Embed Size (px)
Citation preview

Genus Planadina gen. n.Type spe cies: Roundya plana Helms, 1959 from the Platyclymenia annulata Zone, Bohlen near Saalfeld.Der i va tion of name: Taken from the name of the type spe cies.
Di ag no sis. — Triramous in all S el e ments, M el e ment with strongly bent, rib bon−like ex ter nal pro cessand trans versely ori ented undenticulated in ner process.
Re marks. — This ge nus is un usual in that the denticulated and elon gated pro cess in its M el e ments is notthe in ner one, as in vir tu ally all other ozarkodinid and prioniodontid cono donts, but the outer one. Such a bi −zarre mor phol ogy can be ex plained if the an ces tor of Planadina was like Francodina franconica orSweetodina lagoviensis in this re spect, that is with both pro cesses elon gated and bear ing low and widedenticles. Es pe cially the denticulation of the rib bon−like pro cess in the lat ter spe cies is ap peal ing as a pos si −bly an ces tral state be cause it forms a kind of un du lat ing ridge. A mys te ri ous as pect of this ap pa ra tus is that no un doubted P se ries el e ments have been iden ti fied (ex cept for one ques tion able spec i men of small size; Fig.43A). A pos si bil ity emerges that P1–2 el e ments at tained the same shape as the M el e ments. In fact some ofthem re sem ble plat form se ries el e ments of Sweetodina (Fig. 43F).
Planadina plana (Helms, 1959)(Figs 43 and 126)
Type ho ri zon and lo cal ity: Bed 7 (“Wag ner Bank”) of Platyclymenia annulata Zone, Bohlen near Saalfeld (Helms 1959).
Ma te rial. — 396 spec i mens.Di ag no sis. — As for the ge nus.Re marks. — A sin gle el e ment of a rather ro bust ap pear ance as so ci ated with those of P. plana (Fig. 43A)
may or may not rep re sent P series.Oc cur rence. — Pos si bly the K. crepida Zone at Kadzielnia; wide spread from the C. marginifera to P.
jugosus zones at Łagów, Miedzianka, Ostrówka, Kowala, and Dzikowiec.
Family Polygnathidae Bassler, 1925Di ag no sis. — Triramous S0 el e ments and nar row basal cav ity of P1 el e ments with a ten dency to in ver −
sion, fre quently con nected with de vel op ment of a plat form; ap pa ra tus generalized.Re marks. — The Polygnathidae orig i nated at the be gin ning of the De vo nian from Ozarkodina, a mem −
ber of the Spathognathodontidae, by de vel op ing a me dial pro cess in S0 el e ments and elon ga tion of pro cessesin other S el e ments. Pandorinellina prob a bly rep re sents this first lin eage con tin ued with out sig nif i cantchanges to the Famennian.
Genus Pandorinellina Müller et Müller, 1957Type spe cies: Pandorina insita Stauffer, 1940 from the lat est Givetian Litho graph City For ma tion of Min ne sota (G.
Klapper, per sonal com mu ni ca tion 2005).
Di ag no sis. — P1 el e ments de vel op ing more or less ex panded basal cav ity with vir tu ally flat sur face, lack −ing plat form; rel a tively high−arched M el e ments, other el e ments of a gen er al ized polygnathid appearance.
Re marks. — Hindeodontoides of Rexroad and Merrill (1996), with the Viséan type spe cies H. spiculus,is an al ter na tive no men cla tor ial op tion for spe cies af fil i ated here in Pandorinellina. Dasbergina is rooted inthe Pandorinellina lin eage and the tran si tion from P. vulgaris to D. stabilis seems to be continuous.
Pandorinellina? vogelgnathoides sp. n.(Figs 44 and 125)
Holotype: Spec i men ZPAL cXVI/1912 (Fig. 44A).Type ho ri zon and lo cal ity: Sam ple Ka−5, early Famennian K. crepida Zone at Kadzielnia, Holy Cross Moun tains.Der i va tion of name: Re fer ring to sim i lar ity to Vogelgnathus.
Ma te rial. — 350 spec i mens.Di ag no sis. — Gently arched pro file of P1 el e ments at denticle tips, ro bust S el e ments with straight, rather
short pro cesses; high arched long pro cesses of M element.Re marks. — An al most com plete set of el e ments of the spe cies is of fered by sam ple J−68 from Jabłonna.
At first glance they are sim i lar to non−plat form el e ments of Polygnathus but no sin gle spec i men of this ge nus
70 JERZY DZIK

is rep re sented in the sam ple. Much more nu mer ous but high−di ver sity sam ples from Kadzielnia al low dis tin −guish ing prob a ble P1 and P2 el e ments. The ap pa ra tus some what re sem bles that of the francodinids and gen er −ally short pro cesses give it a paedomorphic ap pear ance. The spe cies can not be an ces tral to the firstfrancodinid, Vogelgnathus, be cause of the large size of el e ments, their straight pro cesses, and too young geo −log i cal age. Among S1 el e ments with short straight pro cesses at Kadzielnia there are spec i mens with an ad di −tional pro cess de vel op ing at the el e ment bend. This is an other francodinid fea ture and it can not be ex cludedthat P.? vogelgnathoides is a case of evolutionary reversal within the francodinids.
Oc cur rence. — The K. crepida Zone at Jabłonna and Kadzielnia.
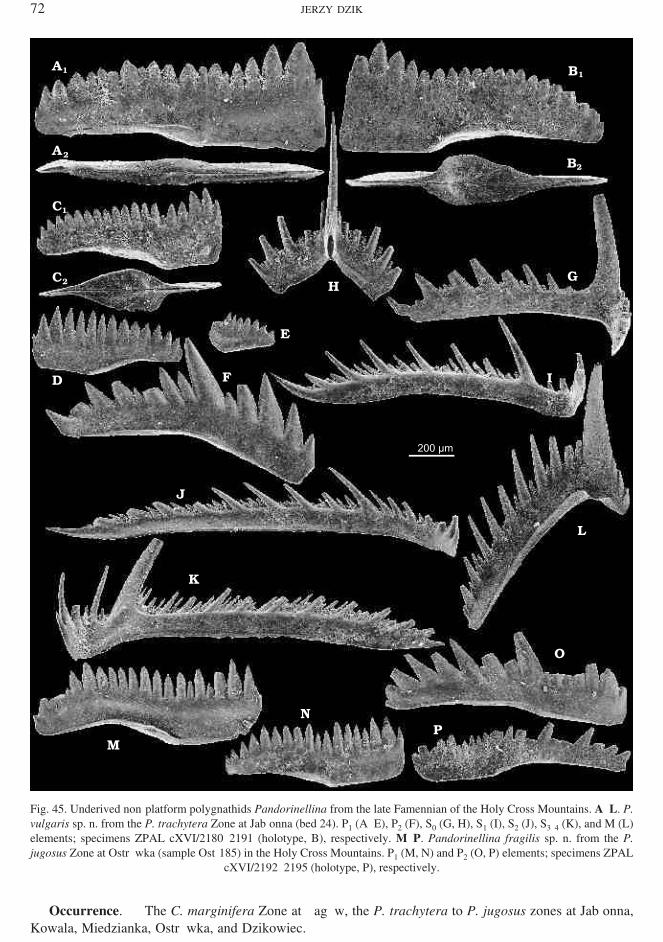
Pandorinellina vulgaris sp. n.(Figs 45A–L and 127)
Holotype: Spec i men ZPAL cXVI/2181 (Fig. 45B).Type ho ri zon and lo cal ity: Bed 24, mid Famennian P. trachytera Zone at Jabłonna, Holy Cross Moun tains.Der i va tion of name: Vulgaris, usual, re fer ring to its in dif fer ent ap pear ance and abun dance.
Ma te rial. — 3,244 spec i mens.Di ag no sis. — Sharp denticles of P1 el e ments of roughly sim i lar height, basal cone with a wil low leaf−like
con tour ex tend ing to about two thirds ot the dor sal pro cess, rest of the base narrow.Re marks. — The spe cies dif fers from the Frasnian P. insita in the more reg u lar dis tri bu tion of height of
denticles, which close to the dor sal tip are ar ranged in a gently con vex pro file. This char ac ter makes the spe ciestran si tional to the Protognathodus and Dasbergina lin eages. The as so ci ated Alternognathus at non−plat formontogenetic stages shows al ready a wide spac ing in dis tri bu tion of denticles in the dor sal part of the P1 element.
The rather large sam ple Ost−12 shows ma ture P1 el e ments with a ro bust ap pear ance, the blade be ingsome what thick ened be low denticles, but never de vel op ing a true plat form. The basal cav ity is very nar row.Ju ve nile spec i mens with their sharp denticles and flat ap pear ance some what re sem ble Mehlina, as well as ju −ve niles of the as so ci ated Mehlina. The main dif fer ence at this stage of on tog eny is in a denser and more ir reg −u lar denticulation in Mehlina transita, which also rel a tively early de vel oped an in cip i ent plat form and in −verted basal cav ity. Some P1 spec i mens of Branmehla may also re sem ble this spe cies but they show widerand shorter basal cavity and are more or less bent.
FAMENNIAN CONO DONTS AND AMMONOIDS 71
Fig. 44. Enigmathic non−plat form polygnathid with pos si ble francodinid af fin i ties Pandorinellina? vogelgnathoides sp. n. fromthe K. crepida Zone at Kadzielnia (A–G, sam ple Ka−5; H, sam ple Ka−3) in the Holy Cross Moun tains. P1 (A), P2 (B), S0 (C), S1 (D
and H), S2 (E), S3–4 (F), and M (G) el e ments; spec i mens ZPAL cXVI/1912–1918 (holotype, A), and 1903, re spec tively.
Oc cur rence. — The C. marginifera Zone at Łagów, the P. trachytera to P. jugosus zones at Jabłonna,Kowala, Miedzianka, Ostrówka, and Dzikowiec.
72 JERZY DZIK
Fig. 45. Un de rived non−plat form polygnathids Pandorinellina from the late Famennian of the Holy Cross Moun tains. A–L. P.vul garis sp. n. from the P. trachytera Zone at Jabłonna (bed 24). P1 (A–E), P2 (F), S0 (G, H), S1 (I), S2 (J), S3–4 (K), and M (L) elements; spec i mens ZPAL cXVI/2180–2191 (holotype, B), re spec tively. M–P. Pandorinellina fragilis sp. n. from the P.jugosus Zone at Ostrówka (sam ple Ost−185) in the Holy Cross Moun tains. P1 (M, N) and P2 (O, P) el e ments; spec i mens ZPAL
cXVI/2192–2195 (holotype, P), re spec tively.

Pandorinellina fragilis sp. n.(Figs 45M–P and 127)
Holotype: Spec i men ZPAL cXVI/2195 (Fig. 45P).Type ho ri zon and lo cal ity: Sam ple Ost−185, late Famennian P. jugosus Zone at Ostrówka, Holy Cross Moun tains.Der i va tion of name: Re fer ring to fra gil ity of spec i mens.
Ma te rial. — 219 spec i mens.Di ag no sis. — Ex tremely flat P2 el e ments with ir reg u lar denticulation.Re marks. — The spe cies is closely sim i lar to P. vulgaris and al most cer tainly is its suc ces sor. It is highly
vari able and sin gle spec i mens may not be spe cif i cally de ter mi na ble. These spe cies seem to oc cur allopatrically.Oc cur rence. — The late P. jugosus to early D. trigonica zones at Ostrówka.
Pandorinellina bituberculata sp. n.(Figs 46 and 127)
Holotype: Spec i men ZPAL cXVI/2199 (Fig. 46A).Type ho ri zon and lo cal ity: Sam ple J−51, late Famennian P. jugosus Zone at Jabłonna, Holy Cross Moun tains.Der i va tion of name: Re fer ring to two tu ber cles de vel op ing on the base.
Ma te rial. — 49 spec i mens.Di ag no sis. — Ro bust blade of P1 el e ments, ma ture el e ments tend to de velop large tu ber cle on each side
of the wid ened basal cone, which is subcircular in out line and re stricted to the mid−length of the blade.Re marks. — The spe cies is an other suc ces sor of P. vulgaris rep re sent ing the op po site to the ten dency in
P. fragilis to wards a ro bust ap pear ance of all el e ments. Ju ve niles of the an ces tor and suc ces sor are indin −stinguishable and can be at trib uted to its spe cies only on the ba sis of co−oc cur rence with adult spec i mens andgeo log i cal age. Be cause of this, the spe cies af fil i a tion of the late Famennian pop u la tions of Pandorinellinaknown ex clu sively from im ma ture spec i mens re mains ten ta tive. This is es pe cially trou ble some with theKowala sec tion, where acid treat ment of lime stone sam ples pro duces mostly ju ve nile spec i mens of Pandori −nellina, whereas wash ing of the in ter ca lat ing clay (e.g., Ko−131) sup plies ro bust ma ture spec i mens. From un −known rea son the pop u la tion dy nam ics of the spe cies de pended on whether limy mud or fine clay was de pos −
FAMENNIAN CONO DONTS AND AMMONOIDS 73
Fig. 46. Non−plat form polygnathid Pandorinellina bituberculata sp. n. from the P. jugosus Zone at Jabłonna (sam ple J−51) in theHoly Cross Moun tains. P1 (A–D), P2 (E), S2 (F), and S3–4 (G) el e ments; spec i mens ZPAL cXVI/2199–2205 (holotype, A),
respectively.

ited. Al though the tax on omy of the P. vulgaris−group is in con ve niently difficult to apply, the distinctionsbetween species seems rather well substantiated.
Oc cur rence. — The L. styriacus to P. jugosus zones at Jabłonna, Ostrówka, Kowala, and Dzikowiec.
Genus Mehlina Youngquist, 1945Type spe cies: Mehlina irregularis Youngquist, 1945 from the late Frasnian In de pend ence Shale (Amana Beds) of Iowa.
Di ag no sis. — Polygnathids with very flat P1 el e ments hav ing ex tremely nar row basal cav ity, in verted inlater on tog eny, lack ing plat form; M el e ments with rel a tively short and trans versely ar ranged inner process.
74 JERZY DZIK
Fig. 47. Non−plat form polygnathids Mehlina. A–R. M. kielcensis sp. n. from the K. triangularis Zone at Płucki (A–G, sam plePł−1) and Jabłonna (H, I, and J, sam ple J−53a), and the K. crepida Zone at Jabłonna (K–R, bed 3) in the Holy Cross Moun tains.P1 A–C, F, J–L, and N), P2 (D, H, I, and K), S0 (M), S1 (P), S2 (O), S3–4 (Q), and M (E, G, and R) el e ments; spec i mens ZPALcXVI/2080–2082 (holotype, A), 2084, 2085, 2083, 2086, 2088, 2089, 2087, 2095, 2090, 2093, 2091, 2095, 2094, 2096, and2097, re spec tively. S–AA. Mehlina robustidentata sp. n. from the K. crepida Zone at Jabłonna (S and BB, bed 8; U–AA, sam pleJ−45a) in the Holy Cross Moun tains. P1 (S, T), P2 (U), S0 (V), S1 (W), S2 (X), S3–4 (Y), and M (Z and AA) el e ments; spec i mens
ZPAL cXVI/2098 (holotype, S), 2100–2106, and 2099, re spec tively.

Re marks. — The morphologic dis tinc tion be tween Pandorinellina and Mehlina is far from be ing ap par −ent and it is fur ther oblit er ated by their pop u la tion vari abil ity. Nev er the less, these are branches of plat −form−lack ing polygnathids sep a rate al ready in the Frasnian and through out their strati graphic oc cur rence giv −ing rise to sev eral lin eages of cono donts with plat form− or icrion−bear ing P1 el e ments. Even if prac ti cal ap pli −ca tion of their tax on omy may cause dif fi cul ties, it is rea son able to dis tin guish them at the ge neric levelbecause of phylogentic consequences.
Mehlina kielcensis sp. n.(Figs 47A–R and 127)
Holotype: Spec i men ZPAL cXVI/2080 (Fig. 47A).Type ho ri zon and lo cal ity: Sam ple Pł−1, early Famennian P. triangularis Zone at Płucki, Holy Cross Moun tains.Der i va tion of name: From latinized name of Kielce, the cap i tal of the Holy Cross Moun tains re gion.
Ma te rial. — 527 spec i mens.Di ag no sis. — Ma ture P1 with rel a tively few (10–15) denticles, all el e ments of gen er al ized mor phol ogy.Re marks. — This is the old est mem ber of the Famennian lin eage of Mehlina. From its prob a ble an ces tor
M. irregularis, it dif fers in hav ing more ro bust and less nu mer ous denticles in P1 el e ments; the lat ter char ac −ter makes it dif fer ent from the suc ces sor, M. robustidentata sp. n. De spite a wide pop u la tion vari abil ity, sam −ples con tain ing these spe cies are rel a tively eas ily dis tin guish able on the ba sis of P1 el e ments. A pos si bly re −lated lin eage is rep re sented by Vogelgnathus variabilis (the old est mem ber of the Francodinidae; Fig. 125),dif fer ent in a paedomorphic appearance and small size of elements.
Oc cur rence. — From the be gin ning of Famennian to the early K. crepida at Płucki, Jabłonna andKadzielnia.
Mehlina robustidentata sp. n.(Figs 47S–AA and 127)
Holotype: Spec i men ZPAL cXVI/2098 (Fig. 47S).Type ho ri zon and lo cal ity: Bed 8, early Famennian K. crepida Zone at Jabłonna, Holy Cross Moun tains.Der i va tion of name: Re fer ring to ro bust ap pear ance of denticulation of the P1 el e ment.
Ma te rial. — 426 spec i mens.Di ag no sis. — P1 el e ment with nu mer ous (15–20) ro bust denticles and long blade.Re marks. — The spe cies shows some sim i lar i ties to Alternognathus pseudostrigosus from slightly youn −
ger strata (the old est mem ber of the Cavusgnathidae; Fig. 137) and may be its an ces tor. The change would in −volve de vel op ment of a some what ir reg u lar plat form in the P1 element.
Oc cur rence. — The late K. crepida to C. marginifera zones at Jabłonna and Łagów.
Mehlina strigosa (Branson et Mehl, 1934)(Figs 48, 49D, E?, and 127)
Type ho ri zon and lo cal ity: Famennian at Dixie, Mis souri (Ziegler 1977, p. 313).
Ma te rial. — 2,050 spec i mens.Di ag no sis. — P1 el e ment with nu mer ous (15–30) sharp−pointed denticles.Re marks. — This is a highly vari able spe cies. From its pre de ces sor, M. robustidentata, it dif fers not only
in the pat tern of denticulation but also in a ten dency to de velop some what wid ened flat basal cone in the P1el e ment. In this re spect it is some what sim i lar to Dasbergina stabilis, from which it dif fers in the much higher sharp denticles on the dor sal pro cess and, of course, the rest of the ap pa ra tus. More over, the wid ened basalcone of Dasbergina ex tends to the dor sal tip of the el e ment. In P1 el e ments of Pandorinellina vulgaris thebasal cone is re stricted to mid−length and is wide. Some ex treme el e ments may be dif fi cult to at trib ute to par −tic u lar spe cies of these gen era. In fact only P1 and M el e ments are diagnostic at the generic level.
S1 el e ments with ad di tional pro cess (Fig. 48O) co−oc cur ring with other el e ments of the spe cies are ten ta −tively at trib uted to it, but sim i lar el e ments are also as so ci ated with Dasbergina micropunctata (Fig. 138).This is sue re mains to be clarified.
Oc cur rence. — Wide spread in the late C. marginifera to P. jugosus zones of the Holy Cross Moun tains.
FAMENNIAN CONO DONTS AND AMMONOIDS 75

Mehlina lunaria sp. n.(Figs 49A, B and 127)
Holotype: Spec i men ZPAL cXVI/2125 (Fig. 49A).Type ho ri zon and lo cal ity: Sam ple Ka−2, early Famennian K. crepida Zone at Kadzielnia, Holy Cross Moun tains.Der i va tion of name: Re fer ring to semilunar out line of the el e ment.
Ma te rial. — Two spec i mens.Di ag no sis. — Whole P1 el e ment trans formed into elab o rate peniculus built of bi fur cat ing lat eral pro −
cesses ra di at ing from the cen ter of element.Re marks. — This is a com monly oc cur ring spe cies with bi zarre el e ments. Sannemann (1955a, pl. 1:10)
re ported it from the K. crepida Zone of the Frankenwald, Wolska (1967) from co eval strata at Jabłonna in theHoly Cross Moun tains. At first glance it does not re sem ble a cono dont at all. When seen from the base (Fig.49A3, B2) it shows bi fur ca tion of lat eral pro cesses that re sulted in de vel op ing so un usual pat tern of thepeniculus. Also the ba sally pointed out line in lat eral view is con sis tent with the characterictic ap pear ance ofP1 el e ments of Mehlina. A peniculus de vel oped widely in other mem bers of the Mehlina–Polynodosusbranch (Figs 47I, 48C, F, 49C–E, and 50A, G), as well as in the pos si bly re lated Frasnian Ancyrodella
76 JERZY DZIK
Fig. 48. Non−plat form polygnathids Mehlina strigosa (Branson et Mehl, 1934) from the P. trachytera Zone at Kowala (A–C,sam ple Ko−15) and the P. jugosus Zone at Ostrówka (D–I, sam ple Ost−185) and Jabłonna (J–R, bed 27) in the Holy Cross Moun −tains. P1 (A, B, DF, and J–L), P2 (C, G, M, N), S0 (M), S1 (M and triramous O), S2 (P), S3–4 (Q), and M (I and R) el e ments; spec i −
mens ZPAL cXVI/2107–2124, re spec tively.

(Lindström and Ziegler 1965) and the dif fer ence be tween them and M. lunaria is ac tu ally not great. An al −most com plete gra da tion can be as sem bled. The rel a tively sta ble mor phol ogy of P1 el e ments of M. lunariasp. n. (shown by both mine and ear lier pub lished spec i mens) and the morphologic gap be tween its mor phol −ogy and vari abil ity ex pressed by other known peniculi indicates that this is a separate species.
Oc cur rence. — The K. crepida Zone at Kadzielnia.
Mehlina sudetica sp. n.(Figs 49F–L and 127)
Holotype: Spec i men ZPAL cXVI/2983 (Fig. 49H).Type ho ri zon and lo cal ity: Sam ple Dz−54, late Famennian P. jugosus Zone at Dzikowiec, Sudetes.Der i va tion of name: From the Sudetes, where the type pop u la tion oc curs.
Ma te rial. — 285 spec i mens.Di ag no sis. — P1 el e ment with rel a tively short blade armed with nu mer ous sharp but rel a tively ro bust
denticles with a ten dency to de velop peniculus at late ontogenetic stages.Re marks. — This is clearly a close rel a tive and suc ces sor of M. strigosa. Co−oc cur rence of triramous S1
el e ments may also be an ex pres sion of prox im ity to that spe cies. The main dif fer ence is in P se ries el e ments,with P1 el e ment be ing more com pact and ro bust than in M. strigosa, whereas in P2 el e ments the ten dency tode velop peniculus seems to be not rep re sented (not all spec i mens of M. strigosa show it).
Oc cur rence. — The P. jugosus and D. trigonica zones at Dzikowiec.
Genus Polynodosus Vorontzova, 1993Type spe cies: Polygnathus nodocostata Branson et Mehl, 1934 from the Famennian at Dixie or Noel (not spec i fied) of Mis −
souri (Branson and Mehl 1934, Helms 1961).
Di ag no sis. — Plat form of P1 el e ments with tu ber cles tend ing to be ar ranged in ridges par al lel to theblade, other el e ments ro bust, sim i lar to those of Mehlina.
Re marks. — That Polynodosus is a de riv a tive of Mehlina is sug gested not only by the sim i lar ity of theirap pa ra tuses, in fact quite gen er al ized, but also by the com mon de vel op ment of a peniculus in at least somespe cies (P. nodoundatus and P. diversus). There are at least four sep a rate lin eages within the ge nus. The least de rived is rep re sented by P. transitus and P. lauriformis, with rel a tively small and sim ple plat form. In the lin −
FAMENNIAN CONO DONTS AND AMMONOIDS 77
Fig. 49. Spe cies of Mehlina bear ing peniculus at adult stages and patho log i cal polygnathid el e ments. A, B. Mehlina lunaria sp. n. from the K. crepida Zone at Kadzielnia (sam ple Ka−2). P1 el e ments; spec i mens ZPAL cXVI/2125, 2126 (holotype, A). C–E. Ex −treme icrion−bear ing el e ments of Polynodosus transitus sp. n. from the P. trachytera Zone at Jabłonna (C, bed 24) and Mehlinastri gosa from the C. quadrantinodosa Zone at Miedzianka (D, E, sam ple Md−2). P1 (D), P2 (C and E); spec i mens ZPALcXVI/2130, and 2178, 2179. F–L. Mehlina sudetica sp. n. from the P. jugosus Zone at Dzikowiec (F, L, sam ple Dz−10; G–K,sam ple Dz 54). P1 (F–H), P2 (I, J), S1 (K), and M (L) el e ments; spec i mens ZPAL cXVI/2981, 2984, 2983 (holotype, H), 2985,
2986, and 2982, re spec tively.

eage of P. confluens ® P. triphyllatus the plat form is ex ten sive, and de vel ops a con stric tion at mid−length.Even ear lier in on tog eny the plat form in the P. diversus ® P. nodoundatus lin eage orig i nated. P. perplexusmay or may not be a con tin u a tion of the lat ter, but the tran si tion has not been traced. Prob a bly it gave rise to
78 JERZY DZIK
Fig. 50. Plat form−bear ing de riv a tive of Mehlina, a pop u la tion of ro bust Polynodosus transitus sp. n., in clud ing pos si ble P. lauri −formis (Dreesen et Dusar, 1974) (D, E), from the C. quadrantinodosa Zone at Łagów (sam ple Ł−26) in the Holy Cross Moun −tains. P1 (A–E), P2 (F, G), S0 (H), S1 (I), S2 (J), S3–4 (K), and M (L and M) el e ments; spec i mens ZPAL cXVI/2145 (holo type, A),
2144, 2146, 2156, 2155, 2147–2152, 2154, and 2153, re spec tively.

Hemilistrona, in which an elab o rate plat form de vel oped in the P1 el e ments. Late mem bers of Polynodosusare al most homeomorphic to Polygnathus and they were ear lier be lieved to orig i nate from that lineage ofancient platform−bearing polygnathids.
Polynodosus transitus sp. n.(Figs 50, 51A–M, and 127)
Holotype: Spec i men ZPAL cXVI/2145 (Fig. 50A).Type ho ri zon and lo cal ity: Sam ple Ł−26, mid Famennian C. quadrantinodosa Zone at Łagów, Holy Cross Moun tains.Der i va tion of name: Re fer ring to tran si tional po si tion be tween Mehlina and Polynodosus.
Ma te rial. — 5,272 spec i mens.
FAMENNIAN CONO DONTS AND AMMONOIDS 79
Fig. 51. Plat form−bear ing de riv a tive of Mehlina, Polynodosus from the Famennian of the Holy Cross Moun tains. A–M. Latepop u la tion of gracile Polynodosus transitus sp. n. from the P. trachytera Zone at Jabłonna (bed 24). P1 (A–E), P2 (F, G, and I),S0 (H), S1 (J), S2 (L), S3–4 (M), and M (K) el e ments; spec i mens ZPAL cXVI/2131–2135, 2137–2139, 2136, 2140, 2143, and2141, 2142, re spec tively. N, O. P. lauriformis (Dreesen et Dusar, 1974) from the L. styriacus Zone at Ściegnia (sam ple Wzd−13).
P1 (O) and P2 (N) el e ments; spec i mens ZPAL cXVI/2260 and 2259.

Di ag no sis. — Plat form of the P1 el e ment of vari able width, usu ally weakly de vel oped and re stricted tothe mid dle part of the el e ment, its dor sal tip sharply pointed.
Re marks. — This is a spe cies of a gen er al ized mor phol ogy and par tic u lar el e ment types of closely sim i −lar mor phol ogy oc cur in sev eral other polygnathids. It is the whole ap pa ra tus com po si tion (Mehlina−like), not the P1 el e ment alone, that al lows to dis tin guish it from, say, Ctenopolygnathus angustidens.
Ju ve nile spec i mens of the spe cies are closely sim i lar to those of Mehlina and an in cip i ent plat formdevelopes rel a tively late in their on tog eny in a form of ridges at the mid dle height of the blade. The ten dencyto de velop peniculus, so char ac ter is tic for Mehlina is ap par ent at early ontogenetic stages (Fig. 51A and C).The mor phol ogy of P1 el e ments is very vari able which makes de lim i ta tion of the spe cies, both within sam −ples and in re spect to its strati graphic range, dif fi cult. From M. lauriformis (Dreesen et Dusar, 1974) prob a bly rep re sent ing a more ad vanced stage in the de vel op ment of the lin eage, it dif fers in hav ing a heart−shaped out −line of the plat form. How ever, even in rel a tively early pop u la tions of P. transitus, spec i mens with elon gatedplat form of laurus−leaf out line oc cur (Fig. 50D, E). They may or may not rep re sent a sep a rate spe cies. In fact, the pop u la tion from the Lower Łagów Beds is rep re sented by a sam ple dom i nated by ro bust plat form el e −ments. Prob a bly this is a re sult of low ju ve nile mor tal ity, in op po si tion to the youn ger pop u la tion fromJabłonna, where the platform is missing in most specimens of generally small size.
Polygnathus aff. fallax Helms et Wolska, 1967 from the C. marginifera Zone of Guangxi (Wang 1989, p.110, pl. 39: 5) rep re sents the same spe cies.
Oc cur rence. — Wide spread in the Holy Cross Moun tains in the C. quadrantinodosa to P. trachytera zones.
80 JERZY DZIK
Fig. 52. Plat form−bear ing de riv a tive of Mehlina, Polynodosus lauriformis (Dreesen et Dusar, 1974) from the L. styriacus Zone at Ostrówka (A–E and J, K, sam ple Ost 12; F–I and L, sam ple Ost 11) in the Holy Cross Moun tains. P1 (A–E), P2 (F, G), S0 (L),S1 (H), S2 (I), S3–4 (J), and M (K) el e ments; spec i mens ZPAL cXVI/2162, 2161, 2160, 2163, 2164, 2167, 2166, 2168–2170, 2165,
and 2171, re spec tively.

Polynodosus lauriformis (Dreesen et Dusar, 1974)(Figs 51N, O, 52, and 127)
Type ho ri zon and lo cal ity: Sam ple 3 from the Hamoir Néblon supérieur sec tion near Liege, Bel gium (Dreesen and Dusar 1974).
Ma te rial. — 367 spec i mens.Di ag no sis. — Laurus leaf−shaped plat form or na mented with nodes, re stricted to its mar gin in ju ve nile
spec i mens and cov er ing the whole plat form sur face in adults.Re marks. — From the co eval Lagovignathus fallax dif fers in the coarse or na men ta tion of the plat form
and the com po si tion of the ap pa ra tus, from its prob a ble an ces tor P. transitus in a more reg u lar out line of theplat form. In sam ples Ł−24 and Ł−26 spec i mens mor pho log i cally in dis tin guish able from P. lauriformis (Fig.50D–F) co−oc cur with those of P. transitus and tran si tional spec i mens are rare. Whether these are sympatricspe cies or just a case of pop u la tion vari abil ity, re mains to be clar i fied. There is a pos si bil ity that P.lauriformis is a more off−shore coun ter part of P. transitus.
Oc cur rence. — From the C. marginifera to L. styriacus zones at Kowala, Wzdół, Miedzianka, andOstrówka.
Polynodosus confluens (Ulrich et Bassler, 1926)(Fig. 53)
Type ho ri zon and lo cal ity: “Hardin sand stone” near Mount Pleas ant, Ten nes see (Hud dle 1968).
Ma te rial. — 1,011 spec i mens.Di ag no sis. — Plat form of P1 el e ments leaf−like in out line and slightly con stricted at mid−length, bears
more or less dis tinctly de vel oped ridges par al lel to the blade in its ven tral part and is uni formly tuberculatedin the dor sal part; P2 el e ments ro bust but with peniculus at early stages.
Re marks. — The ap par ent sep a ra tion of the dor sal part of the plat form from the ven tral part by an in cip i −ent con stric tion, vis i ble in the topotype ma te rial (Hud dle 1968, pl. 141−4), leaves lit tle doubt that this is a rel −a tive of P. triphyllatus. Peniculus and plat form with rows of tu ber cles par al lel to the blade in ju ve niles (Fig.53B and F) dis close af fin i ties with Polynodosus transitus. The sim i lar ity of adult P1 spec i mens to those ofPolylophodonta is thus only convergent.
In the Holy Cross Moun tains and Thuringian sam ples from the early Cheiloceras Stufe, to gether withspec i mens closely sim i lar to the lectotype an other morphotypes oc cur, clas si fied by Helms (1961) in hisPolygnathus flaccida. Such forms have not been il lus trated by Hud dle (1968) among the topotype Amer i canspec i mens. This may be due to the small num ber of ex clu sively ma ture spec i mens rpresented in his sam ple.In fact, the in com pletely pre served holotype of Polygnathus pennatuloideus of Holmes (1928), co−oc cur ringwith P. confluens in the Chat ta nooga Shale near Huntsville, Al a bama may rep re sent a ro bustly or na mentedmorphotype of the same population.
There is a pos si bil ity of some what bi modal (di mor phic) dis tri bu tion of morphologies of this el e ment but it is ob scured by a very wide range of pop u la tion vari abil ity. Pos si bly the spec i mens with finely tuberculateddor sal part of the plat form are counterpairs of those with wide strongly ribbed plat form but there is a com mon re ver sal in asymmetry within the pair.
Oc cur rence. — The K. crepida Zone at Kadzielnia and Kowala.
Polynodosus triphyllatus (Ziegler, 1962)(Figs 54A–I and 128)
Type ho ri zon and lo cal ity: Peb ble in the late Perm ian con glom er ate of Rossenray in the Rhenish Slate Moun tains.
Ma te rial. — 1,106 spec i mens.Di ag no sis. — Plat form of P1 el e ment strongly con stricted in the midlength.Re marks. — Ac cord ing to Helms (1961) and Vorontzova (1993) a few chronospecies of polygnathids
with or nate plat form pre ceded this spe cies in its lin eage. In my ma te rial there seems to be a con ti nu ity be −tween P. confluens and P. triphyllatus and grad ual in crease in depth of plat form con stric tion, the char ac terhighly vari able within each sample.
Ju ve nile el e ments de scribed as Polygnathus cf. P. diversus by Metzger (1994), who iden ti fied P2 el e −ments and il lus trated their on tog eny, prob a bly be long to a pop u la tion tran si tional be tween P. confluens andP. triphyllatus, as sug gested by co−oc cur rence with ma ture P1 elements.
FAMENNIAN CONO DONTS AND AMMONOIDS 81

82 JERZY DZIK
Fig. 53. Elab o rate plat form−bear ing Polynodosus confluens (Ulrich et Bassler, 1926) from the K. crepida Zone at Kadzielnia(A and J, sam ple Ka−7; B and D, sam ple Ka−6; C, E–I, K, L, and N, sam ple Ka−4; M, sam ple Ka−10) in the Holy Cross Moun tains. P1 (A–E), P2 (F, G, and L), S0 (H), S1 (I), S2 (J and N), S3–4 (M), and M (K) el e ments; spec i mens ZPAL cXVI/2634, 2630, 2621,
2631, 2624, 2622, 2623, 2625, 2626, 2633, 2628, 2629, 2632, and 2627, re spec tively.

No men cla tor ial prob lems con nected with this spe cies have been dis cussed by Glenister and Klapper(1966, p. 834).
Oc cur rence. — The C. quadrantinodosa to C. marginifera zones at Kowala, Łagów and Miedzianka.
Polynodosus nodoundatus (Helms, 1961)(Figs 54M–O and 128)
Type ho ri zon and lo cal ity: Grey, thick−bed ded lime stone of the Prolobites Zone at Großen Buschteich near Rödersdorf,Thuringia (Helms 1961).
FAMENNIAN CONO DONTS AND AMMONOIDS 83
Fig. 54. Ad vanced spe cies of Polynodosus. A–I. P. triphyllatus (Ziegler, 1960) from the C. quadrantinodosa Zone at Łagów (A, B,sam ple Ł−26; C–I, sam ple Ł−27) in the Holy Cross Moun tains. P1 (A–C), P2 (D), S0 (E), S1 (ab er rant with ad di tional pro cess, I),S2 (F), S3–4 (G), and M (H) el e ments; spec i mens ZPAL cXVI/2157, 2158, 2496–2500, 2502, and 2501, re spec tively. J–L. Polyno −dosus diversus (Helms, 1959) from the L. styriacus Zone at Miedzianka (sam ple Md−14). P1 el e ments; spec i mens ZPALcXVI/2635–2637, re spec tively. M–O. Polynodosus nodoundatus (Helms, 1961) from the C. quadrantinodosa Zone at Łagów
(sam ple ŁSł73−3). P1 el e ments; spec i mens ZPAL cXVI/2638, 2640, and 2639, re spec tively.

Ma te rial. — 276 spec i mens.Di ag no sis. — Blunt apex of wid ened plat form in ma ture P1 el e ment tend ing to de velop a de pres sion in
the mid dle, or na mented with ir reg u larly dis trib uted tubercles.Re marks. — In no sam ple avail able to me is the spe cies the only plat form−bear ing polygnathid and is al −
most al ways sub or di nate in num ber. Only in sam ple Md−12 a sig nif i cant num ber of P1 el e ments have beenen coun tered. They co−oc cur with ro bust P2 el e ments of mor phol ogy in dis tin guish able from those at trib utedto P. triphyllatus. This speaks in fa vor of the pro posed ge neric af fil i a tion of P. nodoundatus. Pre sum ably thisis an other derivartive of P. confluens.
Oc cur rence. — The C. marginifera Zone at Łagów, Kowala, Miedzianka, and Ostrówka.
Polynodosus diversus (Helms, 1959)(Figs 54J–L and 128)
Type ho ri zon and lo cal ity: With S. velifer 4.5 m be low the top of bed 5 at Bohlen near Saalfeld, Thuringia (Helms 1959).
Ma te rial. — 58 spec i mens.Di ag no sis. — Ju ve nile P1 el e ments with a ro bust peniculus, ma ture el e ments with rows of par al lel nodes
along the oval plat form with great est width in its dorsal part.Re marks. — Only subadult spec i mens of the spe cies have been known pre vi ously (Helms 1959; Aus tin
et al. 1985), but in sam ple Md−14 from Miedzianka there is a gra da tion from morphs with a rather nar rowplat form to frag men tary large spec i mens with mul ti ple rows of denticles. The spe cies may be re lated to P.nodoundatus.
Oc cur rence. — The C. marginifera and L. styriacus Zone at Miedzianka.
Genus Ctenopolygnathus Müller et Müller, 1957
Type spe cies: Polygnathus angustidiscus Youngquist, 1945 from the Frasnian In de pend ence Shale of Iowa.
Di ag no sis. — Polygnathids with ro bustly ap pear ing plat form that does not reach the dor sal (“pos te rior”)tip in most of the P1 el e ments even at late ontogenetic stages.
Re marks. — The mor phol ogy of Ctenopolygnathus is so sim ple and un de rived that there is a greatchance of its con ver gent or i gin. If this is true, the only guide to tax o nomic iden tity of spe cies in cluded in thege nus is their stratophenetically traced re la tion ship to the type species.
Bardashev et al. (2002) in tro duced sev eral ge neric rank names for early polygnathids, de fined on themor phol ogy of the plat form in P1 el e ments, with out ref er ence to the ap pa ra tus com po si tion. They are ap par −ently un der stood as polyphyletic grades in the evo lu tion of the branch. Among them is the re de finedCtenopolygnathus in a mean ing rather fun da men tally dif fer ent from that applied by myself (Dzik 2002).
Ctenopolygnathus brevilamina (Branson et Mehl, 1934)(Figs 55, 56, and 129)
Type ho ri zon and lo cal ity: Early Famennian at Dixie and Noel, Mis souri (Branson and Mehl 1934, p. 246).
Ma te rial. — 933 spec i mens.Di ag no sis. — P1 el e ments with subparallel mar gins of the plat form al most ap proach ing the tip of the
blade in ma ture spec i mens.Re marks. — This spe cies was ear lier il lus trated by my self (Dzik 2002) as Ctenopolygnathus sp. A from
the Kellwasserakalk and C. sp. B from the basal Famennian at Płucki. Non−plat form el e ments of the ap pa ra −tus are of a rather prim i tive mor phol ogy, with short pro cesses. Their growth was ter mi nal, as in di cated byhooked tips of pro cesses, but ter mi na tion may have taken place at very dif fer ent sizes. Gen er ally, spe cies ofCtenopolygnathus are ex tremely vari able and their iden ti fi ca tion re quires sam ples with many spec i mens. The Frasnian P. angustidiscus had a gen er ally shorter and even more vari able plat form of P1 el e ments. How ever,even in C. brevilamina there are spec i mens which did not de velop plat form even late in their on tog eny (Fig.56A), oth ers have a wide plat form re sem bling that of Immognathus (Fig. 56K). Quite fre quently the dor saltip of the el e ment, which lacks a plat form, developed a prominent denticulation of the blade.
The un sta ble mor phol ogy of el e ments of the ap pa ra tus sug gests that Ctenopolygnathus orig i nated bypaedomorphosis, pos si bly from rather ad vanced polygnathids. In its Famennian evo lu tion the orig i nal de −rived sta tus seems to be par tially re−established.
84 JERZY DZIK
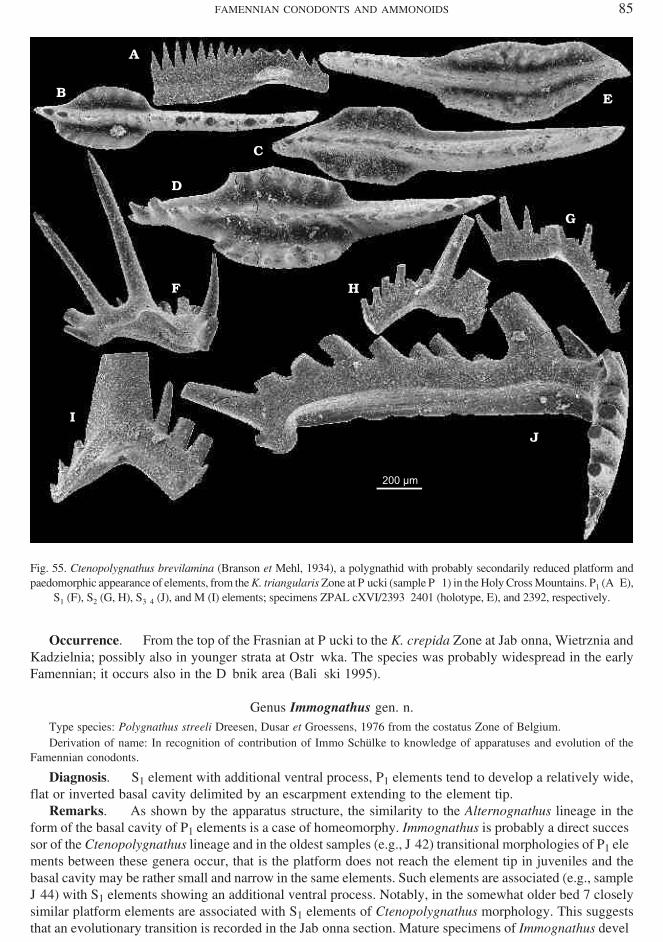
Oc cur rence. — From the top of the Frasnian at Płucki to the K. crepida Zone at Jabłonna, Wietrznia andKadzielnia; pos si bly also in youn ger strata at Ostrówka. The spe cies was prob a bly wide spread in the earlyFamennian; it oc curs also in the Dębnik area (Baliński 1995).
Genus Immognathus gen. n.Type spe cies: Polygnathus streeli Dreesen, Dusar et Groessens, 1976 from the costatus Zone of Bel gium.Der i va tion of name: In rec og ni tion of con tri bu tion of Immo Schülke to knowl edge of ap pa ra tuses and evo lu tion of the
Famennian conodonts.
Di ag no sis. — S1 el e ment with ad di tional ven tral pro cess, P1 el e ments tend to de velop a rel a tively wide,flat or in verted basal cav ity de lim ited by an es carp ment ex tend ing to the element tip.
Re marks. — As shown by the ap pa ra tus struc ture, the sim i lar ity to the Alternognathus lin eage in theform of the basal cav ity of P1 el e ments is a case of homeomorphy. Immognathus is prob a bly a di rect suc ces −sor of the Ctenopolygnathus lin eage and in the old est sam ples (e.g., J−42) tran si tional morphologies of P1 el e −ments be tween these gen era oc cur, that is the plat form does not reach the el e ment tip in ju ve niles and thebasal cav ity may be rather small and nar row in the same el e ments. Such el e ments are as so ci ated (e.g., sam ple J−44) with S1 el e ments show ing an ad di tional ven tral pro cess. No ta bly, in the some what older bed 7 closelysim i lar plat form el e ments are as so ci ated with S1 el e ments of Ctenopolygnathus mor phol ogy. This sug geststhat an evo lu tion ary tran si tion is re corded in the Jabłonna sec tion. Ma ture spec i mens of Immognathus de vel −
FAMENNIAN CONO DONTS AND AMMONOIDS 85
Fig. 55. Ctenopolygnathus brevilamina (Branson et Mehl, 1934), a polygnathid with prob a bly sec ond arily re duced plat form andpaedomorphic ap pear ance of el e ments, from the K. triangularis Zone at Płucki (sam ple Pł−1) in the Holy Cross Moun tains. P1 (A–E),
S1 (F), S2 (G, H), S3–4 (J), and M (I) el e ments; spec i mens ZPAL cXVI/2393–2401 (holotype, E), and 2392, re spec tively.

oped asym me try in plat form shape sim i lar to that in Polygnathus webbi. Polygnathus rarus of Baliński(1995) from the Dębnik area may be another member of this branch.
Immognathus streeli (Dreesen, Dusar et Groessens, 1976)(Figs 57 and 129)
Type ho ri zon and lo cal ity: Sam ple YG 9 taken at theYves−Gomezée road sec tion, Bel gium with Icriodus costatus, lowercostatus Zone (Dreesen et al. 1976).
Ma te rial. — 738 spec i mens.Di ag no sis. — Plat form of P1 el e ments or na mented with mar ginal tu ber cles ex tend ing admedially as short
trans verse ridges; in ju ve niles plat form does not reach the dor sal tip of the blade.Re marks. — Bi fur ca tion of the ven tral pro cess in S1 el e ments may be some what ir reg u lar and ad di tional
bi fur ca tion may oc cur (e.g., in sample J−44).
86 JERZY DZIK
Fig. 56. Sim pli fied polygnathid Ctenopolygnathus brevilamina (Branson et Mehl, 1934) from the K. triangularis Zone atKowala (A, B, F–J, sam ple Ko−154; C–E, sam ple Ko−143) and Jabłonna (K–M, sam ple J−53a) and the K. crepida Zone atKadzielnia (N, sam ple Ka−3) in the Holy Cross Moun tains. P1 (A–D, K, L), P2 (E), S0 (F), S1 (G and M), S2 (H and N), S3–4 (I), and
M (J) el e ments; spec i mens ZPAL cXVI/2406, 2405, 2402–2404, 2407–2411, 2414, 2412, 2413, and 2415, re spec tively.

Oc cur rence. — The K. crepida to P. trachytera zones at Jabłonna, Kowala and Miedzianka.
Immognathus rhabdotus (Schäfer, 1976)(Figs 58B, D–L, and 129)
Type ho ri zon and lo cal ity: Bed 21 in trench dug at Dasberg near Hövel, Rhenish Slate Moun tains (Schäfer 1976).
Ma te rial. — 47 spec i mens.Di ag no sis. — P1 el e ments with trans versely ribbed plat form tend ing to be come flat dor sally in ma ture
spec i mens and with par al lel sides over most of its length.
FAMENNIAN CONO DONTS AND AMMONOIDS 87
Fig. 57. Polygnathid with Siphonodella−like basal cav ity, Immognathus streeli (Dreesen, Dusar et Groessens, 1976) from theK. crepida Zone at Wietrznia (A–E, H, J, L, M, Q and O, sam ple Wtr−21) and Jabłonna (G, I, K, N, P, and R, sam ple J−45a) in theHoly Cross Moun tains. P1 (A–G), P2 (H, I), S0 (J), S1 (K, L), S2 (M, N), S3–4 (O, P), and M (Q, R) el e ments; spec i mens ZPALcXVI/2417, 2418, 2427, 2425, 2416, 2428, 2426, 2420, 2429, 2419, 2430, 2421, 2422, 2431, 2423, 2432, 2424, and 2433,
respectively.

Re marks. — In sam ple Ost−3 P1 el e ments of this spe cies dom i nate among polygnathids, be ing as so ci ated with ro bust ramiform el e ments of the Ctenopolygnathus type (Fig. 58I, J) that sup ports the ge neric place ment of the spe cies. Subadult P1 el e ments of this spe cies are vir tu ally homeomorphic with those of Pinaco −gnathus? praesulcatus. The dif fer ence is best vis i ble in ju ve niles, hav ing ven trally wider plat form than in P.praesulcatus and in larger el e ments, with basal cav ity of less an gu lar sec tion. If the par tial ap pa ra tus re con −struc tion of the Tournaisian Pinacognathus? sulcatus pro posed by my self is cor rect (Dzik 1997), all thesespe cies may be long to the same lin eage. The ge neric name Immognathus may then ap pear ap pro pri ate tothem and is to be trans ferred to the family Elictognathidae to ensure its monophyly.
From stratigraphically older Polygnathus? pennatulus the spe cies dif fers in a less tri an gu lar out line of theplat form and in lan ceo late basal cone.
The P1 el e ments of sim i lar mor phol ogy from the bound ary strata be tween the Famennian and Tour nai −sian, were iden ti fied by Bouckaert and Groessens (1976), Over (1992), and Sanz−López et al. (1999) asPolygnathus inornatus (with a Tournaisian type pop u la tion, clas si fied in Pinacognathus by Dzik 1997) andas Pseudopolygnathus graulichi by Aus tin et al. (1985). The lin eage con tin ued to the Car bon if er ous pos si bly rep re sented there by Pinacognathus inornatus (E.R. Branson, 1934) (e.g., Dzik 1997). P. graulichi, with avery large basal cone, is rather a mem ber of the Dasbergina lin eage. Re la tion ships and no men cla ture of these lat est Famennian and early Tournaisian forms requires clarification.
Oc cur rence. — The L. styriacus to D. trigonica zones at Ostrówka.
Genus Polygnathus Hinde, 1879
Type spe cies: Polygnathus dubius Hinde, 1879 from the North Ev ans Lime stone at Eighteenmile Creek in New York(Klapper and Philip 1971).
88 JERZY DZIK
Fig. 58. Late Famennian Siphonodella−like polygnathids. A, C. Polygnathus pennatulus Ulrich et Bassler, 1926 from the L. sty −riacus Zone at Ostrówka (sam ple Ost−12) in the Holy Cross Moun tains, pos si bly end mem bers of pop u la tion vari abil ity ofHemilistrona perplexa. P1 el e ments; spec i mens ZPAL cXVI/2653–2654. B, D–L. Immognathus rhabdotus (Schäfer, 1976) from the D. trigonica Zone at Ostrówka (B, D, sam ple Ost−293; E–J, sam ple Ost−3) and Kowala (K, sam ple Ko−98) in the Holy CrossMoun tains. P1 (A–G, K, L), P2 (H), S2 (I), and S3–4 (J) el e ments; spec i mens ZPAL cXVI/2673, 2720–2722, 2726, 2723–2725,
2391, and 2388, re spec tively.

Polygnathus praecursor Matyja, 1993(Figs 59, 60, 61, and 130)
Type ho ri zon and lo cal ity: Depth 4636–4637 m in the bore hole Unisław−2, West ern Pomerania, Lower or Mid dletriangularis Zone (Matyja 1993).
Ma te rial. — 5,369 spec i mens.Di ag no sis. — Strong asym me try in pairs of P1 el e ments, ma ture el e ments with ca rina dis tinct to its tip.Re marks. — The ap pa ra tus of an ear li est Famennian pop u la tion of the spe cies from the Montagne Noire
was re stored by Schülke (1999) un der the name Polygnathus brevilaminus. The type pop u la tion of the lat terspe cies is dif fer ent from P. praecursor in show ing a more ir reg u lar de vel op ment of the plat form in P1 el e mentsand more ro bust ap pear ance of the whole ap pa ra tus, and was clas si fied in Ctenopolygnathus by Dzik (2002).
The ear li est Famennian pop u la tion of true Polygnathus from the Holy Cross Moun tains was iden ti fied bymy self (Dzik 2002) as be long ing to the Polygnathus semicostatus lin eage. At that time only ju ve nile plat form spec i mens were known. More nu mer ous ma te rial from that lo cal ity and other sites in the Holy Cross Moun −tains shows a sig nif i cant dif fer ence be tween the ear li est and mid Famennian mem bers of the lin eage and their clas si fi ca tion in sep a rate spe cies, as pro posed by Matyja (1993), is here supported.
The spe cies seems to be a con tin u a tion of the Frasnian P. webbi lin eage and dis crim i na tion of theirbound aries with mor phol ogy alone may not be possible.
Oc cur rence. — From the be gin ning of Famennian to the early K. crepida Zone at Płucki, Kowala,Jabłonna, and Kadzielnia.
Polygnathus semicostatus Branson et Mehl, 1934(Figs 8B, 62, and 130)
Type ho ri zon and lo cal ity: Famennian at Dixie, Mis souri (Ziegler 1977, p. 313).
Ma te rial. — 1,896 spec i mens.Di ag no sis. — Tongue−like dor sal end of the plat form in P1 el e ment much nar rower than the main part, or −
na mented with con tin u ous trans verse ribs.
FAMENNIAN CONO DONTS AND AMMONOIDS 89
Fig. 59. Di rect suc ces sor of the Frasnian lin eage of asym met ric plat form−bear ing polygnathids, Polygnathus praecursor Matyja,1993 from the ear li est pre−K. triangularis Zone of the Famennian at Płucki (sam ple Pł−36) in the Holy Cross Moun tains.P1 (A–C, ju ve nile and prob a ble adults; note that sinistral and dextral spec i mens are not mir ror−im ages), P2 (D), S0 (E, F), S1 (G),S2 (H), S3–4 (I), and M (J, K) el e ments; spec i mens ZPAL cXVI/1535, 1533, 1534, 1536–1541, 1543, and 1542, re spec tively.

Re marks. — The P1 el e ments of the spe cies dif fer from su per fi cially sim i lar el e ments of the Eifelian P.linguiformis in the mode of arch ing of the plat form and in hav ing a shorter blade (Dreesen and Or chard1974). The geo log i cally old est spec i mens show ing plat form or na men ta tion typ i cal for this spe cies oc cur in
90 JERZY DZIK
Fig. 60. Typ i cal plat form−bear ing polygnathid Polygnathus praecursor Matyja, 1993 from the K. triangularis Zone at Jabłonna(A–H, sam ple J−53a; I, K–P, sam ple J−58; J, sam ple J−59) in the Holy Cross Moun tains. P1 (A–I, ontogenetic se ries of el e mentpairs), P2 (J, K), S0 (L), S1 (M), S2 (N), S3–4 (O), and M (P) el e ments; spec i mens ZPAL cXVI/2434, 2436, 2435, 2437–2449,
respectively.

sam ples Ka−3 and Ka−10 at Kadzielnia. They are as so ci ated with the dom i nant morph, in which tu ber clesnear the dor sal end of the plat form tend to be ar ranged into densely dis trib uted trans verse ribs, clas si fied byMatyja (1974) in her P. szulczewskii. The type spec i men of that spe cies seems to be a dextral el e ment of theearly P. semicostatus, as sug gested by its co−oc cur rence in the same sam ple (Minkowice 1 bore hole depth2385.4–2391.4; cf. Matyja 1974 and Matyja and Żbikowska 1974, pl. 6:8) with a sinistral el e ment of P.semicostatus. Such ini tially pro found ap pa ra tus asym me try tends to dis ap pear in the evo lu tion of P. semi −costatus. This is con nected with a wider ap pear ance of the ven tral part of the plat form, which no lon ger de −vel ops ver ti cal mar gins. The same re fers to P. planirostratus Dreesen et Dusar, 1974. Polygnathus limbatusMatyja, 1993 from the lower marginifera zone of the Chojnice bore hole in west ern Pomerania seems to be aju ve nile of P. semicostatus.
There is an in ter est ing as pect of changes in the pat tern of vari abil ity in the evo lu tion of P. semicostatus.In the pre ced ing P. praecursor the asym me try in the P1 el e ment pair is sta ble, that is the sinistral el e ment is nar rower and with no ap par ent el e va tion of the an te rior mar gin of the plat form. In early pop u la tions of P.semicostatus rare dextral el e ments with “sinistral” mor phol ogy (and op po site) ap pear. In late pop u la tionssuch an in ver sion in asym me try is very com mon. In ef fect both dextral and sinistral el e ments be came di −mor phic, with some in ter me di ate forms, too. This loss of strict con trol of asym me try is co eval with thegen eral in crease in pop u la tion vari abil ity. Ap par ently the developmental mechanisms were losing theirrigidity.
P. semicostatus is prob a bly the most vari able of all the Famennian polygnathids (Dreesen and Or chard1974). This re fers also to the or ga ni za tion of its basal cav ity. In sam ples Ko−15, ŁSł73−2 and Ł−27 spec i menswith a de pres sion in the base (the char ac ter used to de fine Neopolygnathus) oc cur (Fig. 70H–J), oth er wisethey do not dif fer from the as so ci ated typ i cal el e ments of the spe cies. More over, spec i mens with a vir tu allysmooth nar row part of the plat form oc cur, sim i lar to Polygnathus sp. A of Matyja and Żbikowska (1974)from the crepida Zone of the bore hole Minkowice 1 in the Lublin area or Polygnathus padovanii of Perri andSpalletta (1990) from co eval strata of the Carnic Alps. Com bi na tion of these two char ac ters in spec i mensfrom Łagów (sam ple Ł−27) re sults in a mor phol ogy vir tu ally in dis tin guish able from Neopolygnathus com −munis. Such spec i mens have been clas si fied as Polygnathus depressus by Metzger (1989).
FAMENNIAN CONO DONTS AND AMMONOIDS 91
Fig. 61. Late pop u la tion of Polygnathus praecursor Matyja, 1993 from the late K. triangularis Zone at Kadzielnia (sam ple Ka−16)in the Holy Cross Moun tains. P1 el e ments; spec i mens ZPAL cXVI/2487, 2486, 2488, 2490, 2489, and 2491, re spec tively.

As in di cated by the dis tri bu tion of im prints of ameloblasts, the dor sal part of the plat form was more in −tensely se creted than the ven tral one.
Oc cur rence. — The late K. crepida to L. styriacus zones at Jabłonna, Kadzielnia, Wietrznia, Kowala,Miedzianka, and Ostrówka. The spe cies oc curs also in the Dębnik area (Baliński 1995).
92 JERZY DZIK
Fig. 62. Polygnathus semicostatus Branson et Mehl, 1934 from the P. trachytera Zone at Kowala (A, B, F, G, I–O, sam ple Ko−15),K. crepida Zone at Kadzielnia (C–E, H, sam ple Ka−3), and the C. quadrantinodosa Zone at Łagów (T, X, sam ple Ł−32; Q, sam pleŁ−27) in the Holy Cross Moun tains. P1 (A–J, P–R), P2 (K), S1 (L), S2 (M), S3–4 (N), and M (O) el e ments; spec i mens ZPAL
cXVI/2517, 2516, 2493, 2495, 2492, 2514, 2515, 2494, 2495, 2519–2509, 2520–2524, 2526, 2570, and 2525, re spec tively.

Polygnathus? experplexus Sandberg et Ziegler, 1979(Figs 63 and 130)
Type ho ri zon and lo cal ity: Up per styriacus Zone, Box Mem ber of Percha Shale east of Santa Rita, New Mex ico (Sandbergand Ziegler 1979).
Ma te rial. — 554 spec i mens.Di ag no sis. — P1 el e ments with two tuberculated rostral ridges and lobate plat form or na mented with tu −
ber cles.Re marks. — The or i gin of the spe cies re mains un set tled. P1 el e ments show some sim i lar ity to Hemili −
strona perplexa, but the ap pa ra tus is rather dif fer ent and its mor phol ogy may sug gest an or i gin fromPolygnathus praecursor. In early pop u la tions of prob a ble P.? experplexus the ma ture P1 el e ments are con vex cen trally (near the cusp), wheareas in typ i cal pop u la tions the cor re spond ing part of the platform is gentlyconcave.
Oc cur rence. — The L. styriacus and P. jugosus zones at Jabłonna, Ostrówka, Kowala, and Dzikowiec.
FAMENNIAN CONO DONTS AND AMMONOIDS 93
Fig. 63. Prob lem atic Polygnathus? experplexus (Sandberg et Ziegler, 1979) from the L.styriacus Zone at Ostrówka (A, D–F,sam ple Ost−12; B, C, I–K, sam ple Ost−293) in the Holy Cross Moun tains and Dzikowiec (L, sam ple Dz−71) in the Sudetes.P1 (A–D, L), P2 (E), S0 (F), S1 (J?), S2 (G, K?), S3–4 (H, L), and M (M?) el e ments; spec i mens ZPAL cXVI/2656, 2680, 2681, 2672,
2664–2668, 2683–2685, and 2971, re spec tively.

Polygnathus extralobatus Schäfer, 1976(Figs 64A–K and 130)
Type ho ri zon and lo cal ity: Bed 65 in the trench at Dasberg near Hövel, Rhenish Slate Moun tains (Schäfer 1976).
94 JERZY DZIK
Fig. 64. Late Famennian polygnathids from the P. jugosus Zone at Ostrówka (D, N, O–R, sam ple Ost−265) and Jabłonna (E–J,bed 27) in the Holy Cross Moun tains, Gołogłowy (K–M, Q, sam ple Goł−3) and Dzikowiec (A–C, sam ple Dz−53) in the Sudetes.A–K. Polygnathus extralobatus Schäfer, 1976. P1 (A, D, E, K), P2 (F, G), S0 (B, H), S3–4 (I), and M (C, G) el e ments; spec i mensZPAL cXVI/2968–2970, 2686–2690, and 2715, re spec tively. L–W. Polygnathus znepolensis Spassov, 1965. P1 (L–N, U),P2 (O, T), S3–4 (P, S), and M (Q, R, V) el e ments; spec i mens ZPAL cXVI/2697, 2698, 2691, 2717, 2699−2700, 2718, 2719, 2975,
2973, 2972, 2974, and 2976, re spec tively.

Ma te rial. — 1,014 spec i mens.Di ag no sis. — Wid ened dor sal part of the plat form in P1 el e ment cov ered with obliquely run ning trans −
verse ribs, dis con tin u ous near the carina.Re marks. — The spe cies is so close to its strati graphic pre de ces sor, P. obliquicostatus Ziegler, 1962 that
the an ces tor−de scen dant re la tion ship be tween them seems al most cer tain (Schäfer 1976). That spe cies has not been en coun tered in the ma te rial avail able to me so the lin eage prob a bly en tered the Holy Cross Moun tainsaf ter the wid en ing of the plat form lobe was com pleted. P. semicostatus seems to be a likely an ces tor of P.obliquicostatus.
Oc cur rence. — The P. jugosus and D. trigonica zones at Kowala, Jabłonna, and Gołogłowy.
Polygnathus znepolensis Spassov, 1965(Figs 64L–W and 130)
Type ho ri zon and lo cal ity: Sum mit of Krast hill at Berainci, Bul garia (Klapper in Ziegler 1975).
Ma te rial. — 445 spec i mens.Di ag no sis. — An te rior part of the plat form in P1 el e ment or na mented with oblique ribs, pos te rior lobe
wide and gently con vex, smooth.Re marks. — This is a spe cies widely oc cur ring in shal low−wa ter Late Famennian en vi ron ments (Schäfer
1976). In the holotype of P. znepolensis re−il lus trated by Klapper (in Ziegler 1975, pl. 5: 7) the pos te rior lobeof the plat form bears ro bust ra di at ing ridges, which have not been encoutered in any of the ma ture spec i mensin the Pol ish ma te rial. Pos si bly the holotype be longs thus to P. extralobatus, its close rel a tive and prob a blean ces tor. Both spe cies co−oc cur at Gołogłowy and Ostrówka. In the old est sam ples con tain ing the spe ciesonly min ute ju ve nile spec i mens oc cur. This may re flect ei ther a high ju ve nile mor tal ity in ex treme en vi ron −ments atypical for the species or its paedomorphic origin.
Oc cur rence. — The late L. styriacus and P. jugosus zones at Jabłonna, Ostrówka, Kowala, Gołogłowy,and Dzikowiec. The spe cies oc curs also in the Dębnik area (Baliński 1995).
Polygnathus procerus Sannemann, 1955(Figs 65A–C and 131)
Type ho ri zon and lo cal ity: Black lime stone with Nehdenites verneuili at Breitengrund, Frankenwald (Sannemann 1955a).
Ma te rial. — 832 spec i mens.Di ag no sis. — P1 el e ments with al most sym met ri cal, wil low−leaf−shaped plat form, rel a tively flat in ma −
ture el e ments, or na mented with tubercles. Re marks. — The P1 el e ments of the spe cies dif fer from ju ve niles of P. volhynicus in a less con cave
occlusal sur face and less asym met ric out line of the plat form, from P. kadzielniae in its lan ceo late out line.Ap pa ra tus of the spe cies was re stored by Schülke (1995).
Oc cur rence. — The K. triangularis and K. crepida zones at Płucki, Jabłonna, Kadzielnia, Wietrznia, andMiedzianka.
Polygnathus volhynicus Drygant, 1986(Figs 65D–O and 131)
Type ho ri zon and lo cal ity: Depth 725–780 m in the bore hole Volodymyr Volynsky, Ukraine, K. crepida Zone (Drygant 1986).
Ma te rial. — 1,893 spec i mens.Di ag no sis. — P1 el e ments with asym met ric plat form, in ju ve niles lan ceo late and nar row; its raised mar −
gins or na mented by trans verse ribs; no ap par ent asym me try in pairs; ca rina sep a rate to its tip even in matureelements.
Re marks. — Ju ve niles show a deeper sep a ra tion of the blade from mar ginal parts of the plat form (Drygant1986) than in P. procerus, its prob a ble an ces tor. From P. praecursor, as so ci ated in some sam ples, the spe ciesdif fers in the lack of asym me try of el e ments and their nar row ap pear ance at ju ve nile (but not ear li est) stages.Spec i mens of P. squalidus Drygant, 1986, co−oc cur ring with those of P. volhynicus in the bore hole VolodymyrVolynski are closely sim i lar to ma ture spec i mens from Karczówka, and these names may be synonyms.
Oc cur rence. — The late K. triangularis Zone at Karczówka. The spe cies oc curs also in the Dębnik area(Baliński 1995).
FAMENNIAN CONO DONTS AND AMMONOIDS 95

96 JERZY DZIK
Fig. 65. Early Famennian nar row−plat form polygnathids from K. triangularis Zone of the Holy Cross Moun tains. A–C. Poly −gnathus procerus Sannemann, 1955 from Jabłonna (A, B, sam ple J−58; C, sam ple J−59). P1 el e ments; spec i mens ZPALcXVI/2450–2452. D–O. Polygnathus volhynicus Drygant, 1986 from Karczówka. P1 (D–I), P2 (J), S0 (K), S1 (L), S2 (M),
S3–4 (N), and M (O) el e ments; spec i mens ZPAL cXVI/2453–2464, re spec tively.

Polygnathus kadzielniae sp. n.(Figs 66 and 131)
Holotype: Spec i men ZPAL cXVI/2479 (Fig. 66O).Type ho ri zon and lo cal ity: Bed 8, early Famennian K. crepida Zone at Jabłonna, Holy Cross Moun tains.Der i va tion of name: The gen i tive case is ap plied for the name of the lo cal ity Kadzielnia, fol low ing the tra di tion ini ti ated by
Gürich (1896).
Ma te rial. — 1,458 spec i mens.Di ag no sis. — Mar gins of the main part of the plat form in ma ture el e ments par al lel, the ven tral part of the
plat form occlusal sur face gently con cave, or na mented with weak trans verse ribs and tubercles. Re marks. — This is a late mem ber of the P. volhynicus lin eage, dif fer ent in hav ing the plat form more
widely ex panded and in a more del i cate and reg u lar denticulation of P2 el e ments. Iden ti fi ca tion of a so gen er −al ized Polygnathus spe cies is a dif fi cult task be cause of its great pop u la tion vari abil ity and few di ag nos ticchar ac ters. Per haps there is al ready a name for the spe cies avail able in the lit er a ture, but I have not been ableto identify it with confidence.
Oc cur rence. — The mid K. crepida Zone at Kadzielnia and Jabłonna.
Polygnathus? pennatulus Ulrich et Bassler, 1926(Figs 58A, C and 129)
Type ho ri zon and lo cal ity: Hardin Sand stone at Mount Pleas ant, Ten nes see (Klapper in Ziegler 1975).
Ma te rial. — 77 spec i mens.Di ag no sis. — Deeply con cave plat form occlusal sur face with strong trans verse ribs, out line of the plat −
form tri an gu lar in ju ve nile spec i mens but its sides be come par al lel at later stages in the ventral half.Re marks. — A few spec i mens of this spe cies occuring with out other plat form−bear ing polygnathids il −
lus trated by Metzger (1989) from the Shef field−Ma ple Mill For ma tion of Ne braska, sup port its sep a ratestatus.
Al though the P1 el e ments are closely sim i lar to those of Immognathus rhabdotus in the oral view, theirbases are typ i cal for Polygnathus. This makes re la tion ship to the Immognathus lin eage less likely than to P.kadzielniae, al though as pects of the basal cav ity are vari able in these cono donts. The avail able ma te rial is too small to en able ap pa ra tus re con struc tion, so I pre serve the tra di tional lo ca tion of the species provisionally.
Oc cur rence. — The L. styriacus and P. jugosus zones at Ostrówka, Kowala, Miedzianka, and Dzikowiec.
Genus Hemilistrona Chauff et Dombrowski, 1977Type spe cies: Hemilistrona depkei Chauff et Dombrowski, 1977 from the basal shale mem ber of the Sul phur Springs For −
ma tion near Pevely, Mis souri, lower costatus Zone.Di ag no sis. — Polygnathids with elab o rate plat form de vel oped also in P2 el e ments; plat form of P1 el e −
ments with ridges par al lel to blade in rostral ven tral part and with tuberculated con vex and strongly asym met −ric dor sal part; re main ing el e ments of the ap pa ra tus sim i lar to Polynodosus.
Re marks. — The an ces try of Hemilistrona re mains un solved. The mor phol ogy of P2 el e ments in its old −est spe cies, H. perplexa, sug gests re la tion ship to the early Famennian Polygnathus. Prom i nent ridges back −ing the blade in the P1 el e ment are also sim i lar to those in P. webbi group but, even if they are in asym met ricpairs, this is ob scured by the tre men dous pop u la tion vari abil ity. More over, the ridges ex tend much far therdor sally than in Polygnathus, in small est ju ve niles be ing rather like that of Polynodosus. Also the shape of Mel e ments with trans versely ori ented pro cesses speaks in fa vor of a re la tion ship to the Mehlina–Polynodosuslin eage, as sug gested al ready with re spect to H. perplexa by Vorontzova (1993). Her idea of Polynodosus asin clud ing all or nate Famennian polygnathids is, how ever, in con sis tent with the di ver sity of their ap pa ra tusstruc ture. A re la tion ship to the Frasnian polygnathids of sim i lar plat form ornamentation remains to be testedby apparatus reconstructions.
Hemilistrona perplexa (Thomas, 1949)(Figs 67 and 128)
Type ho ri zon and lo cal ity: Ma ple Mill shale at roadcut (lo cal ity 14 of Thomas 1949) in Wash ing ton County, Iowa (same asfor Pelekysgnathus inclinatus; see Klapper in Ziegler 1975).
Ma te rial. — 787 spec i mens.
FAMENNIAN CONO DONTS AND AMMONOIDS 97

98 JERZY DZIK

Di ag no sis. — Ma ture P2 el e ments with in cip i ent plat form and straight pro cesses, P1 el e ments with thepos te rior tongue of the plat form de vel oped only in ma ture specimens.
Re marks. — The spe cies ac tu ally does not show the wide plat form in P2 el e ments di ag nos tic for the ge −nus but is also dif fer ent from Polynodosus in not de vel op ing a peniculus (or at least the un du la tion of the pro −cess con nected with it is not rec og niz able in larger specimens).
Ramiform el e ments of the ap pa ra tus may be in dis tin guish able from those of as so ci ated plat form−bear ingMehlina. The holotype of the spe cies is a ju ve nile P1 el e ment as so ci ated in the same sam ple (lo cal ity 14 ofThomas 1949) with ap par ently conspecific ma ture el e ments named Palmatolepis? irregularis and Ancyro −gnathus irregularis, ro bust P2 el e ments named Bryantodus? sp., and low−arched ro bust M el e ments namedEuprioniodina iowaensis and E. lateralis. Co−oc cur ring Branmehla inornata, Polygnathus semicostatus,Palmatolepis perlobata, and Conditolepis falcata are sug ges tive of the P. trachytera Zone (un less this is acase of re worked ma te rial). Thomas (1949) il lus trated also a P2 el e ment of ad vanced Hemilistrona from thesame lo cal ity, but in Chauff and Dombrowski (1977) it is re ferred to as com ing from an other lo cal ity near
FAMENNIAN CONO DONTS AND AMMONOIDS 99
Fig. 66. Polygnathus kadzielniae sp. n. from the K. crepida Zone at Kadzielnia (A–D,F, G, sam ple Ka−2; E, H, I–M, sam pleKa−13) and Jabłonna (N–U, bed 8) in the Holy Cross Moun tains. P1 (A–E, N, O), P2 (F, G, P), S0 (I, J, Q), S1 (K, R), S2 (M, S),S3–4 (L, T), and M (H, U) el e ments; spec i mens ZPAL cXVI/2466, 2467, 2465, 2468, 2471, 2469, 2470, 2477, 2473, 2472, 2474,
2476, 2475, and 2478–2485 (holotype, O), re spec tively.
Fig. 67. Hemilistrona perplexa (Thomas, 1949), a ridged plat form−bear ing polygnathid, from the C. marginifera Zone atŚciegnia (Wzdół Plebański; sam ple Wzd−10) in the Holy Cross Moun tains. P1 (A, B), P2 (C), S0 (D), S1 (E), S2 (F), S3–4 (G), and
M (H) el e ments; spec i mens ZPAL cXVI/2641–2644, 2648, 2645, 2647, and 2646, re spec tively.
®

100 JERZY DZIK
Fig. 68. Polygnathids with plat form−bear ing P2 el e ment. A–K, O. Hemilistrona homoirregularis (Ziegler, 1971) from the L.styriacusZone at Ostrówka (A–K, sam ple Ost−12) in the Holy Cross Moun tains and Gołogłowy (O, sam ple Goł−3) in the Sudetes. P1 (A–G, J,O), P2 (H, I, K), el e ments; spec i mens ZPAL cXVI/2649, 2652, 2657, 2658, 2659, 2660, 2659, 2662–2668, and 2716, re spec tively.L–N. Hemilistrona pulchra Chauff and Dombrowski, 1977 from the L. styriacus (M, N, sam ple Ko−6) and the P. jugosus (L, sam ple
Ko−131) zones at Kowala. P1 (M, N) and P2 (L) el e ments, spec i mens ZPAL cXVI/2692 and 2678–2674, re spec tively.

Kalona. It is thus as sumed here that also the early pop u la tions of the lin eage from the C. marginifera Zone(velifer Zone) belong to this chronospecies.
Oc cur rence. — The C. marginifera to L. styriacus zones at Miedzianka, Kowala, Jabłonna, Ostrówka,and Gołogłowy.
Hemilistrona homoirregularis (Ziegler, 1971)(Figs 68A–K, O and 128)
Type ho ri zon and lo cal ity: Ma ple Mill Shale at road cut south east of Kolona in Iowa (Ziegler 1975).
Ma te rial. — 810 spec i mens.Di ag no sis. — Ma ture P1 and P2 el e ment with pos te rior plat form tongue bent down ward and or na mented
with tubercles.Re marks. — A rel a tively large col lec tion of this spe cies from sam ple Ost−12 con tains el e ments of var i −
ous ontogenetic ages. Ex cept for the ma ture P1 el e ments, they show a puz zling vari abil ity in vir tu ally all as −pects of the plat form shape and or na men ta tion. This is prob a bly the most vari able pop u la tion of polygnathids in the Famennian of the Holy Cross Mts. There is a tran si tion to morphologies clas si fied here as Hemilistrona margaritata, Polygnathus? experplexus and even Neopolygnathus communis. Un til pop u la tion and ap pa ra tus stud ies on much larger and tax o nom i cally less di verse ma te rial are per formed, this classification can betreated only as provisional.
Oc cur rence. — The P. trachytera to L. jugosus zones at Ostrówka, Miedzianka, Kowala and Jabłonna.
Hemilistrona pulchra Chauff et Dombrowski, 1977(Figs 68L–N and 128)
Type ho ri zon and lo cal ity: Basal shale mem ber of the Sul phur Springs For ma tion near Pevely, Mis souri, lower costatusZone (re worked ma te rial).
Ma te rial. — 14 spec i mens.Re marks. — Lon gi tu di nal ribs on the plat form of P2 el e ments char ac ter ize this spe cies. Out side Mis −
souri, it has been known from Thuringia (Helms 1961). In the Holy Cross Moun tains only a few el e ments
FAMENNIAN CONO DONTS AND AMMONOIDS 101
Fig. 69. Hemilistrona margaritata (Schäfer, 1976), a polygnathid with or nate plat form, from the L. styriacus Zone at Ostrówka(A–C, sam ple Ost−12; D, E, sam ple Ost−293) in the Holy Cross Moun tains; spec i mens ZPAL cXVI/2651, 2655, 2661, 2669, and
2674, re spec tively.

have been found and they hardly of fer any thing new re gard ing the ap pa ra tus com po si tion. In sam ple Ko−6two P1 el e ments of dif fer ent ontogenetic age are as so ci ated. Both show an un usu ally low and si nu soi dalblade and are lack ing a free ca rina (blade), sug gest ing that this is the end−mem ber of the lin eage, well rep re −sented in the material of Chauff and Dombrowski (1977).
Oc cur rence. — The L. styriacus and P. jugosus zones at Kowala.
Hemilistrona margaritata (Schäfer, 1976)(Fig. 69)
Type ho ri zon and lo cal ity: Bed 18 in trench dug at Dasberg near Hövel, Rhenish Slate Moun tains (Schäfer 1976).
Ma te rial. — 14 spec i mens.Re marks. — Lon gi tu di nal rows of tu ber cles on the plat form of lan ceo late P1 el e ments are typ i cal for the
spe cies. The mor phol ogy of this kind may be a part of pop u la tion vari abil ity of H. homoirregularis. How −ever, there seems to be a change in mor phol ogy of lon gi tu di nally or na mented P1 el e ments of Hemilistrona.In geo log i cally youn ger sam ples the tu ber cles are less prom i nent and the plat form more reg u larly lan ceo late,giv ing the P1 el e ments as pects some what sim i lar to Dasbergina kayseri.
Oc cur rence. — The L. styriacus Zone at Ostrówka.
Genus Neopolygnathus Vorontsova in Barskov et al., 1991
Type spe cies: Polygnathus communis Branson et Mehl, 1934 from the Tournaisian Bushberg sand stone at Brickeys, Mis souri.
Di ag no sis. — Plat form of P1 el e ments smooth at least in its dor sal part, oval in out line, usu ally a dis tinctly sep a rated ju ve nile basal cone lo cated in a de pres sion of the flat basal cav ity; high arched M element.
Re marks. — The de pres sion sep a rat ing lar val basal cone (in ter preted here as a dis tinct lar val stage pre −ced ing the de vel op ment of plat form) from the rest of the basal cav ity is seems to ap pear sev eral times in theevo lu tion of var i ous polygnathids and can hardly be used alone as the di ag nos tic char ac ter of the ge nus. Itdis ap peared in some late spe cies and is not char ac ter is tic of prob a ble old est pop u la tions of the main lin eage.The same re fers to the smooth, ro bust plat form. The con cept of the ge nus must thus re main pro vi sional un tilits or i gin and evo lu tion is elu ci dated. I sup pose that P. semicostatus is an ces tral and that a few paedomorphiclineages originated independently from it.
Ad vanced Tournaisian mem bers of the ge nus are homeomorphic in the de vel op ment of the plat form of P1el e ments to some spe cies of Lagovignathus gen. n.
Neopolygnathus communis (Branson et Mehl, 1934)(Figs 9B, 70A–S, and 130)
Type ho ri zon and lo cal ity: Tournaisian Bushberg sand stone at Brickeys, Mis souri.
Ma te rial. — 1,447 spec i mens.Di ag no sis. — Mar gins of the plat form of P1 el e ments raised adorally, with more or less dis tinct lon gi tu di −
nal crests in its ven tral area; only in ex treme spec i mens tuberculation of var i ous kinds may develop.Re marks. — Ac cord ing to Vorontzova (1996) pop u la tions of a nar row−plat form form of N. communis
from the K. crepida Zone are an ces tral to a clade of var i ously or na mented forms united only by their de −pressed basal pit. I am in clined rather to the al ter na tive in ter pre ta tion, that the sep a ra tion of pre−plat formstage from the fast grow ing plat form mar gin ally is rather a de vel op men tal as pect not re quir ing fun da men talchange in con trol ling mech a nisms and thus ap pear ing in de pend ently in un re lated lin eages. This may be onlyan ex pan sion of the lar val stage (sinuosa larva), poorly con trolled by de vel op men tal reg u la tory mech a nisms,as sug gested by great pop u la tion vari abil ity in early pop u la tions. The range of vari abil ity in cludes forms with nar row and wide plat forms, com pletely smooth and or na mented with trans verse ridges, with out any crests,bear ing crests par al lel to the blade or at an an gle to it. This morphologic lability ceases in the lat estFamennian, where a morphologic gap de vel ops be tween N. communis and N. vogesi.
Oc cur rence. — The geo log i cally old est el e ments of Neopolygnathus mor phol ogy have been en coun −tered in the Holy Cross Moun tains in the C. quadrantinodosa Zone at Łagów (sam ple Ł−27) where theyco−oc cur with Polygnathus semicostatus and may grade into its mor phol ogy, and in the C. marginifera Zoneat Jabłonna. Un ques tion able re cords of the spe cies start from the P. jugosus Zone of all stud ied lo cal i ties andcon tinue into the Carboniferous.
102 JERZY DZIK

Neopolygnathus vogesi (Ziegler, 1962)(Figs 70T–X and 130)
Type ho ri zon and lo cal ity: Sam ple 7 from the late Famennian mid dle costatus Zone at the Hönnetalstraße sec tion in theRhenish Slate Moun tains (Ziegler 1962).
Ma te rial. — 595 spec i mens.Di ag no sis. — Plat form of P1 el e ments with prom i nent oblique crests in the ven tral part.Re marks. — Pop u la tions of this spe cies show a rather sta ble mor phol ogy, sim i lar to co−oc cur ring P.
communis, but un like early pop u la tions of the latter. In a few sam ples from the P. jugosus Zone at Kowala rare ju ve nile spec i mens of a polygnathid have been
found (Fig. 70U, V) of the kind iden ti fied by Perri and Spaletta (1991) with the Tournaisian Polygnathusmargininvolutus Gedik, 1969. In the tri an gu lar shape of the strongly con cave thin plat form these el e mentsare sim i lar to ju ve niles of P. semicostatus. Sim i lar forms oc cur in the Tournaisian, where they prob a bly rep −re sent ju ve niles of Neopolygnathus vogesi (Dzik 1997, fig. 12F). The avail able ma te rial is too small to besure of their nature.
Oc cur rence. — The P. jugosus to D. trigonica zones at Ostrówka and prob a bly Kowala in the HolyCross Moun tains and at Dzikowiec in the Sudetes. Con tin ues into the Car bon if er ous.
FAMENNIAN CONO DONTS AND AMMONOIDS 103
Fig. 70. Famennian pop u la tions of Neopolygnathus. A–S. N. communis (Branson et Mehl, 1934) from the C. quadrantinodosaZone at Łagów (A, B, sam ples Ł−27), the early L. styriacus Zone (C–F, sam ple Ost−12) at Ostrówka, the P. trachytera Zone atKowala (G–J, sam ple Ko−15), the late L. styriacus (sam ple Ost−293, K–N) and the P. jugosus (O–S, sam ple Ost−185) zones atOstrówka in the Holy Cross Moun tains. P1 (A–Q), P2 (R), and M (S) el e ments; spec i mens ZPAL cXVI/2527, 2528, 2693–2696,2518, 2510, 2512, 2513, 2675–2677, 2674, 2703, 2704, 2706, 2708, re spec tively. T–X. N. vogesi (Ziegler, 1962) from the P.jugosus Zone at Ostrówka (T, X, sam ple Ost−185) and Kowala (U, V, sam ple Ko−191) in the Holy Cross Moun tains and
Dzikowiec (sam ple Dz 54, W) in the Sudetes. P1 el e ments; spec i mens ZPAL cXVI/2701, 2994, 2993, 2707, and 2988.

Genus Lagovignathus gen. n.Type spe cies: Polygnathus lagowiensis Helms et Wolska 1967 from the Łagów Beds at Łagów, the Holy Cross Moun tains.
Di ag no sis. — Plat form of P1 el e ments thin, its sur face or na mented with min ute tu ber cles or smooth, lan −ceo late in out line, low arched M element.
Re marks. — The ear li est spe cies of the new ge nus, L. glaber, re sem bles some Car bon if er ous spe cies ofNeopolygnathus (e.g., N. purus) in hav ing a flat and com pletely smooth plat form. It is dif fi cult to di ag nosethis ge nus in a way al low ing its easy sep a ra tion from such forms. The lin eage ap par ently orig i natedpaedomorphically and the P1 el e ments are sec ond arily sim pli fied. This change took place much ear lier thanin the lin eage of N. communis and it is un likely that they are di rectly re lated. How ever, it has to be ad mit tedthat an ces try of both lin eages remains to be supported with evidence.
Lagovignathus glaber (Ulrich et Bassler, 1926)(Figs 71A–L and 131)
Type ho ri zon and lo cal ity: Hardin Sand stone at Mount Pleas ant, Ten nes see.
Ma te rial. — 749 spec i mens.Di ag no sis. — Heart−shaped out line of flat, smooth plat form with mar gins gently raised ab oral ly.Re marks. — The ap pa ra tus com po si tion was re stored by Metzger (1994). It is here as sumed that the type
spec i mens of L. glaber and L. bilobatus rep re sent modal morphologies of their pop u la tions, which are suc −ces sive parts of the same lin eage. The plat form out line, orig i nally used to dis tin guish these spe cies is highlyvari able and iden ti fi ca tion of the spe cies should not be based on sin gle spec i mens. Prob a bly equally use ful isthe chang ing mor phol ogy of P2 el e ments, which be came more arched in L. bilobatus.
Oc cur rence. — The K. crepida Zone at Jabłonna, Kadzielnia, and Wietrznia.
Lagovignathus bilobatus (Ziegler, 1962)(Figs 71M–T and 131)
Type ho ri zon and lo cal ity: Bed 11 in trench II at Sessaker near Oberscheld. Ma te rial. — 3,891 spec i mens.Di ag no sis. — Slight con stric tion in the mid dle of the heart−shaped out line of the flat, smooth plat form.Re marks. — This is clearly a suc ces sor of L. glaber and whether to dis tin guish them at the chronospecies
or chronosubspecies rank is a mat ter of con ve nience. Dif fer ences are mi nor and ranges of pop u la tion vari −abil ity clearly overlap.
Oc cur rence. — The late K. crepida to L. styriacus zones at Kadzielnia, Jabłonna, Kowala, Miedzianka,and Ostrówka.
Lagovignathus fallax (Helms et Wolska, 1967)(Figs 72J–Q and 131)
Type ho ri zon and lo cal ity: Grey, bed ded lime stone of lower marginifera Zone from the old Geipelsch quarry in Schleiz,Thuringia (Helms and Wolska 1967).
Ma te rial. — 1,273 spec i mens.Di ag no sis. — Wil low leaf−shaped plat form or na mented with min ute tu ber cles.Re marks. — Rare spec i mens from the C. quadrantinodosa Zone at Łagów (sam ples Ł−27 and Ł−26), as −
so ci ated with Polynodosus transitus and prob a bly be long ing to that spe cies (Fig. 50D), re sem ble L. fallax inthe min ute tuberculation of the plat form. The reg u lar denticulation and short dor sal pro cess of P2 el e mentsmake any re la tion ships of this spe cies to Polynodosus unlikely.
Basal cone of the plat form−lack ing larva is sig nif i cantly larger in this spe cies than in other co eval spe ciesof Lagovignathus, but in sub se quent growth the bound ary be tween con i cal and in verted basal cav itysmoothened.
Oc cur rence. — The C. marginifera Zone at Łagów, Kowala, and Miedzianka.
Lagovignathus? dissimilis (Helms et Wolska 1967)(Figs 73 and 131)
Type ho ri zon and lo cal ity: Grey−pur ple, thin bed ded lime stone of the Clymenia Zone from Alte Heerstrasse near Oetters −dorf, Thuringia (Helms and Wolska 1967).
104 JERZY DZIK

Ma te rial. — 513 spec i mens.Di ag no sis. — P2 el e ments of ro bust ap pear ance, with in cip i ent plat form, P1 el e ments with usu ally nar row
but ro bust plat form and with ex ten sive asym met ric basal cone.Re marks. — Dense denticulation of ju ve nile P1 el e ments some what re sem bles peniculus and to gether
with the shape of the plat form in P1 el e ments may sug gest af fin i ties with Polynodosus (es pe cially P.lauriformis). No tran si tional forms have been found, how ever. The basal cone, which cor re sponds to an ex −panded sinuosa larva, points to Lagovignathus fallax as the al ter na tive an ces tor. The shape of the plat formand out line of ma ture P1 el e ments fits this pos si bil ity even better. Prob a ble M el e ment is also un like that ofPolynodosus.
FAMENNIAN CONO DONTS AND AMMONOIDS 105
Fig. 71. Spe cies of Lagovignathus with smooth plat form from the Holy Cross Moun tains. A–L. Lagovignathus glaber (Ulrich etBassler, 1926) from the K. crepida Zone at Kadzielnia (A–C, E, sam ple Ka−14; D, F–L, sam ple Ka−4). P1 (A–F), P2 (G), S0 (H),S1 (I), S2 (J), S3–4 (K), and M (L) el e ments; spec i mens ZPAL cXVI/2529, 2532, 2530, 2533, 2531, and 2534–2540, re spec tively.M–T. Lagovignathus bilobatus (Ziegler, 1962) from the C. quadrantinodosa Zone at Łagów (sam ple Ł−32). P1 (M, N), P2 (O),S0 (P), S1 (Q), S2 (R), S3–4 (S), and M (T) el e ments; spec i mens ZPAL cXVI/2542, 2541, 2543, 2547, and 2549, re spec tively.

The evo lu tion ary change with re spect to L. fallax was the prob a ble fur ther ex ten sion of the sinuosa larvastage and de vel op ment of strong asym me try of the basal cone in its P1 el e ment. As a re sult, a sit u a tion de vel −oped closely sim i lar to that in ad vanced spe cies of the palmatolepidid Tripodellus lineage.
106 JERZY DZIK
Fig. 72. Early spe cies of Lagovignathus with tuberculated plat form from the Holy Cross Moun tains. A–I. Lagovignathuslagowiensis (Helms et Wolska, 1967) from the C. quadrantinodosa Zone at Łagów (sam ple ŁSł73−5). P1 (A–C), P2 (D), S0 (E),S1 (H), S2 (F), S3–4 (I), and M (G) el e ments; spec i mens ZPAL cXVI/2552, 2550, 2551, 2553, 2554, 2556, 2558, 2555, and 2557,re spec tively. J–Q. Lagovignathus fallax (Helms et Wolska, 1967) from slightly older part of the same zone at the same lo cal ity(sam ple ŁSł73−2). P1 (J, K, S, T), P2 (L), S0 (M), S1 (N, O), S2 (P), S3–4 (Q), and M (R) el e ments; spec i mens ZPAL cXVI/2562,
2561, 2563, 2564, 2566, 2565, 2568, 2569, 2560, and 2559, re spec tively.

Oc cur rence. — The L. styriacus to P. jugosus zones at Kowala, Ostrówka, Jabłonna and Dzikowiec.
Lagovignathus lagowiensis (Helms et Wolska, 1967)(Figs 72A–I and 131)
Type ho ri zon and lo cal ity: Sacculus bank with Cheiloceras lagowiense and Dimeroceras lentiforme, mixed up per mostrhomboidea and up per marginifera zones ac cord ing to Helms and Wolska (1967).
Ma te rial. — 3,379 spec i mens.Di ag no sis. — Leaf−like, rel a tively wide plat form of P1 el e ments or na mented with small tu ber cles.Re marks. — The spe cies is pre ceded in the Łagów sec tion by the some what sim i lar L. fallax but tran si −
tion be tween these spe cies has not been dem on strated. A much more prob a ble or i gin from L. glaber wouldre quire only de vel op ment of tuberculation of the plat form. This is ac tu ally the only ap par ent dif fer encebetween these species.
Oc cur rence. — The late C. marginifera Zone at Łagów, Kowala, and Miedzianka.
Lagovignathus granulosus (Branson et Mehl, 1934)(Figs 74A–C and 131)
Type ho ri zon and lo cal ity: Famennian shale at Noel, Mis souri (G. Klapper per sonal com mu ni ca tion 2005).
Ma te rial. — 39 spec i mens.Di ag no sis. — Leaf−like, rel a tively wide plat form of P1 el e ments densely or na mented with rel a tively large
tubercles.Re marks. — The spe cies is so sim i lar to the sig nif i cantly older L. lagowiensis that, de spite the dis con tin −
u ous fos sil re cord at Łagów, the an ces tor−de scen dant re la tion ship seems al most cer tain. Spec i mens withsmooth con cave ar eas near the ven tral end of the plat form are tran si tional to L. styriacus, the suc ces sor of L.granulosus in the same lineage.
Oc cur rence. — The P. trachytera Zone at Łagów, Miedzianka, Jabłonna, Kowala, and Wzdół.
Lagovignathus styriacus (Ziegler, 1957)(Figs 74D–F and 131)
Type ho ri zon and lo cal ity: Sam ple 73 from a nod u lar goniatite lime stone col lected in a quarry be tween Steinberg andHitzendorf near Graz, Aus tria.
Ma te rial. — 287 spec i mens.Di ag no sis. — Wide and asym met ric plat form of P1 el e ment with densely tuberculated dor sal part sep a −
rated from smooth ven tral ar eas by a trans verse row of larger tubercles.Re marks. — Early pop u la tions of L. styriacus show great vari abil ity in their plat form out line, some spec −
i mens be ing nar rower even than in the an ces tral L. granulosus. The tran si tion ap par ently took a sig nif i canttime span, the zonal bound ary is thus hardly clear−cut. It is pos si ble that some pop u la tions from Ostrówka
FAMENNIAN CONO DONTS AND AMMONOIDS 107
Fig. 73. Lagovignathus? dissimilis (Helms et Wolska 1967) from the P. jugosus Zone at Jabłonna (sam ple J−51) in the HolyCross Moun tains. P1 (A–C), P2 (D, E), and M (F) el e ments; spec i mens ZPAL cXVI/2172–2177, re spec tively.

co−oc cur ring with P. trachytera ac tu ally rep re sent L. granulosus, not L. styriacus as sug gested by theirvariable morphology.
Oc cur rence. — The nom i nal zone at Jabłonna, Ostrówka, Kowala, and Miedzianka.
Family Ancyrognathidae fam. n.Di ag no sis. — Ro bust and sharply denticulated el e ments with clearly ter mi nal growth, P1 el e ment with re −
duced free blade in adult spec i mens, S1–2 el e ments bear short pro cesses, M el e ment with prom i nent and welldenticulated external process.
Re marks. — The mor phol ogy of the ancyrognathid ap pa ra tus el e ments is in many re spects tran si tionalbe tween that of the Polygnathidae and Palmatolepididae. Like the palmatolepidids, pro cesses of el e ments ofAncyrognathus are rel a tively short and with the clearly ter mi nal growth. Mesotaxis is ap par ently the an ces torof the palmatolepidids al though, ex cept for the M el e ment, it has not de vel oped all their di ag nos tic char ac ters (sym met ri cal el e ment lack ing me dial pro cess, lobate plat form of P1 elements).
Gen era in cluded. — Mesotaxis Klapper and Philip, 1972; Ancyrognathus Branson et Mehl, 1934;Ancyrolepis Ziegler, 1959; Polylophodonta Branson et Mehl, 1934.
Genus Ancyrognathus Branson et Mehl, 1934Type spe cies: Ancyrognathus symmetricus Branson et Mehl, 1934 from the Famennian Saverton Shale at Sees Creek in
Mon roe County, Mis souri (Ziegler 1975).
Di ag no sis. —Elon gated plat form in P1 el e ments, tend ing to de velop a prom i nently tuberculation; trira −mous in early spe cies; other el e ments of the ap pa ra tus tend to de velop a ru di men tary plat form but still of agen er al ized polygnathid morphology.
Ancyrognathus sinelamina (Branson et Mehl, 1934)(Figs 75A–H and 132)
Type ho ri zon and lo cal ity: Famennian shale at Dixie, Mis souri (Ziegler 1977, p. 313).
Ma te rial. — 22 spec i mens.Di ag no sis. — Slen der P1 el e ments vir tu ally with out free blade and any flank lobe of the plat form; min ute
basal cone (pit).Re marks. — The tran si tion from the biramous lat est Frasnian ancyrognathids to this spe cies has been
doc u mented by Schülke (1996); prob a bly the early Famennian type spe cies of the ge nus de rived at the tran si −
108 JERZY DZIK
Fig. 74. Late spe cies of Lagovignathus with tuberculated plat form from the Holy Cross Moun tains. A–C. Lagovignathusgranulosus (Branson et Mehl, 1934) from the P. trachytera Zone at Jabłonna (bed 24). P1 el e ments; spec i mens ZPAL cXVI/2709,2711, and 2710, re spec tively. D–F. Lagovignathus styriacus (Ziegler, 1957) from its nom i nal Zone at Miedzianka (D, sam ple
Md−6) and Ostrówka (E, F, sam ple Ost−265). P1 el e ments; spec i mens ZPAL cXVI/2714, and 2712, 2713, re spec tively.

tion. Schülke (1999) at tempted res to ra tion of the ap pa ra tus but only iden ti fi ca tion of P1 el e ments is sup ported here. Ramiform el e ments sim i lar to those as so ci ated with Frasnian spe cies of Ancyrognathus (Dzik 2002)co−oc cur with plat form el e ments of A. sinelamina in the Holy Cross Mountains.
Oc cur rence. — The late K. triangularis and early K. crepida zones at Jabłonna, Wietrznia, Kadzielnia,and Miedzianka. The spe cies oc curs also in the Dębnik area (Baliński 1995).
Ancyrognathus sp. n.(Figs 9E, 76, and 132)
Ma te rial. — 28 spec i mens.Re marks. — In the early Famennian of the Holy Cross Moun tains rare plat form el e ments sim i lar to those
of Dasbergina kayseri oc cur, dif fer ent from those from their late Famennian type ho ri zon in a less reg u larout line and coarser tuberculation of the plat form. How ever, rare as so ci ated ramiform el e ments prob a bly be −long ing to the spe cies sup port its af fin i ties with Polylophodonta and Ancyrognathus. Per haps this is a suc ces −sor of A. sinelamina.
Oc cur rence. — The K. crepida Zone at Jabłonna and Kadzielnia.
Genus Polylophodonta Branson et Mehl, 1934
Type spe cies: Polygnathus gyratilineatus Holmes, 1928 from the Gassaway Mem ber of the Chat ta nooga Shale, Al a bama.
Di ag no sis. — Wide oval plat form in P1 el e ments; M el e ment pre serves gen er al ized polygnathid mor phol −ogy.
Re marks. — There are some dif fi cul ties with de lin eat ing the bound ary be tween early spe cies of theancyrognathid Polylophodonta and homeomorphic ad vanced plat form−be ring mem bers of the Mehlina lin −eage (Polynodosus) if only plat form el e ments are considered.
FAMENNIAN CONO DONTS AND AMMONOIDS 109
Fig. 75. Typ i cal ancyrognathids from the Holy Cross Moun tains, Ancyrognathus sinelamina (Branson et Mehl, 1934) from thelate K. triangularis Zone at Karczówka (A, B), the K. crepida Zone at Jabłonna (C–F, H, I, bed 3) and Kadzielnia (G, sam pleKa−2). P1 (A–D), P2 (H), S0 (E), S3–4 (F), and M (G, I) el e ments; spec i mens ZPAL cXVI/2604–2607, 2609, 2610, 2612, 2608, and
2611, re spec tively.

Polylophodonta ovata (Helms, 1961)(Figs 77 and 132)
Type ho ri zon and lo cal ity: Lower bed of red lime stone, lower Cheiloceras Zone, in west ern quarry on Kahlleithe nearRödersdorf, Thuringia (Helms 1961).
Ma te rial. — 289 spec i mens.Di ag no sis. — Row of denticles of the blade of rec og niz able to the dor sal end of the plat form in P1 el e −
ments even in ma ture spec i mens.Re marks. — The old est oc cur rence of the spe cies is in bed 4 at Jabłonna, where it oc curs to gether with
Klapperilepis crepida prior to the en trance of Conditolepis tenuipunctata. El e ments from there are of a rather ro bust and ir reg u lar ap pear ance. The typ i cal form oc curs in the some what youn ger sam ples J−45a andWtr−32.
Oc cur rence. — The late K. triangularis and early K. crepida Zone at Jabłonna, Wietrznia, and Mie dzianka. The spe cies oc curs also in the Dębnik area, iden ti fied there as P. confluens by Baliński (1995).
Polylophodonta pergyrata (Holmes in Butts, 1926)(Figs 78 and 132)
Type ho ri zon and lo cal ity: Gassaway Mem ber of the Chat ta nooga Shale at Quicks Mill, Al a bama (Hud dle 1968).
Ma te rial. — 559 spec i mens.Di ag no sis. — Con cen tric ridges form ing a fin ger print−like ap pear ance of the ma ture plat form of P1
elements.Re marks. — Many spe cies names are avail able for ad vanced Polylophodonta. Al though it can not be ex −
cluded that sev eral bi o log i cal spe cies are rep re sented in the stud ied ma te rial, I am not able to find any dis con −ti nu ity in the ob served pop u la tion variability.
Oc cur rence. — The mid K. crepida to P. trachytera zones at Jabłonna, Kowala, Kadzielnia, Łagów,Miedzianka, and Ostrówka.
110 JERZY DZIK
Fig. 76. Ancyrognathus sp. n. a rel ict ancyrognathid from the K. crepida Zone at Jabłonna (A–D, sam ple J−65) and Kadzielnia(E–G, sam ple Ka−5) in the Holy Cross Moun tains.

Family Palmatolepididae Sweet, 1988Di ag no sis. — Ap pa ra tus with M el e ment hav ing straight pro cesses and S0 el e ment with more or less re −
duced me dial pro cess; usu ally lobate plat form−bear ing P1 and P2 elements.
Genus Klapperilepis Dzik, 2002
Type spe cies: Palmatolepis triangularis Sannemann, 1955 from the early Famennian of Frankenwald.
Di ag no sis. — P1 el e ments with raised tip of the dor sal pro cess and un de rived ap pear ance of the re main −ing el e ments of the apparatus.
Re marks. — Dis crim i na tion of early sympatric spe cies of Klapperilepis is a dif fi cult task and while dis −cuss ing their tax on omy in the con text of the end−Frasnian events I failed to find a sat is fac tory so lu tion (Dzik
FAMENNIAN CONO DONTS AND AMMONOIDS 111
Fig. 77. De rived ancyrognathid Polylophodonta ovata (Helms, 1961) from the early K. crepida Zone at Wietrznia (A, D, sam pleWtr−32) and the K. triangularis Zone at Jabłonna (B, E, H, I, J, sam ple J−45a; F, G, sam ple J−44) in the Holy Cross Moun tains.P1 (A–D), P2 (E), S0 (F), S1 (H), S2 (G), S3–4 (I), and M (J) el e ments; spec i mens ZPAL cXVI/2579, 2582, 2580, 2581, 2583,
2584–2588, re spec tively.

112 JERZY DZIK
Fig. 78. De rived ancyrognathid Polylophodonta pergyrata (Holmes in Butts, 1926) from the K. crepida Zone at Kadzielnia (A,H, sam ple Ka−12; B, D, K, sam ple Ka−5; G, J, and M–R, sam ple Ka−2) and Jabłonna (C, E, F, and L, bed 7), and the C.quadrantinodosa Zone at Łagów (I, sam ple Ł−26) in the Holy Cross Moun tains. P1 (A–E, L, O, Q, R), P2 (I, K), S1 (N), S2 (M),S3–4 (P), and M (F–H) el e ments; spec i mens ZPAL cXVI/2596, 2594, 2573, 2595, 2574, 2575, 2613, 2597, 2159, 2617, 2591,
2572, 2619, 2614, 2618, 2620, and 2615, 2616, re spec tively.

2002). No doubt that sev eral new lin eages emerged soon af ter the tax o nomic di ver sity of the palmatolepididswas re duced to the sin gle spe cies, K. ul tima.
This is proven by the oc cur rence of two dis tinct types of M el e ments in sam ple Pł−42: one of the prim i tivelat est Frasnian mor phol ogy, the other of an ap pear ance sim i lar to that of K. quadrantinodosolobata. Otherramiform el e ments of the Klapperilepis ap pa ra tus do not form any dis crete classes and are prob a bly in dis tin −guish able at the spe cies level. It seems rea son able to as sume that the P1 el e ments mor pho log i cally clos est toK. ul tima be long to the same ap pa ra tus as the as so ci ated prim i tive M el e ments. Plat form el e ments of this kind show a rel a tively sin u ous course of the blade. Ju ve niles may re sem ble Conditolepis tenuipunctata orKlapperilepis circularis (in Dzik 2002, ta ble 2 they were pro vi sion ally listed un der these names). Some whatar bi trarily this pop u la tion is here con sid ered to be the old est K. triangularis.
FAMENNIAN CONO DONTS AND AMMONOIDS 113
Fig. 79. An ces tor of Famennian palmatolepidids, Klapperilepis ul tima (Ziegler, 1958) from the pre−K. triangularis Zone strata at Płucki (sam ple Pł−36) in the Holy Cross Moun tains. P1 (A–L), P2 (M–Q), S0 (R, S), S1 (T), S2 (U), S3–4 (V, W), and M (X) el e −ments; spec i mens ZPAL cXVI/1512, 1503, 1506, 1513, 1509, 1505, 1502, 1510, 1508, 1511, 1507, 1504, 1514, 1518, 1516,
1517, 1515, and 1519–1525, re spec tively.

Us ing the same line of rea son ing one would ex pect that P1 el e ments re sem bling those of K. spathula insam ple Pł−42 are conspecific with the ad vanced M el e ments. Al though there is no ap par ent gap be tween therange of its morphologic vari abil ity and that of as so ci ated K. triangularis, such an in ter pre ta tion is con sis tentwith the ap pa ra tus re con struc tion pro posed by Schülke (1999) for the spe cies here named K. schuelkei sp. n.
The re main ing morphotype of plat form el e ments in sam ple Pł−42 is much more clearly de lim ited. In hav −ing a rhomboidal out line of the smooth plat form with raised mar gins and a straight blade it closely re sem blesTripodellus clarki and prob a bly rep re sents its early pop u la tion. There is a grad ual tran si tion to wardsmorphologies with a wide plat form re sem bling K. delicatula.
Klapperilepis ultima (Ziegler, 1958)(Figs 8C, 79, and 135)
Type ho ri zon and lo cal ity: Lat est Frasnian sam ple 27 from Sessacker II trench (Ziegler 1958).
Ma te rial. — 2,084 spec i mens (1,207 from the Famennian).Di ag no sis. — Rel a tively wide and short plat form of P1 el e ments, un de rived ap pear ance of A−shaped M
elements.Re marks. — There is a grad ual evo lu tion ary change in the mor phol ogy of P1 el e ments ob scured by a
wide pop u la tion vari abil ity (Dzik 2002). This vari abil ity makes the lin eage to be of lit tle cor re la tive value.The ap pa ra tus of the spe cies is doc u mented with clus ters of el e ments from the ear li est Famennian at Płucki
114 JERZY DZIK
Fig. 80. Palmatolepidids of the Klapperilepis triangularis (Sannemann, 1955) lin eage from its nom i nal zone strata at Płucki in theHoly Cross Moun tains. A–G. K. triangularis from sam ple Pł−42. H–R. Prob a ble an ces tral pop u la tion of Palmatolepis initialis sp. n.(sam ple Pł−1). P1 (A, B, D, H–L), P2 (C and M), S0 (N), S1 (E and O), S2 (P), S3–4 (F and Q), and M (G and R) el e ments; spec i mensZPAL cXVI/1232, 1233, 1235, 1234, 1237–1239, 1344, 1339, 1340, 1341, 1343, 1345, 1346–1348, and 1350, 11351, re spec tively.

(sam ples Pł−20 and Pł−16), some rep re sent ing sig nif i cant por tions of the ap pa ra tus (Dzik 2002). Palmatolepis praetriangularis Ziegler and Sandberg, 1988 has been pointed out by Klapper et al. (2004) to be a ju nior sub −jec tive syn onym of P. ultima.
Oc cur rence. — The or i gin of the lin eage re mains cryp tic. Pop u la tions from the lat est Frasnian do notseem to dif fer from those from the be gin ning of the Famennian. At Płucki spec i mens with some what elon −gated plat form clas si fied within the suc ceed ing K. triangularis ap pear in a greater num ber in sam ple Pł−42(note, how ever that this char ac ter is size−de pend ent), at Kowala in sample Ko−155.
Klapperilepis triangularis (Sannemann, 1955)(Figs 80A–G, 81A–J, and 135)
Type ho ri zon and lo cal ity: Nod u lar lime stone at Schübelebene in Frankenwald (Sannemann 1955b; Ziegler 1975).
Ma te rial. — 422 spec i mens.Di ag no sis. — P1 el e ments rel a tively elon gated and M el e ments usu ally with rel a tively wide an gle be −
tween processes.Re marks. — P2 el e ments of the ap pa ra tus were iden ti fied by van den Boogaard and Kuhry (1979); the
com plete ap pa ra tus was re stored by Schülke (1999).This is a vari able spe cies, both in re spect to the out line of plat form in P1 el e ments, which ranges from that
typ i cal to K. ul tima to that of early spe cies of Palmatolepis, but also in the prom i nence of the cusp and cur va −ture of P2 el e ments. The mean value of elon ga tion in dex of the P1 el e ments is, how ever, dis tinctly higher than in K. ul tima (Dzik 2002, 2004), re main ing el e ments of the ap pa ra tus be ing hardly dis tin guish able. In fact, todis tin guish these chronospecies a large sam ple is needed and singler el e ments are of lit tle tax o nomic value.
FAMENNIAN CONO DONTS AND AMMONOIDS 115
Fig. 81. Spe cies of the palmatolepidid Klapperilepis from the early Famennian of the Holy Cross Moun tains. A–J. K. trian −gularis (Sannemann, 1955) from its nom i nal Zone strata at Kowala (A, B, H–J, sam ple Ko−153; C–G, sam ple Ko−154).P1 (A–D), P2 (E, F), S0 (G), S1 (H), S3–4 (I), and M (J) el e ments; spec i mens ZPAL cXVI/2732, 2733, 2727, 2728, 2729–2731, and2734–2736, re spec tively. K–N. Klapperilepis regularis (Coo per, 1931) from the late K. triangularis Zone at Karczówka (K, L)and the K. crepida Zone at Kadzielnia (M, Ka−12) and Miedzianka (N, sam ple Md−1). P1 (K, L) and P2 (M, N) el e ments; spec i −
mens ZPAL cXVI/2919, 2820, 2818, and 2955.

Some late pop u la tions (e.g., Pł−1; Fig. 80H–R) show ro bust denticulation of ramiform el e ments and widelyan gu lar pro cesses of the M el e ment typ i cal of Palmatolepis initialis sp. n.
Oc cur rence. — Zone of its own at Płucki, Karczówka, Kowala, and Wietrznia.
Klapperilepis regularis (Cooper, 1931)(Figs 81K–N and 134)
Type ho ri zon and lo cal ity: Wood ford Shale of the Arbuckle Moun tains, Oklahoma (Coo per 1931).
Ma te rial. — 222 spec i mens.Di ag no sis. — Sin u ously curved P1 el e ments with subparallel mar gins of wide plat form; an gu larly bent P2
el e ments with low denticles and pos te ri orly wide platform.Re marks. — Ap pa ra tus was par tially re stored by Schülke (1999) with the M el e ment in di cat ing a re la −
tion ship to Klapperilepis triangularis. P2 el e ments of the same mor phol ogy as those from the MontagneNoire oc cur in the Holy Cross Mountains.
Oc cur rence. — The late K. triangularis and K. crepida zones at Jabłonna, Karczówka, Kadzielnia andKowala.
Klapperilepis protorhomboidea (Sandberg et Ziegler, 1973)(Figs 82A–C and 134)
Type ho ri zon and lo cal ity: 7.5–8.5 m above the base of the West Range Lime stone at Bactrian Moun tain, Ne vada.
Ma te rial. — 32 spec i mens.Di ag no sis. — Plat form in min ute P1 el e ment of vari able, more or less rhomboidal out line and with ir reg u −
larly dis trib uted tu ber cles, mostly on the ven tral an te rior quadrant.Re marks. — Ap pa ra tus was re stored by Schülke (1995). The spe cies changes into K. rhomboidea but the
tran si tion is rather un clear be cause of the great pop u la tion vari abil ity and gen er ally low num ber of el e mentsin studied samples.
Oc cur rence. — The K. triangularis Zone at Płucki, Kowala, Wietrznia and Jabłonna.
Klapperilepis rhomboidea (Sannemann, 1955)(Figs 82D–H and 134)
Type ho ri zon and lo cal ity: Nod u lar lime stone of up per Cheiloceras Stufe at Kirchgattendorf in Frankenwald (Sannemann1955b; Ziegler 1975).
Ma te rial. — 557 spec i mens.Di ag no sis. — Plat form in min ute P1 el e ment of an gu larly oval out line, with out pos te rior lobe, smooth.Re marks. — Metzger (1994) pro posed a P2 el e ment iden ti fi ca tion that seems un likely to be cor rect. The
spe cies is well rep re sented in some sam ples by P1 el e ments but other el e ments of the ap pa ra tus are dif fi cult to col lect be cause of their small size. They are iden ti fied as be long ing to this spe cies mostly on the ba sis of theirun usu ally prim i tive mor phol ogy com pared with the age of the sample.
116 JERZY DZIK
Fig. 82. Palmatolepidids of the K. rhomboidea lin eage from the Holy Cross Moun tains. A–C. Klapperilepis protorhomboidea(Sandberg et Ziegler, 1973) from the mid K. triangularis Zone at Jabłonna (sam ple J−54). P1 el e ments; spec i mens ZPALcXVI/2737–2739. D–H. Klapperilepis rhomboidea (Sannemann, 1955) from the C. quadrantinodosa Zone at Łagów (sam ple
Ł−32). P1 (D), P2 (E), S2 (F), and M (G, H) el e ments; spec i mens ZPAL cXVI/1370–1375, re spec tively.

FAMENNIAN CONO DONTS AND AMMONOIDS 117
Fig. 83. Palmatolepidids of the K. circularis lin eage. A–M. Klapperilepis wolskae (Ovnatanova, 1969) from the late K. trian −gularis Zone at Wietrznia (A–D, F–H, J, K, and M, sam ple Wtr−27) and Karczówka (E, I, and L) in the Holy Cross Moun tains.P1 (A–H), P2 (I, J), S2 (K), and M (M) el e ments; spec i mens ZPAL cXVI/1449, 2795, 1450, 2949, 1451, 2948, 2806, 2796, 2950,1452, 1453, 2951, 1454, and re spec tively. N–T. Klapperilepis circularis (Szulczewski, 1971) from the K. crepida Zone atMiedzianka (N, S, T, sam ple Md−1), Jabłonna (O, P, bed 7), and Łagów? (S, T, sam ple Ł−Mec). P1 (N–Q), P2 (R, S), and S1 (T)
elements; spec i mens ZPAL cXVI/2952, 2958, 2959, 2956, 2957, and 2953, 2954, re spec tively.

Oc cur rence. — The late K. crepida Zone (with C. klapperi) at Kadzielnia and the C. quadrantinodosaZone at Kowala, Łagów, and Miedzianka. The spe cies oc curs also in the Dębnik area (Baliński 1995).
Klapperilepis wolskae (Ovnatanova, 1969)(Figs 83A–M and 133)
Type ho ri zon and lo cal ity: Sandonsk stage at depth 375.4–377.8 m of bore hole Ilmen in Woronezh re gion, Rus sia; crepidaZone (Ziegler 1975).
Ma te rial. — 236 spec i mens.Di ag no sis. — Oval plat form of P1 el e ments with prom i nent nar row ex ten sion of the pos te rior lobe.Re marks. — El e ments of the kind at trib uted to the ap pa ra tus of this spe cies by Metzger (1994) do not oc −
cur in the Holy Cross Moun tains (per haps this is a par tial ap pa ra tus of a new spe cies of Syncladognathus) and a dif fer ent com po si tion is sug gested here, al though the ev i dence re mains very weak. P1 el e ments are ex −tremely vari able in Wietrznia (Wtr−27), rang ing from an out line re sem bling that of K. triangularis to that ofK. circularis (Fig. 83D, H).
Oc cur rence. — The late K. triangularis and the early K. crepida zones at Płucki, Wietrznia, Karczówka,and Jabłonna.
Klapperilepis circularis (Szulczewski, 1971)(Figs 83N–T and 133)
Type ho ri zon and lo cal ity: Sam ple X.4 from a bed of cephalopod lime stone at the Kadzielnia quarry with early K. ter mini,the K. crepida Zone, Holy Cross Moun tains.
Ma te rial. — 312 spec i mens.Di ag no sis. — Subcircular out line of plat form of P1 el e ments with short and wide ex ten sion of the pos te −
rior lobe and blunt dor sal tip.Re marks. — El e ments of the spe cies are gen er ally much smaller than those of the an ces tral K. wolskae,
even at a stage of ap par ent ma tu rity. The main dis tin guish ing char ac ter is the rounded dor sal tip of the plat formand strongly curved blade. No other ap pa ra tus el e ments have been iden ti fied with reasonable confidence.
Oc cur rence. — The K. crepida Zone at Kadzielnia, Jabłonna, Wietrznia, Miedzianka, and Kowala. Thespe cies oc curs also in the Dębnik area (Baliński 1995).
Klapperilepis schuelkei sp. n.(Figs 84A–H and 133)
Holotype: Spec i men ZPAL cXVI/1240 (Fig. 84A).Type ho ri zon and lo cal ity: Sam ple Pł−42, early Famennian ear li est K. triangularis Zone at Płucki, Holy Cross Moun tains.Der i va tion of name: To hon our the con tri bu tion of Immo Schülke to un der stand ing this species.
Ma te rial. — 25 spec i mens.Di ag no sis. — Smooth plat form of P1 el e ments with pointed lobe, M el e ment with fan−like ar range ment of
denticles, P2 el e ments with weakly de vel oped plat form and reg u lar denticulation.Re marks. — The ap pa ra tus of this spe cies was re stored by Schülke (1999) as Palmatolepis arcuata, but
as I al ready pointed out else where (Dzik 2002, p. 624), the type ho ri zon of the holotype of Palmatolepisarcuata of Schülke (1995) is of sig nif i cantly youn ger age and can not serve as the name−bearer for it. In stead,I sug gested Palmatolepis marginata clarki Ziegler, 1962 as pos si bly conspecific with this early mem ber ofthe Klapperilepis lin eage, al though the lat ter spe cies has been synonimized with Tripodellus variabilis bySchülke (1999). With out ad di tional topotype ma te rial large enough to en able ap pa ra tus re con struc tion it ishard to de cide on such sub jects and the only ba sis for this de ci sion was that the type spec i men of K. clarkishows a more rounded ap pear ance of the lobe and its more trans verse di rec tion that is typ i cal for T.variabilis. Ad mit tedly, the three spec i mens from the type ho ri zon il lus trated by Ziegler (1962, pl. 2: 20–25)show a ro bust plat form mar gin typ i cal of early Tripodellus. More over, the co−oc cur rence of the type ma te rialwith the rather ad vanced mem ber of the Klapperilepis lin eage, K. quadrantinodosolobata, sup ports theSchülke’s (1999) idea that P. clarki is a mem ber of an other lin eage, thus be long ing rather to Tripodellus. Toclar ify these no men cla tor ial un cer tain ties, I pro pose a new name for this spe cies, based on the ma te rial fromthe ear li est Famennian and ac cept Schülke’s (1999) in ter pre ta tion of T. clarki.
Oc cur rence. — The ear li est K. triangularis Zone at Płucki.
118 JERZY DZIK

Klapperilepis spathula (Schülke, 1995)(Figs 84I–L and 133)
Type ho ri zon and lo cal ity: Bed 14 at the Up per Coumiac quarry near Cessenon, Montagne Noire (Schülke, 1995).
Ma te rial. — 49 spec i mens.Di ag no sis. — P1 el e ment with long and nar row ex ten sion of the pos te rior lobe, its sur face or na mented
with ir reg u lar tuberculation, P2 el e ments flat, with out plat form, reg u larly denticulated and with pos te ri orlybent dorsal process.
FAMENNIAN CONO DONTS AND AMMONOIDS 119
Fig. 84. Early Famennian lobate palmatolepidids from the Holy Cross Moun tains. A–H. Klapperilepis schuelkei sp. n. from theK. triangularis Zone strata at Płucki (sam ple Pł−42; for lo ca tion and de tails see Dzik 2002). P1 (A–C), P2 (D), S0 (E), S1 (F),S2 (P), S3–4 (G), and M (H) el e ments; spec i mens ZPAL cXVI/1240 (holotype, A), 1242, 1241, and 1243–1247, re spec tively.I–L. Klap perilepis spathula (Schülke, 1995) from the mid K. triangularis Zone at Jabłonna (sam ples J−53, I, J, L and J−53a, K).P1 (I), P2 (J), S3–4 (K), and M (L) el e ments; spec i mens ZPAL cXVI/1262, 1263, 1227 and 1264, re spec tively. M–BB, CC–EE?.Klapperilepis quadrantinodosolobata (Sannemann, 1955) from the late K. triangularis Zone at Jabłonna (M, N, sam ple J−42;O–S, sam ple J−45a) and Wietrznia (T–DD, sam ple Wtr−27). P1 (M, S–W, pos si bly CC–EE), P2 (N and X), S0 (Y), S1 (P and Z),S2 (O), S3–4 (BB), and M (Q, R, AA) el e ments; spec i mens ZPAL cXVI/1412, 1413, 1330, 1329, 1332, 1331, 1327, 1328, 1433,
1441, 1431, 1434, 1435, 1436, 1439, 1438, 1429, 1432, and 1430, re spec tively.

Re marks. — Ap pa ra tus res to ra tion of Schülke (1995) is con firmed by data from the Holy Cross Moun −tains. Apart from the plat form shape, the most char ac ter is tic as pect of this and suc ces sive spe cies of the lin −eage is the step ping ar range ment of al ter nat ing denticles of S0 to S2 el e ments giv ing them a highly or deredap pear ance un known in any other Famennian cono donts. Also P2 el e ments, lack ing plat form and stronglybent, are of much diagnostic value in this lineage.
Oc cur rence. — The late K. triangularis Zone at Płucki, Wietrznia, Karczówka, Jabłonna, and Kowala.
Klapperilepis quadrantinodosolobata (Sannemann, 1955)(Figs 84M–BB, CC–EE?, and 133)
Type ho ri zon and lo cal ity: Black lime stone with Nehdenites verneuili at Breitengrund, Frankenwald (Sannemann 1955a;Ziegler 1975).
Ma te rial. — 4,256 spec i mens.Di ag no sis. — P1 el e ment with prom i nently tuberculated an te rior lobe and a tongue−like ex ten sion of the
smooth pos te rior lobe, flat P2 el e ments with strongly bent dor sal process.Re marks. — Ap pa ra tus com po si tion was re stored by Metzger (1994) and con firmed by Schülke (1995).
This is the end−mem ber of its lin eage shar ing with its older rel a tives the highly reg u lar ap pear ance oframiform elements.
Oc cur rence. — The K. crepida Zone at Jabłonna, Kadzielnia, Wietrznia, Łagów, Miedzianka, and Kowala.
Klapperilepis delicatula (Branson et Mehl, 1934)(Figs 85 and 133)
Type ho ri zon and lo cal ity: Base of the Saverton Shale at Sees Creek in Mon roe County, Mis souri (Ziegler 1975).
Ma te rial. — 208 spec i mens.Di ag no sis. — P1 el e ment with tri an gu lar out line of the plat form ex tend ing into an ex tremely wide pos te −
rior lobe.Re marks. — This is the first, and prob a bly only up to the pres ent Famennian cono dont with the ap pa ra tus
con firmed by a nat u ral as sem blage (Puchkov et al. 1981). Its pres er va tion is not good enough to of fer de tails onmor phol ogy of in di vid ual el e ments but the rel a tively wide an gle be tween pro cesses of the M el e ment re sem blesearly spe cies of Klapperilepis. In sam ple Pł−1 rel a tively nu mer ous P1 el e ments are as so ci ated with those ofPalmatolepis sandbergi and Tripodellus clarki and M el e ments of K. quadrantinodosolobata type. This leaveslit tle choice and most prob a bly K. delicatula be longs to an ad vanced lin eage of Klapperilepis. Per haps the Rus −sian ap pa ra tus rep re sents an early stage in its evo lu tion with char ac ters of the an ces tral K. ultima still preserved.
Oc cur rence. — The K. triangularis Zone at Płucki, Wietrznia, Miedzianka, Jabłonna, and Kowala.
120 JERZY DZIK
Fig. 85. Palmatolepidid Klapperilepis delicatula (Branson et Mehl, 1934) from the K. triangularis Zone at Płucki (A–G, sam plePł−1) in the Holy Cross Moun tains. P1 (A–D), P2 (E–G), S0 (H), S1 (I), S2 (O), S3–4 (J), and M (K, L) el e ments; spec i mens ZPAL
cXVI/1359, 1361, 1342, 1360, 1367, 1368, 1362–1366, and 1369, re spec tively.
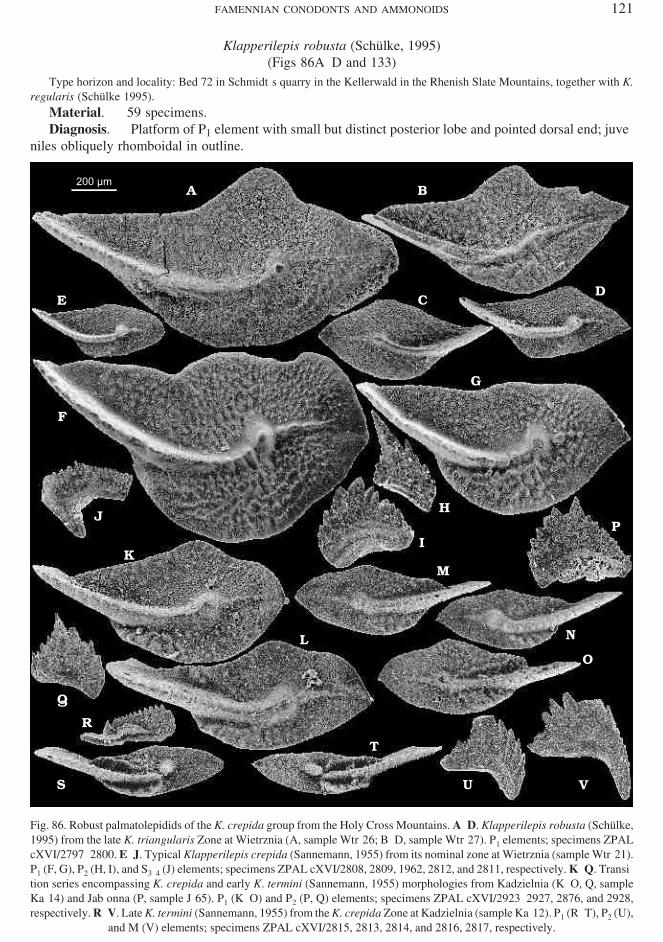
Klapperilepis robusta (Schülke, 1995)(Figs 86A–D and 133)
Type ho ri zon and lo cal ity: Bed 72 in Schmidt’s quarry in the Kellerwald in the Rhenish Slate Moun tains, to gether with K.regularis (Schülke 1995).
Ma te rial. — 59 spec i mens.Di ag no sis. — Plat form of P1 el e ment with small but dis tinct pos te rior lobe and pointed dor sal end; ju ve −
niles obliquely rhomboidal in outline.
FAMENNIAN CONO DONTS AND AMMONOIDS 121
Fig. 86. Ro bust palmatolepidids of the K. crepida group from the Holy Cross Moun tains. A–D. Klapperilepis robusta (Schülke,1995) from the late K. triangularis Zone at Wietrznia (A, sam ple Wtr−26; B–D, sam ple Wtr−27). P1 el e ments; spec i mens ZPALcXVI/2797–2800. E–J. Typ i cal Klapperilepis crepida (Sannemann, 1955) from its nom i nal zone at Wietrznia (sam ple Wtr−21).P1 (F, G), P2 (H, I), and S3–4 (J) el e ments; spec i mens ZPAL cXVI/2808, 2809, 1962, 2812, and 2811, re spec tively. K–Q. Tran si −tion se ries en com pass ing K. crepida and early K. ter mini (Sannemann, 1955) morphologies from Kadzielnia (K–O, Q, sam pleKa−14) and Jabłonna (P, sam ple J−65). P1 (K–O) and P2 (P, Q) el e ments; spec i mens ZPAL cXVI/2923–2927, 2876, and 2928, respectively. R–V. Late K. ter mini (Sannemann, 1955) from the K. crepida Zone at Kadzielnia (sam ple Ka−12). P1 (R–T), P2 (U),
and M (V) el e ments; spec i mens ZPAL cXVI/2815, 2813, 2814, and 2816, 2817, re spec tively.

Re marks. — P2 el e ments were iden ti fied by van den Boogaard and Kuhry (1979, fig. 18), the com pleteap pa ra tus by Schülke (1999). The prim i tive ap pear ance of the M el e ments in di cates a re la tion ship toKlapperilepis triangularis. Plat form of P1 el e ment ex tend ing al most to the end of the free blade is shared bythis spe cies with its suc ces sor, K. crepida.
Oc cur rence. — The lat est K. triangularis Zone at Płucki and Wietrznia.
Klapperilepis crepida (Sannemann, 1955)(Figs 86E–L and 133)
Type ho ri zon and lo cal ity: Black lime stone with Nehdenites verneuili at Breitengrund, Frankenwald (Sannemann 1955a;Ziegler 1975).
Ma te rial. — 840 spec i mens.Di ag no sis. — Plat form of P1 el e ment al most lack ing the pos te rior lobe and with blunt dor sal apex, or na −
mented with ir reg u larly dis trib uted tubercles.Re marks. — P2 el e ments of the ap pa ra tus were iden ti fied by Schülke (1999). This is a highly char ac ter is −
tic spe cies ow ing to the ro bust and some what ir reg u lar ap pear ance of the plat form. The tran si tion from K.robusta to K. crepida seems to be grad ual and iden ti fi ca tion of some sam ples may be dif fi cult be cause of thegreat variability of both species.
Oc cur rence. — Zone of its own at Jabłonna, Kadzielnia, Wietrznia, Kowala, and Miedzianka.
Klapperilepis termini (Sannemann, 1955)(Figs 86M–V and 133)
Type ho ri zon and lo cal ity: Black lime stone with Nehdenites verneuili at Breitengrund, Frankenwald (Sannemann 1955a;Ziegler 1975).
Ma te rial. — 931 spec i mens.Di ag no sis. — Plat form of P1 el e ment with rows of high tu ber cles on both sides of the ven tral blade and
prom i nent, iso lated cusp.Re marks. — The P2 el e ment of the ap pa ra tus was iden ti fied by Schülke (1999) and such el e ments also
oc cur in the Holy Cross Moun tains.As al ready shown by Schülke (1995), early pop u la tions of the spe cies (in my ma te rial sam ples J−65, Ka−14,
Wtr−25, and Md−27) are mor pho log i cally tran si tional to K. crepida and K. robusta, with some el e ments hav ingonly one row of rel a tively low denticles pos te rior to the blade, oth ers lack ing them com pletely and dif fer ingfrom associsted K. crepida only in a nar rower plat form. There is an ap par ent change within the lin eage to wardsa more and more nar rower plat form with a stron ger sep a ra tion of its smooth and flat dor sal area and ex tremelyhigh ridges on both sides of the blade. The ridges do not reach the cusp, which forms an iso lated high con i calstruc ture in the mid dle of the el e ment. This change seems to be of much correlative value.
Oc cur rence. — The K. crepida Zone at Jabłonna, Kadzielnia, Wietrznia, Kowala, and Miedzianka.
Genus Tripodellus Sannemann, 1955Type spe cies: Tripodellus flexuosus Sannemann 1955a (P2 el e ment of T. minutus) from the Famennian of Frankenwald.
Di ag no sis. — Ap pa ra tus with triramous or at least an gu larly bent P2 el e ments, lack ing plat form in un de −rived species.
122 JERZY DZIK
Fig. 87. Ear li est spe cies of the palmatolepidid Tripodellus, T. clarki (Ziegler, 1962) with biramous P2 el e ments from K. trian −gularis Zone strata at Płucki (sam ple Pł−42) in the Holy Cross Moun tains. P1 (A–C) and M (D) el e ments; spec i mens ZPAL
cXVI/1250, 1249, 1248, and 1251, re spec tively.

Tripodellus clarki (Ziegler, 1962)(Figs 87, 88, and 134)
Type ho ri zon and lo cal ity: Bed 23 in trench II at Seßacker near Oberscheld, Rhenish Slate Moun tains (Ziegler 1975).
Ma te rial. — 623 spec i mens.Di ag no sis. — Gently bent P2 el e ment with out plat form, rhomboidal out line of rel a tively wide plat form in
P1 el e ment, short pro cesses of M element.Re marks. — The ap pa ra tus was re stored by Schülke (1995) as Palmatolepis variabilis and P. delicatula
postdelicatula. I failed to dis tin guish be tween these morphologies in my highly vari able ma te rial from theear li est Famennian. Pop u la tions from the late K. triangularis Zone are much more ad vanced to wards themorphologies typ i cal of Tripodellus and they de serve tax o nomic sep a ra tion as T. variabilis.
Oc cur rence. — The K. triangularis Zone at Płucki, Wietrznia, and Jabłonna.
FAMENNIAN CONO DONTS AND AMMONOIDS 123
Fig. 88. Typ i cal Tripodellus clarki (Ziegler, 1962) from K. triangularis Zone strata at Jabłonna (A, C, D, F–H, sam ple J−53; B, C, E, G, J–N, sam ple J−53a; V–CC, sam ple J−58) and Płucki (O–U, sam ple Pł−1) in the Holy Cross Moun tains. P1 (A–C, O, P, V, W), P2 (D–I, T, X–Z), S0 (J, Q, and CC), S1 (K, R, DD), S2 (L, BB), S3–4 (M, S), and M (N, U, and AA) el e ments; spec i mens ZPAL
cXVI/1250, 1249, 1248, and 1251, re spec tively.

Tripodellus variabilis (Sannemann, 1955)(Figs 89A–N and 134)
Type ho ri zon and lo cal ity: Black lime stone with Nehdenites verneuili at Breitengrund, Frankenwald (Sannemann 1955a;Ziegler 1975).
Ma te rial. — 22,931 spec i mens.Di ag no sis. — An gu larly bent P2 el e ment, elon gated plat form in P1 el e ment with short, rounded pos te rior
lobe, elon gate ex ter nal pro cess of M element.Re marks. — It is con ve nient to sep a rate the early part of the Tripodellus lin eage into at least three
chronospecies (T. clarki, T. variabilis, and T. minutus). T. variabilis has been cho sen by Schülke (1999) asthe name for the first spe cies of the lin eage, but there is hardly any ev i dence that the ma te rial of Sannemenn(1955a) truly rep re sents this spe cies as long as ad vanced K. ter mini and K. crepida co−oc cur in the same bed.The P2 el e ments of the palmatolepidids are highly vari able and at this stage of the evo lu tion they are hardly of much diagnostic value.
P. variabilis is a se nior syn onym of Palmatolepis minuta wolskae of Szulczewski (1971), on the ev i denceof ma te rial from a bed at Kadzielnia with early P. ter mini.
Oc cur rence. — The late K. triangularis and K. crepida zones at Jabłonna, Kadzielnia, Wietrznia,Kowala, and Miedzianka.
Tripodellus subtilis (Khalymbadzha et Chernysheva, 1978)(Figs 90 and 134)
Type ho ri zon and lo cal ity: Famennian of the Urals.
124 JERZY DZIK
Fig. 89. Spe cies of the palmatolepidid Tripodellus with biramous P2 el e ments from the K. crepida Zone at Jabłonna (A–O, sam −ple J−45a) and Wietrznia (P, sam ple Wtr−21) in the Holy Cross Moun tains. A–N. T. variabilis (Sannemann, 1955). P1 (A–D, I),P2 (E, F), S0 (G), S1 (H), S2 (J), S3–4 (K), and M (L–N) el e ments; spec i mens ZPAL cXVI/1315–1322, 2757, 1338, 1323, 1325,
1326, and 1324, re spec tively. O–P. T. subgracilis (Bis choff, 1956). P1 el e ments; spec i mens ZPAL cXVI/2834 and 1967.

Ma te rial. — 24 spec i mens.Di ag no sis. — P1 el e ment with nar row, con vex plat form of asym met ric lan ceo late out line, ex tend ing ven −
trally to half length of the blade.Re marks. — Tax o nomic iden ti fi ca tion of the spe cies is based on Metzger (1994). Rare el e ments as so ci ated
with P1 el e ments of the spe cies in clude biramous P2 el e ments, which sug gests its or i gin from T. variabilis. Iftrue, this would then be a rel ict lin eage evolv ing to wards re duc tion of the plat form in P1 elements.
Oc cur rence. — The K. crepida Zone at Wietrznia and Jabłonna.
Tripodellus lobus (Helms, 1963)(Figs 91 and 134)
Type ho ri zon and lo cal ity: A block of Cheiloceras Stufe lime stone from a Car bon if er ous brec cia at Langenaubach, RhenishSlate Moun tains (Ziegler 1975).
Ma te rial. — 653 spec i mens.Di ag no sis. — Plat form in P1 el e ment with prom i nent, nar row pos te rior lobe, in ju ve nile spec i mens ori −
ented some what ven trally; an gu larly bent biramous P2 element.Re marks. — Ap pa ra tus of the spe cies was re con structed by Metzger (1994). Spec i mens of ju ve nile mor phol ogy typ i cal of this spe cies co−oc cur with late pop u la tions of T. minutus
and there seems to be no tran si tion be tween them. This is ap par ently a suc ces sor of T. variabilis that be camesep a rated from the main lin eage rep re sented by T. minutus as a re sult of an allopatric speciation pro cess andsub se quent merg ing of their populations.
Oc cur rence. — The K. crepida Zone at Kadzielnia, Jabłonna, Wietrznia, and Miedzianka.
Tripodellus subgracilis (Bischoff, 1956)(Figs 89O, P and 134)
Type ho ri zon and lo cal ity: K. crepida Zone at Bicken quarry (Bis choff 1956).
Ma te rial. — Three spec i mens.Di ag no sis. — P1 el e ment with very nar row plat form bear ing sharply tri an gu lar small pos te rior lobe.Re marks. — Ap pa ra tus was re stored by Metzger (1994) for a more ad vanced pop u la tion with well
denticulated pos te rior pro cess. Biramous P2 el e ment in di cates a close re la tion ship to T. lobus and the Pol ishspec i mens, lack ing denticulation of the pos te rior pro cess, are tran si tional to those il lus trated by Metzger(1994). Palmatolepis parva Klapper et al., 2004 may be ancestral to this species.
Oc cur rence. — The K. crepida Zone at Jabłonna.
Tripodellus minutus (Branson et Mehl, 1934)(Figs 9D, 92A–J, and 134)
Type ho ri zon and lo cal ity: Famennian at Dixie, Mis souri (Ziegler 1977, p. 313).
Ma te rial. — 2,504 spec i mens.
FAMENNIAN CONO DONTS AND AMMONOIDS 125
Fig. 90. Palmatolepidid Tripodellus subtilis (Khalymbadzha et Chernysheva, 1978) with biramous P2 el e ments from the late K.trian gularis Zone at Wietrznia (A, D–H, sam ple Wtr−27) and the K. crepida Zone at Jabłonna (B, C, sam ple J−45a) in the HolyCross Moun tains. P1 (A–C), P2 (D), S0 (E), S2 (F), S3–4 (G), and M (H) el e ments; spec i mens ZPAL cXVI/1422, 1314, 1313, and
1424–1428, re spec tively.

Di ag no sis. — Triramous P2 el e ment, rhomboidal out line of plat form of P1 al most with out any lobe;straight or slightly asym met ric ju ve nile specimens.
Re marks. — The P2 el e ment was iden ti fied by van den Boogaard and Kuhry (1979). Ap pa ra tus was re −stored by Metzger (1994). P2 el e ments with ad di tional pro cess oc cur rarely but con sis tently in sam ples withT. variabilis, be ing prob a bly end−mem bers of pop u la tion vari abil ity ex pressed mostly in bend ing of the el e −ment. They start to dom i nate the sam ple in the Kadzielnia sec tion between samples Ka−3 and Ka−5.
Oc cur rence. — The K. crepida Zone to the ear li est C. quadrantinodosa Zone at Kadzielnia, Łagów,Jabłonna, Wietrznia, Miedzianka, and Kowala.
Tripodellus donoghuei sp. n.(Figs 92K–Q and 134)
Holotype: Spec i men ZPAL cXVI/2763 (Fig. 92N).
126 JERZY DZIK
Fig. 91. Late palmatolepidid Tripodellus lobus (Helms, 1963) with biramous P2 el e ments from the late K. triangularis Zone atWietrznia (I, J, sam ple Wtr−27), the K. crepida Zone at Jabłonna (A–H,) sam ple J−44, and the late K. crepida (with K.rhomboidea) Zone at Łagów (K–R, sam ple Mak−3−4) in the Holy Cross Moun tains. P1 (A, B, I, J, K, L), P2 (C, M), S0 (D, R), S1
(E, O), S2 (F, P), S3–4 (G, N), and M (H, Q) el e ments; spec i mens ZPAL cXVI/2742, 2741, 2743–2748, 1423, 1421, 2750, 2749,2751, 2755, 2753, 2754, 2756, and 2752, re spec tively.

Type ho ri zon and lo cal ity: Sam ple Ost−12, late Famennian L. styriacus Zone at Ostrówka, Holy Cross Moun tains. Der i va tion of name: To hon our Philip Donoghue for his con tri bu tion to un der stand ing the ar chi tec ture of cono dont
apparatuses.
Ma te rial. — 391 spec i mens.Di ag no sis. — Twisted P1 el e ment with wide, lan ceo late plat form and min ute lar val basal cone (pit). Re marks. — The ap pa ra tus of this spe cies was al ready re stored by Schülke (1999) as Palmatolepis aff. Pal.
gracilis. Spec i mens of sim i lar mor phol ogy oc cur in sam ples with T. minutus but there is no gap in vari abil itysep a rat ing them from the modal mor phol ogy of that spe cies. Such a gap can be iden ti fied only in sam ples with T. schleizius and T. lobus as as so ci ated spe cies, al though rarely ju ve nile spec i mens of T. schleizius of com pa ra blesize are as so ci ated with T. donoghuei in its early pop u la tions. The old est sam ple with ju ve niles of T. schleiziusshow ing a vir tu ally straight blade have been found in Kadzielnia in sam ple Ka−7, where T. donoghuei is missing.
In the out line of the plat form T. donoghuei re sem bles mem bers of the T. gonioclymeniae lin eage but itlacks the stage of “sinuosa” larva with strongly asym met ric basal cone in P1 el e ments. It seems to be a suc −ces sor of T. minutus, which orig i nated allopatrically in re spect to T. schleizius.
Oc cur rence. — The late K. crepida (with C. klapperi) to L. styriacus zones at Kadzielnia, Jabłonna,Kowala, and Ostrówka.
FAMENNIAN CONO DONTS AND AMMONOIDS 127
Fig. 92. Spe cies of the palmatolepidid Tripodellus with triramous P2 el e ments from the Holy Cross Moun tains. A–J. T. minutus(Branson et Mehl, 1934) from the C. quadrantinodosa Zone at Łagów (sam ple Ł−28). P1 (A–C, E), P2 (D), S0 (F), S1 (H), S2 (I),S3–4 (J), and M (G) el e ments; spec i mens ZPAL cXVI/1402–1405, 2940, 1406, 1410, 1408, 1407, and 1409, re spec tively.K–Q. T. do noghuei sp. n. from the early L. styriacus Zone at Ostrówka (K, M, N, sam ple Ost−12) and the P. trachytera Zone atJabłonna (L, O–Q, bed 24). P1 (K–N), P2 (O), S3–4 (P), and M (Q) el e ments; spec i mens ZPAL cXVI/2764, 2758, 2762, 2763
(holotype, N), and 2759–2761, re spec tively.

Tripodellus schleizius (Helms, 1963)(Figs 9C, 93, and 134)
Type ho ri zon and lo cal ity: 6.5 m be low the top of the Famennian at Großen Buschteich near Rödersdorf, Thuringia, up perpart of the Platyclymenia Stufe (Ziegler 1975).
Ma te rial. — 5,112 spec i mens.Di ag no sis. — Clearly sep a rate asym met ric basal cone (pit) and sin u ous blade in ju ve nile P1 el e ment, plat −
form some times dis ap pear ing in front of the tip in adults with an an gu lar pos te rior lobe near the cusp,triramous P2 element.
Re marks. — The tran si tion from T. minutus to T. schleizius can be traced through the Łagów sec tion near the base of the C. quadrantinodosa Zone. In sam ples from the Lower Łagów Beds min ute spec i mens ofTripodellus are lat er ally arched and their basal cone re mains very small. At the base of the Up per ŁagówBeds sigmoidally bent ju ve nile el e ments ap pear with a wide asym met ric basal cone. From some rea son inthese tran si tional sam ples there is a dis tinct bimodality in size fre quency dis tri bu tion and no in ter me di atespec i mens have been found. It looks as if sin u ous morphologies had a higher rate of ju ve nile mor tal ity, per −haps re lated to meta mor pho sis at the end of the sinuosa larva stage. The blade is usu ally more sin u ous inadults than it was in older sam ples but they do not show clear sep a ra tion of the lar val basal cav ity. Ei ther it
128 JERZY DZIK
Fig. 93. Palmatolepidid Tripodellus schleizius (Helms, 1963) with triramous P2 el e ments from the P. trachytera Zone at Kowala(B, F, G, J–L, N, O, sam ple Ko−8a; M, sam ple Ko−9a) and the early Lagovignathus styriacus Zone at Ostrówka (A, C–H, I, P–S,sam ple Ost−12) in the Holy Cross Moun tains. P1 (A–F, K, M), P2 (G, H), S0 (I, J), S1 (N, R), S2 (Q), S3–4 (L, P), and M (O, S) elements; spec i mens ZPAL cXVI/2768, 391, 2769, 2766, 2765, 392, 2770, 2771, 393, 2937, 395, 2944, 394, 396, 2774, 2773,
2772, and 2775, re spec tively.

was resorbed, spec i mens with asym me try rarely sur vived to adult stages, or spe cies of two sep a rate lin eagesco−oc curred. The lat ter pos si bil ity seems un likely, as in sam ples from some what higher beds (e.g., Ł−40) in −ter me di ate spec i mens oc cur show ing con ti nu ity be tween ex treme stages of on tog eny. This is the case alsowith late Famennian sam ples with T. gracilis. They show, how ever, in creased ju ve nile mor tal ity and are inthis re spect dif fer ent from co−oc cur ring cono dont spe cies. Per haps this was an eco log i cally op por tu nis ticspe cies. In the course of evo lu tion of the lineage within the mid Famennian the basal cavity became widerand wider, as documented in the Kowala section.
The bound ary be tween this chronospecies and its suc ces sor T. gracilis is dif fi cult to de fine as the di ag nos −tic char ac ters are very vari able in pop u la tions and seem to spread grad u ally in the course of evo lu tion. Thetype pop u la tion of T. schleizius is un com fort ably close to the end of its range.
Oc cur rence. — The C. quadrantinodosa to P. trachytera zones at Łagów, Jabłonna, Miedzianka,Kowala, and Ostrówka.
Tripodellus gracilis (Branson et Mehl, 1934)(Figs 94A–K and 134)
Type ho ri zon and lo cal ity: Famennian at Dixie, Mis souri (Ziegler 1977, p. 313).Ma te rial. — 11,928 spec i mens.Di ag no sis. — Wide and strongly asym met ric basal cone (pit) of P1 el e ment, its tip bent pos te ri orly,
rounded lobe near the cusp even in large spec i mens.Re marks. — Ap pa ra tus was re stored by van den Boogaard and Kuhry (1979), and con firmed by my self
(Dzik 1991). No spec i mens of the mor phol ogy pro posed to be typ i cal for Tripodellus expansus (Sandberg etZiegler, 1979) are known from the Holy Cross Moun tains but some ma ture el e ments with an ex tremely wideplat form ap proach that mor phol ogy (e.g., in sam ple Ost−5). They share with those from Montana their asym −met ric lar val basal cav ity, which places them in the T. gonioclymeniae lin eage, but they do not show sostrong a wid en ing of the plat form and in occlusal view do not dif fer form T. gracilis. In sam ple Ost−7 thedom i nant el e ments with asym met ric lar val basal cav ity are as so ci ated with rare spec i mens hav ing a widerplat form, which do not show a sep a ra tion of the basal cav ity at all, thus be ing sim i lar in those re spects toolder pop u la tions of T. gracilis. In fact, such asym me try and in cip i ent sep a ra tion of the basal cone is ob serv −able in the pop u la tion of T. gracilis from sam ple Ost−12, but the basal cone is small and well vis i ble only inju ve nile and strongly asym met ric spec i mens. It seems thus that this stage in the evo lu tion of the Tripodellusbranch is marked by ex pan sion of ju ve nile char ac ters to later ontogenetic stages. Palmatolepis deflectenssigmoidalis of Ziegler (1962) seems to be based on a ju ve nile T. gracilis.
Pos si bly, “Palmatolepis expansa” is a morphotype within the range of pop u la tion vari abil ity of T.gracilis. Perri and Spaletta (1991) clas si fied within “P. expansa” spec i mens that do not show asym me try ofthe basal cav ity. This in di cates how mis lead ing may be the con cept of that spe cies and that strati graphic de ci −sions based on it are of questionable value.
Oc cur rence. — The L. styriacus to D. trigonica zones at Jabłonna, Ostrówka, Miedzianka, Kowala,Gołogłowy, and Dzikowiec.
Tripodellus mancus (Helms, 1963)(Figs 94S and 134)
Type ho ri zon and lo cal ity: 3 m be low the top of the red lime stone of the Platyclymenia Stufe at Alte Heerstrasse nearOettersdorf, Thuringia (Helms, 1963).
Ma te rial. — 12 spec i mens.Di ag no sis. — Strongly bent P1 el e ment with ob tuse tip of the plat form and strongly asym met ric ju ve nile
basal cav ity; ap pa ra tus unknown.Re marks. — This is a poorly known spe cies, rep re sented by a few spec i mens of min ute size in all pub −
lished lo cal i ties. Per haps they rep re sent mar ginal pop u la tions of the T. gonioclymeniae lin eage at the stage ofits allopatric sep a ra tion from the main lin eage of Tripodellus. Small size of el e ments seems con sis tent withap par ently paedomorphic evo lu tion of the lin eage, marked by ex pan sion of the sinuosa larva stage to the endof on tog eny. The plat form of some spec i mens from Kowala show a pointed dor sal tip, which makes themsim i lar to T. gonioclymeniae.
Oc cur rence. — The early P. jugosus Zone at Ostrówka and Kowala.
FAMENNIAN CONO DONTS AND AMMONOIDS 129

Tripodellus gonioclymeniae (Müller, 1956)(Figs 94L–R and 134)
Type ho ri zon and lo cal ity: Gray ish brown lime stone from the top of Effenberg near Hachen, Rhenish Slate Moun tains(Ziegler 1975).
Ma te rial. — 243 spec i mens.Di ag no sis. — Strongly bent P1 el e ment with plat form nar row ing to the tip, with its great est width in the
mid dle length of the el e ment, the asym met ric basal cone con tin ues through out on tog eny, other el e ments ofthe ap pa ra tus prob a bly in dis tin guish able from T. gracilis.
Re marks. — No doubt that the lin eage T. mancus ® T. gonioclymeniae orig i nated from T. gracilis.Sandberg and Ziegler (1979) pro posed “Palmatolepis expansa” to be the con nect ing link. As noted above,this mor phol ogy is not rep re sented in the Holy Cross Moun tains and prob a bly the evo lu tion took place else −where, with T. manca and T. gonioclymeniae be ing suc ces sive im mi grants from that source area.
Oc cur rence. — The late P. jugosus to D. trigonica zones at Ostrówka, Kowala, aned Dzikowiec.
130 JERZY DZIK
Fig. 94. Late spe cies of the palmatolepidid Tripodellus with triramous P2 el e ments. A–K. T. gracilis (Branson et Mehl, 1934)from the D. trigonica (A, sam ple Ko−166) and P. jugosus (C, sam ple Ko−3) zones at Kowala and the P. jugosus Zone at Ostrówka (B, D–K, sam ple Ost−2). P1 (A–E), P2 (F), S0 (G), S1 (H), S2 (I), S3–4 (J), and M (K) el e ments; spec i mens ZPAL cXVI/2931, 2780,2934, 2781, 2779, 2782–2787, re spec tively. L–R. T. gonioclymeniae (Müller, 1956) from the P. jugosus Zone at Ostrówka(L, M, Q, sam ple Ost−2) and Kowala (P, sam ple Ko−62) in the Holy Cross Moun tains and the P. jugosus (N, R, sam ple Dz−8) andthe D. trigonica (O, sam ple Dz−7) zones at Dzikowiec in the Sudetes. P1 el e ments; spec i mens ZPAL cXVI/2790, 2777, 2792,2789, 2788, 2778 an 2791, re spec tively. S. T. mancus (Helms, 1963) from the L. styriacus Zone at Ostrówka (sam ple Ost−5).
P1 el e ment; spec i mens ZPAL cXVI/2776.
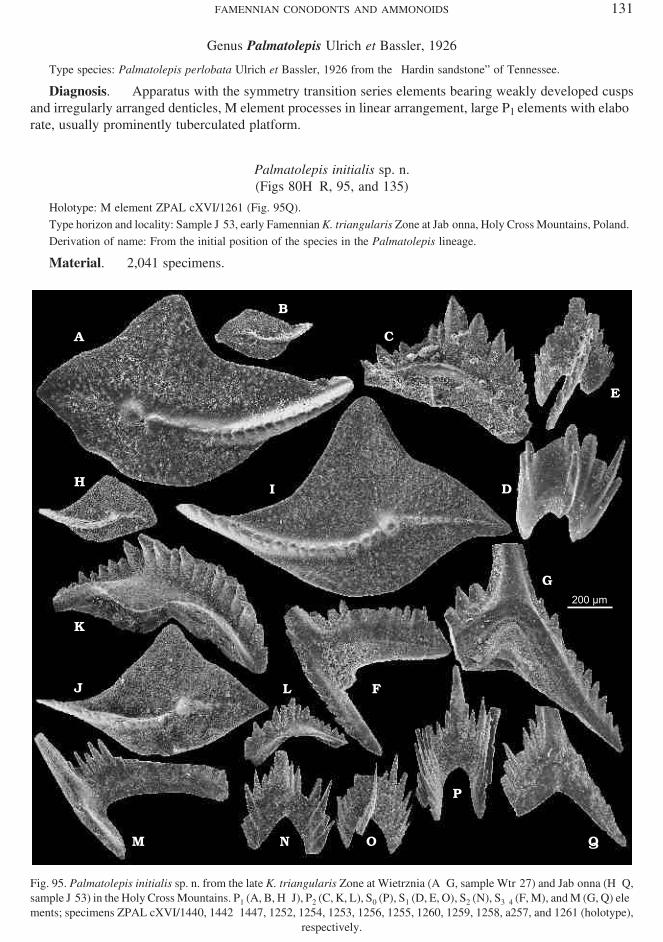
Genus Palmatolepis Ulrich et Bassler, 1926
Type spe cies: Palmatolepis perlobata Ulrich et Bassler, 1926 from the “Hardin sand stone” of Ten nes see.
Di ag no sis. — Ap pa ra tus with the sym me try tran si tion se ries el e ments bear ing weakly de vel oped cuspsand ir reg u larly ar ranged denticles, M el e ment pro cesses in lin ear ar range ment, large P1 el e ments with elab o −rate, usu ally prom i nently tuberculated platform.
Palmatolepis initialis sp. n.(Figs 80H–R, 95, and 135)
Holotype: M el e ment ZPAL cXVI/1261 (Fig. 95Q).
Type ho ri zon and lo cal ity: Sam ple J−53, early Famennian K. triangularis Zone at Jabłonna, Holy Cross Moun tains, Po land.
Der i va tion of name: From the ini tial po si tion of the spe cies in the Palmatolepis lin eage.
Ma te rial. — 2,041 spec i mens.
FAMENNIAN CONO DONTS AND AMMONOIDS 131
Fig. 95. Palmatolepis initialis sp. n. from the late K. triangularis Zone at Wietrznia (A–G, sam ple Wtr−27) and Jabłonna (H–Q,sam ple J−53) in the Holy Cross Moun tains. P1 (A, B, H–J), P2 (C, K, L), S0 (P), S1 (D, E, O), S2 (N), S3–4 (F, M), and M (G, Q) el e −ments; spec i mens ZPAL cXVI/1440, 1442–1447, 1252, 1254, 1253, 1256, 1255, 1260, 1259, 1258, a257, and 1261 (holotype),
re spec tively.

Di ag no sis. — M el e ment with al most straight cusp and short but well denticulated ven tral pro cess form −ing a wide an gle with the prom i nently denticulated dor sal process.
Re marks. — Schülke (1999) pro posed to use the name Palmatolepis abnormis (Branson et Mehl, 1934)for this spe cies. How ever, as pointed out by Klapper et al. (2004, p. 382) the P2 el e ment from the SavertonShale cho sen to be the holotype of this spe cies is undiagnostic and in dis tin guish able from ones that oc cur indif fer ent ap pa ra tus re con struc tions (ac cord ing to Branson and Mehl 1934 there is a “fairy com plete gra da −tion” to other P2 spec i mens in the sam ple) and the spe cies is in valid (a nomen dubium). It is rather the M el e −ment that bears char ac ters di ag nos tic for the spe cies. In its shape, with slightly re clined cusp, it is tran si tionalbe tween the an ces tral mor phol ogy of Klapperilepis and that of Palmatolepis and Conditolepis. The pop u la −
132 JERZY DZIK
Fig. 96. Palmatolepis sandbergi Ji et Ziegler, 1993 from the late K. triangularis (A–K, sam ple J−53a) and early K. crepida(L, sam ple J−43; M, O, P, bed 3; N, bed 5) zones at Jabłonna in the Holy Cross Moun tains. P1 (A–C, N–P), P2 (D, E, M), S0 (F),S1 (G), S2 (I), S3–4 (H), and M (J−L) el e ments; spec i mens ZPAL cXVI/1280−1284, 1286, 1288, 1285, 1289, 1290, 2884, 2883,
2882, and 2880, 2881, re spec tively.

tion from sam ple Pł−1 (Fig. 80H–R) is prob a bly the old est one in my ma te rial show ing fea tures of P. initialissp. n., but the gen er ally small size of el e ments makes it sim i lar rather to Klapperilepis triangularis. P1 el e −ments of these two species seem to be indistinguishable.
Oc cur rence. — The mid K. triangularis Zone at Płucki and Jabłonna.
Palmatolepis sandbergi Ji et Ziegler, 1993(Figs 96 and 135)
Type ho ri zon and lo cal ity: Lali sec tion, south ern China.
Ma te rial. — 803 spec i mens.Di ag no sis. — M el e ment with proclined cusp and short finely denticulated ven tral pro cess form ing a
wide an gle with the prom i nently denticulated dor sal process.Re marks. — The ap pa ra tus was re stored by Schülke (1999). Ex cept for the ap par ent trend to wards typ i −
cal Palmatolepis in the mor phol ogy of M el e ments, the ap pa ra tus is sim i lar to Klapperilepis triangularis. In bed 3 at Jabłonna P1 el e ments some what re sem bling those of Conditolepis lobicornis, but with the pos −
te rior lobe of the plat form dif fer ently ori ented, co−oc cur with P2 and M el e ments closely sim i lar to those of P. sandbergi (Fig. 96M, O, P). Whether the pro posed ap pa ra tus com po si tion is cor rect, and then this is pos si blya new spe cies of Palmatolepis, or the plat form el e ments be long to an other ge nus, has to be tested with amaterial from another locality.
Oc cur rence. — The late K. triangularis and early K. crepida zones at Płucki, Wietrznia, Kowala,Miedzianka, and Jabłonna.
Palmatolepis perlobata Ulrich et Bassler, 1926(Figs 97, 98, and 135)
Type ho ri zon and lo cal ity: “Hardin sand stone” near Mount Pleas ant, Ten nes see (Hud dle 1968).
Ma te rial. — 320 spec i mens.Di ag no sis. — Rel a tively low pop u la tion vari abil ity in P1 el e ments with sin u ous out line and small pos te −
rior lobe of al most smooth plat form, sharply denticulated P2 el e ments with prominent cusp.
FAMENNIAN CONO DONTS AND AMMONOIDS 133
Fig. 97. Palmatolepis perlobata Ulrich et Bassler, 1926 from the K. crepida Zone at Kadzielnia (sam ple Ka−7) in the Holy CrossMoun tains. P1 (A, B), P2 (C), S0 (D), S1 (E), S2 (F), S3–4 (G), and M (H) el e ments; spec i mens ZPAL cXVI/2885–2892, re spec tively.

Re marks. — Ap pa ra tus of the spe cies was re stored by Metzger (1994) and the Holy Cross Moun tainsma te rial sup ports his in ter pre ta tion. P2 el e ments in sam ples from Miedzianka fre quently show bi fur ca tion ofthe ex ter nal pro cess. Their plat form may be very wide and bro ken pieces su per fi cially re sem ble ancyro −gnathid P1 elements.
Oc cur rence. — The late K. crepida to C. marginifera zones at Jabłonna, Wietrznia, Łagów, Kowala, andKadzielnia.
Palmatolepis schindewolfi Müller, 1956(Figs 99 and 135)
Type ho ri zon and lo cal ity: Gray lime stone from a brec cia with ?Platyclymenia from a quarry at Langenaubach near Haiger,Rhenish Slate Moun tains (Ziegler 1975).
Ma te rial. — 6,378 spec i mens.Di ag no sis. — Vari able shape of P1 el e ments with ma ture el e ments show ing fre quently very nar row and
sharp lobes of the plat form, while ju ve niles may lack any pos te rior pro cesses hav ing in stead a large roundedanterior lobe.
Re marks. — P2 el e ments were iden ti fied by van den Boogaard and Kuhry (1979), com plete ap pa ra tus wasre stored by Metzger (1994) but with in cor rectly at trib uted M el e ment (ac tu ally be long ing to Conditolepis).
P. schindewolfi is a vari able spe cies. Even the very large sam ple Ost−12, with al most eight hun dred P1 el −e ments has not helped much in iden ti fi ca tion of pos si ble cryp tic spe cies within the ob served range of vari −abil ity. The an te rior lobe is semi cir cu lar in out line and gently con vex in the dom i nant morphotype (usu allyclas si fied as P. max ima, e.g., Metzger 1994), whereas the pos te rior lobe is well de vel oped and sharplypointed. The vari abil ity ex tends in three main di rec tions. One of the ex treme morphotypes de vel ops acrenulated mar ginal ridge of the an te rior lobe. It is tran si tional to P. trachytera but just a cou ple of such spec −i mens (P. perlobata sigmoidea Ziegler, 1962) have been found and the de mar ca tion of P. schindewolfi and P.
134 JERZY DZIK
Fig. 98. Palmatolepis perlobata Ulrich et Bassler, 1926 from the C. quadrantinodosa Zone at Łagów (A–D, F–I, sam ple Ł−28;E, sam ple Ł−38) in the Holy Cross Moun tains. P1 (A–C), P2 (D), S0 (E, F), S1 (H), S3–4 (G), and M (I) el e ments; spec i mens ZPAL
cXVI/2893, 2894, 1376, 1377, 1414, 1378, 1380, 1379, and 1381, re spec tively.
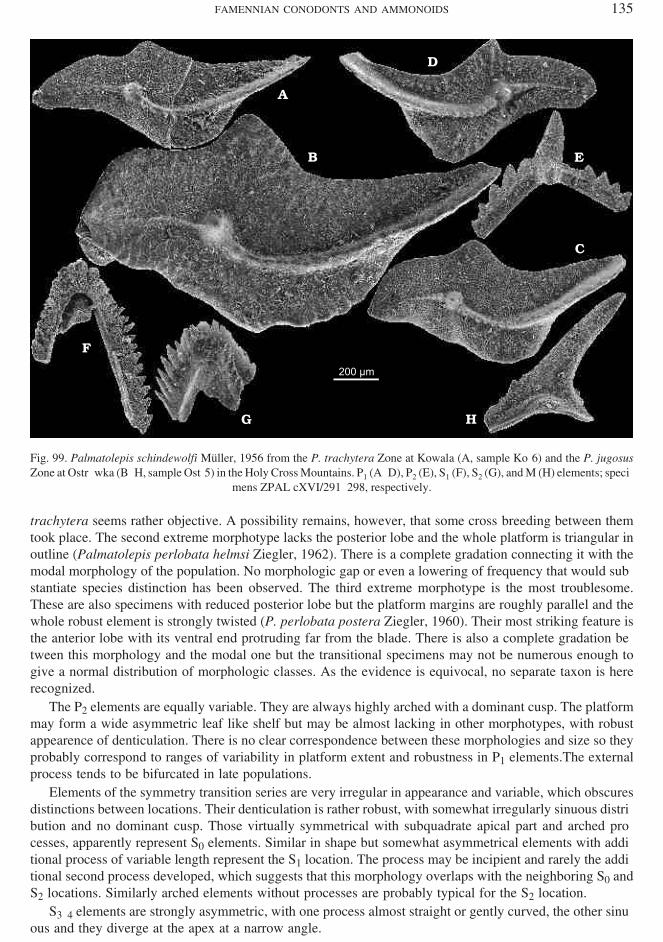
trachytera seems rather ob jec tive. A pos si bil ity re mains, how ever, that some cross−breed ing be tween themtook place. The sec ond ex treme morphotype lacks the pos te rior lobe and the whole plat form is tri an gu lar inout line (Palmatolepis perlobata helmsi Ziegler, 1962). There is a com plete gra da tion con nect ing it with themodal mor phol ogy of the pop u la tion. No morphologic gap or even a low er ing of fre quency that would sub −stan ti ate spe cies dis tinc tion has been ob served. The third ex treme morphotype is the most trou ble some.These are also spec i mens with re duced pos te rior lobe but the plat form mar gins are roughly par al lel and thewhole ro bust el e ment is strongly twisted (P. perlobata postera Ziegler, 1960). Their most strik ing fea ture isthe an te rior lobe with its ven tral end pro trud ing far from the blade. There is also a com plete gra da tion be −tween this mor phol ogy and the modal one but the tran si tional spec i mens may not be nu mer ous enough togive a normal distribution of morphologic classes. As the evidence is equivocal, no separate taxon is hererecognized.
The P2 el e ments are equally vari able. They are al ways highly arched with a dom i nant cusp. The plat formmay form a wide asym met ric leaf−like shelf but may be al most lack ing in other morphotypes, with ro bustappearence of denticulation. There is no clear cor re spon dence be tween these morphologies and size so theyprob a bly cor re spond to ranges of vari abil ity in plat form ex tent and ro bust ness in P1 el e ments.The ex ter nalpro cess tends to be bi fur cated in late populations.
El e ments of the sym me try tran si tion se ries are very ir reg u lar in ap pear ance and vari able, which ob scuresdis tinc tions be tween lo ca tions. Their denticulation is rather ro bust, with some what ir reg u larly sin u ous dis tri −bu tion and no dom i nant cusp. Those vir tu ally sym met ri cal with subquadrate api cal part and arched pro −cesses, ap par ently rep re sent S0 el e ments. Sim i lar in shape but some what asym met ri cal el e ments with ad di −tional pro cess of vari able length rep re sent the S1 lo ca tion. The pro cess may be in cip i ent and rarely the ad di −tional sec ond pro cess de vel oped, which sug gests that this mor phol ogy over laps with the neigh bor ing S0 andS2 lo ca tions. Sim i larly arched el e ments with out pro cesses are prob a bly typ i cal for the S2 location.
S3–4 el e ments are strongly asym met ric, with one pro cess al most straight or gently curved, the other sin u −ous and they di verge at the apex at a nar row angle.
FAMENNIAN CONO DONTS AND AMMONOIDS 135
Fig. 99. Palmatolepis schindewolfi Müller, 1956 from the P. trachytera Zone at Kowala (A, sam ple Ko−6) and the P. jugosusZone at Ostrówka (B–H, sam ple Ost−5) in the Holy Cross Moun tains. P1 (A–D), P2 (E), S1 (F), S2 (G), and M (H) el e ments; spec i −
mens ZPAL cXVI/291–298, re spec tively.

136 JERZY DZIK

M el e ments dif fer from those of Conditolepis in the pres ence of a dom i nant cusp, weak dor sal denticlesand some what an gu lar dis po si tion of pro cesses as seen in profile.
Oc cur rence. — The P. trachytera to P. jugosus zones at Łagów, Ostrówka, Kowala, and Miedzianka.
Palmatolepis ampla Müller, 1956(Figs 100A–G and 135)
Type ho ri zon and lo cal ity: Sam ple 217 of Platyclymenia or Clymenia Stufe from Seßacker near Oberscheld in the RhenishSlate Moun tains (Ziegler 1975).
Ma te rial. — 562 spec i mens.Di ag no sis. — Plat form of strongly sin u ous P1 el e ments or na mented with ran domly dis trib uted tu ber cles
on its pos te rior part and trans verse rows of small tu ber cles on the an te rior lobe, P2 el e ments with low cuspand wide posterior platform.
Re marks. — Ap pa ra tus was re stored by my self un der the name P. rugosa (Dzik 1991). The P1 el e mentsshow a rel a tively nar row range of vari abil ity for a Palmatolepis spe cies and are eas ily dis tin guish able fromas so ci ated P. perlobata.
Oc cur rence. — The C. quadrantinodosa and C. marginifera zones at Łagów, Kowala, and Miedzianka.
Palmatolepis trachytera Ziegler, 1960(Figs 100H–O and 135)
Type ho ri zon and lo cal ity: Sam ple 10 of gray cephalopod lime stone 0.3 m above layer with Prolobites delphinus andPlatyclymenia involuta (Ziegler 1975).
Ma te rial. — 989 spec i mens.Di ag no sis. — Plat form of P1 el e ments with sharp ridge or a row of tu ber cles along its an te rior margin.Re marks. — P1 el e ments are flat and sinous in ap pear ance. They vary much in de vel op ment of the pos te −
rior pointed lobe. In early pop u la tions the pos te rior lobe is sharply pointed and elon gate (Fig. 100H), whereas in later pop u la tions it may be vir tu ally missing.
The P2 el e ments bear a min ute cusp and denticles in prox im ity of it are lower than oth ers. The pro cessesmeet at an an gle in the plane of the plat form, which re sults in its boo mer ang−like shape. Width and or na men −ta tion of the plat form vary and ex tremely smooth, wide−plat form el e ments re sem ble some what el e ments ofConditolepis.
The sym me try transistion se ries el e ments dif fer from those of P. schindewolfi in a more gen er al ized ap −pear ance, that is, in usual dom i na tion of the cusp, sharp denticulation and their less sin u ous ar range ment.Oth er wise they are sim i lar to those of other Palmatolepis species.
Oc cur rence. — Zone of its own at Łagów, Ostrówka, Miedzianka, Kowala, and Jabłonna.
Palmatolepis rugosa Branson et Mehl, 1934(Figs 100PO–S and 135)
Type ho ri zon and lo cal ity: Saverton Shale seven miles north of Mon roe City, Mis souri (Ziegler 1975).
Ma te rial. — 664 spec i mens.Di ag no sis. — P1 el e ments with trans versely ribbed an te rior lobe.Re marks. — There is a gap in oc cur rence be tween P. trachytera and P. rugosa in the Holy Cross Moun −
tains but the lat er ally sin u ous shape of P2 el e ments sug gests a close re la tion ship be tween these spe cies. In somespec i mens of late P. trachytera the an te rior ridge splits into a row of tu ber cles. There would not be much tochange this into the tuberculated an te rior field of P. rugosa, but the change re mains to be domonstrated.
Oc cur rence. — The P. styriacus and P. jugosus zones at Ostrówka, Miedzianka, Kowala, and Jabłonna.
FAMENNIAN CONO DONTS AND AMMONOIDS 137
Fig. 100. Ad vanced spe cies of Palmatolepis from the Holy Cross Moun tains. A–G. Palmatolepis ampla Müller, 1956 from theC. marginifera Zone at Łagów (sam ple Ł−5). P1 (A, B), P2 (C), S1 (D), S2 (E), S3–4 (F), and M (G) el e ments; spec i mens ZPALcXVI/2895–2901, re spec tively. H–O. Palmatolepis trachytera Ziegler, 1960 from its nom i nal zone at Łagów (H, sam ple Ł−40)and Jabłonna (N, bed 24) and the L. styriacus Zone at Ostrówka (I–O, sam ple Ost−12). P1 (H–J), P2 (K, N, O), S1 (L), and M (M)el e ments; spec i mens ZPAL cXVI/2902–2909, re spec tively. P–T. Palmatolepis rugosa Branson et Mehl, 1934 from the P.jugosus Zone at Ostrówka (P–S, sam ple Ost−5) and the late L. styriacus zone at Kowala (T, sam ple Ko−6). P1 (P–R, T), and P2 (S)
el e ments; spec i mens ZPAL cXVI/2919–2922, and 2910, re spec tively.
®