G-agarose foranother 2hours. Beadswerewashedfive timeswith the lysis bufferand boiled in 1� loading buffer for 5 minutes. The retrievedproteins were subjected to SDS-PAGE and detected by Westernblot analysis.
Patient specimens and IHC for tissue microarrayA series of 418 human breast tumor specimens were obtained
from the First Affiliated Hospital of Nanjing Medical University(Nanjing, China). The clinicopathologic information of patientswas obtained from the archive of the pathology department andconfirmed by the medical record of the hospital, and informedconsent was obtained from all patients. The patient studies wereconducted in accordance with Declaration of Helsinki. Five-yearclinical follow-up results were available for 123 patients. The useof these specimens and data for research purposes were approvedby the Ethics Committee of the Hospital. For IHC, as describedpreviously (23), tissueswere stained overnight at 4�Cwith a 1:100dilution of PTBP3 antibody (Santa Cruz Biotechnology, sc-100845) or with a 1:200 dilution of ZEB1 antibody (Santa CruzBiotechnology, sc-25388).
Luciferase assaysThe ZEB1-30UTR fragment was cloned by PCR and inserted into
the luciferase reporter vector psiCHECK2 (Promega) betweenXho1 and Not1 sites. Experiments were performed as describedpreviously (20). Primer sequences for vector cloning are availablein Supplementary Experimental Procedures.
Mammosphere formation assaysExperiments were performed as described previously (20).
Animal workBALB/c nude mice (4–5 weeks old) were purchased from
Beijing HFK Bio-technology. All animal experiments wereapproved by the Animal Care Committee of the Xuzhou MedicalUniversity, Xuzhou, China. Animal experiments were performedas described previously (20).
Statistical analysesStatistical analyses were carried out with SPSS 20.0 software.
The association between PTBP3 staining and the clinicopatho-logic parameters of the breast cancer patients were evaluated by ac2 test. The Kaplan–Meier method and log-rank test were used toevaluate the correlation between PTBP3 expression and patientsurvival. Pearson and Spearman correlation coefficients were usedto calculate the correlation between PTBP3 and ZEB1 in tissuemicroarray (TMA). The unpaired t test was used to determine thestatistical significance of differences between groups. Data werepresented as mean � SD. P < 0.05 was considered statisticallysignificant.
ResultsHigh PTBP3 expression correlates with clinicopathologicparameters and poor 5-year overall survival in breast cancerpatients
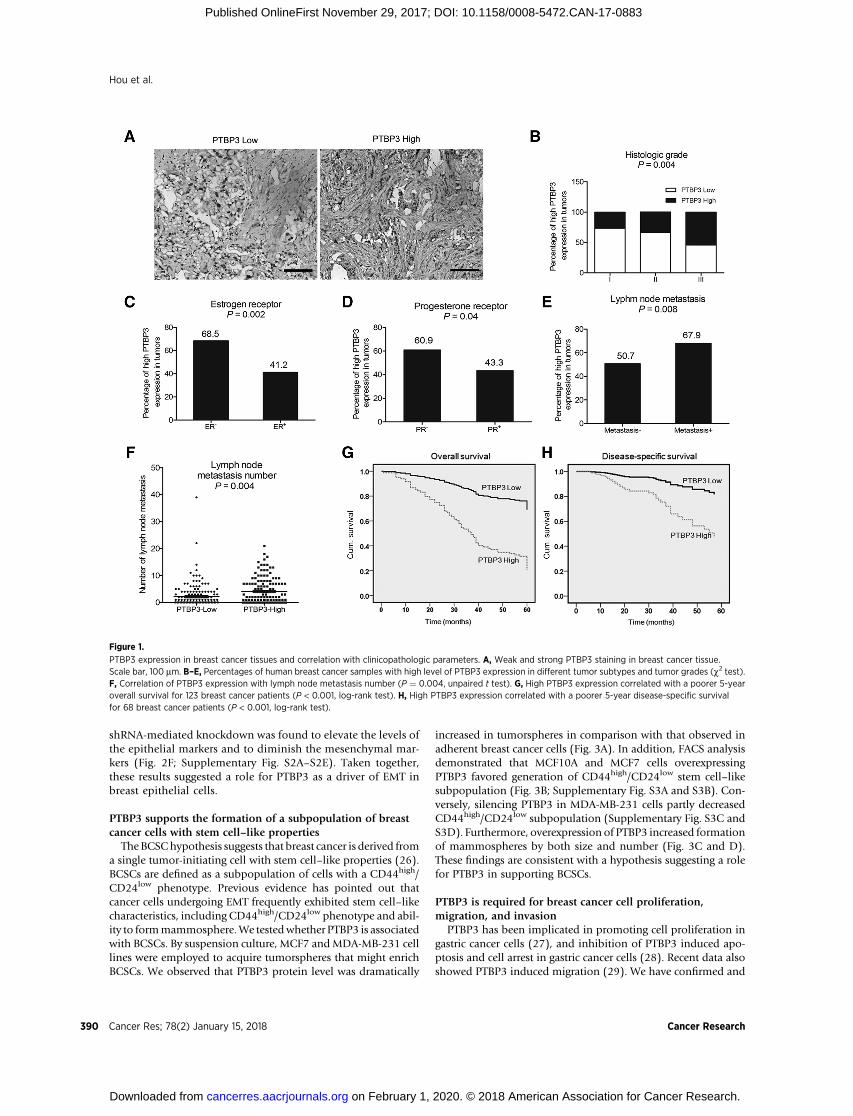
To examine the expression of PTBP3 in breast cancer tissuesamples, IHC was carried out in TMA slides (Fig. 1A). The
correlation between PTBP3 expression and clinicopathologiccharacteristics in 418 breast cancer samples was analyzed (Sup-plementary Table S1). Compared with histology grade 1 and 2,the PTBP3 signal was dramatically increased in histology grade3 (P ¼ 0.004, c2 test, Fig. 1B). In addition, high PTBP3expression is positively correlated with ER negative (P ¼0.002, c2 test, Fig. 1C) and PR negative (P ¼ 0.040, c2
test, Fig. 1D). Interestingly, high PTBP3 expression was signif-icantly correlated with lymph node metastasis (P ¼ 0.008, c2
test, Fig. 1E) and lymph node metastasis numbers (P ¼ 0.004,c2 test, Fig. 1F). The results of Kaplan–Meier survival analysisrevealed that high PTBP3 levels are correlated with poor 5-yearoverall survival (P < 0.001, c2 test, Fig. 1G) and disease-specificpatient survival (P < 0.001, c2 test, Fig. 1H).
Next, we addressed whether PTBP3 expression was an inde-pendent prognostic factor for breast cancer. The univariate Coxregression analyses revealed that PTBP3 expression was an inde-pendent prognostic marker for breast cancer patients overallsurvival (HR, 4.226; 95% confidence interval (CI), 2.406–7.422; P < 0.001) and disease-specific survival (HR, 4.989;95% CI, 2.269–10.970; P < 0.001; Supplementary Table S2). Inmultivariate Cox regression analysis (Supplementary Table S3),we found that PTBP3 expression was also an independent prog-nosticmarker for both 5-year overall survival (HR, 3.768; 95%CI,2.014–7.051; P < 0.001) and disease-specific survival (HR, 3.700;95% CI, 1.495–9.158; P ¼ 0.005).
To further evaluate the clinical link between PTBP3 andbreast carcinomas, PTBP3 mRNA expression level was analyzedwith the Oncomine database (24). Data from 29 (total 29)breast cancer cohorts showed higher mRNA level of PTBP3 inmammary carcinomas than in normal breast tissues (Supple-mentary Table S4), strongly suggesting that PTBP3 expressionwas positively associated with breast cancer. Further analysisindicated that the PTBP3 expression was closely correlated witha very poor clinical outcome (Supplementary Table S5). Theanalysis of PTBP3 mRNA expression levels in a TCGA datasetcontaining 817 breast tumors (25) indicated that high PTBP3expression was positively correlated with a poor overall survivaland disease-free survival (Supplementary Fig. S1A and S1B). Insummary, results from both database and our clinical speci-mens have suggested PTBP3 expression as a predictor of thepoor clinical outcome in breast cancer.
Overexpression of PTBP3 promotes EMT transitionin breast cancer cells
To assess the role of PTBP3 in EMT, a MCF10A-based cell linewas created to stably express PTBP3 at levels comparable withthose observed in breast cancer cell lines and breast tumortissues (Fig. 2A). Forced expression of PTBP3 caused MCF10Ato undergo EMT, as revealed by the loss of cell–cell adhesionand apical–basal polarity, as well as by the gain of mesenchy-mal cell phenotype (Fig. 2B). Consistent with these findings,ectopic overexpression of PTBP3 induced the downregulationof epithelial markers (E-cadherin and b-catenin) and the upre-gulation of mesenchymal markers (vimentin, fibronectin, andN-cadherin) at the protein levels (Fig. 2C and D). Real-timePCR experiments revealed higher levels of mRNA expression,suggesting a predominantly transcription effect (Fig. 2E). Sim-ilar to MCF10A cells, overexpressed PTBP3 in MCF7 cells alsopromotes EMT program; in contrast, decreased expression ofPTBP3 in MCF7, MDA-MB-231, and BT474 cells by siRNA or
PTBP3 Promotes EMT through Regulating ZEB1 mRNA Stability
www.aacrjournals.org Cancer Res; 78(2) January 15, 2018 389
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883
shRNA-mediated knockdown was found to elevate the levels ofthe epithelial markers and to diminish the mesenchymal mar-kers (Fig. 2F; Supplementary Fig. S2A–S2E). Taken together,these results suggested a role for PTBP3 as a driver of EMT inbreast epithelial cells.
PTBP3 supports the formation of a subpopulation of breastcancer cells with stem cell–like properties
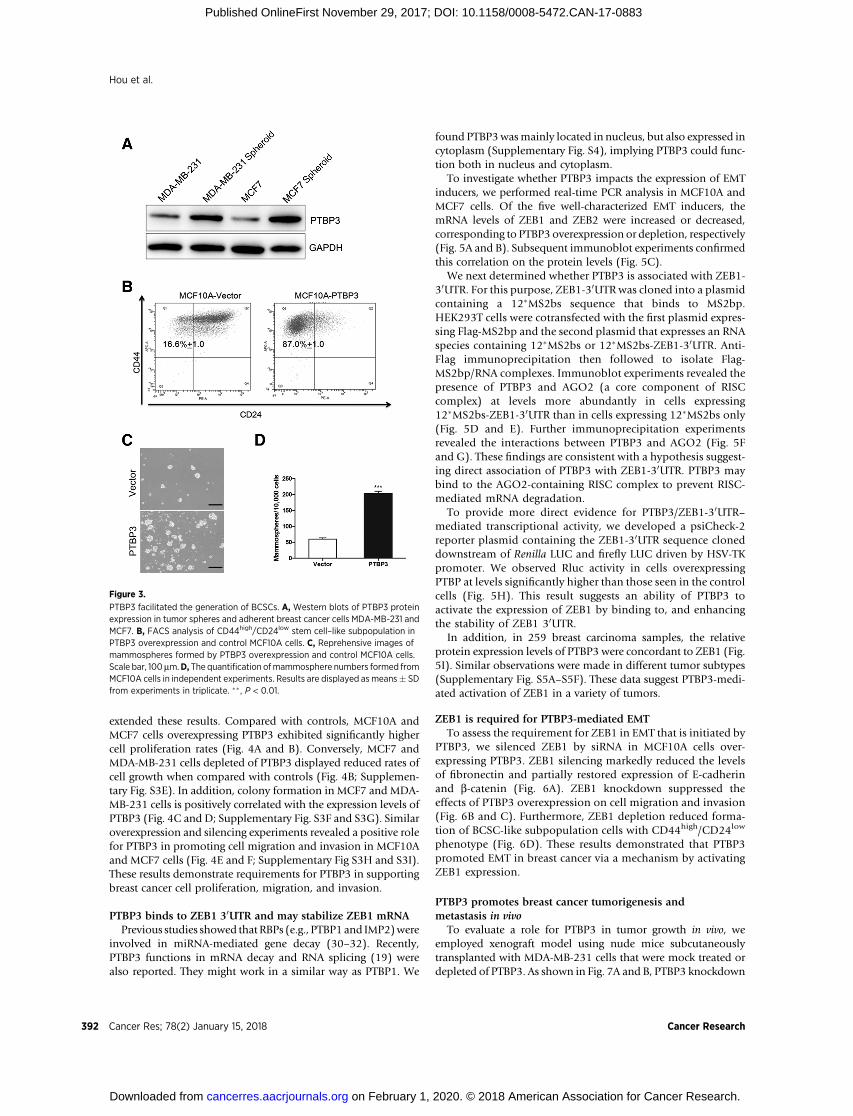
TheBCSChypothesis suggests that breast cancer is derived froma single tumor-initiating cell with stem cell–like properties (26).BCSCs are defined as a subpopulation of cells with a CD44high/CD24low phenotype. Previous evidence has pointed out thatcancer cells undergoing EMT frequently exhibited stem cell–likecharacteristics, including CD44high/CD24low phenotype and abil-ity to formmammosphere.We testedwhether PTBP3 is associatedwith BCSCs. By suspension culture, MCF7 andMDA-MB-231 celllines were employed to acquire tumorspheres that might enrichBCSCs. We observed that PTBP3 protein level was dramatically
increased in tumorspheres in comparison with that observed inadherent breast cancer cells (Fig. 3A). In addition, FACS analysisdemonstrated that MCF10A and MCF7 cells overexpressingPTBP3 favored generation of CD44high/CD24low stem cell–likesubpopulation (Fig. 3B; Supplementary Fig. S3A and S3B). Con-versely, silencing PTBP3 in MDA-MB-231 cells partly decreasedCD44high/CD24low subpopulation (Supplementary Fig. S3C andS3D). Furthermore, overexpression of PTBP3 increased formationof mammospheres by both size and number (Fig. 3C and D).These findings are consistent with a hypothesis suggesting a rolefor PTBP3 in supporting BCSCs.
PTBP3 is required for breast cancer cell proliferation,migration, and invasion
PTBP3 has been implicated in promoting cell proliferation ingastric cancer cells (27), and inhibition of PTBP3 induced apo-ptosis and cell arrest in gastric cancer cells (28). Recent data alsoshowed PTBP3 induced migration (29). We have confirmed and
Figure 1.
PTBP3 expression in breast cancer tissues and correlation with clinicopathologic parameters. A, Weak and strong PTBP3 staining in breast cancer tissue.Scale bar, 100 mm. B–E, Percentages of human breast cancer samples with high level of PTBP3 expression in different tumor subtypes and tumor grades (c2 test).F, Correlation of PTBP3 expression with lymph node metastasis number (P ¼ 0.004, unpaired t test). G, High PTBP3 expression correlated with a poorer 5-yearoverall survival for 123 breast cancer patients (P < 0.001, log-rank test). H, High PTBP3 expression correlated with a poorer 5-year disease-specific survivalfor 68 breast cancer patients (P < 0.001, log-rank test).
Hou et al.
Cancer Res; 78(2) January 15, 2018 Cancer Research390
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883
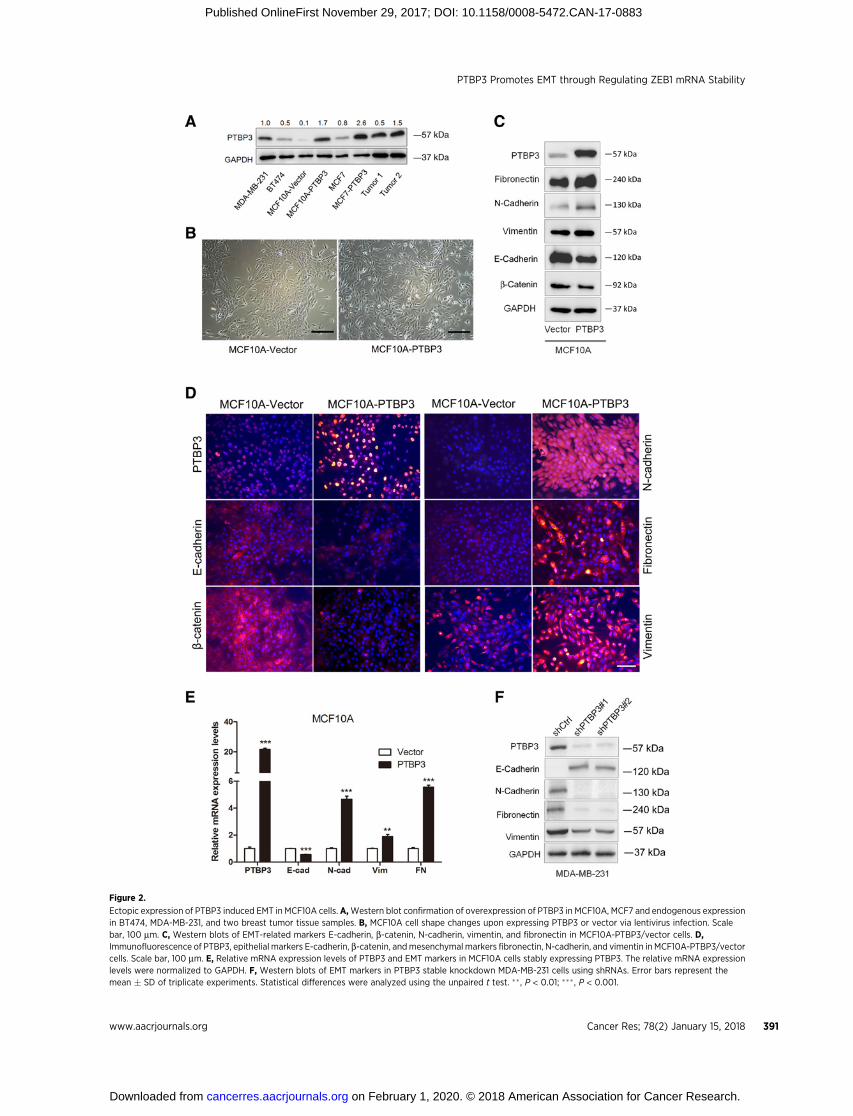
Figure 2.
Ectopic expression of PTBP3 induced EMT in MCF10A cells. A,Western blot confirmation of overexpression of PTBP3 in MCF10A, MCF7 and endogenous expressionin BT474, MDA-MB-231, and two breast tumor tissue samples. B, MCF10A cell shape changes upon expressing PTBP3 or vector via lentivirus infection. Scalebar, 100 mm. C, Western blots of EMT-related markers E-cadherin, b-catenin, N-cadherin, vimentin, and fibronectin in MCF10A-PTBP3/vector cells. D,Immunofluorescence of PTBP3, epithelial markers E-cadherin, b-catenin, andmesenchymal markers fibronectin, N-cadherin, and vimentin in MCF10A-PTBP3/vectorcells. Scale bar, 100 mm. E, Relative mRNA expression levels of PTBP3 and EMT markers in MCF10A cells stably expressing PTBP3. The relative mRNA expressionlevels were normalized to GAPDH. F, Western blots of EMT markers in PTBP3 stable knockdown MDA-MB-231 cells using shRNAs. Error bars represent themean � SD of triplicate experiments. Statistical differences were analyzed using the unpaired t test. �� , P < 0.01; ��� , P < 0.001.
PTBP3 Promotes EMT through Regulating ZEB1 mRNA Stability
www.aacrjournals.org Cancer Res; 78(2) January 15, 2018 391
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883
extended these results. Compared with controls, MCF10A andMCF7 cells overexpressing PTBP3 exhibited significantly highercell proliferation rates (Fig. 4A and B). Conversely, MCF7 andMDA-MB-231 cells depleted of PTBP3 displayed reduced rates ofcell growth when compared with controls (Fig. 4B; Supplemen-tary Fig. S3E). In addition, colony formation in MCF7 and MDA-MB-231 cells is positively correlated with the expression levels ofPTBP3 (Fig. 4C and D; Supplementary Fig. S3F and S3G). Similaroverexpression and silencing experiments revealed a positive rolefor PTBP3 in promoting cell migration and invasion in MCF10Aand MCF7 cells (Fig. 4E and F; Supplementary Fig S3H and S3I).These results demonstrate requirements for PTBP3 in supportingbreast cancer cell proliferation, migration, and invasion.
PTBP3 binds to ZEB1 30UTR and may stabilize ZEB1 mRNAPrevious studies showed that RBPs (e.g., PTBP1 and IMP2)were
involved in miRNA-mediated gene decay (30–32). Recently,PTBP3 functions in mRNA decay and RNA splicing (19) werealso reported. They might work in a similar way as PTBP1. We
found PTBP3wasmainly located in nucleus, but also expressed incytoplasm (Supplementary Fig. S4), implying PTBP3 could func-tion both in nucleus and cytoplasm.
To investigate whether PTBP3 impacts the expression of EMTinducers, we performed real-time PCR analysis in MCF10A andMCF7 cells. Of the five well-characterized EMT inducers, themRNA levels of ZEB1 and ZEB2 were increased or decreased,corresponding to PTBP3 overexpression or depletion, respectively(Fig. 5A and B). Subsequent immunoblot experiments confirmedthis correlation on the protein levels (Fig. 5C).
We next determined whether PTBP3 is associated with ZEB1-30UTR. For this purpose, ZEB1-30UTR was cloned into a plasmidcontaining a 12�MS2bs sequence that binds to MS2bp.HEK293T cells were cotransfected with the first plasmid expres-sing Flag-MS2bp and the second plasmid that expresses an RNAspecies containing 12�MS2bs or 12�MS2bs-ZEB1-30UTR. Anti-Flag immunoprecipitation then followed to isolate Flag-MS2bp/RNA complexes. Immunoblot experiments revealed thepresence of PTBP3 and AGO2 (a core component of RISCcomplex) at levels more abundantly in cells expressing12�MS2bs-ZEB1-30UTR than in cells expressing 12�MS2bs only(Fig. 5D and E). Further immunoprecipitation experimentsrevealed the interactions between PTBP3 and AGO2 (Fig. 5Fand G). These findings are consistent with a hypothesis suggest-ing direct association of PTBP3 with ZEB1-30UTR. PTBP3 maybind to the AGO2-containing RISC complex to prevent RISC-mediated mRNA degradation.
To provide more direct evidence for PTBP3/ZEB1-30UTR–mediated transcriptional activity, we developed a psiCheck-2reporter plasmid containing the ZEB1-30UTR sequence cloneddownstream of Renilla LUC and firefly LUC driven by HSV-TKpromoter. We observed Rluc activity in cells overexpressingPTBP at levels significantly higher than those seen in the controlcells (Fig. 5H). This result suggests an ability of PTBP3 toactivate the expression of ZEB1 by binding to, and enhancingthe stability of ZEB1 30UTR.
In addition, in 259 breast carcinoma samples, the relativeprotein expression levels of PTBP3 were concordant to ZEB1 (Fig.5I). Similar observations were made in different tumor subtypes(Supplementary Fig. S5A–S5F). These data suggest PTBP3-medi-ated activation of ZEB1 in a variety of tumors.
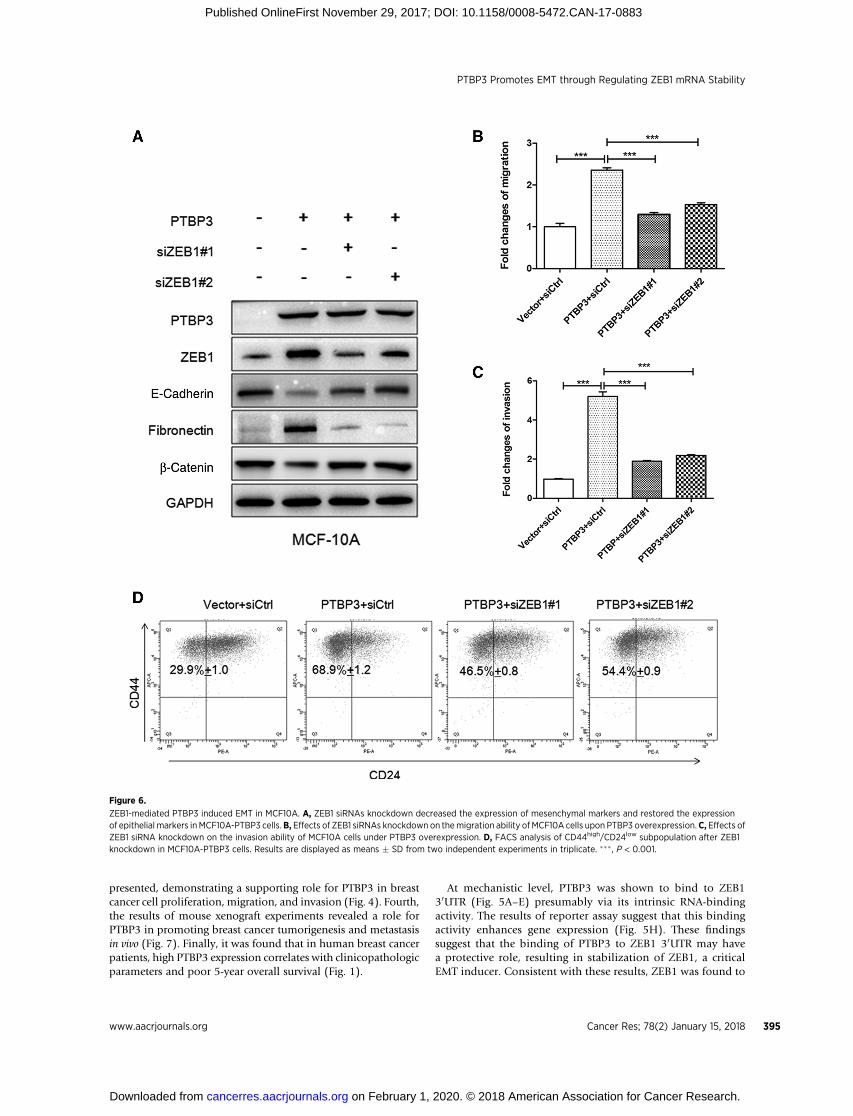
ZEB1 is required for PTBP3-mediated EMTTo assess the requirement for ZEB1 in EMT that is initiated by
PTBP3, we silenced ZEB1 by siRNA in MCF10A cells over-expressing PTBP3. ZEB1 silencing markedly reduced the levelsof fibronectin and partially restored expression of E-cadherinand b-catenin (Fig. 6A). ZEB1 knockdown suppressed theeffects of PTBP3 overexpression on cell migration and invasion(Fig. 6B and C). Furthermore, ZEB1 depletion reduced forma-tion of BCSC-like subpopulation cells with CD44high/CD24low
phenotype (Fig. 6D). These results demonstrated that PTBP3promoted EMT in breast cancer via a mechanism by activatingZEB1 expression.
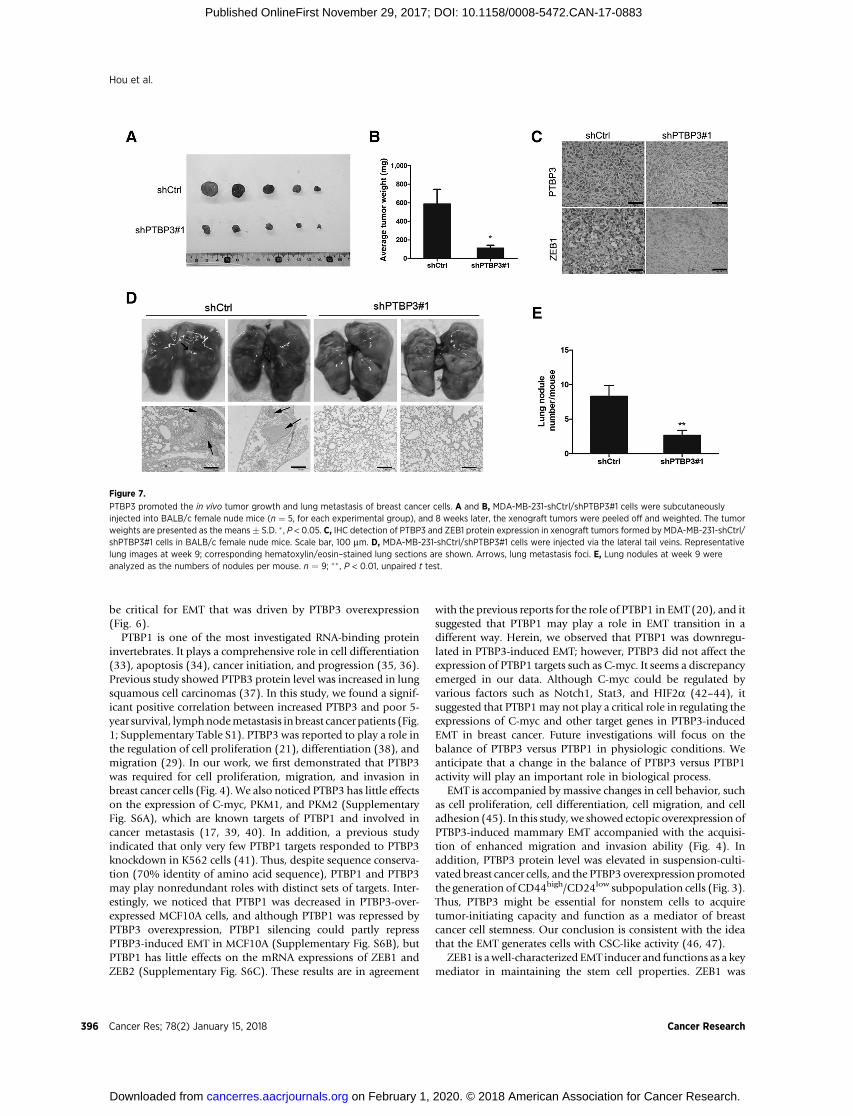
PTBP3 promotes breast cancer tumorigenesis andmetastasis in vivo
To evaluate a role for PTBP3 in tumor growth in vivo, weemployed xenograft model using nude mice subcutaneouslytransplanted with MDA-MB-231 cells that were mock treated ordepleted of PTBP3. As shown in Fig. 7A and B, PTBP3 knockdown
Figure 3.
PTBP3 facilitated the generation of BCSCs. A, Western blots of PTBP3 proteinexpression in tumor spheres and adherent breast cancer cells MDA-MB-231 andMCF7. B, FACS analysis of CD44high/CD24low stem cell–like subpopulation inPTBP3 overexpression and control MCF10A cells. C, Reprehensive images ofmammospheres formed by PTBP3 overexpression and control MCF10A cells.Scale bar, 100 mm.D, The quantification ofmammosphere numbers formed fromMCF10A cells in independent experiments. Results are displayed as means� SDfrom experiments in triplicate. �� , P < 0.01.
Hou et al.
Cancer Res; 78(2) January 15, 2018 Cancer Research392
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

cells formed smaller tumors than the control. Also, the expressionlevels of PTBP3 and ZEB1 were low in the PTBP3 knockdownsubcutaneous tumors (Fig. 7C). Next, we investigated the effectsof PTBP3 in breast cancer cell lung metastasis by the tail veininjectionmodel. Briefly,MDA-MB-231-shPTBP3/shCtrl cellswereinjected via lateral tail vein to 6-week-old nudemice, and themicewere executed to analyze the lung metastasis status 9 weeks later.As shown in Fig. 7D, although multiple large metastatic foci inlung were found in the control group, fewer were observed in thesamples containing MDA-MB-231-shPTBP3 cells (Fig. 7E). Thesefindings provided compelling evidence for the ability of PTBP3 toconfer tumorigenesis and metastasis in vivo.
DiscussionIn this work, we provide a comprehensive set of data suggesting
critical roles for PTBP3 in EMT and in breast cancer progression.First, PTBP3was shown to support the proper function of the EMTtranscription program, acting to downregulate epithelial markers(such as E-cadherin and b-catenin), but upregulating mesenchy-mal markers (such as vimentin, fibronectin, and N-cadherin;Fig. 2). Second, it was revealed that PTBP3mediates the formationof a subpopulation of breast cancer cells named mammospheresthat display a CD44high/CD24low phenotype (Fig. 3), which hasbeen defined as a stem cell–like property. Third, evidence was
Figure 4.
PTBP3 promoted breast cancer cell proliferation, migration, and invasion. A, CCK8 assay analysis of cell proliferation in PTBP3 overexpression and controlMCF10A cells. B, CCK8 assay in PTBP3 overexpression and knockdown MCF7 cells. C and D, Colony formation assay performed in MCF7 cells upon PTBP3overexpressionor knockdown.A relative average ratio (in percentage) of colonynumberswas calculated.E,The ability ofmigrationbyoverexpressionor knockdownof PTBP3 in MCF10A and MCF7 cells. F, Invasion ratio changes upon PTBP3 overexpression or silencing in MCF10A and MCF7. All experiments were carriedout in triplicate. Results are displayed as means � SD from experiments in triplicate. � , P < 0.05; �� , P < 0.01; ���, P < 0.001.
PTBP3 Promotes EMT through Regulating ZEB1 mRNA Stability
www.aacrjournals.org Cancer Res; 78(2) January 15, 2018 393
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

Figure 5.
PTBP3 regulated ZEB1 expression through preventing ZEB1 mRNA degradation. A and B, Relative mRNA expression levels of EMT inducers (Snail, Slug, Twist,ZEB1, and ZEB2) were assessed by real-time PCR in PTBP3-overexpressing or silencing MCF10A and MCF7 cells. The relative mRNA expression levels werenormalized to GAPDH. C,Western blots of ZEB1 and ZEB2 in PTBP3 overexpression or knockdown MCF10A, MCF7, and MDA-MB-231 cells. D,MS2-based RNA pull-down assay showing that PTBP3 and AGO2 were enriched by ZEB1-30UTR RNA. E, Schematic diagram of the MS2-based RNA pull-down strategy. F and G,Immunoprecipitation assays showing PTBP3 and AGO2 binding to each other in MCF7 cells. H, Luciferase activity of ZEB1-30UTR was measured after miR-200c orPTBP3 transfection in MCF7 cells. I, Analysis of the correlation coefficients between PTBP3 and ZEB1 in 259 breast tissue samples. The relative PTBP3 IHCscores were concordant to that of ZEB1. The correlation coefficients were calculated by Pearson and Spearman test. Results are displayed as means � SD fromexperiments in triplicate. ��� , P < 0.001
Hou et al.
Cancer Res; 78(2) January 15, 2018 Cancer Research394
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883
presented, demonstrating a supporting role for PTBP3 in breastcancer cell proliferation, migration, and invasion (Fig. 4). Fourth,the results of mouse xenograft experiments revealed a role forPTBP3 in promoting breast cancer tumorigenesis and metastasisin vivo (Fig. 7). Finally, it was found that in human breast cancerpatients, high PTBP3 expression correlates with clinicopathologicparameters and poor 5-year overall survival (Fig. 1).
At mechanistic level, PTBP3 was shown to bind to ZEB130UTR (Fig. 5A–E) presumably via its intrinsic RNA-bindingactivity. The results of reporter assay suggest that this bindingactivity enhances gene expression (Fig. 5H). These findingssuggest that the binding of PTBP3 to ZEB1 30UTR may havea protective role, resulting in stabilization of ZEB1, a criticalEMT inducer. Consistent with these results, ZEB1 was found to
Figure 6.
ZEB1-mediated PTBP3 induced EMT in MCF10A. A, ZEB1 siRNAs knockdown decreased the expression of mesenchymal markers and restored the expressionof epithelial markers inMCF10A-PTBP3 cells.B, Effects of ZEB1 siRNAs knockdown on themigration ability of MCF10A cells upon PTBP3 overexpression.C, Effects ofZEB1 siRNA knockdown on the invasion ability of MCF10A cells under PTBP3 overexpression. D, FACS analysis of CD44high/CD24low subpopulation after ZEB1knockdown in MCF10A-PTBP3 cells. Results are displayed as means � SD from two independent experiments in triplicate. ��� , P < 0.001.
PTBP3 Promotes EMT through Regulating ZEB1 mRNA Stability
www.aacrjournals.org Cancer Res; 78(2) January 15, 2018 395
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883
be critical for EMT that was driven by PTBP3 overexpression(Fig. 6).
PTBP1 is one of the most investigated RNA-binding proteininvertebrates. It plays a comprehensive role in cell differentiation(33), apoptosis (34), cancer initiation, and progression (35, 36).Previous study showed PTPB3 protein level was increased in lungsquamous cell carcinomas (37). In this study, we found a signif-icant positive correlation between increased PTBP3 and poor 5-year survival, lymphnodemetastasis inbreast cancer patients (Fig.1; Supplementary Table S1). PTBP3 was reported to play a role inthe regulation of cell proliferation (21), differentiation (38), andmigration (29). In our work, we first demonstrated that PTBP3was required for cell proliferation, migration, and invasion inbreast cancer cells (Fig. 4). We also noticed PTBP3 has little effectson the expression of C-myc, PKM1, and PKM2 (SupplementaryFig. S6A), which are known targets of PTBP1 and involved incancer metastasis (17, 39, 40). In addition, a previous studyindicated that only very few PTBP1 targets responded to PTBP3knockdown in K562 cells (41). Thus, despite sequence conserva-tion (70% identity of amino acid sequence), PTBP1 and PTBP3may play nonredundant roles with distinct sets of targets. Inter-estingly, we noticed that PTBP1 was decreased in PTBP3-over-expressed MCF10A cells, and although PTBP1 was repressed byPTBP3 overexpression, PTBP1 silencing could partly repressPTBP3-induced EMT in MCF10A (Supplementary Fig. S6B), butPTBP1 has little effects on the mRNA expressions of ZEB1 andZEB2 (Supplementary Fig. S6C). These results are in agreement
with the previous reports for the role of PTBP1 in EMT (20), and itsuggested that PTBP1 may play a role in EMT transition in adifferent way. Herein, we observed that PTBP1 was downregu-lated in PTBP3-induced EMT; however, PTBP3 did not affect theexpression of PTBP1 targets such as C-myc. It seems a discrepancyemerged in our data. Although C-myc could be regulated byvarious factors such as Notch1, Stat3, and HIF2a (42–44), itsuggested that PTBP1may not play a critical role in regulating theexpressions of C-myc and other target genes in PTBP3-inducedEMT in breast cancer. Future investigations will focus on thebalance of PTBP3 versus PTBP1 in physiologic conditions. Weanticipate that a change in the balance of PTBP3 versus PTBP1activity will play an important role in biological process.
EMT is accompanied by massive changes in cell behavior, suchas cell proliferation, cell differentiation, cell migration, and celladhesion (45). In this study, we showed ectopic overexpression ofPTBP3-induced mammary EMT accompanied with the acquisi-tion of enhanced migration and invasion ability (Fig. 4). Inaddition, PTBP3 protein level was elevated in suspension-culti-vated breast cancer cells, and the PTBP3 overexpression promotedthe generation of CD44high/CD24low subpopulation cells (Fig. 3).Thus, PTBP3 might be essential for nonstem cells to acquiretumor-initiating capacity and function as a mediator of breastcancer cell stemness. Our conclusion is consistent with the ideathat the EMT generates cells with CSC-like activity (46, 47).
ZEB1 is awell-characterized EMT inducer and functions as a keymediator in maintaining the stem cell properties. ZEB1 was
Figure 7.
PTBP3 promoted the in vivo tumor growth and lung metastasis of breast cancer cells. A and B, MDA-MB-231-shCtrl/shPTBP3#1 cells were subcutaneouslyinjected into BALB/c female nude mice (n ¼ 5, for each experimental group), and 8 weeks later, the xenograft tumors were peeled off and weighted. The tumorweights are presented as the means� S.D. � , P < 0.05. C, IHC detection of PTBP3 and ZEB1 protein expression in xenograft tumors formed by MDA-MB-231-shCtrl/shPTBP3#1 cells in BALB/c female nude mice. Scale bar, 100 mm. D, MDA-MB-231-shCtrl/shPTBP3#1 cells were injected via the lateral tail veins. Representativelung images at week 9; corresponding hematoxylin/eosin–stained lung sections are shown. Arrows, lung metastasis foci. E, Lung nodules at week 9 wereanalyzed as the numbers of nodules per mouse. n ¼ 9; �� , P < 0.01, unpaired t test.
Hou et al.
Cancer Res; 78(2) January 15, 2018 Cancer Research396
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

proven to be essential for PTBP3-induced EMT in breast cancerhere. ZEB1 was upregulated upon PTBP3 overexpression, andZEB1 silencing partly reversed the PTBP3-induced EMTcharacters.ZEB1 and ZEB2 were increased at both mRNA and protein levelsupon PTBP3overexpression, while PTBP3had very little influenceon the expression of other EMT-TFs, such as Snail, Slug, and Twist.ZEB1 and ZEB2 were regulated by mir-200 family and formed aZEB/miR-200 feedback loop (48, 49). PTBP3 might indirectlyinfluence ZEB/mir-200 feedback loop through regulating ZEB1.Whether PTBP3 directly regulates ZEB2 expression requires fur-ther investigation.
In conclusion, we have established an important functionalrole for PTBP3 in breast cancer and have also demonstrated thatPTBP3 expression is positively correlated with poor 5-yearsurvival and lymph node metastasis in breast cancer patients.Our clinical analyses point the prospect of PTBP3 as a novelbiomarker for prognosis and a potential molecular therapeutictarget for highly aggressive breast cancer. We also reveal thatPTBP3 regulates an important cell-fate determination event,namely, EMT. In mechanism, PTBP3 regulates ZEB1 by pre-venting ZEB1 30UTR degradation. Although more details stillneed to explore the functions of PTBP3 in a physiologiccondition, our data uncover the importance of PTBP3 in breastcancer progression.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: P. Hou, J. Bai, J. ZhengDevelopment of methodology: P. Hou, L. LiAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): P. Hou, L. Li, F. Chen, H. LiuAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): P. Hou, L. Li, F. Chen, Y. ChenWriting, review, and/or revision of the manuscript: P. Hou, J. Li, J. ZhengAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): P. Hou, L. Li, F. ChenStudy supervision: P. Hou, J. Bai, J. Zheng
AcknowledgmentsThis work was supported by funding from the National Natural Science
Foundation of China (81502280) to Pingfu Hou, 81672845 and 81472663to Jin Bai; The Research Foundation of Xuzhou Medical University(D2015016) to Pingfu Hou; Education Department of Jiangsu Province(no. 15KJA320006) to Jin Bai. This work also was supported by the Project ofInvigorating Health Care through Science, Technology and Education;Jiangsu Provincial Key Medical Discipline (Laboratory); and Jiangsu Pro-vincial Medical Youth Talent Project (QNRC2016776) to Jingjing Li. Theauthors thank Dr. Luis F. Congote and Xiaofei Zheng for providing theplasmids mentioned in Materials and Methods.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
ReceivedMarch26, 2017; revised July 28, 2017; acceptedNovember 20, 2017;published OnlineFirst November 29, 2017.
References1. Fouad TM, Kogawa T, Liu DD, Shen Y, Masuda H, El-Zein R, et al. Overall
survival differences between patients with inflammatory and noninflam-matory breast cancer presenting with distantmetastasis at diagnosis. BreastCancer Res Treat 2015;152:407–16.
2. Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymaltransitions in development and disease. Cell 2009;139:871–90.
3. AcloqueH, AdamsMS, FishwickK, Bronner-FraserM,NietoMA. Epithelial-mesenchymal transitions: the importance of changing cell state in devel-opment and disease. J Clin Invest 2009;119:1438–49.
4. Polyak K, Weinberg RA. Transitions between epithelial and mesenchymalstates: acquisition of malignant and stem cell traits. Nat Rev Cancer2009;9:265–73.
5. Tam WL, Weinberg RA. The epigenetics of epithelial-mesenchymal plas-ticity in cancer. Nat Med 2013;19:1438–49.
6. Puisieux A, Brabletz T, Caramel J. Oncogenic roles of EMT-inducingtranscription factors. Nat Cell Biol 2014;16:488–94.
7. Eger A, Aigner K, Sonderegger S, Dampier B, Oehler S, Schreiber M, et al.DeltaEF1 is a transcriptional repressor of E-cadherin and regulates epithe-lial plasticity in breast cancer cells. Oncogene 2005;24:2375–85.
8. Shirakihara T, Saitoh M, Miyazono K. Differential regulation of epithe-lial and mesenchymal markers by delta EF1 proteins in epithelial-mesenchymal transition induced by TGF-beta. Mol Biol Cell 2007;18:3533–44.
9. Yang J, Weinberg RA. Epithelial-mesenchymal transition: At the crossroadsof development and tumor metastasis. Dev Cell 2008;14:818–29.
10. Wels C, Joshi S, Koefinger P, Bergler H, Schaider H. Transcriptionalactivation of ZEB1 by Slug leads to cooperative regulation of the epithe-lial-mesenchymal transition-like phenotype in melanoma. J Invest Der-matol 2011;131:1877–85.
11. Gregory PA, Bert AG, Paterson EL, Barry SC, Tsykin A, Farshid G, et al. ThemiR-200 family and miR-205 regulate epithelial to mesenchymal transi-tion by targeting ZEB1 and SIP1. Nat Cell Biol 2008;10:593–601.
12. Wang RT, Xu M, Xu CX, Song ZG, Jin H. Decreased expression of miR216acontributes to non-small-cell lung cancer progression. Clin Cancer Res2014;20:4705–16.
13. Chaffer CL, Marjanovic ND, Lee T, Bell G, Kleer CG, Reinhardt F, et al.Poised chromatin at the ZEB1 promoter enables breast cancer cell plasticityand enhances tumorigenicity. Cell 2013;154:61–74.
14. Yang W, Xia Y, Cao Y, Zheng Y, Bu W, Zhang L, et al. EGFR-induced andPKCepsilon monoubiquitylation-dependent NF-kappaB activation upre-gulates PKM2 expression and promotes tumorigenesis. Mol Cell 2012;48:771–84.
15. Ge Z, Quek BL, Beemon KL, Hogg JR. Polypyrimidine tract binding protein1 protects mRNAs from recognition by the nonsense-mediated mRNAdecay pathway. eLife 2016;5:e11155.
16. Cote CA, Gautreau D, Denegre JM, Kress TL, Terry NA, Mowry KL. AXenopus protein related to hnRNP I has a role in cytoplasmic RNAlocalization. Mol Cell 1999;4:431–7.
17. Cobbold LC, Wilson LA, Sawicka K, King HA, Kondrashov AV, Spriggs KA,et al. Upregulated c-myc expression in multiple myeloma by internalribosome entry results from increased interactions with and expressionof PTB-1 and YB-1. Oncogene 2010;29:2884–91.
18. Romanelli MG, Diani E, Lievens PM. New insights into functional roles ofthe polypyrimidine tract-binding protein. Int JMol Sci 2013;14:22906–32.
19. Brazao TF, Demmers J, van IW, Strouboulis J, FornerodM, Romao L, et al. Anew function of ROD1 in nonsense-mediated mRNA decay. FEBS Lett2012;586:1101–10.
20. Hou P, Zhao Y, Li Z, Yao R, Ma M, Gao Y, et al. LincRNA-ROR inducesepithelial-to-mesenchymal transition and contributes to breast cancertumorigenesis and metastasis. Cell Death Dis 2014;5:e1287.
21. Sadvakassova G, Dobocan MC, Difalco MR, Congote LF. Regulator ofdifferentiation 1 (ROD1) binds to the amphipathic C-terminal peptide ofthrombospondin-4 and is involved in its mitogenic activity. J Cell Physiol2009;220:672–9.
22. Yao RS, Jiang H, Ma YH, Wang LP, Wang L, Du J, et al. PRMT7 inducesepithelial-to-mesenchymal transition and promotes metastasis in breastcancer. Cancer Res 2014;74:5656–67.
23. Bai J, YongHM, Chen FF,Mei PJ, LiuH, Li C, et al. Cullin1 is a novelmarkerof poor prognosis and a potential therapeutic target in human breastcancer. Ann Oncol 2013;24:2016–22.
PTBP3 Promotes EMT through Regulating ZEB1 mRNA Stability
www.aacrjournals.org Cancer Res; 78(2) January 15, 2018 397
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

24. Rhodes DR, Yu J, Shanker K, Deshpande N, Varambally R, Ghosh D, et al.ONCOMINE: a cancer microarray database and integrated data-miningplatform. Neoplasia 2004;6:1–6.
25. Ciriello G, Gatza ML, Beck AH, Wilkerson MD, Rhie SK, Pastore A, et al.Comprehensive molecular portraits of invasive lobular breast cancer. Cell2015;163:506–19.
26. ShackletonM,Vaillant F, SimpsonKJ, Stingl J, SmythGK,Asselin-LabatML,et al. Generation of a functional mammary gland from a single stem cell.Nature 2006;439:84–8.
27. Chen B, Zhao AG, Shao J, Mu XY, Jiang L, Liu JW. The effects of PTBP3silencing on the proliferation and differentiation ofMKN45 human gastriccancer cells. Life Sci 2014;114:29–35.
28. Liang X, Shi H, Yang L, Qiu C, Lin S, Qi Y, et al. Inhibition of polypyr-imidine tract-binding protein 3 induces apoptosis and cell cycle arrest, andenhances the cytotoxicity of 5- fluorouracil in gastric cancer cells. Br JCancer 2017;116:903–11.
29. Tano K, Mizuno R, Okada T, Rakwal R, Shibato J, Masuo Y, et al. MALAT-1enhances cell motility of lung adenocarcinoma cells by influencing theexpression of motility-related genes. FEBS Lett 2010;584:4575–80.
30. Xue Y,Ouyang K,Huang J, Zhou Y,OuyangH, LiH, et al. Direct conversionof fibroblasts to neurons by reprogramming PTB-regulated microRNAcircuits. Cell 2013;152:82–96.
31. Degrauwe N, Schlumpf TB, Janiszewska M, Martin P, Cauderay A, ProveroP, et al. TheRNAbinding protein IMP2preserves glioblastoma stem cells bypreventing let-7 target gene silencing. Cell Rep 2016;15:1634–47.
32. Engels B, Jannot G, Remenyi J, Simard MJ, Hutvagner G. Polypyrimidinetract binding protein (hnRNP I) is possibly a conserved modulator ofmiRNA-mediated gene regulation. PLoS One 2012;7:e33144.
33. Shibayama M, Ohno S, Osaka T, Sakamoto R, Tokunaga A, Nakatake Y,et al. Polypyrimidine tract-binding protein is essential for early mousedevelopment and embryonic stem cell proliferation. FEBS J 2009;276:6658–68.
34. Cui J, PlaczekWJ. PTBP1modulation ofMCL1 expression regulates cellularapoptosis induced by antitubulin chemotherapeutics. Cell Death Differ2016;23:1681–90.
35. He X, Pool M, Darcy KM, Lim SB, Auersperg N, Coon JS, et al. Knockdownof polypyrimidine tract-binding protein suppresses ovarian tumor cellgrowth and invasiveness in vitro. Oncogene 2007;26:4961–8.
36. WangC,Norton JT,Ghosh S, Kim J, FushimiK,Wu JY, et al. Polypyrimidinetract-binding protein (PTB) differentially affects malignancy in a cell line-dependent manner. J Biol Chem 2008;283:20277–87.
37. Ooi AT, Gower AC, Zhang KX, Vick JL, Hong L, Nagao B, et al. Molecularprofiling of premalignant lesions in lung squamous cell carcinomasidentifies mechanisms involved in stepwise carcinogenesis. Cancer PrevRes 2014;7:487–95.
38. YamamotoH, Tsukahara K, Kanaoka Y, Jinno S, OkayamaH. Isolation of amammalian homologue of a fission yeast differentiation regulator. MolCell Biol 1999;19:3829–41.
39. David CJ, Chen M, Assanah M, Canoll P, Manley JL. HnRNP proteinscontrolled by c-Myc deregulate pyruvate kinase mRNA splicing in cancer.Nature 2010;463:364–8.
40. Huang J, Zhang A, Ho TT, Zhang Z, Zhou N, Ding X, et al. Linc-RoRpromotes c-Myc expression through hnRNP I and AUF1. Nucleic Acids Res2016;44:3059–69.
41. Tan LY, Whitfield P, Llorian M, Monzon-Casanova E, Diaz-Munoz MD,Turner M, et al. Generation of functionally distinct isoforms of PTBP3 byalternative splicing and translation initiation. Nucleic Acids Res 2015;43:5586–600.
42. Palomero T, Lim WK, Odom DT, Sulis ML, Real PJ, Margolin A, et al.NOTCH1 directly regulates c-MYC and activates a feed-forward-looptranscriptional network promoting leukemic cell growth. Proc Natl AcadSci U S A 2006;103:18261–6.
43. Bowman T, Broome MA, Sinibaldi D, Wharton W, Pledger WJ, SedivyJM, et al. Stat3-mediated Myc expression is required for Src transfor-mation and PDGF-induced mitogenesis. Proc Natl Acad Sci U S A2001;98:7319–24.
44. Gordan JD, Bertout JA, Hu CJ, Diehl JA, Simon MC. HIF-2alpha promoteshypoxic cell proliferation by enhancing c-myc transcriptional activity.Cancer Cell 2007;11:335–47.
45. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell2011;144:646–74.
46. Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, et al. Theepithelial-mesenchymal transition generates cells with properties of stemcells. Cell 2008;133:704–15.
47. Morel AP, LievreM, ThomasC,HinkalG, Ansieau S, Puisieux A.Generationof breast cancer stem cells through epithelial-mesenchymal transition.PLoS One 2008;3:e2888.
48. Brabletz S, Brabletz T. The ZEB/miR-200 feedback loop–amotor of cellularplasticity in development and cancer? EMBO Rep 2010;11:670–7.
49. Hill L, Browne G, Tulchinsky E. ZEB/miR-200 feedback loop: at thecrossroads of signal transduction in cancer. Int J Cancer 2013;132:745–54.
Cancer Res; 78(2) January 15, 2018 Cancer Research398
Hou et al.
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

2018;78:387-398. Published OnlineFirst November 29, 2017.Cancer Res Pingfu Hou, Lin Li, Fang Chen, et al.
Mesenchymal Transition in Breast Cancer−Epithelial PTBP3-Mediated Regulation of ZEB1 mRNA Stability Promotes
Updated version
10.1158/0008-5472.CAN-17-0883doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2017/11/29/0008-5472.CAN-17-0883.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/78/2/387.full#ref-list-1
This article cites 49 articles, 9 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/78/2/387To request permission to re-use all or part of this article, use this link
on February 1, 2020. © 2018 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst November 29, 2017; DOI: 10.1158/0008-5472.CAN-17-0883

























![91-302 [Multilateral CSA Staff Notice] - BCSC](https://img.dokumen.tips/doc/110x75/6219f7ccfa4b8f19181bc819/91-302-multilateral-csa-staff-notice-bcsc.jpg)



