Embed Size (px)
Citation preview

Biological Conservation 51 (1990) 287-297
Pollination of the Orchid Dactylorhiza fuchsii by Longhorn Beetles in Primeval Forests of Northeastern
Poland
Jerzy M. G u t o w s k i
Forest Research Institute, Department of Nature Protection, 17-230 Bialowie~a, Poland
(Received 22 August 1988; revised version received 21 May 1989; accepted 23 June 1989)
ABSTRACT
The pollination of the orchid Dactylorhiza fuchsii ( Druce ) Sob by Alosterna tabacicolor (Deg.) (Coleoptera: Cerambycidae) is recorded for the first time. This longhorn beetle--particularly the males-- was found to be the main pollinator of orchids in the Bialowie~a and Augustbw Forests, NE Poland. The mechanism of pollination is described. Pseudocopulation was not observed; however, the possibility of sexual attraction of male A. tabacicolor to the orchids was not excluded. A certain dependence of D. fuchsii on the beetle species both ecologically (they appear simultaneously and are almost exclusively pollinated by these beetles) and geographically was observed. The consequences of the disappearance of one of these symbiotic partners are analysed. The evolution of longhorn beetle adaptation to flower pollination and problems of the preservation of orchids and their pollinators are discussed.
I N T R O D U C T I O N
Orchids (Orchidaceae) have developed an advanced adaptation to pollination of individual species by specific animals, primarily insects. The orchid pollen is not free but is formed into masses, the so-called pollinia. Pollinia stick to the insects' bodies by the viscid matter at their bases and in this way are carried to the next flowers. The very complex construction of most orchid species means that they can only be pollinated by a specific, relatively small,
287 Biol. Conserv. 0006-3207/90/$03"50 O 1990 Elsevier Science Publishers Ltd, England. Printed in Great Britain

288 Jerzy M. Gutowski
group of insects, for example, a family, genus or even a single species. The size of the pollinating insects and the construction of their mouthparts are important determining factors. There are also several cases of characteristic attraction of males of a specific group of insects to orchids which have perianths imitating the female, either by shape or inside design as well as by scent (Olberg, 1951; Pervukhina, 1970; Sundermann, 1975; Halvorsen, 1986). Cases of pseudocopulation by males have been recorded in some Hymenoptera (Kullenberg, 1961; Ford, 1967; Proch~izka & Velisek, 1983).
Orchids are usually pollinated by Hymenoptera, and less often by Lepidoptera, Diptera and others. Very rarely beetles (Coleoptera), mainly from the families Scarabaeidae, Staphylinidae, Nitidulidae and Oedeme- ridae, take part in their pollination (Silrn, 1906a; Olberg, 1951; Kullenberg, 1961; Proch~izka & Velisek, 1983; Borg-Karlson, 1987).
The purpose of this work was an exploratory investigation as to the insect species that pollinate the protected orchid Dactylorhizafuchsii in forests of northeastern Poland, how this process is done under natural conditions and to analyse how specific are the connections between orchids and longhorn beetles.
STUDY AREA AND METHODS
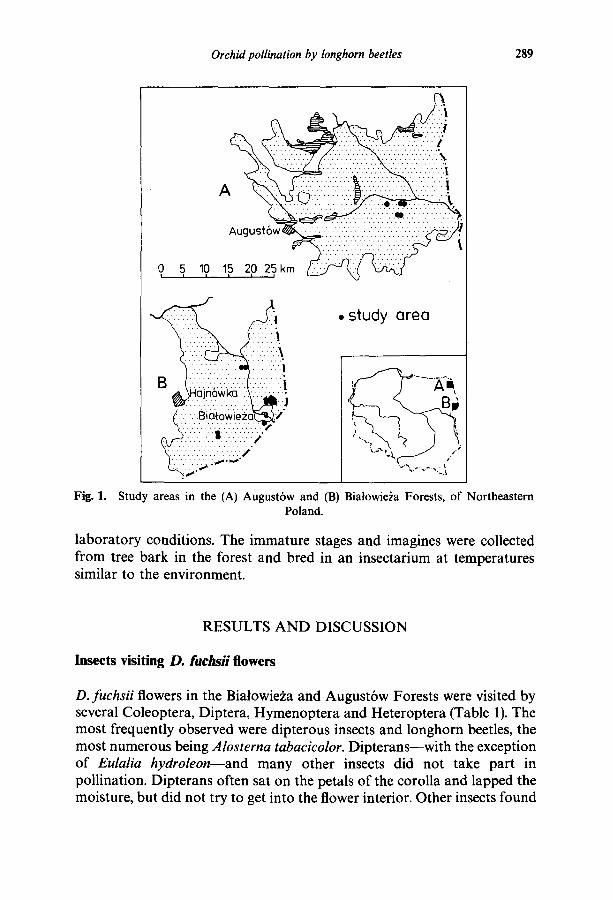
Observations were carried out from 1979 to 1982 and from 1985 to 1987 in areas with many D. fuchsii in the Biatowie~a Forest (Biatowie2a National Park--7 localities; forest units: Czerlonka--2, Gruszki--2, Podolany--1) and in the Augustbw Forest (near the village of Jazy--2, Staro2yn reserve-- 2, Perku6 reserve--l) (Fig. 1).
D.fuchsii usually grows on the edges of forests, sometimes in clearings, in rich forest associations, for example Tilio-Carpinetum stachyetosum Traczyk 1962, T.-C. caricetosum remotae Sokotowski 1968, T.-C. circaeaetosum alpinae Sokotowski 1969, Circaeo-Alnetum Oberdorfer 1953.
In the BiatowieZa Forest D. fuchsii usually blossoms between 5 and 30 June, during which time interaction with insects was observed. The number of longhorn beetles (those with pollinia on their bodies were considered separately) observed near the orchid localities was counted during 16 half- hour sampling periods. During such a period an average of 100 longhorn beetle specimens was noted. In addition all insects visiting the specified group of flowering orchids were observed during 31 half-hour sampling periods. Their behaviour and method of pollination were noted. Some were collected for identification.
The biology of the longhorn beetle Alosterna tabacicolor, which appeared to be the main pollinator of D.fuchsii, was also examined under natural and
Orchid pollination by longhorn beetles 289
0 519
. . . .
A ' ~ i
" +. i :, ~ ; i : : " . . . . . . . . . . . . . . . . . . . . . .~
~':~~:::fiS:::::::::::::::::::::::::::::::::::::::::::::::::::: v.-.rtrl
1,s 20 2 S k ~ ~
Fig. 1.
B ::::::::::::::i -::: " i:::)
• study area
Study areas in the (A) August6w and (B) Biatowie~a Forests, of Northeastern Poland.
laboratory conditions. The immature stages and imagines were collected from tree bark in the forest and bred in an insectarium at temperatures similar to the environment.
RESULTS AND DISCUSSION
Insects visiting D. fuehsii flowers
D. fuchsii flowers in the Biatowie£a and August6w Forests were visited by several Coleoptera, Diptera, Hymenoptera and Heteroptera (Table 1). The most frequently observed were dipterous insects and longhorn beetles, the most numerous being Alosterna tabacicolor. Dipterans--with the exception of Eulalia hydroleon--and many other insects did not take part in pollination. Dipterans often sat on the petals of the corolla and lapped the moisture, but did not try to get into the flower interior. Other insects found

290 Jerzy M. Gutowski
TABLE 1 Species and Numbers of Insects Visiting Dactylorhizafuchsiiin the August6w and Biatowie~
Forests, NE Poland
Species Numbers
Carrying pollinia
Coleoptera Cerambycidae
Alosterna tabacicolor (Deg.) 38~c~, 16~, 33 sex non det. Anastrangalia reyi (Heyd.) 6~c~, 1~ Pachytodes cerambyciformis (Schrk.) 4 Judolia sexmaculata (L.) 1 Leptura aethiops Poda 1
Oedemeridae Oedemera virescens (L.) 1
Diptera Stratiomyidae
Eulalia hydroleon (L.) 1
Not carrying pollinia
Diptera Undetermined 38 Empididae
Empis sp. 33 Anthomyiidae 10
Coleoptera Melyridae
Dasytes plumbeus O. F. Miill. 14 Cerambycidae
Brachyleptura maculicornis (Deg.) 1 Elateridae
Ectinus aterrimus (L.) 1 Malachiidae
Malachius bipustulatus (L.) 1 Cryptophagidae
Antherophagus pallens (Fabr.) 1 Anaspididae 1 Undetermined 1
Hymenoptera Aculeata undetermined 7
Heteroptera Undetermined 2 Miridae
Asthenarius quercus (Kirsch.) 1

Orchid pollination by longhorn beetles 291
on the orchids were probably there accidentally (Heteroptera, Ectinus aterrimus, Malachius bipustulatus and others). The transfer of pollen by aculeates (Hymenoptera: Aculeata) was not seen, although these insects are the most common pollinators of other orchid species (Kullenberg, 1961; Proch/lzka & Velisek, 1983). Alosterna tabacicolor, a small longhorn beetle (body length 5-2-8"0 mm), occurs in large numbers, and in some areas even masses. They have head and mouthparts that are adapted to feeding on the pollen or nectar of flowers, a characteristic of the entire Lepturinae subfamily to which A. tabacicolor belongs (Grinfeld, 1975). Except for one record in Finland (Sil6n, 1906b), the domination of longhorn beetles in pollinating orchids was not previously known. In rare instances pollination by Scarabaeidae has been recorded (Kullenberg, 1961; Borg-Karlson, 1987). Sil6n (1906b) recorded Brachyleptura maculicornis (Deg.) and Stenurella melanura (L.) as the main pollinators of Orchis maculata L. Brachyleptura maculicornis was also noted on Listera ovata (L.) R. Br. Pijl, and Dodson (1966, cited in Nilsson, 1979) stated that Leptura and Grammoptera spp. were regularly seen on Herminium monorchis (L.) R. Br. blossoms. However, Nilsson (1979) considered this to be a misinterpretation.
Data on pollinators of D. fuchsii are very scarce and uncertain because this orchid was only recently separated from Orchis maculata. Available information suggests that dipterans are common pollen vectors of D.fuchsii throughout its range (Sil6n, 1906a, b; Hagerup, 1951; Nilsson, 1980; Prochfizka & Velisek, 1983). Longhorn beetles may, however, prevail locally, and even hymenopterans cannot be excluded as possible pollinators of minor importance.
Mechanism of pollination
The mechanism of pollination observed in A. tabacicolor was as follows. Insects flew above the flowers, came closer to them, almost hung in the air like a hummingbird near the perianth, searching for food with antennae pointing diagonally forwards. An insect would look inside several flowers (Fig. 2C), after which it flew away to repeat the action. After a while it might return to the same inflorescence. While thrusting the head and prothorax inside the flower it pressed its frons and pronotum on the anthers, inside which are the pollinia. The latter stick to the insect's body by means of retinacula (Fig. 3) and are transported with it to the next flower. At first the pollinia project almost perpendicularly from the surface of the frons or pronotum. Later they bend forwards and when the insect repeatedly visits flowers it comes in direct contact with the stigma of the pistil, causing pollination. The pollinia attached to the insect look like the conidia of some entomopathogenous fungus, or myxomycete. With D. fuchsii, in order for

292 Jerzy M. Gutowski
A B
Fig. 2.
f
Dactylorhiza fuchsii: (A), overall view; (B), individual flower from the front with pollinia (a); (C), flower being pollinated by Alosterna tabacicolor beetle.

Orchid pollination by longhorn beetles 293
Fig. 3. Alosterna tabacicolor: (A), with D. fuchsii pollinia stuck to the pronotum (a); (B), detail of pollinia stuck to the head of A. tabacicolor beetle.
pollination to be successful, the insects can neither be too small (e.g. Dasytes plurnbeus) nor too big. Probably A. tabacicolor is the optimum size. This function is carried out less frequently by somewhat larger insects, e.g. Anastrangalia reyi or Pachytodes cerambyciformis (Table 1).
Phenological observations and possible methods of attracting pollinators
During 16 half-hour observation periods in the Bialowie~a Forest 0-8-16.7% of the A. tabacicolor individuals observed had pollinia stuck to

294 Jerzy M. Gutowski
them, depending on the blossoming phase of D.fuchsii. This proportion was greatest at the beginning of blossoming (5-15 June) and gradually decreased.
Imagines of A. tabacicolor were already appearing in the Biatowie~a Forest at the end of May, and emergence usually ended during the first days of July. They fed primarily on pollen of Aegopodium podagraria L. (Umbelliferae). This did not deviate from the general preference of longhorn beetles--or of all anthophilous beetles--which usually select umbels that are white or greenish-white (Faegri & Pijl, 1982). However, in places where D. fuchsii occurred they also visited the pinkish red flowers of this orchid. Attraction is probably not to colour but to the scent which this plant emits. The insects seek out the centre of the flowers, where they expect to find pollen or nectar.
From observations of A. tabacicolor on Aegopodium podagraria it was seen that individuals carrying orchid pollinia behave normally--they are active, feed on the pollen of this species, and copulate. The majority of longhorn beetles with pollen collected from A. podagraria flowers were males. Sex of this species is difficult to differentiate in the field, so was not determined in some individuals. Perhaps during copulation the orchid pollinia adhering to the females are rubbed off by the mouthparts and the first pair of legs of the males. The large majority of males with pollinia, both at some distance from the orchids and directly on the D.fuchsii flowers, may suggest that the orchids emit an odorous substance that imitates the female sexual scent and is attractive to A. tabacicolor males. It is, however, impossible to prove this without experimental testing. Only Borg-Karlson (1987) has mentioned orchid scents which are sexually attractive to beetles, in this case scarabs attracted to Ophrya scolopax Cav. However, even when there are sexually attractive scents, beetles already on flowers do not try to copulate, but look for food. The shape and flower colour or the perianth parts of D. fuchsii do not resemble female A. tabacicolor.
The evolution of beetle adaptation to pollination
Beetles as flower pollinators are considered to be very primitive and are clearly less efficient than some hymenopterans or lepidopterans adapted for this purpose (Kevan & Baker, 1983). In spite of this it is felt that beetles were one of the first insects to pollinate angiosporous plants. It has been suggested by Diels (1916) that the first angiosporous plants were pollinated exclusively by Coleoptera. This hypothesis has been criticized (Pervukhina, 1970; Grinfeld, 1975) and the present opinion is that cantharophily was parallel to pollination by such groups as Orthoptera, Neuroptera, Thysanoptera, Diptera and Hymenoptera. The feeding preference of beetles, including longhorns, for pollen and perianth parts, rather than for nectar, is

Orchid pollination by longhorn beetles 295
considered a primitive trait. They are not well-adapted morphologically (cf. Aculeata and Lepidoptera) nor is there a narrow specialization for pollinating only specific species or genera of plants. On the other hand the example given above of A. tabacicolor pollinating D.fuchsii indicates a fairly advanced specialization on the part of longhorn beetles. The pollination of orchids--a relatively young group phylogenetically--is a complicated process and is commonly given as an example of the most advanced specialization of insect and flower symbiosis (e.g. Pervukhina, 1970).
Geographical distribution of D. fuchsii and A. tabacicolor
Since there is a certain dependence between both species, it is useful to compare their geographical location. The distribution centre of the genus Dactylorhiza Necker ex Nevski is in Asia Minor. However, D. fuchsii is distributed in Western Europe through Central and Eastern Europe even as far as Mongolia and Eastern Siberia (Proch/~zka & Velisek, 1983). The distribution centre of the genus Alosterna Muls. was probably also in the Caucasian region and northern parts of Asia Minor. Alosterna tabacicolor can be found in Europe, Caucasus, Trans-Caucasus, northern Turkey, northern Iran, Siberia, northern Mongolia, northern China, Korea and Japan ,(Danilevskii & Miroshnikov, 1985). Therefore, the distribution of both ecologically related species is similar and it is likely that pollination of an orchid by Alosterna tabacicolor beetles may also take place in the other parts of its range.
Preservation of orchids and their pollinators
Imagines of A. tabacicolor have many other plants on which to feed (mainly Umbelliferae). Therefore the disappearance or destruction of localities of this orchid does not significantly affect the population numbers of these longhorn beetles. However, the disappearance of A. tabacicolor near the localities studied may locally threaten the orchid. The disappearance of these longhorn beetles is already occurring in areas with intensive forest management where there is a lack of large deciduous trees. The larvae of this species of beetle develop in the outer bark of very large, live trees--trees that are too young, with thin bark, are not able to provide the proper environment for the development of the immature stages. However, where there are mature deciduous stands (e.g. the Bialowie/~a National Park) this beetle occurs in masses.
At present practical conservation most often concerns a single species as a unit, particularly in the case of flowers and insects. Moreover, the importance of insects in pollination or dispersal of rare plants is very poorly

296 Jerzy M. Gutowski
recognized and both conservation biologists and managers seem little concerned about insects (see Ginsberg, 1987). Therefore, protection of the whole biocenoses within reserves of adequate size is the best strategy to try to maintain the high diversity of species and their relationships.
A C K N O W L E D G E M E N T S
I thank Mr Stanistaw Batazy for his help in unravelling the 'mysterious growths' (i.e. pollinia) on the bodies of A. tabacicolor. I also thank Mrs Bogumita J¢drzejewska for her help and valuable comments on the text. For their insect and plant determination the following persons are sincerely thanked: M. Jagietto (orchids), B. Soszyfiski (Diptera), A. Herczek and J. Gorczyca (Heteroptera) and L. Buchholz (Elateridae). Thanks also go to M. Waszkiewicz for drawing the figures and M. Androsiuk who did some of the technical work.
REFERENCES
Borg-Karlson, A. K. (1987). Chemical basis for the relationship between Ophrys orchids and their pollinators, III. Volatile compounds of species in the Ophrys sections Fuciflorae and Bombyliflorae as insect mimetic attractants/excitants. Chem. Scripta, 27, 313-25.
Danilevskii, M. L. & Miroshnikov, A. I. (1985). Zhuki-drovoseki Kavkaza (Coleoptera, Cerambycidae) Opredelitel. Krasnodar.
Diels, L. (1916). K/iferblumen bei den Ranales und ihre Bedeutung fiir die Phylogenie der Angiospermen. Ber. dt. Bot. Ges., 34, 69.
Faegri, K. & Pijl, L. (1982). Osnovy ekologii opyleniya (The Principles of Pollination Ecology). Mir, Moskva [translation].
Ford, E. B. (1967). Genetyka ekologiczna. PWRiL, Warszawa. Ginsberg, J. R. (1987). What is conservation biology? Tree, 2(9), 262-4. Grinfeld, E. K. (1975). Antofiliya zhukov (Coleoptera) i kritika kantarofil'noi
gipotezy. Ent. Obozrenie, 54, 507-14. Hagerup, O. (1951). Pollination in the Faroes--in spite of rain and poverty in
insects. Dan. Biol. Medd., 18, 15. Halvorsen, R. (1986). Hanner av gravevepsslekten Argogorytes pa flueblomst
(Ophrys insectifera) i Telemark. Blyttia, 44, 53-6. Kevan, P. G. & Baker, H. G. (1983). Insects as flower visitors and pollinators. Ann.
Rev. Ent., 28, 407-53. Kullenberg, B. (1961). Studies in Ophrys,pollination. Zool. Bid., 34, 1-340. Nilsson, L. A. (1979). The pollination ecology ofHerminium monorchis (Orchidaceae).
Bot. Not., 132, 537-49. Nilsson, L. A. (1980). The pollination ecology of Dactylorhiza sambucina
(Orchidaceae). Bot. Not., 133, 367-85. Olberg, G. (1951). Blfite and lnsekt. Neue Brehm-Biicherei, Leipzig.

Orchid pollination by longhorn beetles 297
Pervukhina, N. V. (1970). Problemy morfologii i biologii tsvetka. Izdaterstvo 'Nauka', Leningrad.
Pijl, L. & Dodson, C. H. (1966). Orchid Flowers, Their Pollination and Evolution. Coral Gables, Florida.
Proch~zka, F. & Velisek, V. (1983). Orchideje nagi p~irody. (~SAV, Praha. Sil6n, F. (1906a), Blombiologiska iakttagelser i Kittil~i Lappmark. Meddel. Soc.
Fauna Flora Fenn., 31, 80-99. Sil6n, F. (1906b). Blombiologiska iakttagelser i s6dra Finland. Meddel. Soc. Fauna
Flora Fenn., 32, 120-34. Sundermann, H. (1975). Europiiische und mediterrane Orchideen. Eine Bestimmungs-
flora mit Beriicksichtigung der Okologie, 2nd edn. Briicke Verlag, K. Schmersow, Hildesheim.




















![Primeval and Other Times [excerpt]](https://img.dokumen.tips/doc/110x75/5468f203af795974338b5ef6/primeval-and-other-times-excerpt.jpg)








