Embed Size (px)
Citation preview

Pietrusewsky M.Department of Anthropology, University of Hawaii2424 Maile Way, Saunders 346Honolulu, Hawaii 96822 U.S.A.Email: [email protected]
Craniometric variation in Southeast Asia and neighboring regions: a multivariate analysis of cranial measurements
Introduction
Southeast Asia, East Asia, and Oceania, which comprise the greater part of the re-gion known as the Indo-Pacific, have a complicated history. Given their geographical proximity and shared antiquity, Australia and Tasmania are sometimes further included in unraveling the prehistory of the region. Because the colonization of the region, nota-
Key words: Southeast Asia, Pacific, East Asia, multivariate analysis, cra-nial measurements
Vol. 23 - n.1-2 (49-86) - 2008HUMAN EVOLUTION
Stepwise discriminant function analysis and Mahalanobis’ generalized distance are applied to 27 landmark measurements recorded in 2,595 male crania representing 56 modern and near modern cranial series from Mainland and Island South-east Asia, East Asia, Australia, and the Pacific. This analysis examines models (e.g., agricultural colonization and continuity models) based on archaeological, historical linguistic, and bio-logical data that attempt to explain the presence of the modern inhabitants of Southeast Asia and the surrounding regions of East Asia, the Pacific, and Australia beginning approximately 40,000 years ago.The results, including the inspection of jackknifed classifica-tion results, canonical plots, and the construction of diagrams of relationship based on Mahalanobis’ distances indicate:• The primary division is between Asian and Australian-Mela-nesian series.• With the exception of the Southern Moluccas series, South-east Asian cranial series form a separate branch with island and mainland subgroupings.• Cranial series representing East Asia and North Asia occupy a branch well separated from Southeast Asia, an association that argues for long term continuity within these regions rather than intrusion or replacement in Southeast Asia. • Island Southeast Asia is the likely homeland for the ancestors of the Polynesians and other inhabitants of remote Oceania.• Several interesting associations (e.g., New Zealand Maori and Southern Moluccas) between Remote Oceania and Island Southeast Asia are discussed.This study demonstrates that human skeletal and dental remains are an important source of information for evaluating models based on archaeological and historical linguistic data and for evaluating the hotly debated LB1 hominin from Liang Bua Cave on Flores Island, Indonesia.

50 PIETRuSEWSky
bly Island Southeast Asia, by modern humans commenced in late Pleistocene and early Holocene, searching for the origins and dispersions of the inhabitants of this immense island world has long guided research in this region. Geologically, during this time, Is-land Southeast Asia was divided into the Sunda and Sahul shelves which separated the islands of Borneo, Sumatra, Java and smaller islands in the west from Australia, New Guinea, Tasmania, and adjoining smaller islands in the east.
The very first human presence in the region can be traced to Homo erectus in Java approximately 1.6 to 1 million years ago. The initial appearance of modern Homo sa-piens coincides with the human crossing of Wallacea during late Pleistocene times (ca 40,000 – 60,000 BP) (Bellwood and Glover, 2004), an event which ultimately led to the peopling of the Sahul Shelf and surrounding islands extending as far east as the Bis-marck Archipelago and Solomon Islands, an area popularly referred to by archaeologists as Near Oceania (Green, 1991a).
The majority of modern inhabitants of the region that extends from Madagascar in the west to Easter Island in the east, however, are the result of a very recent popula-tion expansion beginning approximately 3,500 years BP, one that is tied to the dispersal of people speaking Austronesian languages and the spread of agriculture. This second major colonization event has been linked with the Lapita Cultural Complex, which is as-sociated with the presumed immediate ancestors of the Polynesians and other inhabitants of Remote Oceania (see e.g., Green, 1979; kirch, 1997).
Hence, the modern inhabitants of Southeast Asia and Oceania have been influenced by both very old and very recent movements of people into the region that have shaped their culture and biology. Although the vast majority of the inhabitants of the region have been phenotypically identified as Asian or Mongoloid, the region also includes several short-stature groups (Negritos) found, for example, in Malaysia, Philippines, and Anda-man Islands, as well as Melanesians who extend east and west of New Guinea.
In recent years archaeologists, such as Bellwood (1996, 2005), Glover and Higham (1996), and Higham (1996, 2001) have championed models that argue against both the in situ agricultural development and the diffusion of agricultural technology to the in-digenous hunter-gathering populations in late Pleistocene Southeast Asia, in favor of an agricultural colonization model. This view has also received support, as well as added insights, from linguistics (e.g., Bayard, 1996; Bellwood, 1993, 2001, 2005; Blust, 1996; Diamond and Bellwood, 2003).
Studies in physical anthropology have made substantial contributions to under-standing the biological relationships and origins of the people who occupy Oceania (in-cluding Australasia), and the neighboring continental regions represented by East Asia and Southeast Asia. This corpus of biological data, too numerous to review in this paper, includes measurements and somatological studies of living people, genetic studies be-ginning with traditional blood group antigen marker data, and more recently, molecular genetic evidence such as mtDNA and y-chromosome data. Another source of biological data is that provided by the human skeletal record, both ancient and modern.
50

51CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
The studies that utilize human biological data for the region have advanced alterna-tive views to explain the population history of Southeast Asia and East Asia, models that mainly favor regional continuity and in situ adaptation in Southeast Asia. For example, Turner (1987, 1989, 1990) using dental non-metric traits, has proposed that Southeast Asia was the ultimate source, rather than the recipient of a “southern Mongoloid” (Sun-dadont) population, that ultimately spread northward to give rise to a “northern Mon-goloid” (Sinodont) dental complex. Others who have supported the continuity model include Bulbeck (1982), Hanihara (1993, 1994), Pietrusewsky (1994, 1999, 2004, 2005, 2006a), Pietrusewsky and Douglas (2002) and Pope (1992). In contrast to these studies, the work of Matsumura (1995, 2001, 2006), Matsumura and Hudson (2005), and Mat-sumura et al. (2001) favor admixture models.
Studies of human skeletal remains that span the transition from hunting and gather-ing to agriculture in Southeast Asia, as well as studies of living people and modern and near modern skeletal assemblages, should help demonstrate whether the indigenous peo-ples of Southeast Asia and the neighboring regions of Near Oceania were displaced by later colonists or if population continuity, characterized by a common genetic heritage, bridged the technological and social transition in this region. Alternatively, the evidence from physical anthropology may require a more complex scenario.
The primary focus of this study is to investigate, using craniometric data re-corded in modern and near modern humans, the historical-biological relationships of the modern indigenous peoples of Southeast Asia, East Asia, Australia, and the Pacific Islands. Comparisons of these results with those based on genetic, archaeological, and historical linguistic data will be used to evaluate some of the competing hypotheses that relate to the settlement and colonization of the Pacific and the biological connections within the Asian continent that may inform on population movements in this region of world.
Cranial Morphology and Biodistance Studies
Cranial morphology has played a central role in understanding biological re-lationships and evolution of human populations in physical anthropology. The earliest studies, modeled primarily on racial typological classifications, failed to achieve their predicted goal primarily because of flawed theory and unrefined methodology. Since these earlier attempts there have been improvements in analytical methods, especially the development of multivariate statistical procedures (Mahalanobis, 1936; Mahalanobis et al., 1949; Rao, 1948), which now provide a much more objective means for comparing human groups and for classifying individual specimens (Howells, 1973, 1989, 1995). Likewise, the elucidation of the mechanisms of evolution and the genetics of populations now provide a much more rigorous basis for understanding human population structure and past biological relationships.
51

52 PIETRuSEWSky
Cranial measurements remain an important and valuable source of information for examining relatedness between and within populations, past and present (see e.g., How-ells, 1973, 1989, 1995; Pietrusewsky, 2000; Van Vark and Howells, 1984). The continued interest in metric variation is the result of the precision and repeatability of measurement techniques, the conservative nature of continuous variation, the direct link with the past, the demonstration that craniometric traits have a genetic component (e.g., Droessler, 1981; kohn, 1991; Sjøvold, 1984), and the amenability of measurements to multivariate statistical procedures.
While morphological variation, including craniometric variation, is subject to non-genetic or environmental influences, this category of variation is generally viewed as reflecting genetic similarity and provides the basis for biodistance studies (Buikstra et al., 1990; Larsen, 1997:302-332). Concordance in results based on anthropometric and quantitative genetic analyses strengthens this assertion (konigsberg and Ousley, 1995), which allows distances based on metric data to be interpreted within a population genetic framework.
Material and Methods
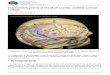
Cranial SeriesA total of 2,595 male crania representing fifty-six cranial series are included in this
study (Table 1, Figure 1). The cranial series represent modern and near modern indig-enous inhabitants of Remote Oceania (Polynesia, Guam, and island eastern Melanesia), Near Oceania (New Guinea, Bismarck Archipelago, Solomon Islands), Australia, Island Southeast Asia, Mainland Southeast Asia, East Asia, and North Asia. The samples, in-cluding their sizes and other details, are given in Table 1. Although exact dates for these specimens are often not known, the majority were collected in the late 19th and early 20th centuries placing them in time and context to the indigenous peoples currently living in these regions. A predominance of the crania from Island Southeast Asia, including Indo-nesia, was examined by the author in museums located in Germany (Leipzig, Dresden, Berlin, Bremen, Göttingen, and Freiburg) and in the Musée de l’Homme in Paris.
The inhabitants of the eastern Lesser Sunda (Nusa Tenggara) and Moluccas Islands to the west of New Guinea show considerable variation. However, because of the limited number of specimens available for this region, they have been combined with specimens from the string of islands extending east of Java from Bali to Aru to represent a Lesser Sunda series. Similarly, crania from Buru and Seram are combined to form the Southern Moluccas series used in the present study.

53CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
Cranial MeasurementsTwenty-seven standard measurements (see Table 2) of the cranial vault and face,
the largest number of measurements comparable to all the series, are used in the present study. The methods used to record these cranial measurements primarily follow those of Martin and Saller (1957) and Howells (1973).
Multivariate Statistical ProceduresStepwise discriminant function analysis and Mahalanobis’ generalized distance sta-
tistic (Mahalanobis, 1936) are applied to the cranial measurements. These methods and the clustering algorithm used to construct the diagram of relationships (dendrograms) are explained in Pietrusewsky (1994, 1997, 1999, 2000).
Stepwise Discriminant Function (Canonical) AnalysisThe major purpose of discriminant function, or canonical, analysis is to maximize
differences between groups by producing a linear array of weighted variables, referred to as discriminant functions or canonical variates, from the original measurements (Tat-suoka, 1971). Typically, the first few functions, or canonical variates, account for most of the variation among groups. In this analysis, the original measurements were selected in a stepwise manner such that, at each step, the measurement that adds most to the separation of the groups was the one entered into the discriminant function in advance of the others (Dixon and Brown, 1979:711). This procedure allows identification of those variables that are most responsible for the observed differentiation between individuals of the various groups. Interpretations of discriminant functions and the patterns of group separation are based on an inspection of standardized canonical coefficient values.
At the end of the stepping process, each individual specimen is classified into one of the original groups based on the discriminant scores it receives through the calcula-tion of posterior (regular classification) and/or typicality (jackknifed classification) prob-abilities (Van Vark and Schaafsma, 1992:244-255). Jackknifed classification represents a common cross-validation procedure in multiple discriminant analysis, where cases are classified without using misclassified individuals in computing the classification func-tion. The ‘correct’ and ‘incorrect’ classification results provide a general guide for as-sessing the homogeneity or heterogeneity of the original series. Another useful feature of this procedure is that it allows group means to be plotted on the first few canonical variates, thus allowing visualization of intergroup relationships. The computer program, BMDP-7M (Dixon and Brown, 1979) is used to perform the stepwise discriminant func-tion analysis, while two-dimensional and three-dimensional plots are made using the SyGRAPH module of SySTAT (Wilkinson, 1992).

54 PIETRuSEWSky
Mahalanobis’ Generalized DistanceMahalanobis’ generalized distance, or the sum of squared differences, provides a
single quantitative measure of dissimilarity (distance) between groups using several variables while removing the correlation between the variables (Mahalanobis, 1936). The significance of these distances is determined using the method of Rao (1952:245), a procedure recommended by Buranarugsa and Leach (1993:17).
The average linkage within group clustering algorithm, or unweighted Pair Group Method Algorithm- uPGMA (Sneath and Sokal, 1973), is the clustering procedure used to construct the diagrams of relationship, or dendrograms, using Mahalanobis’ distances. This latter algorithm combines clusters so that the average distance among all cases in the resulting cluster is as small as possible and the distance between two clusters is taken to be the average among all possible pairs of cases in the cluster. The NTSyS-pc compu-ter software program is used to construct the dendrograms (Rohlf, 1993).
Results
Stepwise Discriminant Function AnalysisStepwise discriminant function was applied to twenty-seven cranial measurements
recorded in fifty-six male cranial series using the computer program, BMDP-7M (Dixon, 1992), written for the mainframe computer.
A summary of the measurements, ranked according to the F-values [tests of equality of group means using classical one-way analysis of variance] received in the final step of discriminant function analysis provides an indication of the discriminatory power of the original variables (Table 2). In this analysis the variables that are ranked the highest (i.e. they contribute the most to the discrimination produced) include three breadth mea-surements (maximum cranial breadth, biorbital breadth, and minimum cranial breadth), basion-nasion length, and nasion-alveolare height.
Eigenvalues, which represent the amount of variance accounted for by each func-tion or canonical variate, expressed as the percentage of total dispersion and level of significance (Rao, 1952:323) for the twenty-seven canonical variates indicate that the first three canonical variates account for 64.0 per cent of the total variation (Table 3). The first twenty eigenvalues are significant at the 1 per cent level, indicating significant heterogeneity for these functions.
Canonical coefficients, those values by which an individual’s measurements may be multiplied to obtain its score, for twenty-seven measurements, for the first three canoni-cal variates are presented in Table 4. Biorbital breadth, nasion-alveolare height, nasio-occipital length, and nasal height (those variables with the highest coefficients regardless of sign) are the most important variables in producing group separation in the first canon-ical variate. This first variate may, therefore, be defined as a biorbital breadth, facial and nasal height, and cranial length discriminator. Minimum cranial breadth, orbital height,

55CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
nasal height, and alveolar breadth, are most responsible for group separation produced in the second canonical variate. Maximum cranial length, nasal height, biorbital breadth, and nasion-alveolare height are primarily responsible for the discrimination produced in the third canonical variate.
A summary of the group classification results, regular and jackknifed (Table 5), in-dicate that Mongolia, Easter Island, Swanport (Australia), Chatham Island, Tuamotu, Pu-rari Delta, Ainu, Tasmania, Western Australia, and Guam are among the series having the best classification results. The poorest jackknifed classification results (less than 20 % of the cases correctly classified to their original group) are found for the Lesser Sunda Is., Solomon Is., Sulawesi, Hainan Is., Sumatra, New Ireland, Borneo, and Nanjing. Three of the groups with the poorest classification results represent Island Southeast Asia.
Closer inspection of some of the jackknifed classification results (Table 6) for fifty-six groups reveals where the most frequent mis-classifications occur for each group.
These classification results serve to highlight those regions that exhibit some of the greatest heterogeneity and possibly where contact with outsiders was the most intense or long-term. It is significant that several of the Island Southeast Asian series, e.g., Lesser Sunda Islands, Sulawesi, and the Southern Moluccas, have some of the poorest classifi-cation results. Only six of the specimens originally assigned to the Lesser Sunda Islands are reclassified to that group. Seven of the Lesser Sunda Island specimens are reclassi-fied to Borneo, four to New Zealand Maori, and three each to Atayal, Java, Sulawesi, Philippines, and Cambodia-Laos. Six each of the Southern Moluccas specimens are reas-signed to two Polynesian series, Marquesas and New Zealand Maori, and five each are reclassified as Sepik and New Ireland.
Five of the New Zealand Maori crania are reclassified as Southern Moluccas and three each are reassigned to the Marquesas, Tuamotu, and Loyalty Island series. Two more each are reclassified as Chatham Island, Solomon Is., and New Ireland. The Solo-mon Island and New Ireland misclassifications are mainly to other cranial series from is-land Melanesia that neighbor the Solomon Island chain and the Bismarck Archipelago.
When the fifty-six group means are plotted on the first two canonical variates (Fig-ure 2), three separate clusters are apparent. Cranial series from Australia, New Guinea, and geographical Melanesia form a coherent group. The cranial series representing Poly-nesia and Guam form a second isolated constellation. The remaining series, from Main-land and Island Southeast Asia and East Asia, form a relatively dense third major group-ing in the same diagram. The cranial series from the Southern Moluccas, Admiralty Islands, and Lesser Sunda Islands are more peripheral members of this latter grouping, occupying an intermediate position between the Polynesian and Australo-Melanesian assemblages.
When the fifty-six group means are plotted on the first three canonical variates (Fig-ure 3), the patterning seen in the previous plot becomes even more obvious. The Aus-tralian and island Melanesian series hover on one side of this diagram. The remaining series form a dense forest opposite these. Several of the Island Southeast Asian series,

56 PIETRuSEWSky
such as Borneo, Lesser Sunda Islands, Southern Moluccas, and the Admiralty Islands are centrally placed; New Zealand Maori align with these. To further facilitate viewing of these results, removal of the Australian series results in the grouping seen in Figure 4. The majority of the Polynesian series are dispersed mainly among cranial series from Mainland and Island Southeast Asia. The groups closest to New Zealand Maori include Sumatra, Borneo, Lesser Sunda Islands, Southern Moluccas, Atayal, and Admiralty Is-lands. The Chinese, Japanese and other northern Asian series tend to separate from the remaining series. The cranial series from New Guinea and island Melanesia form a sepa-rate grouping in this diagram.
Mahalanobis’ Generalized DistanceApplying the uPGMA clustering algorithm to the distances for fifty-six groups
results in the dendrogram shown in Figure 5. Two major branches are evident in this diagram one that includes all the Australian and Melanesian series and a second which, with the exception of New Zealand and the Southern Moluccas, includes all the Poly-nesian, Southeast Asian and East Asian series. The cranial series representing Australia and Tasmania are further differentiated from cranial series representing New Guinea and island Melanesia. Likewise, subgroupings of Polynesian (and Guam), Southeast Asian, and East Asian series are evident in the second major division. Significantly, New Zea-land Maori, Southern Moluccas, and the Admiralty Islands series form a cluster that ultimately links with the Australian-Melanesian branch of this diagram.
Inspection of the ten smallest distances for each of the fifty-six groups (Table 7) gives additional information on biological relatedness that is not readily apparent in the diagrams that result from clustering algorithms. using distance size as a measure of closeness, the groups closest to the southeast Asian series include other Southeast Asian se-ries, most exclusively so. One major exception, however, is the Southern Moluccas series. Based on distance size, the groups closest to the Southern Moluccas include New Zealand (Maori), Biak Is., New Ireland, Solomon Islands, Admiralty, and the Sepik series.
Examining the Polynesian series, eight of the ten closest distances to Tonga-Samoa series in western Polynesia are from Island and Mainland Southeast Asia. The Southern Moluccas and Lesser Sunda Islands are found among the smallest distance for several of the Polynesian series. The groups closest to New Zealand and Chatham Islands, are Southern Moluccas and Lesser Sunda Islands and, for New Zealand, series that occupy the region extending from the Admiralty Islands and Bismarck Archipelago through to the Solomon Islands chain.
For the cranial series located within geographical Melanesia, the Southern Moluc-cas and the Lesser Sunda Islands series appear among the smallest distances in more than half of these series. The smallest distances for the Australian and Tasmanian series are invariably Australian and Melanesian series.

57CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
Discussion
Australia/Melanesia versus Southeast/East Asia and Remote Oceania As shown in previous craniometric analyses (e.g., Hanihara, 1993, 1994; Howells,
1973, 1989, 1995; Pietrusewsky, 1990a, 1994, 1999, 2000, 2005, 2006a, 2006b) the re-sults of this new multivariate analysis of craniometric data demonstrate the presence of two major divisions of modern and near modern humans inhabiting the wider Asian-Pa-cific region. All cranial series from Australia, Tasmania, New Guinea, and geographical island Melanesia, represent one of these divisions. The second major division is repre-sented by all the cranial series from East/North Asia, Mainland and Island Southeast Asia, and Remote Oceania. The striking difference between these two divisions argues for separate origins for the indigenous inhabitants of these two regions.
The indigenous inhabitants of Australia, Tasmania, and geographical Melanesia share a common origin, one that is unrelated to the ultimate origin of the modern in-habitants of Southeast Asia and East Asia. The sharp contrast between Polynesian and Australo-Melanesians series further lends support for archaeological and linguistic mod-els that hypothesize an earlier colonization of Australia, New Guinea, and neighboring regions of Near Oceania and a much later colonization that led to the peopling of previ-ously uninhabited Remote Oceania.
Southeast Asia and North/East AsiaThe results of the present analysis of cranial measurements also allow an examina-
tion of some of the current archaeological models that have been advanced to explain the population history of Southeast Asia and East Asia. For example, Bellwood (1996, 1997, 2005) and Diamond and Bellwood (2003) have recently argued for a population displacement to account for the people who now inhabit the Indo-Malaysian Archipela-go. Higham (1996, 2001) has advanced similar explanations to account for the influx of people and culture associated with rice agriculture in Mainland Southeast Asia. Specifi-cally, these models maintain that the indigenous inhabitants of Southeast Asia were re-placed by an immigrant group of people of a more northern origin, or, to use Bellwood’s terminology, “Australoids” were displaced by “Mongoloids”. Such a scenario should, at least in theory, result in the presence of a somewhat hybridized population living in this region. The work of Matsumura (1994, 1995, 2001, 2006), Matsumura and Hudson (2005), and Matsumura et al. (2001) using dental and cranial traits favors a similar inter-pretation of population history.
Alternative models that stress population continuity beginning in the late Pleisto-cene to account for the present day inhabitants of Southeast Asia, have also been issued. Important among these is the work of Turner (1987, 1989, 1990, 1992a, 1992b), focusing on dental non-metric traits, and the recognition of two polar dental complexes, Sunda-donty for Southeast Asia and Polynesia, and Sinodonty for the inhabitants of East Asia. Others whose work supports continuity in the region include Bulbeck (1982), Hanihara

58 PIETRuSEWSky
(1993, 1994), Pietrusewsky (1994, 1999, 2004, 2005, 2006a), Pietrusewsky and Douglas (2002) and Pope (1992).
The diagram of relationship based on Mahalanobis distances (Figure 5) shows a clear separation between East/North Asian and Southeast Asian cranial series. Likewise, these results show a clear connection between Mainland and Island Southeast Asian series.
Generally, the distances closest to modern (non-Ainu) Japanese series are other Jap-anese series and those from korea and Manchuria in northeast Asia. Similarly, the groups closest to modern Chinese are other modern Chinese series and those from northeast Asia. One Southeast Asian series, Vietnam, is relatively close to several of the modern Chinese series but there is otherwise little evidence for a connection between North/East Asia and Southeast Asia. Matsumura’s (2006) study of earlier and later humans finds ad-ditional evidence for admixture between Northeast Asians and Southeast Asians.
With the exception of Cambodia-Laos and the Philippines, Island and a Mainland Southeast Asian series fall into their respective branches in the dendrogram seen in Fig-ure 5. Inspection of the smallest distances (Table 7) confirms that the groups closest to the Southeast Asian series are other Southeast Asian series and not the East Asian se-ries. An exception is the Southern Moluccas series, which has as its closest group, New Zealand Polynesian Maori, followed by three cranial series from Melanesia: Biak, New Ireland, and Solomon Islands.
Closer inspection of the jackknifed classification results (Table 6) further reveals that only a few of the East and Northeast Asian specimens reclassify as Southeast Asians. The classifications for Hainan Island are an exception with ten of the misclassified Hain-an Island crania being assigned to a Southeast Asian series in this table.
In summary, the present craniometric results indicate a clear distinction between the inhabitants of East/North Asia and Southeast Asia (mainland and insular), a distinc-tion that implies long term in-situ development within each of these regions and argues against displacement to account for the present day inhabitants of Southeast Asia. Simi-lar conclusions have been reached by Turner (1987, 1989, 1990, 1992a, 1992b) using dental morphology and Hanihara (1993) using craniometric data.
Island Southeast Asia/Polynesian Homeland?The results of this craniometric analysis also support an ancestral Polynesian home-
land in East/Southeast Asia and not one within geographically-adjacent Melanesia. The groups closest (i.e. have the smallest distances) to Polynesian series include several Is-land Southeast Asian series (e.g., Lesser Sunda Islands, Sulawesi, Southern Moluccas).
While the Polynesian series form a discrete and isolated cluster in the present study, one unexpected association found is the connection between New Zealand Maori (a Polynesian series) and the Southern Moluccas cranial series from eastern Indonesia, seen in the dendrogram of Mahalanobis distances in Figure 5. These two groups connect with the Admiralty Islands forming an isolated branch peripheral to a larger grouping that includes all of the remaining series from island Melanesia and New Guinea. These affini-ties are confirmed by the distance and classification results. Inspection of the smallest

59CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
distances (Table 7) indicates that New Zealand is the group closest to Southern Moluc-cas. Likewise the Southern Moluccas is the group closest to New Zealand in this table. The classification results (Table 6) included reassignment of several Southern Moluccas crania to New Zealand, Marquesas, and Hawaii, all series from Polynesia.
Recent studies that use molecular genetic data (e.g., kayser et al., 2000; Hagelberg, 1998; Lum and Cann, 2000; Melton et al., 1995; Merriwether et al., 1999; Oppenheimer and Richards, 2001a, 2001b; Redd et al., 1995; Richards et al., 1998; Su et al., 2000) generally support an East Asian or Southeast Asian origin of the Polynesians. More spe-cifically, Oppenheimer (1998) and Oppenheimer and Richards (2001a, 2001b, 2003) point to a Polynesian homeland somewhere in eastern Island Southeast Asia, a view which is consistent with the results presented in this paper.
These results also demonstrate connections between the Southern Moluccas and coastal New Guinea (e.g., Biak Is., Sepik, and Admiralty Is.) and cranial series from New Ireland through the Solomon Island chain. These biological relationships suggest a shared ancestry as well as possible admixture between eastern Indonesia and groups further to the east in New Guinea and Melanesia. This connection lends support to ar-cheological models that favor intrusion of new people and ideas along with interaction between the earliest Austronesian-speakers and the indigenous peoples in a “voyaging corridor” that stretches from eastern Indonesia to the Bismarck and Solomon Islands (Irwin, 1992; Green, 1991b). The results of the present craniometric analysis also dem-onstrate that the patterning within the islands of Indonesia is clinal in nature running west to east along this island chain.
Ainu, Ryukyu Islanders, and Atayal Contrary to the view expressed by Brace and colleagues (Brace and Tracer, 1992;
Brace et al., 1990) a close biological connection between the Ainu and Polynesians is not supported by the present multivariate craniometric results. Rather, the Ainu are members (albeit marginal) of a greater East/North Asian division and do not connect directly with any of the Polynesian series. Similar conclusions has been reached by several different researchers using skeletal evidence (see e.g., Hanihara, 1993).
Further, the present results do not support an eastern Asian (i.e., Taiwan) ancestral homeland for the Polynesians. None of the Atayal (Taiwan Aboriginal) specimens re-classify as Polynesian in Table 6. None of Polynesians series were found to reclassify as Atayal in the same table.
There is some evidence of an association between the Ryukyu Islands and the Viet-nam cranial series in the results of this analysis. The group closest to the Ryukyu in the distance analysis is Vietnam. Four of the Ryukyu crania are reclassified to Vietnam in the jackknifed classification results. However, like the Ainu series, the Ryukyu Island cranial series is part of a greater East Asian/North Asian grouping. There is no evidence in the present results connecting the Ainu and Ryukyu Islanders.
As was the case for the Ryukyu Island series, the Atayal connect to several cranial series from eastern and northern Asia.

60 PIETRuSEWSky
34
Table 1: Fifty-six male cranial series used in the present study.
Series Name(abbrev.)
No. ofCrania
Location1
and Number ofCrania
Remarks
Polynesia
1. Tonga-Samoa(TOG)
19 BER-3; AMS-2;DRE-1; PAR-1BPB-4; AIM-2;AUK-5; SIM-1
Fourteen specimens are from Tonga and five are fromSamoa. Included in the Tongan series are three skulls fromPongaimotu excavated by McKern in 1920; two from To-At-1,2 excavated by Janet Davidson in 1965; and five from To-At-36 excavated by Dirk Spennemann in 1985/6. Theremaining specimens are from museums in Berlin, Paris andSydney. Although the exact dates for a few specimens arenot known, the majority are believed to be prehistoric.
2. EasterIsland(EAS)
50 BER-5; DRE-9;PAR-36
Most of the crania in Paris were collected by Pinart in 1887at Vaihu and La Perouse Bay, Rapa Nui (Easter Island). Theexact dates of these specimens are not known.
3. Hawaii(HAW)
60 BPB-20; HON-20;SIM-20
An equal number of specimens have been randomly chosenfrom three different skeletal series: Mokapu (Oahu),Honokahua (Maui), and Kauai. All specimens are presumedto be prehistoric (pre-1778).
4. Marquesas(MRQ)
63 PAR-49; LEP-1;BLU-1; BPB-12
Crania are from four islands, Fatu Hiva, Tahuata, Nuku Hivaand Hiva Oa. The exact dates of these specimens are notknown.
5. NewZealand(NZ)
50 BRE-3; PAR-21;SAM-1; AIM-13;GOT-1; ZUR-5;DRE-6
A representative sample of New Zealand Maori crania fromthe North and South Islands of New Zealand. The exactdates of these specimens are not known.
6. ChathamIsland (CHT)
45 DUN-8; OTM-2WEL-4; CAN-10AIM-3; DRE-5AMS-2; DAS-3GOT-4; PAR-4
Moriori crania from the Chatham Island, New Zealand. Theexact dates of these specimens are not known.
7. SocietyIslands(SOC)
44 PAR-33; BPB-11 Crania are from the island of Tahiti, Society Islands. Theexact dates of these specimens are not known.
8. TuamotuArchipelago(TUA)
18 PAR-18 The majority of the specimens are from Makatea in theTuamotu Archipelago. The exact dates of these specimensare not known.
IslandMelanesia
9. Fiji(FIJ)
42 BER-1; SAM-3 ;QMB-1; DRE-4FRE-3; CHA-1;BPB-11; PAR-7AMS-3; DUN-6;SIM-2
Crania are from all major islands including the Lau Group inthe Fiji Islands. The exact dates of these specimens are notknown.
10. Vanuatu(VAN)
47 BAS-47 Most of the specimens were collected by Felix Speiser in1912 from Malo, Pentecost and Espirtu Santo Islands.Vanuatu. The exact dates of these specimens are not known.

61CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
35
11. LoyaltyIslands(LOY)
50 BAS-43; PAR-7 Crania are from Mare, Lifou, and Ouvea Island Groups,Loyalty Islands The exact dates of these specimens are notknown.
12. NewCaledonia(NCL)
50 BAS-34; PAR-16 Crania are from several coastal and inland locations on NewCaledonia. The majority of these specimens were collectedin the late 19th century. The exact dates of these specimensare not known.
13. SantaCruz Islands(SCR)
46 SAM-4; AMS-2;BAS-40
The crania in Basel were collected by Felix Speiser in 1912(Speiser, 1928). The exact dates of these specimens are notknown.
14. SolomonIslands(SOL)
49 DRE-3; BER-1;NMV-1; QMB-3;AMS-16; DAS-10;BAS-14;GOT-1
Crania are from New Georgia (5), Guadalcanal (9), SanCristobal Island (7), and other locations in the SolomonIslands. The exact dates of these specimens are not known.
15. NewBritain(NBR)
50 CHA-20; DRE-30 The specimens from New Britain in Dresden were collectedby A. Baessler in 1900 and those in Berlin were collected byR. Parkinson in 1911.These specimens were collected fromtrading posts near Rabul in the Gazelle Peninsula and mostlikely represent Tolai crania (see Pietrusewsky, 1990b: 236-237; Howells, 1973: 24-25). The exact dates of thesespecimens are not known.
16. NewIreland(NIR)
53 AMS-4; BER-2;BLU-6; DRE-18;GOT-15; QMB-1;SAM-6; TUB-1
Most of the crania in Dresden were collected by Pöhl in1887-1888 from the northern end of the island; thespecimens in Göttingen were collected during the SüdseeExpedition in 1908. The exact dates of these specimens arenot known.
17. AdmiraltyIslands(ADR)
50 DRE-20; GOT-9;CHA-6; TUB-15;
Specimens from Hermit, Kaniet and Manus Islands of theAdmiralty Islands. The exact dates of these specimens arenot known.
New Guinea
18. Sepik R.(SEP)
50 DRE-33; GOT-10;TUB-7
The specimens in Dresden were collected by OttoSchlaginhaufen in 1909 from various locations along theSepik River, Papua New Guinea. The exact dates of thesespecimens are not known.
19. BiakIsland(BIK)
48 DRE-48 Most (45) of the specimens were collected by A.B. Meyer in1873 on Biak Island (Mysore), Geelvink Bay, Irian Jaya.The exact dates of these specimens are not known.
20. PurariDelta(PUR)
50 DRE-50 Decorated (engraved) skulls obtained by Gerard andWebster between 1900 and 1902 are from along the PurariRiver and Purari Delta regions, Papua New Guinea. Theexact dates of these specimens are not known.
Australia/Tasmania
21. MurrayR. (MRB)
50 AIA-39; DAM-11 Australian Aboriginal crania were collected by G.M. Blackalong the Murray River (Chowilla to Coobool) in New SouthWales between 1929-1950. The exact dates of thesespecimens are not known.
22. NewSouth Wales(NSW)
62 AMS-21; DAS-41 Australian Aboriginal crania from the coastal locations inNew South Wales. The exact dates of these specimens arenot known.

62 PIETRuSEWSky
36
23.Queensland(QLD)
54 AMS-21; DAS-3;QMB-30
Australian Aboriginal crania from the southeastern andmiddle-eastern regions of Queensland. The exact dates ofthese specimens are not known.
24. NorthernTerritory(NT)
50 AIA-4; AMS-3;MMS-1; NMV-38;QMB-1; SAM-3
Australian Aboriginal crania from Port Darwin (39) andArnhemland (36) in the Northern Territory, Australia. Theexact dates of these specimens are not known.
25. Swanport,S.A. (SAS)
36 SAM-36 Australian Aboriginal crania representing the Tarildekaldand Warki-Korowalde tribes in the lower Murray Riverbasin. The specimens were collected by F. R. Zeitz in 1911from an aboriginal cemetery located approximately 10 kmsoutheast of the Murray Bridge in South Australia (Howells,1973:21). The exact dates of these specimens are not known.
26. WesternAustralia(WA)
47 WAM-47 Australian Aboriginal crania from central (20), eastern (4),northern (14), and southern (9) regions of Western Australia.The exact dates of these specimens are not known.
27. Tasmania(TAS)
26 THM-22; CHA-1;SAM-2; NMV-1
The crania represent Tasmanian Aborigines. The exact datesof these specimens are not known.
Micronesia
28. Guam(GUA)
46 BPB-42; PAR-4 Pre-Spanish Chamorro crania associated with latte structurescollected in the 1920's by Hans Hornbostel along TumonBeach, Tumon Bay, Guam. The majority of these specimensrepresent prehistoric (pre-1521) Chamorro.
IslandSoutheastAsia
29. Sumatra(SUM)
39 BER-1; BRE-1;DRE-5; LEP-4;PAR-3; ZUR-25
The specimens in Zurich are designated “Battak”, specificlocations within the island of Sumatra are not known. Theexact dates of these specimens are not known.
30. Java(JAV)
50 BER-1; BLU-8;CHA-9; DRE-1;LEP-24; PAR-7
Crania were collected from several different localities inJava. The exact dates of these specimens are not known.
31. Borneo(BOR)
34 BER-2; BRE-2;DRE-6; FRE-4;LEP-8; PAR-12
A great many of the specimens are indicated as representingDayak tribes, some have elaborate decorations. The exactdates of these specimens are not known.
32. Sulawesi(SLW)
41 BAS-7; BER-10;DRE-4; FRE-7;LEP-5; PAR-8
An exact location is known for many of these specimens.The exact dates of these specimens are not known.
33. LesserSunda Islands(LSN)
61 BAS-5; BER-15;BLU-2; CHA-1;DRE-24; LEP-1;PAR-6; ZUR-7
Crania from Bali (13), Flores (9), Sumba (1), Lomblem (2),Alor (2), Timor (11), Wetar (2), Leti (4), Barbar (1),Tanimbar (13), Kai (2) and Aru (1) Islands of the LesserSunda Islands. The exact dates of these specimens are notknown.
34. SouthernMoluccasIslands(SML)
65 FRE-48; DRE-17 Crania are from Seram (48) and Buru (17) Islands of theSouthern Molucca Islands. The exact dates of thesespecimens are not known.
35. Sulu(SUL)
38 LEP-1; PAR-37 The specimens in Paris were collected by Montano-Reycirca 1900. The exact dates of these specimens are notknown.

63CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
37
36.Philippines(PHL)
28 BER-9; DRE-19 Most specimens are from Luzon Island. The exact dates ofthese specimens are not known.
MainlandSoutheastAsia
37. Vietnam(VTN)
49 HCM-49 Near modern crania from Hanoi (Van Dien Cemetery) andHo Chi Minh City.
38. BachucVillage,(BAC)
51 BAC-51 Victims of the 1978 Khmer Rouge massacre in BachucVillage in western Angiang Province, Vietnam.
39. Cambodia& Laos(CML)
40 PAR-40 A combined sample of crania from various locations inCambodia and Laos collected between 1877 and 1920. Theexact dates of these specimens are not known.
40. Thailand(THI)
50 SIR-50 Most of the specimens represent dissecting room cases fromBangkok.
41. Burma(BUR)
16 ZUR-16 The crania in Zurich are from a series (Cat. Nos. 93-125) ofskulls collected in Mandalay, Myanmar (Burma), describedin a catalogue dated circa 1900. The exact dates of thesespecimens are not known.
East Asia
42. Kanto(KAN)
50 CHB-50 A dissecting room population of modern Japanese from theKanto District of eastern Honshu. The majority of theindividuals were born during the Meiji period (1868-1911)and died well before 1940.
43. Tohoku(TOH)
53 SEN-53 Dissecting room specimens of modern Japanese from theTohoku District in northern Honshu Island.
44. Kyushu(KYU)
51 KYU-51 Modern Japanese which derive mostly from FukuokaPrefecture in Kyushu Island. Other specimens are fromYamaguchi, Saga, Nagasaki and adjoining prefectures.
45. Ainu(AIN)
50 SAP-18; TKM-5;TKO-27
Modern to near modern skeletons collected by Koganei in1888-89 from abandoned Ainu cemeteries in Hokkaido(Koganei, 1893-1894).
46. RyukyuIslands(RYU)
60 KYO-18; KAN-21;RYU-8; KYU-5;TKO-8
Eighteen near modern crania are from Tokunoshima Islandof the Amami Islands located north of the Okinawa Group inthe central Ryukyu Islands; twenty-one specimens are fromtwo different locations on Kume Island, an island locatedwest of Okinawa Island: Yattchi (17) and Hiyajo (4);twenty-one specimens are from five separate islands in theSakishima Group of the southern Ryukyu Islands: HaterumaIsland (2); Miyako (4); Iriomote Island (2); Ishigaki Island(1), and Yonaguni Island (12).
China/ E. &N.E. Asia
47. Shanghai(SHA)
50 SHA-50 The specimens are mostly from post-Qing (pre-1911)cemeteries in Shanghai.
48. Nanjing(NAJ)
49 SHA-49 The series represents near modern crania exhumed from themodern city of Nanjing, Jiangsu Province, eastern China.

64 PIETRuSEWSky
38
49. Chengdu(CHD)
53 SHA-10; CHE-43 A majority of these specimens date to the Ch'en Dynasty(A.D. 1644-1911) and are from Chengdu, Sichuan Provincein western China. Ten crania are from Leshan, LizhongCounty, Sichuan Province.
50. HongKong (HK)
50 HKU-50 Specimens represent individuals who died in Hong Kongbetween 1978-1979.
51. Taiwan(TAI)
47 TPE-47 Modern Chinese living in Taiwan who trace their immediateorigins to Fujian and Guangdong Provinces on the mainlandof China.
52. HainanIsland(HAI)
47 TPE-47 Near modern Chinese whose ancestors began migrating fromthe Canton region of China to Hainan Island around 200B.C. (Howells, 1989:108). This material was excavated byTakeo Kanaseki in Haikou City on Hainan Island.
53.Manchuria(MAN)
50 TKO-50 Many of the specimens are from northeastern China or theregion formerly referred to as "Manchuria," which todayincludes Heilongjiang and Jilin Provinces and adjacentnorthern Korea. A great many of these specimens areidentified as soldiers, or cavalrymen, who died in battle inthe late 19th century A.D.
54. Korea(KOR)
32 KYO-7; SEN-3,TKM-2; TKO-20
Specific locations in Korea are known for most of these nearmodern specimens.
55. Mongolia(MOG)
50 SIM-50 The skulls are identified as coming from Ulaanbaatar (Urga),Mongolia and were purchased by A. Hrdli_ka in 1912.
56. Atayal(ATY)
36 TPE-28; TKM-7;TKO-1
The Atayal are the second largest surviving Aboriginal tribein Taiwan. The specimens are Atayal slain in the Wusheincident in 1930. The specimens were collected by TakeoKanaseki in 1932 (Howells, 1989:109).
1 AIM, Auckland Institute and Museum, Auckland, New Zealand; AIA, Australian Institute ofAnatomy, Canberra, Australia; AMS, The Australian Museum, Sydney, Australia; AUK,University of Auckland, Auckland, New Zealand; BAC, Bachuc Village, Angiang Province,Vietnam; BAS, Naturhistorisches Museum, Basel, Switzerland; BER, Museum für Naturkunde,Berlin, Germany; BLU, Anatomisches Institut, Universität Göttingen, Göttingen, Germany; BPB,B. P. Bishop Museum, Honolulu, U.S.A.; BRE, Über-see Museum, Bremen, Germany; CAN,Canterbury Museum, Christchurch, New Zealand; CHA, Anatomisches Institut der Chairté,Humboldt Universität, Berlin, Germany; CHB, Chiba University School of Medicine, Chiba,Japan; CHE, Dept. of Anatomy, Chengdu College of Traditional Chinese Medicine, Chengdu,China; DAM, Dept. of Anatomy, University of Melbourne, Melbourne, Australia; DAS, Dept. ofAnatomy, University of Sydney, Sydney, Australia; DUN, Dept. of Anatomy, University of Otago,Dunedin, New Zealand; DRE, Museum für Völkerkunde, Dresden, Germany; FRE, Institut fürHumangenetik und Anthropologie, Universität Freiburg, Freiburg im Breisgau, Germany ; GOT,Institut für Anthropologie, Universität Göttingen, Göttingen, Germany; HCM, Faculty ofMedicine, Ho Chi Minh City, Viet Nam; HON, Honokahua, Maui, Hawaii, U.S.A.; HKU,University of Hong Kong, Hong Kong; KAN, Kanegusuku Storage Room, Board of EducationCultural Division, Kanegusuku, Okinawa, Japan; KYO, Physical Anthropology Laboratory,Faculty of Science, Kyoto University, Kyoto, Japan; KYU, Dept. of Anatomy, Faculty ofMedicine, Kyushu University, Fukuoka, Japan; LEP, Anatomisches Institut, Karl Marx Universität,Leipzig, Germany; MMS, Macleay Museum, University of Sydney, Sydney, Australia; NMV,National Museum of Victoria, Melbourne, Australia; OTM, Otago Museum and Art Gallery,Otago, New Zealand; PAR, Musée de l'Homme, Paris, France; QMB, Queensland Museum,Brisbane, Australia; RYU, University of the Ryukyus, Naha, Okinawa Island, Japan; SAM, SouthAustralian Museum, Adelaide, Australia; SAP, Dept. of Anatomy, Sapporo Medical College,
38
49. Chengdu(CHD)
53 SHA-10; CHE-43 A majority of these specimens date to the Ch'en Dynasty(A.D. 1644-1911) and are from Chengdu, Sichuan Provincein western China. Ten crania are from Leshan, LizhongCounty, Sichuan Province.
50. HongKong (HK)
50 HKU-50 Specimens represent individuals who died in Hong Kongbetween 1978-1979.
51. Taiwan(TAI)
47 TPE-47 Modern Chinese living in Taiwan who trace their immediateorigins to Fujian and Guangdong Provinces on the mainlandof China.
52. HainanIsland(HAI)
47 TPE-47 Near modern Chinese whose ancestors began migrating fromthe Canton region of China to Hainan Island around 200B.C. (Howells, 1989:108). This material was excavated byTakeo Kanaseki in Haikou City on Hainan Island.
53.Manchuria(MAN)
50 TKO-50 Many of the specimens are from northeastern China or theregion formerly referred to as "Manchuria," which todayincludes Heilongjiang and Jilin Provinces and adjacentnorthern Korea. A great many of these specimens areidentified as soldiers, or cavalrymen, who died in battle inthe late 19th century A.D.
54. Korea(KOR)
32 KYO-7; SEN-3,TKM-2; TKO-20
Specific locations in Korea are known for most of these nearmodern specimens.
55. Mongolia(MOG)
50 SIM-50 The skulls are identified as coming from Ulaanbaatar (Urga),Mongolia and were purchased by A. Hrdli_ka in 1912.
56. Atayal(ATY)
36 TPE-28; TKM-7;TKO-1
The Atayal are the second largest surviving Aboriginal tribein Taiwan. The specimens are Atayal slain in the Wusheincident in 1930. The specimens were collected by TakeoKanaseki in 1932 (Howells, 1989:109).
1 AIM, Auckland Institute and Museum, Auckland, New Zealand; AIA, Australian Institute ofAnatomy, Canberra, Australia; AMS, The Australian Museum, Sydney, Australia; AUK,University of Auckland, Auckland, New Zealand; BAC, Bachuc Village, Angiang Province,Vietnam; BAS, Naturhistorisches Museum, Basel, Switzerland; BER, Museum für Naturkunde,Berlin, Germany; BLU, Anatomisches Institut, Universität Göttingen, Göttingen, Germany; BPB,B. P. Bishop Museum, Honolulu, U.S.A.; BRE, Über-see Museum, Bremen, Germany; CAN,Canterbury Museum, Christchurch, New Zealand; CHA, Anatomisches Institut der Chairté,Humboldt Universität, Berlin, Germany; CHB, Chiba University School of Medicine, Chiba,Japan; CHE, Dept. of Anatomy, Chengdu College of Traditional Chinese Medicine, Chengdu,China; DAM, Dept. of Anatomy, University of Melbourne, Melbourne, Australia; DAS, Dept. ofAnatomy, University of Sydney, Sydney, Australia; DUN, Dept. of Anatomy, University of Otago,Dunedin, New Zealand; DRE, Museum für Völkerkunde, Dresden, Germany; FRE, Institut fürHumangenetik und Anthropologie, Universität Freiburg, Freiburg im Breisgau, Germany ; GOT,Institut für Anthropologie, Universität Göttingen, Göttingen, Germany; HCM, Faculty ofMedicine, Ho Chi Minh City, Viet Nam; HON, Honokahua, Maui, Hawaii, U.S.A.; HKU,University of Hong Kong, Hong Kong; KAN, Kanegusuku Storage Room, Board of EducationCultural Division, Kanegusuku, Okinawa, Japan; KYO, Physical Anthropology Laboratory,Faculty of Science, Kyoto University, Kyoto, Japan; KYU, Dept. of Anatomy, Faculty ofMedicine, Kyushu University, Fukuoka, Japan; LEP, Anatomisches Institut, Karl Marx Universität,Leipzig, Germany; MMS, Macleay Museum, University of Sydney, Sydney, Australia; NMV,National Museum of Victoria, Melbourne, Australia; OTM, Otago Museum and Art Gallery,Otago, New Zealand; PAR, Musée de l'Homme, Paris, France; QMB, Queensland Museum,Brisbane, Australia; RYU, University of the Ryukyus, Naha, Okinawa Island, Japan; SAM, SouthAustralian Museum, Adelaide, Australia; SAP, Dept. of Anatomy, Sapporo Medical College,
39
Sapporo, Japan; SEN, Dept. of Anatomy, School of Medicine, Tohoku University, Sendai, Japan;SHA, Institute of Anthropology, College of Life Sciences, Fudan University, Shanghai, China;SIM, National Museum of Natural History, Smithsonian Institution, Washington, D.C., U.S.A.;SIR, Dept. of Anatomy, Siriraj Hospital, Bangkok, Thailand; THM, Tasmanian Museum and ArtGallery, Hobart, Australia; TKM, Medical Museum, University Museum, University of Tokyo,Tokyo, Japan; TKO, University Museum, University of Tokyo, Tokyo, Japan; TPE, AcademiaSinica, Nankang, Taipei, Taiwan; TUB, Institut für Anthropologie u. Humangenetik, UniversitätTübingen, Tübingen, Germany; WAM, Western Australian Museum, Perth, Australia; WEL,National Museum of New Zealand, Wellington, New Zealand; ZUR, Anthropologisches Institut,Universität Zürich, Zürich, Germany.
1

65CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
40
Table 2. Summary ranking of cranial measurements according to F-Values received in the final step ofdiscriminant function analysis (56 male groups, 27 measurements)
Step No. Measurement1,2 F-Value d.f.B/d.f.W3 P4
1 Maximum cranial breadth, M-8 47.5 55/2539 *2 Biorbital breadth, H-EKB 21.8 55/2538 *3 Minimum cranial breadth, M-14 27.0 55/2537 *4 Basion-nasion length, M-5 18.6 55/2536 *5 Nasion-alveolare height, M-48 14.5 55/2535 *6 Maximum cranial length, M-1 13.0 55/2534 *7 Basion-bregma height, M-17 14.2 55/2533 *8 Biauricular breadth, M-11b 12.1 55/2532 *9 Basion-prosthion length, M-48 11.6 55/2531 *10 Nasal height, H-NLH 11.1 55/2530 *11 Nasio-occipital length, M-1d 9.0 55/2529 *12 Nasal breadth, M-54 8.5 55/2528 *13 Bijugal breadth, M-45(1) 7.9 55/2527 *14 Bifrontal breadth, M-43 7.5 55/2526 *15 Alveolare breadth, M-61 7.1 55/2525 *16 Cheek height, H-WMH 7.2 55/2524 *17 Mastoid height, H-MDL 7.0 55/2523 *18 Nasion-bregma chord, M-29 6.0 55/2522 *19 Orbital height left, M-52 5.5 55/2521 *20 Bimaxillary breadth, (M-46 5.3 55/2520 *21 Orbital breadth left, M-51a 5.1 55/2519 *22 Bistephanic breadth, H-STB 4.8 55/2518 *23 Maximum frontal breadth, M-10 5.4 55/2517 *24 Minimum frontal breadth, M-9 4.4 55/2516 *25 Bregma-lambda chord, M-30 3.7 55/2515 *26 Biasterionic breadth, M-12 3.5 55/2514 *27 Mastoid width, H-MDB 3.4 55/2513 *
1 M = Martin and Saller (1957)2 H = Howells (1973)3 d.f.B/d.f.W = degrees of freedom between/degrees of freedom within.4 *p 0.01; n.s. = not significant.

66 PIETRuSEWSky
41
Table 3. Eigenvalues, percentage of total dispersions, cumulative percentage of dispersion, and level ofsignificance for 27 canonical variates resulting from stepwise discriminant function analysis (56 malegroups, 27 measurements)
CanonicalVariate
Eigenvalue % Dispersion Cumulative % Dispersion
d.f.1 p2
1 3.49674 42.3 42.3 81 *2 1.14815 13.9 56.2 79 *3 0.64221 7.8 64.0 77 *4 0.57765 7.0 71.0 75 *5 0.37842 4.6 75.5 73 *6 0.27800 3.4 78.9 71 *7 0.22743 2.8 82.0 69 *8 0.20280 2.5 84.1 67 *9 0.17987 2.2 86.3 65 *10 0.14690 1.8 88.1 63 *11 0.14288 1.8 89.8 61 *12 0.12746 1.6 91.3 59 *13 0.11735 1.3 92.8 57 *14 0.09288 1.0 93.9 55 *15 0.08609 0.9 94.9 53 *16 0.06993 0.7 95.8 51 *17 0.06566 0.7 96.6 49 *18 0.06112 0.7 97.3 47 *19 0.04594 0.5 97.9 45 *20 0.03874 0.4 98.3 43 *21 0.02910 0.3 98.7 41 n.s.22 0.02818 0.3 99.0 39 n.s.23 0.02184 0.2 99.3 37 n.s.24 0.01914 0.2 99.5 35 n.s.25 0.01792 0.2 99.7 33 n.s.26 0.01323 0.1 99.9 31 n.s.27 0.00927 0.1 100.0 29 n.s.
1 d.f. = degrees of freedom = (p + q – 2), (p + q – 4)…2 *p 0.01 when eigenvalues are tested for significance according to criterion [N – _ (p + q)] loge (_+1),where N = total number of crania, p = number of variables, q = number of groups, _= eigenvalue, all ofwhich are distributed approximately as chi-square (Rao, 1952:373).

67CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
42
Table 4. Canonical coefficients of 27 cranial measurements for the first three canonical variates that resultfrom stepwise discriminant function analysis of 56 male groups
Canonical Variate 1 Canonical Variate 2 Canonical Variate 3Maximum cranial length, M-1 0.09531 -0.07784 -0.12924Nasio-occipital length, M-1d -0.10506 0.07835 0.00962Basion-nasion length, M-5 -0.04867 0.03290 -0.06390Basion-bregma height, M-17 -0.02739 0.04489 0.09491Maximum cranial breadth, M-8 -0.05486 -0.00729 0.01122Maximum frontal breadth, M-10 -0.00760 -0.06073 -0.02385Minimum frontal breadth, M-9 0.05815 -0.02206 -0.04411Bistephanic breadth, H-STB -0.04076 0.05167 0.04837Biauricular breadth, M-11b -0.01449 0.05899 -0.07105Minimum cranial breadth, M-14 -0.09765 -0.12480 -0.00312Biasterionic breadth, M-12 0.01597 -0.03996 -0.01639Basion-prosthion length, M-40 0.09912 0.05992 0.03905Nasion-alveolare height, M-48 -0.12187 -0.09167 -0.10230Nasal height, H-NLH 0.10344 0.11183 0.11901Nasal breadth, M-54 -0.01427 -0.09988 0.06281Orbital height left, M-52 -0.03882 0.11779 -0.04946Orbital breadth left, M-51a 0.09511 0.03776 -0.08626Bijugal breadth, M-45(1) -0.01542 0.09059 -0.07226Alveolar breadth, M-61 0.06238 -0.10229 0.02753Mastoid height, H-MDL 0.04426 0.07174 0.01476Mastoid width, H-MDB -0.03702 0.03580 0.03347Bimaxillary breadth, M-46 -0.06772 0.00117 0.03203Bifrontal breadth, M-43 -0.04462 -0.07269 0.00327Biorbital breadth, H-EKB 0.14769 -0.03228 0.10389Cheek height, H-WMH -0.07830 0.09058 0.01884Nasion-bregma chord, M-29 0.05047 0.03779 -0.00023Bregma-lambda chord, M-30 0.01757 -0.01794 0.02564

68 PIETRuSEWSky
43
Table 5. Classification results (Regular and Jackknifed) arranged by groups with the best to the poorestresults showing percentage of correctly assigned cases.
Regular Classification Results Jackknifed Classification Results
Group % Group %
Mongolia 84.0 Mongolia 80.0
Easter Is. 82.0 Easter Is. 76.0
Swanport 80.6 Chatham Is. 75.6
Tuamotu Arch. 77.8 Swanport 75.0
Purari Delta 76.0 Purari Delta 66.0
Chatham Is. 75.6 Western Australia 59.6
Ainu 74.0 Guam 58.7
Tasmania 73.1 Ainu 58.0
Guam 67.4 Tasmania 53.8
Atayal 63.9 Hong Kong 52.0
Taiwan 63.8 Taiwan 51.1
Western Australia 63.8 Tuamotu Arch. 50.0
Burma 62.5 Atayal 50.0
Chengdu 58.5 Chengdu 49.1
Loyalty Is. 58.0 Hawaii 48.3
Tonga-Samoa 57.9 Admiralty Is. 48.0
Hawaii 56.7 Santa Cruz Is. 45.7
Santa Cruz Is. 56.5 Cambodia and Laos 45.0
Hong Kong 56.0 Bachuc 43.1
New Caledonia 56.0 New Caledonia 42.0
Admiralty Is. 56.0 New Zealand 40.0
Cambodia and Laos 55.0 Murray R. 40.0
Murray R. 52.0 Tonga-Samoa 36.8
Tohoku 50.9 Ryukyu Is. 36.7
Bachuc 49.0 Sepik 36.0
Ryukyu Is. 48.3 Manchuria 36.0
Manchuria 48.0 Tohoku 34.0
Sepik 48.0 Kyushu 31.4
Sulu 47.4 New South Wales 30.6
Shanghai 44.0 New Britain 30.0
Korea 43.8 Northern Territory 30.0
New Zealand 42.0 Thailand 30.0
Thailand 42.0 Society Is. 29.5
Biak Is. 41.7 Sulu 28.9

69CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
44
Regular Classification Results Jackknifed Classification Results
Group % Group %
Queensland 40.7 Kanto 28.0
Philippines 39.3 Marquesas 27.0
Kyushu 39.2 Vietnam 26.5
New South Wales 38.7 Shanghai 26.0
Vanuatu 38.3 Biak 25.0
New Britain 38.0 Korea 25.0
Northern Territory 38.0 Burma 25.0
Kanto 38.0 Queensland 24.1
Vietnam 36.7 Loyalty Is. 24.0
Marquesas 36.5 Java 24.0
Fiji 35.7 Fiji 23.8
Borneo 35.3 Southern Moluccas Is. 23.1
Hainan Is. 31.9 Philippines 21.4
Society Is. 31.8 Vanuatu 21.3
Java 30.0 Nanjing 18.4
Southern Moluccas Is. 29.2 Borneo 17.6
New Ireland 28.3 New Ireland 17.0
Nanjing 26.5 Sumatra 15.4
Solomon Is. 24.5 Hainan Is. 12.8
Sumatra 23.1 Solomon Is. 12.2
Sulawesi 19.5 Sulawesi 12.2
Lesser Sunda Is. 14.8 Lesser Sunda Is. 9.8

70 PIETRuSEWSky
45
Tab
le 6
. S
om
e o
f th
e ja
ckk
nif
ed c
lass
ific
atio
n r
esu
lts
ob
tain
ed f
rom
ste
pw
ise
dis
crim
inan
t fu
nct
ion
an
aly
sis
sho
win
g t
he
case
s re
-cla
ssif
ied
at
the
end
of
the
step
pin
g p
roce
ss (
nu
mb
ers
in p
aren
thes
es r
epre
sen
t th
e n
um
ber
of
cran
ia o
rig
inal
ly a
ssig
ned
to
eac
h n
um
ber
of
gro
up
). S
ee T
able
1 f
or
exp
lan
atio
n
of
abb
rev
iati
on
s.
To
ng
a-S
amo
a
(19
)
Eas
ter
Is.
(50
)
Haw
aii
(60
)
Mar
qu
esas
(63
)
New
Zea
lan
d
(50
)
Ch
ath
am I
s.
(45
)
So
ciet
y I
s.
(44
)
Tu
amo
tu
(18
)
TO
G7
RA
P3
8H
AW
29
MR
Q1
7N
Z2
0C
HT
34
SO
C1
3T
UA
9
HA
W3
HA
W2
RA
P4
TU
A8
SM
L5
NZ
2T
UA
11
SO
C3
FIJ
2S
OC
2M
RQ
4S
OC
7M
RQ
3G
UA
2H
AW
5R
AP
2
SO
C1
NIR
2T
OG
3N
Z5
TU
A3
AIN
2M
RQ
2T
OG
1
TA
I1
MR
Q1
CH
T3
FIJ
5L
OY
3JA
V1
CH
T2
NZ
1
GU
A1
NZ
1S
OC
3H
AW
3C
HT
2S
EP
1N
CL
2S
ML
1
JAV
1C
HT
1G
UA
3L
SN
2S
OL
2B
IK1
NIR
2S
UL
1
CM
L1
TU
A1
KA
N2
NIR
2N
IR2
TO
G1
KY
U2
KA
N0
KA
N1
FIJ
1A
DR
1R
YU
2K
YU
1H
AW
1R
AP
1T
OH
0
RY
U1
LO
Y1
TA
I1
RA
P2
RY
U1
RA
P0
FIJ
1K
YU
0
Fij
i
(42
)
Van
uat
u
(47
)
Lo
yal
ty I
s.
(5
0)
New
Cal
edo
nia
(50
)
San
ta C
ruz
(46
)
So
lom
on
Is.
(4
9)
New
Bri
tain
(5
0)
Ad
mir
alty
(50
)
FIJ
10
VA
N1
0N
CL
14
NC
L2
1S
CR
21
NB
R7
NB
R1
5A
DR
24
LO
Y7
NB
R9
LO
Y1
2L
OY
9V
AN
6S
OL
6V
AN
10
SE
P5
CH
T3
BIK
5S
CR
4N
IR3
SE
P5
FIJ
4N
T5
BIK
4
SO
L3
NS
W3
FIJ
3N
T3
NT
4N
IR4
FIJ
3S
ML
3
SU
L2
TA
S3
SO
L3
VA
N2
LO
Y3
SE
P4
SO
L2
BO
R2
SC
R2
LO
Y2
QL
D3
NB
R2
NB
R2
LO
Y3
NIR
2K
AN
1
NB
R2
NC
L2
NZ
2S
EP
2P
UR
2N
CL
3S
EP
2N
AJ
1
TO
G2
NIR
2N
IR2
FIJ
1N
CL
1S
CR
2P
UR
2V
TN
1
WA
1S
AS
2B
IK2
SO
L1
SO
L1
NZ
1N
SW
1S
UL
1
SM
L1
SM
L2
MR
Q1
BIK
1N
IR1
SO
C1
TA
S1
TA
S1
Sep
ik
(50
)
Bia
k
(48
)
Pu
rari
(50
)
Mu
rray
(50
)
New
So
uth
Wal
es
(62
)
Qu
een
slan
d
(54
)
N.T
erri
tory
(50
)
Sw
anp
ort
(36
)
SE
P1
8B
IK1
2P
UR
33
MR
B2
0N
SW
19
QL
D1
3N
T1
5S
AS
27
BIK
6S
EP
6S
OL
3W
A9
QL
D1
4N
SW
10
MR
B7
QL
D3
PU
R4
FIJ
5F
IJ2
NS
W5
SA
S6
MR
B8
WA
6M
RB
2
SO
L4
NZ
3S
CR
2Q
LD
4N
T5
NT
7N
SW
5V
AN
1
NIR
3V
AN
3S
EP
1N
T4
MR
B3
WA
5Q
LD
4N
BR
1
SU
L3
NB
R3
BIK
1V
AN
2S
ML
2V
AN
2S
CR
4N
SW
1
NB
R2
NIR
3M
RB
1L
OY
1S
CR
2S
OL
2S
OL
2A
IN1
AD
R2
PU
R3
NT
1N
CL
1B
IK2
PU
R2
FIJ
2S
LW
0
VA
N1
NS
W2
LS
N1
SE
P1
WA
2G
UA
1V
AN
1V
TN
0
LO
Y1
MR
B1
BA
C1
PU
R1
JAV
1N
BR
1N
CL
1C
ML
0

71CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
46
Tab
le 6
. S
om
e o
f th
e ja
ckk
nif
ed c
lass
ific
atio
n r
esu
lts
ob
tain
ed f
rom
ste
pw
ise
dis
crim
inan
t fu
nct
ion
an
aly
sis
sho
win
g t
he
case
s re
-cla
ssif
ied
at
the
end
of
the
step
pin
g p
roce
ss (
nu
mb
ers
in p
aren
thes
es r
epre
sen
t th
e n
um
ber
of
cran
ia o
rig
inal
ly a
ssig
ned
to
eac
h n
um
ber
of
gro
up
). S
ee T
able
1 f
or
exp
lan
atio
n
of
abb
rev
iati
on
s. C
on
tin
ued
W.
Au
stra
lia
(47
)
Tas
man
ia
(26
)
Gu
am
(46
)
Su
mat
ra
(39
)
Jav
a
(50
)
Bo
rneo
(34
)
Su
law
esi
(41
)
Les
ser
Su
nd
as
(6
1)
WA
28
TA
S1
4G
UA
27
SU
M6
JAV
12
BO
R6
CM
L7
BO
R7
TA
S3
NS
W2
HA
W5
BU
R3
CM
L6
SL
W4
SL
W5
LS
N6
NT
3B
OR
1T
OG
3B
OR
3S
LW
4L
SN
3T
HI
3N
Z4
VA
N2
SM
L1
MR
Q3
TO
H2
LS
N3
JAV
2L
SN
3A
TY
3
NS
W2
WA
1C
ML
2S
HA
2B
AC
3S
OL
2B
OR
3C
ML
3
SA
S2
MR
B1
SU
L1
JAV
2T
HI
3S
UL
2V
TN
2P
HL
3
KY
U1
SE
P1
SU
M1
LS
N2
SU
L3
BIK
1N
IR2
SL
W3
AT
Y1
AD
R1
NZ
1S
UL
2K
AN
1A
DR
1T
OG
2JA
V3
MR
B1
NZ
1N
IR1
CH
D2
AIN
1H
AW
1JA
V2
AIN
2
VA
N1
KO
R1
AIN
1C
HD
1N
CL
1S
UL
2V
TN
2
S.
Mo
lucc
as
(65
)
Su
lu
(38
)
Ph
ilip
pin
es
(28
)
Vie
tnam
(49
)
Bac
hu
c
(51
)
Cam
bo
dia
/Lao
s
(40
)
Th
aila
nd
(50
)
Bu
rma
(16
)
SM
L1
5S
UL
11
PH
L6
VT
N1
3B
AC
22
CM
L1
8T
HI
15
BU
R4
NZ
6C
ML
6V
TN
5P
HL
10
CM
L4
SU
L5
BA
C6
CM
L3
MR
Q6
JAV
3B
AC
2A
TY
4T
HI
4S
OL
3K
AN
4T
OH
2
SE
P5
BO
R3
SL
W2
RY
U4
HA
I3
HA
W2
PH
L3
JAV
1
NIR
5S
LW
3K
YU
1H
K3
VT
N2
SL
W2
CM
L3
SL
W1
BIK
2B
AC
1S
HA
1S
HA
3T
AI
2S
UM
1H
K2
LS
N1
AD
R2
TU
A1
CM
L1
BA
C2
HK
2V
TN
1K
OR
2S
UL
1
NC
L2
MR
Q1
SU
M1
TH
I2
KO
R2
SM
L1
HA
I2
MA
N1
LO
Y2
HA
W1
KO
R1
TA
I1
BU
R2
BO
R1
SL
W2
AD
R1
HA
W2
AT
Y1
SO
L1
SH
A1
CH
D1
VT
N2
NA
J1
Kan
to
(50
)
To
ho
ku
(53
)
Ky
ush
u
(51
)
Ain
u
(50
)
Ry
uk
yu
(60
)
Sh
ang
hai
(50
)
Nan
jin
g
(49
)
Ch
eng
du
(53
)
KA
N1
4T
OH
18
KY
U1
6A
IN2
9R
YU
22
SH
A1
3N
AJ
9C
HD
26
KY
U5
KY
U6
KA
N8
KA
N5
AIN
5N
AJ
9S
HA
7N
AJ
5
TO
H4
KO
R5
TO
H4
TO
H3
VT
N4
HK
5M
AN
6M
OG
4
AT
Y3
AIN
4K
OR
4K
YU
3T
OH
3K
AN
4C
HD
5H
K3
HK
3M
AN
3R
YU
3L
SN
2T
AI
3T
HI
4H
K4
VT
N3
MA
N2
AT
Y3
HK
3R
YU
2K
YU
2C
HD
4K
OR
2S
HA
3
LS
N2
TA
I2
AT
Y2
FIJ
1T
HI
2M
OG
2M
OG
2C
HT
1
TH
I2
KA
N2
TA
I2
HK
1A
TY
2M
AN
2S
UM
2M
AN
1
KO
R2
LS
N2
HA
I2
CH
T1
KO
R2
JAV
1R
YU
2T
AI
1
AIN
1R
YU
1M
AN
2H
AW
1H
AI
2T
OG
1S
OC
1K
OR
1

72 PIETRuSEWSky
47
Tab
le 6
. S
om
e o
f th
e ja
ckk
nif
ed c
lass
ific
atio
n r
esu
lts
ob
tain
ed f
rom
ste
pw
ise
dis
crim
inan
t fu
nct
ion
an
aly
sis
sho
win
g t
he
case
s re
-cla
ssif
ied
at
the
end
of
the
step
pin
g p
roce
ss (
nu
mb
ers
in p
aren
thes
es r
epre
sen
t th
e n
um
ber
of
cran
ia o
rig
inal
ly a
ssig
ned
to
eac
h n
um
ber
of
gro
up
). S
ee T
able
1 f
or
exp
lan
atio
n
of
abb
rev
iati
on
s. C
on
tin
ued
New
Ire
lan
d
(53
)
HK
(50
)
Tai
wan
(4
7)
Hai
nan
(47
)
Man
chu
ria
(50
)
Ko
rea
(32
)
Mo
ng
oli
a
(50
)
Ata
yal
(36
)
NIR
9H
K2
6T
AI
24
HA
I6
MA
N1
8K
OR
8M
OG
40
AT
Y1
3
NB
R7
MA
N4
TO
H4
KO
R6
CH
D4
HA
I5
SH
A3
KA
N3
BIK
6S
HA
3H
AI
3T
AI
4H
K3
TO
H3
CH
D2
KO
R2
AD
R4
NA
J2
KO
R2
TH
I4
KO
R3
KY
U3
RY
U1
KY
U2
NC
L3
KY
U2
VT
N2
BU
R4
RY
U3
NA
J2
TH
I1
AIN
2
LO
Y2
TH
I2
BA
C2
RY
U4
SH
A3
HA
W2
JAV
1S
ML
1
PU
R2
TA
I2
KA
N1
HK
2T
AI
2B
OR
1H
AI
1P
HL
1
SO
L2
TO
G1
KY
U1
MA
N2
KA
N2
VT
N1
TO
H1
TO
H1
SE
P2
CH
T1
RY
U1
GU
A2
TO
H2
AT
Y1
RY
U1
SM
L2
AD
R1
CH
D1
PH
L2
KY
U2
SU
M1
CH
D1

73CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
48
Table 7. The smallest Mahalanobis’ distances for 56 male cranial groups using 27 measurements. Alldistances are significant p # 0.01 unless indicated otherwise
1
Tonga-Samoa Easter Island Hawaii Marquesas
HAW 6.646 NZ 7.365 TOG 6.646 SOC 3.553
JAV 7.806 MRQ 8.645 MRQ 7.340 NZ 4.309
SLW 8.248* HAW 10.682 SUL 7.921 TUA 7.141
CML 8.450* SOC 10.792 JAV 8.100 HAW 7.340
SUL 8.581* FIJ 11.517 NZ 8.505 SML 8.161
LSN 8.765 LOY 12.208 GUA 8.906 CHT 8.214
GUA 8.942 TOG 12.482 SLW 8.943 EAS 8.645
BOR 9.991 NCL 12.627 LSN 9.128 FIJ 9.839
THI 10.247 SOL 12.980 CHT 9.674 LSN 10.703
SUM 10.534 SML 13.002 CML 9.893 GUA 10.705
New Zealand Chatham Island Society Islands Tuamotu
SML 3.557 NZ 7.437 MRQ 3.553 SOC 4.672**
MRQ 4.309 MRQ 8.214 TUA 4.672** MRQ 7.141
SOL 5.577 HAW 9.674 NZ 8.682 NZ 10.793
NIR 6.820 GUA 12.193 HAW 9.937 HAW 14.589
BIK 7.073 SOC 13.740 EAS 10.792 TOG 15.033**
EAS 7.365 SML 14.445 TOG 10.888 EAS 15.061
CHT 7.437 LSN 14.598 FIJ 11.483 SML 15.731
LSN 7.504 SUL 14.724 NCL 12.774 GUA 16.979
FIJ 7.692 SUM 15.134 SOL 13.172 ADR 17.025
ADR 8.097 FIJ 15.136 GUA 13.271 SOL 17.093
Fiji Vanuatu Loyalty Islands New Caledonia
BIK 4.268 NBR 1.741* NCL 3.300 LOY 3.300
SOL 5.026 BIK 4.201 FIJ 5.209 NBR 5.912
LOY 5.209 NIR 4.532 SOL 5.716 NIR 6.174
NIR 5.944 SOL 4.780 NIR 6.721 SOL 6.202
NCL 6.784 SCR 5.182 BIK 6.901 VAN 6.593
VAN 6.971 NSW 5.319 VAN 6.938 SCR 6.609
SML 7.442 QLD 5.895 NBR 7.402 FIJ 6.784
NZ 7.692 NT 5.903 SCR 8.056 SEP 8.005
SEP 7.751 NCL 6.593 QLD 8.475 BIK 8.389
LSN 8.230 SML 6.711 NT 8.862 NT 8.793
Santa Cruz Solomon Islands New Britain New Ireland
NBR 4.722 NIR 2.377** VAN 1.741** SOL 2.377*
NT 5.110 SEP 3.836 NIR 3.461 NBR 3.461
VAN 5.182 BIK 4.112 SOL 4.230 BIK 3.865
SOL 5.904 NBR 4.230 SCR 4.722 SEP 3.966
NCL 6.609 SML 4.371 NCL 5.912 SML 4.233
BIK 6.785 VAN 4.780 NT 6.039 VAN 4.532
NIR 6.814 FIJ 5.026 BIK 6.120 FIJ 5.944
SEP 6.832 LSN 5.448 NSW 6.983 NCL 6.174
PUR 7.548 NZ 5.577 SEP 7.209 LSN 6.207
LOY 8.056 SCR 5.904 LOY 7.402 LOY 6.721

74 PIETRuSEWSky
49
Table 7. The smallest Mahalanobis’ distances for 56 male cranial groups using 27 measurements. Alldistances are significant p # 0.01 unless indicated otherwise (continued)
Admiralty Sepik Biak Purari
SML 5.892 SOL 3.836 NIR 3.865 SEP 6.247
SEP 7.385 NIR 3.966 SML 3.870 SOL 6.385
NIR 7.395 BIK 4.025 SEP 4.025 BIK 7.130
SOL 7.410 SML 5.988 SOL 4.112 SCR 7.548
NZ 8.097 PUR 6.247 VAN 4.201 NT 8.457
BOR 8.134 SCR 6.832 FIJ 4.268 NIR 8.629
BIK 8.345 LSN 7.093 NBR 6.120 FIJ 8.916
LSN 8.493 NBR 7.209 LSN 6.184 NBR 9.018
SUL 8.493 VAN 7.297 SCR 6.785 VAN 9.471
SUM 9.020 ADR 7.385 LOY 6.901 NSW 10.555
Murray River New South Wales Queensland Northern Territory
QLD 2.728* QLD 1.567** NSW 1.567** QLD 2.535*
NT 2.863* MRB 2.868 NT 2.535* MRB 2.863*
NSW 2.868 NT 3.218 MRB 2.728* NSW 3.218
VAN 6.714 VAN 5.319 VAN 5.895 SCR 5.110
WA 6.841 WA 5.892 WA 6.245 VAN 5.903
SAS 7.139 SAS 6.124 SOL 7.277 NBR 6.039
NBR 8.372 NBR 6.983 NBR 7.629 SOL 6.824
TAS 8.778 SOL 7.068 SAS 8.063 WA 7.080
SCR 9.394 BIK 8.745 FIJ 8.433 BIK 7.901
SOL 11.382 TAS 8.967 LOY 8.475 PUR 8.457
Western Australia Swanport Tasmania Guam
NSW 5.892 NSW 6.124 QLD 8.496 HAW 8.906
QLD 6.245 MRB 7.139 VAN 8.497 TOG 8.942
MRB 6.841 QLD 8.063 MRB 8.778 NZ 9.430
NT 7.080 VAN 8.345 NSW 8.967 LSN 9.537
VAN 9.804 NT 9.766 NBR 10.650 BOR 9.944
TAS 10.721 NBR 10.650 WA 10.721 SUM 10.051
SAS 11.019 WA 10.721 NT 12.114 SUL 10.139
NBR 11.154 LOY 12.719 SAS 12.889 MRQ 10.705
FIJ 11.524 TAS 12.889 BIK 14.244 SLW 11.666
BIK 11.765 BIK 13.851 SML 14.778 CML 11.719
Sumatra Java Borneo Sulawesi
BOR 2.207** SLW 2.357** LSN 1.792** JAV 2.357**
LSN 2.577* CML 3.470 SUM 2.207** CML 2.587**
SLW 3.069** LSN 3.611 SLW 2.666** BOR 2.666**
VTN 4.351 SUL 3.781 SUL 4.067** LSN 2.697**
JAV 4.372 THI 4.310 JAV 4.501 SUL 2.826**
SUL 4.730 SUM 4.372 PHL 4.812** SUM 3.069**
PHL 4.950* BOR 4.501 VTN 4.916 PHL 3.485**
KAN 5.820 PHL 5.483 CML 5.180 VTN 5.467
THI 5.858 BAC 5.994 SML 5.313 BAC 5.880
RYU 6.242 VTN 6.246 SOL 6.966 SML 6.311

75CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
50
Table 7. The smallest Mahalanobis’ distances for 56 male cranial groups using 27 measurements. Alldistances are significant p # 0.01 unless indicated otherwise (continued)
Lesser Sunda Is. Southern Moluccas Sulu Philippines
BOR 1.792** NZ 3.557 CML 2.711** VTN 2.358**
SUM 2.577* BIK 3.870 SLW 2.826** SLW 3.485**
SLW 2.697** NIR 4.233 JAV 3.781 LSN 3.749*
JAV 3.749 SOL 4.371 BOR 4.067** BOR 4.812**
PHL 4.663** LSN 4.864 SUM 4.730 THI 4.834
VTN 4.663 BOR 5.313 LSN 4.775 SUM 4.950*
SUL 4.775 ADR 5.892 BUR 6.183** JAV 5.483
SML 4.864 SEP 5.988 PHL 6.821 CML 5.976
SOL 5.448 SLW 6.311 SML 6.881 BAC 6.150
CML 5.913 SUM 6.401 BAC 7.193 RYU 6.775
Vietnam Bachuc Cambodia and Laos Thailand
PHL 2.358** THI 3.109 SLW 2.587** BAC 3.109
RYU 4.245 HAI 3.691 SUL 2.711** SLW 3.981
THI 4.282 VTN 5.081 JAV 3.470* VTN 4.282
SUM 4.351 CML 5.753 BOR 5.180 JAV 4.310
LSN 4.663 SLW 5.880 THI 5.501 PHL 4.834*
BOR 4.916 JAV 5.994 BAC 5.753 HAI 5.412
KOR 5.068 PHL 6.150 LSN 5.913 CML 5.501
BAC 5.081 KOR 6.229 PHL 5.976 KAN 5.769
HAI 5.135 HK 6.887 SUM 6.339 SUM 5.858
SLW 5.467 SUL 7.193 BUR 6.953 KOR 5.963
Burma Kanto Tohoku Kyushu
SUL 6.183** KYU 2.758* KYU 2.155** TOH 2.155**
SUM 6.435** TOH 3.323 KOR 3.182** KAN 2.758*
CML 6.953** KOR 4.157* KAN 3.323 KOR 2.811**
SLW 7.166** THI 5.769 MAN 4.606 RYU 4.736
JAV 7.301* SUM 5.820 ATY 5.363 HAI 4.858
BAC 7.408* AIN 6.309 AIN 6.003 ATY 5.056
THI 7.654* MAN 6.445 NAJ 6.115 MAN 5.146
VTN 9.194 HAI 6.509 RYU 6.243 AIN 5.310
BOR 9.286** HK 6.525 SUM 6.553 HK 6.163
HAI 9.467 VTN 6.694 HAI 6.657 SUM 6.372
Ainu Ryukyu Shanghai Nanjing
KYU 5.310 VTN 4.245 NAJ 1.984** SHA 1.984**
TOH 6.003 KYU 4.736 CHD 3.815 CHD 2.565*
RYU 6.256 KOR 5.219 HK 3.878 MAN 3.264
KAN 6.309 HAI 5.219 KOR 5.853 HK 4.006
SUM 6.764 TAI 5.837 MAN 5.875 KOR 4.347
LSN 8.449 LSN 6.016 SUM 6.228 TOH 6.115
VTN 9.535 SUM 6.242 VTN 6.811 VTN 6.593
KOR 9.790 TOH 6.243 THI 7.114 HAI 6.778
BOR 11.048 AIN 6.256 KAN 7.241 SUM 6.953
ATY 11.395 THI 6.513 HAI 7.503 KYU 7.019

76 PIETRuSEWSky
51
Table 7. The smallest Mahalanobis’ distances for 56 male cranial groups using 27 measurements. Alldistances are significant p # 0.01 unless indicated otherwise (continued)
Chengdu Hong Kong Taiwan Hainan Is.
NAJ 1.984* SHA 3.878 HAI 3.189* KOR 2.396**
SHA 3.815 NAJ 4.006 KOR 4.047* TAI 3.189*
MAN 5.667 VTN 5.670 RYU 5.837 BAC 3.691
KOR 7.102 MAN 5.761 VTN 6.023 KYU 4.858
HK 7.742* KOR 6.073 MAN 6.247 VTN 5.135
VTN 8.131 KYU 6.163 KYU 6.454 THI 5.412
TAI 8.916 THI 6.213 TOH 7.076 RYU 5.583
HAI 9.203 HAI 6.279 ATY 7.275 MAN 5.772
TOH 9.421 KAN 6.525 NAJ 7.559 HK 6.279
MOG 9.680 BAC 6.887 THI 7.944 ATY 6.296
Manchuria Korea Mongolia Atayal
NAJ 3.264 HAI 2.396** SHA 9.438 KYU 5.056
KOR 3.610* KYU 2.811** NAJ 11.209 TOH 5.363
TOH 4.606 TOH 3.182** SUM 13.331 KOR 5.361*
KYU 5.146 MAN 3.610* KOR 14.839 HAI 6.296
CHD 5.667 TAI 4.047* SLW 15.071 VTN 6.799
HK 5.761 KAN 4.157* MAN 15.236 LSN 6.985
HAI 5.772 NAJ 4.343 RYU 15.852 RYU 7.207
SHA 5.875 VTN 5.068 VTN 16.459 SUM 7.227
TAI 6.247 RYU 5.219 THI 16.889 KAN 7.269
KAN 6.445 ATY 5.631* BUR 17.328 TAI 7.275
1 Where the quantity (ni nj/ni + nj) D
2ij is distributed as chi-square with p degrees of freedom (ni = sample
size of group i; nj = sample size of group j); D2
ij = square of the generalized distance between groups i and j,
and p = number of variables.
* distances significant at 5% level, ** distances not significant at 1% or 5% levels.

77CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
Figure 1. Map showing the approximate locations of the fifty-six male cranial series used in this analysis.

78 PIETRuSEWSky
Figure 2. Plot of 56 group means on the first two canonical variates using 27 cranial measurements. [See Table 1 for explanation of group abbreviations]
Figure 3. Plot of 56 group means on the first three canonical variates using 27 cranial measurements. [See Table 1 for explanation of group abbreviations]

79CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
Figure 4. Plot of 45 of the 56 group means on the first three canonical variates using 27 cranial measurements. [See Table 1 for explanation of group abbreviations]

80 PIETRuSEWSky
Figu
re 5
. Dia
gram
of r
elat
ions
hip
(den
drog
ram
) bas
ed o
n a
clus
ter
anal
ysis
(UPG
MA)
of M
ahal
anob
is’ g
ener
aliz
ed d
ista
nces
usi
ng 2
7 cr
ania
l mea
sure
men
ts
reco
rded
in 5
6 m
ale
grou
ps.

81CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
Conclusions
The results of this new multivariate analysis of craniometric data help explicate the biological relationships of the modern and near modern inhabitants of Southeast Asia and surrounding regions. The main points of this new craniometric analysis are presented here: • Inhabitants of Australia, Tasmania, and Melanesia sharply contrast with the peoples of East Asia, Southeast Asia and Remote Oceania, a division that implies separate origins for the indigenous inhabitants of these two regions. • The marked separation of East/North Asian and Southeast Asian cranial series supports models that posit long term continuity rather than population intrusion and/or replace-ment.• Biological connections between Island and Mainland Southeast Asian cranial series are clearly evident in these results.• The inhabitants of Remote Oceania, while part of the greater Asian division show con-nections with several Island Southeast Asian series supporting archaeological, linguistic, and genetic models for the peopling of Remote Oceania.• Within Island Southeast Asia, including modern-day Indonesia, a west to east pattern-ing within the island chain is apparent. The cranial series from the Southern Moluccas reveals biological affinity with adjacent Melanesian as well as Polynesian series further to the east.• There is no support for an Ainu-Pacific or Atayal-Pacific connection in these results.• Finally, understanding the craniometric variability of the modern humans of the region will provide important context for evaluating the hotly debated LB1 hominin from Liang Bua Cave on Flores Island, Indonesia.
AcKnowledgments - My thanks to Rona Ikehara-Quebral for assistance in various stages of data analy-sis, and to Rhea Hood, Atsuko Dillon, and karen kadohiro for assistance with the construction of the tables and manuscript preparation. Dr. Michele Toomay Douglas gave helpful advice and comments on earlier drafts of this paper. Ms. Billie Ikeda is responsible for the figures. An earlier version of this paper was given during the International Seminar on Southeast Asia Paleoanthropology in yogyakarta, Indonesia, July 23 – 29, 2007, organized by the late Professor Teuku Jacob.

82 PIETRuSEWSky
References
Bayard, D.T. 1996. Linguistics, archaeologists, and Austronesian origins: comparative and sociolin-guistic aspects of the Meacham-Bellwood debate. Bulletin of the Indo-Pacific Prehistory Associa-tion 15:71-85.
Bellwood, P. 1993. Cultural and biological differentiation in Peninsular Malaysia: the last 10,000 years. Asian Perspectives 32: 37-60.
Bellwood, P. 1996. Early agriculture and the dispersal of the southern Mongoloids. In T. Akazawa and E.J.E. Szathmáry, eds. Prehistoric Mongoloid dispersals. Oxford: Oxford university Press. p. 287-302.
Bellwood, P. 1997. Prehistory of the Indo-Malaysian Archipelago. Revised edition. Honolulu: univer-sity of Hawaii Press.
Bellwood, P. 2001. Early agriculturalist population diasporas? Farming, languages, and genes. Annual Review of Anthropology 30:381-207.
Bellwood, P. 2005. First farmers: the origins of agricultural societies. Oxford/Malden (MA): Black-well.
Bellwood, P. and Glover, I. 2004. Southeast Asia. Foundations for an archaeological history. In: I. Glover and P. Bellwood, eds. Southeast Asia: From prehistory to history. London: RoutledgeCurzon. P. 4-20.
Blust, R. 1996. Beyond the Austronesian homeland: the Austric hypothesis and its implications for ar-chaeology. In W. Goodenough, ed. Prehistoric settlement of the Pacific. Philadelphia: Transactions of the American Philosophical Society. p. 117-140.
Brace, C. L., Brace, M.C., Dodo, y., Leonard, W.R., yongyi, L., Sangvichien, S., Xiangqing, S., and Zhang, Z. 1990. Micronesians, Asians, Thais and relations: a craniofacial and odontometric perspec-tive. Micronesica Supplement 2:247-269.
Brace, C. L. and Tracer, D.C. 1992. Craniofacial continuity and change: a comparison of late Pleisto-cene and recent Europe and Asia. In T. Akazawa, k. Aoki, and T. kimura, eds. The evolution and dispersal of modern humans in Asia. Tokyo: Hokusen-Sha. p. 439-471.
Buikstra, J.E., Frankenberg, S.R., and konigsberg, L.W. 1990. Skeletal biological distance studies in American physical anthropology: recent trends. American Journal of Physical Anthropology 82:1-7.
Bulbeck, D. 1982. A re-evaluation of possible evolutionary processes in Southeast Asia since the late Pleistocene. Bulletin of the Indo-Pacific Prehistory Association 3:1-21.
Buranarugsa, M. and Leach, D.P. 1993. Coordinate geometry of Moriori crania and comparisons with Maori. Man and Culture in Oceania 9:1-43.
Diamond, J. and Bellwood, P. 2003. Farmers and their languages: the first expansions. Science 300:597-603.
Dixon, W.J., ed. 1992. BMDP. Statistical software manual, vol. 1. Berkeley: university of California Press.
Dixon, W.J. and Brown, M.B., eds. 1979. BMDP-79. Biomedical computer programs p-series. Berke-ley: university of California Press.
Droessler, J.B. 1981. Craniometry and biological distance: biocultural continuity and change at the late-Woodland-Mississippian interface. Center for American archeology research series, vol. 1. Evanston: Center for American Archeology.
Glover, I.C. and Higham, C.F.W. 1996. New evidence for early rice cultivation in South, Southeast, and East Asia. In D.R. Harris, ed. The origins and spread of agriculture and pastoralism in Eurasia. Washington, D. C: Smithsonian Institution Press. p. 413-441.

83CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
Green, R.C. 1979. Lapita. In J. Jennings, ed. The prehistory of Polynesia. Cambridge, Massachusetts: Harvard university Press. p. 27-60.
Green, R.C. 1991a. Near and Remote Oceania -disestablishing “Melanesia” in culture history. In A. Pawley, ed. Man and a half. Essays in Pacific anthropology and ethnobiology in honour of Ralph Bulmer. Auckland: The Polynesian Society. p.491-502.
Green, R.C. 1991b. The Lapita Cultural Complex: current evidence and proposed models. In P. Bell-wood, ed. Indo-Pacific prehistory 1990, volume 2. Canberra and Jakarta: Bulletin of the Indo-Pacific Prehistory Association. p. 295-305.
Hagelberg, E. 1998. Genetic perspectives on the settlement of the Pacific. In C.M. Steveson, G. Lee, and F.J. Morin, eds. Easter Island in Pacific context: South Seas symposium. Proceedings of the Fourth International Conference on Easter Island and East Polynesia, Los Osos: The Easter Island Foundation Occasional Paper 4.
Hanihara, T. 1993. Population prehistory of East Asia and the Pacific as viewed from craniofacial morphology: the basic populations in East Asia, VII. American Journal of Physical Anthropology 91:173-187.
Hanihara, T. 1994. Craniofacial continuity and discontinuity of Far Easterners in the late Pleistocene and Holocene. Journal of Human Evolution 27: 417-441.
Higham, C.F.W. 1996. The bronze age of Southeast Asia. Cambridge: Cambridge university Press. Higham, C.F.W. 2001. Prehistory, language and human biology: is there a consensus in East and South-
east Asia? In L. Jin, M. Seielstad, and C. Xiao, eds. Genetic, linguistic and archaeological perspec-tives on human diversity in Southeast Asia. New Jersey: World Scientific. p 3-16.
Howells, W.W. 1973. Cranial variation in man. Papers of the Peabody Museum of Archaeology and Ethnology, vol. 67. Cambridge, Massachusetts: Harvard university.
Howells, W.W. 1989. Skull shapes and the map: craniometric analyses in the dispersion of modern Homo. Papers of the Peabody Museum of Archaeology and Ethnology, volume 79. Cambridge, Mas-sachusetts: Harvard university.
Howells, W.W. 1995. Who’s who in skulls: ethnic identification of crania from measurements. Papers of the Peabody Museum of Archaeology and Ethnology, vol. 82. Cambridge: Harvard university Press.
Irwin, G. 1992. The prehistoric exploration and colonisation of the Pacific. Cambridge: Cambridge university Press.
kayser, M.S., Brauer, S.,Weiss, G., underhill, P.A., Roewer, L., Schiefenhövel, W., and Stoneking, M. 2000. Melanesian origin of Polynesian y chromosomes. Current Biology 10:1237-1246.
kirch, P.V. 1997. The Lapita peoples: ancestors of the Oceanic world. Oxford: Blackwell Scientific. koganei, y. (1893-94) Beiträge zur physischen Anthropologie der Aino. Mittheilungen aus der Medici-
nischen Facultät der Kaiserlich-Japanischen Universität zu Tokyo 2:1-249, 251-404.kohn, L.A. 1991. The role of genetics in craniofacial morphology and growth. Annual Review of An-
thropology 20: 261-278.konigsberg, L.W. and Ousley, S.D. 1995. Multivariate quantitative genetics of anthropometric traits
from the Boas data. Human Biology 67:481-498.Larsen, C.S. 1997. Bioarchaeology. Interpreting behavior from the human skeleton. Cambridge: Cam-
bridge university Press.Lum, J.k. and Cann, R.L. 2000. mtDNA lineage analyses: origins and migrations of Micronesians and
Polynesians. American Journal of Physical Anthropology 113:151–168.Mahalanobis, P.C. 1936. On the generalized distance in statistics. Proceedings of the National Institute
of Sciences, Calcutta 2:49-55.

84 PIETRuSEWSky
Mahalanobis, P.C., Majumdar, D.N., and Rao, C.R. 1949. Anthropometric survey of the united Prov-inces, 1941: a statistical study. Sankhya 9: 89-324.
Martin, R. and Saller, k. 1957. Lehrbuch der anthropologie. Stuttgart: Gustav Fischer Verlag.Matsumura, H. 1994. A microevolutional history of the Japanese people from a dental characteristics
perspective. Anthropological Science 102:93–118.Matsumura, H. 1995. Dental characteristics affinities of the prehistoric to modern Japanese with the
East Asians, American natives and Australo-Melanesians. Anthropological Science 103:235-261.Matsumura, H. 2001. Differentials of yayoi immigration to Japan as derived from dental metrics. Homo
52:135-156.Matsumura, H. 2006. The population history of Southeast Asia viewed from morphometric analyses of
human skeletal and dental remains. In M.R. Oxenham and N. Tayles, eds. Bioarchaeology of South-east Asia. Cambridge: Cambridge university Press. p.33-58.
Matsumura, H. and Hudson, M. 2005. Dental perspectives on population history of Southeast Asia. American Journal of Physical Anthropology 127: 182-209.
Matsumura, H. Cuone, N.L., Thuy, N. k., and Anezaki, T. 2001. Dental morphology of the early Hoabinhian, the Neolithic Da But and the Metal Age Dong Son cultural people in Vietnam. Zeitshrift für Morphologie und Anthropologie 83: 59-73.
Melton, T., Peterson, R., Redd, A.J., Saha, N., Sofro, A.S.M., Martinson, J., and Stoneking, M. 1995. Polynesian genetic affinities with Southeast Asian populations as identified by mtDNA analysis. American Journal of Human Genetics 57:403–414.
Merriwether, D.A., Friedlaender, J.S., Mediavilla, J., Mgone, C., Gentz, F., and Ferrell, R.E. 1999. ‘Mi-tochondrial DNA variation is an indicator of Austronesian influence in Island Melanesia’. American Journal of Physical Anthropology 110(3): 243-270.
Oppenheimer, S.J. 1998. Eden in the east: the drowned continent of Southeast Asia. London: Weiden-feld and Nicolson.
Oppenheimer, S.J. and Richards, M. 2001a. Polynesian origins: slow boat to Melanesia? Nature 410:166-167.
Oppenheimer, S.J. and Richards, M. 2001b. Fast trains, slow boats, and the ancestry of the Polynesian Islanders. Science Progress 84(3):157-181.
Oppenheimer, S.J. and Richards, M. 2003. Polynesians: devolved Taiwanese rice farmers or Wallacean maritime traders with fishing, foraging and horticultural skills? In P. Bellwood and C. Renfrew, eds. Examining the farming/language dispersal hypothesis. Cambridge: McDonald Institute for Ar-chaeological Research. p. 287–297.
Pietrusewsky, M. 1990a. Craniofacial variation in Australasian and Pacific populations. American Jour-nal of Physical Anthropology 82:319-340.
Pietrusewsky, M. 1990b. Cranial variation in New Guinea and neighboring populations of the Pa-cific: a multivariate study of specimens in the Museum für Völkerkunde Dresden and the German Democratic Republic. Abhandlungen und Berichte des Staatlichen Museums für Völkerkunde 45: 233-257
Pietrusewsky, M. 1994. Pacific-Asian relationships: a physical anthropological perspective. Oceanic Linguistics 33:407-430.
Pietrusewsky, M. 1997. The people of Ban Chiang: An early bronze-age site in northeast Thailand. Bul-letin of the Indo-Pacific Prehistory Association 16:119-148.
Pietrusewsky, M. 1999. A multivariate craniometric investigation of the inhabitants of the Ryukyu Islands and comparisons with cranial series from Japan, Asia, and the Pacific. Anthropological Sci-ence 107:255-281.

85CRANIOMETRIC VARIATION IN SOuTHEAST ASIA
Pietrusewsky, M. 2000. Metric analysis of skeletal remains: methods and applications. In M.A. katzen-berg and S.R. Saunders, eds. Biological anthropology of the human skeleton. New york: Wiley-Liss. p. 375-415.
Pietrusewsky, M. 2004. Multivariate comparisons of female cranial series from the Ryukyu Islands and Japan. Anthropological Science 112:199-211.
Pietrusewsky, M. 2005. The physical anthropology of the Pacific, East Asia, and Southeast Asia: a mul-tivariate craniometric analysis. In L. Sagart, R. Blench, and A. Sanchez-Mazas, eds. The peopling of East Asia: putting together archaeology, linguistics, and genetics. Jakarta, Indonesia: Indonesian Institute of Sciences, LIPI Press. London: Routledge Curzon. p. 201-229.
Pietrusewsky, M. 2006a. Chapter 3. A multivariate craniometric study of the prehistoric and modern inhabitants of Southeast Asia, East Asia, and surrounding regions: a human kaleidoscope? In M.R. Oxenham and N. Tayles, eds. Bioarchaeology of Southeast Asia. Cambridge: Cambridge university Press. p. 59-90.
Pietrusewsky, M. 2006b. The initial settlement of remote Oceania: The evidence from physical an-thropology. In T. Simanjuntak, I.H.E. Pojoh, and M. Hisyam, eds. Austronesian diaspora and the ethnogenesis of people in Indonesian Archipelago. Proceedings of the International Symposium. p. 320-347.
Pietrusewsky, M. and Douglas, M.T. 2002. Ban Chiang, a prehistoric site in Northeast Thailand I: the human skeletal remains. University monograph 111. Philadelphia: university of Pennsylvania Mu-seum of Archaeology and Anthropology.
Pope, G.G. 1992. Replacement versus regionally continuous models: the paleobehavioral and fossil evidence from East Asia. In T. Akazawa, k. Aoki, and T. kimura, eds. The evolution and dispersal of modern humans in Asia. Tokyo: Hokusen-sha. p. 3-14.
Rao, C.R. 1948. The utilization of multiple measurements in problems of biological classification. Journal of the Royal Statistical Society B 10:159-193.
Rao, C.R. 1952. Advanced statistical methods in biomedical research. New york: John Wiley.Redd, A.J., Takezaki, N., Sherry, S.T., McGarvey, S.T., Sofro, A.S.M. and Stoneking, M. 1995. Evolu-
tionary history of the COII/tRNALys intergenic 9-base-pair deletion in human mitochondrial DNAs from the Pacific. Molecular Biology and Evolution 12:604 615.
Richards, M., Oppenheimer, S., and Sykes, B. 1998. mtDNA suggests Polynesian origins in eastern Indonesia. American Journal of Human Genetics 63:1234-1236.
Rohlf, F.J. 1993. NTSyS-pc. Numerical taxonomy and multivariate analysis system, version 1.80. Se-tauket, New york: Exeter Software.
Sjøvold, T. 1984. A report on the heritability of some cranial measurements and non-metric traits. In G.N. Van Vark and W.W. Howells, eds. Multivariate statistics in physical anthropology. Dordrecht: D. Reidel. p. 223-246.
Sneath, P.H.A and Sokal, R.R. 1973. Numerical taxonomy. San Francisco: Freeman.Speiser, F. 1928. Anthropologische messungen aus dem St. Cruz-Inseln. Archiv für Anthropologie 19:
89-146.Su, B., Jin, L., underhill, P., Marthinson, J., Saha, N. McGarvey, S.T., Shriver, M.D., Chui, J., Oefner,
P., Chakraborty, R., and Deka, R. 2000. ‘Polynesian origins: insights from the y chromosome’. Proceedings of the National Academy of Sciences (U.S.A.) 96(15): 8225-8228.
Tatsuoka, M.M. 1971. Multivariate analysis: techniques for educational and psychological research. New york: John Wiley and Sons.
Turner C.G.II. 1987. Late Pleistocene and Holocene population history of East Asia based on dental variation. American Journal of Physical Anthropology 73:305-321.

86 PIETRuSEWSky
Turner C.G.II. 1989. Teeth and prehistory in Asia. Scientific American 262:88-96.Turner, C.G.II. 1990. Major features of sundadonty and sinodonty including suggestions about East
Asian microevolution, population history, and late Pleistocene relationships with Australian Ab-origines. American Journal of Physical Anthropology 82:295-317.
Turner, C.G.II. 1992a. Sundadonty and sinodonty in Japan: the dental basis for a dual origin hypothesis for the peopling of the Japanese Islands. In k. Hanihara, ed. Japanese as a member of the Asian and Pacific populations. kyoto: International Research Center for Japanese Studies. p. 96-112.
Turner, C.G.II. 1992b. Microevolution of East Asian and European populations: a dental perspective. In T. Akazawa, k. Aoki, and T. kimura, eds. The evolution and dispersal of modern humans in Asia. Tokyo: Honkusen-Sha. p. 415-438.
Van Vark, G.N. and Howells, W.W. eds. 1984. Multivariate statistics in physical anthropology. Dor-drecht: D. Reidel.
Van Vark, G.N., and Schaafsma, W. 1992. Advances in the quantitative analysis of skeletal morphology. In M.A. katzenberg and S.R. Saunders, eds. Skeletal biology of past peoples: research methods. New york: Wiley-Liss.
Wilkinson L. 1992. Systat for windows, version 5. Evanston: Systat, Inc.