Embed Size (px)
Citation preview

ABSTRACT We used comparisons among T. thermophila and its close relatives to determine the importance of the locations and sequence features of fragmentation sites. The target of this study was a 4.5 Mb region - supercontigs 6 and 11 -on Mic chromosome 3L of T. thermophila. The work included A) sequence alignment using database sequences of micronuclear/macronuclear DNA of T. thermophila and macronuclear only –no Mic sequences available - from three relatives of T. thermophila (T. malaccensis, T. elliotti and T. borealis), and B) experimental investigation of Chromosome breakage sequences between pairs of adjoining macronuclear chromosomes in relatives of T. thermophila.
Results of sequence alignments showed that all macronuclear chromosomes were co-linear with the T. thermophila Mic supercontigs 6 and 11, with the exception of supercontigs 14 and 61 of T. elliotti. These two supercontigs aligned at two noncontiguous positions on the T. thermophila Mic supercontig 11. Supercontigs 14 and 61 contain an almost 6 kb inverted repeat. The most likely explanation of the observed alignment is a micronuclear inversion in T. elliotti.
Findings related to the conservation of chromosome breakage sites are shown in a companion poster.���
A probable inversion in the micronucleus of Tetrahymena elliotti compared to chromosome 3 of T. thermophila
P. E. Huvos, E. Orias E. P. Hamilton, Department of Biochemistry and Molecular Biology, Southern Illinois University, Carbondale, IL; Department of Molecular, Cellular and Developmental Biology, University of California at Santa Barbara, Santa Barbara,
1
T. therm MAC scaffolds
T. mal MAC scaffolds
T. el MAC scaffolds
T. bor MAC scaffolds
4517 4504R 4410R 4437
299 250 208 49
14A-1 119 R 30CR
172R 171 160 47R
CBS4=3L-‐18 533kb AAAGAGGTTGGTTTT CBS5=3L-‐17
685k
44(+71) kb
52 (+73) kb
100 kb
107 kb
100 kb
97 kbp
100 kb
107 kb
139 kb
140 kb
140 kb
144 kb
548 kb
556 kb
622 kb
61-AR
10 kb
30BR
598 kb
CBS3=3L-‐19 431kb TAAACCAACCTCTTT
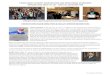
Alignment of T. thermophila Mic supercontig 11 to Mac chomosomes of thermophila and three of its relatives. The positions and lengths of the scaffolds shown are approximately right, but the scale is different in the different slides to make viewing more convenient. The chromosome maps were drawn based on the database (http://www.broadinstitute.org/annotation/genome/Tetrahymena/Regions.html), but some information derived from the experimental work described here is also shown. The Micronuclear supercontings and macronuclear scaffolds are represented by horizontal lines, with numbers above the lines showing the scaffold numbers, and those below the lines showing their lengths. Dashed lines indicate that the chromosome extends beyond the slide. Short vertical lines crossing the line representing the Mic chromosome show the positions of chromosome breakage sites (Cbs). Short thick blue lines show the presence of telomeres assembled in the database at the ends chromosomes. Thick purple lines show telomeres identified in this work by sequencing, dashed purple lines show telomeres identified only by the existence of the relevant PCR products when combining primers from the chromosome ends with telomere primers. A thinner purple line shows the existence of a telomere based on indirect evidence for the neighboring Cbs being functional. Green squares represent regions of Cbs in T. thermophila that were investigated in other species (T. malaccensis, T. elliotti, T. borealis in increasing phylogenetic distance). Red squares show where homologous regions were successfully amplified in other species. Purple squares denote PCR products containing functional Cbs sequences derived from contiguous chromosome sections according to the database.
CBS7=3L-‐15 1521k TAAACCAACCTCTTT
CBS6=3L-‐16 1284 AAAACCAACCTCTTT
T. therm. MAC scaffolds
35R
656 kb
166R
4610
14-A2R
92
T. mal. MAC scaffolds
T. elliotti MAC scaffolds
T. borealis MAC scaffolds
4588
129 47R
49
30BR
4437
556 kb
598 kb
622 kb
202 kb
269 kb
548 kb 214 kb
125 kb
367 (+427) kb
663 kb
61-B 30AR
207 kb 543 kb
1,073 kb
T. therm MAC scaffolds 4582R 4415 4517
217bp
T. mal MAC scaffolds 16R 16 R
1,047 kbp 216 kbp
14C T. el MAC scaffolds
1,123 kb
T. bor MAC scaffolds 18 133R 172R
1,016 kbp 249 kb 107 kb
T. Therm. MIC Supercont. 11-‐-‐1 983 kb CBS1=3L-‐21 Pos 79 bk TAAACCAACCTCTTT
CBS2= 3L-‐20 305 kb AAAACCAACCTCTTT
3894
71 kb 45 kb
61A
299 276R
73 kbp 51 kbp
14B 14A-1
238 kb 100 kb
Supercontigs 14 and 61 contain a 5.7 kb repeat in inverted orientation with 85% sequence identity (with relatively long sections of almost 100% identity). It therefore seemed possible that the chromosomes had been misassembled in the database. To test for this, probes derived from noncontiguous sections of the two T. elliotti MAC supercontigs were hybridized to MAC chromosomes on CHEF gels to see which of those would co-hybridize.
The results show that the two parts of supercontig 61 (probes 1 and 2) hybridize to the same band at around 550 kb, whereas the separated part of supercontig 14 (probe 3) hybridizes to a smaller band somewhere between 225 and 285 kb. That is, the assembly presented in the database is correct. If the alternative were true, probe 2, derived from the beginning supercont 6, would hybridize to a shorter chromosome extending from CBS2 that was identified in the present study, and probe 3 would hybridize with a 6-700 kb band with the rest of supercontig 61.
The repeats in spctg 14 and 61 have been assigned different but perhaps related coding sequences
The black arrows show the positions of the repeat in spctg 14 and 61.
Only T. elliotti has the repeat at the position of the ancestral spctg 61
A ‘patched’ ancestral T. elliotti spctg 61 was constructed by fusing positions 110kb-130kb from spctg14R to positions 13k-30k from spctg 61. The 36 kb construct was aligned with homologous scaffolds from the other three species (scaffold 8254610 of T. thermophila, spctg 35 of T. malaccensis and spctg 92 of T. borealis). The area corresponding to the repeat, i.e. the end of the spctg 14 (124kb-129.6 kb), was missing in all species. The alignment with T. thermophila (Y coordinate) is shown.
Alignment of the inverted repeats with the homologous region in the other three species does not reveal much difference in evolutionary constraints on the two repeat copies.
61 repeat 14 repeat % Query coverage % Identity Query coverage % Identity
T. therm. 8254517 64 74 65 75
T. mal. spctg 299 72 72 73 73
T. bor. spctg 172 25 70 27 71
Results of noncontiguous alignments of two T. elliotti chromosomes to T. thermophila chromosomes are best explained by a micronuclear inversion. The presence of 5.7 kb inverted repeats near the expected breakpoints in the two chromosomes would explain the mechanism of inversion. Other, more complex, explanations have not been excluded.
Whether the additional copy could have a function is unclear.
Model of the micronuclear inversion in T. elliotti
Aligment of T. elliotti spctgs 14 and 61 (X coordinates) to T. thermophila Mic spctg 11 (Y coordinates)
14 19 18 30A 17 30B 16 30C 15 119 21 20 14B 61 14A
14 15 16 30A 17 30B 18 30C 19 119 21 20 14B 61 14A 22
22
14C
14C
Current/ ancestral T.ell sp30A DNA
Current sp14, ancestral T.ell sp61 DNA
Current/ ancestral T.ell sp14A DNA Current/
ancestral T.ell sp119 DNA
Current/ancestral T.ell sp61 DNA
Current sp61, ancestral T.ell sp14 DNA
T. thermophila configuration, i.e. ancestral T. elliotti configuration
Derived, i.e. current T. elliotti configuration