Embed Size (px)
Citation preview

Journal of Ecology
2001
89
, 1054–1060
© 2001 British Ecological Society
Blackwell Science Ltd
Plant traits and temporal scale: evidence from a 5-year invasion experiment using native species
KEN THOMPSON, JOHN G. HODGSON, J. PHILIP GRIME and MICHAEL J. W. BURKE*
Department of Animal and Plant Sciences, The University, Sheffield S10 2TN, UK; and
*
English Nature, Northminster House, Peterborough PE1 1UA, UK
Summary
1
A long-term experiment was established in 1990 in which seeds of 54 native species, notoriginally present at the site, were sown into a fertility
×
disturbance matrix establishedin unproductive limestone grassland at the Buxton Climate Change Impacts Laboratory(BCCIL). The objective was to examine the roles of productivity and disturbance asmajor factors controlling the invasibility of plant communities, and to identify the func-tional characteristics of successful invaders in response to different types of invasionopportunity. The results of the first 2 years of the study have already been published.
2
After 2 years, invasion was strongly promoted by disturbance and less so by increasedfertility. Three years later the cover of invaders had declined over most of the matrix,and the greatest cover of sown species was where the highest levels of fertility and dis-turbance coincided. However, no part of the fertility–disturbance matrix was immuneto invasion and the area of the matrix occupied by each of the sown species that suc-cessfully established was unique. Abundance of invaders was reduced by low soil pH.
3
The identity and distribution of the successful invaders changed as the early stages ofinvasion gave way to a later stage of consolidation. After 2 years regenerative traits (seedmass and germination characteristics) were the best predictors of success. After 5 yearsthese traits were unrelated to success of the invaders, the most successful invaders wereperennial grasses, and no single trait was a good predictor of invasiveness.
4
Our results are consistent with the hypothesis that invasions are promoted by anincrease in the availability of resources, either through addition of extra resources or areduction in their use by the resident vegetation.
Key-words
: BCCIL, disturbance, fertility, functional types, herbivory, invasibility,limestone grassland, plant traits, seed size
Journal of Ecology
(2001)
89
, 1054–1060
Introduction
The past 10–15 years have seen an unprecedentedinterest in biological invasions, partly deriving froman increasing realization that, against a backgroundof increasing human population and global climatechange, invasive species are likely to become one of thekey environmental problems of the coming century.Much of this work has centred on attempts to identifythe key ecological characteristics of invasive speciesand potentially invasible ecosystems (Crawley 1986,1987; Usher 1988; Roy 1990; Lodge 1993; Williamson
1996; Levine & D’Antonio 1999; Lonsdale 1999).However, almost all this effort has been expended onobservations of invasions in natural systems (e.g. casestudies in de Waal
et al
. 1994; Py
s
ek
et al
. 1995; Brock
et al
. 1997). There have been few attempts to studyinvasion experimentally, by manipulating either thecharacteristics of the invaded community or the iden-tity of potential invaders (Peart & Foin 1985; Robinson
et al
. 1995; Bastl
et al
. 1997; Tilman 1997; Crawley
et al
. 1999; Knops
et al
. 1999; Levine 2000). Still morerarely have such experimental manipulations been con-ducted for relatively long periods of time. Experimentalstudies, including the one reported here, have focusedon grasslands and other herbaceous communities,since the time scales involved in woody plant com-munities are generally too long. Although importantinsights have begun to emerge from the analysis of the
Correspondence: Ken Thompson, Department of Animaland Plant Sciences, The University, Sheffield S10 2TN,UK (tel. 0114 2224314; fax 0114 2220015; [email protected]).
JEC_627.fm Page 1054 Wednesday, November 21, 2001 9:10 AM

1055
Plant traits and temporal scale
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1054–1060
large body of accumulated observational data (Lonsdale1999; Levine & D’Antonio 1999), important questionsremain unanswered and perhaps unanswerable by suchan approach. It seems likely that further progress willcome from experimental manipulation of disturbance,fertility, climatic variables, herbivory and other factorsthat might determine the outcome of invasions.
In September 1990, we established an experimentwith the objective of examining the roles of productivityand disturbance as major controlling factors of theinvasibility of plant communities, and to identify thefunctional characteristics of successful invaders inresponse to different types of invasion opportunity.The results of the first 2 years of this study have alreadybeen published (Burke & Grime 1996); here we reporton the changes that occurred in the next 3 years. Theexperiment continues.
Methods
The site and methods are described in detail in Burke &Grime (1996). Briefly, the experiment was located at theBuxton Climate Change Impacts Laboratory (BCCIL),near Buxton, Derbyshire, UK. The vegetation wasunproductive species-rich grassland, previously sheeppasture but now derelict. For more on the site, seehttp://www.shef.ac.uk/~nuocpe/bccil/.
In September 1990, 36 2
×
2 m plots were markedout within the established vegetation, in a 6 by 6square with 1-m paths between plots. Within each plot,orthogonally opposed five-step gradients of soil fertilityand physical disturbance resulted in a matrix withineach plot of 25 subplot combinations of productivityand disturbance, each 30
×
30 cm. The plot matrix wasrandomly allocated to one of eight possible orientations.The central 1.5
×
1.5 m area was marked as the vegeta-tion to be monitored, surrounded by an unmonitored25-cm ‘buffer’ zone that received the same treatment asthe adjacent subplot. Samples of the upper 5 cm of soilwere collected from every subplot in October 1992 onlyand the pH determined, after mixing with deionizedwater, with a Kent EIL 7055 pH meter calibrated withbuffers at pH 4 and 7.
Agricultural grade granular 20 : 10 : 10 NPK fertiliserwas applied at 0, 60, 120, 180 and 240 kg ha
–1
in October1990, April 1991 and October 1991, and subsequentlyin every March or April. The highest rate thereforecorresponds to 48 kg N, 10.5 kg P and 20 kg K ha
–1
, orapproximately one-third of the average rate applied toBritish agricultural grasslands in 1999 (Anonymous2000). Two methods were chosen to create the disturb-ance gradient: cutting gaps in the turf, and mowingthe vegetation at different heights, although only the 18plots from the gap-cutting treatment are consideredhere. Circular gaps were created within the turf andthe soil disturbed to a depth of approximately 50 mm.On the first occasion, all shoot and root material wasremoved from the gaps, but material was left
in situ
on subsequent occasions. Two gap sizes were used;
a ‘small’ gap that measures 49 mm in diameter (area1885 mm
2
), and a ‘large’ gap, six times the area of thesmall gap measuring 119 mm in diameter (area11 122 mm
2
). These gaps mimic the small naturaldisturbances caused by the hooves of grazing animals,or the slightly larger gaps created by mole hills, thatthis grassland would normally experience. It wouldnot normally experience any larger-scale disturbance.
The disturbance gradient was achieved by increasinggap density within the subplots, from zero to 48 small(small gap treatment), 8 large (large gap treatment) or24 small and 4 large gaps (mixed gaps treatment) (sixplots per treatment). In each case, the highest gap treat-ment had the potential to create 100% bare ground, butas gaps were allowed to overlap, the maximum bareground created was usually rather less than this. Levelsof bare ground across the disturbance gradient wereidentical in small, large and mixed gap treatments.Gaps were placed randomly within each subplot, andwere allowed to overlap each other and the borders ofthe subplot. All gap treatments were applied for thefirst time in October 1990 and have been repeated inOctober in every subsequent year.
The advantages of the matrix approach describedabove are: (i) the elimination of edge effects (even theedge of the matrix is bounded by a border receivingidentical treatment), as any treatment combination isadjacent to similar treatments; (ii) the continuousgradients allow plants to expand quickly from theirpoint of establishment to the part of the matrix mostconducive to their survival, thus reducing the chance ofrandom establishment failure determining the outcome;and (iii) the opportunity to create a wide range of envir-onments in a very compact space. A disadvantageis that disturbance and fertilizer treatments are notstatistically independent. For further discussion seeCampbell & Grime (1992).
A standard inoculum of 54 species, none originallypresent at the site, was sown into all plots in October1990. All species were common, widespread nativeBritish species found in the Sheffield area, in habitatswith climatic and altitudinal characteristics similar tothose of the experimental site. The choice included abroad range of ecological types with respect to bothregenerative and established stages of the life cycle.Large seeds were applied at lower densities than smallones. Nomenclature follows Stace (1997).
Plots were censused by placing a 1.5
×
1.5 m quadratover the centre of the plot. This quadrat was subdi-vided into a 5
×
5 grid to mark the positions of the25 subplots. At the first census in 1991, recording waslimited to presence of all species within each subplot,regardless of where it was rooted. In subsequent
JEC_627.fm Page 1055 Wednesday, November 21, 2001 9:10 AM
1056
K. Thompson
et al.
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1054–1060
censuses, in 1992, 1993 and 1995, the percentage coverof each species within each subplot was recorded, usingthe following scale: absent, 0; 1–5%, 1; 5–25%, 2; 25–50%, 3; 50–75%, 4; 76 + %, 5. All censuses took placeat the time of maximum plant biomass, in July, Augustor September.
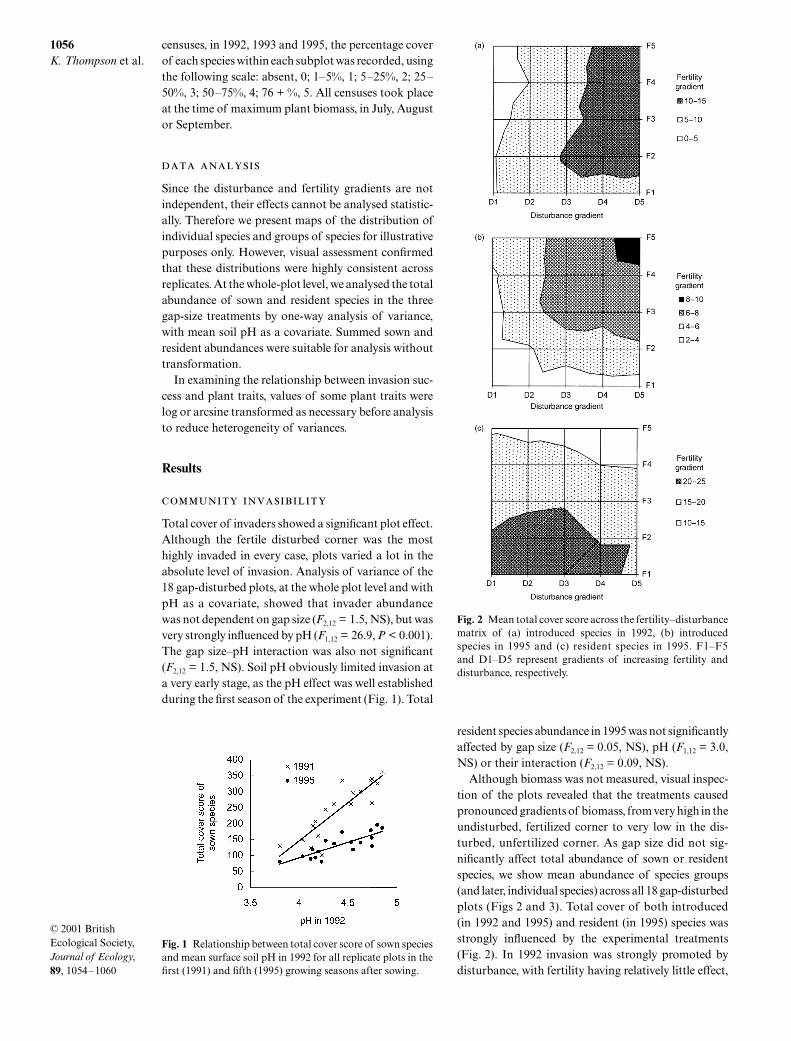
Since the disturbance and fertility gradients are notindependent, their effects cannot be analysed statistic-ally. Therefore we present maps of the distribution ofindividual species and groups of species for illustrativepurposes only. However, visual assessment confirmedthat these distributions were highly consistent acrossreplicates. At the whole-plot level, we analysed the totalabundance of sown and resident species in the threegap-size treatments by one-way analysis of variance,with mean soil pH as a covariate. Summed sown andresident abundances were suitable for analysis withouttransformation.
In examining the relationship between invasion suc-cess and plant traits, values of some plant traits werelog or arcsine transformed as necessary before analysisto reduce heterogeneity of variances.
Results
Total cover of invaders showed a significant plot effect.Although the fertile disturbed corner was the mosthighly invaded in every case, plots varied a lot in theabsolute level of invasion. Analysis of variance of the18 gap-disturbed plots, at the whole plot level and withpH as a covariate, showed that invader abundancewas not dependent on gap size (
F
2,12
= 1.5, NS), but wasvery strongly influenced by pH (
F
1,12
= 26.9,
P
< 0.001).The gap size–pH interaction was also not significant(
F
2,12
= 1.5, NS). Soil pH obviously limited invasion ata very early stage, as the pH effect was well establishedduring the first season of the experiment (Fig. 1). Total
resident species abundance in 1995 was not significantlyaffected by gap size (
F
2,12
= 0.05, NS), pH (
F
1,12
= 3.0,NS) or their interaction (
F
2,12
= 0.09, NS).Although biomass was not measured, visual inspec-
tion of the plots revealed that the treatments causedpronounced gradients of biomass, from very high in theundisturbed, fertilized corner to very low in the dis-turbed, unfertilized corner. As gap size did not sig-nificantly affect total abundance of sown or residentspecies, we show mean abundance of species groups(and later, individual species) across all 18 gap-disturbedplots (Figs 2 and 3). Total cover of both introduced(in 1992 and 1995) and resident (in 1995) species wasstrongly influenced by the experimental treatments(Fig. 2). In 1992 invasion was strongly promoted bydisturbance, with fertility having relatively little effect,
Fig. 1 Relationship between total cover score of sown speciesand mean surface soil pH in 1992 for all replicate plots in thefirst (1991) and fifth (1995) growing seasons after sowing.
Fig. 2 Mean total cover score across the fertility–disturbancematrix of (a) introduced species in 1992, (b) introducedspecies in 1995 and (c) resident species in 1995. F1–F5and D1–D5 represent gradients of increasing fertility anddisturbance, respectively.
JEC_627.fm Page 1056 Wednesday, November 21, 2001 9:10 AM

1057
Plant traits and temporal scale
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1054–1060
particularly at low intensities of disturbance (Burke& Grime 1996; Fig. 2a). By 1995, disturbance andfertility appeared to have equally large positive effectson cover of sown species (Fig. 2b). Cover of residentspecies, on the other hand, was lowest at high fertility(Fig. 2c).
The overall pattern in Fig. 2(b) conceals consider-able variation between individual sown species. Thediversity of patterns of invasion of the sown species isillustrated in Fig. 3. Some species, generally of ratherfertile habitats, appeared to respond positively to bothincreased fertility and disturbance, including the mostabundant invader,
Holcus lanatus
. Others respondedpositively to disturbance (e.g.
Cynosurus cristatus
) butappeared to be indifferent to fertility, while others (e.g.
Arrhenatherum elatius
) responded only to fertility. Afew sown species responded negatively to disturbance,fertility or both (e.g.
Vicia cracca
and
Brachypodiumpinnatum
). Finally
Taraxacum officinale
appeared tobe relatively indifferent to both treatments and waspresent throughout the matrix, with a broad peakunder conditions of low to moderate disturbance andfertility. Note the similarity of these distributions tothose in Buckland
et al
. (2001). The strong associationbetween total cover of invaders and high levels of fer-tility and disturbance (Fig. 2b) was the net effect ofthese individual distributions.
The dominant indigenous species in 1992 remained themost abundant in 1995. In contrast, ranking of intro-duced species in 1992 was a poor predictor of rankingin 1995 (
r
2
= 0.4). If the criterion of success of aninvader is an ability not only to persist, but also toincrease in cover over a period of years, then most ofthe introduced species were failures, at least in themedium term. Over most of the matrix, the abundanceof introduced species declined between 1992 and 1995(Figs 1 and 2), and only eight species increased in meancover during this period. All were perennial grasses, mostnotably
Dactylis glomerata
and
Alopecurus pratensis
.Correlation of change in cover of introduced species
between 1992 and 1995, across all treatments, with arange of plant traits involving seed size, current status,life history, ecological strategy, habitat, germination,morphology and growth, showed no significant rela-tionships (Table 1). One of the most striking results ofthe analysis of the first 2 years of the experiment wasclear evidence of the role of large seed size in facilitatinginvasion of dense turf, i.e. in the fertile, undisturbedquadrant (Burke & Grime 1996). Ability to increase inabundance under these conditions between 1992 and1995 was no longer related to seed size (
r
2
= 0.018,
P
= 0.27).
Fig. 3 Summed cover scores of six sown species across the fertility–disturbance matrix in 1995. F1–F5 and D1–D5 representgradients of increasing fertility and disturbance, respectively.
JEC_627.fm Page 1057 Wednesday, November 21, 2001 9:10 AM

1058
K. Thompson
et al.
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1054–1060
Discussion
One of the principal obstacles to increased understandingof invasions is that success is influenced by threefactors: the number of propagules (propagule pressure);the characteristics of the invading species; and thesusceptibility of the environment to invasion (Crawley
et al
. 1996; Williamson & Fitter 1996; Williamson 1996;Lonsdale 1999). In this case, all species were sown inlarge numbers on a single occasion, and therefore prop-agule pressure was uniform across all treatments. Wecan also discount differences in the identities of theinvaders available to different parts of the matrix, as allparts of every matrix were sown with the same widetaxonomic and functional range of species. The observedpattern of invasion is therefore a direct quantificationof susceptibility to invasion (invasibility). Furthermore,
many
post hoc
investigations of natural invasions arehampered by lack of knowledge of the identities of theunsuccessful invaders, i.e. species that were introducedbut never became established, and thus remained un-detected. Here we had complete knowledge of all thesown species, and so we were able to attempt to relateinvasion success to plant traits.
There are those who would argue that invasions byexotic species are a distinct ecological phenomenon,quite separate from the normal processes of regenera-tion, colonization and succession by native species(Elton 1958; Dukes & Mooney 1999). However, detailedstudy of the traits of exotic invaders and native colon-izers in NW Europe showed that the two groups wereessentially indistinguishable (Thompson
et al
. 1995).We would therefore argue that the processes that facil-itate invasion by exotic plant species and colonizationsby native species are fundamentally the same. This viewhas also been adopted by other workers (Huston 1994;Levine & D’Antonio 1999; Davis
et al
. 2000), includ-ing many of those who have undertaken experimentalstudies of invasions (Robinson
et al
. 1995; Tilman 1997;Crawley
et al
. 1999; Knops
et al
. 1999).
By 1992, at the time of the earlier analysis (Burke &Grime 1996), most individuals of the introduced spe-cies were still juveniles. Their distribution was stillpredominantly a function of the response of the initialinoculum to the opportunities for establishment affordedby the experimental treatments. Not surprisingly, thedistribution of invaders was mainly dependent ondisturbance, i.e. the creation of bare ground and debil-itation of the pre-existing vegetation (Fig. 2a), andsuccess or failure were largely explicable in terms ofregenerative traits, e.g. seed size and germination char-acteristics (Burke & Grime 1996).
Over the following 3 years, the focus of invasionshifted towards the more fertile parts of the matrix anddisturbance alone appeared to become less important(Fig. 2b). This seems to mark a transition from theearly stages of invasion (seed germination and seedlingestablishment) to a later stage of consolidation. Asmany of the species successful in this second stage wererobust, perennial grasses of fertile habitats, it is not sur-prising that high fertility became more important. Theincreasing importance of high fertility is probably anamalgamation of two effects. First, the time requiredfor large perennial invaders to accumulate a large bio-mass. Secondly, the sustained accumulation of mineralnutrients in the initially infertile soil. The early findingthat invasion was promoted more by small than bylarge gaps (Burke & Grime 1996) was no longer sup-ported after 5 years.
Overall, the results are consistent with the hypothe-sis, proposed by Davis
et al
. (2000), that invasibility iscorrelated with the availability of unused resources.Both disturbance and fertilizer addition increase the
Table 1 Pearson correlation coefficients between change incover of introduced species between 1992 and 1995 andselected plant traits
r n
Seed mass (mg)1 0.059 39Current status3 0.254 39Minimum temperature (°C) required for
50% germination1–0.220 34
Maximum temperature (°C) required for 50% germination1
0.070 34
Temperature range (°C) for 50% germination (i.e. max.–min.)1
0.162 34
Life history2 –0.005 39Canopy height class2 0.106 39Lateral spread class2 0.105 39Association with grassland2 –0.175 39Number of days required for 50%
germination in the light10.104 31
Germination percentage in the light1 –0.072 31Germination percentage in the dark1 –0.108 31Difference between germination
percentage in light and dark10.064 31
Rmax (week–1)4 0.090 27S score5 –0.019 39Competitive ability6 0.404 24Specialism index7 –0.099 39Palatability index8 –0.520 19
1Data from Grime et al. (1981). 2Data from Grime et al. (1988) and Hodgson et al. (1995). 3An index of current status based on recent patterns of increase or decrease in England, the Netherlands and western Germany, from Thompson et al. (1997). 4Maximum potential relative growth rate. Data from Grime & Hunt (1975). 5A measure of proximity to the S (stress-tolerant) corner of the CSR strategic triangle, calculated according to the protocol in Hodgson et al. (1999). 6A dominance index derived from extensive vegetation surveys, calculated as described in Hodgson et al. (1999). 7A measure of the range of habitats utilized by a species (high values = high specialization), calculated as described in Thompson et al. (1998). 8An index of palatability to the generalist mollusc herbivore Helix aspersa. Data from Grime et al. (1996). None of these correlations is significant at the Bonferroni-adjusted P-value of 0.05/18 = 0.003.
JEC_627.fm Page 1058 Wednesday, November 21, 2001 9:10 AM

1059
Plant traits and temporal scale
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1054–1060
availability of resources, and invasibility was clearlygreatest where both were combined. Bastl
et al
. (1997)found that, among a variety of treatments applied to aninfertile wet meadow, only large additions of manurepromoted the successful establishment of the invasiveannual
Impatiens glandulifera
. Nevertheless, invasibilityis predictable only in a statistical sense, as althoughincreased fertility and disturbance promoted invasionoverall, no part of the matrix was immune to invasion,and a minority of species (e.g.
Vicia cracca
and
Brach-ypodium pinnatum
, Fig. 3) did not conform to the generaltrend.
Although the experimental plots were established onlimestone, the high rainfall and low temperatures haveled to surface leaching and thus the pH of surface soilwas variable and sometimes very low. This had a majorimpact on invasion, with low pH clearly reducing theoverall level of invasion (Fig. 1). The effect of pH isexplicable in terms of the known tolerances of thepotential invaders; only one of the sown species (
Des-champsia flexuosa
) was a plant of acidic soils, and thegreat majority had pH optima
≥
6 (Grime
et al
. 1988).Increased soil acidity is often employed in habitat res-toration to inhibit invasion by weeds of neutral soils(e.g. Owen & Marrs 2000). pH clearly had a majorimpact on invasion at an early stage; later, the impactappeared to decrease (Fig. 1). This decrease could bedue to the pH being altered by fertiliser addition, or bythe later stages of invasion being less sensitive to pHthan seedling establishment. As pH was measured onlyonce, near the beginning of the experiment, it is notpossible to distinguish between these possibilities.
Although the factors promoting invasibility were clear(Fig. 2), our results confirm the well-known difficultyof predicting the identity of successful invaders (Crawley1986, 1987; Crawley
et al
. 1996; Williamson & Fitter1996; Williamson 1996), despite notable successes insome taxa, e.g.
Pinus
(Rejmánek & Richardson 1996).Among all the sown species, no single trait was signif-icantly associated with success (Table 1). Perhaps thisis not too surprising, given the variety of opportunitiesoffered by the experimental matrix and the individual-istic way in which those opportunities were exploitedby different species (Fig. 3). No doubt the distribution ofthe less-competitive invaders was strongly influenced bythe more-competitive species. Of all the traits examinedin Table 1, we suggest that palatability is worthy of furtherinvestigation, in the light of other evidence that herbivoryis important in preventing invasion by palatable species(e.g.
Urtica dioica
) at this site (Fraser 1998).
The recent review by Lonsdale (1999) illustrates thedifficulty of drawing unambiguous conclusions from
the study of natural invasions. Successful invasions areinfluenced by the interaction between propagule avail-ability, ecosystem invasibility and the characteristics ofthe potential invaders. Usually all these are knownincompletely, if at all. The results reported here cannotbe compared with most of the generalizations dis-covered by Lonsdale that concern invasion at the largescale, i.e. the effects of latitude, biome and islands vs.mainland. Lonsdale and others (Planty-Tabacchi
et al
.1996; Stohlgren
et al
. 1999) found that species-richcommunities were more invaded, but our plots were allsimilar in this respect, at least initially. If the treatmentsinduced changes in the distribution and diversity ofindigenous species in different parts of the matrix, theimpacts of these changes on potential invaders wereinseparable from the effects of the treatments themselves.It is also possible that the success of some invaders wasinhibited by the growth of other, more competitiveinvaders, but again such effects cannot be distinguishedfrom direct treatment effects.
Other experimental studies of invasion are few andhave tended to focus on the role of species richness ofthe invaded community (Peart & Foin 1985; Robinson
et al
. 1995; Tilman 1997; Levine 2000). The results ofsuch studies have been contradictory; Peart & Foin(1985) and Robinson
et al
. (1995) added seeds of fivegrass species and
Eschscholzia californica
, respectively,to Californian grasslands and found in each case thatthe most diverse patches were most invasible. The leastdiverse plots at Cedar Creek are the most invasible(Tilman 1997; Knops
et al
. 1999), but Crawley
et al
.(1999) have attributed a similar result to the ‘samplingeffect’. The most likely explanation for such contradic-tions is that invaders are responding to environmentalvariables that often covary with species richness, ratherthan to species richness itself (Levine & D’Antonio1999; Levine 2000). At variance with our results, andconsistent with the hypothesis that the identity of suc-cessful invaders depends strongly on the invaded envir-onment, Tilman (1997) found that perennial grasses werethe least successful invaders in the strongly nitrogen-limited system at Cedar Creek. Here N-fixing legumeswere the most successful invaders.
Acknowledgements
This work was supported by the Natural EnvironmentResearch Council. Thanks to past and present mem-bers of UCPE, Peter Mitchell for statistical advice,Jerry Tallowin for fertiliser data and to Mark William-son and several referees for much helpful advice.
References
Anonymous (2000)
The British Survey of Fertiliser Practice.Fertiliser Use on Farm Crops for Crop Year 1999
. The BSFPAuthority, The Fertiliser Manufacturers’ Association,Peterborough.
Bastl, M., Kocár, P., Prach, K. & Py
s
ek, P. (1997) The effect ofsuccessional age and disturbance on the establishment of
JEC_627.fm Page 1059 Wednesday, November 21, 2001 9:10 AM

1060
K. Thompson
et al.
© 2001 British Ecological Society,
Journal of Ecology
,
89
, 1054–1060
alien plants in man-made sites: an experimental approach.
Plant Invasions: Studies from North America and Europe
(eds J.H. Brock, M. Wade, P. Py
s
ek & D. Green), pp. 191–201.Backhuys, Leiden.
Brock, J.H., Wade, M., Py
s
ek, P. & Green, D. (1997)
PlantInvasions: Studies from North America and Europe
. Back-huys, Leiden.
Buckland, S.M., Thompson, K., Hodgson, J.G. & Grime, J.P.(2001) Grassland invasions: effects of manipulations ofclimate and management.
Journal of Applied Ecology
,
38,301–309.
Burke, M.J.W. & Grime, J.P. (1996) An experimental study ofplant community invasibility. Ecology, 77, 776–790.
Campbell, B.D. & Grime, J.P. (1992) An experimental test ofplant strategy theory. Ecology, 73, 15–29.
Crawley, M.J. (1986) The population biology of invaders.Philosophical Transactions of the Royal Society of London.B Biological Sciences, 314, 711–731.
Crawley, M.J. (1987) What makes a community invasible?Colonization, Succession and Stability (eds A.J. Gray, M.J.Crawley & P.J. Edwards), pp. 429–453. Blackwell, Oxford.
Crawley, M.J., Brown, S.L., Heard, M.S. & Edwards, G.R.(1999) Invasion-resistance in experimental grasslandcommunities: species richness or species identity? EcologyLetters, 2, 140–148.
Crawley, M.J., Harvey, P.H. & Purvis, A. (1996) Comparativeecology of the native and alien floras of the British Isles.Philosophical Transactions of the Royal Society of London.B Biological Sciences, 351, 1251–1259.
Davis, M.A., Grime, J.P. & Thompson, K. (2000) Fluctuatingresources in plant communities: a general theory of invas-ibility. Journal of Ecology, 88, 528–534.
Dukes, J.S. & Mooney, H.A. (1999) Does global changeincrease the success of biological invaders? Trends in Ecologyand Evolution, 14, 135–139.
Elton, C.S. (1958) The Ecology of Invasions by Animals andPlants. Methuen, London.
Fraser, L.H. (1998) Top-down vs. bottom-up control influ-enced by productivity in a North Derbyshire, UK, dale.Oikos, 81, 99–108.
Grime, J.P., Cornelissen, J.H.C., Thompson, K. & Hodgson,J.G. (1996) Evidence of a causal connection between anti-herbivore defence and the decomposition rate of leaves.Oikos, 77, 489–494.
Grime, J.P., Hodgson, J.G. & Hunt, R. (1988) ComparativePlant Ecology. Unwin Hyman, London.
Grime, J.P. & Hunt, R. (1975) Relative growth-rate: its rangeand adaptive significance in a local flora. Journal of Ecology,63, 393–422.
Grime, J.P., Mason, G., Curtis, A.V., Rodman, J., Band, S.R.,Mowforth, M.A.G., Neal, A.M. & Shaw, S. (1981) A com-parative study of germination characteristics in a localflora. Journal of Ecology, 69, 1017–1059.
Hodgson, J.G., Grime, J.P., Hunt, R. & Thompson, K. (1995)The Electronic Comparative Plant Ecology. Chapman &Hall, London.
Hodgson, J.G., Wilson, P.J., Hunt, R., Grime, J.P. & Thomp-son, K. (1999) Allocating C-S-R plant functional types: asoft approach to a hard problem. Oikos, 85, 282–296.
Huston, M.A. (1994) Biological Diversity. The Coexistence ofSpecies on Changing Landscapes. Cambridge UniversityPress, Cambridge.
Knops, J.M.H., Tilman, D., Haddad, N.M., Naeem, S.,Mitchell, C.E., Haarstad, J., Ritchie, M.E., Howe, K.M.,Reich, P.B., Siemann, E. & Groth, J. (1999) Effects of plantspecies richness on invasion dynamics, disease outbreaks,
insect abundances and diversity. Ecology Letters, 2, 286–293.
Levine, J.M. (2000) Species diversity and biological invasions:relating local process to community pattern. Science, 288,852–854.
Levine, J.M. & D’Antonio, C.M. (1999) Elton revisited: areview of evidence linking diversity and invasibility. Oikos,87, 15–26.
Lodge, D. (1993) Biological invasions: lessons for ecology.Trends in Ecology and Evolution, 8, 133–137.
Lonsdale, W.M. (1999) Global patterns of plant invasions andthe concept of invasibility. Ecology, 80, 1522–1536.
Owen, K.M. & Marrs, R.H. (2000) Creation of heathland onformer arable land at Minsmere, Suffolk, UK: the effectsof soil acidification on the establishment of Calluna andruderal species. Biological Conservation, 93, 9–18.
Peart, D.R. & Foin, T.C. (1985) Analysis and prediction ofpopulation and community change: a grassland case study.The Population Structure of Vegetation (ed. J. White),pp. 313–339. Junk, Dordrecht.
Planty-Tabacchi, A., Tabacchi, E., Naiman, R.J., Deferrari, C.& Décamps, H. (1996) Invasibility of species-rich commun-ities in riparian zones. Conservation Biology, 10, 598–607.
Pysek, P., Prach, K., Rejmánek, M. & Wade, P.M. (1995a)Plant Invasions: General Aspects and Special Problems. SPBAcademic Publishing, Amsterdam.
Rejmánek, M. & Richardson, D.M. (1996) What attributesmake some plant species more invasive? Ecology, 77, 1655–1661.
Robinson, G.R., Quinn, J.F. & Stanton, M.L. (1995) Invas-ibility of experimental habitat islands in a California winterannual grassland. Ecology, 76, 786–794.
Roy, J. (1990) In search of the characteristics of plant invaders.Biological Invasions in Europe and the Mediterranean (edsF. di Castri, A.J. Hansen & M. Debussche), pp. 335–352.Kluwer, Dordrecht.
Stace, C.A. (1997) New Flora of the British Isles, 2nd edn.Cambridge University Press, Cambridge.
Stohlgren, T.J., Binkley, D., Chong, G.W., Kalkhan, M.A.,Schell, L.D., Bull, K.A., Otsuki, Y., Newman, G., Bashkin,M. & Son, Y. (1999) Exotic plant species invade hot spots ofnative plant diversity. Ecological Monographs, 69, 25–46.
Thompson, K., Hodgson, J.G. & Gaston, K.J. (1998) Abun-dance–range size relationships in the herbaceous flora ofcentral England. Journal of Ecology, 86, 439–448.
Thompson, K., Hodgson, J.G. & Rich, T.C.G. (1995) Nativeand alien invasive plants: more of the same? Ecography, 18,390–402.
Thompson, K., Parkinson, J.A., Band, S.R. & Spencer, R.E.(1997) A comparative study of leaf nutrient concentrationsin a regional herbaceous flora. New Phytologist, 136, 679–689.
Tilman, D. (1997) Community invasibility, recruitment lim-itation, and grassland biodiversity. Ecology, 78, 81–92.
Usher, M.B. (1988) Biological invasions of nature reserves: asearch for generalisations. Biological Conservation, 44,119–135.
de Waal, L.C., Child, L.E., Wade, P.M. & Brock, J.H. (1994)Ecology and Management of Invasive Riverside Plants. Wiley,Chichester.
Williamson, M.H. (1996) Biological Invasions. Chapman &Hall, London.
Williamson, M.H. & Fitter, A. (1996) The characters of suc-cessful invaders. Biological Conservation, 78, 163–170.
Received 23 October 2000 revision accepted 3 July 2001
JEC_627.fm Page 1060 Wednesday, November 21, 2001 9:10 AM





























