Embed Size (px)
Citation preview

PL PA 601 (2004) Lecture 2THE MOST IMPORTANT MOLECULAR BIOLOGICAL TOOLS FORPHYTOPATHOGENIC BACTERIA
Prelude: A brief history of molecular plant pathology from the perspective of evolving technologiesA. A schematic of the production of a typical disease factor and the need to:
1. locate (tag) genes2. clone genes3. direct mutations into known genes
B. Mutagenesis overview1. random chemical2. random transposon3. directed by homologous recombination
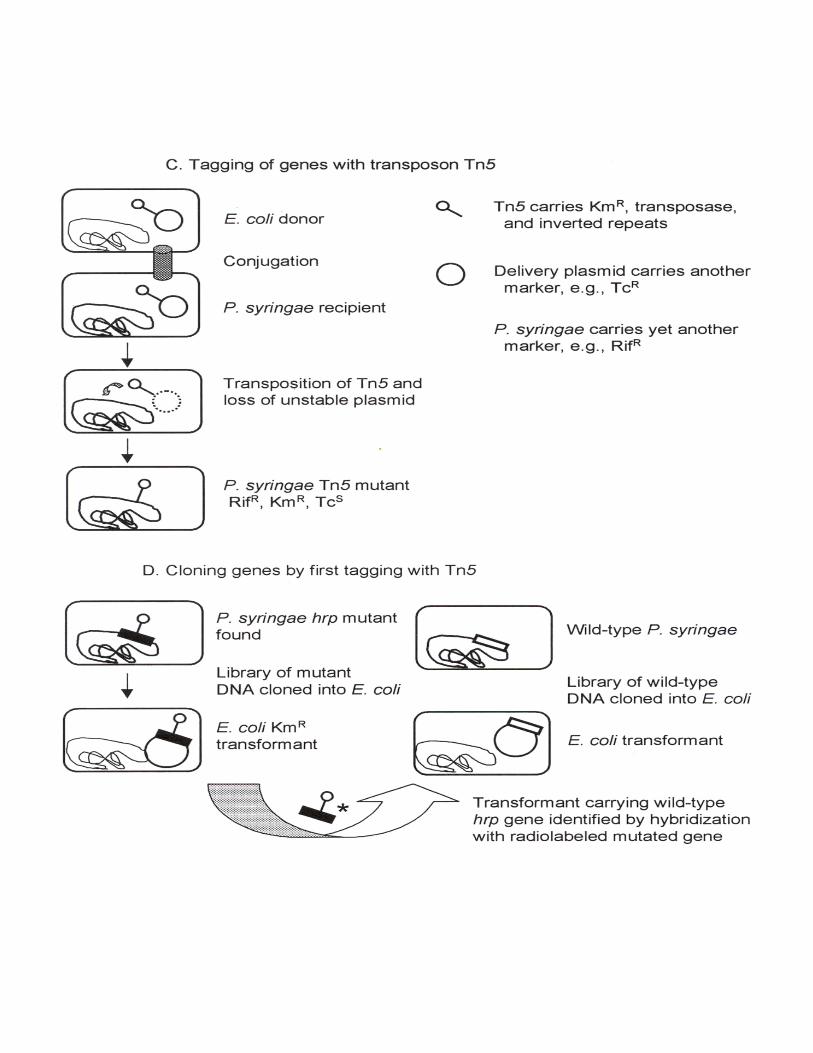
C. Mutagenesis (and tagging) of genes with transposon Tn5 delivered on a conjugative, suicidevector (Mills 1985)
1. "random" transposition2. generally one insertion per chromosome3. strong (and polar) mutations4. mutated gene tagged with resistance to kanamycin (and other antibiotics) and Tn5 DNA,
whose location can be probedD. Cloning genes (e.g., hrp) by first tagging with Tn5 (Lindgren et al. 1986)E. Cloning genes (e.g., pel) by screening E. coli clones for a new phenotype or production of a newantigen (Collmer 1987 pp255-258)F. Cloning genes (e.g., avr) by conjugation of a DNA library contained in a broad host range cosmidvector into a related strain followed by screening of transconjugants for a new phenotype (Staskawiczet al. 1984)G. Directed (=marker-exchange) mutagenesis (Ruvkun and Ausubel 1981; Collmer 1987 Fig. 1)H. Reporter gene transposons and their use
1. Tn3HoHo1 (Stachel et al. 1985)I. Altering gene expression with foreign promoters (e.g., pel; Keen and Tamaki 1986)J. DNA sequencing and the new era of bacterial phytopathogen genomics (see
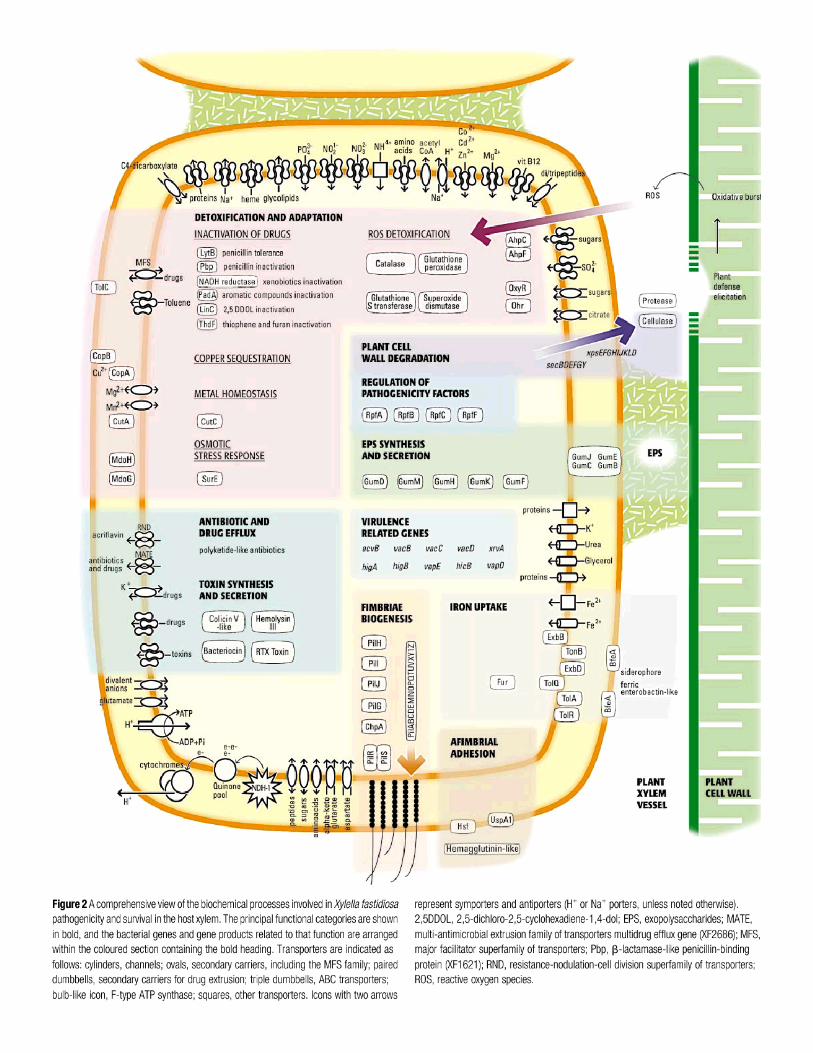
http://www.tigr.org/%7Evinita/PPwebpage.html) for the TIGR plant pathogens web page1. Whole-genome analyses and identification of pathogenicity islands and other horizontally-acquired regions (Lawrence and Roth 1996; Ochman and Moran 2001)2. Xylella fastidiosa: the first plant pathogenic bacterium to be sequenced – a black box openedwide (Simpson et al. 2000)3. Ralstonia solanacearum and Xanthomonas axonopodis pv. citri vs. X. campestris pv.campestris – early lessons from comparative genomics (Salanoubat et al. 2002; Da Silva et al.2002)4. Pseudomonas syringae pv. tomato DC3000 – the power of combining computational andexperimental approaches: experiment-based identification of virulence genes > training set >recognition of pattern useful for genomewide search > comprehensive identification of candidatesworthy of further experimental testing > refined training set (Fouts et al. 2002; Petnicki-Ocwiejaet al.. 2002; Buell et al. 2003; Collmer et al. 2002; Guttman et al. 2002; Greenberg and Vinatzer2003)5. Microarray analysis: global gene expression profiling: early lessons from Erwiniachrysanthemi (Cummings and Relman 2000; Okinaka et al. 2002).6. Clues to function increasingly found through improved tools for finding motifs, folds, andother predictive features in protein sequences (Thornton 2001; Shao et al. 2002).

REFERENCES
Buell, C. R., Joardar, V., Lindeberg, M., Selengut, J., Paulsen, I. T., Gwinn, M. L., Dodson, R. J.,Deboy, R. T., Durkin, A. S., Kolonay, J. F., Madupu, R., Daugherty, S., Brinkac, L., Beanan, M.J., Haft, D. H., Nelson, W. C., Davidsen, T., Liu, J., Yuan, Q., Khouri, H., Fedorova, N., Tran, B.,Russell, D., Berry, K., Utterback, T., Vanaken, S. E., Feldblyum, T. V., D'Ascenzo, M., Deng,W.-L., Ramos, A. R., Alfano, J. R., Cartinhour, S., Chatterjee, A. K., Delaney, T. P., Lazarowitz,S. G., Martin, G. B., Schneider, D. J., Tang, X., Bender, C. L., White, O., Fraser, C. M., andCollmer, A. 2003. The complete sequence of the Arabidopsis and tomato pathogen Pseudomonassyringae pv. tomato DC3000. Proc. Natl. Acad. Sci. USA 100:10181-10186.
Collmer, A. 1987. Pectic enzymes and bacterial invasion of plants. Pages 253-284 in: Plant-MicrobeInteractions: Molecular and Genetic Perspectives, Vol. 2. T. Kosuge and E. W. Nester, ed.Macmillan, New York.
Collmer, A., Lindeberg, M., Petnicki-Ocwieja, T., Schneider, D. J., and Alfano, J. R. 2002. Genomicmining type III secretion system effectors in Pseudomonas syringae yields new picks for allTTSS prospectors. Trends in Microbiol. 10:462-469.
Cummings, C. A., and Relman, D. A. 2000. Using DNA microarrays to study host-microbeinteractions. Emerg Infect Dis 6:513-25.
Da Silva, A. C., Ferro, J. A., Reinach, F. C., Farah, C. S., Furlan, L. R., Quaggio, R. B., Monteiro-Vitorello, C. B., Sluys, M. A., Almeida, N. F., Alves, L. M., Do Amaral, A. M., Bertolini, M. C.,Camargo, L. E., Camarotte, G., Cannavan, F., Cardozo, J., Chambergo, F., Ciapina, L. P.,Cicarelli, R. M., Coutinho, L. L., Cursino-Santos, J. R., El-Dorry, H., Faria, J. B., Ferreira, A. J.,Ferreira, R. C., Ferro, M. I., Formighieri, E. F., Franco, M. C., Greggio, C. C., Gruber, A.,Katsuyama, A. M., Kishi, L. T., Leite, R. P., Lemos, E. G., Lemos, M. V., Locali, E. C.,Machado, M. A., Madeira, A. M., Martinez-Rossi, N. M., Martins, E. C., Meidanis, J., Menck, C.F., Miyaki, C. Y., Moon, D. H., Moreira, L. M., Novo, M. T., Okura, V. K., Oliveira, M. C.,Oliveira, V. R., Pereira, H. A., Rossi, A., Sena, J. A., Silva, C., De Souza, R. F., Spinola, L. A.,Takita, M. A., Tamura, R. E., Teixeira, E. C., Tezza, R. I., Trindade Dos Santos, M., Truffi, D.,Tsai, S. M., White, F. F., Setubal, J. C., and Kitajima, J. P. 2002. Comparison of the genomes oftwo Xanthomonas pathogens with differing host specificities. Nature 417:459-463.
Keen, N. T., and Tamaki, S. 1986. Structure of two pectate lyase genes from Erwinia chrysanthemiEC16 and their high level expression in Escherichia coli. J. Bacteriol. 168:595-606.
Fouts, D. E., Abramovitch, R. B., Alfano, J. R., Baldo, A. M., Buell, C. R., Cartinhour, S., Chatterjee,A. K., D'Ascenzo, M., Gwinn, M. L., Lazarowitz, S. G., Lin, N.-C., Martin, G. B., Rehm, A. H.,Schneider, D. J., van Dijk, K., Tang, X., and Collmer, A. 2002. Genomewide identification ofPseudomonas syringae pv. tomato DC3000 promoters controlled by the HrpL alternative sigmafactor. Proc. Natl. Acad. Sci. USA 99:2275-2280.
Greenberg, J. T., and Vinatzer, B. A. 2003. Identifying type III effectors of plant pathogens andanalyzing their interaction with plant cells. Curr. Opin. Microbiol. 6:20-28.
Guttman, D. S., Vinatzer, B. A., Sarkar, S. F., Ranall, M. V., Kettler, G., and Greenberg, J. T. 2002.A functional screen for the Type III (Hrp) secretome of the plant pathogen Pseudomonassyringae. Science 295:1722-1726.
Lawrence, J. G., and Roth, J. R. 1996. Selfish operons: horizontal transfer may drive the evolution ofgene clusters. Genetics 143:1843-1860.
Lindgren, P. B., Peet, R. C., and Panopoulos, N. J. 1986. Gene cluster of Pseudomonas syringae pv.'phaseolicola' controls pathogenicity of bean plants and hypersensitivity on nonhost plants. J.Bacteriol. 168:512- 522.
Mills, D. 1985. Transposon mutagenesis and its potential for studying virulence genes in plantpathogens. Ann. Rev. Phytopathol. 23:297-320.

Ochman, H., and Moran, N. A. 2001. Genes lost and genes found: evolution of bacterial pathogenesisand symbiosis. Science 292:1096-1099.
Okinaka, Y., Yang, C. H., Perna, N. T., and Keen, N. T. 2002. Microarray profiling of Erwiniachrysanthemi 3937 genes that are regulated during plant infection. Mol Plant Microbe Interact15:619-29.
Pallen, M. J. 1999. Microbial genomes. Mol. Microbiol. 32:907-912.Petnicki-Ocwieja, T., Schneider, D. J., Tam, V. C., Chancey, S. T., Shan, L., Jamir, Y., Schechter, L.
M., Buell, C. R., Tang, X., Collmer, A., and Alfano, J. R. 2002. Genomewide identification ofproteins secreted by the Hrp type III protein secretion system of Pseudomonas syringae pv.tomato DC3000. Proc. Natl. Acad. Sci. USA 99:7652-7657.
Ruvkun, G. B., and Ausubel, F. M. 1981. A general method for site-directed mutagenesis inprokaryotes. Nature 289:85-88.
Salanoubat, M., Genin, S., Artiguenave, F., Gouzy, J., Mangenot, S., Arlat, M., Billault, A., Brottier,P., Camus, J. C., Cattolico, L., Chandler, M., Choisne, N., Claudel-Renard, C., Cunnac, S.,Demange, N., Gaspin, C., Lavie, M., Moisan, A., Robert, C., Saurin, W., Schiex, T., Siguier, P.,Thebault, P., Whalen, M., Wincker, P., Levy, M., Weissenbach, J., and Boucher, C. A. 2002.Genome sequence of the plant pathogen Ralstonia solanacearum. Nature 415:497-502.
Shao, F., Merritt, P. M., Bao, Z., Innes, R. W., and Dixon, J. E. 2002. A Yersinia effector and aPseudomonas avirulence protein define a family of cysteine proteases functioning in bacterialpathogenesis. Cell 109:575-88.
Simpson, A. J. G., et al. 2000. The genome sequence of the plant pathogen Xylella fastidiosa. Nature406:151-157.
Stachel, S. E., An, G., Flores, C., and Nester, E. W. 1985. A Tn3 lacZ transposon for the randomgeneration of beta-galactosidase gene fusions: application to the analysis of gene expression inAgrobacterium. EMBO J. 4:891- 898.
Staskawicz, B. J., Dahlbeck, D., and Keen, N. T. 1984. Cloned avirulence gene of Pseudomonassyringae pv. glycinca determines race specific incompatibility of Glycine max (L.) Merr. Proc.Proc. Natl. Acad. Sci. U.S.A. 81:6024- 28.
Thornton, J. M. 2001. From genome to function. Science 292:2095-2097.

+
Signal
Disease
Plant cell
Bacterium
Nucleus
yfg

Library of pathogen DNA containingyfg is cloned in E. coli
Identify cloned yfg in wild-type library by:•hybridization with known homologous DNA•detecting protein product or phenotype in E. coli•heterologous expression in organism related topathogen
Cloned yfg
Mutated yfg
Random transposon (Tn) mutagenesis
Homologous recombination ofyfg::marker (or reporter) mutationinto pathogen genome
Identify yfg+ clone in wild-typeDNA library by hybridization withcloned yfg::Tn or bycomplementation of mutationwith cloned wild-type yfg
Uses of cloned yfg:
Bioassay: is yfg product sufficient for some biologicalfunction?
Manipulate overexpression to facilitate biochemicalstudy
Explore relatedness and diversity
Detect and identify pathogen
Uses of yfg mutant:
Bioassay: is yfg necessary for some biologicalfunction?
Monitor yfg expression to understand regulation


Library of pathogen DNA containingyfg is cloned in E. coli
Identify cloned yfg in wild-type library by:•hybridization with known homologous DNA•detecting protein product or phenotype in E. coli•heterologous expression in organism related topathogen
Cloned yfg
Mutated yfg
Random transposon (Tn) mutagenesis
Homologous recombination ofyfg::marker (or reporter) mutationinto pathogen genome
Identify yfg+ clone in wild-typeDNA library by hybridization withcloned yfg::Tn or bycomplementation of mutationwith cloned wild-type yfg
Uses of cloned yfg:
Bioassay: is yfg product sufficient for some biologicalfunction?
Manipulate overexpression to facilitate biochemicalstudy
Explore relatedness and diversity
Detect and identify pathogen
Uses of yfg mutant:
Bioassay: is yfg necessary for some biologicalfunction?
Monitor yfg expression to understand regulation
+
Signal
Disease
Plant cell
Bacterium
Nucleus
yfg

Da Silva 2002
Salanoubat 2002

Collmer et al. 2002
Greenberg and Vinatzer 2003

Figure 1. Measuring relative gene expression by usingDNA microarrays. Capillary printing is used to array DNAfragments onto a glass slide (upper right). RNA is preparedfrom the two samples to be compared, and labeled cDNA isprepared by reverse transcription, incorporating either Cy3(green) or Cy5 (red)(upper left). The two labeled cDNAmixtures are mixed and hybridized to the microarray, andthe slide is scanned. In the resulting pseudocolor image, thegreen Cy3 and red Cy5 signals are overlaid--yellow spotsindicate equal intensity for the dyes. With the use of imageanalysis software, signal intensities are determined for eachdye at each element of the array, and the logarithm of theratio of Cy5 intensity to Cy3 intensity is calculated (center).Positive log(Cy5/Cy3) ratios indicate relative excess of thetranscript in the Cy5-labeled sample, and negativelog(Cy5/Cy3) ratios indicate relative excess of thetranscript in the Cy3-labeled sample. Values near zeroindicate equal abundance in the two samples. After severalsuch experiments have been performed, the dataset can beanalyzed by cluster analysis (bottom). In this display, redboxes indicate positive log(Cy5/Cy3) values, and greenboxes indicate negative log(Cy5/Cy3) values, with intensityrepresenting magnitude of the value. Black boxes indicatelog(Cy5/Cy3) values near zero. Hierarchical clustering ofgenes (vertical axis) and experiments (horizontal axis) hasidentified a group of coregulated genes (some shown here)and has divided the experiments into distinct classes.(Illustration by J. Boldrick, Stanford University).
Cummings and Relman 2000
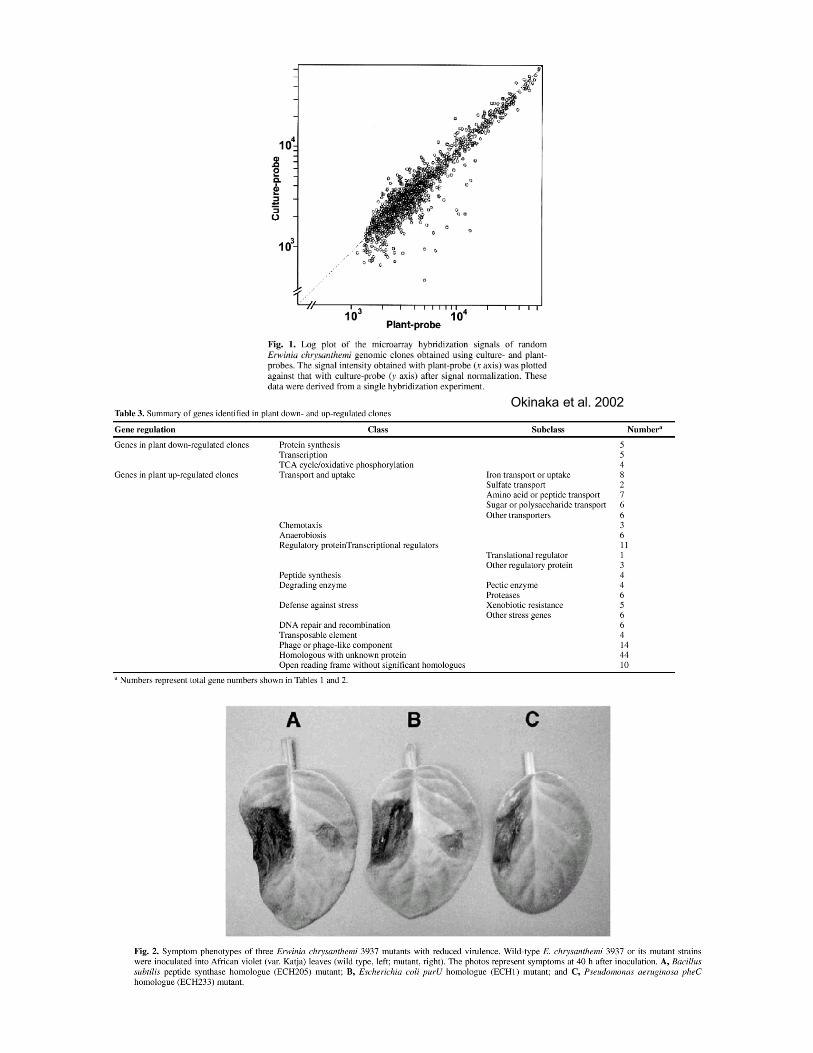
Okinaka et al. 2002

Shao et al. 2002





























