Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 89, pp. 1968-1972, March 1992Cell Biology
Proliferation of totipotent hematopoietic stem cells in vitro withretention of long-term competitive in vivo reconstituting ability
(bone marrow transplantation/self-renewal/retroviral gene transfer)
CHRISTOPHER C. FRASER, STEPHEN J. SZILVASSY, CONNIE J. EAVES, AND R. KEITH HUMPHRIES*Terry Fox Laboratory, British Columbia Cancer Agency, and the Departments of Medical Genetics and Medicine, Universityof British Columbia, Vancouver, BC, Canada V5Z 1L3
Communicated by Eugene P. Cronkite, December 4, 1991
ABSTRACT Marrow cells from male mice pretreated with5-fluorouracil were infected with helper-free neomycin-resistant (neor) recombinant retrovirus and then used to initiatelong-term cultures (LTC) on irradiated adherent marrowfeeder layers. Four weeks later LTC cells were harvested andinjected into lethally irradiated female recipients either aloneor together with 2 x 105 female marrow cells with selectivelycompromised long-term repopulating potential to assay fortotipotent and competitive repopulating units (CRU), respec-tively. A total of 46 unique clones were detected in recipients 5wk to 7 mo after transplant. Half of these clones (22 of 46)included both lymphoid and myeloid progeny. Eight of the 22lympho-myeloid clones were represented in multiple recipi-ents, in some cases after ijfection of limiting numbers of CRU,thus indicating repopulation from sibling totipotent stem cellsgenerated during the initial 4-wk period in LTC. Serial analysisof cells released into the nonadherent fraction of LTC for up to7 wk provided additional evidence of the continuing prolifer-ation in LTC of totipotent stem cells with long-term repopu-lating potential. The frequency of CRU determined fromlimiting-dilution analyses ofLTC-derived cells was the same forrecipients analyzed at 5 wk or 7 mo after transplantation andwas also the same whether marrow or thymus repopulation wasassessed. These assays showed that concurrent with the expan-sion of some totipotent cells revealed by retroviral marking,there was a slow but net 6.5-fold decrease in total CRUnumbers after 4 wk in LTC. These results show the capacity ofsome totipotent hematopoietic stem cells to be maintained andamplified over extensive time periods in vitro without diminu-tion of their long-term in vivo repopulating potential. Theseresults also set the stage for analogous studies of human stemcell selection and expansion in vitro, which may be importantfor future gene therapy protocols.
The regenerative capacity and life-long maintenance of thehematopoietic system depends on a primitive subpopulationof stem cells with extensive self-renewal, proliferative, anddifferentiation potential. Totipotent hematopoietic stem cellsthat can clonally repopulate lymphoid and myeloid tissues inmyelo-ablated recipients have been documented in bothmouse and human by several experimental strategies. Thesestrategies include the use of radiation-induced chromosomalmarkers (1), naturally occurring genetic markers (2, 3), andmarkers introduced by retroviral insertion (4-10). Althoughsuch cells may represent the most primitive hematopoieticpopulation in adult marrow, considerable evidence points toa hierarchy of transplantable hematopoietic cells with differ-ent potentials for sustaining hematopoiesis in vivo (11-16).Cells with long-term hematopoietic reconstituting ability canbe distinguished by a number of physical and biologicalproperties from cells that generate mature progeny in short-
term in vivo or in vitro clonogenic assays (17-22). Timecourse studies of the pattern of hematopoietic reconstitutionin recipients of retrovirally marked cells have revealed fre-quent examples oflong-term reconstitution by small numbersof totipotent stem cells under conditions where multiple,fluctuating short-lived clones are seen for up to 4 mo aftertransplantation (23, 24). These results are consistent with ahierarchical structure in the stem cell populations trans-planted, although they also do not exclude that environmen-tal or stochastic mechanisms may lead to heterogeneity in thebehavior exhibited by biologically similar primitive hemato-poietic cells.Methods for maintaining, expanding, and following the fate
of the most primitive hematopoietic cells in vitro representkey requirements for investigating these questions. A systemfor the long-term culture (LTC) of mouse bone marrowintroduced by Dexter et al. (25) has been shown to maintaincells capable of lymphoid and myeloid reconstitution inirradiated recipients (26), although a diminution in colony-forming unit-spleen (CFU-S) self-renewal capacity (27) andcompetitive erythroid repopulating ability (28) of LTC-derived cells compared with fresh marrow cells has beenreported. In this study we have combined the use of aquantitative assay for competitive repopulating units (CRU)with retroviral marking of the initial cell population to dem-onstrate the ability of totipotent hematopoietic cells withlong-term (7 mo) repopulating ability to undergo extensiveamplification as their overall numbers decline over a periodof 4 wk in the LTC system.
MATERIALS AND METHODSVirus. Recombinant Tkneol9 virus at a titer of 1 x 106 per
ml was generated from a T-2 producer line maintained in 10%calf serum/Dulbecco's modified Eagle's medium (26). Ab-sence of helper virus in Tkneol9 producer cells or infectedLTC (see below) was verified by failure to serially transfervirus conferring G418 resistance to NIH 3T3 cells (26).
Retroviral Infection and LTC. Marrow cells from adultmale (C57BL/6J x C3H/HeJ) F1 (B6C3F1) mice injected i.v.4 days earlier with 5-fluorouracil (5-FU, 150 mg/kg of bodyweight) were infected with Tkneol9 virus using a supernatantinfection protocol in which 3-5 x 106 marrow cells werecultured for 24 hr in 100-mm Petri dishes in 10 ml of virussupernatant containing Polybrene at 4 jug/ml, 5% pokeweedmitogen-stimulated spleen cell-conditioned medium, and10%o agar-stimulated human leukocyte-conditioned mediumas described (26, 29). Cells were then recovered by gentleagitation and scraping of dishes with a rubber policeman,
Abbreviations: 5-FU, 5-fluorouracil; CRU, competitive repopulatingcells; LTC, long-term cultures; IL-3, interleukin 3.*To whom reprint requests should be addressed at: Terry FoxLaboratory, British Columbia Cancer Research Centre, 601 West10th Avenue, Vancouver, BC, Canada V5Z 1L3.
1968
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 89 (1992) 1969
washed in 2% fetal calf serum/a modification of minimalessential Eagle medium (Sigma) and resuspended in LTCmedium [a medium/10% horse serum/10% fetal calf serum/10-6 M sodium hydrocortisone hemisuccinate/10-4 M 2-mer-captoethanol]. Aliquots of 3 x 106 cells were overlaid onpreviously established 3-wk-old long-term B6C3F1 femalemarrow adherent layers that had been irradiated with 15 Gy[250 kilovolt peak (kVp) x-rays] to inactivate persistinghematopoietic cells (26). LTC were maintained by weeklyremoval of half of the medium and nonadherent cells andrestoration of the volume with fresh LTC medium. To assaycells in LTC for repopulating cells, adherent layers wereremoved with a rubber policeman, suspended by passagethrough a 21-gauge needle and then combined with thenonadherent cells. Cells were washed once in 2% fetal calfserum/a-minimal essential medium, and aliquots from indi-vidual culture flasks were then injected i.v. into irradiated (8Gy, 250 kVp x-rays) female recipients. In some cases, 2 x 105female B6C3F1 marrow cells that had been previously sub-jected to two cycles of transplantation and regeneration werealso injected to allow quantitative measurements of CRU inthe retrovirally marked test cells to be obtained (29, 30). Forcultures that were used to assess recovery of repopulatingcells in the nonadherent fraction over time, all of the mediumand nonadherent cells were removed weekly and replacedeither with fresh medium alone or with LTC medium con-taining 25 units per ml of recombinant mouse interleukin-3(IL-3) (Biogen). The nonadherent cells removed after 3, 5, 6,and 7 weeks of culture were then injected into irradiatedrecipients.
Hematopoietic Tissue Analysis. Recipients were sacrificedeither 5 wk or between 5 and 7 mo after transplantation ofcultured cells. DNA was routinely extracted from marrow,spleen, and thymus. In some cases, DNA was also extractedfrom lymph nodes and from various subpopulations of mar-row and spleen. Highly enriched (>90%) mast cell popula-tions were generated by culturing marrow or spleen cells for3 wk in WEHI-3B-supplemented medium (as a source ofIL-3), and highly enriched (>95% Mac-i-positive) macro-phage populations were generated by culturing marrow orspleen cells for 48 hr in medium supplemented with 10%WEHI-3B-conditioned medium and then for 7-10 days inmedium supplemented with 35% human leukocyte-condi-tioned medium.
Highly enriched (>90% Thy-i-positive) T-cell populationswere obtained by elution of nonadherent cells after loading ofa half-spleen equivalent onto a 3-ml nylon wool column andincubating it for 1 hr at 37°C (31). Nylon wool adherent cellswere then released by gentle agitation for 2-3 min in phos-phate-buffered saline/10 mM EDTA and a highly enriched(>90% B220-positive) population of B lymphocytes subse-quently isolated from this fraction by panning for 1 hr at 37°Cin dishes precoated with unpurified rabbit anti-mouse immu-noglobulin (<108 cells per dish) and selective removal of theadherent cells (32).
Southern Blot Analysis. High-molecular-weight DNA wasdigested with HindIII or EcoRI, which cuts once in theproviral genome and releases a fragment unique to theintegration site. Ten micrograms of digested DNA (5 ,g formale control lanes) was electrophoresed and analyzed bySouthern blotting (33) as described (29) with a 32P-labeledprobe specific for the neor gene sequence in the provirus fromplasmid pMC1 (34). HindIII-digested blots were stripped andreprobed with a Y chromosome-specific fragment from plas-mid pY2 (20, 29, 35).
RESULTS
In Vitro Recovery of Long-Term Repopulating Cells. Todetermine whether totipotent cells capable of long-term re-
population could be demonstrated in 4-wk-old LTC initiatedwith retrovirally infected 5-FU cells, aliquots were injectedinto irradiated recipients. These mice were then sacrificed 5mo later and assessed for the presence of common clones ofLTC-derived cells in various myeloid and lymphoid cellpopulations. Results from two representative recipients areshown in Fig. 1 A and B. Fig. 1A shows the presence of adominant clone in both myeloid (unseparated marrow, mar-row-derived mast cells, and marrow- or spleen-derived mac-rophages) and lymphoid cell populations (isolated splenic Tand B cells, as well as thymus and lymph node cells).Comparison of the intensity of the provirus band with thatseen in a clonally derived 3T3 cell line generated afterinfection with the same virus indicates that the retrovirallymarked stem cell clone in this mouse was contributing to atleast 80% of total hematopoiesis at sacrifice. In the othermouse shown (Fig. 1B) at least two marked clones werepresent in all hematopoietic cell populations examined.Again, intensities of the bands of the major clone indicates adominant contribution to reconstitution in each.
Quantitation of CRU in LTC. To quantitate changes in thetotal number of repopulating cells present after 4 wk in LTC,we used the CRU assay (29, 30). Data from three experimentsare plotted in Fig. 2, and the derived CRU values are shownin Table 1 together with published values for CRU frequen-cies in freshly isolated day-4 5-FU marrow cells (30). Thenumber of CRU in 4-wk-old LTC is the same whether thecalculations are based on marrow or thymus repopulation, asshown previously for fresh or partially purified marrow cells(30). When combined, an overall decrease in CRU frequency
A
- -
L'7~~~0- ;: 6. -
V
B
~~ -
-'a t -a 7 - "
= r $ -. =at Lr :,- z S
S
0
FIG. 1. Presence of specific retroviral insertion fragments in bonemarrow (BM), spleen (Spl), separately isolated macrophage (0) andmast cell populations (mast), separated splenic T and B lymphocytepopulations (Spl B, Spl T), thymus (Thy), and lymph node (LN)tissues of two mice (A and B) detected by Southern blot analysis ofHindIll-digested DNA. Membranes were hybridized to a neor-specific probe. Lethally irradiated mice received 1-2 x 106 cells fromretrovirally infected 4-wk-old LTC (i.e., approximately one-tenth ofa culture initiated from 3 x 106 day4 5-FU marrow cells) and wereanalyzed 5 mo after transplant. Lane M is DNA from the viralproducer cell line Tkneol9l-2 containing multiple proviral integra-tions. All lanes contain 5-10 jig of DNA.
Cell Biology: Fraser et al.
-1
- :7. -
o",

Proc. Natl. Acad. Sci. USA 89 (1992)
10000 20000 30000
Initial Cells per LTC
FIG. 2. Proportion of mice (four to eight animals per group)negative for reconstitution (<5%) of bone marrow (Left) or thymus(Right) with male cells 5 wk after transplantation with variousproportions of a 4-wk-old LTC initiated with day-4 5-FU cells,together with 2 x 105 compromised female marrow cells. Theproportion of reconstitution by male cells was determined by South-ern blot analysis ofmarrow and thymus cell DNA from each recipientwith the Y-specific probe pY2. Circles, squares, and trianglesrepresent results from analyses of three separate experiments. Astraight-line fit to the combined data based on maximum-likelihoodanalysis is shown by the solid line. The broken line representspublished (30) data for similar analyses of fresh day-4 5-FU marrowcells.
after 4 wk in LTC of 6.5-fold is apparent (1/2000 in day-45-FU marrow vs. 1/13,000 in 4-wk-old LTC based on pooleddata for bone marrow and thymus). In two experiments, CRUfrequencies in the 4-wk-old LTC were also determined inadditional recipients given aliquots of the same cells butsacrificed 7 mo after transplant. Because fewer recipientswere available for evaluation at this later time, the derivedCRU frequencies are less precise. Nevertheless, the CRUvalues from 7-mo tissue repopulation data were again notsignificantly different whether marrow or thymus was ana-lyzed and were also not significantly different from the valuesobtained from mice analyzed only 5 wk after transplant(Table 1).Evidence of Totipotent Stem Cell Amplification in Vitro.
Because all LTC in these studies were established withretrovirally infected 5-FU marrow cells and the contents of
Table 1. Frequency of CRU in 4-wk-old long-term bonemarrow cultures
Hemopoietic tissueanalyzed for LTC-derived hemopoiesis*
Bone marrowPosttransplan- and thymustation time Bone marrow Thymus (pooled data)
5 wk 12,000 14,500 13,000(8,800-18,000) (8,900-23,600) (9,600-17,800)
7 mo 25,000 21,000 23,400(14,000-45,400) (10,200-43,100) (14,800-37,000)
Cells from three LTC experiments were analyzed in recipientssacrificed 5 wk after transplant (20-28 mice per experiment). For twoof these experiments, CRU measurements were also derived fromassessment of recipients sacrificed 7 mo after transplant (16-22 miceper experiment).*Values are the reciprocal of CRU frequency expressed relative tonumber of cells used to initiate the LTC 4 wk earlier. Confidencelimits of 95% defined by +2 SEM are shown in parentheses inmiddle two columns. Reciprocal CRU frequencies in fresh day-45-FU marrow have been previously determined to be 2700 (1300-5700) and 1300 (540-3100) from bone marrow or thymus repopula-tion analyzed separately, or 2000 (1200-3500) when both read-outsare combined (30).
individual cultures were then assayed in multiple recipients,the number and distribution of marked clones in differentanimals could be determined and, in some cases, the patternsobserved could be related to the number of CRU injected.Table 2 summarizes the characteristics of all marked clonesregenerated in recipients of cells from eight individual 4-wkLTC individually assayed in multiple recipients. Of a total of46 specifically identifiable clones, nine were seen to contrib-ute to repopulation of two or more mice, and eight of theseclones included clones that repopulated both myeloid andlymphoid tissues (amplified clones I-IX, Table 2). A further37 clones were identified in only one recipient, but 14 oftheseclones showed contributions to both myeloid and lymphoidtissues (single clones, Table 2).
Fig. 3 shows four examples of a common clone contribut-ing to the hematopoietic reconstitution of multiple recipientsof cells from a single flask, indicating amplification of totipo-tent stem cells in the original retrovirus-infected input pop-ulation. For simplicity we will refer to such clones as siblingstem cell clones because they are derived from a singleparental precursor.
Fig. 3A shows an experiment in which reconstitution oflymphoid and/or myeloid tissues by sibling clones (clone VIIin Table 2) was detected in two CRU assay mice sacrificed 5wk after transplant (mouse 1/150A and mouse 1/iSOB) aswell as in two of six recipients injected with cells from thesame LTC (1/20th of a flask), but without additional com-promised cells, and sacrificed 7 mo after transplant (mouse1/20A and mouse 1/20B). Mouse 1/iSOB, which was com-petitively repopulated after injection of limiting numbers ofCRU (two per recipient) demonstrates repopulation of mar-row, spleen, and thymus by a common stem cell. In mouse1/150A and in mouse 1/20A, cells with the same clonalmarker were detectable only in the marrow and spleen. Inmouse 1/20B, the sibling clone detected appeared restrictedto the spleen and thymus.
Further evidence of expansion of totipotent stem cells inLTC is provided by the recipient data shown in Fig. 3B. Here,analysis oftissues from two mice that had been injected undercompetitive conditions with approximately two CRU fromthe same flask revealed progeny of sibling clones (clone V inTable 2) in the marrow, spleen, and thymus at both 5 wk and7 mo after transplant. These results also extend the CRUquantitation studies indicating detection of the same type oftotipotent cell at either 5 wk or 7 mo after transplant when theCRU assay is used.
In recipients of cells from some LTC, analysis of retroviralinsertion fragments revealed marked sibling clones that
Table 2. Tissue distribution of marked clones
Repopulation pattern, no. of mice
Marrow and Spleen and Marrow andspleen* Spleen thymust thymust
Single clones 11 8 4 14Amplified clonesClone I 1 1Clone II 1 1Clone III 2Clone IV 1 1Clone V 1 3Clone VI 1 1Clone VII 2 3Clone VIII 3Clone IX 3 1
Total 17 13 6 27
*Apparently myeloid-restricted population.tApparently lymphoid-restricted repopulation.tMyeloid and lymphoid repopulation.
0a.(4
0
Z 0.374)
0C.0a.
0.1
Initial Cells per LTC
1970 Cell Biology: Fraser et al.
Proc. Natl. Acad. Sci. USA 89 (1992) 1971
5 week 7 month
1/150 A 1/150 B 1/20 A 1/20BI is S T' 'B S T' IB S T' 'B STF
"-4,t ,_
neo
.~~~~~~.
B 5 -leekI/ 150(
l s 'l',l
7 monhli
I /5II0
I't sTi II
aU S8*if
IA 0
C 1/45 A 1/450 B
I 1 s.1'1 l
its,.r 7
neo
w
wV..
so_ Y-s
y
D A It
.lI _s
n _o
_ _
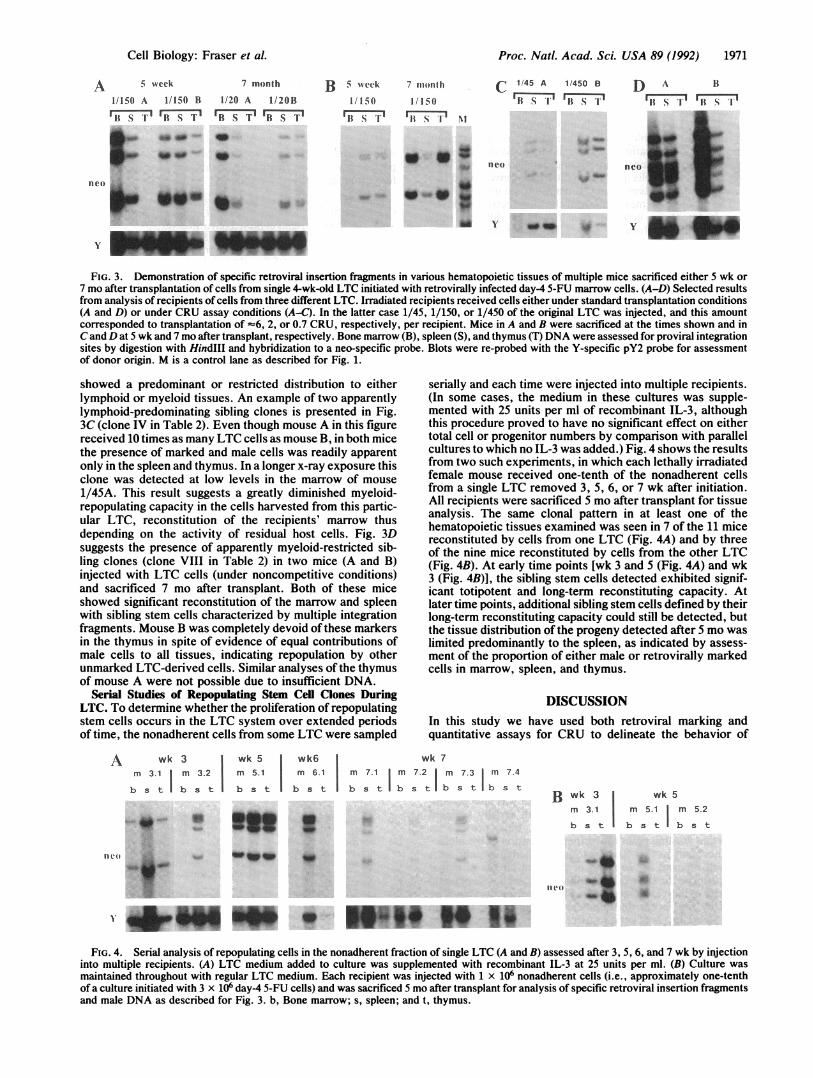
FIG. 3. Demonstration of specific retroviral insertion fragments in various hematopoietic tissues of multiple mice sacrificed either 5 wk or7 mo after transplantation of cells from single 4-wk-old LTC initiated with retrovirally infected day-4 5-FU marrow cells. (A-D) Selected resultsfrom analysis of recipients of cells from three different LTC. Irradiated recipients received cells either under standard transplantation conditions(A and D) or under CRU assay conditions (A-C). In the latter case 1/45, 1/150, or 1/450 of the original LTC was injected, and this amountcorresponded to transplantation of -6, 2, or 0.7 CRU, respectively, per recipient. Mice in A and B were sacrificed at the times shown and inC and D at 5 wk and 7 mo after transplant, respectively. Bone marrow (B), spleen (S), and thymus (T) DNA were assessed for proviral integrationsites by digestion with HindIII and hybridization to a neo-specific probe. Blots were re-probed with the Y-specific pY2 probe for assessmentof donor origin. M is a control lane as described for Fig. 1.
showed a predominant or restricted distribution to eitherlymphoid or myeloid tissues. An example of two apparentlylymphoid-predominating sibling clones is presented in Fig.3C (clone IV in Table 2). Even though mouse A in this figurereceived 10 times as many LTC cells as mouse B, in both micethe presence of marked and male cells was readily apparentonly in the spleen and thymus. In a longer x-ray exposure thisclone was detected at low levels in the marrow of mouse1/45A. This result suggests a greatly diminished myeloid-repopulating capacity in the cells harvested from this partic-ular LTC, reconstitution of the recipients' marrow thusdepending on the activity of residual host cells. Fig. 3Dsuggests the presence of apparently myeloid-restricted sib-ling clones (clone VIII in Table 2) in two mice (A and B)injected with LTC cells (under noncompetitive conditions)and sacrificed 7 mo after transplant. Both of these miceshowed significant reconstitution of the marrow and spleenwith sibling stem cells characterized by multiple integrationfragments. Mouse B was completely devoid of these markersin the thymus in spite of evidence of equal contributions ofmale cells to all tissues, indicating repopulation by otherunmarked LTC-derived cells. Similar analyses of the thymusof mouse A were not possible due to insufficient DNA.
Serial Studies of Repopulating Stem Cell Clones DuringLTC. To determine whether the proliferation of repopulatingstem cells occurs in the LTC system over extended periodsof time, the nonadherent cells from some LTC were sampled
serially and each time were injected into multiple recipients.(In some cases, the medium in these cultures was supple-mented with 25 units per ml of recombinant IL-3, althoughthis procedure proved to have no significant effect on eithertotal cell or progenitor numbers by comparison with parallelcultures to which no IL-3 was added.) Fig. 4 shows the resultsfrom two such experiments, in which each lethally irradiatedfemale mouse received one-tenth of the nonadherent cellsfrom a single LTC removed 3, 5, 6, or 7 wk after initiation.All recipients were sacrificed 5 mo after transplant for tissueanalysis. The same clonal pattern in at least one of thehematopoietic tissues examined was seen in 7 of the 11 micereconstituted by cells from one LTC (Fig. 4A) and by threeof the nine mice reconstituted by cells from the other LTC(Fig. 4B). At early time points [wk 3 and 5 (Fig. 4A) and wk3 (Fig. 4B)], the sibling stem cells detected exhibited signif-icant totipotent and long-term reconstituting capacity. Atlater time points, additional sibling stem cells defined by theirlong-term reconstituting capacity could still be detected, butthe tissue distribution of the progeny detected after 5 mo waslimited predominantly to the spleen, as indicated by assess-ment of the proportion of either male or retrovirally markedcells in marrow, spleen, and thymus.
DISCUSSIONIn this study we have used both retroviral marking andquantitative assays for CRU to delineate the behavior of
A wk 3I3.1 m 3.2
b st b s t
wk 5m 5.1
b s t
wk6
m 6.1
b s t
wk 7
m 7.1 m 7.2 m 7.3 m 7.4
b s t b s t b st b s t
4'.." C *a a
0- - I
I|'< ~ - w___qqop 4
" .0
Y_ _s
i _ _ sA
B wk 3
m 3.1
bast
wk 5
m 5.1 m 5.2
b s t b s t
FIG. 4. Serial analysis of repopulating cells in the nonadherent fraction of single LTC (A and B) assessed after 3, 5, 6, and 7 wk by injectioninto multiple recipients. (A) LTC medium added to culture was supplemented with recombinant IL-3 at 25 units per ml. (B) Culture was
maintained throughout with regular LTC medium. Each recipient was injected with 1 x 106 nonadherent cells (i.e., approximately one-tenthof a culture initiated with 3 x 106 day-4 5-FU cells) and was sacrificed 5 mo after transplant for analysis of specific retroviral insertion fragmentsand male DNA as described for Fig. 3. b, Bone marrow; s, spleen; and t, thymus.
A
Cell Biology: Fraser et al.
Inrlwlw

Proc. Natl. Acad. Sci. USA 89 (1992)
totipotent hematopoietic stem cells under conditions of LTC.Analysis of mice injected with cells from 4-wk-old culturesrevealed the consistent presence of marked progeny in re-constituted tissues for 7 mo. A high proportion of all clonesdetected (50O, Table 2) were represented in both lymphoidand myeloid populations at analysis. In some instances thisresult occurred even when limiting numbers of CRU wereinjected (-two cells per recipient), thus providing directevidence of totipotent stem cell maintenance in LTC.The repeated demonstration of the same retroviral inser-
tion fragments in hematopoietic tissues of different miceinjected with cells from the same LTC further showed that atleast some of the totipotent cells in day-4 5-FU marrowundergo clonal expansion in vitro with preservation of boththeir long-term and competitive lymphoid- and myeloid-repopulating ability. The fact that such marked sibling clonescould be detected in CRU assay recipients of -two CRUassessed either 5 wk or 7 mo after transplant is consistentwith previous evidence that CRU in day-4 5-FU marrow canbe readily infected with the supernatant procedure used andthat most, if not all, CRU are totipotent cells capable ofsustaining hematopoiesis for at least 7 mo. This hypothesis isfurther supported by the finding of the same CRU frequencyregardless of time of recipient assessment from 5 wk to 7 moafter transplant and regardless of whether the marrow orthymus was used to assess repopulating potential. Thus, thetransplant conditions embodied in the CRU assay allow stemcells with long-term repopulating potential to be accuratelyquantitated as early as 5 wk after transplant. Unlike previousstudies in which severe combined immunodeficiency (scid)mice were used as recipients for stem cell quantitation (36),we did not find enrichment in LTC of repopulating stem cellswith lymphoid-restricted differentiation potential. Neither,however, did we observe a consistent decline in the numberof repopulating stem cells active at 5 wk and 7 mo aftertransplant as described (24). One explanation for both ofthese differences is that the number and genotype of cellspresent during the initial period of hematologic recovery mayinfluence both type of progeny produced and rapidity withwhich they appear. Evidence in support of this possibilitycomes from the different rates of appearance of +/+ cells indeficient lineages after their transplantation into W/W' mice(37). In addition, allophenic mice created from two differinggenotypes show a consistently unequal contribution of eachgenotype to hematopoiesis (10).
Quantitation of CRU numbers after 4 wk in LTC revealeda slow decline to -15% of the input value. Because a highproportion of those present after 4 wk could be shown torepresent clonal derivatives of initially marked CRU, thebehavior of individual CRU in LTC may be very heteroge-neous: some undergo extensive amplification even in the faceof concurrent mechanisms leading to a net loss of CRU,presumably due to their differentiation and/or death. Evi-dence for the continuous turnover and self-renewal of totipo-tent stem cells in LTC over several weeks was also demon-strated by analysis of mice injected with serially samplednonadherent LTC cells. These results, thus, provide animportant starting point for further delineation of the molec-ular mechanisms that support totipotent hematopoietic stemcell self-renewal in vitro and of potential effects of clonalexpansion on the subsequent proliferative and developmentalcapacity of individual stem cells. Such experiments should beaided by the recent development of methods for attainingeven higher levels of gene transfer efficiency to repopulatingcells (38).The present studies also have implications for clinical bone
marrow transplantation. Persistence in LTC of very primitivehuman hematopoietic cells with long-term in vitro repopu-lating ability has been shown to be analogous to the kineticsof CRU maintenance in murine LTC described here (39), and
it is not unreasonable to assume the operation in human LTCof similarly competing mechanisms of stem cell proliferationand decline. It should, therefore, also be possible by appro-priate manipulation to optimize conditions that favor expan-sion of transplantable human hematopoietic stem cells invitro, particularly where selection of clones over prolongedperiods may be therapeutically advantageous-for example,for certain gene therapy protocols.
This work was supported by the National Cancer Institute ofCanada (NCIC) and Medical Research Council of Canada. C.C.F.and S.J.S. were recipients of Steve Fonyo Studentships ofthe NCIC.C.J.E. is a Terry Fox Cancer Research Scientist of the NCIC. Theexpert technical assistance ofP. Rosten and the secretarial assistanceof K. Windham are gratefully acknowledged.1. Abramson, S., Miller, R. G. & Phillips, R. A. (1977) J. Exp. Med. 145,
1567-1579.2. Mintz, B., Anthony, K. & Litwin, S. (1984) Proc. Natl. Acad. Sci. USA
81, 7835-7839.3. Turhan, A. G., Humphries, R. K., Phillips, G. L., Eaves, A. C. &
Eaves, C. J. (1989) N. Engl. J. Med. 320, 1655-1661.4. Keller, G., Paige, C., Gilboa, E. & Wagner, E. F. (1985) Nature (London)
318, 149-154.5. Dick, J. E., Magli, M. C., Huszar, D., Phillips, R. A. & Bernstein, A.
(1985) Cell 42, 71-79.6. Lemischka, 1. R., Raulet, D. H. & Mulligan, R. C. (1986) Cell 45,
917-927.7. Snodgrass, R. & Keller, G. (1987) EMBO J. 6, 3955-3960.8. Capel, B., Hawley, R., Covarrubias, L., Hawley, T. & Mintz, B. (1989)
Proc. Natl. Acad. Sci. USA 86, 4564-4568.9. Capel, B., Hawley, R. G. & Mintz, B. (1990) Blood 75, 2267-2270.
10. Van Zant, G., Chen, J.-J. & Scott-Micus, K. (1991) Blood 77, 756-763.11. Hellman, S., Botnick, L. E., Hannon, E. C. & Vigneulle, R. M. (1978)
Proc. Natl. Acad. Sci. USA 75, 490-494.12. Cudkowicz, G., Upton, A. C. & Shearer, G. M. (1964) Nature (London)
201, 165-167.13. Ross, E. A. M., Anderson, N. & Micklem, H. S. (1982) J. Exp. Med.
155, 432-444.14. Jones, R. J., Celano, P., Sharkis, S. J. & Sensenbrenner, L. L. (1989)
Blood 73, 397-401.15. Mauch, P. & Hellman, S. (1989) Blood 74, 872-875.16. Harrison, D. E., Stone, M. & Astle, C. M. (1990) J. Exp. Med. 172,
431-437.17. Hodgson, G. S. & Bradley, T. R. (1979) Nature (London) 281, 381-382.18. Visser, J. W. M., Bauman, J. G. J., Mulder, A. H., Eliason, J. F. & de
Leeuw, A. M. (1984) J. Exp. Med. 59, 1576-1590.19. Spangrude, G. J., Heimfeld, S. & Weissman, I. L. (1988) Science 241,
58-62.20. Szilvassy, S. J., Lansdorp, P. M., Humphries, R. K., Eaves, A. C. &
Eaves, C. J. (1989) Blood 74, 930-939.21. Ploemacher, R. E. & Brons, R. H. C. (1989) Exp. Hematol. 17,263-266.22. Jones, R. J., Wagner, J. E., Celano, P., Zicha, M. S. & Sharkis, S. J.
(1990) Nature (London) 347, 188-189.23. Keller, G. & Snodgrass, R. (1990) J. Exp. Med. 171, 1407-1418.24. Jordan, C. T. & Lemischka, I. R. (1990) Genes Dev. 4, 220-232.25. Dexter, T. M., Allen, T. D. & Lajtha, L. G. (1977) J. Cell Physiol. 91,
335-344.26. Fraser, C. C., Eaves, C. J., Szilvassy, S. J. & Humphries, R. K. (1990)
Blood 76, 1071-1076.27. Mauch, P., Greenberger, J. S., Botnik, L., Hannon, E. & Hellman, S.
(1980) Proc. Natl. Acad. Sci. USA 77, 2927-2930.28. Harrison, D. E., Lerner, C. P. & Spooncer, E. (1987) Blood 69, 1021-
1025.29. Szilvassy, S. J., Fraser, C. C., Eaves, C. J., Lansdorp, P. M., Eaves,
A. C. & Humphries, R. K. (1989) Proc. Natl. Acad. Sci. USA 86,8798-8802.
30. Szilvassy, S. J., Humphries, R. K., Lansdorp, P. M., Eaves, A. C. &Eaves, C. J. (1990) Proc. Natl. Acad. Sci. USA 87, 8736-8740.
31. Dougherty, G. J., Allen, C. A. & Hogg, N. M. (1986) in Handbook ofExperimental Immunology: Applications of Immunological Methods inBiomedical Sciences, eds. Weir, D. M., Herzenberg, L. A., Blackwell,C. & Herzenberg, L. A. (Blackwell Scientific, Oxford), Vol. 4, pp.125.1-125.12.
32. Chan, P.-Y. & Takei, F. (1989) J. Immunol. 142, 1727-1736.33. Southern, E. M. (1975) J. Mol. Biol. 98, 503-517.34. Thomas, K. R. & Capecchi, M. R. (1987) Cell 51, 503-512.35. Lamar, E. E. & Palmer, E. (1984) Cell 37, 171-177.36. Fulop, G. M. & Phillips, R. A. (1989) Blood 74, 1537-1544.37. Barker, J. E., Braun, J. & McFarland-Starr, E. C. (1988) Proc. Natl.
Acad. Sci. USA 85, 7332-7335.38. Bodine, D. M., Karlsson, S. & Nienhuis, A. W. (1989) Proc. Nail. Acad.
Sci. USA 86, 8897-8901.39. Sutherland, H. J., Eaves, C. J., Lansdorp, P. M., Thacker, J. D. &
Hogge, D. E. (1991) Blood 78, 666-672.
1972 Cell Biology: Fraser et al.





























