Embed Size (px)
Citation preview

Structure
Article
Protein-RNA and Protein-Protein Recognitionby Dual KH1/2 Domainsof the Neuronal Splicing Factor Nova-1Marianna Teplova,1,7 LucyMalinina,1,7,9 Jennifer C. Darnell,3,7 Jikui Song,1Min Lu,6 Ruben Abagyan,5 KiranMusunuru,3,4,8
Alexei Teplov,1 Stephen K. Burley,2,4,10 Robert B. Darnell,2,3 and Dinshaw J. Patel1,*1Structural Biology Program, Memorial Sloan-Kettering Cancer Center, New York, NY 10021, USA2Howard Hughes Medical Institute3Laboratory of Molecular Neuro-Oncology4Laboratory of Molecular Biophysics
Rockefeller University, New York, NY 10021, USA5Department of Molecular Biology, The Scripps Research Institute, La Jolla, CA 92037, USA6Biochemistry Department, Weill Medical College of Cornell University, New York, NY 10021, USA7CIC bioGUNE, Technology Park of Bizkaia, 48160 Derio-Bilbao, Spain8Department of Stem Cell and Regenerative Biology, Harvard University, Cambridge, MA 02138, USA9These authors contributed equally to this work10Present address: Research & Development, Eli Lilly & Co, Indianapolis, IN 46285, USA
*Correspondence: [email protected]
DOI 10.1016/j.str.2011.05.002
SUMMARY
Nova onconeural antigens are neuron-specific RNA-binding proteins implicated in paraneoplastic opso-clonus-myoclonus-ataxia (POMA) syndrome. Novaharbors three K-homology (KH) motifs implicated inalternate splicing regulation of genes involved ininhibitory synaptic transmission. We report the crys-tal structure of the first two KH domains (KH1/2)of Nova-1 bound to an in vitro selected RNA hairpin,containing a UCAG-UCAC high-affinity binding site.Sequence-specific intermolecular contacts in thecomplex involve KH1 and the second UCAC repeat,with the RNA scaffold buttressed by interactionsbetween repeats. Whereas the canonical RNA-binding surfaceofKH2 in theabovecomplexengagesin protein-protein interactions in the crystalline state,the individual KH2 domain can sequence-specificallytarget the UCAC RNA element in solution. Theobserved antiparallel alignment of KH1 and KH2domains in thecrystal structureof thecomplexgener-ates a scaffold that could facilitate target pre-mRNAlooping on Nova binding, thereby potentially explain-ing Nova’s functional role in splicing regulation.
INTRODUCTION
The generation of a versatile repertoire of functionally diverse
proteins is critically dependent on alternate splicing of pre-
mRNAs. Alternate splicing of the subunits of all the main neuro-
transmitter receptors can influence their localization, ligand-
binding, signal-transducing, and electrophysiological properties.
Aberrant splicing of mRNAs in highly specialized cells, such as
930 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All right
neurons, can impact proteins critical for neuronal function,
leading to the onset of neurological disease (Dredge and Darnell,
2003; Dredge et al., 2001; Licatalosi and Darnell, 2006). The
proteins of the Nova (neuro-oncological ventral antigen) family,
which are exclusively expressed in central nervous system
(CNS) neurons, are important regulators of neuronal RNAmetab-
olism. The Nova proteins are target antigens in the autoimmune
disorder paraneoplastic opsoclonus-myoclonus ataxia (POMA)
(Musunuru and Darnell, 2001). POMA is a neurodegenerative
syndrome that originates when systemic malignant tumors
express proteins normally sequestered in the central nervous
system (Albert and Darnell, 2004). The immune system recog-
nizes these antigens to be nonself, and the ensuing response
results in neuronal degeneration. Homozygous Nova-1 knockout
mice appear to exhibit normal gestation and development, but
the resulting litters show apoptosis of motor neurons in the brain
stem and spinal cord and become incapacitated by progressive
motor system failure (Jensen et al., 2000a).
K-homology (KH) domains are RNA-binding elements (Grishin,
2001; Siomi et al., 1993), identified in hnRNP proteins involved in
RNA stabilization and translational control (Burd and Dreyfuss,
1994; Ostareck-Lederer et al., 1998; Valverde et al., 2008).
Nova-1 and Nova-2 proteins each contain three proteolytically
stable KH domains (Figure 1A), connected by flexible linkers.
The crystallographic structures of Nova KH3 domains exhibit a
three-stranded antiparallel b sheet backed by three a helices
(Lewis et al., 1999). RNA targets for the Nova KH3 domain (and
adjacent C-terminal residues) have been identified by in vitro
selection experiments (Jensen et al., 2000b). The optimal RNA
target exhibits a YCAY tetranucleotide element (Y is a pyrimidine)
as part of an accessible 12-residue loop segment within the
context of a RNA hairpin. The crystal structure of this complex
has been determined at 2.4 A resolution, with the majority of
the intermolecular contacts restricted to the extended UCAC
segment of the RNA loop, which is pinioned between the
invariant Gly-X-X-Gly motif and a variable loop of the KH domain
s reserved

A
B
D
C
E
Figure 1. Sequences of Nova KH Domains
and RNA Hairpin Target and the Structure
of the Nova-1 KH1/2-RNA Hairpin Complex
in the Crystal
(A) Schematic of three KHdomains and intervening
linkers in Nova proteins. Red-colored numbers
indicate the length of linker segments in Nova-1.
(B) The Nova-1 KH1/2 construct used in the
current project.
(C) The sequence of the in vitro selected 25-mer
RNA hairpin. The tandem UCAN sites are colored
in gold (U9–G12) and cyan (U13–C16). Structural
studies were also undertaken on a sequence
containing a 5BrU2,A24 pair.
(D) Sequence and secondary structure (Lewis
et al., 1999) alignments of Nova KH1, KH2, and
KH3 domains, with conserved residues shown in
red. The GXXG motif and so-called variable loop,
are denoted as I and V, respectively.
(E) Ribbon representation of the structure of the
Nova-1 KH1/2-RNA hairpin complex in the crystal.
The stoichiometry is KH1/2:RNA hairpin of 2:2. The
color codes are RNA in green, KH1 in gold and KH2
in cyan. The UCAC motif interacting with KH1 is
colored inblue. See also Table 1 and FiguresS1–S3.
Structure
Structure of Nova-1 KH1/2-RNA Complex
(Lewis et al., 2000). This CA segment, which is most intolerant to
mutations, is recognized by Watson-Crick-like hydrogen bond-
ing patterns from protein residues, whereas the flanking nucleo-
tides are restricted to pyrimidine residues.
Full-length Nova proteins containing three KH domains have
beenshown to targetand regulatealternativesplicingeventswithin
neuronal pre-mRNAs that contain repeats of the (YCAY) binding
element (Buckanovich and Darnell, 1997; Dredge and Darnell,
2003; Dredge et al., 2005; Licatalosi et al., 2008; Ule et al., 2006;
Zhang et al., 2010). RNA aptamers generated against full length
Nova protein adopt a RNA hairpin loop scaffold containing
a conserved [YCAY-(N)0-2]3 sequence in the loop segment (Buck-
anovich andDarnell, 1997; Yang et al., 1998). The published struc-
ture of the Nova KH3-RNA hairpin complex (Lewis et al., 2000)
doesnot addresshowmultipleKHdomainsofNovaproteinsmight
cooperate in recognizing tandem YCAY repeat modules. To
address this issue in a systematicmanner, we set out to determine
the structure of aNovacomplex containing adjacent KH1andKH2
domains (designated KH1/2; Figure 1B) targeted to a pair of
tandem YCAY RNA sites. Such an opportunity was available
because in vitro selection experiments had identified an RNA
hairpin architecture (Figure 1C), containing a potential 15-mer
Structure 19, 930–944, July 13, 2011
loop within which is nestled an UCAG-
UCAC segment, which binds the Nova-1
KH1/2 construct (Figure 1B) with high
affinity (Musunuru and Darnell, 2004).
Mutagenic analysis established that both
CAstepswithin theUCAG-UCACsegment
aremost intolerant tomutations and there-
fore represent key residues involved either
inprotein recognition, and/or inbuttressing
the recognition interface.
We now report the crystal structure
of the Nova-1 KH1/2 construct bound to
the UCAG-UCAC segment-containing RNA hairpin. Strikingly,
the RNA hairpin is targeted solely by the KH1 domain, with
both loop UCAG and UCAC tetranucleotide sites contributing
to recognition. Both the crystal structure and binding studies
establish that the KH2 domain does not interact with the
UCAG-UCAC-containing RNA hairpin. In the crystal, the RNA-
binding surface of the KH2 domain instead engages in protein-
protein interactions with symmetry-related KH2 domain. By
contrast, Nuclear magnetic resonance (NMR) chemical shift
perturbations, gel-shift, and ITC binding studies demonstrate
that the KH2 domain does target UCAY tetranucleotide-contain-
ing RNAs in a sequence specificmanner in solution. Nova’s func-
tional role in splicing regulation could be explained by the
observed antiparallel alignment of KH1 and KH2 domains in
the crystal structure of the complex, thereby providing an ideal
scaffold to induce target pre-mRNA looping on Nova binding.
RESULTS
Crystallization and Structure Determination of ComplexA Nova-1 KH1/2-expression construct (Figure 1B) was engi-
neered lacking a 24-amino acid segment between the KH1 and
ª2011 Elsevier Ltd All rights reserved 931

Table 1. Crystal Data, Data Collection, and Refinement Statistics
for Nova-1 KH1/2-RNA Hairpin Aptamer Complex
Crystal data Type I Type II
Space group C 2 C 2
Unit cell parameters
a (A) 155.3 159.5
b (A) 37.2 37.9
c (A) 34.8 38.5
b� 94.4 96.6
Asymmetric unit
RNA bases 23 25
KH1+KH2 residues 148 155
Solvent: H2O / ion 77/3 (K+, 2Mg2+) 155/3 (K+, 2Mg2+)
Data collection
X-ray source
detector
APS, SBC - 19ID
APS-1 CCD
NSLS, X4A
Quantum IV
Wavelength (A) 1.0080 0.919826
Resolution (A)
Range 20–2.3 20–1.94
Last shell 2.38–2.30 2.00–1.94
Unique reflections 8981 (873) 16,202 (1,513)
Redundancy 3.44 6.55
Completeness (%) 99.7 (97.9) 94.2 (89.6)
Mosaicity 0.88 1.32
R–mergea 0.068 (0.44) 0.117 (0.49)
Refinement (F>0)
Work/free reflections 8551/430 15,372/830
R-factor/R-free 0.193/0.248 0.227/0.268
Rmsd
Bond lengths (A) 0.011 0.013
Bond angles (�) 1.335 1.682
Average B-factors (A2) 27.9 25.0
Values in parentheses correspond to the last resolution shell.a R-merge = ShklSiII(hkl)i - <I(hkl)>I/ShklSi<I(hkl)i> over i observations.
Structure
Structure of Nova-1 KH1/2-RNA Complex
KH2 domains (Figure 1A, dashed line), which is encoded by an
alternatively spliced exon. The protein was expressed and puri-
fied as described in Supplemental Experimental Procedures
available online. RNA in vitro selection carried out previously
against this Nova-1 KH1/2 construct identified a RNA hairpin
composed of a 5-base pair stem and a 15-nucleotide loop con-
taining conserved 8-base UCAG-UCAC sequence (Figure 1C)
(Musunuru and Darnell, 2004).
We crystallized complexes of Nova-1 KH1/2 protein with two
25-nucleotide RNA aptamer hairpins, which differed at the
2,24 base pair position in the stem segment. One of the RNA
hairpins (type I) contained aG2,C24 pair (Figure 1C; Figure S1A),whereas the other (type II) contained a 5BrU2,A24 pair (Fig-
ure S1B). The complexes of Nova-1 KH1/2 with type I and type
II RNA hairpins were both crystallized in space group C2, with
similar unit cell dimensions, except for a 10.5% difference in
the unit cell c dimension (Table 1). Structures were determined
via molecular replacement (MR) using the published structure
of a KH domain from the Nova-2 KH3-RNA aptamer hairpin
complex (Lewis et al., 2000) as a search model (see Supple-
932 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All right
mental Experimental Procedures). The KH1 and KH2 domains
in the Nova-1 KH1/2-RNA hairpin complex adopt the same
baabba topology (Figure 1D), reported earlier for the KH3 domain
of Nova-1 and Nova-2 in the free state (Lewis et al., 1999) and in
the Nova-2 KH3-RNA hairpin complex (Lewis et al., 2000). The
superimposed Nova-1 KH1 and KH2 domains in the RNA bound
state exhibit a pair-wise Ca rmsd of 1.1 A for residues 6–79
(excluding residues 44–52 of the variable loop). The asymmetric
unit contains one KH1/2 molecule and one RNA molecule, with
similar overall packing arrangement for both complexes. The
differences in the structures of type I and type II complexes are
outlined in the Supplemental Experimental Procedures and
Figure S1.
Stoichiometry of Complex in Crystal and in SolutionThe crystal structure of two Nova-1 KH1/2-RNA hairpin complex
molecules related by 2-fold crystallographic symmetry is shown
in Figure 1E. The KH1 (gold) and KH2 (cyan) domains are aligned
in an antiparallel orientation. The UCAG-UCAC-containing RNA
hairpin (green) is targeted solely by the KH1 domain, whereas
the canonical RNA-binding surface of KH2 engages in protein-
protein interactions with symmetry-related KH2. The packing
arrangement of KH1/2-RNA hairpin complexes in the crystal
and the details of KH2-KH2 interface are outlined in the Supple-
mental Experimental Procedures and Figure S2.
In contrast, analytical ultracentrifugation measurements indi-
cate that KH1/2 exists as a monomer in solution at 10–200 mM
concentration range, and the KH1/2-RNA complex sediments
as a 1:1 complex under these conditions (for details see Supple-
mental Experimental Procedures and Figure S3).
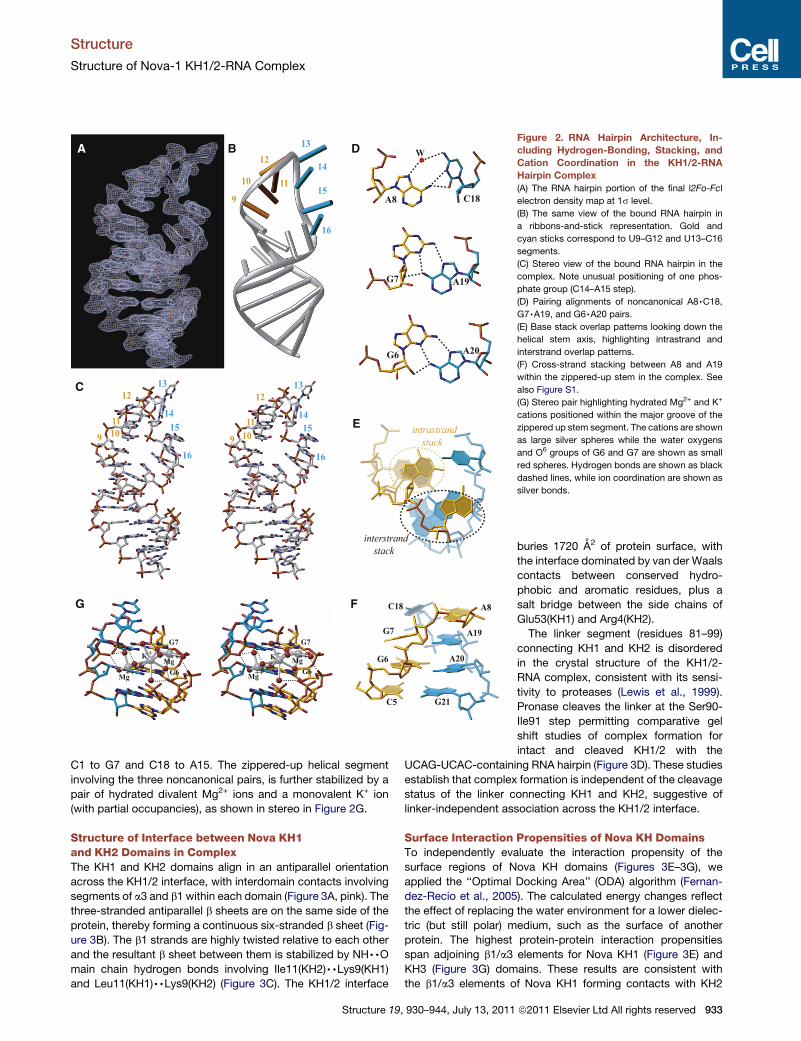
RNA Hairpin Architecture in the Bound StateThe entire UCAG-UCAC-containing RNA hairpin (Figure 2A) can
be traced in the crystal structure of the type II complex, whereas
the 50-C1-G2 segment is disordered in the crystal structure of
the type I complex. The simulated annealing 2Fo-Fc omit map
of the UCAC RNA site at 1s level for the type I complex is shown
in Figure S1C. The conformation of the UCAG-UCAC is of great-
est interest, due to the potential of each tetranucleotide element
serving as a KH domain binding site. All four-hairpin residues
within the gold-colored U9 to G12 segment (Figure 2B) form
a stacked array. By contrast, residues U13 and C14 loop out,
whereas residues A15 and C16 form a stacked pair within the
cyan-colored U13 to C16 segment (Figure 2B). The conformation
of the bound RNA hairpin is shown in stereo in Figure 2C and
establishes extensive stacking interactions that propagate
from the paired stem segment into both arms of the hairpin,
with the stacking discontinuities restricted to the G12 to A15
segment.
The Watson-Crick paired stem of the RNA hairpin zippers up
through formation of G6,A20, G7,A19, and A8,C18 nonca-
nonical pairs (Figure 2D). The two noncanonical G,A pairs are
of the sheared type, whereas the A8,C18 noncanonical pair is
buckled and involves bifurcated and water-mediated hydrogen
bonds. The small twist angle at the G6-G7 step and the large
twist angle at the G7-A8 step, results in a pronounced cross-
strand stack between residues A8 and A19 (Figures 2E and
2F). This results in a continuous helical stack that spans G25 to
A19 and A8 to G12. The other continuous helical stack spans
s reserved
A B
C
13
14
15
16
12
1110
9
13
1415
16
12
11109
13
1415
16
12
11109
Mg++
K+
G6
G7
Mg++
Mg++
K+
G6
G7
Mg++
G
G7 A19
D
A8 C18
W
G6 A20
C5
G6
G7
A8
G21
A20
A19
C18
intrastrand stack
interstrand stack
E
F
Figure 2. RNA Hairpin Architecture, In-
cluding Hydrogen-Bonding, Stacking, and
Cation Coordination in the KH1/2-RNA
Hairpin Complex
(A) The RNA hairpin portion of the final l2Fo-Fcl
electron density map at 1s level.
(B) The same view of the bound RNA hairpin in
a ribbons-and-stick representation. Gold and
cyan sticks correspond to U9–G12 and U13–C16
segments.
(C) Stereo view of the bound RNA hairpin in the
complex. Note unusual positioning of one phos-
phate group (C14–A15 step).
(D) Pairing alignments of noncanonical A8,C18,
G7,A19, and G6,A20 pairs.
(E) Base stack overlap patterns looking down the
helical stem axis, highlighting intrastrand and
interstrand overlap patterns.
(F) Cross-strand stacking between A8 and A19
within the zippered-up stem in the complex. See
also Figure S1.
(G) Stereo pair highlighting hydrated Mg2+ and K+
cations positioned within the major groove of the
zippered up stem segment. The cations are shown
as large silver spheres while the water oxygens
and O6 groups of G6 and G7 are shown as small
red spheres. Hydrogen bonds are shown as black
dashed lines, while ion coordination are shown as
silver bonds.
Structure
Structure of Nova-1 KH1/2-RNA Complex
C1 to G7 and C18 to A15. The zippered-up helical segment
involving the three noncanonical pairs, is further stabilized by a
pair of hydrated divalent Mg2+ ions and a monovalent K+ ion
(with partial occupancies), as shown in stereo in Figure 2G.
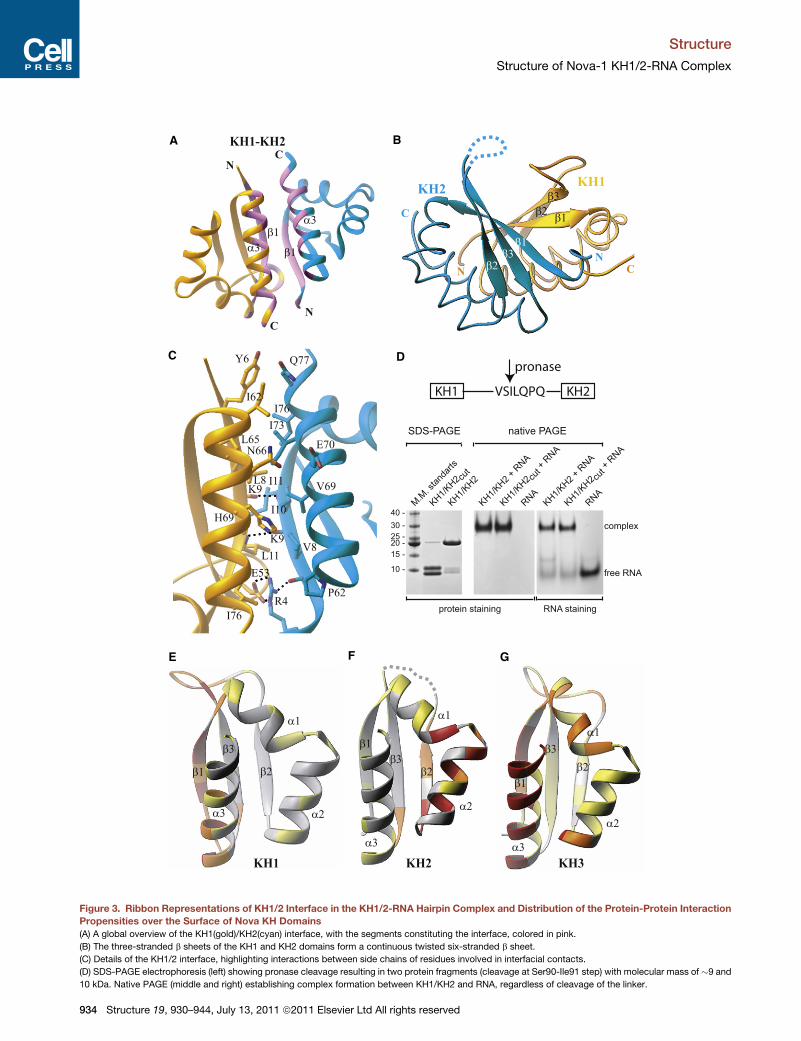
Structure of Interface between Nova KH1and KH2 Domains in ComplexThe KH1 and KH2 domains align in an antiparallel orientation
across the KH1/2 interface, with interdomain contacts involving
segments of a3 and b1 within each domain (Figure 3A, pink). The
three-stranded antiparallel b sheets are on the same side of the
protein, thereby forming a continuous six-stranded b sheet (Fig-
ure 3B). The b1 strands are highly twisted relative to each other
and the resultant b sheet between them is stabilized by NH,,Omain chain hydrogen bonds involving Ile11(KH2),,Lys9(KH1)and Leu11(KH1),,Lys9(KH2) (Figure 3C). The KH1/2 interface
Structure 19, 930–944, July 13, 2011
buries 1720 A2 of protein surface, with
the interface dominated by van der Waals
contacts between conserved hydro-
phobic and aromatic residues, plus a
salt bridge between the side chains of
Glu53(KH1) and Arg4(KH2).
The linker segment (residues 81–99)
connecting KH1 and KH2 is disordered
in the crystal structure of the KH1/2-
RNA complex, consistent with its sensi-
tivity to proteases (Lewis et al., 1999).
Pronase cleaves the linker at the Ser90-
Ile91 step permitting comparative gel
shift studies of complex formation for
intact and cleaved KH1/2 with the
UCAG-UCAC-containing RNA hairpin (Figure 3D). These studies
establish that complex formation is independent of the cleavage
status of the linker connecting KH1 and KH2, suggestive of
linker-independent association across the KH1/2 interface.
Surface Interaction Propensities of Nova KH DomainsTo independently evaluate the interaction propensity of the
surface regions of Nova KH domains (Figures 3E–3G), we
applied the ‘‘Optimal Docking Area’’ (ODA) algorithm (Fernan-
dez-Recio et al., 2005). The calculated energy changes reflect
the effect of replacing the water environment for a lower dielec-
tric (but still polar) medium, such as the surface of another
protein. The highest protein-protein interaction propensities
span adjoining b1/a3 elements for Nova KH1 (Figure 3E) and
KH3 (Figure 3G) domains. These results are consistent with
the b1/a3 elements of Nova KH1 forming contacts with KH2
ª2011 Elsevier Ltd All rights reserved 933
V8
Q77
I76I73
I11
I10
P62
V69
Y6
I76
H69
N66L65
I62
L8
L11
NC
CN
α3
α3
α1
α2α3
α1
α2
α3
β1
β1
β2β1
E70
K9
K9
E53
R4
KH1-KH2
KH1 KH2
KH1KH2
N
C
CN
β1
β2β3
β1
β3β2
M.M. s
tanda
rts
KH1/KH2 cu
t
KH1/KH2
KH1/KH2 +
RNA
KH1/KH2 cu
t + R
NA
RNA
protein staining RNA staining
SDS-PAGE native PAGE
free RNA
complex
10 -
15 -20 -25 -30 -40 -
KH1/KH2 +
RNA
KH1/KH2 cu
t + R
NA
RNA
VSILQPQKH1 KH2
pronase
β2β3
β3α1
α2
α3
β2β1
KH3
β3β1
A B
C D
E F G
Figure 3. Ribbon Representations of KH1/2 Interface in the KH1/2-RNA Hairpin Complex and Distribution of the Protein-Protein Interaction
Propensities over the Surface of Nova KH Domains
(A) A global overview of the KH1(gold)/KH2(cyan) interface, with the segments constituting the interface, colored in pink.
(B) The three-stranded b sheets of the KH1 and KH2 domains form a continuous twisted six-stranded b sheet.
(C) Details of the KH1/2 interface, highlighting interactions between side chains of residues involved in interfacial contacts.
(D) SDS-PAGE electrophoresis (left) showing pronase cleavage resulting in two protein fragments (cleavage at Ser90-Ile91 step) with molecular mass of �9 and
10 kDa. Native PAGE (middle and right) establishing complex formation between KH1/KH2 and RNA, regardless of cleavage of the linker.
Structure
Structure of Nova-1 KH1/2-RNA Complex
934 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All rights reserved

A11
C14
A15
R54L41
I39
Q32
w1
w2
α1
α2G24
G18
U13
R54C14
S44K45
G12
A11
C10
K43A15Q32
w3
w2
w1
K40 C16
U9
1716
15
10
11
9
12
13
14
CC C
U
STEM
A AC
UG
U13
O2'O3'
O5' C16
VDWK23
K40G22
G18
A
B
C D
Figure 4. KH1-RNA Interactions in Complex
(A) Overview of interactions between RNA hairpin loop (U9–G12 in gold and
the U13–C16 in cyan; see inset) and the N-terminal half of the KH1 domain
(Tyr15–Lys45 in gold, encompassing a1, loop I, a2, and b2 segments). Bridging
water molecules at the interface are labeled w1, w2, and w3.
(B) Details of intermolecular contacts involving the C14-A15 segment of the
RNA, and amino acids Arg54, Lys41, and Gln32 of the KH1 domain. Note
buttressing interactions between the base and sugar of A11 and C14. The side
chain of Lys40 is removed for clarity.
(C) Details of intermolecular contacts involving U13 base and sugar hydroxyl
group of the RNA, and backbone atoms of amino acids Gly18, Gly22, and
Lys23 of the KH1 domain.
(D) Intermolecular contacts between the Watson-Crick edge of C16 and the
side chain amine (hydrogen bonding) and methylene groups (van der Waals)
of Lys40.
Structure
Structure of Nova-1 KH1/2-RNA Complex
(Figures 3A) in the Nova KH1/2-RNA hairpin complex, and with
the b1/a3 elements of Nova KH3 forming contacts with
symmetry-related KH3 in the crystal (Lewis et al., 2000). By
contrast, the highest surface interaction propensities span
adjoining b2/a2 elements, the loop between a1- and a2-helices,
and the C terminus of a1-helix for Nova KH2 (Figure 3F). These
elements are located on the canonical RNA-binding surface of
KH domains and it is this surface of KH2 that participates in
protein-protein interactions with symmetry-related KH2 in the
crystal (Figure S2B).
Recognition between KH1 Domain of Nova KH1/2and RNA HairpinProtein-RNA recognition in the Nova-1 KH1/2-RNA hairpin
complex involves only one of the two KH domains and both
UCAG-UCAC tetranucleotide elements within the RNA hairpin.
Specifically, the KH1 domain primarily targets the U13 to C16
segment, and the C10 base of the preceding U9 to G12 segment
(Figure 4A), with C14 and A11 buttressing each other through
hydrogen bonding interactions (Figure 4B). The U13 base loops
out of the hairpin and interacts with the KH1 domain through
both its base and sugar residues (Figure 4A). The U13 base stacks
over theGly18-Ser19peptidebond,withO4ofU13 formingvander
Waals contact with Cb of Ser19 and O2 of U13 forming a weak
hydrogen bond with main chain NH of Lys23 (Figure 4C). In addi-
tion, the 20-OH of the U13 sugar forms a bifurcated hydrogen
bond with the main chain O atoms of Gly18 and Gly22. The C14
base forms two hydrogen bonds through its Watson-Crick edge
with the guanidinium group of Arg54, with C14 buttressed in posi-
tion through a network of base-sugar/sugar-base-phosphate
hydrogen bondswith A11 of the adjacent U9 toG12 segment (Fig-
ure 4B). The next residue, A15 forms a pair of hydrogen bonds to
themain chainNHandOatomsof Leu41, and is further buttressed
in place by a bifurcated water-mediated hydrogen bond from its
base (N3) and sugar (20-OH) atoms to the side chain of Gln32.
The network of hydrogen bonds and water-mediated bridges
(w1 and w2) that direct the observed intermolecular recognition
and buttressing events, are specific for cytosine at position 14
and adenine residues at positions 15 and 11 in the complex (Fig-
ure 4B). The Lys40 and Lys43 side chains are directed toward
the C16 and C10 bases, with the specific intermolecular contacts
involving these lysine side chains differing between the type I and
type II complexes (Figure 4A). The Lys43 3-amino group forms
a hydrogen bond with either N3 atom of C16 or O2 atom of C10.
When Lys43 is occupied in the interaction with C10, the N3 atom
of C16 is hydrogen bonded to the Lys40 3-amino group, with the
rest of the Lys40 side chain forming van der Waals contacts with
O2 atom of C16 (Figure 4D).
Binding of Nova KH1/2 Domainsto UCAG-UCAC-Containing RNA HairpinWe have used filter-binding assays to measure the binding
affinities of the individual KH2 and KH3 domains, KH1/2 dual
(E–G) Distribution of the protein-protein interaction propensities over the protein surface as calculated by the optimal docking area (ODA) approach for three
different KH domains. Ribbon representation of KH1 (E) (this study), KH2 domains of the Nova-1 KH1/2 protein (F) (this study), and Nova-2 KH3 domain (G)
(Lewis et al., 2000). Structures are colored by the absolute magnitude of the ODA signal from the strongest in red, through medium in orange and weak in yellow,
to the weakest in white. See also Figure S5.
Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All rights reserved 935

1 101 102 103 104
0
20
40
60
80
100
1 10 102 103 104
1 101 102 103 104
0
20
40
60
80
100
1 10 102 103 104 1 10 102 103 104
KH1/2 RNA KH1/2 RNA
KH1/2 WT
K40,43Q S14E
Nova KH1/2 Nova KH2 Nova KH3
KH1/2 RNA KH3 RNA
KH1/2/3 RNA KH1/2 RNA KH3 RNA
KH1/2/3 RNA
KH1/2 KH2 KH3
KH1/2/3
KH1/2 RNA KH3 RNA
KH1/2/3 RNA
kc1
% to
tal R
NA
boun
d
[protein], nM [protein], nM
[protein], nM
% to
tal R
NA
boun
d
3'5'GAGGA
CUCCC
CC
A UG A
U
CAC
KH3RNA hairpin
C3'5'
GCGCGGA
GCGCGAAC
UC
G CU
A
CCA
KH1/2RNA hairpin
G3'5'
ACUC
AC
G
CUGAG
AAC
UC
UC
U
A
UU
C UA
GG
C
CA
KH1/2/3RNA hairpin
A B C
D E
F G H
Figure 5. Filter Binding Assays for Nova KH
Domain-RNA Complexes
(A) The sequence of the Nova-2 KH3-binding
20-nt RNA hairpin containing a single UCAN site
(Lewis et al., 2000).
(B) The sequence of the Nova-1 KH1/2-binding
25-nt RNA hairpin containing a (UCAN)2 site (Mu-
sunuru and Darnell, 2004) that was used in this
study.
(C) The sequence of the Nova-1 KH1/2/3-binding
32-nt RNA hairpin containing a (UCAN-N)3 site
(Buckanovich and Darnell, 1997).
(D) Filter-binding assays for complex formation of
Nova KH1/2/3 (open circles), KH1/2 (black
circles), KH2 (red circles), and KH3 (green circles)
with KH1/2 RNA hairpin containing a (UCAN)2site (B).
(E) Filter-binding assays for complex formation of
wild-type Nova KH1/2 (black circles), Ser14Glu
mutant (green circles), and Lys40Gln, Lys43Gln
dual mutant (red circles) with KH1/2 RNA hairpin
containing a (UCAN)2 site (B).
(F) Filter-binding assays for complex formation of
Nova KH1/2 with RNA hairpins containing a single
UCAN site (A) (red circles), (UCAN)2 site (B) (black
circles), and (UCAN-N)3 site (C) (green circles).
(G) Filter-binding assays for complex formation of
Nova KH2 with RNA hairpins containing a single
UCAN site (A) (red circles), (UCAN)2 site (B) (black
circles), (UCAN-N)3 site (C) (green circles), and
a RNA hairpin specific for FMRP KH2 domain
(open circles).
(H) Filter-binding assays for complex formation of
Nova KH3 with RNA hairpins containing a single
UCAN site (A) (red circles), (UCAN)2 site (B) (black
circles), and (UCAN-N)3 site (C) (green circles).
Structure
Structure of Nova-1 KH1/2-RNA Complex
domains, and full-length KH1/2/3 Nova protein to a panel of
three RNA hairpin targets. These include the KH3-binding RNA
aptamer containing a single UCAC site (Figure 5A), the KH1/2-
binding RNA aptamer containing a UCAG-UCAC site (Figure 5B),
and the KH1/2/3-binding RNA aptamer containing a (YCAY-N)3site (Figure 5C). Thus, although the intact KH1/2/3 Nova protein
(Figure 5D, open circles) and KH1/2 dual domains (Figure 5D,
black circles) bind the UCAG-UCAC-containing RNA aptamer
target (Figure 5B) with mM affinity, the KH3 domain (Figure 5D,
green circles) exhibits reduced binding affinity, whereas the
KH2 domain (Figure 5D, red circles) exhibits no measurable
binding affinity.
We have used filter-binding assays to compare the binding
affinities of wild-type (KD of 0.42 ± 0.11 mM; Figure 5E, black
circles) and mutant KH1/2 dual domains for the KH1/2-binding
UCAG-UCAC-containing RNA hairpin (Figure 5B). Replacement
of Ser14 in KH1 by its Glu counterpart observed at this position in
KH3, within the context of the KH1/2 construct, resulted in an
�4-fold decrease in RNA hairpin binding affinity to 1.6 ±
0.2 mM (Figure 5E, green circles). Replacement of Lys40 and
Lys43 in KH1 by their Gln counterparts observed at these posi-
tions in KH2, within the context of the KH1/2 construct, resulted
in no measurable RNA hairpin binding affinity (Figure 5E, red
circles).
936 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All right
Binding of Nova KH Domains to (YCAY)n-ContainingRNA HairpinsFilter-binding assays have also been used to measure the
binding affinities of the KH2 and KH3 individual domains (KH1
is unstable as an individual domain) and KH1/2 dual domains
to RNA aptamer hairpin targets containing (YCAY)n sites, where
n = 1,2,3 (Figures 5A–5C). The KH1/2 dual domains bind most
tightly to the RNA aptamers containing UCAG-UCAC (KD =
0.42 ± 0.11 mM; Figure 5F, black circles) sites and (YCAY-N)3(KD = 1.9 ± 0.3 mM; Figure 5F, green circles), with weaker binding
affinity to the RNA aptamer containing a single UCAC site (Fig-
ure 5F, red circles).
The KH2 domain binds very weakly to all three RNA aptamers
(Figure 5G), with the residual binding observed for the RNA
aptamer containing a (YCAY-N)3 site (Figure 5G, green circles)
being nonspecific, because a similar binding curve is observed
for an unrelated FMRP KH2 domain-binding RNA aptamer, kc1
(Figure 5G, open circles).
Finally, the KH3 domain binds most tightly to its own RNA
aptamer containing a single UCAC site (KD = 0.58 ± 0.09 mM;
Figure 5H, red circles), less tightly to the RNA aptamer containing
a (YCAY-N)3 site (KD = 3.4 ± 0.6 mM; Figure 5H, green circles),
and weakest to the RNA aptamer containing a UCAG-UCAC
site (Figure 5H, black circles).
s reserved
1H (ppm)
15N
(pp
m)
7.95 7.90 7.85 7.80 7.75
125.5
125.0
124.5 V28f
1H (ppm)7.95 7.90 7.85 7.80 7.75
125.5
125.0
124.5 V28f
V28c
1H (ppm)7.95 7.90 7.85 7.80 7.75
125.5
125.0
124.5
V28c
(ppm
)av
eδΔ
0
0.2
0.4
0.6
0.8
1
1.2α3β3α1 α2 β2β1
6 9 14 17 20 23 27 30 33 36 39 42 46 49 53 56 59 63 66 69 72 75 79
N
C
β1β3
β2α2
α1
Figure 8
N
C
β1β3
β2
α2
α1
KH2 KH1
KH2
A
B
C
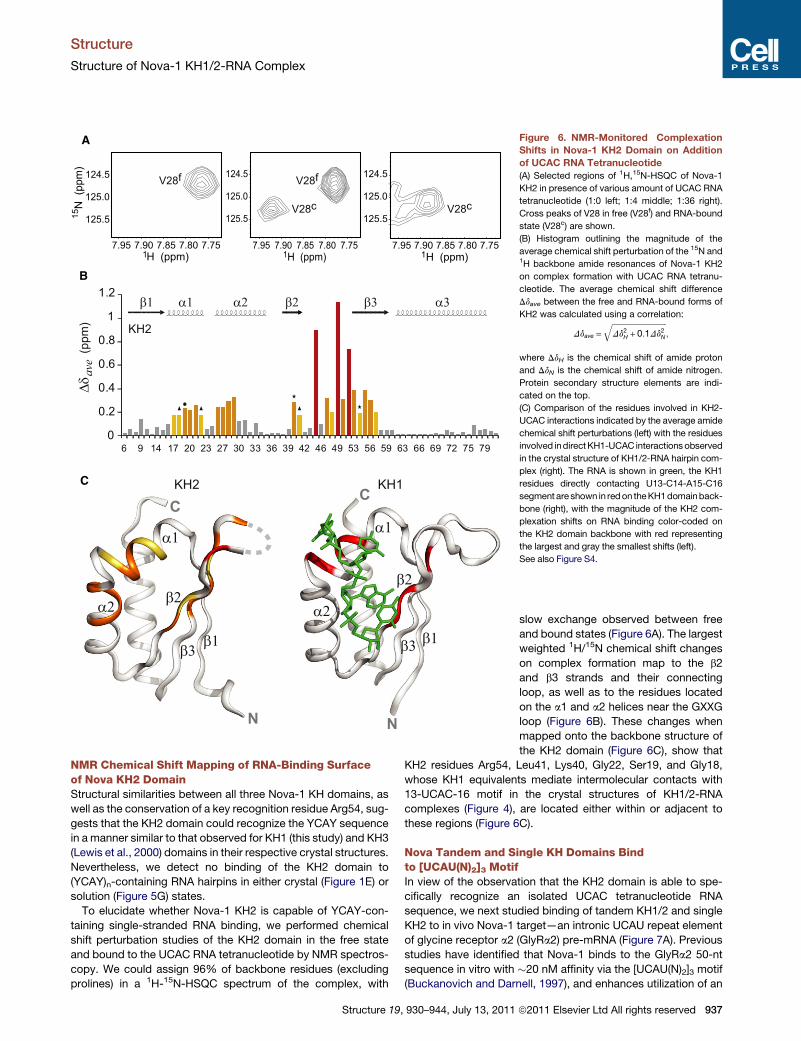
Figure 6. NMR-Monitored Complexation
Shifts in Nova-1 KH2 Domain on Addition
of UCAC RNA Tetranucleotide
(A) Selected regions of 1H,15N-HSQC of Nova-1
KH2 in presence of various amount of UCAC RNA
tetranucleotide (1:0 left; 1:4 middle; 1:36 right).
Cross peaks of V28 in free (V28f) and RNA-bound
state (V28c) are shown.
(B) Histogram outlining the magnitude of the
average chemical shift perturbation of the 15N and1H backbone amide resonances of Nova-1 KH2
on complex formation with UCAC RNA tetranu-
cleotide. The average chemical shift difference
Ddave between the free and RNA-bound forms of
KH2 was calculated using a correlation:
Ddave =ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiDd2H + 0:1Dd2N
q;
where DdH is the chemical shift of amide proton
and DdN is the chemical shift of amide nitrogen.
Protein secondary structure elements are indi-
cated on the top.
(C) Comparison of the residues involved in KH2-
UCAC interactions indicated by the average amide
chemical shift perturbations (left) with the residues
involved indirectKH1-UCAC interactionsobserved
in the crystal structure of KH1/2-RNA hairpin com-
plex (right). The RNA is shown in green, the KH1
residues directly contacting U13-C14-A15-C16
segment are shown in redon theKH1domainback-
bone (right), with the magnitude of the KH2 com-
plexation shifts on RNA binding color-coded on
the KH2 domain backbone with red representing
the largest and gray the smallest shifts (left).
See also Figure S4.
Structure
Structure of Nova-1 KH1/2-RNA Complex
NMR Chemical Shift Mapping of RNA-Binding Surfaceof Nova KH2 DomainStructural similarities between all three Nova-1 KH domains, as
well as the conservation of a key recognition residue Arg54, sug-
gests that the KH2 domain could recognize the YCAY sequence
in a manner similar to that observed for KH1 (this study) and KH3
(Lewis et al., 2000) domains in their respective crystal structures.
Nevertheless, we detect no binding of the KH2 domain to
(YCAY)n-containing RNA hairpins in either crystal (Figure 1E) or
solution (Figure 5G) states.
To elucidate whether Nova-1 KH2 is capable of YCAY-con-
taining single-stranded RNA binding, we performed chemical
shift perturbation studies of the KH2 domain in the free state
and bound to the UCAC RNA tetranucleotide by NMR spectros-
copy. We could assign 96% of backbone residues (excluding
prolines) in a 1H-15N-HSQC spectrum of the complex, with
Structure 19, 930–944, July 13, 2011
slow exchange observed between free
and bound states (Figure 6A). The largest
weighted 1H/15N chemical shift changes
on complex formation map to the b2
and b3 strands and their connecting
loop, as well as to the residues located
on the a1 and a2 helices near the GXXG
loop (Figure 6B). These changes when
mapped onto the backbone structure of
the KH2 domain (Figure 6C), show that
KH2 residues Arg54, Leu41, Lys40, Gly22, Ser19, and Gly18,
whose KH1 equivalents mediate intermolecular contacts with
13-UCAC-16 motif in the crystal structures of KH1/2-RNA
complexes (Figure 4), are located either within or adjacent to
these regions (Figure 6C).
Nova Tandem and Single KH Domains Bindto [UCAU(N)2]3 MotifIn view of the observation that the KH2 domain is able to spe-
cifically recognize an isolated UCAC tetranucleotide RNA
sequence, we next studied binding of tandem KH1/2 and single
KH2 to in vivo Nova-1 target—an intronic UCAU repeat element
of glycine receptor a2 (GlyRa2) pre-mRNA (Figure 7A). Previous
studies have identified that Nova-1 binds to the GlyRa2 50-nt
sequence in vitro with �20 nM affinity via the [UCAU(N)2]3 motif
(Buckanovich and Darnell, 1997), and enhances utilization of an
ª2011 Elsevier Ltd All rights reserved 937
Res
idua
ls
-0.040.000.04
41.0 42.0 43.0 44.0
-1.2
-0.8
-0.4
0.0
0.4
ln A
294
nm
Radius squared (cm2)
KH222 krpm17.1 kDa
GlyR 20
Complex
[KH1/2]:[GlyR20] [KH2]:[GlyR20]0 0.5 1 2 0 0.5 1 2 3 4 83
GlyR20 UCUCAUCAUCAUUUUCAUUU
0.0 0.5 1.0 1.5
-50
-40
-30
-20
-10
0
-0.4
-0.3
-0.2
-0.1
0.0
-10 0 10 20 30 40 50 60 70 80
Time (min)
µcal
/sec
kcal
/mol
e of
inje
ctan
t
0.0 0.5 1.0 1.5
-40
-30
-20
-10
0-0.4
-0.3
-0.2
-0.1
0.0
-10 0 10 20 30 40 50 60 70 80Time (min)
µcal
/sec
kcal
/mol
e of
inje
ctan
t
0.0 0.5 1.0 1.5 2.0-40
-30
-20
-10
0-0.5
-0.4
-0.3
-0.2
-0.1
0.0
0.1-10 0 10 20 30 40 50 60 70 80
Time (min)
µcal
/sec
[GlyR20]/[KH1/2] [GlyR20]/[KH2] [GlyR20]/[KH3]
kcal
/mol
e of
inje
ctan
t
n (sites)
0.89
0.43
0.38
Protein
KH1/2
KH2
KH3
RNA
GlyR20
GlyR20
GlyR20
0.13 0.01
0.30 0.02
0.78 0.02
KD (μM)
-38.5 0.1
-45.6 0.3
-49.5 0.5
ΔH (kcal mol-1)
-98
-125
-136
0.86KH1/2 RNA 0.59 0.04 -40.0 0.3 -106KH1/2
ΔS (cal K-1 mol-1)
A
B
D
G
E F
C
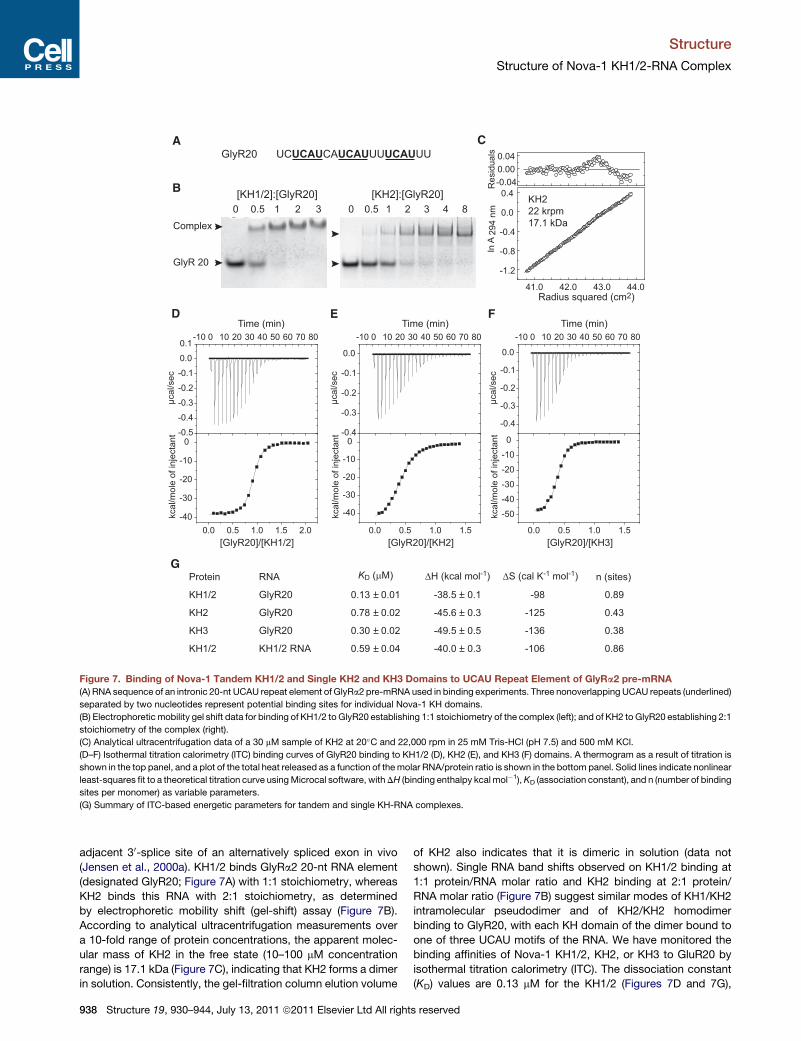
Figure 7. Binding of Nova-1 Tandem KH1/2 and Single KH2 and KH3 Domains to UCAU Repeat Element of GlyRa2 pre-mRNA
(A) RNA sequence of an intronic 20-nt UCAU repeat element of GlyRa2 pre-mRNA used in binding experiments. Three nonoverlapping UCAU repeats (underlined)
separated by two nucleotides represent potential binding sites for individual Nova-1 KH domains.
(B) Electrophoreticmobility gel shift data for binding of KH1/2 to GlyR20 establishing 1:1 stoichiometry of the complex (left); and of KH2 to GlyR20 establishing 2:1
stoichiometry of the complex (right).
(C) Analytical ultracentrifugation data of a 30 mM sample of KH2 at 20�C and 22,000 rpm in 25 mM Tris-HCl (pH 7.5) and 500 mM KCl.
(D–F) Isothermal titration calorimetry (ITC) binding curves of GlyR20 binding to KH1/2 (D), KH2 (E), and KH3 (F) domains. A thermogram as a result of titration is
shown in the top panel, and a plot of the total heat released as a function of themolar RNA/protein ratio is shown in the bottom panel. Solid lines indicate nonlinear
least-squares fit to a theoretical titration curve usingMicrocal software, withDH (binding enthalpy kcal mol�1),KD (association constant), and n (number of binding
sites per monomer) as variable parameters.
(G) Summary of ITC-based energetic parameters for tandem and single KH-RNA complexes.
Structure
Structure of Nova-1 KH1/2-RNA Complex
adjacent 30-splice site of an alternatively spliced exon in vivo
(Jensen et al., 2000a). KH1/2 binds GlyRa2 20-nt RNA element
(designated GlyR20; Figure 7A) with 1:1 stoichiometry, whereas
KH2 binds this RNA with 2:1 stoichiometry, as determined
by electrophoretic mobility shift (gel-shift) assay (Figure 7B).
According to analytical ultracentrifugation measurements over
a 10-fold range of protein concentrations, the apparent molec-
ular mass of KH2 in the free state (10–100 mM concentration
range) is 17.1 kDa (Figure 7C), indicating that KH2 forms a dimer
in solution. Consistently, the gel-filtration column elution volume
938 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All right
of KH2 also indicates that it is dimeric in solution (data not
shown). Single RNA band shifts observed on KH1/2 binding at
1:1 protein/RNA molar ratio and KH2 binding at 2:1 protein/
RNA molar ratio (Figure 7B) suggest similar modes of KH1/KH2
intramolecular pseudodimer and of KH2/KH2 homodimer
binding to GlyR20, with each KH domain of the dimer bound to
one of three UCAU motifs of the RNA. We have monitored the
binding affinities of Nova-1 KH1/2, KH2, or KH3 to GluR20 by
isothermal titration calorimetry (ITC). The dissociation constant
(KD) values are 0.13 mM for the KH1/2 (Figures 7D and 7G),
s reserved

Structure
Structure of Nova-1 KH1/2-RNA Complex
0.78 mM for the KH2 (Figures 7E and 7G) and 0.30 mM for the KH3
(Figures 7F and 7G) complexes with GlyR20. The n value (a
number of binding sites per protein monomer) estimated from
ITC curves using a single site binding model is 0.89 for the
KH1/2-RNA complex, whereas n = 0.43 for KH2-RNA complex,
consistent with 1:1 and 2:1 protein/RNA stoichiometries of these
respective complexes (Figures 7D, 7E, and 7G). The n value of
0.38 estimated from ITC curve (Figures 7F and 7G) for the
KH3-RNA complex indicates that stoichiometry of KH3-GlyR20
complex is either 2:1 or 3:1. These experiments demonstrate
that individual Nova KH2 and KH3 domains bind [UCAU(N)2]3motif of single stranded RNA with comparable (2.6-fold differ-
ence) binding affinity, while dual KH1/2 domain binds this ssRNA
with 6-fold higher affinity compared to a single KH2 domain.
We have also monitored binding of KH1/2 to GlyR20 RNA by
NMR spectroscopy. We observed significant chemical shift
perturbations between free and RNA-bound states (Figure S4A).
The number of KH1/2 resonances that experienced chemical
shift changes is at least twice the number observed for individual
KH2 (Figure 6B), suggesting that both KH2 and KH1 domains are
bound to GlyR20 in the complex. For example, in addition to
Gly22 and Gly25 of KH2 domain, there are two other cross-
peaks in the same region of the KH1/2 1H-15N-HSQC spectrum
that undergo significant chemical shift changes on RNA binding
(compare the selected regions of the spectra of KH2 and KH1/2
in Figures S4B and S4C, respectively).
DISCUSSION
KH domains, which constitute one of the most abundant RNA-
binding motifs (Messias and Sattler, 2004), are of particular
interest because of their role in regulating gene expression at
both transcriptional and translational levels, in alternate splicing,
and in the maturation of mRNA (Valverde et al., 2008). Eukary-
otic KH domains bind their mRNA targets with sequence spec-
ificity and affinities spanning the mM to nM range, and target tet-
ranucleotide sequences where the first and last bases are
pyrimidines, with structures of complexes available where the
third base is an adenine or cytosine. Our current understanding
of the principles underlying KH-RNA recognition emerged
following determination of the crystal structure of Nova-2 KH3
domain bound to a UCAC element within the hairpin segment
of a RNA stem-loop (Lewis et al., 2000), the NMR structure of
the STAR (signal transduction and activation of RNA) member
KH domain augmented by a QUA1 (quaker homology 1) domain
of SF1 (splicing factor 1) bound to a UAAC segment in a single-
stranded RNA context (Liu et al., 2001), the complex between
NusA, a key regulator of bacterial antitermination, and RNAs
derived from the antitermination region (Beuth et al., 2005),
and the crystal structures of KH1 domain of poly(C)-binding
protein 2 (PCBP2) bound to C-rich telomeric DNA and RNA
(Du et al., 2007). In each case, the RNA tetranucleotide is grip-
ped in a molecular vise composed of a hydrophobic a-helix/
b strand platform, an invariant GXXG motif and a variable
loop, thereby aligning the Watson-Crick edge of the adenine
at the third position for hydrogen-bonding with the peptide
backbone.
The current contribution reports the crystal structure of dual
eukaryotic KH domains of Nova-1 targeted to tandemRNA tetra-
Structure 19,
nucleotide repeats, where KH1 and KH2 domains form an intra-
molecular pseudodimer, with KH1 solely involved in RNA recog-
nition and KH2 involved in protein-protein contacts. These
results highlight the ability of KH domains to participate in
protein-RNA and protein-protein recognition and outline how
RNA-RNA buttressing interactions expand on our current under-
standing of KH-RNA tetranucleotide recognition events.
Comparison of Nova KH1/2 Interface with Other KH-KHInterfacesOur structure of the complex establishes that KH1 and KH2 have
extensive interdomain interactions, forming an intramolecular
pseudodimer. Two recently reported structures revealed a
similar arrangement of tandem KH domains in the RNA-free
state: the NMR structure of KH1/2 domains of human PCBP2
(Figure S5A) (Du et al., 2008), and the crystal structure of IMP1
mRNA-Binding Protein 1 (also called ZBP1) KH3/4 domains
(Figure S5B), whereas three other structures of tandem KH
domains in the free state revealed different modes of arrange-
ment of their respective KH domains (Figures S5C–S5E) (Diaz-
Moreno et al., 2010; Gopal et al., 2001; Valverde et al., 2007;
Worbs et al., 2001). A detailed comparison of the Nova KH1/2
interface with these KH-KH interfaces is given in the Supple-
mental Information.
Nova KH1/2-RNA Recognition Involving TandemYCAY RepeatsThe three KH domains of Nova proteins have been shown to
target [YCAY-(N)2]3 sites in natural a-2 glycine receptor subunit,
GABAA and Nova-1 pre-mRNAs (Buckanovich and Darnell,
1997; Dredge and Darnell, 2003; Dredge et al., 2005) and
[YCAY-(N)0-2]3 sites in RNA hairpin aptamers identified through
in vitro selection (Buckanovich and Darnell, 1997). Nevertheless,
it was not clear whether individual KH domains target one or
more tandem YCAY sites or whether one or more KH domains
target a single YCAY site. The structure of the Nova-1 KH1/2-
RNA hairpin complex reported in the present study addresses
this issue by revealing a novel recognition mode, where the
KH1 domain differentially recognizes adjacent tandem UCAG
and UCAC sites, mediated by specific buttressing interactions
between the two sites. The intermolecular interactions are pri-
marily between the KH1 domain and all four nucleotides of the
second UCAC tetranucleotide (U13–C16) site, with a few addi-
tional interactions with C10 and its phosphate of the first
UCAG tetranucleotide (U9–G12) site in the complex (Figure 4A).
Both UCAG and UCAC sites are involved in sculpting the RNA
scaffold due to the extensive base-sugar and base-phosphate
interactions associated with the mutual buttressing of A11 of
the first tetranucleotide repeat and C14 of the second tetranu-
cleotide repeat in the complex (Figure 4B).
The recognition elements between the KH1 domain and the
second UCAC tetranucleotide (U13–C16) site in the Nova-1
KH1/2-RNA hairpin complex reported in this study are very
similar to those reported previously between the KH3 domain
and the single UCAC site in the Nova-2 KH3-RNA hairpin com-
plex (Lewis et al., 2000). In both complexes, the sequential uracil,
cytosine and adenine residues of the UCAC segment are posi-
tioned atop an a/b platform and pinioned in place by interactions
with the invariant Gly-X-X-Gly motif and the variable loop. The
930–944, July 13, 2011 ª2011 Elsevier Ltd All rights reserved 939

K33 K33
K75
W38KH3
KH1
KH2 KH2
K23
K23
K45
K45
K43K43
R54
K40
K40
K23K23
K44K44
K29
Q43
Q40
K9
R54
K9
R54
W38
K67
K29
Q43Q40
K75
R54
K26
R75
K23
R52
K44
K43
R54E14
R38
K89
R83
Q40
R55
K5
R38
K23
R52
K44
K43
R54E14
K89R83
C3'5'
GCGCGGA
GCGCGAAC
UC
G CU
A
CCA
3'5'GAGGA
CUCCC
CC
A UG A
U
CAC
KH1/2RNA hairpin
KH3RNA hairpin
A B
C D
E F H
G
Figure 8. Two Perpendicular Views of the
Electrostatic Surface of Nova KH1 (with
Bound RNA), KH2, and KH3 (with Bound
RNA) Domains
(A and B) Two alternate views of KH1 domain of
KH1/2 complexed with RNA hairpin containing
tandem UCAN-sites within the loop segment.
(C and D) Two alternate views of KH2 domain of
KH1/2, that does not form a complex with RNA
hairpin.
(E and F) Two alternate views of KH3 domain
complexed with RNA hairpin containing single
UCAC site (Lewis et al., 2000; PDB entry 1EC6).
Blue and red patches are associated with posi-
tively- and negatively-charged KH surface seg-
ments. The RNA backbone is shown in a ribbon
and the bases in a slab representation. The slabs
are colored as follows: U, blue; C, magenta; A, red;
G, yellow.
(G) The sequence of the Nova-1 KH1/2-binding
25-nt RNA hairpin containing a (UCAN)2 site (Mu-
sunuru and Darnell, 2004) that was used in this
study.
(H) The sequence of the Nova-2 KH3-binding
20-nt RNA hairpin containing a single UCAN site
(Lewis et al., 2000).
Structure
Structure of Nova-1 KH1/2-RNA Complex
cytosine pairs with R54 usingWatson-Crick-like hydrogen bond-
ing alignments and the adenine pairs with the backbone using
Watson-Crick-like hydrogen bonding alignments (Figure 4B).
We highlight below a common intermolecular recognition
feature that has been observed in the eukaryotic Nova-2 KH3-
RNA hairpin (Lewis et al., 2000) and Nova-1 KH1/2-RNA hairpin
(this study) complexes, as well as in the splicing factor 1-intron
branch site RNA complex (Liu et al., 2001) and the bacterial
NusA KH1/2 complex with a GAACUCAAUAG RNA sequence
(Beuth et al., 2005). All these complexes have one key element
in common, namely that the main chain of the b2-strand forms
two hydrogen bonds with the Watson-Crick edge of the adenine
base (Figure 4B). A more detailed comparison of Nova KH1/2-
RNA hairpin and NusA KH1/2-ssRNA complexes is provided in
the Supplemental Experimental Procedures.
Comparison of RNA Hairpin Topologies in Nova KH1/2and KH3 ComplexesThe Nova KH1/2 and KH3 domains bind preferentially their re-
spective RNA hairpin aptamers, whereas they bind each other’s
aptamers significantly weaker (Figures 5F and 5H), implying that
not only the presence of primary UCAY recognition site but also
the RNA hairpin loop structure determine binding affinity and
specificity of individual Nova KH domains. The charge distribu-
tions are somewhat different on the canonical RNA-binding
940 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All rights reserved
surfaces of the three Nova KH domains
(Figure 8). The electrostatics of the
RNA-binding KH1 surface in the KH1/2-
RNA hairpin complex (Figures 8A and
8B) appears to be designed to favor
binding by a compact RNA loop (contains
nine bases), as reflected in a sharp turn
facilitated by the Lys45 side chain. The conformation of the
primary U13-C14-A15-C16 target site (Figure 4A, cyan) together
with the sharp turn results in the mutual buttressing of C14 by
A11 of the neighboring site (Figure 4B). At the same time, two
appropriately located positively charged residues Lys40 and
Lys43, which adopt finger-like conformations, recognize cyto-
sine bases C16 and C10, respectively (Figure 4A). This network
of intermolecular and buttressing interactions results in both
the primary UCAC target site and the CA step of the preceding
U9-C10-A11-G12 site participating in KH1 domain recognition
and stabilization.
The electrostatics of the RNA-binding KH3 surface in the
KH3-RNA hairpin complex (Lewis et al., 2000) appears to be
designed to favor a less-restrictive RNA loop (contains 10 bases)
topology, as reflected in no sharp turns in the loop trajectory (Fig-
ure 8E). Two important differences between the electrostatics of
the RNA-binding surfaces of KH1 (Figure 8A) and KH3 (Figure 8E)
are the presence of Glu14 and Lys44 in the latter complex. This
distribution of charged amino acids appears to contribute to
the separation of the 50- and 30-halves of the loop in the KH3
complex, such that the loop topology adopts a more gradual
turn trajectory (Figure 8E). The positively charged residues
Arg83 and Arg52 together with the negatively charged Glu14
side chain direct the RNA backbone toward the C terminus of
the KH3 domain (Figure 8E). No buttressing interactions are

Structure
Structure of Nova-1 KH1/2-RNA Complex
observed across the loop in the KH3-RNA hairpin complex, due
to the increased separation between the opposite sides of
the loop.
The importance of the electrostatics of the RNA interacting
surface of the Nova KH domains to recognition is reinforced
from the observed reduction in binding affinities associated
with the KH1/2-RNA hairpin complex for the KH1 Ser14Glu
single mutant and the KH1 Lys40,43Gln dual mutant (Figure 5E).
In the single mutant, neutral serine in KH1 is replaced by nega-
tively-charged glutamate observed at this position in KH3,
whereas in the dual mutant, positively-charged lysines observed
at these positions in KH1 are replaced by neutral glutamines
observed at these positions in KH2.
The KH2 domain does not participate in RNA recognition in the
Nova-1 KH1/2-RNA hairpin complex (Figure 1E), and it does not
appear to bind either UCAG-UCAC- or UCAC-containing RNA
hairpin aptamers with high affinity and binds very weakly to a
RNA hairpin containing (YCAY-N)3 sites (Figure 5G). The charge
distributions are somewhat different on the canonical RNA-
binding face of the KH2 domain (Figures 8C and 8D), due to
Gln40 and Gln43 replacing their lysine counterparts in the KH1
domain (Figures 8A and 8B). We attempt to explain why the
canonical RNA-binding surface of the KH1 domain as part of
a KH1/2 construct (Figure 5D, black circles), but not the KH2
domain (Figure 5D, red circles), targets the Nova-1 KH1/2-RNA
hairpin. We put forward two potential contributors to such
discriminative recognition following a detailed structural analysis
of protein-RNA contacts in the complex, as outlined in the
Supplemental Experimental Procedures.
All Nova KH Domains Bind UCAY RepeatSingle-Stranded RNAWe demonstrated that all three Nova-1 KH domains bind
[UCAU(N)2]3 repeats of a single-stranded RNA, with comparable
binding affinities between individual KH2 and KH3 domains (Fig-
ure 7). NMR chemical shift perturbation studies (Figures 6A and
6B) establish that Nova KH2 binds the UCAC RNA tetranucleo-
tide in slow exchange, a feature characteristic of a tightly bound
complex in solution. These results on a single-stranded RNA that
spans the KH domain binding pocket contrasts with no measur-
able binding when the same tetranucleotide element, or its
tandem repeats, is part of the loop segment of RNA hairpins (Fig-
ure 5G). The chemical shift changes in the KH2 domain on UCAC
binding are mainly distributed within the nucleic acid-binding
channel (Figure 6C), and their magnitude is in the range reported
for Nova KH3 binding to ssRNA pentanucleotide sequences
containing UCAN (N is either pyrimidine or purine) elements
(Beuth et al., 2007).
The ability of KH2 to recognize UCAC motif is fully expected
based on structural similarity of its RNA-binding surface to that
of the other two Nova KH domains, as well as the conservation
of a key recognition residue Arg54. A 2.6-fold lower binding
affinity of KH2 than KH3 for [UCAU(N)2]3 repeat ssRNA is consis-
tent with less positively charged RNA binding surface of KH2
domain compared to that of KH3 domain (Figures 8C–8F). We
have established that KH2 domain in an isolated state forms
a dimer in solution. Because hydrophobic interactions dominate
the KH1/KH2 interface of the dual KH1/2 domain, the KH2
domain in the absence of KH1 binding partner likely forms the
Structure 19,
KH2/KH2 homodimer through the same hydrophobic b1/a3
surface. Such very similar dimeric interfaces have been previ-
ously observed between KH1 domains of PCBP2 in the crystal
structures of its complexes with poly(C) nucleic acids (Du
et al., 2007) and between the tandem KH1 and KH2 domains
of the protein (Du et al., 2008) (Figure S5A). Therefore, it is
possible that KH2/KH2 homodimer and KH1/KH2 intramolecular
pseudodimer bind RNA in a similar way, such that each KH
domain of a dimer bind the two terminal UCAU motifs of
GlyR20, consistent with 1:1 (RNA/dimer) stoichiometry deter-
mined for each complex. The NMR chemical shift perturbations
observed for the intramolecular KH1/2 pseudodimer on binding
GlyR20 (Figure S4) are also consistent with the involvement of
both KH domains in the interaction with the RNA. This model is
stereochemically possible as the 8 nucleotide intervening
sequence is of sufficient length to connect the two terminal
UCAU motifs simultaneously bound by the two KH domains of
a dimer.
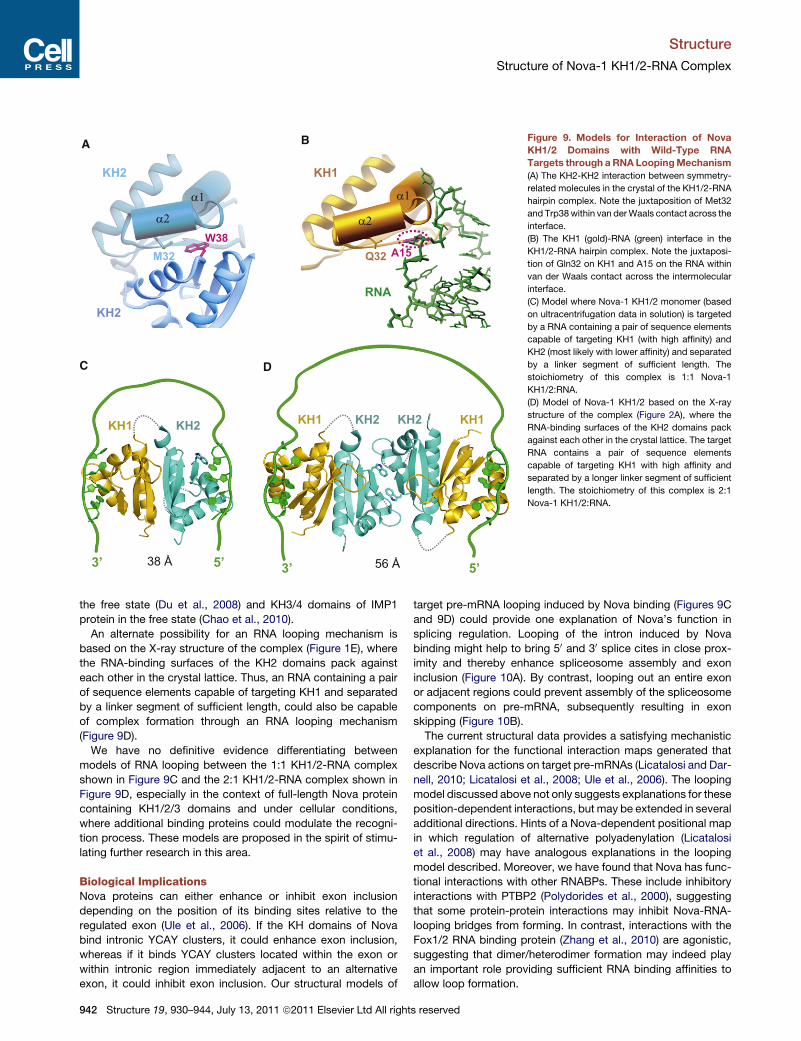
Nova KH Domain Surface Mimicry Associatedwith Protein and RNA RecognitionThe same canonical RNA-binding surface of the KH domain
participates in protein-protein interactions in the crystal (KH2
domain) (Figure 9A, top left) or in RNA recognition (KH1 domain)
(Figure 9B, top left) in the Nova-1 KH1/2-RNA hairpin complex.
A competing molecular recognition duality has also been re-
ported for the Y14 protein, whose canonical RNA-binding
surface, is also involved in protein-protein interactions with
Mago, with both Y14 and Mago being components of the exon
junction complex (Fribourg et al., 2003; Lau et al., 2003; Shi
and Xu, 2003).
Equally striking is the extension of the structural mimicry to an
element of the interacting partner. Thus, the indole ring of a Trp
residue mediates the protein-protein contacts involving KH2
domains in the crystal (Figure 9A), whereas the purine ring of
the key adenine base occupies the same interfacial position in
the KH1-RNA interaction (Figure 9B) in the Nova-1 KH1/2-RNA
hairpin complex.
Alternate Models Involving RNA Looping on the Surfaceof Nova-1 KH1/2Our ultracentrifugation studies imply that Nova-1 KH1/2 forms
a monomer in the free state (Figure S3A) and when bound to
its RNA aptamer target (Figure S3B) in dilute solution. The KH1
and KH2 domains interact with each other through an extensive
interface (Figures 3A and 3C), thereby positioning their canonical
RNA-binding surfaces at opposite ends of the molecule and
accessible for recognition. Thus, an RNA containing a pair of
sequence elements capable of targeting KH1 (with high affinity)
and KH2 (most likely with lower affinity) and separated by a linker
segment of sufficient length, should be capable of complex
formation through a RNA looping mechanism (Figure 9C), as
initially proposed for splicing regulation from the structure of
tandem RNA-binding domains RBD3/4 of polypyrimidine tract-
binding protein bound to RNA (Oberstrass et al., 2005). An
RNA looping mechanism has also been proposed for RNA
recognition based on the structure of tandem zinc-finger
domains of Muscleblind-like protein bound to RNA (Teplova
and Patel, 2008), KH1/2 domains of polyC-binding protein in
930–944, July 13, 2011 ª2011 Elsevier Ltd All rights reserved 941
38 Å 5’3’ 56 Å 5’3’
KH2
α2
α1
KH2
W38M32
α2
α1
Q32 A15
KH1
RNA
KH1 KH2 KH1 KH2 KH1KH2
A B
C D
Figure 9. Models for Interaction of Nova
KH1/2 Domains with Wild-Type RNA
Targets through aRNA LoopingMechanism
(A) The KH2-KH2 interaction between symmetry-
related molecules in the crystal of the KH1/2-RNA
hairpin complex. Note the juxtaposition of Met32
and Trp38within van derWaals contact across the
interface.
(B) The KH1 (gold)-RNA (green) interface in the
KH1/2-RNA hairpin complex. Note the juxtaposi-
tion of Gln32 on KH1 and A15 on the RNA within
van der Waals contact across the intermolecular
interface.
(C) Model where Nova-1 KH1/2 monomer (based
on ultracentrifugation data in solution) is targeted
by a RNA containing a pair of sequence elements
capable of targeting KH1 (with high affinity) and
KH2 (most likely with lower affinity) and separated
by a linker segment of sufficient length. The
stoichiometry of this complex is 1:1 Nova-1
KH1/2:RNA.
(D) Model of Nova-1 KH1/2 based on the X-ray
structure of the complex (Figure 2A), where the
RNA-binding surfaces of the KH2 domains pack
against each other in the crystal lattice. The target
RNA contains a pair of sequence elements
capable of targeting KH1 with high affinity and
separated by a longer linker segment of sufficient
length. The stoichiometry of this complex is 2:1
Nova-1 KH1/2:RNA.
Structure
Structure of Nova-1 KH1/2-RNA Complex
the free state (Du et al., 2008) and KH3/4 domains of IMP1
protein in the free state (Chao et al., 2010).
An alternate possibility for an RNA looping mechanism is
based on the X-ray structure of the complex (Figure 1E), where
the RNA-binding surfaces of the KH2 domains pack against
each other in the crystal lattice. Thus, an RNA containing a pair
of sequence elements capable of targeting KH1 and separated
by a linker segment of sufficient length, could also be capable
of complex formation through an RNA looping mechanism
(Figure 9D).
We have no definitive evidence differentiating between
models of RNA looping between the 1:1 KH1/2-RNA complex
shown in Figure 9C and the 2:1 KH1/2-RNA complex shown in
Figure 9D, especially in the context of full-length Nova protein
containing KH1/2/3 domains and under cellular conditions,
where additional binding proteins could modulate the recogni-
tion process. These models are proposed in the spirit of stimu-
lating further research in this area.
Biological ImplicationsNova proteins can either enhance or inhibit exon inclusion
depending on the position of its binding sites relative to the
regulated exon (Ule et al., 2006). If the KH domains of Nova
bind intronic YCAY clusters, it could enhance exon inclusion,
whereas if it binds YCAY clusters located within the exon or
within intronic region immediately adjacent to an alternative
exon, it could inhibit exon inclusion. Our structural models of
942 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All right
target pre-mRNA looping induced by Nova binding (Figures 9C
and 9D) could provide one explanation of Nova’s function in
splicing regulation. Looping of the intron induced by Nova
binding might help to bring 50 and 30 splice cites in close prox-
imity and thereby enhance spliceosome assembly and exon
inclusion (Figure 10A). By contrast, looping out an entire exon
or adjacent regions could prevent assembly of the spliceosome
components on pre-mRNA, subsequently resulting in exon
skipping (Figure 10B).
The current structural data provides a satisfying mechanistic
explanation for the functional interaction maps generated that
describe Nova actions on target pre-mRNAs (Licatalosi and Dar-
nell, 2010; Licatalosi et al., 2008; Ule et al., 2006). The looping
model discussed above not only suggests explanations for these
position-dependent interactions, but may be extended in several
additional directions. Hints of a Nova-dependent positional map
in which regulation of alternative polyadenylation (Licatalosi
et al., 2008) may have analogous explanations in the looping
model described. Moreover, we have found that Nova has func-
tional interactions with other RNABPs. These include inhibitory
interactions with PTBP2 (Polydorides et al., 2000), suggesting
that some protein-protein interactions may inhibit Nova-RNA-
looping bridges from forming. In contrast, interactions with the
Fox1/2 RNA binding protein (Zhang et al., 2010) are agonistic,
suggesting that dimer/heterodimer formation may indeed play
an important role providing sufficient RNA binding affinities to
allow loop formation.
s reserved

splicing enhancem ent
U1U2
U2U1
splicing repression
1 2
3
E x o n
1 2
3
5 ’
5 ’
3 ’
3 ’
U1U2
A
B
Figure 10. Models of RNA Looping Induced by Nova KH1-3 Domains
Binding to Splicing Enhancers and Splicing Silencers Predicted by
Nova RNA Map of Splicing Regulation
(A) Model based on the examples of Nova upregulation of exon inclusion by
binding to intronic splicing enhancer elements (red circles) to enhance
spliceosome assembly.
(B) Model based on the examples of Nova inhibition of exon inclusion by
binding to exonic splicing silencing element (blue circle) and blocking U1
snRNP (U1) assembly on the pre-mRNA, and Nova inhibition of exon inclusion
by binding to intronic splicing silencing element immediately upstream of
alternative exon by blocking recognition of the 30 splice site by U2 snRNP (U2).
KH1, KH2, and KH3 are colored gold, cyan, and purple, respectively. The
linkers between KH1 and KH2, and between KH2 and KH3 are represented by
the dotted gray lines. The green line represents RNA loop.
Structure
Structure of Nova-1 KH1/2-RNA Complex
EXPERIMENTAL PROCEDURES
Detailed procedures for protein and RNA preparation, ITC measurements,
crystallization and data collection, structure determination and refinement,
NMR sample preparation, NMR spectroscopy and chemical shift assign-
ments, NMR relaxation measurements, gel electrophoretic mobility shift
binding assays, and filter binding assays are listed in the Supplemental Exper-
imental Procedures.
ACCESSION NUMBERS
Atomic coordinates and structure factors for the type I and type II Nova-1 KH1/
KH2-RNA hairpin complexes have been deposited in the Protein Data Bank
(www.rutgers.edu/pub) under accession codes 2ANN and 2ANR, respectively.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures
and five figures and can be found with this article online at doi:10.1016/j.str.
2011.05.002.
Structure 19,
ACKNOWLEDGMENTS
D.J.P. received support from NIH grant CA49982, J.C.D. from NIH grant
HD40647, R.B.D. from NIH grant NS34389, and K.M., S.K.B., and R.B.D.
from NIH grant NS40955 and the Howard Hughes Medical Institute. K.M.
received support from the Weill Cornell/Rockefeller/Sloan-Kettering Tri-Insti-
tutional MD-PhD program and NIH MSTP grant GM07739.
Received: February 1, 2011
Revised: April 29, 2011
Accepted: May 2, 2011
Published: July 12, 2011
REFERENCES
Albert, M.L., and Darnell, R.B. (2004). Paraneoplastic neurological degenera-
tions: keys to tumour immunity. Nat. Rev. Cancer 4, 36–44.
Beuth, B., Pennell, S., Arnvig, K.B., Martin, S.R., and Taylor, I.A. (2005).
Structure of a Mycobacterium tuberculosis NusA-RNA complex. EMBO J.
24, 3576–3587.
Beuth, B., Garcia-Mayoral, M.F., Taylor, I.A., and Ramos, A. (2007). Scaffold-
independent analysis of RNA-protein interactions: the Nova-1 KH3-RNA
complex. J. Am. Chem. Soc. 129, 10205–10210.
Buckanovich, R.J., and Darnell, R.B. (1997). The neuronal RNA binding protein
Nova-1 recognizes specific RNA targets in vitro and in vivo. Mol. Cell. Biol. 17,
3194–3201.
Burd, C.G., and Dreyfuss, G. (1994). Conserved structures and diversity of
functions of RNA-binding proteins. Science 265, 615–621.
Chao, J.A., Patskovsky, Y., Patel, V., Levy, M., Almo, S.C., and Singer, R.H.
(2010). ZBP1 recognition of beta-actin zipcode induces RNA looping. Genes
Dev. 24, 148–158.
Diaz-Moreno, I., Hollingworth, D., Kelly, G., Martin, S., Garcia-Mayoral, M.,
Briata, P., Gherzi, R., and Ramos, A. (2010). Orientation of the central domains
of KSRP and its implications for the interaction with the RNA targets. Nucleic
Acids Res. 38, 5193–5205.
Dredge, B.K., and Darnell, R.B. (2003). Nova regulates GABA(A) receptor
gamma2 alternative splicing via a distal downstream UCAU-rich intronic
splicing enhancer. Mol. Cell. Biol. 23, 4687–4700.
Dredge, B.K., Polydorides, A.D., and Darnell, R.B. (2001). The splice of life:
alternative splicing and neurological disease. Nat. Rev. Neurosci. 2, 43–50.
Dredge, B.K., Stefani, G., Engelhard, C.C., and Darnell, R.B. (2005). Nova
autoregulation reveals dual functions in neuronal splicing. EMBO J. 24,
1608–1620.
Du, Z., Lee, J.K., Fenn, S., Tjhen, R., Stroud, R.M., and James, T.L. (2007).
X-ray crystallographic and NMR studies of protein-protein and protein-nucleic
acid interactions involving the KH domains from human poly(C)-binding
protein-2. RNA 13, 1043–1051.
Du, Z., Fenn, S., Tjhen, R., and James, T.L. (2008). Structure of a construct of
a human poly(C)-binding protein containing the first and second KH domains
reveals insights into its regulatory mechanisms. J. Biol. Chem. 283, 28757–
28766.
Fernandez-Recio, J., Totrov, M., Skorodumov, C., and Abagyan, R. (2005).
Optimal docking area: a new method for predicting protein-protein interaction
sites. Proteins 58, 134–143.
Fribourg, S., Gatfield, D., Izaurralde, E., and Conti, E. (2003). A novel mode
of RBD-protein recognition in the Y14-Mago complex. Nat. Struct. Biol. 10,
433–439.
Gopal, B., Haire, L.F., Gamblin, S.J., Dodson, E.J., Lane, A.N.,
Papavinasasundaram, K.G., Colston, M.J., and Dodson, G. (2001). Crystal
structure of the transcription elongation/anti-termination factor NusA from
Mycobacterium tuberculosis at 1.7 A resolution. J. Mol. Biol. 314, 1087–1095.
Grishin, N.V. (2001). KH domain: one motif, two folds. Nucleic Acids Res. 29,
638–643.
930–944, July 13, 2011 ª2011 Elsevier Ltd All rights reserved 943

Structure
Structure of Nova-1 KH1/2-RNA Complex
Jensen, K.B., Dredge, B.K., Stefani, G., Zhong, R., Buckanovich, R.J., Okano,
H.J., Yang, Y.Y., and Darnell, R.B. (2000a). Nova-1 regulates neuron-specific
alternative splicing and is essential for neuronal viability. Neuron 25, 359–371.
Jensen, K.B., Musunuru, K., Lewis, H.A., Burley, S.K., and Darnell, R.B.
(2000b). The tetranucleotide UCAY directs the specific recognition of RNA
by the Nova K-homology 3 domain. Proc. Natl. Acad. Sci. USA 97, 5740–5745.
Lau, C.K., Diem, M.D., Dreyfuss, G., and Van Duyne, G.D. (2003). Structure of
the Y14-Magoh core of the exon junction complex. Curr. Biol. 13, 933–941.
Lewis, H.A., Chen, H., Edo, C., Buckanovich, R.J., Yang, Y.Y., Musunuru, K.,
Zhong, R., Darnell, R.B., and Burley, S.K. (1999). Crystal structures of Nova-
1 and Nova-2 K-homology RNA-binding domains. Structure 7, 191–203.
Lewis, H.A., Musunuru, K., Jensen, K.B., Edo, C., Chen, H., Darnell, R.B., and
Burley, S.K. (2000). Sequence-specific RNA binding by a Nova KH domain:
implications for paraneoplastic disease and the fragile X syndrome. Cell 100,
323–332.
Licatalosi, D.D., and Darnell, R.B. (2006). Splicing regulation in neurologic
disease. Neuron 52, 93–101.
Licatalosi, D.D., and Darnell, R.B. (2010). RNA processing and its regulation:
global insights into biological networks. Nat. Rev. Genet. 11, 75–87.
Licatalosi, D.D., Mele, A., Fak, J.J., Ule, J., Kayikci, M., Chi, S.W., Clark, T.A.,
Schweitzer, A.C., Blume, J.E., Wang, X., et al. (2008). HITS-CLIP yields
genome-wide insights into brain alternative RNA processing. Nature 456,
464–469.
Liu, Z., Luyten, I., Bottomley, M.J., Messias, A.C., Houngninou-Molango, S.,
Sprangers, R., Zanier, K., Kramer, A., and Sattler, M. (2001). Structural basis
for recognition of the intron branch site RNA by splicing factor 1. Science
294, 1098–1102.
Messias, A.C., and Sattler, M. (2004). Structural basis of single-stranded RNA
recognition. Acc. Chem. Res. 37, 279–287.
Musunuru, K., and Darnell, R.B. (2001). Paraneoplastic neurologic disease
antigens: RNA-binding proteins and signaling proteins in neuronal degenera-
tion. Annu. Rev. Neurosci. 24, 239–262.
Musunuru, K., and Darnell, R.B. (2004). Determination and augmentation of
RNA sequence specificity of the Nova K-homology domains. Nucleic Acids
Res. 32, 4852–4861.
Oberstrass, F.C., Auweter, S.D., Erat, M., Hargous, Y., Henning, A.,Wenter, P.,
Reymond, L., Amir-Ahmady, B., Pitsch, S., Black, D.L., and Allain, F.H. (2005).
944 Structure 19, 930–944, July 13, 2011 ª2011 Elsevier Ltd All right
Structure of PTB bound to RNA: specific binding and implications for splicing
regulation. Science 309, 2054–2057.
Ostareck-Lederer, A., Ostareck, D.H., and Hentze, M.W. (1998). Cytoplasmic
regulatory functions of the KH-domain proteins hnRNPs K and E1/E2. Trends
Biochem. Sci. 23, 409–411.
Polydorides, A.D., Okano, H.J., Yang, Y.Y., Stefani, G., and Darnell, R.B.
(2000). A brain-enriched polypyrimidine tract-binding protein antagonizes
the ability of Nova to regulate neuron-specific alternative splicing. Proc. Natl.
Acad. Sci. USA 97, 6350–6355.
Shi, H., and Xu, R.M. (2003). Crystal structure of the Drosophila Mago nashi-
Y14 complex. Genes Dev. 17, 971–976.
Siomi, H., Matunis, M.J., Michael, W.M., and Dreyfuss, G. (1993). The pre-
mRNA binding K protein contains a novel evolutionarily conserved motif.
Nucleic Acids Res. 21, 1193–1198.
Teplova, M., and Patel, D.J. (2008). Structural insights into RNA recognition by
the alternative-splicing regulator muscleblind-like MBNL1. Nat. Struct. Mol.
Biol. 15, 1343–1351.
Ule, J., Stefani, G., Mele, A., Ruggiu, M., Wang, X., Taneri, B., Gaasterland, T.,
Blencowe, B.J., andDarnell, R.B. (2006). An RNAmap predicting Nova-depen-
dent splicing regulation. Nature 444, 580–586.
Valverde, R., Pozdnyakova, I., Kajander, T., Venkatraman, J., and Regan, L.
(2007). Fragile X mental retardation syndrome: structure of the KH1-KH2
domains of fragile X mental retardation protein. Structure 15, 1090–1098.
Valverde, R., Edwards, L., and Regan, L. (2008). Structure and function of KH
domains. FEBS J. 275, 2712–2726.
Worbs, M., Bourenkov, G.P., Bartunik, H.D., Huber, R., andWahl, M.C. (2001).
An extended RNA binding surface through arrayed S1 and KH domains in
transcription factor NusA. Mol. Cell 7, 1177–1189.
Yang, Y.Y., Yin, G.L., and Darnell, R.B. (1998). The neuronal RNA-binding
protein Nova-2 is implicated as the autoantigen targeted in POMA patients
with dementia. Proc. Natl. Acad. Sci. USA 95, 13254–13259.
Zhang, C., Frias, M.A., Mele, A., Ruggiu, M., Eom, T., Marney, C.B., Wang, H.,
Licatalosi, D.D., Fak, J.J., and Darnell, R.B. (2010). Integrative modeling
defines the Nova splicing-regulatory network and its combinatorial controls.
Science 329, 439–443.
s reserved























