Embed Size (px)
Citation preview

Enzymologie : aspects fondamentaux, passé et présent
Pr Michèle Reboud-Ravaux
UMR 8256 - Institut de Biologie Paris-Seine
Université Pierre et Marie Curie (UPMC)
Cours Franco-Québécois d’Enzymologie AvancéeEnzymologie Moléculaire & Mécanistique
UE 5EV107 (UPMC)
& BMC 6225 (Université de Montréal)
23 septembre 2016
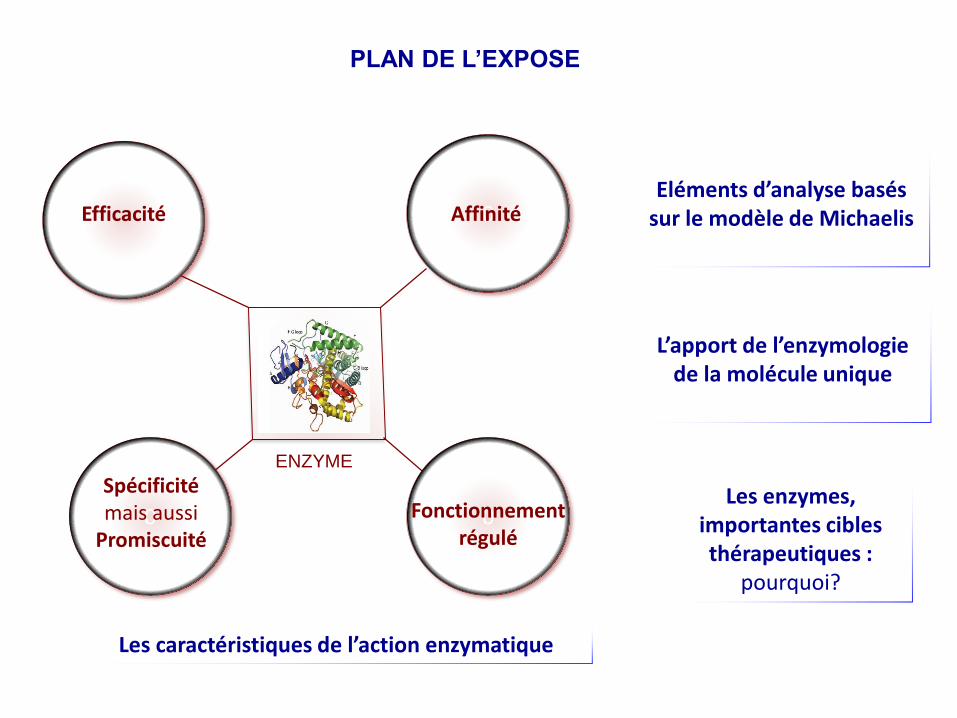
PLAN DE L’EXPOSE
Efficacité
ENZYME
Eléments d’analyse basés sur le modèle de Michaelis
Les enzymes, importantes cibles thérapeutiques :
pourquoi?
Les caractéristiques de l’action enzymatique
L’apport de l’enzymologie de la molécule unique
0Affinité
0Fonctionnement régulé
0
Spécificité mais aussi
Promiscuité

Pourquoi les enzymes sont-elles des cibles thérapeutiques ?
Enzymes 47%
Protéines G couplées aux récepteurs 30%
Canaux ioniques 7%
Autres récepteurs 2%
Récepteurs nucléaires 2%
Divers 1%
Intégrines 1%
ADN 1%
Cibles biochimiques des médicaments(adapté de Hopkins et Groom, 2002, Nature Rev. Drug Discov., 1, 727).
Les enzymes demeurent des cibles essentielles
Représentent toujours 50-75% des efforts recherche pour
découvrir de nouveaux médicaments.

Aspirine
Rivatigmine BortezomibOseltamivir
CaptoprilLovastatine
Fluorouracile
Acyclovir
Méthotrexate
Raltegravir
Tipranavir
AZT (zidovudine)

Attractiveness of enzymes as drug targets
- Essentiality of their activity.
- By their nature (proteins) and catalytic mechanism, are
highly amenable to inhibition by small molecules :
enzymes are primary targets.
- Related enzymes to primary targets must be studied to
avoid unwanted side effects.
Example: HIV protease inhibitors/ human aspartic proteases
(renin, pepsin, cathepsins D and E).
- Enzymes are implicated in drug metabolism and
pharmacokinetics that drive metabolic transformations.
Critical for the drug discovery and development process:
. a good understanding of the structure and mechanisms of action of enzymes
. quantitative studies of enzyme-ligand interactions.
Adapted from R. A. Copeland. Evaluation of Enzyme Inhibitors in Drug Discovery (2013)
- Involved in prodrug conversion to active
species.
- Enzymes form the basis of a number of
diagnostic tests that are in current clinical use.
- Some enzymes are used as therapeutic agents:
rt-PA (recombinant tissue plasminogen activator),
ingestion of pancreatic enzymes.
- Genes that encode specific enzymes are also
potential therapeutic agents for enzyme-based
therapies.

The chosen microstate is stabilized leading to a shift in the distribution of microstates towards the
deeper potential well (i.e. lower potential energy) of the preferred microstate.
Its degree of stabilization is directly related to what is called the ‘affinity’ of the ligand for this state.
The structure of the free enzyme and enzyme ligand-complexes is dynamic:
a manifold conformational substates, or microstates, that are interconvertible.
Po
ten
tia
l e
nerg
y
Microstate
Enzyme Complex ES
Microstate
Enzymes are dynamic entities
(conformational fluctuations)
Po
ten
tia
l e
nerg
y
Unique opportunity
for interaction
A ligand binds specifically to
a specific microstate
(best complementarity
between pocket and ligand)
Conformational fluctuations can be exploited for drug interactions:
large molecular diversity for the ligands of an enzyme
*

The Henri-Michaelis-Menten equation
See Deichman et al. 2014, FEBS J, 281, 426
v Vmax [S]
[S]KM
kcat[E][S]
[S]KM
Victor Henri
Leonor MichaelisCornish-Bowden 2014, FEBS J, 587, 2715
Maud Leonora Menten
Citations to Michaelis and Menten’s paper of 1913
. Henri V (1902) Théorie générale de l’action de quelques diastases, C. R. Hebd. Séances Acad Sci. 153, 916-919;
(1903) Lois générales de l’action des diastases, Herman.
. Michaelis L & Menten M. L. (1913) Kinetik der Invertinwirkung, Biochem. Z. 49, 333-369.

Characterization of enzymatic reactions
Inhibition
6679
Activation
195
After Mc Donal & Tipton 2014, FEBS J, 281, 583
Michaelis-
Menten
18594
Kinetic data statistics in SABIO-RK
Inhibition
6,679
Activation
195
Michaelis-
Menten
18,594
Hill cooperativity
1,702
Number of parameters Distribution of kinetic law types
(number of database entries)
Km
34,485
kcat
17,611
Vmax
13,237
kcat/Km
13,237
Ki
6,775
Modèle
michaélien

Efficacité : kcat & kcat/KM
Acétylcholinestérase : kcat = 25.000 s-1
2
+
Choline
AchE
Ach
AchE + Acétylcholine (Ach)
Acide acétique
E + S ES E-S’
P1
P2
Non covalent Covalent
E + P2
P1
kcat = VM/[E]0 (s-1)Anhydrase carbonique : kcat = 1.00.000 s-1
kcat (temps-1) = activité moléculaire spécifique ou nombre d’actes catalytiques/unité de
temps/molécule d’enzyme ou ‘turnover number’
k1
k-1k2
k3
kcat k2k3
k2 k3
kcat/KM( temps-1. M-1) = efficacité ‘vraie’ ou efficacité dynamique
Relié à KM
Acetyl-enzyme
E + S ES E+Pk2k1
k-1
kcat = k2

[S]0 < < KM [S]0 = KM [S]0 >> KM
[E]0 << [S]0
[S]2 > [S]1[S]1[S]3 > [S]2
Enzyme saturée
vi = Vmax/2 vi = Vmax
Affinité
1/KM = AFFINITE APPARENTE de l’enzyme pour son substrat
Plus KM est faible, plus l’enzyme aura ‘de l’affinité’ pour son substrat

Le modèle de Michaelis : sens physique et schéma cinétique
Equation de Henri-Michaelis-Menten
[P]
Temps (min)
[S]0
v
vi1
vi2
vi3
vi4
vi5 = vi
6 = Vmax
[E]0 << [S]0
Etat stationnaire pour les formes enzymatiques
E + S ES E+Pk2k1
k-1
0
0.2
0.4
0.6
0.8
1
1.2
0 5 10 15 20 25
vi
vi1
vi2
vi3
vi4
vi5 vi
6
[S]0
Schéma cinétique minimum
Vmax
Vmax/2
(Ici, kcat = k2)
v Vmax [S]
[S]KM
kcat[E][S]
[S]KM
Vmax = kcat [E]0a
v
e
c
Etape physique
de liaisonEtape chimique
de catalyse
KM
Vi = vitesse initiale
KM = (k-1 + k2)/k1
Saturation de
l’enzyme

Various data types that may be used to estimate the kinetic parameters
obtained from the classical Michaelis–Menten equation.
After Tummler et al. 2014, FEBS J, 281, 549
Estimation of kinetic parameters
Flux measurements
- Nutrient
uptake rates
- 13C-labeling
(steady-state
and dynamics)
Enzyme concentrations
-- Quantitive
proteomics
- Qualitative
proteomics
- Expression data
Kinetic parameters
Substrate concentrations
- In vitro assays
- Biacore
- Existing models
- Structure
calculations
- Metabolomics
(steady-state and
dynamic)
- Proteomics for
signaling systems
v [E]0kcat[S]
Km [S]
v [E ]0 kcat[S]
Km [S]

Towards single-molecule enzymology
But both enzyme
conformation and catalytic
activity fluctuate with time
. Are they methods to measure
precisely this stochasticity?
. Is the Michaelis model still valid ?
11-mer
TRAP
12-mer
TRAP
Matsunada et al. PLOS One , 2012, é50011

Enzymologie de la molécule unique
Mesure directe de la distribution en fonction du temps d’une propriété moléculaire
pour une seule molécule et pour un temps long (plutôt que mesure pour la moyenne d’un
ensemble de molécules) : construction d’histogrammes d’une variable particulière et analyse
statistique des données.
L’analyse statistique des trajectoires d’une seule molécule donne une information
dynamique détaillée sur le fonctionnement de cette molécule.
Possibilité de suivre des processus biochimiques en temps réel et de capturer des
intermédiaires.
Vers l’élucidation de mécanismes réactionnels.
Nécessite :
(1) un important travail théorique;
(2) des techniques particulières pour conduire les essais expérimentaux : elles sont
en continuelle évolution et sont utilisables y compris dans les cellules.
De nouvelles expériences révolutionnaires permettent d’observer et de manipuler des molécules
uniques et de suivre la dynamique d’une seule molécule enzymatique :
. il y a des fluctuations pour la production du produit.
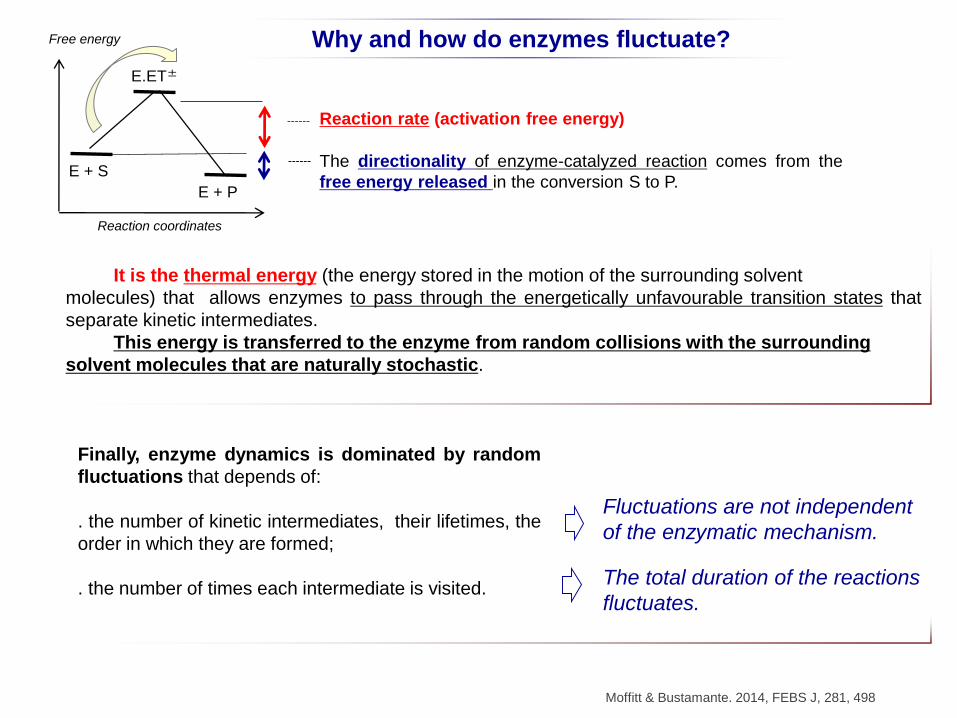
Why and how do enzymes fluctuate?
Moffitt & Bustamante. 2014, FEBS J, 281, 498
It is the thermal energy (the energy stored in the motion of the surrounding solvent
molecules) that allows enzymes to pass through the energetically unfavourable transition states that
separate kinetic intermediates.
This energy is transferred to the enzyme from random collisions with the surrounding
solvent molecules that are naturally stochastic.
E + S
E + P
E.ET±
The directionality of enzyme-catalyzed reaction comes from the
free energy released in the conversion S to P.
Free energy
Reaction coordinates
Reaction rate (activation free energy)
Finally, enzyme dynamics is dominated by random
fluctuations that depends of:
. the number of kinetic intermediates, their lifetimes, the
order in which they are formed;
. the number of times each intermediate is visited.
Fluctuations are not independent
of the enzymatic mechanism.
The total duration of the reactions
fluctuates.

How are the fluctuations measured and quantified?
Moffitt & Bustamante. 2014, FEBS J, 281, 498
Regardless the method used, measurements of the exact time at which the enzyme completed each cycle = ‘cycle
completion time’ or ‘dwell time’: total time that an enzyme takes to complete a cycle = sum of the time spent in each of its
different enzymatic states. The times spent between each state are often termed ‘waiting times’.
Formalization of the concept by defining the probability that the enzyme will visit a given state at least
once in the cycle.
p0 = 1: for on-way states through which the enzyme must pass.
p0 < 1: for states that are not necessarily visited each cycle (ex. EI)
Single molecule fluorescence
microscopy. Burst for the production
of fluorescent product
Change in position of a molecular motor on
its polymer track detected by the movement of
optically or magnetically trapped bead
Change in a DNA molecule length
strechted between two optical trapped
beads to reveal a nucleic acid motor
E + S ES E + P
E + S ES E + P
IEI
Fluctuations

Theoritical understanding of single-molecule kinetics
Steady state approximation
with
Ensemble averaged kinetics Single-molecule kinetics
C C [S]
[P]
C [S] Cste
[P] Cste
Obtention of waiting time distribution
normalized such as
Adapted from Kou et al. 2005, J Phys Chem B 109, 19068
E + S ES E0+P E0 Ek2k1
k-1
d
with at t = 0: PE(0) = 1, PES(0) = 0, PE0 (0) = 0
and PE(t) + PES(t) + PE0 (t) = 1
Probability
density of the
waiting time
Vitesse
initiale

Ensemble averaged kinetics Single-molecule kinetics
Each emission intensity peak corresponds
to a burst of enzymatic turnover.
Bursts
Blank
v
0
0.2
0.4
0.6
0.8
1
1.2
0 5 10 15 20 25
vi
[S]0
KM
Vmax
Vmax/2
[P]
Time (min)
[S]0
vi1
vi2
vi3
vi4
vi5 = vi
6 = Vmax
Michaelis-Menten equation
Real-time single-molecule recordings
of enzymatic turnovers
Statistical analysisProbability density of waiting time
Michaelis-Menten equation?

Probability density of the waiting time, f(t)
with k1 = 107 M-1 s-1, k2 = 250 s-1, k-1 = 50 s-1
Exponential rise followed by an
exponential decay corresponding to the
generation of the intermediate ES.
Average reaction rate 1/<t> or its equivalent v/[E]T as a function of [S]:
Single molecule
Michaelis-Menten
equation
v/[E]T = 1/<t>
KM = 30 µM
Single-Molecule Michaelis-Menten Kinetics in the Absence of Dynamic Disorder
Characteristic hyperbolic profile of the classical
Michaelis-Menten saturation curve.
Adapted from Kou et al. 2005, J Phys Chem B 109, 19068
<t>: mean waiting
time for the reaction
Consistency between the single-molecule
and ensemble averaged kinetics

Single-Molecule Michaelis-Menten Kinetics in the Presence of Dynamic Disorder
General applicability of the Michaelis-
Menten equation to single-molecule kinetics
even in the presence of dynamic disorder.
Multiexponentiality attributed to dynamic disorder.
The three states of the enzyme E, ES and E0 can each
exist in two interconverting conformations.
Two state model
Single molecule Michaelis-Menten equation holds
even under the condition of dynamic disorder.
Multistate model
Real enzyme systems are likely to interconvert among a
large number of conformational substates.
The value of kcat fluctuates.
Rate limiting step:
. at low [S], binding of the substrate to the enzyme;
. at high [S], dissociation of the ES to product.
Adapted from Kou et al. 2005, J Phys Chem B 109, 19068

Concentration initiale de substrat [S]0
Vit
esse
init
iale
[E]0 << [S]0
Conditions d’état stationnaire
Dans ce type d’expérience, les concentrations
d’enzyme et de substrat sont maintenues constantes
(comme dans les cellules vivantes)
D’après Xie Sunney X 2013 Science 342 1457
Equation de Henri-Michaelis-Menten :
(1) Quantification des cinétiques enzymatiques;
(2) Moyen pratique pour caractériser une enzyme
en termes de kcat et KM.
Indicateurs de l’efficacité enzymatIque :
haut kcat, bas KM et haut rapport kcat /KM .
Volume d’intérêt
En résumé
Pour une cinétique donnée, [S] diminue
Cinétiques
rapides(stopped-flow, flux
continu)
Mise en
évidence de la formation
d’intermédiaires
Donne en plus des informations cruciales sur la
formation d’intermédiaires et le désordre dynamique.
Etablissement d’état stationnaire
‘Turnover’ d’une molécule uniquePas d’état stationnaire

Enthalpie libre DGEtats de transition ET‡
Coordonnées réactionnelles
DGcc‡
DGnc‡
DG0
DGenz‡
Non catalysé
Catalyse chimique
Catalyse enzymatique
RéactifsE + S
ComplexeES Complexe
EP
ProduitsE + P
Non catalysé
Catalyseur chimique
Catalyseur enzymatique
ES, EP : complexes non covalents
Diagramme traditionnel d’enthalpie libre pour une réaction enzymatiqueComparaison avec une réaction non catalyése ou catalysée par un réactif chimique
DG‡ Enthalpie libre d’activation
DG0 Enthalpie libre de la réaction
DGnc‡DGcc
‡DGenz‡ < <
Le catalyseur stabilise l’état de transition de la réaction catalysée par rapport à la réaction non catalysée : DG‡ DIMINUE et la constante de vitesse k AUGMENTE d’où l’accéleration.
Relation de Eyring: k = (kBT/h) e-DG‡ /RT
L’équilibre de la réaction n’est pas modifié par le catalyseur : DG0 est INCHANGE. DG0 = -RT ln Keq
kB : constante de Boltzman; h : constante de Planck; R : 2 cal/mol/degré
[ES]‡
[S]‡

Free
energy
Reaction coordinate
Transition state
Products
Reactants
A- + B-C
A-----B-----C
d d
A-B + C-
Free energy profile of a chemical reaction
Cleavage of B-C bond
Formation of
A-B bond
A- + B-CReactants
Products
A-B + C-
Transition state
A-B--C
Unstable
inermedaite
A- + B+ + C-
Unstable
intermediate
Free energy surface
Reactants
Products
1
2
21

Enzyme fluctuations: standard free energy lanscape
Hammes-Schiffer 2013 Biochemistry, 52, 2012.
S + E1 ES1 P + E10 E1
S + E2 ES2 P + E20 E2
S + En ESn P + En0 En
… … …
Kinetic scheme of the multistate model
involving n interconverting conformers.
Schematic representation of the standard free energy
landscape for the catalytic network of an enzyme reaction
Parallel catalytic
pathways
The reaction paths can slide along and between both coordinates:
fluctuations of kcat
kcat

Single Molecule Fluorescence Assays by Fluorescent Measurements
. Assays with fluorescent active sites (cholesterol oxidase).
. Assays with fluorescently labeled substrates (single molecule DNA sequencing).
. Assays based on a fluorogenic substrate converted to a fluorescent product molecule by an
enzymatic reaction.
. Assays with fluorescent resonance energy transfer (FRET) pairs that report conformational changes
triggered by enzyme tunrovers (staphylococcal nuclease, a ribozyme, T4 lysozyme).
. Assays based on photo-induced electron transfer used between excited states (flavin and tyrosine
residues to probe conformational motions associated with enzymatic reactions of flavoenzymes).
- Complementary to FRET, probes conformational changes at shorter distances (several Å) than FRET
probes (several nm).
. Assays based on rotational and translational trajectories of molecular motors by recording movies
of fluorescent labels (relative rotation of RNA polymerase and RNA usisng a magnetic and fluorescent
bead; translation of possessive motor enzymes such as kinesin, myosin, DNA polymerase, helicase).
Flow Stretching Assay
Other single molecule manipulation methods
Single molecule tools for enzymology
Xie 2001, Single Mol 2, 4, 229 ; Widom et al. 2014 Arch Toxicology 88, 1965
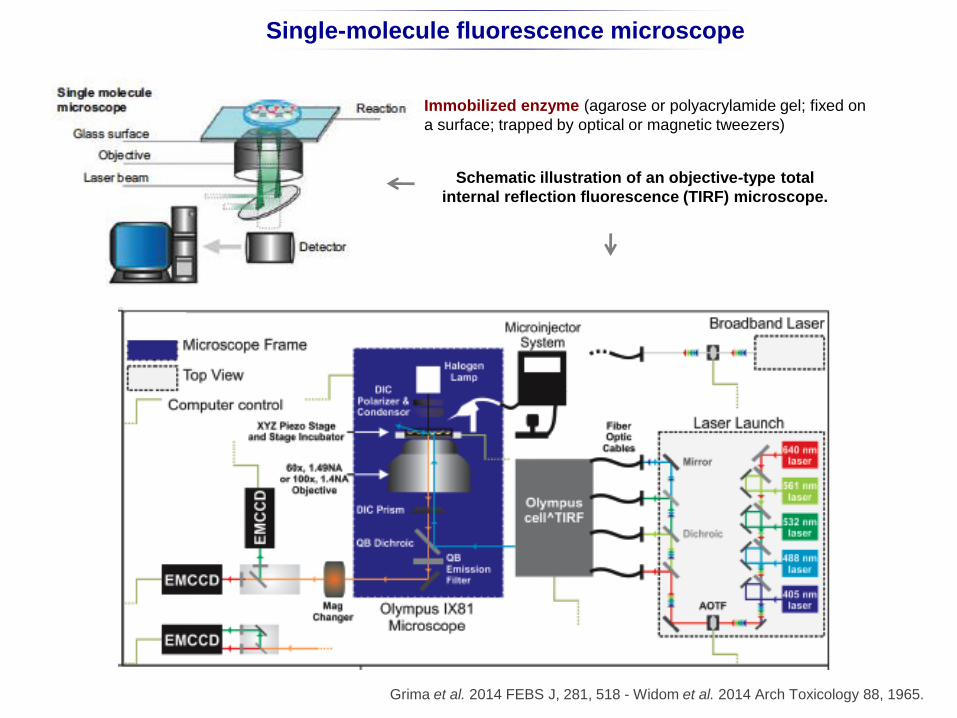
Single-molecule fluorescence microscope
Schematic illustration of an objective-type total
internal reflection fluorescence (TIRF) microscope.
Grima et al. 2014 FEBS J, 281, 518 - Widom et al. 2014 Arch Toxicology 88, 1965.
c
Immobilized enzyme (agarose or polyacrylamide gel; fixed on
a surface; trapped by optical or magnetic tweezers)

Real-time single-molecule recordings of enzymatic turnovers
A fluorogenic substrate is converted into fluorescent product. Each emission intensity peak corresponds to a burst
of enzymatic turnover.
Binding and unbinding of fluorescently labeled probes to an immobilized oligonucleotide.
Grima et al. 2014 FEBS J, 281, 518 - Widom et al. 2014 Arch Toxicology 88, 1965.
Recordings
Experimental setup Fluorescent intensity trace
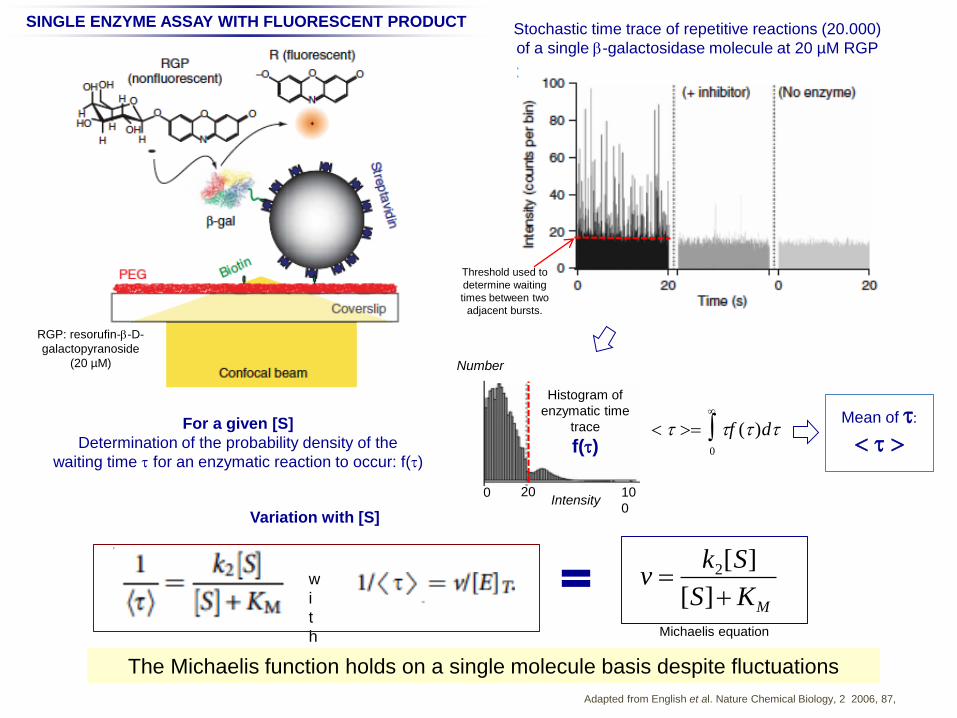
Adapted from English et al. Nature Chemical Biology, 2 2006, 87,
Stochastic time trace of repetitive reactions (20.000)
of a single b-galactosidase molecule at 20 µM RGP
b-galactosidase
Threshold used to
determine waiting
times between two
adjacent bursts.
Histogram of
enzymatic time
trace
f(t)
20 10
0
0Intensity
Mean of t:
<t>
SINGLE ENZYME ASSAY WITH FLUORESCENT PRODUCT
RGP: resorufin-b-D-
galactopyranoside
(20 µM)
For a given [S]
Determination of the probability density of the
waiting time t for an enzymatic reaction to occur: f(t)
v k2[S]
[S]KM
Variation with [S]
w
i
t
h Michaelis equation
Number
The Michaelis function holds on a single molecule basis despite fluctuations
< t > tf (t )dt0

Waiting time fluctuations are explained by enzyme conformation fluctuations.
Aim: create conformations that able to accommodate the substrate.
. Spontaneous or thermal activated fluctuations (ubiquituous).
. Nonequilibrium process driven by substrate binding or chemical reactions.
Techniques used to study conformational changes
- FRET (fluorescence resonance energy transfer), only sensitive to the distance
change on the order of nanometers (Weiss 2000);
- method based on photo-induced single-electron transfer (Yang et al. 2003);
- method based on motion hindering in the excitated stated (Luo et al 2007)
Two types
conformational
changes
Single molecule conformational dynamics

Preuve de la dynamique d’une molécule unique par transfert d’électrons photoinduit
hn
hn :excitation laser UV à faible
let forte fréquence
g
g :vitesse de recombinaison
radiative (réémission d’un photon
à une fréquence plus basse)
kg: vitesse de recombinaison non
radiative (désexcitation)
kg
kEt
Influence du
milieu extérieur
Un microscope confocal est utilisé pour détecter
le signal de fluorescence d’une seule molécule.
kEt: transfert d’énergie sur une autre molécule (par
ex, entre un donneur et un accepteur)
Diagramme d’énergie de l’état excité Et
Très sensible à la fluctuation de la distance entre
donneur et accepteur (échelle de l’Å) qui se traduit par
une fluctuation du temps de fluorescence.
Donneurs naturels : Tyr, Trp, guanine du DNA
Accepteurs naturels : flavine ou colorants artificiels
comme la rhodamine

Single-Molecule Conformational Dynamics Probed by Electron Transfer
Structure of flavin reductase Fluorescence decays of the wild type
and mutants of flavin reductase
By probing the fluorescence lifetime of the single flavin:
variation of flavin-tyrosine distance over time thus conformational
fluctuations at multiple time scales spanning from hundreds of µs to s.
Yang et al. 2003 Science, 302, 262
Oxidized flavin Reduced flavin
NADH NAD
Favin reductase
Y35S/FMN
Free FMN
2 H+,
2 e-
Reduction
Pulse laserDrastic increase of fluorescence lifetime only
observed for the Y35S complex (red curve), proving
the quenching by tyrosine Y35.
Existence of multiple interconverting conformers related to the fluctuating catalytic activity.

Fluorescence intensity of Cy3 is increased when the nonradiative decay rate of Cy3 is decreased due to hindered torsional motions of the excited state of Cy3 molecule: reflects closing of the fingers domain.
Probing Non-equilibrium Conformational Motions by Fluorescence Intensity Changes of a Hindered Fluorophore
Primed DNA duplex
labeled with Cy3 dye
molecule
Exonuclease part
Closing of DNA Polymerase fingers domain
dNTP
Primed DNA
With dNTP
Li et al 1998 EMBO J, 17, 7514; Luo et al.
2007 PNAS, 104, 12610
closedopen
Ternary
complex
Detection real time
conformational changes

Assays with fluorescent active sites
Cholesterol oxidase
Crystal structureFluorescence image
(8 x 8 µm) of single E
molecules immobilized
in a agarose gel
Distribution of on-timesReal-time observation of enzymatic turnovers of a single
enzyme catalyzing oxidation of cholesterol moleculesLu et al. 1998, Science, 282, 1877
E + S ES Productk2k1
on
on
on off
off off
Occu
ren
ce

Flow Stretching Assay
Van Oijen et al , 2003, Science, 301, 1235,
Monitoring the activity of individual nucleic acid enzymes on DNA simultaneously
Two digestion traces showing complete enzymatic
conversion of l-phage dsDNA into ssDNA The coiling of l-phage ssDNA causes it
to be shorter than dsDNA
Model of the l
exonuclease complex
dsDN
A
ssDNA
ssDNA
dsDN
A

Other single molecule manipulation methods
Novo folding of ribosome-bound nascent proteins
Bustamante et al , 2014, Ann Rev Biophys, 43, 119
Optical tweezer measurements

Spécificité
. Types de réaction catalysée
. Nature du (des) substrat(s) pour un type de réaction donnée
. Selon l’enzyme, la spécificité est plus ou moins étroite: notion de promiscuité
La spécificité dépend d’un arrangement très précis des atomes du centre actif

The specificity is related to the chiral structure of the enzyme molecule, including the three-dimensional folding of the polypeptide backbone and the orientation of
the amino acid side chains in the folded enzyme .
Total chemical synthesis
of D-enzymes
Milton et al. Science 1992
256, 1445 – Kent J Peptide
Science, 2003, 574
L-HIV-1 protease D-HIV-1 protease
L-PR
+
L-Sub
L-PR
+
D-Sub
Time (min)Time (min)
D-PR
+
L-Sub
D-PR
+
D-Sub
Flu
ore
scen
ce
Each enzyme eniantomer cuts only the corresponding substrate eniantomer:
reciprocal chiral specificity on peptide substrates
Flu
ore
scen
ce

Promiscuité enzymatique
L’enzyme peut effectuer des réactions non prévues (si on se base sur sa spécificité connue)
Promiscuité de conditions réactionnelles.
Enzymes ayant une activité catalytique dans diverses conditions réactionnelles qui sont différentes
de ses conditions naturelles (par exemple : milieu anhydre, températures ou pH extrêmes).
Promiscuité dans la gamme de substrats reconnus.
Enzymes ayant une spécificité très large.
Promiscuité catalytique.
Enzymes catalysant de manière distincte différentes transformations chimiques qui présentent des
états de transition différents.
Elle peut être
a. accidentelle : réaction annexe catalysée par l‘enzyme.
b. induite : une nouvelle réaction est établie suite à une ou plusieurs mutations ce qui change la
nature de la réaction par rapport à l’enzyme sauvage (ingéniérie des protéines).

Induction de promiscuité
catalytique
Glycosidase
(type sauvage)
Glycosynthase
Thioglycoligase
Thioglycosynthase
Acide aminé
muté
See also Hancock et al 2005 ChemBioChem, 6, 866 and references therein.

F. Stoll et al. 2011 Drug Discovery Today,
16
Promiscuity of ligand binding of cytochrome P450 2C9
Superposition of several X-ray structures of
complexes enzyme-ligands
Active site

Fonctionnement régulé des enzymes
. Régulation de l’activité par changement de conformation.
Rétro-contrôle; effecteurs allostériques
A B C ------ Y
. Régulation par modification de la structure covalente de l’enzyme.
Modifications covalentes réversibles : phosphorylation, adénylation, protéines G (GDP/GTP).Modifications irréversibles : protéolyse (activations protéolytiques), ADP-ribosylation.
. Contrôle exercé par des inhibiteurs naturels.
Si déficient : polymérisation et induction de pathologies conformationnelles.
. Régulation par des molécules exogènes d’intérêt pharmacologique
-E1 E2 E3 En

The proteolytic cascade of kallikreins (hKs) involved in
epidermidermal homeostasis
LEKT
I
hK5pro hK5
hK14
hK14pro
Cornification Desquamation Innate immunity Inflammation
hK7
hK7pro
CathelicidinProfilaggrin PAR-2
IL - 1 β
Desmoglein (DSG-
1)
Desmoplakin
Desmocollin 1-3
hK5
hK5
Matriptase LEKTI: natural inhibitor
(145 kDa)
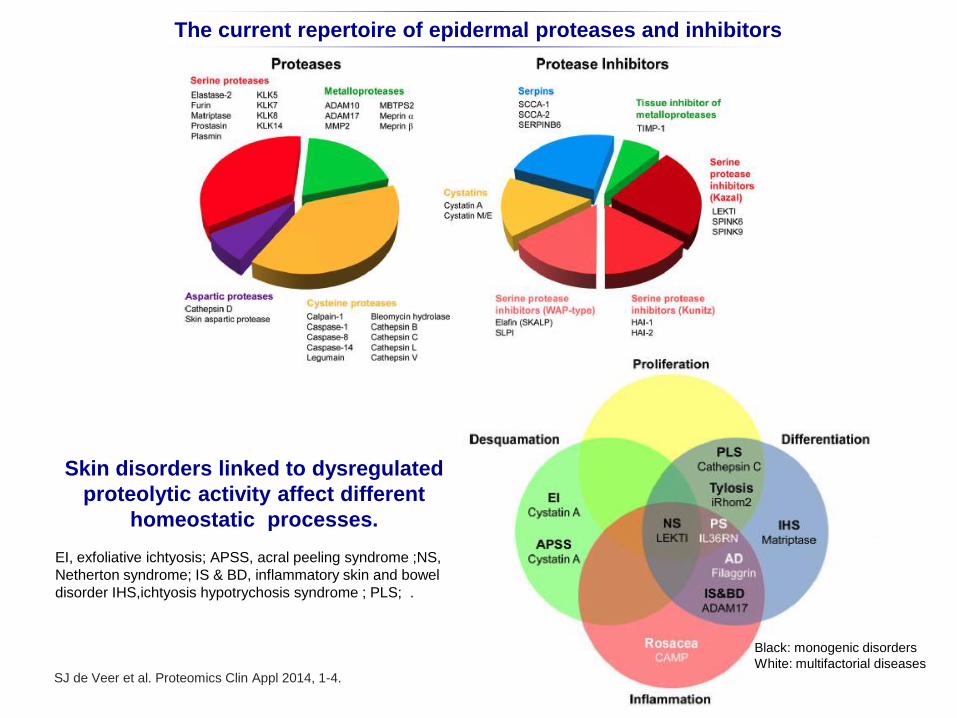
The current repertoire of epidermal proteases and inhibitors
SJ de Veer et al. Proteomics Clin Appl 2014, 1-4.
Skin disorders linked to dysregulated
proteolytic activity affect different
homeostatic processes.
EI, exfoliative ichtyosis; APSS, acral peeling syndrome ;NS,
Netherton syndrome; IS & BD, inflammatory skin and bowel
disorder IHS,ichtyosis hypotrychosis syndrome ; PLS; .
Black: monogenic disorders
White: multifactorial diseases

Mauvais repliement
Trafic incorrect
(Emphysème pulmonaire)
Repliement toxique : agrégats fibrillaires
(Amyloïdoses)
Maladies conformationnelles ou du mauvais
repliement des protéines
Probabilité augmentée de dégradation
(caveolin-3, LGMD-1C; CFTR, mucoviscidose)
Protéine dépliée Protéine native
fonctionnelle
La perte d’activité des inhibiteurs peut être due à un
mauvais repliement
a1-PI(a1-antitrypsine)
Recyclage et
perte de
fonction
Synthèse et
acquisition de
fonction
Etat cellulaire normalEquilibre entre synthèse et
recyclage des protéines
(pour une protéine donnée)
From Nature Chem Biol, nov 2014)
Lomas & Mahadeva J
Clin Invest 2002, 110,
1585

Bibliography
Special Issue. Enzyme Catalysis and Allostery: A Century of Advances in Molecular
Understanding. FEBS J, 2014, 281.
References cited in the diaporama.
Website: http://bernstein.harvard.edu/research/singlemolecule.html
Books
Athel Cornish-Bowden
Fundamental of enzyme kinetics (2004; 2012); Ed. Wiley-Blackwell.
Robert A. Copeland
Enzymes: A pratical introduction to structure, mechanisms and data analysis (2000)
Evaluation of Enzyme Inhibitors in Drug Discovery (2013); Ed. Wiley-VCH.
Daniel L. Purlich
Enzyme kinetics: catalysis and control (2010) Elsevier.
Alan Fehrst
Enzyme structure and mechanism in protein science: a guide to enzyme analysis and protein folding (1999 );
Ed. WH Freeman.
The selected papers of Sir Alan Fersrt: development of protein engeneering. ICP selected papers (2010).
Jeannine Yon-Kahn & Guy Hervé.
Molecular and cellular enzymology (2010); Ed. Springer-Verlag.
Irwin H. Segel
Enzyme kinetics: behavior and analysis of rapid equilinbrium and steady-state enzyme systems (1993); Ed.
Wiley Classics Library.





























