Embed Size (px)
Citation preview

1
����
����
Phytochemical and Biosynthetic Studies of Lignans
����
with a Focus on Indonesian Medicinal Plants

2

3
RIJKSUNIVERSITEIT GRONINGEN
Phytochemical and Biosynthetic Studies of Lignans with a Focus on Indonesian Medicinal Plants
Proefschrift
ter verkrijging van het doctoraat in de Wiskunde en Natuurwetenschappen aan de Rijksuniversiteit Groningen
op gezag van de Rector Magnificus, dr. F. Zwarts, in het openbaar te verdedigen op
maandag 26 juni 2006 om 14.45 uur
door
Elfahmi
geboren op 25 april 1969 te Padang, Indonesië

4
Promotor : Prof. Dr. W.J. Quax Copromotores : Dr. H.J. Woerdenbag Dr. O. Kayser Beoordelingscommissie: Prof. Dr. F.A.J. Muskiet Prof. Dr. T.J. Schmidt Prof. Dr. R. Verpoorte

5
Paranimfen: M.K. Julsing M. Chalid
To my parents, M. Yaman and Emma
my lovely wife, Yulia Helmi my kids, Fathiya Mufidah, Nurul Rahimah and Ahmad Muzakki Imanullah

6
This research project was financially supported by the QUE Project Batch II, Department of Biology, Institut Teknologi Bandung ITB, Indonesia, under contract No. 3028-IX/P3S-1/KON-QUE II/2000; IBRD Loan No. 4193-IND, and partially by a scholarship from Universiy of Groningen, the Netherlands. The Graduate School for Drug Exploration (GUIDE) is gratefully acknowledged for supporting part of the printing costs of the thesis. Printing: Facilitair Bedrijf, University of Groningen, the Netherlands © 2006 by Elfahmi. All rights reserved. No part of this book may be reproduced or transmitted in any forms or by any means without permission from the author. ISBN: 90-902-0771-6

7
Contents
Chapter 1 Aims and scope of the thesis 9
Chapter 2 Jamu: The Indonesian traditional herbal medicine 13
Chapter 3 Lignans from cell suspension cultures of Phyllanthus niruri, an Indonesian medicinal plant 35
Chapter 4 Lignan profile of Piper cubeba, an Indonesian medicinal plant 45
Chapter 5 Essential oil constituents of Piper cubeba from Indonesia 53
Chapter 6 Reduced coniferin and enhanced 6-methoxypodophyllotoxin production in Linum flavum cell suspension cultures after treatment with Na2EDTA 61
Chapter 7 Cloning of glucosyltransferase genes from cell suspension cultures of Linum flavum in E. coli 71
Chapter 8 Production of a cytotoxic arylnaphthalene lignan in genetically transformed root cultures of Linum leonii 83
Summary and concluding remarks 93
Samenvatting (Dutch) 97
Ringkasan (Indonesian) 101
References 105
Curriculum vitae 123
Acknowledgements 125

8

9
Chapter 1
Aims and scope of the thesis

���������
10
Aims and scope of the thesis The use of plants, plant extracts and plant-derived chemicals in the treatment of diseases, in
supplementing foods and in making cosmetics is firmly rooted in the past and still developing. Many drugs used in contemporary medicine have been derived from plants and were originally discovered through the traditional use by indigenous people. Podophyllotoxin, vincristine, vinblastin, camptothecin, taxol, artemisinin, aspirin, atropine, ephedrine, quinine, reserpin and digoxin are well known examples of such drugs.
Progress in science and technology boosts the further development of medicinal plants as valuable sources of drugs and drug leads. Modern analytical methods, biotechnology approaches, genomics, proteomics and metabolomics are nowadays applied in medicinal plant research and contribute to the advancement of the field. Quite frequently a relatively low yield of active components and difficulties in standardization are bottlenecks in medicinal plant exploitation. Efforts have been made worldwide to enhance the production of bioactive component using a biotechnology approach. In this thesis we aim to study phytochemical and biosynthetic aspects of lignans in selected medicinal plants used in Indonesia and in European Linum species. In chapter 2, we review jamu, the traditional Indonesian medicine based on the popular use of medicinal plants. Indonesia is one of the biggest biological diversities in the word. Indonesian people have been using jamu to treat various diseases since long. An introduction to jamu, its present status in Indonesia, legislative and regulatory aspects, economical perspective and rational therapy with jamu are highlighted as well as ethical considerations that should be taken into account in the development of medicinal plants. This review aims to give comprehensive information about the biological activity and therapeutic value of the most commonly used medicinal plants (and plant constituents) in jamu. This knowledge can be used to further develop jamu in Indonesia in a rational way.
In the chapter 3 we studied lignans from cell suspension cultures of Phyllanthus niruri, an important medicinal plant in Indonesia. Feeding experiments using the biosynthetic precursors of lignans, caffeic acid and ferulic acid, were carried out in order to enhance the lignan production and to study their biosynthetic pathway. A phytochemical study of another important medicinal plant in Indonesia, Piper cubeba, was conducted as well. A systematic profiling of lignans was made using TLC, HPLC, GC and GC-MS of different parts of this plant (chapter 4). So far, only fruits from P. cubeba are used medicinally and as a food supplement. To complete the phytochemical study we also analyzed the essential oil composition of P. cubeba using GC and GC-MS (chapter 5). Lignans and the essential oil are the main classes of secondary metabolites of P. cubeba.
The discovery of podophyllotoxin, the starting compound for the semisynthetic anticancer drugs etoposide, teniposide and etophopos stimulates the research of lignans including derivatives of podophyllotoxin and its biosynthetic intermediates. In chapter 6 we present the enhancement of the production of 6-methoxypodophyllotoxin in cell suspension cultures of Linum flavum using ethylenediamine tetra acetate (EDTA) and other glucosyltransferase inhibitors. The enzyme glucosyltransferase converts coniferyl alcohol to coniferin that accumulates in the cells in high amounts. Coniferyl alcohol is a biosynthetic precursor of 6-methoxypodophyllotoxin and other lignans. We carried out feeding experiment in order to inhibit the coniferin formation and to enhance the 6-methoxypodophyllotoxin production. The lignan biosynthesis in cell suspension cultures of L. flavum was also investigated at a genetic level. We tried to clone the glucosyltransferase in Escherichia coli and to study the enzyme further in order to better understand the biosynthesis pathway of lignans (chapter 7). The ultimate goal is to develop a tool to enhance the production of cytotoxic lignans.
The production of a cytotoxic lignan, justicidin B, using hairy roots cultures of genetically modified Linum leonii (transformation by Agrobacterium rhizogenes) is described in chapter 8. Justicidin B is an arylnaphthalene lignan which exerts cytotoxic, antiviral, fungicidal, antiprotozoal and

� ��� ���� ����� ��������������� �
11
antiplatelet properties. In addition to the production we also study the cytotoxic activity of justicidin B against three chronic myeloid leukemia-derived LAMA-84, K-562 and SKW human cell lines.
This thesis covers a broad range of lignan studies including phytochemistry, biotechnology and genetic engineering in order to better understand and to enhance the production of lignans. Subjects of our study were two selected Indonesian plants used in jamu, and two European Linum species that contain cytotoxic lignans.

���������
12

13
Chapter 2
Jamu: The Indonesian traditional herbal medicine Elfahmi, Komar Ruslan, Rein Bos, Oliver Kayser, Herman J. Woerdenbag, Wim J. Quax Submitted

����������
14
Abstract Jamu is the Indonesian traditional herbal medicine that has been practiced for many centuries in the Indonesian community to maintain good health and to treat diseases. Although modern (western) medicine is becoming increasingly important in Indonesia, jamu is still very popular in rural as well as in urban areas. Based on its traditional use jamu is being developed to a rational form of therapy, from herbal practitioners to drugs in pharma industries. Jamu has acquired a potential benefit, both economically and clinically. We survey the most frequently used plants in jamu that in addition have been investigated as to their constituents and pharmacological effects. Many isolated compounds have potent biological activities. Examples are: curcumin (Curcuma longa) as anticancer, antihipertensive, antidiabetes and immunostimulating agent; andrographolide (Andrographis paniculata) as anticancer, antiviral and cardioprotective agent; 1'-acetoxychavicol (Alpinia galanga) as anticancer, antimicrobial, antifungal and gastroprotective agent; lignans (Phyllanthus niruri) as antiviral and hepatoprotective agent. The Indonesian government has divided the preparation of medicinal plants into three categories, i.e. jamu, standardized herbal medicines and fitofarmaka. As the biological activity ascribed to jamu is largely based on empirical data, more research is needed to scientifically prove efficacy and to assure safety. In the further development of jamu, ethical issues such as intellectual property right, benefit sharing, biodiversity and conservation should be considered. This paper aims to review the state-of-art of jamu and to give comprehensive views that can be used for the further improvement of the utility of jamu in curing illnesses and maintaining good health.

� � ���������������� �������������������������������
15
Introduction Following the Amazon rain forests, Indonesia has the second biggest biodiversity in the world
expressed by a high number of indigenous medicinal plants. Based on this rich source the use of medicinal plants is very important, and in the rural areas medicinal plants are even the first choice to treat diseases. Most of the Indonesian people have ever used traditional herbal medicines which are popularly known as jamu. Jamu is a word in Javanese tribe language, meaning the traditional medicine from plants. Today, jamu has been adopted into Bahasa Indonesia with the similar meaning (Riswan and Roemantyo, 2002). Nowadays jamu is being developed from traditional handling to industrial (larger scale) production. Worldwide however, jamu is less known than, e.g., Traditional Chinese Medicine (TCM), Japanese Kampo and Indian Ayuverda. Jamu gendong is a kind of traditional jamu sold without label, and freshly prepared (not preserved) from plant material in warung, the ubiquitous stalls along the streets in Indonesia (Limyati and Juniar, 1998, Suharmiati, 2003). Jamu gendong is instantly served to whom orders this jamu. The sellers must bring the jamu from door to door. The word gendong itself means to carry something on the back of a body. The fresh jamu is put inside each bottle in bamboo or rattan basket. And they use a long wide shawl called selendang for carrying the basket on the back (Risman and Roemantyo, 2002).
A scientific approach is essential to further develop the rational use of jamu. The Indonesian government, industry and academia put considerable efforts on it. Various groups of secondary metabolites are known to be active components in jamu, including alkaloids, flavonoids, steroids, terpenoids, coumarins, and lignans. They contribute to the therapeutic effect as single active compounds as well as in combination with others.
This article reviews the use of Indonesian medicinal plants in jamu including its history, current status, economical prospective, development, scientific approach, and summarizes the potential developments in the future. Both online and offline literature searches have been done to compile this review. Pubmed (Medline) and ISI Web of Science were used to retrieve any online publications. About 5,000 species of medicinal plants have been retrieved from the Medicinal Herbs Index in Indonesia and the plants that are most frequently used as constituents of jamu are discussed in this paper. Indonesian medicinal plants Biodiversity
Biodiversity is defined as the variety of all life forms on earth, along with the interactions between them and their physical environment. As an archipelagic state with thousands of islands, Indonesia is endowed with a rich and unique biodiversity. The area of Indonesian tropical forests covers about 143 million hectares and is inhabited by about 80% of the world’s medicinal plants. It is estimated that the Indonesian tropical forests inhabit 28,000 plant species. There are various reports concerning the inventory of higher plant in Indonesia. The Indonesian Country Study on Biodiversity (ICSBD 1993) puts the number of flowering plants species in Indonesia between 25,000 and 30,000. Some 40 million Indonesians depend directly on the country’s biodiversity, and the Indonesian community makes use of around 6,000 plant species. Data of the number of medicinal plants also vary. PT Eisei (1995) published the Dictionary of Indonesian Medicinal Herbs containing more than 2,500 plants species which potentially, while Zuhud et al. (2001) identified 1,845 species with medicinal potential in the forests of Indonesia. These numbers are potentially to be updated due to the continuing inventory and investigation of yet unidentified species. According to the National Agency of Drug and Food Control (NADFC/BPOM), 283 plant species have been officially registered for their medicinal use; the larger remaining part is used traditionally.

����������
16
To facilitate the activities on the conservation and sustainable use of biodiversity, the Indonesian Government, through the National Development Planning Agency (BAPPENAS), has launched the Indonesian Biodiversity Strategy and Action Plan 2003-2020 (IBSAP). IBSAP is based on the evaluation of the previous action plan from 1993 called BAPI (Biodiversity Action Plan for Indonesia), formulated in collaboration between the Indonesian Government (BAPPENAS), the Ministry of Environment, research institutes and non-governmental stakeholders with the support of the international developments institutions. Recent research development and research communities
Evaluation of jamu as a rational phytotherapy has to cover different research topics including social, cultural, economic, and ethical aspects. Phytochemical studies including extraction, isolation and characterization of secondary plant metabolites have been developed to date. Biological activity studies have been conducted in vitro and in vivo, and even a few clinical studies are available. To coordinate and to conduct research directed to the development of medicinal plants, many institutions in Indonesia are engaged. They comprise governmental institutions such as the Ministry of Health, Ministry of Forestry, Ministry of Environment, Ministry of Agriculture, the National Development Planning Agency (BAPPENAS), the National Agency of Drug and Food Control (NADFC or BPOM). The universities are actively involved through the related faculties or departments from different areas like medicine, pharmacy, chemistry, biology, agriculture, forestry, marine, environment and engineering. National research institutions such as the Indonesian Institute of Science and the Herbarium Bogoriense, are involved as well as non-governmental institutions such as KEHATI (Indonesian Biodiversity Foundation�, WALHI, SKEPHI, and various industrial companies (Bermawie et al., 2005). Economical prospective
Since the 1980s, small size jamu producers have grown sufficiently to introduce larger scale and modern production methods (Beer, 2001). The jamu producing industry now has an annual growth of 25-30%. According to Pramono (2002) there are about 810 companies active in Indonesian traditional medicine of which 87 are classified as IOT (Industri Obat Tradisional, Traditional Medicine Industry) and 723 as IKOT (Industri Kecil Obat Tradisional, Small Industry of Traditional Medicine). In 2005, 872 companies in this field have been registered at BPOM. In addition, 462 companies from foreign countries also play a role in the production of Indonesian traditional medicine. About 20 local companies are the major players. The examples of jamu products from these companies are shown in Table 1. The industry revenues in 2000 was estimated to be 150 million USD. However, taking into account the possibilities on the international market and the richness of the country regarding its natural resources, this amount can potentially be increased (Pramono, 2002). In the period between January to June 2005, the export of medicinal plants such as Amomum cardamomum, Cinnamomum burmani, Piper spp. and many others used to make jamu reached an amount of 126.8 million USD (Ministry of Industry, Republic of Indonesia). Ethical considerations in the development of medicinal plants
Intellectual property right (IPR), indigenous knowledge, benefit sharing, efficacy and safety are issues that must be considered in the further development of Indonesian medicinal plants. Jamu has been handed over from generation to generation based on the traditional knowledge and experience of the community. When new plant-derived therapeutics based on indigenous knowledge are being explored, it is important that the companies return benefits to the native population and the local governments from which the research material was obtained (King et al., 1996). When individuals or institutions from

� � ���������������� �������������������������������
17
biotechnologically developed countries wish to obtain indigenous raw material from a biotechnologically less developed country, an agreement for the procurement of such material may be negotiated. In article 19.2 of the Rio Convention (1992) there is an agreement about the handling of biotechnology and distribution of its benefits. It is mentioned that each contracting party shall take all practicable measures to promote and advance priority access on a fair and equitable basis by contracting parties, especially developing countries, to the results and benefits arising from biotechnologies based upon genetic resources provided by those contracting parties. Such access shall be on mutually agreed terms. Goodwill to maintain such a flow may be achieved through appropriate scientific and monetary compensation, both in real time and in long-term sharing of the benefits of discovery (Soejarto, 1996). Harvesting too much and/or cultivating too little may render medicinal plants into endangered species. It is important to take into account that the individuals or institutions exploring the medicinal plant material also have responsibilities for the conservation. Most of the current knowledge that jamu can maintain health and/or can cure diseases comes from the people who have experienced success in curing illness by taking jamu, it remains to be proved that jamu fulfills the generally accepted criteria of safety and efficacy in order to protect the patients. Jamu as a way of traditional healing Rational phytotherapy with jamu and phytomedicine
Jamu, as traditional medicine arising from experiences of the past and embedded in the culture of society cannot stand still but constantly changes and develops. Along with allopathic medicine it shares issues in appropriate and rational use. These include qualification and licensing of the provider, proper use of good quality products, good communication between traditional medicine providers and patients and provision of scientific information and guidance to the public (WHO, 2002). Although pharmacological effects of jamu constituents have been recorded, there is an apparent lack of records or written data reporting the effectiveness of jamu, especially of jamu gendong. To assure the proper use of such products, the Indonesian government has divided the medicinal plants in three categories based on the way they are prepared and based on their efficacy; i.e. jamu, standardized herbal medicines, and fitofarmaka (phytomedicines). All preparations have to meet basic safety criteria. The therapeutic effects of jamu have to be supported by empirical data. The efficacy of standardized herbal medicines has to be proved in pre-clinical trials and standardization on active ingredients is required. For fitofarmaka clinical trials have to be available. The Indonesian government has launched the Centre for Development and Application of Traditional Treatment (Sentra P3T) in 1995. The Centre’s activities include research, testing, education, training and service of traditional treatment. Other programmes include selecting, testing, certifying, registration/licensing, inventory, screening, clinical testing, utilization and evaluation of traditional medicine, and compilation of laws applicable to traditional treatment. Preparation of jamu
Original jamu (jamu gendong) exists in the form of a decoction and is sold by ladies carrying jamu on their back. Jamu gendong is produced by household scale industries in a simple and traditional way. Traditional jamu makers also care about hygiene, sanitation and chemical contaminations from biological or non-biological sources. They try to protect raw materials and products from contamination, although this is far from international industrial standards. The way of preparation is often different from producer to producer, and production steps like selection of raw materials, sorting, grating, scraping, crushing, mixing and cooking, followed by boiling of the plant material in a hygienic way can differ significantly. From this background professional training was necessary to introduce certain standards like standardization of the raw materials used in jamu according to the Materia Medika Indonesia (MMI). Jamu makers have to be trained on hygienic production methods and for semi-modern

����������
18
technologies. The most important aspect of the training is the introduction of scientific aspects of jamu. From household scale industries jamu has been developed and is now produced by the industries called IKOT and IOT. To prepare jamu, IKOT and IOT use the modern technologies and their activities are based on a scientific approach. They have to follow the directions for good manufacturing production (GMP). Today jamu made by the industry is not anymore only in the form of a decoction but also in the form of a tablet, pill, powder, pastille, capsule, extract, cream, and ointment. Legislative aspects of jamu and phytomedicines in Indonesia
The Indonesian government, through the Ministry of Health and BPOM, has regulated jamu and phytomedicines (fitofarmaka). The regulations are aimed to develop herbal medicinal products, to protect the people from unwanted (adverse) effects, and to watch over the quality including efficacy and efficiency. Three types of Indonesian medicinal plants that mentioned before have been regulated by BPOM through a regulation nr. HK.00.05.4.2411, 2004. For the production of traditional medicine in Indonesia, the industries have to refer to good manufacturing practice guidelines for traditional medicine, called CPOTB (Cara Pembuatan Obat Tradisional yang Baik). CPOTB is regulated by the Ministry of Health (regulation nr. 659/MENKES/SK/X/1991). This regulation has been renewed by BPOM in 2005 with regulation nr. HK.00.05.4.1380. CPOTB includes all aspects of production such as raw material, production process, quality control, factory building, workers, management, instrument, sanitation, etc. CPOTB is also to be applied in the industries to produce standardized herbal medicines and phytomedicine. The traditional medicine industry (IOT and IKOT) as well as the products have to be registered in the BPOM (246/MENKES/Per/V/90 and HK.00.05.41.1384, 2005). Using this regulation, the production and distribution of traditional medicine could be controlled to fulfill the requirements according CPOTB. There are several forms of traditional medicine such as powders, pills, capsules, crude extracts, tablets, liquids. These products have to be produced according to the description published in regulation number 661/MENKES/SK/VII/1994. To develop the traditional medicines, the Indonesian government has established the Centre for Development of Traditional Medicine (Sentra P3T). The Centre is supported by regulation number 0584/MENKES/SK/VI/1995.
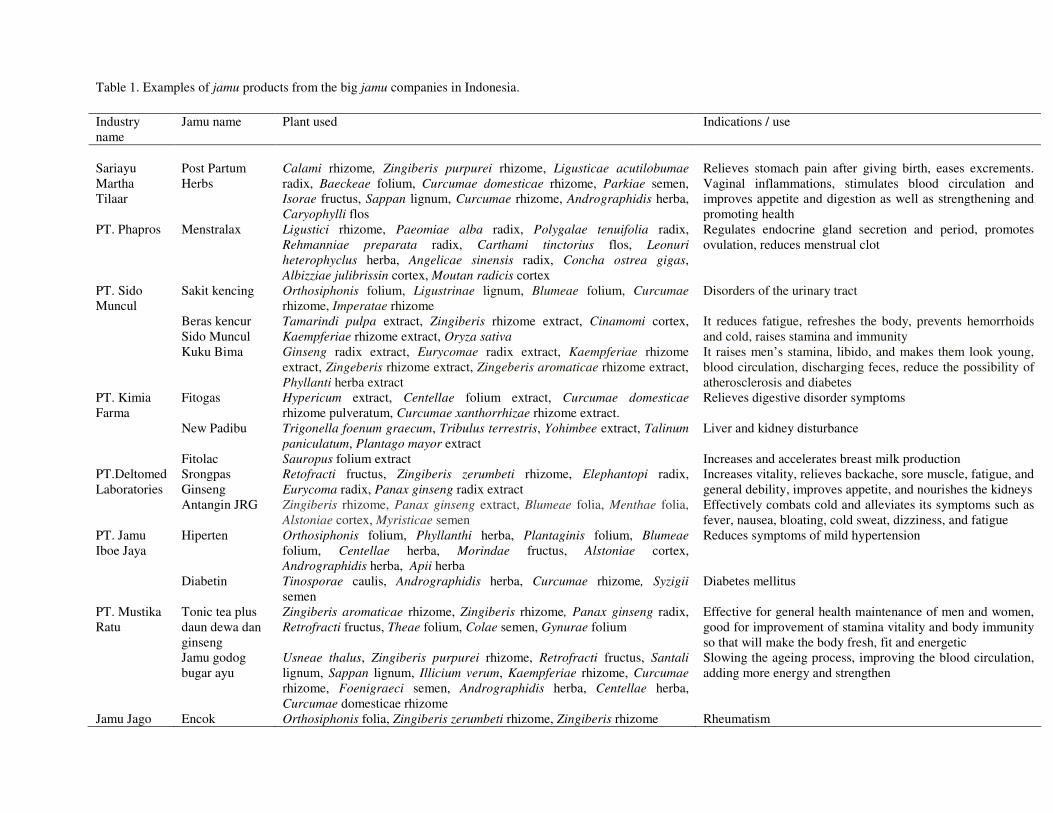
19
Table 1. Examples of jamu products from the big jamu companies in Indonesia. Industry name
Jamu name Plant used Indications / use
Sariayu Martha Tilaar
Post Partum Herbs
Calami rhizome, Zingiberis purpurei rhizome, Ligusticae acutilobumae radix, Baeckeae folium, Curcumae domesticae rhizome, Parkiae semen, Isorae fructus, Sappan lignum, Curcumae rhizome, Andrographidis herba, Caryophylli flos
Relieves stomach pain after giving birth, eases excrements. Vaginal inflammations, stimulates blood circulation and improves appetite and digestion as well as strengthening and promoting health
PT. Phapros Menstralax Ligustici rhizome, Paeomiae alba radix, Polygalae tenuifolia radix, Rehmanniae preparata radix, Carthami tinctorius flos, Leonuri heterophyclus herba, Angelicae sinensis radix, Concha ostrea gigas, Albizziae julibrissin cortex, Moutan radicis cortex
Regulates endocrine gland secretion and period, promotes ovulation, reduces menstrual clot
PT. Sido Muncul
Sakit kencing Orthosiphonis folium, Ligustrinae lignum, Blumeae folium, Curcumae rhizome, Imperatae rhizome
Disorders of the urinary tract
Beras kencur Sido Muncul
Tamarindi pulpa extract, Zingiberis rhizome extract, Cinamomi cortex, Kaempferiae rhizome extract, Oryza sativa
It reduces fatigue, refreshes the body, prevents hemorrhoids and cold, raises stamina and immunity
Kuku Bima Ginseng radix extract, Eurycomae radix extract, Kaempferiae rhizome extract, Zingeberis rhizome extract, Zingeberis aromaticae rhizome extract, Phyllanti herba extract
It raises men’s stamina, libido, and makes them look young, blood circulation, discharging feces, reduce the possibility of atherosclerosis and diabetes
PT. Kimia Farma
Fitogas Hypericum extract, Centellae folium extract, Curcumae domesticae rhizome pulveratum, Curcumae xanthorrhizae rhizome extract.
Relieves digestive disorder symptoms
New Padibu Trigonella foenum graecum, Tribulus terrestris, Yohimbee extract, Talinum paniculatum, Plantago mayor extract
Liver and kidney disturbance
Fitolac Sauropus folium extract Increases and accelerates breast milk production PT.Deltomed Laboratories
Srongpas Ginseng
Retofracti fructus, Zingiberis zerumbeti rhizome, Elephantopi radix, Eurycoma radix, Panax ginseng radix extract
Increases vitality, relieves backache, sore muscle, fatigue, and general debility, improves appetite, and nourishes the kidneys
Antangin JRG Zingiberis rhizome, Panax ginseng extract, Blumeae folia, Menthae folia, Alstoniae cortex, Myristicae semen
Effectively combats cold and alleviates its symptoms such as fever, nausea, bloating, cold sweat, dizziness, and fatigue
PT. Jamu Iboe Jaya
Hiperten Orthosiphonis folium, Phyllanthi herba, Plantaginis folium, Blumeae folium, Centellae herba, Morindae fructus, Alstoniae cortex, Andrographidis herba, Apii herba
Reduces symptoms of mild hypertension
Diabetin Tinosporae caulis, Andrographidis herba, Curcumae rhizome, Syzigii semen
Diabetes mellitus
PT. Mustika Ratu
Tonic tea plus daun dewa dan ginseng
Zingiberis aromaticae rhizome, Zingiberis rhizome, Panax ginseng radix, Retrofracti fructus, Theae folium, Colae semen, Gynurae folium
Effective for general health maintenance of men and women, good for improvement of stamina vitality and body immunity so that will make the body fresh, fit and energetic
Jamu godog bugar ayu
Usneae thalus, Zingiberis purpurei rhizome, Retrofracti fructus, Santali lignum, Sappan lignum, Illicium verum, Kaempferiae rhizome, Curcumae rhizome, Foenigraeci semen, Andrographidis herba, Centellae herba, Curcumae domesticae rhizome
Slowing the ageing process, improving the blood circulation, adding more energy and strengthen
Jamu Jago Encok Orthosiphonis folia, Zingiberis zerumbeti rhizome, Zingiberis rhizome Rheumatism
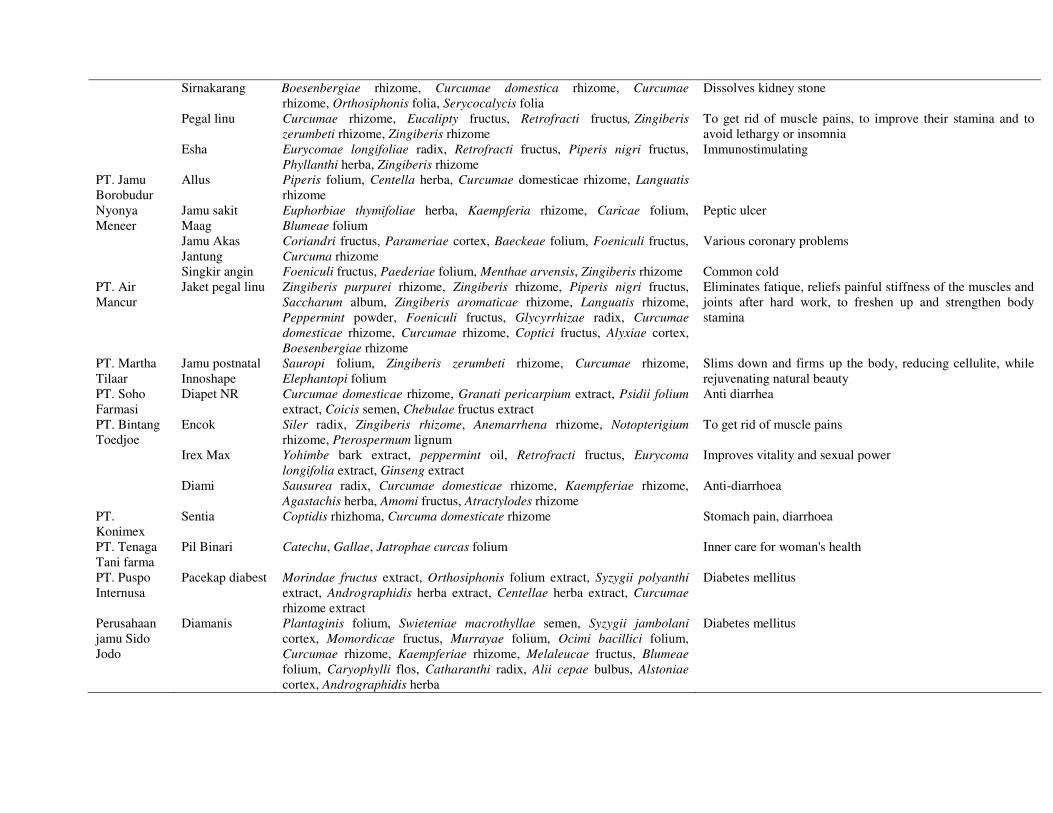
20
Sirnakarang Boesenbergiae rhizome, Curcumae domestica rhizome, Curcumae rhizome, Orthosiphonis folia, Serycocalycis folia
Dissolves kidney stone
Pegal linu Curcumae rhizome, Eucalipty fructus, Retrofracti fructus, Zingiberis zerumbeti rhizome, Zingiberis rhizome
To get rid of muscle pains, to improve their stamina and to avoid lethargy or insomnia
Esha Eurycomae longifoliae radix, Retrofracti fructus, Piperis nigri fructus, Phyllanthi herba, Zingiberis rhizome
Immunostimulating
PT. Jamu Borobudur
Allus Piperis folium, Centella herba, Curcumae domesticae rhizome, Languatis rhizome
Nyonya Meneer
Jamu sakit Maag
Euphorbiae thymifoliae herba, Kaempferia rhizome, Caricae folium, Blumeae folium
Peptic ulcer
Jamu Akas Jantung
Coriandri fructus, Parameriae cortex, Baeckeae folium, Foeniculi fructus, Curcuma rhizome
Various coronary problems
Singkir angin Foeniculi fructus, Paederiae folium, Menthae arvensis, Zingiberis rhizome Common cold PT. Air Mancur
Jaket pegal linu Zingiberis purpurei rhizome, Zingiberis rhizome, Piperis nigri fructus, Saccharum album, Zingiberis aromaticae rhizome, Languatis rhizome, Peppermint powder, Foeniculi fructus, Glycyrrhizae radix, Curcumae domesticae rhizome, Curcumae rhizome, Coptici fructus, Alyxiae cortex, Boesenbergiae rhizome
Eliminates fatique, reliefs painful stiffness of the muscles and joints after hard work, to freshen up and strengthen body stamina
PT. Martha Tilaar
Jamu postnatal Innoshape
Sauropi folium, Zingiberis zerumbeti rhizome, Curcumae rhizome, Elephantopi folium
Slims down and firms up the body, reducing cellulite, while rejuvenating natural beauty
PT. Soho Farmasi
Diapet NR Curcumae domesticae rhizome, Granati pericarpium extract, Psidii folium extract, Coicis semen, Chebulae fructus extract
Anti diarrhea
PT. Bintang Toedjoe
Encok Siler radix, Zingiberis rhizome, Anemarrhena rhizome, Notopterigium rhizome, Pterospermum lignum
To get rid of muscle pains
Irex Max Yohimbe bark extract, peppermint oil, Retrofracti fructus, Eurycoma longifolia extract, Ginseng extract
Improves vitality and sexual power
Diami Sausurea radix, Curcumae domesticae rhizome, Kaempferiae rhizome, Agastachis herba, Amomi fructus, Atractylodes rhizome
Anti-diarrhoea
PT. Konimex
Sentia Coptidis rhizhoma, Curcuma domesticate rhizome Stomach pain, diarrhoea
PT. Tenaga Tani farma
Pil Binari Catechu, Gallae, Jatrophae curcas folium Inner care for woman's health
PT. Puspo Internusa
Pacekap diabest Morindae fructus extract, Orthosiphonis folium extract, Syzygii polyanthi extract, Andrographidis herba extract, Centellae herba extract, Curcumae rhizome extract
Diabetes mellitus
Perusahaan jamu Sido Jodo
Diamanis Plantaginis folium, Swieteniae macrothyllae semen, Syzygii jambolani cortex, Momordicae fructus, Murrayae folium, Ocimi bacillici folium, Curcumae rhizome, Kaempferiae rhizome, Melaleucae fructus, Blumeae folium, Caryophylli flos, Catharanthi radix, Alii cepae bulbus, Alstoniae cortex, Andrographidis herba
Diabetes mellitus

� � ���������������� �������������������������������
21
Biological activity of the most common plants in jamu Biological activities of the most common plants in jamu as reported in the literature are summarized in Table 2. In the following sections in more details are discussed. Anticancer
Plants from the family Zingiberaceae are the most often used ingredient of jamu. Eleven Curcuma species (Curcuma aeruginosa, C. aurantiaca, C. colorata, C. domestica (synonym: C. longa), C. euchroma, C. mangga, C. petiolata, C. purpurascens, C. soloensis, C. xanthorrhizae, and C. zedoria) have been used traditionally as a spice and to treat several illness such as appendicitis, asthma, itch, rheumatism, abdominalgia, anemia, hypertension, diarrhea, and dysentery. Curcumin is a main phenolic constituent of the genus especially in the rhizome of tumeric (Curcuma domestica). Although C. domestica, that is also called C. longa, has not been used traditionally for anticancer purposes, recent investigations show that this plant has promising effects in this area, mainly to be ascribed to curcumin (Fig. 1). The mechanism of action of curcumin has been partly elucidated. Inducing apoptosis plays an important role. Furthermore, it reduces the cell cycle progression thereby preventing cancerous cell growth (Chattopadhay et al., 2004, Karunagaran et al., 2005). In vitro and in vivo, it suppressed carcinogenesis of the liver, kidney, colon, and breast (Okazaki et al., 2005, Kirana et al., 2003). Preclinical and clinical studies with curcumin in relation to its anticancer potential have been reviewed. Human clinical trials indicated no dose-limiting toxicity up to 10 g/day taken orally. These studies carried out so far suggest that curcumin has potential in the prevention and therapy of cancer (Aggarwal et al., 2003, Sharma et al., 2005). For C. xanthorrhiza that is used traditionally as antibacterial, anticancer and anti-inflammatory agent scientific proof has been given for its antiproliferative and anticancer activities. These activities are largely attributed to the sesquiterpene compound xanthorrhizol isolated from this plant. It significantly increased apoptosis in HeLa cells (Ismail et al., 2005). Ginger (Zingiber officinale Rose) that contains the phenolic ketones gingerol and paradol has been launched in Indonesia as fitofarmaka (phytomedicine) for malignancies (antineoplasma). Anticancer activity of the ginger extract has been reported in vitro and in vivo. The strongest anticancer activity has been shown for another Zingiberaceae species, Zingiber aromaticum (Kirana et al., 2003, Manju and Nalini, 2005). It has been suggested that Z. aromaticum containing the sesquiterpene zerumbone, also has the potential to be developed as fitofarmaka with anticancer properties. Panduratin, a chalcone derivative isolated from Kaemferia pandurata rhizome has been reported to suppress carcinogenesis in human colon cancer cell lines (Kirana et al., 2003, Yun et al., 2005). Pinostrobin, a flavonoid from this plant showed cytotoxic activity against human mammary carcinoma cells (Sukardiman et al., 2000). Ethyl trans-cinnamate and ethyl 4-methoxy-trans-cinnamate from galanga root oil (Alpinia galanga) induced the activity of the detoxifying enzyme, glutathione S-transferase (GST), a major mechanism for chemical carcinogen detoxification (Zheng et al., 1993). Another isolated compound from this plant, l'-acetoxychavicol acetate has been found to suppress chemical- and virus-induced tumor initiation and promotion. Although the mechanism is not fully understood, this compound inhibits activation of NF-κB and NF-κB-regulated gene expression. This may explain its ability to enhance apoptosis and to inhibit invasion (Ichikawa et al., 2005). Isolated compounds from jamu showed antioxidative activity in vitro using H4IIE rat hepatoma cells. Kaempferol and luteolin protected these cells against oxidative stress. The ability of kaempferol and luteolin to inhibit oxidative DNA strand breaks supports their suggested role as protective agents against diseases such as cancer (Steffan et al., 2005). Three anthraquinone glycosides (pulmatin, chrysophanein and physcionin) isolated from Rheum palmatum roots exhibited moderate cytotoxic activity against HeLa epitheloid cells and inhibited the growth of BT-20 human breast carcinoma cells (Kubo et al., 1992). The in vitro cytotoxicity of the plumieride, an iridoid compound which was isolated

����������
22
from methanol extract of the bark of Plumeria bicolor and several analogues was determined in radiation-induced fibrosarcoma (RIF) tumor cells. The analogues gave stronger activity than plumieride itself (Dobhal et al., 2004). An ethanol extract of the bark of Alstonia scholaris enhanced the anticancer activity of berberine in the Ehrlich ascites carcinoma-bearing mice. This extract also showed cytotoxic activity to HeLa cells. Compared to the active principle echitamine, present in Alstonia scholaris, the extract was more powerful to kill HeLa cells. The cytotoxic activity of the extract depends on the season of collection of the plant bark. The extract of bark collected in the summer season has the highest activity (Jagetia and Baliga, 2004, 2005). Usually this plant to be used in jamu, is collected during the dry season (also considered as the summer season). Andrographis paniculata that is called sambiloto by local people in Indonesia has been intensively investigated for its anticancer activity. The diterpenoid compounds 14-deoxyandrographolide and 14-deoxy-11,12-didehydroandrographolide isolated from aerial parts of this plant showed marked activity against a human breast carcinoma cell lines (Tan et al., 2005). The consumption of Ardisia compressa tea (aqueous extract) resulted in complete inhibition of the chemically-induced hepatocarcinogenesis in Wistar rats (De Mejia and Ramirez, 2004). Catharanthus roseus that has been used to treat cancer (Eisei, 1995) contains the clinically used anticancer drugs vincristine, vinblastin and other vinca alkaloids (Cragg and Newman, 2005). A water extract of Centella asiatica significantly reduced the multiplicity of neoplasms in the small intestine. This result suggests that C. asiatica has a chemopreventive effect on colon tumorigenesis in male F344 rats (Bunpo et al., 2004). 2'-Hydroxycinnamaldehyde isolated from Cinnamomum cassia bark, strongly inhibited the in vitro growth of a broad panel human cancer cells and the in vivo growth of the SW-620 human tumor xenograft (Lee et al., 1999). Coriandrum sativum was shown to act protectively against the deleterious effects in lipid metabolism in experimental colon cancer (Chithra and Leelamma, 2000). Ganopoly, an aqueous polysaccharide fraction extracted from the fruiting bodies of Ganoderma lucidum has antitumor activity combined with immunomodulating activity. Ganopoly significantly reduced the tumor weight in a dose-dependent manner, with inhibition rates of 32.3, 48.2, and 84.9% at doses of 20, 50, and 100 mg/kg, respectively in mice. It may represent a novel promising immunotherapeutic agent or a lead for cancer treatment (Gao et al., 2005). Immunomodulating effects that may be useful in the treatment of cancer have been reported for ethanolic extracts of aerial parts of Phyllanthus niruri (Ma’at, 2002). Combination of anticancer drugs such as paclitaxel with the herbal extracts e.g. from Glycyrrhizae radix, Rhei rhizome, Scutellariae radix, Zizyphi fructus and Zingiberis rhizome enhanced the paclitaxel sensitivity in HeLa cells via the inhibition of multidrug resistance. These extract suppressed the growth of HeLa cells concentration dependently. The results concluded that the combination of anticancer drugs with some herbal extracts contributes to the improvement of clinical outcomes in cancer chemotherapy (Takara et al., 2005). Alkaloids and quassinoids from Eurycoma longifolia, iridoids and lignans from Plumeira rubra showed cytotoxic activity to human breast, colon, fibrosarcoma, lung, melanoma, KB, KB-V1 cancer cell lines and in murine lymphocytic leukemia (Kardono et al., 1990, 1991). Antiviral
Hundreds of medicinal plants used in jamu have been tested for antiviral activity in vitro and in vivo. But the antiviral efficacy of such herbal medicine has seldom been tested in rigorous clinical trial. The methanol extracts of plants used in jamu e.g Andrographis paniculata, Swietinia mahagoni and Curcuma aeruginosa showed anti-HIV activity using HIV-I-infected MT-4 cells. With the dose range of 4.2 to 175 µg mL-1 they inhibited the HIV-protease (Otaka et al., 1995). Methanol extracts of Melaleuca leucadendron fruit and Annona muricata stembark collected in Indonesia have been reported to be active against herpes simplex virus-1 in vitro. M. leucadendron significantly prolonged the development of skin lesions and reduced the mortality (Padma et al., 1998, Nawawi et al., 1999). Aqueous extracts, tannin, lignan and other isolated compounds from Phyllanthus species have been tested for their anti-

� � ���������������� �������������������������������
23
HIV activity in vitro and in vivo. They inhibited the HIV-key enzymes e.g. integrase, reverse transcriptase and protease (Calixto et al., 1998, Notka et al., 2004). The genus Phyllanthus has been intensively studied clinically for its antiviral effects. A systematic review of 22 randomized clinical trial showed that Phyllanthus species have positive effect on antiviral activity and show positive effects on liver biochemistry in chronic hepatitis B virus infection (Liu et al., 2001, Calixto et al., 1998). Andrographis paniculata was also clinically tested for its antiviral activity. A phase I clinical trial of andrographolide from A. paniculata was conducted in 13 HIV positive patients and five HIV uninfected, healthy volunteers. This trial concluded that andrographolide may inhibit HIV induced cell cycle deregulation, leading to a rise in CD4 (+) lymphocyte levels in HIV-1 infected individuals (Calabrese et al., 2000). Helicterins A-F (Fig. 1), dimeric (7.5',8.2')-neolignans with a bicyclo[2.2.2]octene C-framework isolated from Helicteres isora showed a mild inhibitory activity against reverse transcriptase from avian myeloblastosis virus (Tezuka et al., 2000). Antimalaria and antiparasitic
Most of the plants mentioned here have been traditionally used as antimalarial agent in Indonesia. Methanol extracts prepared from stem bark of Alstonia scholaris, A. macrophylla and A. glaucescense have been assessed for antiplasmodial activity against multidrug-resistant K1 strain of Plasmodium falciparum cultured in human erythrocytes. The active indole alkaloids from these extracts, in contrast to chloroquine, had a significantly higher affinity to the K1 strain than to the T9-96 strain (Keawpradub et al., 1999). 1,2-Dihydroxy-6,8-dimethoxy-xanthone, isolated from Andrographis paniculata possessed in vitro activity against P. falciparum. In vivo it gave a reduction (62%) in parasitemia after treating the Swiss Albino mice with P. berghei (Dua et al., 2004). The petroleum ether extracts of the rind of Carica papaya and Citrus sinensis also showed antimalarial activity against strain P. falciparum FCK 2 in vitro (Bhat and Surolia, 2001). Screening of plant extracts that are traditionally used for the treatment of malaria on Java showed strong antimalarial and antibabesial activitiy. They include Achillea millefolium, Baeckea frutenscens, Brucea javanica, Curcuma xanthorrhiza, Strychnos lucida, Swietenia macrophylla and Phyllantus niruri (Trimurningsih et al., 2005, Subeki et al., 2005). Antibabesial activity was also found for protoberberine alkaloids and 20-hydroxyecdysone from Arcangelisia flava against Babesia gibsoni (Subeki et al., 2005). An in vitro study on traditionally used malaria remedies in the Kenyah of the Apo Kayan, East Kalimantan (Indonesian Borneo) concluded that plants such as Lansium domesticum and Carica papaya are more likely to be effective antimalarials. These herbal remedies were found to have activity against chloroquine-resistant P. falciparum (Leaman et al., 1995). Eurycomanone and 7-methoxy-β-carboline-1-propionic acid from Eurycoma longifolia, triterpenoid lansioides from Lansium domesticum, and Azadirachta indica collected from Kalimantan demonstrated significant antimalarial activity (Kardono et al., 1991, Omar et al., 2003). Anti-inflammatory, antirheumatic, antipyretic and analgesic
The anti-inflammatory effects of extract of Morinda officinalis (noni or mengkudu) in vitro and in vivo have been shown by inhibition of the production of nitric oxide, prostaglandin E-2 and tumor necrosis factor-alpha in lipopolysaccharide-stimulated RAW 264.7 macrophages (Kim et al., 2005). The inhibition of the prostaglandin E2 production has been also shown by Aloe vera gel. Another effect of A. vera was the inhibition on reactive oxygen metabolites in the human colorectal mucosa. This finding may have a therapeutic relevance in inflammatory bowel disease (Langmead et al., 2004). The aqueous extract of tempe (fermented soja-beans) which is a popular food in Indonesia have been reported to have anti-inflammatory, antioxidant and antithrombotic activity in an experimental photochemical thrombogenesis model using rat femoral artery (Rilantono et al., 2000). Cinnamomom cortex that was collected in Indonesia inhibited the rise in vascular permeability and edema induced by acetic acid,

����������
24
carrageenin, serotonin and arachidonic acid. The effect was also shown on secondary lesions in the development of adjuvant-induced arthritis (Kubo et al., 1996). Hydroxypanduratin A and panduratin A isolated from Kaempferia pandurata rhizome showed significant topical anti-inflammatory activity in the assay of TPA-induced ear edema in rats. The presence of these compounds may very well be related to the uses of this plant in traditional medicine (Tuchinda et al., 2002). The lignans niranthin, phyltetralin and nirtetralin isolated from aerial parts of Phyllanthus amarus exhibited marked anti-inflammatory properties and suggest that these lignans are the main active principles responsible for the traditional application of this plant for anti-inflammatory properties (Kassuya et al., 2005). Screening of 75 medicinal plants collected in Indonesia showed that many of them had the inhibitory effects on the nitric oxide (NO) production in lipopolysaccharide-stimulated RAW264.7 macrophages as well as antioxidant activity through the evaluation of free radical scavenging effect and reducing power (Choi and Hwang, 2005). NO is widely recognized as an important messenger and effective molecule in a variety of biological system. The NO production is inhibited by the nitric oxidase inhibitors that can be used as therapeutic agents for inflammatory diseases (Tinker and Wallace, 2006). Chrubasik et al. (2005) comprehensively reviewed on the effects of an ethanol extract of ginger (Zingiber officinale rhizome) and its efficacy profiles in vitro, in vivo and in clinical studies. The ginger extracts showed a pain relieving effects in which up to 0.3 g ginger/day for musculoskeletal pain in human were administered. This review, however, suggested the further studies to prove the efficacy and to find an optimum dosage of ginger preparations in the treatment of osteoarthritic pain. The ethanol extract of ginger (Z. officinale) together with Alpinia galanga, Curcuma longa, Camellia sinensis and Uncaria tomentosa had also a statistically significant effect on reducing symptoms of osteoarthritis of the knee of patients (Altman and Marcussen, 2001, Ahmed et al., 2005). Hepatoprotective
Herbal preparations containing Andrographis panuculata and Phyllanthus amarus for various liver disorders have been proved to have antihepatotoxic activity (Ram, 2001). The ethanol extract and isolated diterpenes andrographolide and neoandrographolide from the aerial parts of Andrographis paniculata showed significant antihepatotoxic action in Plasmodium berghei K173-induced hepatic damage in Mastomys natalensis (Chander et al., 1995). The ethanol extract of Carica papaya seeds caused elevation of rat serum levels of acid phosphatase (ACP), alkaline phosphatase (ALP), and aspartate amino transferase (AST). Also mild to severe metaplasia of hepatocytes was revealed in a dose-related manner as well as proliferation of Kupfer cells and hepatic cells cirrhosis. These biochemical and pathological changes indicated liver cell damage and malfunction (Udoh and Udoh, 2005). A hepatoprotective effect of ethanol extracts of turmeric together with sesquiterpenes and curcuminoid containing fractions has been shown to be related to the suppression of alanin and aspartate aminotransferase and lactate dehydrogenase level on D-galactosamin induced liver injury in rats (Miyakoshi et al., 2004). The hepatoprotective effect of Alstonia scholaris bark on liver injuries induced by the carbon tetrachloride (CCl4), β-D-galactosamine, acetaminophen and ethanol were investigated by means of serum-biochemical and histopathological examinations. Ethanol extracts of A. scholaris bark significantly lowered β-D-galactosamine induced serum transaminases elevation in the serum-biochemical analysis in rats (Lin et al., 1996). (CCl4)-induced hepatotoxicity in the liver of rats, as judged by the raised serum enzymes, glutamate oxaloacetate transaminase and glutamate pyruvate transaminase, was prevented by pretreatment with the extracts of Phyllanthus niruri, demonstrating its hepatoprotective action (Harish and Shivanandappa, 2006).

� � ���������������� �������������������������������
25
Antidiabetic
Diabetes mellitus is recognized by chronic elevation of the glucose level in the blood and often accompanied by symptoms of severe thirst, profuse urination, polyuria, weigh loss, and stupor. Medicinal plants that are used clinically to treat diabetes have shown their antidiabetes activity in vitro, in vivo and in clinical studies. The methanol and aqueous extracts derived from Alpinia galanga caused highly significant reduction in the blood glucose levels of normal rabbits (Akhtar et al., 2002). The glucosidic compounds 4'-O-methylpiceid and rhapontin, isolated from kelembak (Rheum palmatum) roots that were collected from the market in Indonesia exhibited moderate α-glucosidase inhibitory activity in vitro. The inhibition of α-glucosidase activity may be effective in controlling abnormal levels of blood glucose in metabolic diseases such as diabetes (Kubo et al., 1991). Hypolipidemic effects have been shown for aqueous extracts of cumin seeds (Cuminum cyminum) on alloxan-induced diabetic, triton and cholesterol fed hyperlipemic rats. Hyperlipidemia is an associated complication of diabetes mellitus. In this study, administering cumin extract to diabetic rats significantly reduced the blood glucose level. The mechanism may be by potentiating the insulin effect or by increasing the pancreatic secretion of insulin from the cells (Dhandapani et al., 2002). Guazuma ulmifolia leaves and Trigonella fonum graceum seeds that are used clinically against diabetes mellitus have been studied for their antihyperglycemic effect. Aqueous extract of these plants reduced hyperglycemic peak in the rabbits (Alarcon-Aguilara et al., 1998). Ganoderma lucidum, the water and ethanol extracts of Piper betle and dianex, a polyherbal formulation consisting of the aqueous extracts of Gymnema sylvestre leaves, Eugenia jambolana seeds, Momordica charantia fruits, Azadirachta indica leaves, Cassia auriculata flowers, Aegle marmelose fruits, Withania somnifera roots, and Curcuma longa rhizome had hypoglycemic activity in normal and streptozotocin induced diabetic mice and rats (Yang et al. 2004, Mutalik et al., 2005, Arambewela et al., 2005). Most of those plants are used in jamu. Preclinical evaluation consisting of animal studies, acute and subacute toxicity testing and evaluation of the antidiabetic effect of Eugenia jambolana seed powder in streptozotocin-diabetic rats was adequate for approval to start phase 2 clinical trials to evaluate this seed powder as complementary therapy in type 2 diabetes. The study showed that E. jambolana possibly acts as a hypoglycemic agent by increasing insulin levels. Toxicity studies showed no evidence of mortality or abnormality (Sridhar et al., 2005). The total triterpenoid fraction from aerial parts of Centella asiatica, has been studied in the patients with diabetic microangiopathy. It was shown that this fraction is useful in diabetic microangiopathy by improving microcirculation and decreasing capillary permeability and protects against the deterioration of microcirculation due to diabetic microangiopathy (Cesarone et al., 2001). Antimicrobial and antifungal
Antibacterial and antifungal activities have been shown by aqueous extracts, andrographolide and arabinogalactan proteins isolated from Andrographis paniculata and are comparable (in term of growth inhibition of Bacillus subtilis, Escherichia coli, Pseudomonas aeruginosa and Candida albicans) to some known antibiotics, streptomycin, gentamycin and nystatin (Singha et al., 2003). Extracts of Alstonia scholaris, Anacardium occidentale (hexane), and Carica papaya seeds (methanol and buthanol) have been reported to possess a broad spectrum of antibacterial activity (Bouttier et al., 2002, Khan et al., 2003, Dawkins et al., 2003). The growth-inhibiting activity of cinnamaldehyde isolated from Cinnamomum cassia toward human intestinal bacteria (Clostridium perfringens, Bacteroides fragilis and Bifidobacterium bifidum) was shown using an impregnated paper disk method and compared with that of tetracycline and chloramphenicol (Lee and Ahn, 1998). The essential oils of Coriandrum sativum and Foeniculum vulgare were reported to possess antibacterial activity to Escherichia coli and Bacillus megaterium in vitro (Lo Cantore et al., 2004). The essential oil from Cuminum cyminum and the isolated compounds, p-mentha-1,4-dien-7-al, cumin aldehyde, γ-terpinene, and β-pinene, showed antibacterial

����������
26
activity against the genera Clavibacter, Curtobacterium, Rhodococcus, Erwinia, Xanthomonas, Ralstonia, and Agrobacterium (Iacobellis et al., 2005). The essential oils from Cymbopogon citratus, C. nardus, and C. schoenanthus showed a fungistatic effect against superficial mycosis (Koba et al., 2003). The essential oil of Cinnamomum burmanni (bark and leaves) and Tagetes erecta (leaves) that were collected in Indonesia have been reported to exhibit antimicrobial activity against Bacillus subtilis and Salmonella typhimurium and antifungal activity against Candida albicans in vitro (Sukandar et al., 1999, Hartati et al., 1999). An ethyl acetate extract of Curcuma longa has been reported to have antibacterial activity and the potential to restore the effectiveness of β-lactams against methicillin-resistant Staphylococcus aureus (MRSA), and inhibit the MRSA invasion of human mucosal fibroblasts (Kim et al., 2005). A study of Indonesian plants with ethnomedical uses showed that the methylene chloride and methanol extracts of Terminalia catappa, Swietenia mahagoni, Phyllanthus acuminatus, Ipomoea spp., Tylophora asthmatica and Hyptis brevipes have the antibacterial activities against Eschericia coli, Staphylococcus aureus, Xanthomonas campestris and Bacillus subtilis and the antifungal activities against C. albicans, Pythium ultimum, Rhizoctonia solani and Sclerotium rolfsii (Goun et al., 2003). The ethanol extracts from several plant species belonging to the Zingiberaceae family used in Kenyah (Indonesian Borneo), especially Alpinia galanga, Curcuma zedoaria and Zingiberis purpureum, were found to have pronounced inhibitory activities against a wide variety of human pathogenic fungi, including strains resistant to the common antifungals amphotericin B and ketoconazole. As members of the Zingiberaceae are generally regarded as safe for human consumption, these species are excellent candidates for development as novel therapeutic (Ficker et al., 2003). 1'-Acetoxychavicol acetate, an active compound from Alpinia galanga has antifungal activity against Trichophyton mentagrophytes, T. rubrum, T. concentricum, Rhizopus stolonifer and Aspergillus niger with a concentration 14 mg mL-1 (Janssen and Scheffer, 1985) Gastroprotective
1S-1'-Acetoxychavicol acetate and 1S-1'-acetoxyeugenol acetate, isolated from Alpinia galanga markedly inhibited the ethanol-induced gastric mucosal lesions in rats. The action of 1S-1'-acetoxychavicol was attenuated by pretreatment with indomethacin and N-ethylmalcimide and significantly increased the glutathione (GSH) levels of gastric mucosa in rats. GSH acts as antioxidant and is important for maintaining the mucosal integrity in the stomach (Matsuda et al., 2003). A different mechanism of hepatoprotective activity was shown by asiaticoside, an active triterpenoid constituent of Centella asiatica and its extract. They were found to promote angiogenesis and stimulate blood vessel formation and mucosal cell regeneration during the gastric ulcer healing stage that are important parts of the wound healing. Angiogenesis in granulation tissues improves circulation to the wound site thus providing oxygen and nutrients are essential for the healing process (Cheng et al., 2004). The effect of an ethanolic extract of Aloe vera on acute gastric mucosal lesions induced by 0.6 M HCl and acid output was studied in pylorus ligated and lumen perfused rats, respectively. Aloe vera is endowed with gastric acid anti-secretory activity and could protect the gastric mucosa at low concentrations against injurious agents (Yusuf et al., 2004). Morinda citrifolia (noni) inhibits gastric emptying in male rats via a mechanism involving stimulation of cholecystokinin and its receptor activation. Cholecystokinin is a peptide hormone of the gastrointestinal system responsible for stimulating the digestion of fat and protein. It delays gastric emptying and inhibits gastric acid and plasma gastrin responses (Konturek et al., 1994, Pu et al., 2004). Ethanol and water extract of Abrus cantoniensis, Saussurea lappa, Eugenia caryophyllata, Magnolia officinalis and Ligusticum species strongly inhibited the growth of Helicobacter pylori which is an important etiologic impetus leading usually to chronic active gastritis and gastric ulcer (Li et al., 2005).

� � ���������������� �������������������������������
27
Cardioprotective
A study on the edible plants common in Asian diets such as Ipomoea batatas, Piper betle, Anacardium occidentale, Gynandropsis gynandra, Carica papaya, and Mentha arvensis extracts showed that they exhibited more than 50% relaxing effect on aortic ring preparations. Piper betle and Cymbopogon citratus showed comparable vasorelaxation on isolated perfuse mesenteric artery preparation (Runnie et al., 2004). 14-deoxy-11,12-didehydroandrographolide from Andrographis paniculata was shown to have bradycardia-inducing and β-adrenoceptor antagonistic properties in vivo using anesthetized Sprague-Dawley rats (Zhang et al., 1998). A cardioprotective effect of Centella asiatica on the antioxidant tissue defense system during doxorubicin induced cardiac damage in rats has been reported. The water extracts of this plant resulted in significant reduction in the levels of lactate dehydrogenase, creatine phosphokinase, glutamate oxaloacetate transaminase and glutamate pyruvate transaminase. Increased activity in serum of these enzymes is a well-known diagnostic marker of myocardial function. As active ingredients triterpenes (asiatic acid and asiaticoside) may be responsible for the cardioprotective effect of C. asiatica extracts (Gnanapragasam et al., 2004). The cardioprotection provided by ligustrazine is related to a reduction of TNF-alpha content by inhibition of free radical production in isolated rat hearts. It was known that TNF-alpha can contribute to myocardial damage during ischemia-reperfusion (Zhou et al., 2004). The studies of the antioxidative and cytoprotective effects using H9c2 cardiac myoblasts showed that Phyllanthus urinaria have a protective activity against doxorubicin cardiotoxicity. This protection was mediated through multiple pathways such as enhancement of survival factor through elevation of glutathione, activation of catalase/superoxide dismutase activity and inhibition of lipid peroxidation. This plant may serve as an alternative source of antioxidants for prevention of doxorubicin cardiotoxicity (Chularojmontri et al., 2005). Antihypertensive
The ethanol extracts of fresh matured fruits of Carica papaya markedly depressed the blood pressure and heart rate in mineralocorticoid salt and in renal hypertensive rats when compared with the normotensive controls. The extracts (20 mg/kg i.v.) decreased the blood pressure by about 20.1%, 50.7% and 54.5% in normotensive, renal and deoxycortocosterone acetate-salts hypertensive rats, respectively. The extract appeared to be more potent than hydrallazine (200µg/kg i.v.), a well known antihypertensive (vasodilator) agent that decreased the blood pressure by about 10.7%, 22.8% and 26.4% in those three types of hypertension (Eno et al., 2000). The total triterpenoid fraction of Centella asiatica, a venoactive drug acting on the microcirculation and on capillary permeability, has been tested in three groups of patients with venous hypertension. The improvement of signs and symptoms by extracts observed in venous hypertensive patients correlated well with the improvement of the variation of capillary filtration rate and ankle edema (De Sanctis et al., 2001). The vasodilatory effect of Curcuma herbs has been studied. C. longa induced endothelium-independent vasodilatation. It was concluded that Curcuma herbs have hypotensive and protective effects on the endothelium in spontaneously hypertensive rats, and its mechanism is thought to be related to a radical scavenging effect and improvement of hemorheology (Goto et al., 2005). A major constituent in the water decoction of Orthosiphon aristatus leaves, methylripariochromene A (a benzochromene), exhibited a continuous decrease in systolic blood pressure after subcutaneous administration in conscious stroke-prone spontaneously hypertensive rats. This plant is popular as kumis kucing in Indonesia. Javanese people prescribe the leaves in their jamu, mainly for treatment of hypertension (Matsubara et al., 1999). Anti-asthma, antitussive and anti-allergic
A study of selected medicinal folklore plants that are traditionally used for asthma treatment in Indonesia indicated that alcoholic extracts, from Plantago major leaves, from Eucalyptus globulus

����������
28
leaves and fruits, from Cinnamomum massoiae cortex and from Vitex trifolia leaves and two hexane extracts -Eucalyptus globulus leaves and Vitex trifolia leaves- inhibited IgE-dependent histamine release from RBL-2H3 cells. This suggested that extracts contain active compounds which inhibit mast cell degranulation, and may be used in the development of new drugs for treating asthma and/or allergic disease (Ikawati et al., 2001). An ethanol extract of Alstonia scholaris leaves induced pronounced bronchodilatory activity in anaesthetized rats with the probable involvement of prostaglandins (Channa et al., 2005). Clinical studies with Andrographis paniculata suggested that this plant may be effective as an early treatment of uncomplicated acute upper respiratory tract infection on the patients tested. The ethanol extract of A. paniculata alone or in combination with the ethanol extract of A. senticocus appear to be more effective than placebo (Poolsup et al., 2004). The active constituents of A. paniculata, andrographolide and neoandrographolide, have been reported to have anti-allergic effect. This effect is due to its mast cell stabilizing activity, the same as for the antiallergic drug, disodium cromoglycate. Neoandrographolide was more potent than andrographolide in this study. All three compounds demonstrated significant inhibition of passive cutaneous anaphylaxis (Gupta et al., 1998). An antitussive effect of liquiritin apioside, liquritin and liquiritigenin, isolated from Glychirrhiza radix (licorice) has been reported. The effect of liquiritin apioside may depend on both peripheral (modulation of ATP-sensitive K+ channels) and a central mechanism (modulation of serotonergic system) (Kamei et al., 2005). An aqueous extract of Alpinia galanga rhizome was found to inhibit the release of β-hexosaminidase, a marker of antigen-IgE-mediated degranulation in RBL-2H3 cells. Isolated compounds, 1'S-1'-acetoxychavicol acetate and 1'S-1'-acetoxyeugenol acetate inhibited β-hexosaminidase and a passive coetaneous anaphylaxis reactions in mice and the antigen-IgE-mediated TNF-alpha and IL-4 production, both of which participate in the late phase of type I allergic reactions (Matsuda et al., 2003). Immunostimulating
An immunostimulating effect has been reported from pule (Alstonia scholaris) that is used in South East Asia mainly as antimalarial and antidysentery agents, in BALB/c mice. The bark aqueous extract stimulated non specific immune response, restored the reduction of phagocytic action induced by prednisolone and protected the body from the opportunistic infection caused by Escherichia coli (Iwo et al., 2000). This effect was also shown by curcumin from Curcuma longa in BALB/c mice (Antony et al., 1999). A polysaccharide extract of Alpinia galanga rhizome showed a marked stimulating effect on the reticulo-endothelial system (RES) and increased the number of peritoneal exudate cells, and spleen cells of mice (Bendjeddou et al., 2003). Immunomodulating effects were also shown by a methanol extract, andrographolide, 14-deoxyandrographolide and 14-deoxy-11,12-didehydroandrographolide isolated from Andrograpis paniculata. They enhanced the proliferation and interleukin-2 (IL-2) induction in human peripheral blood lymphocytes (Kumar et al., 2004). The different mechanism of immunostimulating effect was shown by the hexane and aqueous extract of Carica papaya seeds and its bioactive fractions. They significantly enhanced the phytohemagglutinin responsiveness of lymphocytes and inhibited the classical complement-mediated hemolytic pathway (Mojica-Henshaw et al., 2003). Central nervous system (CNS) activity
The essential oil from the fruits of Cuminum cyminum, that is used traditionally as a stimulant exhibited anticonvulsant activity. This effect was shown in both pentylenetrazole- and maximal electroshock-induced seizures in male NMRI mice (Sayyah et al., 2002). Recently, also antidepressant effects have been reported for curcumin. This effect may be mediated by actions in the central monoaminergic neurotransmitter systems (Xu et al., 2005). Glycyrrhizae radix, together with other medicinal plants has been tested to the patients who were exhibiting tremor, a symptom of

� � ���������������� �������������������������������
29
antipsychotic-induced parkinsonism. The results concluded that the combination of those plants was effective against tremor from parkinsonism (Ishikawa et al., 2000). Alstonia macrophylla has been reported to have a CNS depressant activity. It caused a significant reduction in spontaneous activity, a remarkable decrease in exploratory behavioral pattern, a reduction in muscle relaxant activity and also significantly potentiated phenobarbital sodium-induced sleeping time (Chattopadhyay et al., 2004). Other activities
Various other activities have been reported from the medicinal plants which are used in jamu. Grosvenor et al. (1995) surveyed the medicinal plants in Riau Province, Indonesia. Out of one hundred and fourteen species of flowering plants belonging to 51 families, and claimed to have medicinal uses, 50% were recorded to be used to combat fever, 33% for diarrhea and 31% for other gastrointestinal problems. Unny et al. (2003) reviewed about 161 medicinal plants which are a potential source of new contraceptive principles. The review contains the isolated compounds and the mechanism of actions. Some of them are used in jamu, e.g, Foeniculum vulgare, Abrus precatorius, Muraya paniculata, Punica granatum, Curcuma longa, and C. zedoria. They inhibited implantation and increased fetal loss in mice and reduced secretory activity and weight of accessory sex glands. The aqueous extracts of Carica papaya and Ananas comosus have been reported to possess diuretic activity. Both plant extracts gave similar profiles of urinary electrolyte excretion to that of the hydrochlorothiazide. The analysis of the urinary osmolality and electrolyte excretion per unit time, together with the plant salt contents, may help to differentiate the mechanism by which these plants acts as diuretic. The results indicated that the diuretic activity of Ananas comosus was intrinsic and not a result of the salt loading effect, whereas C. papaya extracts may have resulted from a high salt content of this extracts. This activity correlated well with the maximum volume, the highest osmolality, and the amount of electrolytes excreted during urine collection (Sripanidkulchai et al., 2001). The methanol extracts of Areca catechu, Brucea sumatrana, Allamanda cathartica, collected in Sumateran rainforests showed strong antinematodal activity against Bursaphelenchus xylophilus (Alen et al., 2000). Known risks and side effects of medicinal plants used in jamu It is generally assumed by the public, and also even by some medical practitioners, that plant drugs are harmless and therefore are preferable. Put in such general terms this clearly is not always true. In fact various medicinal plants also induce side effects. Powerful herbal drugs like Digitalis, Strychnos, Belladonna and Colchicum can even be highly dangerous. Between 1988 and 2002, 70 patients with a diagnosis of digoxin intoxication at the National Cheng Kung Hospital, Hongkong have been studied. The symptoms that were caused by digoxin overdose included nausea, vomiting, anorexia, weakness, syncope, dizziness and a change in consciousness (Chen et al., 2004). Digitalis may induce the toxicity of licorice (the root of Glycyrrhiza glabra) by drug interaction via licorice-associated electrolyte imbalance, particularly in elderly. Licorice itself may be a cause for exogenously induced hypertension, hypokalemia, hypernatremia, or suppression of the renin-aldosterone system (Harada et al., 2002). Deadly nightshade (Atropa belladonna) intoxication has been infrequently reported in both children and adults. Caksen et al. (2003) showed that meaningless speech, lethargy, coma, and absence of tachycardia were ominous signs in deadly nightshade intoxication in childhood. Huntley et al. (2005) have reviewed the adverse effect of Echinacea species reported between 1950 an 2002. Some adverse effects such as headache, dizziness, tiredness, occasional nausea and abdominal pain have been suffered by people after taking Echinacea. Data from clinical trials suggest that the most commonly experienced adverse effects of Panax ginseng are headache, sleep and gastrointestinal disorder. The possibility of more serious adverse effects even was found in combination products containing ginseng as one of the constituents (Coon and Ernst,

����������
30
2002). Kava-kava (Piper methysticum) may cause tiredness, low energy, headache, gastrointestinal symptoms. A 50-year-old woman was seen with papules and plaques on the face and later on her dorsal and ventral thorax and arms after taking a kava product for 3 weeks (Stevinson et al., 2002). Although ginger (Zingiberis officinale) shows a very broad range pharmacological effect, there are still undesirable effects such as causing heartburn. In the quantities exceeding 6 g dried ginger may act as a gastric irritant. Inhalation of dust from ginger may produce IGE-mediated allergy (Chrubasik et al., 2005).
The risk of herbal medicines producing an adverse reaction depends not only on the medicine and its dosage but also on consumer-related parameters, such as age, genetics, concomitant diseases and co-medication (herb-herb and herb-drug interactions). Reports of herbal medicinal products affected by contamination, adulteration or substitution of botanical material have repeatedly caused concern. Asian herbal medicinal products including jamu are most often implicated (Ernst and Pittler, 2002). One report was found that mentioned the microbial contamination of raw material and end product of jamu gendong (Limyati and Juniar, 1998). Agranulocytosis and citrobacterial infection have been found after using jamu containing phenylbutazone (Paul et al., 2005). A study of 23 commercial jamu showed the occurrence natural aflatoxins that exhibit carcinogenic, teratogenic and mutagenic properties (Ali et al., 2005). A side effect of Morinda citrifolia in a 45-year-old patient who has highly elevated transaminases and lactate dehydrogenase has been reported. This gave rise to the suspicion of herbal toxicity, which was confirmed by taking a liver biopsy (Millonig et al., 2005). Conclusion The in vitro, in vivo and clinical studies on medicinal plants that are used in jamu have scientifically proved their claimed biological activities in part. Species belonging to the family Zingiberaceae such as Curcuma, Zingiber, Kaempferia, are the most frequently used plants in jamu. These species have also been studied intensively for their secondary metabolites and biological activity. Curcumin and panduratin are typical examples of bioactive secondary metabolites from these plants. As members of the Zingiberaceae are generally regarded as safe for human consumption, these species are excellent candidates for development as novel therapeutic. BPOM has done systematic and comprehensive research on 9 priority medicinal plants in Indonesia, i.e. ginger (Zingiber officinale) and king of bitter (Andrographis paniculata) as antineoplasma; turmeric (Curcuma domestica), Java turmeric (C. xanthorrhiza) and Guazuma ulmifolia, as antihyperlipidemic, Java noni (Morinda citrifolia) and Syzygium polyanthum, as antidiabetic and Piper retrofractum as androgenic. Other medicinal plants have been launched as fitofarmaka such as Phyllanthus niruri as immunostimulating, C. xanthorrhiza as antirheumatic, Psidium guajava and C. domestica as antidiarrhea (Bermawie et al., 2005). Based on the literature search, we conclude that there are many more medicinal plants that have potential to be developed as fitofarmaka or as sources of new therapeutic agents. Although the commonly used plants in jamu have been investigated scientifically for their biological activities, the jamu makers or the industries still have to standardize the formulae of jamu in order to assure the efficacy and safety.
Jamu has the potential to develop because it is economically prospective and used to maintain the health and to cure diseases. Compared to Traditional Chinese Medicine (TCM), jamu still needs considerable efforts to reach optimum beneficiary. The scientific study of the common plants should be continued. Exploration of medicinal plants which are the indigenous knowledge of the Indonesian community should also consider ethical issues, such as efficacy, safety, IPR, benefit sharing and biodiversity conservation.
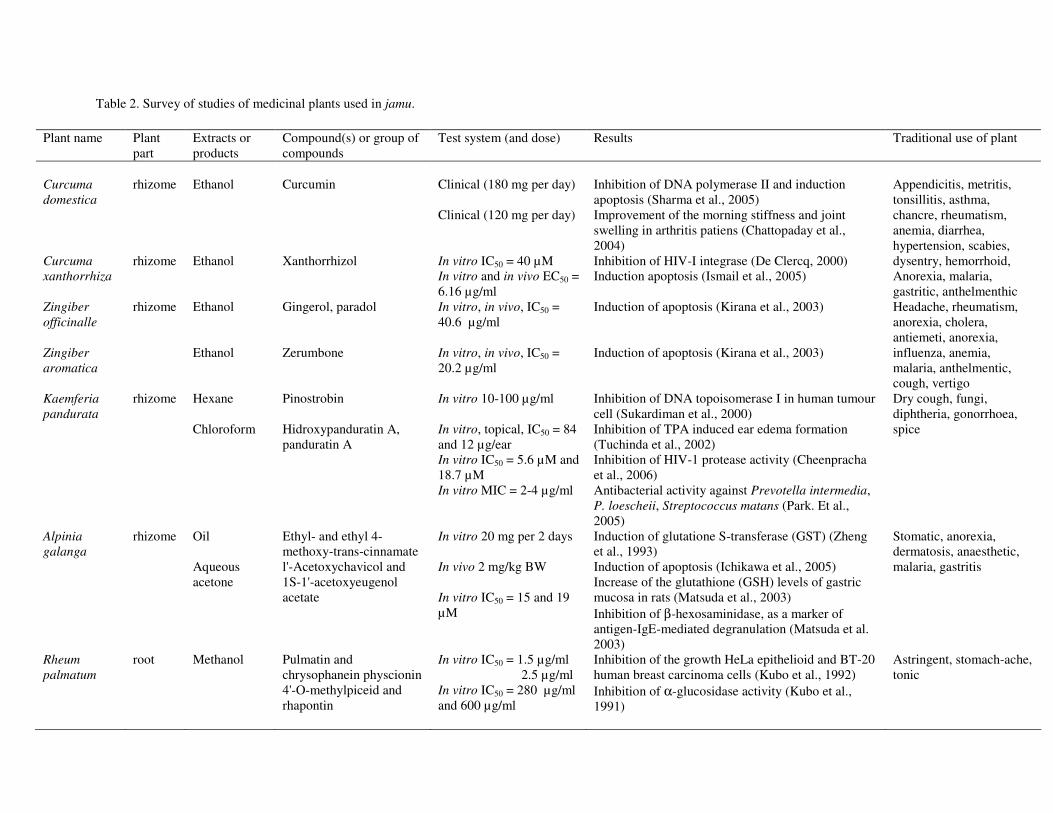
31
Table 2. Survey of studies of medicinal plants used in jamu.
Plant name Plant part
Extracts or products
Compound(s) or group of compounds
Test system (and dose) Results Traditional use of plant
Curcuma domestica Curcuma xanthorrhiza
rhizome rhizome
Ethanol Ethanol
Curcumin Xanthorrhizol
Clinical (180 mg per day) Clinical (120 mg per day) In vitro IC50 = 40 µM In vitro and in vivo EC50 = 6.16 µg/ml
Inhibition of DNA polymerase II and induction apoptosis (Sharma et al., 2005) Improvement of the morning stiffness and joint swelling in arthritis patiens (Chattopaday et al., 2004) Inhibition of HIV-I integrase (De Clercq, 2000) Induction apoptosis (Ismail et al., 2005)
Appendicitis, metritis, tonsillitis, asthma, chancre, rheumatism, anemia, diarrhea, hypertension, scabies, dysentry, hemorrhoid, Anorexia, malaria, gastritic, anthelmenthic
Zingiber officinalle Zingiber aromatica
rhizome Ethanol Ethanol
Gingerol, paradol Zerumbone
In vitro, in vivo, IC50 = 40.6 µg/ml In vitro, in vivo, IC50 = 20.2 µg/ml
Induction of apoptosis (Kirana et al., 2003) Induction of apoptosis (Kirana et al., 2003)
Headache, rheumatism, anorexia, cholera, antiemeti, anorexia, influenza, anemia, malaria, anthelmentic, cough, vertigo
Kaemferia pandurata
rhizome Hexane Chloroform
Pinostrobin Hidroxypanduratin A, panduratin A
In vitro 10-100 µg/ml In vitro, topical, IC50 = 84 and 12 µg/ear In vitro IC50 = 5.6 µM and 18.7 µM In vitro MIC = 2-4 µg/ml
Inhibition of DNA topoisomerase I in human tumour cell (Sukardiman et al., 2000) Inhibition of TPA induced ear edema formation (Tuchinda et al., 2002) Inhibition of HIV-1 protease activity (Cheenpracha et al., 2006) Antibacterial activity against Prevotella intermedia, P. loescheii, Streptococcus matans (Park. Et al., 2005)
Dry cough, fungi, diphtheria, gonorrhoea, spice
Alpinia galanga
rhizome Oil Aqueous acetone
Ethyl- and ethyl 4-methoxy-trans-cinnamate l'-Acetoxychavicol and 1S-1'-acetoxyeugenol acetate
In vitro 20 mg per 2 days In vivo 2 mg/kg BW In vitro IC50 = 15 and 19 µM
Induction of glutatione S-transferase (GST) (Zheng et al., 1993) Induction of apoptosis (Ichikawa et al., 2005) Increase of the glutathione (GSH) levels of gastric mucosa in rats (Matsuda et al., 2003) Inhibition of β-hexosaminidase, as a marker of antigen-IgE-mediated degranulation (Matsuda et al. 2003)
Stomatic, anorexia, dermatosis, anaesthetic, malaria, gastritis
Rheum palmatum
root Methanol Pulmatin and chrysophanein physcionin 4'-O-methylpiceid and rhapontin
In vitro IC50 = 1.5 µg/ml 2.5 µg/ml In vitro IC50 = 280 µg/ml and 600 µg/ml
Inhibition of the growth HeLa epithelioid and BT-20 human breast carcinoma cells (Kubo et al., 1992) Inhibition of α-glucosidase activity (Kubo et al., 1991)
Astringent, stomach-ache, tonic
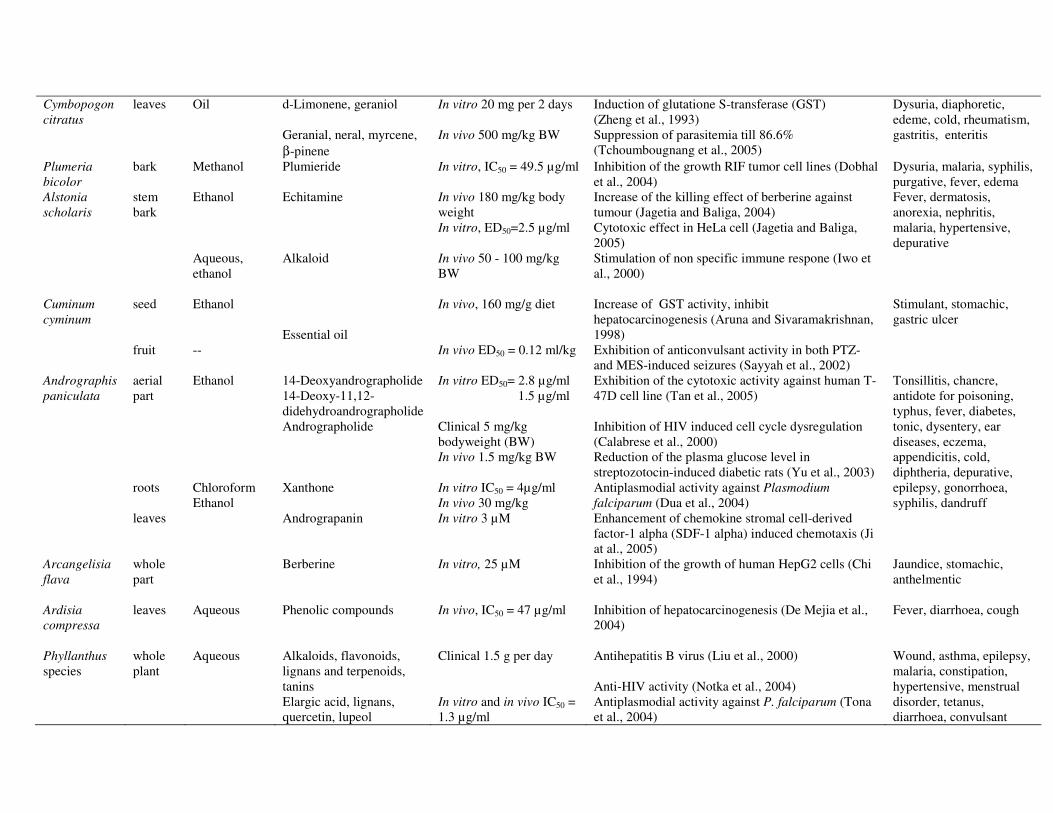
32
Cymbopogon citratus
leaves Oil
d-Limonene, geraniol Geranial, neral, myrcene, β-pinene
In vitro 20 mg per 2 days In vivo 500 mg/kg BW
Induction of glutatione S-transferase (GST) (Zheng et al., 1993) Suppression of parasitemia till 86.6% (Tchoumbougnang et al., 2005)
Dysuria, diaphoretic, edeme, cold, rheumatism, gastritis, enteritis
Plumeria bicolor
bark Methanol Plumieride In vitro, IC50 = 49.5 µg/ml Inhibition of the growth RIF tumor cell lines (Dobhal et al., 2004)
Dysuria, malaria, syphilis, purgative, fever, edema
Alstonia scholaris
stem bark
Ethanol Aqueous, ethanol
Echitamine Alkaloid
In vivo 180 mg/kg body weight In vitro, ED50=2.5 µg/ml In vivo 50 - 100 mg/kg BW
Increase of the killing effect of berberine against tumour (Jagetia and Baliga, 2004) Cytotoxic effect in HeLa cell (Jagetia and Baliga, 2005) Stimulation of non specific immune respone (Iwo et al., 2000)
Fever, dermatosis, anorexia, nephritis, malaria, hypertensive, depurative
Cuminum cyminum
seed fruit
Ethanol --
Essential oil
In vivo, 160 mg/g diet In vivo ED50 = 0.12 ml/kg
Increase of GST activity, inhibit hepatocarcinogenesis (Aruna and Sivaramakrishnan, 1998) Exhibition of anticonvulsant activity in both PTZ- and MES-induced seizures (Sayyah et al., 2002)
Stimulant, stomachic, gastric ulcer
Andrographis paniculata
aerial part roots leaves
Ethanol Chloroform Ethanol
14-Deoxyandrographolide 14-Deoxy-11,12-didehydroandrographolide Andrographolide Xanthone Andrograpanin
In vitro ED50= 2.8 µg/ml 1.5 µg/ml Clinical 5 mg/kg bodyweight (BW) In vivo 1.5 mg/kg BW In vitro IC50 = 4µg/ml In vivo 30 mg/kg In vitro 3 µM
Exhibition of the cytotoxic activity against human T-47D cell line (Tan et al., 2005) Inhibition of HIV induced cell cycle dysregulation (Calabrese et al., 2000) Reduction of the plasma glucose level in streptozotocin-induced diabetic rats (Yu et al., 2003) Antiplasmodial activity against Plasmodium falciparum (Dua et al., 2004) Enhancement of chemokine stromal cell-derived factor-1 alpha (SDF-1 alpha) induced chemotaxis (Ji at al., 2005)
Tonsillitis, chancre, antidote for poisoning, typhus, fever, diabetes, tonic, dysentery, ear diseases, eczema, appendicitis, cold, diphtheria, depurative, epilepsy, gonorrhoea, syphilis, dandruff
Arcangelisia flava
whole part
Berberine In vitro, 25 µM Inhibition of the growth of human HepG2 cells (Chi et al., 1994)
Jaundice, stomachic, anthelmentic
Ardisia compressa
leaves Aqueous Phenolic compounds In vivo, IC50 = 47 µg/ml Inhibition of hepatocarcinogenesis (De Mejia et al., 2004)
Fever, diarrhoea, cough
Phyllanthus species
whole plant
Aqueous Alkaloids, flavonoids, lignans and terpenoids, tanins Elargic acid, lignans, quercetin, lupeol
Clinical 1.5 g per day In vitro and in vivo IC50 = 1.3 µg/ml
Antihepatitis B virus (Liu et al., 2000) Anti-HIV activity (Notka et al., 2004) Antiplasmodial activity against P. falciparum (Tona et al., 2004)
Wound, asthma, epilepsy, malaria, constipation, hypertensive, menstrual disorder, tetanus, diarrhoea, convulsant
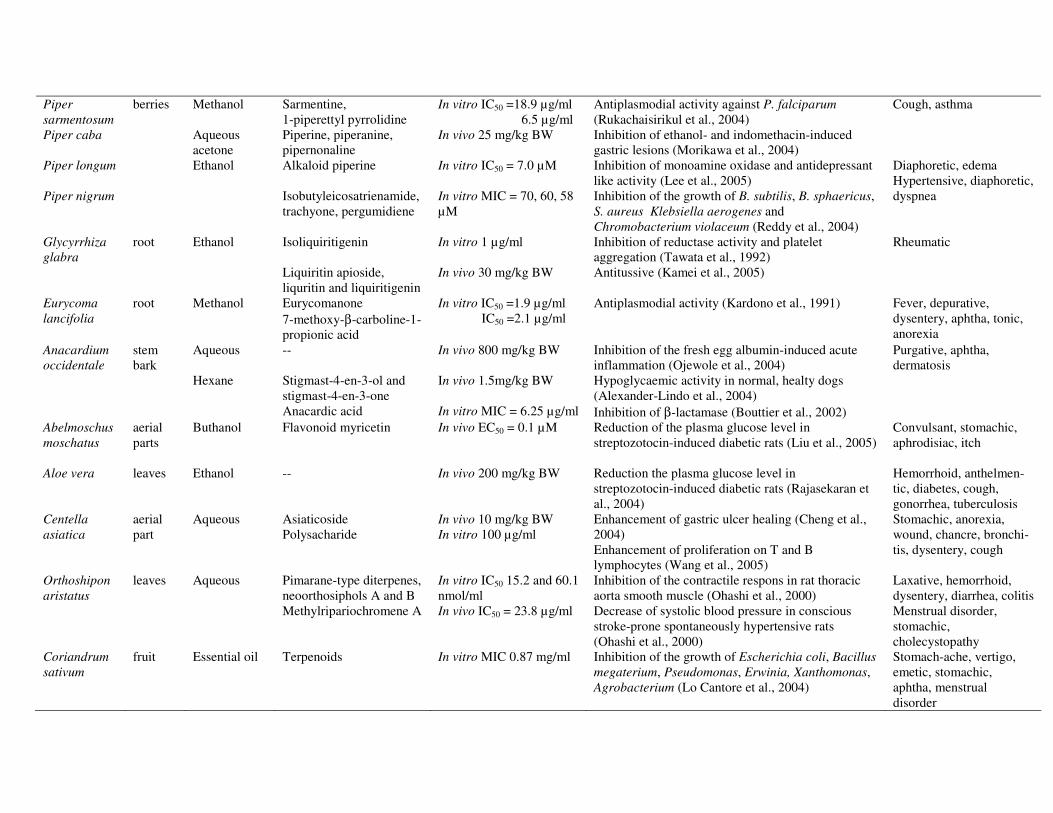
33
Piper sarmentosum Piper caba Piper longum Piper nigrum
berries
Methanol Aqueous acetone Ethanol
Sarmentine, 1-piperettyl pyrrolidine Piperine, piperanine, pipernonaline Alkaloid piperine Isobutyleicosatrienamide, trachyone, pergumidiene
In vitro IC50 =18.9 µg/ml 6.5 µg/ml In vivo 25 mg/kg BW In vitro IC50 = 7.0 µM In vitro MIC = 70, 60, 58 µM
Antiplasmodial activity against P. falciparum (Rukachaisirikul et al., 2004) Inhibition of ethanol- and indomethacin-induced gastric lesions (Morikawa et al., 2004) Inhibition of monoamine oxidase and antidepressant like activity (Lee et al., 2005) Inhibition of the growth of B. subtilis, B. sphaericus, S. aureus Klebsiella aerogenes and Chromobacterium violaceum (Reddy et al., 2004)
Cough, asthma Diaphoretic, edema Hypertensive, diaphoretic, dyspnea
Glycyrrhiza glabra
root Ethanol Isoliquiritigenin Liquiritin apioside, liquritin and liquiritigenin
In vitro 1 µg/ml In vivo 30 mg/kg BW
Inhibition of reductase activity and platelet aggregation (Tawata et al., 1992) Antitussive (Kamei et al., 2005)
Rheumatic
Eurycoma lancifolia
root Methanol Eurycomanone 7-methoxy-β-carboline-1-propionic acid
In vitro IC50 =1.9 µg/ml IC50 =2.1 µg/ml
Antiplasmodial activity (Kardono et al., 1991) Fever, depurative, dysentery, aphtha, tonic, anorexia
Anacardium occidentale
stem bark
Aqueous Hexane
-- Stigmast-4-en-3-ol and stigmast-4-en-3-one Anacardic acid
In vivo 800 mg/kg BW In vivo 1.5mg/kg BW In vitro MIC = 6.25 µg/ml
Inhibition of the fresh egg albumin-induced acute inflammation (Ojewole et al., 2004) Hypoglycaemic activity in normal, healty dogs (Alexander-Lindo et al., 2004) Inhibition of β-lactamase (Bouttier et al., 2002)
Purgative, aphtha, dermatosis
Abelmoschus moschatus
aerial parts
Buthanol Flavonoid myricetin In vivo EC50 = 0.1 µM Reduction of the plasma glucose level in streptozotocin-induced diabetic rats (Liu et al., 2005)
Convulsant, stomachic, aphrodisiac, itch
Aloe vera
leaves
Ethanol
--
In vivo 200 mg/kg BW
Reduction the plasma glucose level in streptozotocin-induced diabetic rats (Rajasekaran et al., 2004)
Hemorrhoid, anthelmen-tic, diabetes, cough, gonorrhea, tuberculosis
Centella asiatica
aerial part
Aqueous Asiaticoside Polysacharide
In vivo 10 mg/kg BW In vitro 100 µg/ml
Enhancement of gastric ulcer healing (Cheng et al., 2004) Enhancement of proliferation on T and B lymphocytes (Wang et al., 2005)
Stomachic, anorexia, wound, chancre, bronchi-tis, dysentery, cough
Orthoshipon aristatus
leaves Aqueous Pimarane-type diterpenes, neoorthosiphols A and B Methylripariochromene A
In vitro IC50 15.2 and 60.1 nmol/ml In vivo IC50 = 23.8 µg/ml
Inhibition of the contractile respons in rat thoracic aorta smooth muscle (Ohashi et al., 2000) Decrease of systolic blood pressure in conscious stroke-prone spontaneously hypertensive rats (Ohashi et al., 2000)
Laxative, hemorrhoid, dysentery, diarrhea, colitis Menstrual disorder, stomachic, cholecystopathy
Coriandrum sativum
fruit Essential oil Terpenoids In vitro MIC 0.87 mg/ml Inhibition of the growth of Escherichia coli, Bacillus megaterium, Pseudomonas, Erwinia, Xanthomonas, Agrobacterium (Lo Cantore et al., 2004)
Stomach-ache, vertigo, emetic, stomachic, aphtha, menstrual disorder
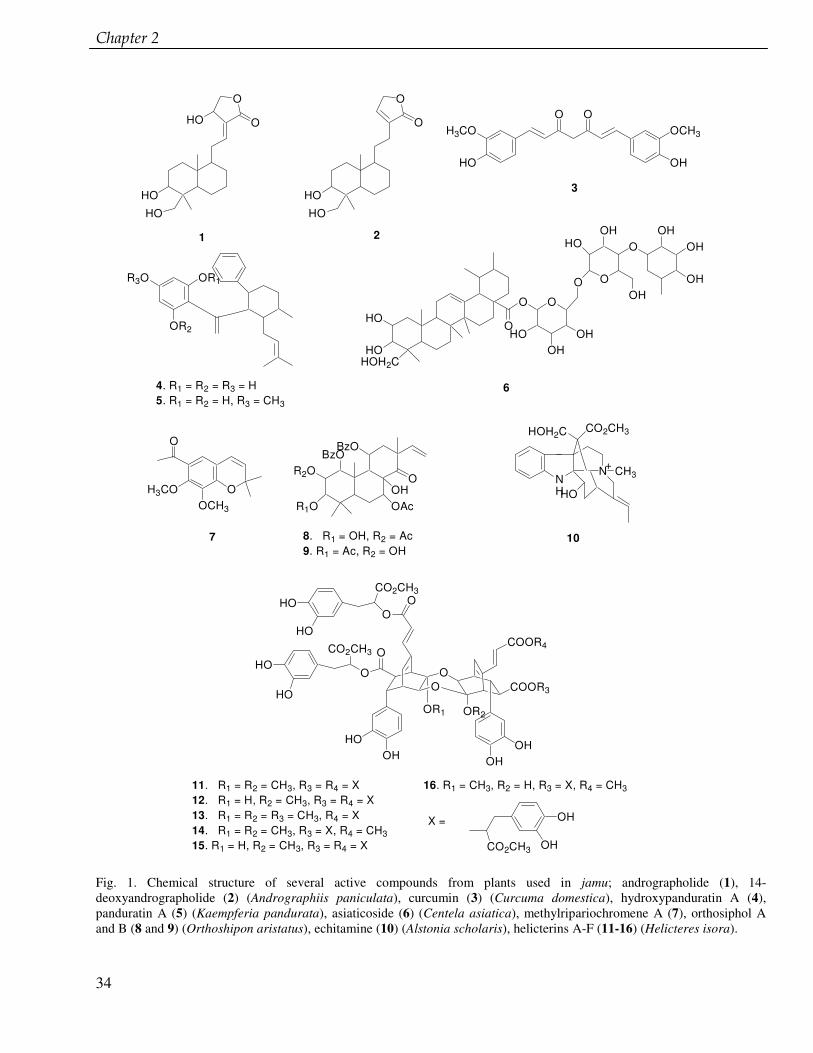
����������
34
H3CO
HO OH
OCH3
O OO
O
HO
HO
HO
O
O
HO
HO
OR1R3O
OR2
O
CO2CH3O
OO
COOR4
OR2OR1
O
CO2CH3
HO
HO
HO
HO
OHHO
OHOH
O
COOR3
CO2CH3
OH
OH
HO
HO
HOH2C
O
O
O
O O
O
HOOH
OH
HOOH
OHOH
OHOH
O
O
H3COOCH3
R2O
R1O
BzOBzO
OAcOH
O NH
N CH3
HOH2C CO2CH3
HO
3
1 2
4. R1 = R2 = R3 = H5. R1 = R2 = H, R3 = CH3
11. R1 = R2 = CH3, R3 = R4 = X12. R1 = H, R2 = CH3, R3 = R4 = X13. R1 = R2 = R3 = CH3, R4 = X14. R1 = R2 = CH3, R3 = X, R4 = CH315. R1 = H, R2 = CH3, R3 = R4 = X
16. R1 = CH3, R2 = H, R3 = X, R4 = CH3
X =
6
7 8. R1 = OH, R2 = Ac9. R1 = Ac, R2 = OH
+
10
Fig. 1. Chemical structure of several active compounds from plants used in jamu; andrographolide (1), 14-deoxyandrographolide (2) (Andrographiis paniculata), curcumin (3) (Curcuma domestica), hydroxypanduratin A (4), panduratin A (5) (Kaempferia pandurata), asiaticoside (6) (Centela asiatica), methylripariochromene A (7), orthosiphol A and B (8 and 9) (Orthoshipon aristatus), echitamine (10) (Alstonia scholaris), helicterins A-F (11-16) (Helicteres isora).

35
Chapter 3 Lignans from cell suspension cultures of Phyllanthus niruri, an Indonesian medicinal plant
Elfahmi, Sieb Batterman, Albert Koulman, Thomas Hackl, Rein Bos, Oliver Kayser, Herman J. Woerdenbag, Wim J. Quax
This chapter is based on Journal of Natural Products, 2006, 69, 55-58

����������
36
Abstract Cell suspension cultures of Phyllanthus niruri L. were used to study the lignan profiles and
biosynthesis. Suspension cultures yielded two lignans: the new dihydrocubebin-dimethylether (1) and urinatetralin (2), a new lignan from P. niruri, but reported earlier from P. urinaria. This is the first report of cell suspension cultures of P. niruri that successfully produce lignans. Feeding 0.5 mM ferulic acid or 0.5 mM caffeic acid, being early precursors of lignan biosynthesis, resulted in an increase up to 0.7 mg g-1 DW of 1 (control value 0.1 mg g-1 DW) and up to 0.3 mg g-1 DW of 2 (control value 0.2 mg g-1 DW). Comparison of the lignan profiles of cell suspensions, callus cultures, aerial plant parts, roots and seeds showed significant differences.

� ������ ����������� � ��� ����������� �������������� �������
37
Introduction Phyllanthus niruri L. (Euphorbiaceae) is a small plant widely distributed in tropical and
subtropical regions in Central and South America and Asia (including India and Indonesia). Whole plants have been used in traditional medicine for treatment of jaundice, asthma, hepatitis and malaria and because of diuretic, antiviral, and hypoglycemic properties (Eisei, 1995, Mehrota et al., 1990, Calixto et al., 1998). To scientifically support the traditional use, several pharmacological studies have been carried out with plant extracts and with isolated compounds. P. niruri extract shows potential therapeutic actions in the management of hepatitis B (Mehrota et al., 1990, Calixto et al., 1998). Its antiviral activity extends to HIV-1 RT inhibition (Ogata et al., 1992, Naik and Juvekar, 2003). Its role in urolithiasis is related to the inhibition of calcium oxalate endocytosis by renal tubular cells (Campos and Schor, 1999, Freitas et al., 2002). In vitro antiplasmodial activity of this plant extract has been described (Tona et al., 2001). P. niruri extract also showed inhibitory activities against angiotensin converting enzyme (ACE) and rat lens aldose reductase (AR), which play a significant role in the reduction of aldose to alditol under abnormal conditions such as diabetes (Shimizu et al., 1989). Immunomodulating effects in treatment of cancer by influencing the function and activity of the immune system (Ma’at, 2002) and lipid lowering activity (Khanna, 2002) have been reported. An extract of the callus culture of P. niruri showed analgesic activity (Santos et al., 1994)
The aerial parts of P. niruri have been reported to contain alkaloids, flavonoids, phenols, coumarins, tannins, terpenoids and lignans. Several of these isolated compounds have been tested for their pharmacological activities (Ishimaru et al., 1992, Calixto et al., 1998, Huang et al., 2003, Naik and Juvekar, 2003). Lignans from this plant have been studied most intensively; 17 different lignans have been found so far (Fig. 1). Several of these lignans were tested for cytotoxicity and other biological activities in vitro. Phyllanthin and hypophyllanthin were protective against carbon tetrachloride- and galactosamine-induced cytotoxicity in primary cultured rat hepatocytes (Syamasundar et al., 1985). 3,4-methylenedioxybenzyl-3�,4�-dimethoxybenzylbutyrolactone has been reported to possess antitumor activity (Satyanarayana and Venkateswarlu, 1991). Nirtetralin and niranthin were tested against human hepatitis B virus in vitro (Huang et al., 2003).
The aims of this study were to establish the cell suspension cultures of P. niruri, to study the production and biosynthesis of lignans and to compare the lignan profile of cell suspensions with callus cultures, aerial plant parts, roots and seeds.
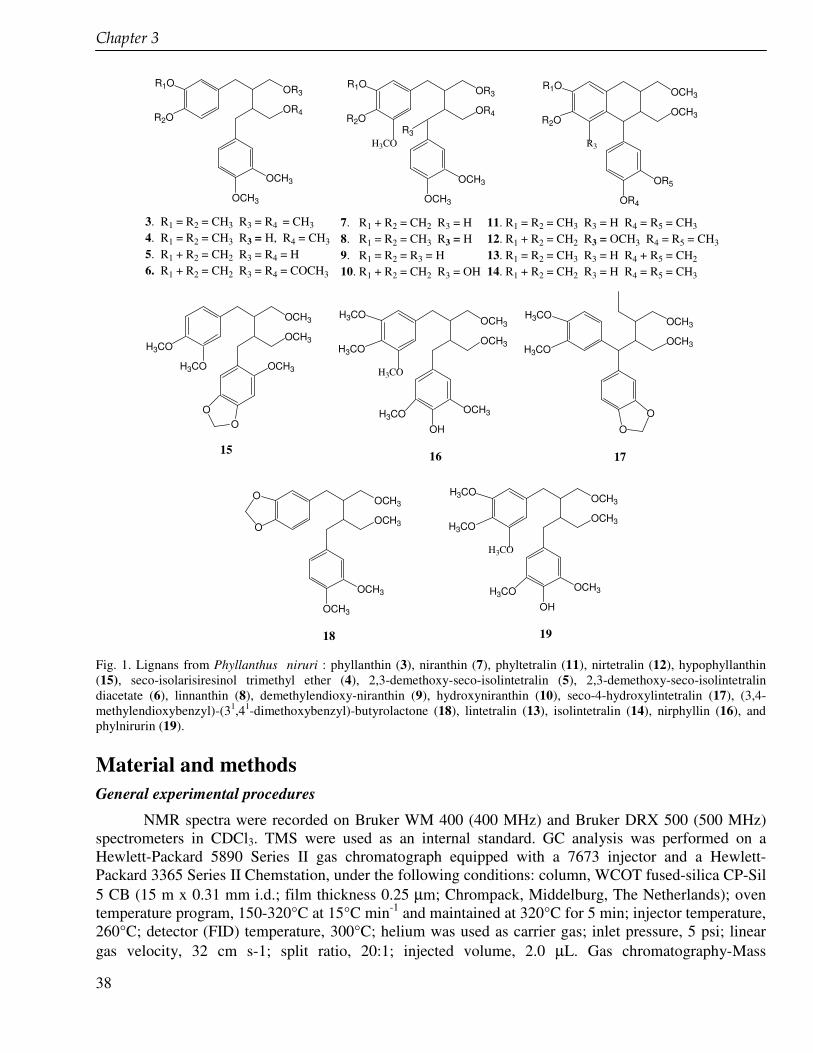
����������
38
OR4
OR3R1O
R2O
OCH3
OCH3
OR4
OR3R1O
R2O
OCH3
OCH3
R3H3CO
OCH3
OCH3R1O
R2O
OR4
OR5
R3
OCH3
OCH3
H3CO
H3CO OCH3
OO
OCH3
OCH3H3CO
H3CO
OH
OCH3H3CO
H3CO
OCH3
OCH3H3CO
H3CO
O
O
OCH3
OCH3
OCH3
OCH3
O
OOCH3
OCH3H3CO
H3CO
OH
OCH3H3CO
H3CO
3. R1 = R2 = CH3 R3 = R4 = CH3
4. R1 = R2 = CH3 R3 = H, R4 = CH3
5. R1 + R2 = CH2 R3 = R4 = H6. R1 + R2 = CH2 R3 = R4 = COCH3
7. R1 + R2 = CH2 R3 = H8. R1 = R2 = CH3 R3 = H9. R1 = R2 = R3 = H10. R1 + R2 = CH2 R3 = OH
11. R1 = R2 = CH3 R3 = H R4 = R5 = CH3
12. R1 + R2 = CH2 R3 = OCH3 R4 = R5 = CH3
13. R1 = R2 = CH3 R3 = H R4 + R5 = CH2
14. R1 + R2 = CH2 R3 = H R4 = R5 = CH3
15 16 17
18 19 Fig. 1. Lignans from Phyllanthus niruri : phyllanthin (3), niranthin (7), phyltetralin (11), nirtetralin (12), hypophyllanthin (15), seco-isolarisiresinol trimethyl ether (4), 2,3-demethoxy-seco-isolintetralin (5), 2,3-demethoxy-seco-isolintetralin diacetate (6), linnanthin (8), demethylendioxy-niranthin (9), hydroxyniranthin (10), seco-4-hydroxylintetralin (17), (3,4-methylendioxybenzyl)-(31,41-dimethoxybenzyl)-butyrolactone (18), lintetralin (13), isolintetralin (14), nirphyllin (16), and phylnirurin (19). Material and methods General experimental procedures
NMR spectra were recorded on Bruker WM 400 (400 MHz) and Bruker DRX 500 (500 MHz) spectrometers in CDCl3. TMS were used as an internal standard. GC analysis was performed on a Hewlett-Packard 5890 Series II gas chromatograph equipped with a 7673 injector and a Hewlett-Packard 3365 Series II Chemstation, under the following conditions: column, WCOT fused-silica CP-Sil 5 CB (15 m x 0.31 mm i.d.; film thickness 0.25 µm; Chrompack, Middelburg, The Netherlands); oven temperature program, 150-320°C at 15°C min-1 and maintained at 320°C for 5 min; injector temperature, 260°C; detector (FID) temperature, 300°C; helium was used as carrier gas; inlet pressure, 5 psi; linear gas velocity, 32 cm s-1; split ratio, 20:1; injected volume, 2.0 µL. Gas chromatography-Mass

� ������ ����������� � ��� ����������� �������������� �������
39
spectrometry (GC-MS) analysis was performed on a Shimadzu QP5000 GC-MS system equipped with a 17A GC, an AOC-20i autoinjector, and the GC-MS solution software 1.10. The GC conditions were the same as for GC analysis. MS conditions: ionization energy, 70 eV; ion source temperature, 250°C; interface temperature, 280°C; scan speed, 2 scans s-1; mass range, 34-600 u. The HPLC system consisted of a Spectra-Physics (model SP 8810) liquid pressure pump, a Rheodyne high pressure valve equipped with a 100 µL sample loop and a Lichrospher RP-18 column (250 x 4 mm i.d., Merck, Damstradt, Germany). The mobile phase consisted of C2H3N: H2O (45:55; v/v) containing 0.1 % H3PO4 at a flow of 1.0 mL min-1 and a Shimadzu photodiode array UV-Vis SPD-M6A detector (Shimadzu, ‘s-Hertogenbosch, The Netherlands) at 290 nm. Analytical thin layer chromatography (TLC) was performed using silica gel 60-F254 (5 x 10 cm, 0.25 mm thickness, Merck, Darmstadt, Germany) and toluene: acetone (50:1) as the eluent. The elution length was 8 cm in a saturated chamber. The bands were detected by 254 nm UV light. Plant material, solvents and chemicals
Phyllanthus niruri L. (Euphorbiaceae) was collected from a wild habitat in Bandung, West Java, Indonesia, and authenticated at the Department of Biology, Institut Teknologi Bandung (Indonesia), based on the Flora of Java (Backer and Van den Brink, 1968) A voucher specimen (HBG10PN12) is deposited in the Herbarium Bandungense. The age of the plant material was 0.5 – 1 year. All solvents and chemicals were purchased from Sigma-Aldrich (Zwijndrecht, The Netherlands). All media and supplements used to grow the callus and the suspension culture were obtained from Duchefa (Haarlem, the Netherlands). Cell suspension cultures of P. niruri
Seeds of P. niruri were grown in a plant container under an L/D regime (16/8 h: 3,000 lux) at 25°C. Small plants developed after 3-4 weeks. The leaves were sterilized by dipping them into a 3% w/v NaOCl solution for 3 min, followed by bathing in sterile H2O for 10 min. They were rinsed three times with sterile twice-deionized H2O. The sterile leaves were cut into slices and callus induction was obtained using media with different compositions. These media were modifications of the Murashige and Skoog medium (Murahige and Skoog, 1962) or the Gamborg’s B5 medium (Gamborg et al., 1968). Murashige and Skoog (MS) medium was supplemented with 1.0 mg L-1 indole-3-acetic acid and 1.0 mg L-1 6-benzylaminopurin. Gamborg’s B5 (B5) was supplemented with 4 mg L-1 �-naphtalene acetic acid. The so-called MS-B5, a combination of the macro nutrients of MS and the micro nutrients of B5 medium, was also used. It was supplemented with 3 mg L-1 �-naphtalene acetic acid. All media were supplemented with 4 % (w/w) sucrose and solidified with 0.9% agar. The callus cultures were grown under an L/D regime (16/8 h: 3,000 lux) at 26°C. Callus developed after 6 weeks. Friable callus appeared as clumps, varying in color from dark brown to yellow.
Cell suspension cultures were initiated by transferring callus clumps into 100 mL sterile conical flasks with 50 mL of liquid medium of the same composition as described above but without agar. Cultures were incubated on a rotary shaker (175 rpm) at 26°C under an L/D regime (16/8 h: 3,000 lux, daylight L 36W/10, OSRAM, Germany). After 1 month, 50 mL of cell suspension cultures were transferred into a 500 mL conical flask, with fresh medium yielding a total volume of 300 mL. Subcultures were prepared every 3 weeks by adding 100 mL of a full-grown cell suspension culture to 200 mL of fresh medium. Feeding experiment
Caffeic and ferulic acid were added to reach three different final concentrations: 0.1, 0.5 and 1 mM. These compounds were dissolved using 0.2 mL of 96% EtOH and transferred to a sterile 500 mL

����������
40
Erlenmeyer flask containing the cell suspension cultures of P. niruri after inoculation into fresh medium (at the start of growth cycle). Suspension-grown cells were harvested each 2 days during the growth cycle of 21 days. Samples of about 10 mL were taken aseptically and transferred into a calibrated conical tube and centrifuged for 5 min at 1,500 g. To monitor the viability and growth of the cell cultures, medium pH and conductance were routinely measured in the supernatant. Cells were filtered using a Büchner funnel. FW was determined and the cells were put overnight into the freezer and then freeze-dried. DW was also determined. Lignans were analyzed by GC and GC-MS. Analysis
Lignan profiling was performed according to Koulman et al. (2001). Dried and powdered material (100 mg) was weighed in a Sovirel tube. MeOH (2 mL, 80%) was added and the mixture was sonicated for 1 h. CH2Cl2 (4 mL) and distilled water (4 mL) were added. The tube was closed, vortexed and centrifuged at 1,500 g for 5 min. The aqueous layer was discarded and 2 mL of the organic layer was transferred into a 2 mL micro tube. The CH2Cl2 was evaporated gently using a flow of nitrogen and the residue was redissolved in 200 µL of MeOH. The tube was centrifuged at 15,000 g for 5 min in an Eppendorf 5414 centrifuge, and part of the liquid was transferred into a 0.8 mL crimp neck vial (Art. No. 98819, Alltech/Applied Science B.V. Breda, The Netherlands) and sealed immediately. For quantitative determination of 1 and 2, calibration curves were prepared using MeOH solutions with concentrations ranging from 2.5 to 25 µg mL-1. Each concentration was done in triplicate. Regression equations were y = 143229 x – 264 (R2 = 0.9902) and y = 123516 x – 218 (R2 = 0.9885) for 1 and 2. The limit of detection (LOD) was established as the amount of analyte that provided a signal-to-noise ratio of 3. LOD were 0.12 µg for 1 and 0.25 for 2. The limit of quantification (LLOQ) was defined as the lowest calibration standard that could be quantified with an accuracy of 90-110% and a precision of 15%. LLOQ was 2.5 µg for both 1 and 2. Extraction and isolation
For the isolation of lignans, three-weeks-old suspension cultures were harvested and filtered through miracloth (Lot B43936, Calbiochem, Behring diagnostics, La Jolla, CA). A total of 500 g of fresh cells were extracted (3x) using 1 L of 80% MeOH each time. To disrupt cells, mixtures were sonicated for 1 h. The MeOH extracts were combined and the volume was reduced to 100 mL under reduced pressure. The remaining extract was partitioned (3x) between 250 mL of H2O and 250 mL of CH2Cl2. All organic phases were combined and concentrated. The resulting extract was fractionated on a column (75 x 2.5 cm) filled with silica gel 60 (70-230 mesh, Merck, Darmstadt, Germany). Fractions were eluted by subsequent use of the following eluents: n-hexane: CH2Cl2 (9:1, 500 mL), CH2Cl2 (500 mL), and CH2Cl2: MeOH (25:1, 500 mL). Fractions of 20 mL each were collected and were profiled by TLC and GC-MS. Fractions with corresponding profiles were combined and concentrated. This resulted in a total of 16 fractions. The lignan-containing fraction was further separated by preparative TLC on preparative silica gel plates (20x20 cm, 1 mm thickness, Merck, Darmstadt, Germany) using toluene: acetone (50:1) as eluent. The elution length was 18 cm in a saturated chamber. Bands were detected by 254 nm UV light. Three bands were scraped off and analyzed by GC and GC-MS. Rf values were 0.28, 0.31 and 0.43, respectively.
For further purification of the lignans obtained by preparative TLC, semipreparative HPLC was used. Eluting substances were collected in glass tubes according to their retention times. Organic solvents were evaporated and the aqueous residue was partitioned with CH2Cl2. The CH2Cl2 fraction yielded pure compound 1 (3.0 mg) and 2 (2.5 mg).

� ������ ����������� � ��� ����������� �������������� �������
41
Dihydrocubebin-dimethylether (1) was obtained as an amorphous solid: 1H and 13C NMR data, see Table 1; MS (EI, 70 eV) m/z (rel. int.): 386 [M]+(2), 354(3), 322 (2), 187 (19), 135 (100), 77 (29), 45 (56); HRMS: m/z = 386.1723 [M+] (calc. for C22H26O6: 386.1729).
Urinatetralin (2) was obtained as an amorphous solid: 1H and 13C NMR data, see Table 1; MS (EI, 70 eV) m/z (rel. int.): 384 [M]+(5), 352(6), 320 (7), 185 (12), 135 (26), 77 (8), 45 (100) Results and discussion
From the various medium compositions tested, the B5 medium resulted in the best growth of callus and cell suspension cultures of P. niruri. This medium was used in all experiments. Cell suspension cultures of P. niruri had a growth cycle of 21 days. During this time, conductivity decreased from 3.2 ± 0.01 mS on day 1 to 1.5 ± 0.02 mS on day 15 and slightly increased to 2.0 mS on day 21. This indicates that cell lysis occurred between day 15 and 21. During the growth cycle, a slow increase in pH of the growth medium was observed. Fresh weight (FW) increased from 105.2 ± 3.2 g L-1 to 202.8 ± 19.2 g L-1 and dry weight (DW) increased from 6.0 ± 1.1 g L-1 to 12.9 ± 0.1 g L-1.
We found qualitatively and quantitatively different lignan profiles comparing seeds, roots, aerial plant parts, callus and suspension cultures as determined by GC-MS. All lignans found in aerial plant parts (phyllantin, 3,4-methylenedioxybenzyl-3�,4�-dimethoxybenzylbutyrolactone, niranthin, phyltetralin, seco-isolarisiresinol trimethylether, nirtetralin, lintetralin, isolintetralin, hypophyllantin, seco-4-hydroxylintetralin), seed (phyllantin, niranthin and nirtetralin), root (demethylenedioxyniranthin, lintetralin and isolintetralin) and callus culture (3,4-methylenedioxybenzyl-3�,4�-dimethoxybenzylbutyrolactone) have been described previously in P. niruri (Row and Srinivasalu, 1964, Anjaneyulu et al., 1973, Satyanarayana and Venkateswarlu, 1991, Calixto et al., 1998). None of these lignans was present in cell suspension cultures. However, four other lignans were found in cell suspension cultures. Compound 1 was a lignan that was not found in the plants before and compound 2 has been reported earlier as new compound from P. urinaria (Chang et al., 2003). Two more compounds with MS fragmentation patterns resembling lignans were found. These last two compounds could not be separated. Compounds 1 and 2 were purified by preparative HPLC and their structures determined and confirmed (Fig. 2). This is another example that plant cell cultures do not always accumulate, either qualitatively or quantitatively, the same compounds found in the plant from which they are established.
Compound 2 was identified as urinatetralin that was confirmed by comparison of NMR and MS spectra with published data (Chang et al., 2003). The relative configuration derived from the 1H NMR coupling constants was in agreement with the (7�R*,8S*,8�S*) configuration of 2 from P. urinaria (Chang et al., 2003).
Compound 1 was a white amorphous solid with a molecular formula of C22H26O6, as shown by HRMS ([M+]: m/z 386.1723). The molecular mass of 1 is two mass units higher than that of compound 2. Fragments at m/z 354 and 322 correspond to the loss of two molecules of MeOH [M-HOCH3], as shown by HRMS. The base peak at m/z 135 corresponded to a methylenedioxy benzyl fragment (C8H7O2), typical for lignan structures. These data indicated an urinatetralin-like structure for 1, i.e. lacking the carbocylic ring at C-2 and C-7�. The 1H NMR data exhibited two methylenedioxy groups at �H 5.92 (4H, s), two methoxy groups at �H 3.28 (6H, s) and six aromatic protons (�H 6.5 – 6.7) as two ABM spin systems. Due to the symmetry of the molecule only one set of signals is observed. The structure for 1 was further supported by H,H-COSY, HMQC and HMBC spectroscopic analysis. This compound, called dihydrocubebin-dimethylether, is a new natural product but has been synthesized from dihydrocubebin (Anjaneyulu et al., 1981). Although the relative configuration could not be determined unambigously, a (8S*,8'S*) configuration is assumed from a common biogenesis of 1 and 2. This is the first report of cell suspension cultures of P. niruri that successfully produce lignans.

����������
42
O
O
OO
OCH3
OCH3O
O
OO
OCH3
OCH3
1 2
1
23
45
6
7
7'
1'2'
5'
6'
3'
4'
8
8'
9
9'
1
23
4
56 7
7'
1'2'
3'
4'5'
6'
8
8'
9
9'
Fig. 2. Lignans from cell suspension cultures of P. niruri: dihydrocubebin-dimethylether (1) and urinatetralin (2).
Table 1. 1H and 13C NMR data of dihydrocubebin-dimethylether (1) and urinatetralin (2) (500 MHz, TMS, CDCl3).
Position 1 2
1H (J/Hz) 13C 1H (J/Hz) 13C 1 134.9 s 130.1 s 2 6.7 d (7.8) 121.8 d 133.4 s 3 108.1 d 6.21 s 109.5 d 4 145.6 s 145.81 s 5 6.59 d (1.3) 147.5 d 145.81 s 6 6.56 dd (7.9, 1.6) 109.5 d 6.56 s 108.0 d 7 3.26 m, 3.30 m 35.1 t 2.78 m 33.8 t 8 2.02 m 40.8 d 2.13 m 36.4 d 9 2.55 dd (13.9, 8.0) 72.7 t 3.40 dd (6.5, 9.3) 75.5 t 2.65 dd (13.5, 5.9) 3.45 dd (4.0, 9.3) 1' 134.9 s 139.4 s 2' 6.59 d (1.3) 109.5 d 6.55 d (1.7) 108.6 d 3' 147.5 s 146.4 s 4' 145.6 s 147.9 s 5' 6.7 d (7.8) 108.1 d 6.74 d (7.9) 107.6 d 6' 6.56 dd (7.85, 1.55) 121.8 d 6.64 dd (7.9, 1.7) 122.6 s 7' 3.26 m, 3.30 m 35.1 t 3.93 d (10.8) 47.7 d 8' 2.02 m 40.8 d 1.78 ddt (10.8, 10.8, 3.2) 45.1 d 9' 2.55 dd (13.9, 8.0) 72.7 t 3.10 dd (9.6, 3.4) 71.2 t 2.65 dd (13.5, 5.9) 3.36 m 9-O Me 3.28 s 58.8 q 3.35 s 59.0 q 9'-O Me 3.28 s 58.8 q 3.26 s 59.1 q 4,5-OCH2O- 5.92 s 100.7 t 5.83 s 99.8 t 3',4'-OCH2O- 5.92 s 100.7 t 5.93 s 100.2 t
1 signals coincide.
Feeding of early precursors of lignans to cell suspension cultures, 0.5 mM caffeic acid or 0.5 mM ferulic acid, enhanced the production of 1 and 2. While feeding 0.1 mM of either compounds had no effect, 1 mM appeared to be toxic for cell suspension cultures (causing growth inhibition). In the control cells, 1 reached a maximum at 0.46 mg g-1 DW on day 1 after inoculation, decreased to 0.18 mg
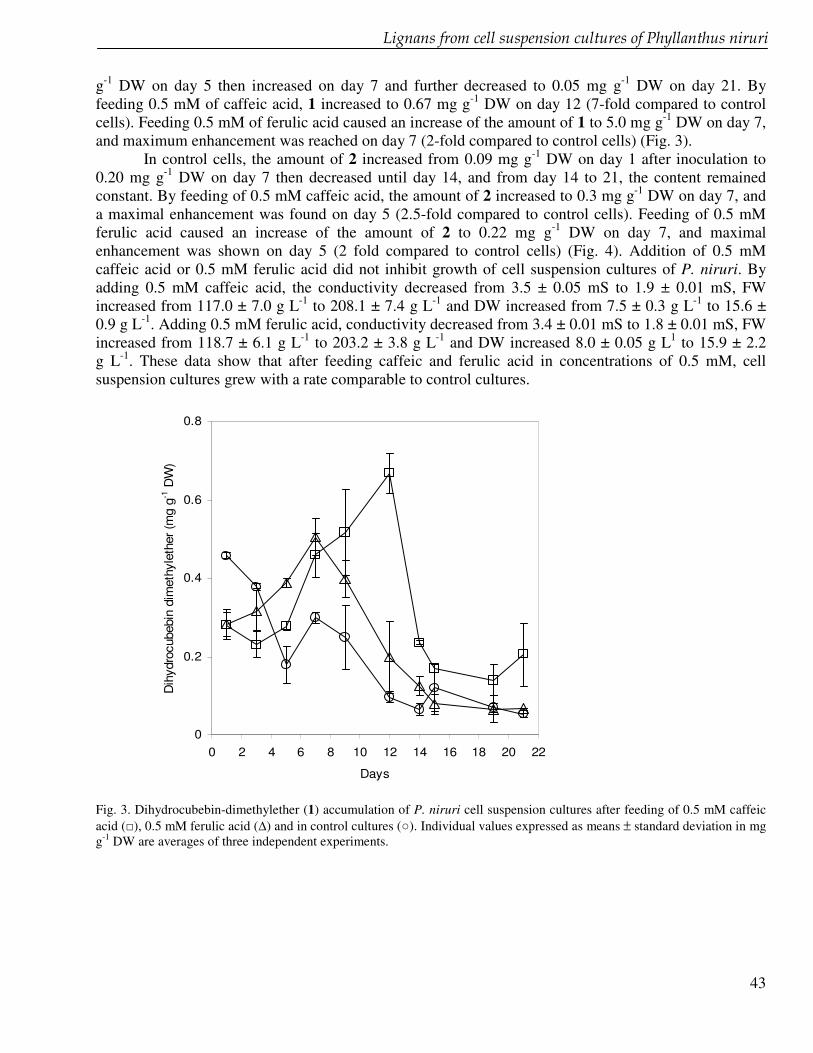
� ������ ����������� � ��� ����������� �������������� �������
43
g-1 DW on day 5 then increased on day 7 and further decreased to 0.05 mg g-1 DW on day 21. By feeding 0.5 mM of caffeic acid, 1 increased to 0.67 mg g-1 DW on day 12 (7-fold compared to control cells). Feeding 0.5 mM of ferulic acid caused an increase of the amount of 1 to 5.0 mg g-1 DW on day 7, and maximum enhancement was reached on day 7 (2-fold compared to control cells) (Fig. 3).
In control cells, the amount of 2 increased from 0.09 mg g-1 DW on day 1 after inoculation to 0.20 mg g-1 DW on day 7 then decreased until day 14, and from day 14 to 21, the content remained constant. By feeding of 0.5 mM caffeic acid, the amount of 2 increased to 0.3 mg g-1 DW on day 7, and a maximal enhancement was found on day 5 (2.5-fold compared to control cells). Feeding of 0.5 mM ferulic acid caused an increase of the amount of 2 to 0.22 mg g-1 DW on day 7, and maximal enhancement was shown on day 5 (2 fold compared to control cells) (Fig. 4). Addition of 0.5 mM caffeic acid or 0.5 mM ferulic acid did not inhibit growth of cell suspension cultures of P. niruri. By adding 0.5 mM caffeic acid, the conductivity decreased from 3.5 ± 0.05 mS to 1.9 ± 0.01 mS, FW increased from 117.0 ± 7.0 g L-1 to 208.1 ± 7.4 g L-1 and DW increased from 7.5 ± 0.3 g L-1 to 15.6 ± 0.9 g L-1. Adding 0.5 mM ferulic acid, conductivity decreased from 3.4 ± 0.01 mS to 1.8 ± 0.01 mS, FW increased from 118.7 ± 6.1 g L-1 to 203.2 ± 3.8 g L-1 and DW increased 8.0 ± 0.05 g L1 to 15.9 ± 2.2 g L-1. These data show that after feeding caffeic and ferulic acid in concentrations of 0.5 mM, cell suspension cultures grew with a rate comparable to control cultures.
0
0.2
0.4
0.6
0.8
0 2 4 6 8 10 12 14 16 18 20 22
Days
Dih
ydro
cube
bin
dim
ethy
leth
er (m
g g-1
DW
)
Fig. 3. Dihydrocubebin-dimethylether (1) accumulation of P. niruri cell suspension cultures after feeding of 0.5 mM caffeic acid (�), 0.5 mM ferulic acid (�) and in control cultures (�). Individual values expressed as means ± standard deviation in mg g-1 DW are averages of three independent experiments.
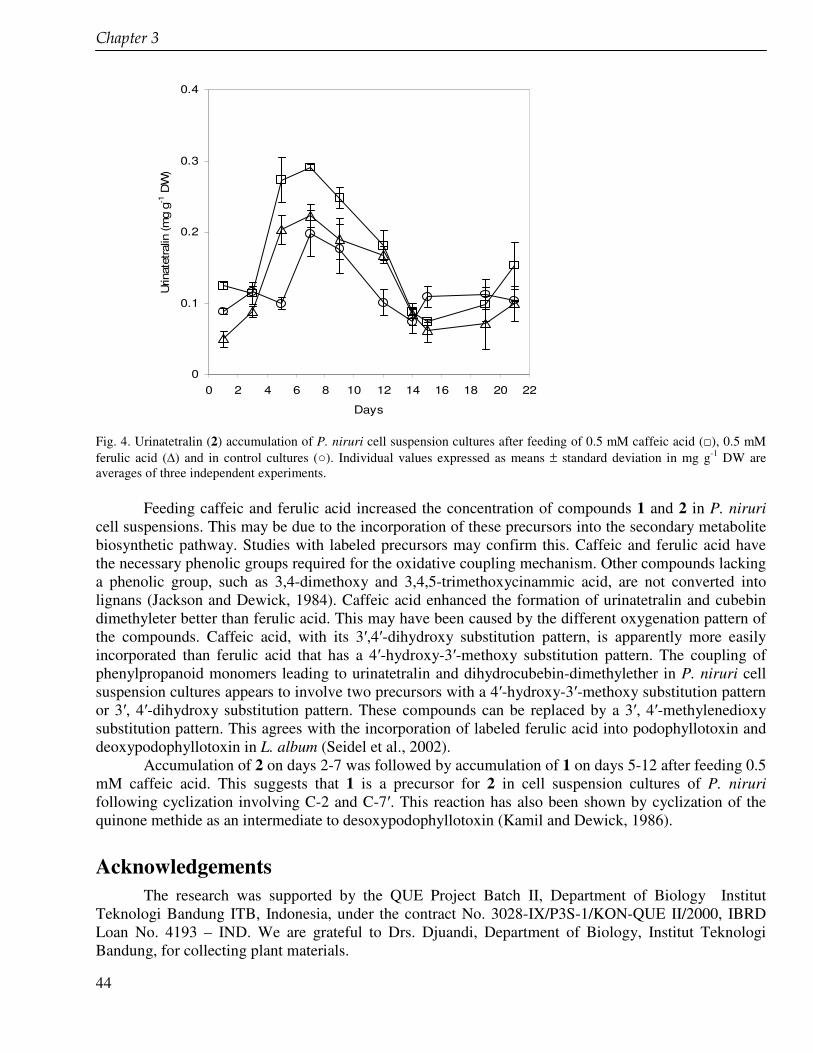
����������
44
0
0.1
0.2
0.3
0.4
0 2 4 6 8 10 12 14 16 18 20 22
Days
Urin
atet
ralin
(mg
g-1 D
W)
Fig. 4. Urinatetralin (2) accumulation of P. niruri cell suspension cultures after feeding of 0.5 mM caffeic acid (�), 0.5 mM ferulic acid (�) and in control cultures (�). Individual values expressed as means ± standard deviation in mg g-1 DW are averages of three independent experiments.
Feeding caffeic and ferulic acid increased the concentration of compounds 1 and 2 in P. niruri
cell suspensions. This may be due to the incorporation of these precursors into the secondary metabolite biosynthetic pathway. Studies with labeled precursors may confirm this. Caffeic and ferulic acid have the necessary phenolic groups required for the oxidative coupling mechanism. Other compounds lacking a phenolic group, such as 3,4-dimethoxy and 3,4,5-trimethoxycinammic acid, are not converted into lignans (Jackson and Dewick, 1984). Caffeic acid enhanced the formation of urinatetralin and cubebin dimethyleter better than ferulic acid. This may have been caused by the different oxygenation pattern of the compounds. Caffeic acid, with its 3�,4�-dihydroxy substitution pattern, is apparently more easily incorporated than ferulic acid that has a 4�-hydroxy-3�-methoxy substitution pattern. The coupling of phenylpropanoid monomers leading to urinatetralin and dihydrocubebin-dimethylether in P. niruri cell suspension cultures appears to involve two precursors with a 4�-hydroxy-3�-methoxy substitution pattern or 3�, 4�-dihydroxy substitution pattern. These compounds can be replaced by a 3�, 4�-methylenedioxy substitution pattern. This agrees with the incorporation of labeled ferulic acid into podophyllotoxin and deoxypodophyllotoxin in L. album (Seidel et al., 2002).
Accumulation of 2 on days 2-7 was followed by accumulation of 1 on days 5-12 after feeding 0.5 mM caffeic acid. This suggests that 1 is a precursor for 2 in cell suspension cultures of P. niruri following cyclization involving C-2 and C-7�. This reaction has also been shown by cyclization of the quinone methide as an intermediate to desoxypodophyllotoxin (Kamil and Dewick, 1986). Acknowledgements
The research was supported by the QUE Project Batch II, Department of Biology Institut Teknologi Bandung ITB, Indonesia, under the contract No. 3028-IX/P3S-1/KON-QUE II/2000, IBRD Loan No. 4193 – IND. We are grateful to Drs. Djuandi, Department of Biology, Institut Teknologi Bandung, for collecting plant materials.

45
Chapter 4 Lignan profile of Piper cubeba, an Indonesian medicinal plant Elfahmi, Komar Ruslan, Sieb Batterman, Rein Bos, Oliver Kayser, Herman J. Woerdenbag, Wim J. Quax Submitted

�������� �
46
Abstract The lignan profiles of aerial part of Piper cubeba L. (Piperaceae) was determined using GC, GC-
MS and HPLC. The number of lignans found in the leaves was 15, followed by berries and the stalks with respectively 13 and 5 lignans. This is the first investigation of lignans from the leaves and the stalks of P. cubeba. Cubebin, hinokinin, yatein, isoyatein are common lignans in the genus Piper and appeared as major components in all part of P. cubeba investigated.

� � ������������������������������
47
Introduction Piper cubeba L. (Piperaceae) inhabits Java, Sumatra, Southern Borneo, and other isles in the
Indian Ocean. The berries of P. cubeba are commonly known as cubeb (in Indonesia known as kemukus) and used in Indonesian traditional medicine to treat gonorrhea, dysentery, syphilis, abdominal pain, diarrhea, enteritis and asthma (Eisai, 1995, Satroamidjojo, 2001). Phytochemical and biological investigations have been carried out in order to prove its traditional use. In comparison to other species of genus Piper, P. cubeba has received less attention so far. Only three groups of secondary metabolites have been reported from the berries of P. cubeba, i.e. alkaloids, lignans and terpenoids (essential oil). The lignans and the essential oil have been more intensively investigated, since P. cubeba accumulates both groups of compounds in relatively high amounts. Economically, P. cubeba is important as a source of pepper (the dried berries) for the worldwide spice market (Usia et al., 2005). Piperine is an abundant alkaloid in the berries of this species (Parmar et al., 1997).
Twenty four lignans have so far been reported from P. cubeba (Fig. 1) (Prabhu and Mulchandani, 1985, Badheka et al., 1986, 1987, Koul et al., 1996, Parmar et al., 1997, Usia et al,. 2005). Lignans are an important group of secondary metabolites that are also assumed to contribute to the biological activity. Some of these lignans showed inhibitory activity against cytochrome P450 enzymes that are involved in the metabolism of all currently used drugs (Usia et al., 2005a, 2005b). Yatein, hinokinin, cubebin, dihydrocubebin have been reported to have antifeedant activity against a number of stored product insects. This activity is comparable to podophyllotoxin (Harmatha and Nawrot, 2002). Hinokinin has been reported to have anti-inflammatory and analgesic effect. Because of the structural relationship, hinokinin can be synthesized using cubebin as precursor (Da Silva et al. 2005). Cubebin has been shown to possess anti-inflamatory, analgesic and trypanocidal activities (Borsato et al., 2000, Bastos et al., 2001, De Souza et al., 2005). Yatein is also an interesting lignan due to its biological activity and its function as a biosynthetic precursor of deoxypodophyllotoxin and podophyllotoxin that are well known for their anticancer properties. Methanol and water extract of P. cubeba berries have been shown to display an inhibitory effect against the hepatitis C virus (Hussein et al., 2000). Anti-inflammatory, antioxidant, anti-allergic and analgesic activities of P. cubeba have been studied using chemically-induced edema and arthritis in vivo (Choi and Hwang, 2003 and 2005).
In this study we made a lignan profile of aerial parts of P. cubeba using GC, GC-MS and HPLC and compared the data from berries, stalks and leaves. Materials and methods Plant material, solvents and chemicals
Piper cubeba L. (Piperaceae) was collected in April 2002 from Jatiroto, Temanggung, Central Java, Indonesia, and authenticated at the Department of Biology, Institut Teknologi Bandung (Indonesia), based on the Flora of Java (Backer and Van de Brink, 1968). A voucher specimen (HBG10PC01) is deposited at the Herbarium Bandungense, Indonesia. The collected plant material was air-dried. All solvents and chemicals were purchased from Sigma-Aldrich (Zwijndrecht, The Netherlands). Extraction and isolation
One gram of dried plant material (either berries, leaves or stalks) was grinded together with quartz (Merck, Darmstadt, Germany) and extracted under sonification (1 h) using 2 mL of 80% MeOH. The resulting extract was fractionated using 4.0 mL of CH2Cl2 and 4 mL of water, shaken for 5 s and centrifuged at 1,500 g for 5 min. For the determination of lignans, 2.0 mL of CH2Cl2 phase were taken and evaporated to dryness. The residue was redissolved in 1.0 mL methanol and centrifuged. These

�������� �
48
samples were ready to be analyzed by GC, GC-MS, HPLC. The isolation of lignans was achieved by preparative thin layer chromatography (TLC) using silica gel F254 plate (Merck, Germany) and toluene : acetone (85 : 15 v/v) as eluent. Bands corresponding to lignans were scraped off and extracted with 2 mL MeOH. The methanolic solutions were submitted to further analysis. The most abundant lignans, cubebin, hinokinin and yatein, were isolated as >95% pure compounds (as checked by GC and HPLC) and used for quantitative purposes. Analysis of lignans using GC, GC-MS, HPLC, TLC
Gas chromatography (GC) analysis was performed on a Hewlett-Packard 5890 Series II gas chromatograph equipped with a 7673 injector and a Hewlett Packard 3365 Series II Chemstation, under the following conditions: column, WCOT fused-silica CP-Sil 5 CB (15 m x 0.31 mm i.d.; film thickness 0.25 µm; Chrompack, Middelburg, The Netherlands); oven temperature program, 150-320°C at 15°C min-1 and maintained at 320°C for 5 min; injector temperature, 260°C; detector (FID) temperature, 300°C; helium was used as carrier gas; inlet pressure, 5 psi; linear gas velocity, 32 cm s-1; split ratio, 20:1; injected volume, 2.0 µL.
GC-MS analysis was performed on a Shimadzu QP5000 GC-MS system equipped with a 17A GC, an AOC-20i autoinjector, and the GCMS solution software 1.10. GC conditions: WCOT fused silica CPsil 5 CB lowbleed/MS, CP7810, film thickness 0.10 �m; 15 m x 0.25 mm ID (Varian Middelburg, The Netherlands). Temperature program: 150°C – 320 °C at 15 °C min-1. Injector temperature: 275°C; inlet pressure: 75 kPa; column flow: 2.1 mL min-1: linear velocity: 75.5 cm sec-1; split ratio: 20:1; total flow: 46.2 mL min-1; carrier flow (He): 46.2 mL min-1; injection volume: 2 µL; temperature program 35 min at 90 °C, 90-170 °C at 4 °C min-1. MS conditions: ionization energy, 70 eV; ion source temperature, 250°C; interface temperature, 250°C; scan speed, 2 scans s-1; mass range, 34-600 u
HPLC analysis was performed using a Shimadzu-VP system (Shimadzu, ‘s-Hertogenbosch, The Netherlands) consisting of a LC-10AT vp pump, a Kontron 360 auto sampler, a SPD-M10A vp DAD detector (200 – 340 nm, band with: 4 nm) , a FCV-10AL vp low pressure gradient mixer, a SCL-10A vp system controller, a FIAtron systems CH-30 column heater (USA), and CLASS-VP software, version 6.12SP4. ProFill 25 mm syringe HPLC filters, nature, PTFE, pore size 0.45 µm, part nr. AF223471- 13208 (Alltech/Applied Science Group, Breda) were used, together with 2 mL autosampler vials, cat. nr. 151123; Brown Chromatography Supplies). As crimp seals, 11 mm with rubber/PTFE septa (cat. nr. 151216; Brown Chromatography Supplies) were used.
The column was a Luna C18(2), 250 x 4.6 mm, 5µm, 00G-4252-E0, nr.: 279423-28, together with a Phenomenex guard cartridge C18 (ODS, 4x3 mm), AJ0-4287. (Phenomenex, Bester, the Netherlands). The injection volume was 20 �L with a flow rate of 1 mL min-1 using a time program of 30 minutes consisting of 5 min 95% solvent A (5 mM ammonium formate (0.05% formic acid) : acetonitrile = 800 : 156 (w/w)) and 5% solvent B (acetonitrile : MeOH (0.05% formic acid) : 5 mM ammonium formate (0.05% formic acid) = 585 : 40 : 200 (w/w)), followed by a gradient to 100% solvent B in 19 min, 2 min 100% solvent B, a gradient back to 5 % solvent B in 2 min and remaining on this final concentration for another 2 min, based on a method described by Walsky and Obach (2004)
Analytical thin layer chromatography (TLC) was performed using silica gel 60-F254 (5 x 10 cm, 0.25 mm thickness, Merck, Darmstadt, Germany) and toluene: acetone (85:15) as the eluent. The elution length was 8 cm in a saturated chamber. The bands were detected by UV light at 254 nm.
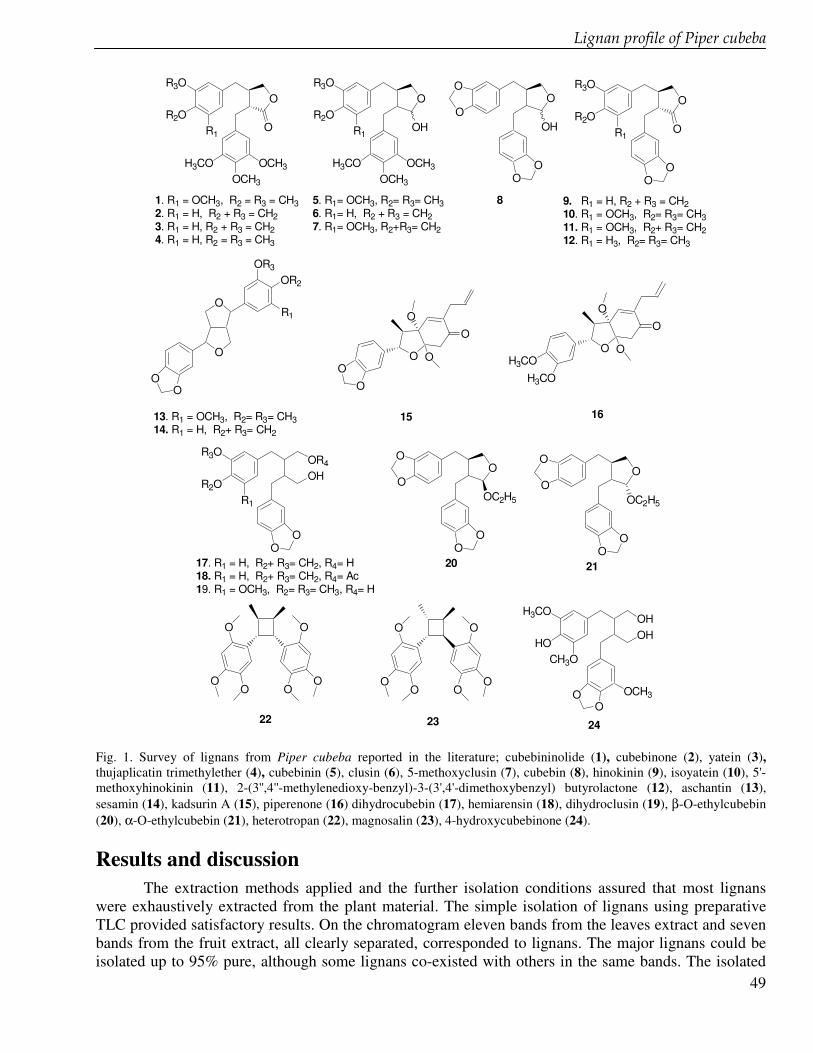
� � ������������������������������
49
O
H3COOCH3
OCH3
R3O
R2OR1
O
O
H3COOCH3
OCH3
R3O
R2OR1
OH
O
O
OO
OR3
OR2
R1
R3O
R2OR1
OO
OHOR4 O
OO
O
OOC2H5
O
OO
O
OOC2H5
O
OO
OH
O
O
OO
O
OO
O
OO
O
OO
OH3CO
HOCH3O
OHOH
OO
OCH3
OR3O
R2OR1
OO
O
H3CO
H3CO
O
O
O
OO
O
O
OO
O
1. R1 = OCH3, R2 = R3 = CH32. R1 = H, R2 + R3 = CH2 3. R1 = H, R2 + R3 = CH24. R1 = H, R2 = R3 = CH3
5. R1= OCH3, R2= R3= CH36. R1= H, R2 + R3 = CH27. R1= OCH3, R2+R3= CH2
13. R1 = OCH3, R2= R3= CH314. R1 = H, R2+ R3= CH2
15
17. R1 = H, R2+ R3= CH2, R4= H18. R1 = H, R2+ R3= CH2, R4= Ac19. R1 = OCH3, R2= R3= CH3, R4= H
20 21
16
22 23
9. R1 = H, R2 + R3 = CH210. R1 = OCH3, R2= R3= CH311. R1 = OCH3, R2+ R3= CH212. R1 = H3, R2= R3= CH3
8
24 Fig. 1. Survey of lignans from Piper cubeba reported in the literature; cubebininolide (1), cubebinone (2), yatein (3), thujaplicatin trimethylether (4), cubebinin (5), clusin (6), 5-methoxyclusin (7), cubebin (8), hinokinin (9), isoyatein (10), 5'-methoxyhinokinin (11), 2-(3'',4''-methylenedioxy-benzyl)-3-(3',4'-dimethoxybenzyl) butyrolactone (12), aschantin (13), sesamin (14), kadsurin A (15), piperenone (16) dihydrocubebin (17), hemiarensin (18), dihydroclusin (19), β-O-ethylcubebin (20), α-O-ethylcubebin (21), heterotropan (22), magnosalin (23), 4-hydroxycubebinone (24). Results and discussion
The extraction methods applied and the further isolation conditions assured that most lignans were exhaustively extracted from the plant material. The simple isolation of lignans using preparative TLC provided satisfactory results. On the chromatogram eleven bands from the leaves extract and seven bands from the fruit extract, all clearly separated, corresponded to lignans. The major lignans could be isolated up to 95% pure, although some lignans co-existed with others in the same bands. The isolated

�������� �
50
lignans were used to identify lignan peaks in the GC, GC-MS and HPLC chromatograms. Mass spectra of the analyzed lignans were compared to earlier published data (see Table 2). Different parts of P. cubeba (leaves, berries, stalks) showed different lignan profiles (see Table 2) and identification took place based on comparison of the mass fragmentation patterns. Thirteen lignans were detected in the berries, fifteen in the leaves and only five lignans in the stalks. The stereochemsitry of these compounds surveyed in Fig. 1 was taken from the literature. Mass spectral data can not be used to determine the stereochemistry of the isolated compounds.
The structures of the lignans found in P. cubeba show a broad variation. All types of lignan structures that are commonly distinguished could be identified. Furanofuran lignans such as cubebin, hinokinin, yatein, isoyatein that are common lignans found in the genus Piper also appeared as major lignans in all parts of P. cubeba. Neolignans with an unusual structure such as kadsurin A and piperenone that have also been reported from P. cubeba (Koul et al., 1996) could not be identified in our material. Yatein was the most abundant lignan found in the berries at concentrations 2.0 times higher than hinokinin and 1.3 times higher than cubebin. Hinokinin was the most abundant lignan in the leaves and the seeds (see Table 1).
For the quantitative determination of cubebin, hinokinin and yatein, calibration curves were prepared using MeOH solutions of the isolated lignans at concentrations ranging from 2.5 to 25 µg mL-1. Each concentration was prepared in triplicate. Regression equations were y = 123516 x – 218 (R2 = 0.9885), y = 117773x - 6397.4 (R2 = 0.997) and y = 111833x - 558.67 (R2 = 0.995) for cubebin, hinokinin and yatein, respectively. The limit of detection (LOD) was established as the amount of analyte that provided a signal-to-noise ratio of 3. LOD were 0.1 µg for cubebin, and 0.2 for hinokinin and 0.25 for yatein. The limit of quantification (LLOQ) was defined as the lowest calibration standard that could be quantified with an accuracy of 90-110% and a precision of 15%. LLOQ was 0.3 µg for cubebin, 0.9 µg for hinokinin and 0.1 µg for yatein
So far, phytochemical and pharmacological studies have only been done with the berries of P. cubeba that are used in traditional medicine, including jamu. Based on the present inventory of lignans in the aerial parts of the plant we conclude that in addition to the berries also the leaves may be used for medical purposes. This knowledge can be used for the further development of (rationally designed) phytomedicines from P. cubeba. It might be attractive from an economical point of view to harvest the leaves earlier, in a stage in which the plant has not yet developed flowers and later berries. This may be of economical benefit. It should however be taken into account that the concentrations of the major lignans in the leaves are considerably lower than in the berries.
Table 1. Concentration of major lignans in Piper cubeba berries, leaves, and stalks (means from three independent determinations).
Concentration (mg DW-1) Compound
Berries (± SD) Leaves (± SD) Stalks (± SD) Yatein 23.9 ± 4.9 2.4 ± 0.1 0.2 ± 0.0 Hinokinin 18.9 ± 5.6 3.7 ± 0.4 0.6 ± 0.0 Cubebin 12.3 ± 1.9 2.8 ± 0.3 0.4 ± 0.0
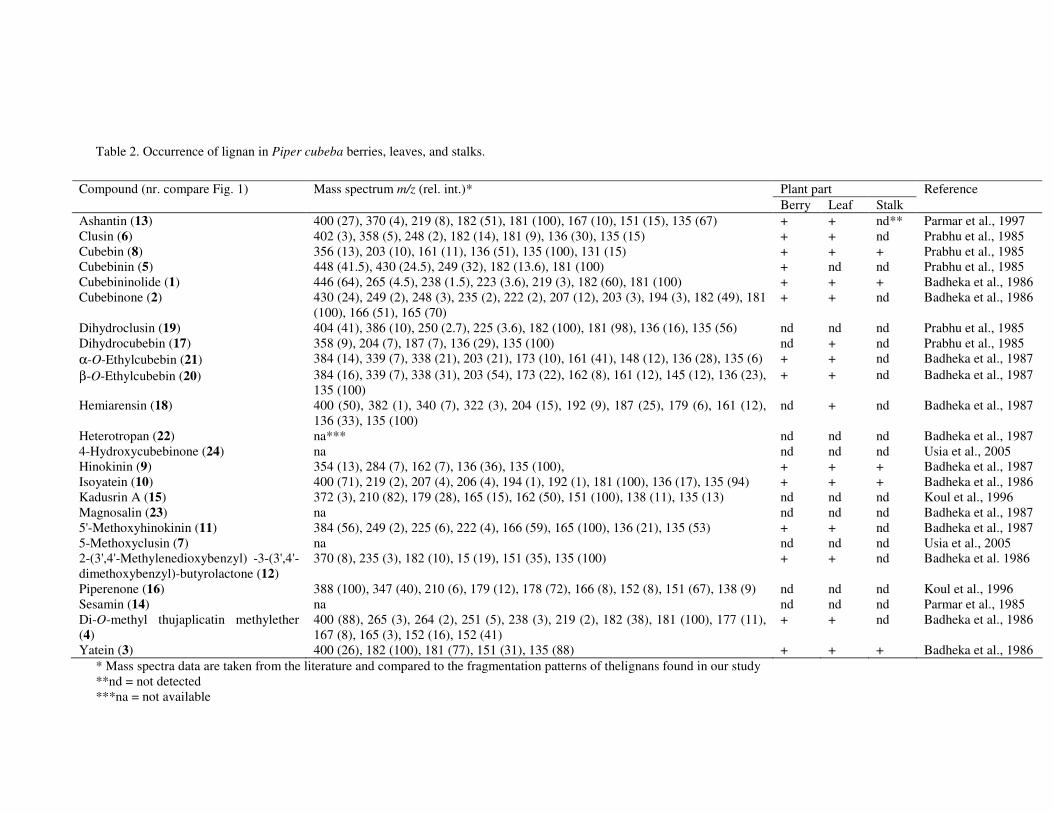
51
Table 2. Occurrence of lignan in Piper cubeba berries, leaves, and stalks.
* Mass spectra data are taken from the literature and compared to the fragmentation patterns of thelignans found in our study **nd = not detected ***na = not available
Plant part Compound (nr. compare Fig. 1) Mass spectrum m/z (rel. int.)* Berry Leaf Stalk
Reference
Ashantin (13) 400 (27), 370 (4), 219 (8), 182 (51), 181 (100), 167 (10), 151 (15), 135 (67) + + nd** Parmar et al., 1997 Clusin (6) 402 (3), 358 (5), 248 (2), 182 (14), 181 (9), 136 (30), 135 (15) + + nd Prabhu et al., 1985 Cubebin (8) 356 (13), 203 (10), 161 (11), 136 (51), 135 (100), 131 (15) + + + Prabhu et al., 1985 Cubebinin (5) 448 (41.5), 430 (24.5), 249 (32), 182 (13.6), 181 (100) + nd nd Prabhu et al., 1985 Cubebininolide (1) 446 (64), 265 (4.5), 238 (1.5), 223 (3.6), 219 (3), 182 (60), 181 (100) + + + Badheka et al., 1986 Cubebinone (2) 430 (24), 249 (2), 248 (3), 235 (2), 222 (2), 207 (12), 203 (3), 194 (3), 182 (49), 181
(100), 166 (51), 165 (70) + + nd Badheka et al., 1986
Dihydroclusin (19) 404 (41), 386 (10), 250 (2.7), 225 (3.6), 182 (100), 181 (98), 136 (16), 135 (56) nd nd nd Prabhu et al., 1985 Dihydrocubebin (17) 358 (9), 204 (7), 187 (7), 136 (29), 135 (100) nd + nd Prabhu et al., 1985 α-O-Ethylcubebin (21) 384 (14), 339 (7), 338 (21), 203 (21), 173 (10), 161 (41), 148 (12), 136 (28), 135 (6) + + nd Badheka et al., 1987 β-O-Ethylcubebin (20) 384 (16), 339 (7), 338 (31), 203 (54), 173 (22), 162 (8), 161 (12), 145 (12), 136 (23),
135 (100) + + nd Badheka et al., 1987
Hemiarensin (18) 400 (50), 382 (1), 340 (7), 322 (3), 204 (15), 192 (9), 187 (25), 179 (6), 161 (12), 136 (33), 135 (100)
nd + nd Badheka et al., 1987
Heterotropan (22) na*** nd nd nd Badheka et al., 1987 4-Hydroxycubebinone (24) na nd nd nd Usia et al., 2005 Hinokinin (9) 354 (13), 284 (7), 162 (7), 136 (36), 135 (100), + + + Badheka et al., 1987 Isoyatein (10) 400 (71), 219 (2), 207 (4), 206 (4), 194 (1), 192 (1), 181 (100), 136 (17), 135 (94) + + + Badheka et al., 1986 Kadusrin A (15) 372 (3), 210 (82), 179 (28), 165 (15), 162 (50), 151 (100), 138 (11), 135 (13) nd nd nd Koul et al., 1996 Magnosalin (23) na nd nd nd Badheka et al., 1987 5'-Methoxyhinokinin (11) 384 (56), 249 (2), 225 (6), 222 (4), 166 (59), 165 (100), 136 (21), 135 (53) + + nd Badheka et al., 1987 5-Methoxyclusin (7) na nd nd nd Usia et al., 2005 2-(3',4'-Methylenedioxybenzyl) -3-(3',4'-dimethoxybenzyl)-butyrolactone (12)
370 (8), 235 (3), 182 (10), 15 (19), 151 (35), 135 (100) + + nd Badheka et al. 1986
Piperenone (16) 388 (100), 347 (40), 210 (6), 179 (12), 178 (72), 166 (8), 152 (8), 151 (67), 138 (9) nd nd nd Koul et al., 1996 Sesamin (14) na nd nd nd Parmar et al., 1985 Di-O-methyl thujaplicatin methylether (4)
400 (88), 265 (3), 264 (2), 251 (5), 238 (3), 219 (2), 182 (38), 181 (100), 177 (11), 167 (8), 165 (3), 152 (16), 152 (41)
+ + nd Badheka et al., 1986
Yatein (3) 400 (26), 182 (100), 181 (77), 151 (31), 135 (88) + + + Badheka et al., 1986

��������
52
Aknowledgment The research was supported by the QUE Project Batch II. Department of Biology, Institut
Teknologi Bandung ITB, Indonesia, under contract No. 3028-IX/P3S-1/KON-QUE II/2000; IBRD Loan No. 4193-IND. We are grateful to Drs. Djuandi, Department of Biology, Institut Teknologi Bandung, for collecting and identifying the plant material.

53
Chapter 5 Essential oil constituents of Piper cubeba from Indonesia
Elfami, Rein Bos, Komar Ruslan, Herman J. Woerdenbag, Oliver Kayser, and Wim J. Quax Submitted

��������!
54
Abstract
The chemical composition of the essential oil of ripe berries (11.8 % v/w) and leaves (0.9 % v/w) of Piper cubeba L. Fils. (Piperaceae) was investigated by GC and GC-MS. Sabinene (9.1%), �-elemene (9.4%), caryophyllene (3.1%), epi-cubebol (4.3%), and cubebol (5.6%) were the main components of the berries oil. Trans-sabinene hydrate (8.2%), E-caryophyllene (5.0%), epi-cubebol (4.2%), -cadinene (16.6%), and cubebol (4.8%) were the main components of the leaves oil. No large qualitative differences were found in the composition between berries and leaves oil, although the berries contained a considerable amount of constituents in traces (<0.05%) that were not found in the leaves. The principal difference was of a quantitative nature.

� � " ���������������������������
55
Introduction The genus Piper belongs to the Piperaceae, a family with more than 700 species throughout the
tropical and subtropical regions of the world. Piper cubeba (in Indonesia known as kemukus), is a plant native to Java and Borneo that produces spicy berries (cubeb berries). It is now also cultivated in several other tropical areas, including East Africa. In Indonesia P. cubeba is valued as a medicinal plant (Eisei 1995, Sastroamidjojo, 2001). Many species of the genus Piper are used in traditional herbal medicine, and have shown antifungal, insecticidal, anthelminthic and antitumor activities. They are also used for the treatment of cough, bronchitis, intestinal diseases, rheumatism (Sumathykutty et al., 1999). A number of polyhydroxy cyclohexanes have been isolated from Piper cubeba and shown to display tumor inhibitory, antileukemic and antibiotic activities (Taneja et al., 1991). Essential oil investigations of a number of Piper species have been reported, but only one gives a detailed overview of the essential oil of cubeb berries (Martins et al., 1998, Sumathykutty et al., 1999, Jirovetz et al., 2002, Orav et al., 2004, Oyedeji et al., 2005). The aim of the present study was to investigate of the essential oil composition of P. cubeba berries and leaves from Indonesia. Materials and methods Plant material
Piper cubeba L. (Piperaceae) was collected in April 2002 from Jatiroto, Temanggung, Central Java, Indonesia, and authenticated at the Department of Biology, Institut Teknologi Bandung (Indonesia), based on the Flora of Java (Backer and Van de Brink, 1968). A voucher specimen (HBG10PC01) is deposited at the Herbarium Bandungense. The collected material was air-dried. Isolation procedure
The essential oil sample was isolated from 20.0 g of air-dried and freshly ground (1 mm) leaves and fruit material by hydrodistillation for 4 h in 300 mL-1 water, according to the determination of the essential oil content in vegetable drugs, using the apparatus described in the Nederlandse Farmacopee, 6th ed., 2nd printing (Anonymous, 1966). Xylene (100 µL) was used as the collection liquid, and the essential oil was stored at -20° C until analyzed. The essential oil was diluted 50 times with cyclohexane prior to GC and GC-MS analysis.
In addition, the oil was separated into two fractions with hydrocarbons and oxygen-containing compounds, respectively, by eluting 250 µL of oil on a Bakerbond SPE column, filled with 1 g of silica gel (7086-07, J.T. Baker, Deventer, The Netherlands), with subsequently 5 mL-1 n-hexane and 5 mL-1 diethyl ether. After gentle evaporation of the solvents of both fractions, 50 µL of each residue were diluted with 950 µL cyclohexane and injected to GC and GC-MS analysis. Gas chromatography
GC analysis was performed on a Hewlett-Packard 5890 Series II gas chromatograph equipped with a 7673 injector and a Hewlett Packard 3365 Series II Chemstation, under the following conditions: column, WCOT fused-silica (J & W) DB-5 (30 m x 0.26 mm; film thickness 0.25 µm); oven temperature programme, 60°-290°C at 3°C min-1; injector temperature, 250°C; detector (FID) temperature, 300°C; carrier gas, He; inlet pressure, 18 psi; linear gas velocity, 31.8 cm s-1; split ratio, 56:1; injected volume, 1.0 µL.

��������!
56
Gas chromatography-mass spectrometry
A Shimadzu GCMS QP5000 system was used equipped with a GC-17A gas chromatograph, an AOC-20i auto injector, and GCMS solution version 1.10 software. The GC conditions were: column, WCOT fused-silica (J & W) DB-5 (30 m x 0.26 mm; film thickness 0.25 µm); oven temperature programme, 60°-240°C at 3°C min-1; injector temperature, 275°C; carrier gas, He; inlet pressure, 75 pKa; linear gas velocity, 81.4 cm s-1; column flow, 2.5 mL min-1; total flow, 56.7 mL min-1; split ratio, 21:1; injected volume, 1.0 µL. MS conditions: ionization energy, 70 eV; ion source temperature, 250°C; interface temperature, 250°C; scan speed, 3 scans s-1; mass range, 34-350 u.
The identity of the components was assigned by comparison of their retention indices, relative to C9-C22 n-alkanes, and mass spectral databases and from the literature (Adams, 2001, Joulain and König, 1998). The percentages of the components were calculated from the GC peak areas, using the normalization method. Results and discussion
Hydrodistillation of the berries of Piper cubeba yielded 11.8% (w/w) and the leaves 0.9% (v/w) oil. In total 105 components could be identified in the berries, dealing with 63.1% of the oil. In the leaves oil, 63 components could be identified, accounting for 78.0% of the oil. As far as we know, this is the first time the essential oil composition of P. cubeba leaves has been investigated.
Comparing the composition of the berries and the leaves oil, no big differences were found for the monoterpene and sesquiterpene hydrocarbon fractions, as well as for the oxygenated monoterpene and sesquiterpene fractions (see Table 1). The total amount of monoterpenes was comparable in both oils (17.2% and 17.0%, for berries and leaves, respectively). The main monoterpenes in the berries oil were �-thujene (2.5%), �-pinene (1.8%), sabinene (9.1%), and limonene (2.3%), while �-pinene (3.2%), sabinene (3.8%), �-pinene (3.8%) and limonene (3.4%) were the principal monoterpenes in the leaves oil. In the oxygenated monoterpene fractions (3.6% and 10.6%, respectively, for the berries and leaves oil), trans-sabinene hydrate was the main component (2.5% and 8.2%, respectively). �-Copaene (3.8%), �-elemene (9.4%), E-caryophyllene (2.5%), caryophyllene (3.1%), were the main sesquiterpenes (26.8%) in the berries oil, where E-caryophyllene (5.0%), and -cadinene (16.6%) were the main sesquiterpenes (31.0%) in the leaves oil. Remarkable is the high content of -cadinene in the leaves oil, whereas it was present in the berries oil in only small amounts (0.1%). From the oxygenated sesquiterpenes (15.5% and 18.6%, respectively in berries and leaves), epi-cubebol (4.6% and 4.2%) and cubebol (5.6% and 4.8%), were the main components in both oils. Other major components were guaiol (2.9%) in the berries oil, and -cadinol (2.7%) and �-cadinol (1.9%) in the leaves oil.
Two sesquiterpenes in a relatively low concentration (0.05%) with retention indices of 1566 (compound I) and 1570 (compound II) were detected, but could not be identified. Comparing the mass fragmentation patterns of compound I, m/z(%): 41(70); 55(29); 69(3); 77(11); 91(74); 105(58); 117(9); 133(100); 145(3); 157(2); 187(1); 202(M+, 12), and compound II, m/z(%): 41(67); 55(29); 65(10); 77(14); 91(87); 105(62); 117(10); 133(100); 187(2); 202(M+, 14), we conclude that these two sesquiterpenes are isomers. Also seven sesquiterpene epoxides with retention indices between 1600 and 1700 were detected, but could not be identified either.
From the literature only three older and more limited studies concerning the essential oil composition of P. cubeba berries are known. In a commercial sample from Indonesia (without further specifications), the main components were copaene (10.4%), �-cubebene (11.0%), and cubebol with 10.0%, where cubenol and epi-cubenol were present with 3.5% (Lawrence, 1980). In the P. cubeba berries oil (16.8%) from Sri Lanka the main components were cubebol (31%), �-cubebene (5.1%) and �-copaene (8.1%) (Violon et al., 1991). In P. cubeba berries oil (14.5%) from India the main compounds

� � " ���������������������������
57
were cubebol (23.6%), �-pinene (18.2%), �-elemene (7.3%), �-cubebene (5.6%), and �-cadinene (4.7%) (Sumathykutty et al., 1999).
Comparing the results of our present study with those from the three older reports, we come to the following conclusions. Cubebol is one of the main components in the berries oil, but the amount seems to depend on the origin of the material. Indonesian samples contained about 10%, the Indian and Sri Lankese samples considerably more. We identified cubebol together with epi-cubebol. The other studies do not discriminate between the two epimers. Less abundant in our samples, compared with the previous studies, were �-cubebene, �-copaene, and �-cubebene.
Lawrence (1980) found cubenol in the berries oil from India. We did not find this compound, but detected 1,10-di-epi-cubenol (trace), and epi-cubenol (0.3%) in the berries oil from Indonesia. These differences may be used as a tool for the characterization of P. cubeba oils from different origin, although further systematic studies are needed to prove this. Acknowledgements
The research was supported by the QUE Project Batch II, Department of Biology, Institut Teknologi Bandung ITB, Indonesia, under contract No. 3028-IX/P3S-1/KON-QUE II/2000; IBRD Loan No. 4193-IND. We are grateful to Drs. Djuandi, Department of Biology, Institut Teknologi Bandung, for collecting plant materials.
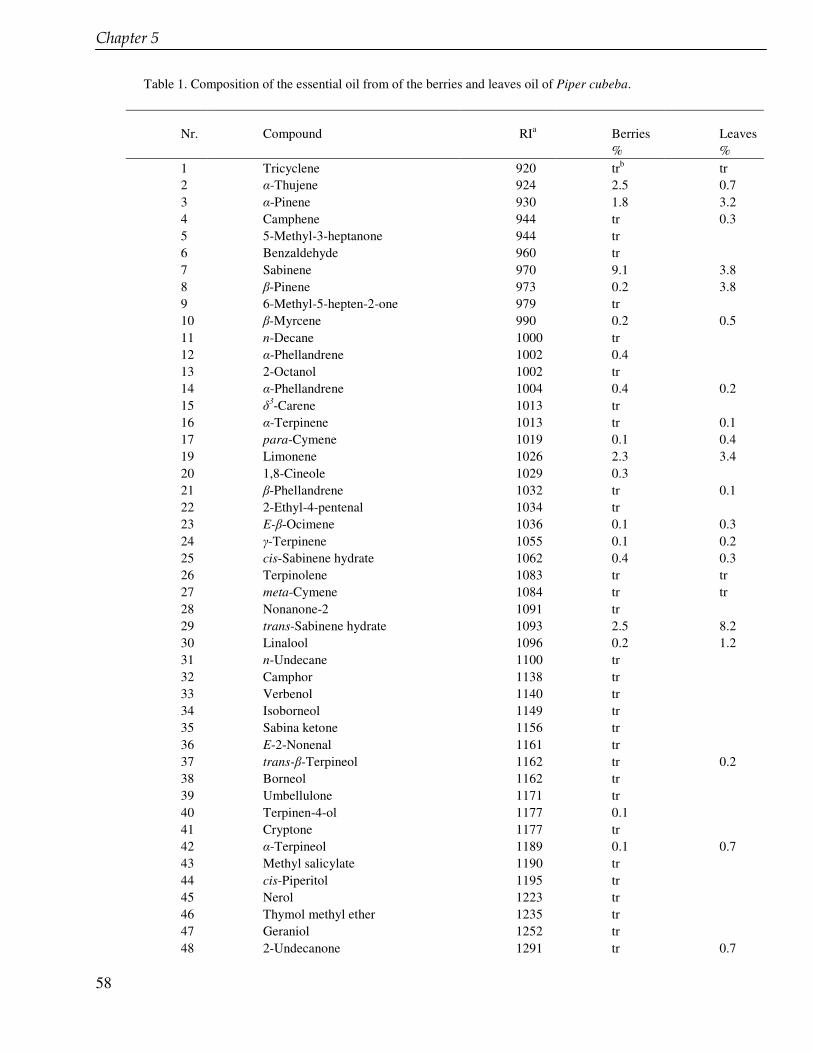
��������!
58
Table 1. Composition of the essential oil from of the berries and leaves oil of Piper cubeba.
Nr. Compound RIa Berries Leaves % % 1 Tricyclene 920 trb tr 2 �-Thujene 924 2.5 0.7 3 �-Pinene 930 1.8 3.2 4 Camphene 944 tr 0.3 5 5-Methyl-3-heptanone 944 tr 6 Benzaldehyde 960 tr 7 Sabinene 970 9.1 3.8 8 �-Pinene 973 0.2 3.8 9 6-Methyl-5-hepten-2-one 979 tr 10 �-Myrcene 990 0.2 0.5 11 n-Decane 1000 tr 12 �-Phellandrene 1002 0.4 13 2-Octanol 1002 tr 14 �-Phellandrene 1004 0.4 0.2 15 �
3-Carene 1013 tr 16 �-Terpinene 1013 tr 0.1 17 para-Cymene 1019 0.1 0.4 19 Limonene 1026 2.3 3.4 20 1,8-Cineole 1029 0.3 21 �-Phellandrene 1032 tr 0.1 22 2-Ethyl-4-pentenal 1034 tr 23 E-�-Ocimene 1036 0.1 0.3 24 �-Terpinene 1055 0.1 0.2 25 cis-Sabinene hydrate 1062 0.4 0.3 26 Terpinolene 1083 tr tr 27 meta-Cymene 1084 tr tr 28 Nonanone-2 1091 tr 29 trans-Sabinene hydrate 1093 2.5 8.2 30 Linalool 1096 0.2 1.2 31 n-Undecane 1100 tr 32 Camphor 1138 tr 33 Verbenol 1140 tr 34 Isoborneol 1149 tr 35 Sabina ketone 1156 tr 36 E-2-Nonenal 1161 tr 37 trans-�-Terpineol 1162 tr 0.2 38 Borneol 1162 tr 39 Umbellulone 1171 tr 40 Terpinen-4-ol 1177 0.1 41 Cryptone 1177 tr 42 �-Terpineol 1189 0.1 0.7 43 Methyl salicylate 1190 tr 44 cis-Piperitol 1195 tr 45 Nerol 1223 tr 46 Thymol methyl ether 1235 tr 47 Geraniol 1252 tr 48 2-Undecanone 1291 tr 0.7
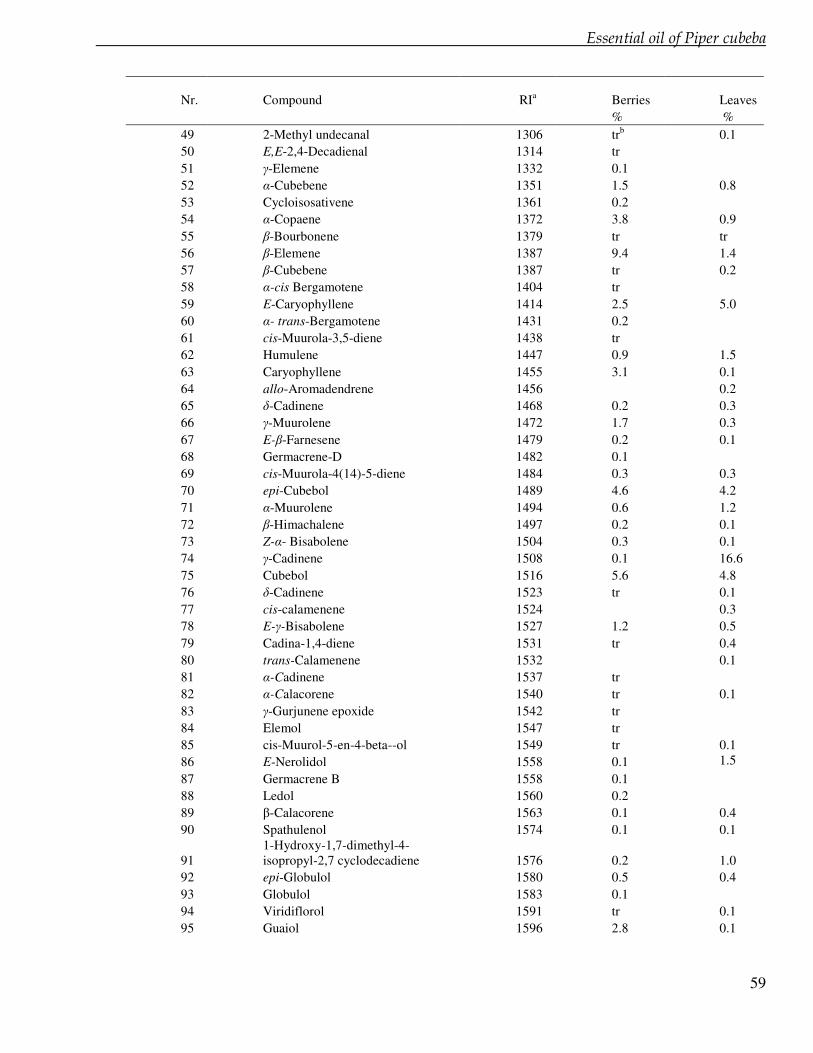
� � " ���������������������������
59
Nr. Compound RIa Berries Leaves % % 49 2-Methyl undecanal 1306 trb 0.1 50 E,E-2,4-Decadienal 1314 tr 51 �-Elemene 1332 0.1 52 �-Cubebene 1351 1.5 0.8 53 Cycloisosativene 1361 0.2 54 �-Copaene 1372 3.8 0.9 55 �-Bourbonene 1379 tr tr 56 �-Elemene 1387 9.4 1.4 57 �-Cubebene 1387 tr 0.2 58 �-cis Bergamotene 1404 tr 59 E-Caryophyllene 1414 2.5 5.0 60 �- trans-Bergamotene 1431 0.2 61 cis-Muurola-3,5-diene 1438 tr 62 Humulene 1447 0.9 1.5 63 Caryophyllene 1455 3.1 0.1 64 allo-Aromadendrene 1456 0.2 65 �-Cadinene 1468 0.2 0.3 66 �-Muurolene 1472 1.7 0.3 67 E-�-Farnesene 1479 0.2 0.1 68 Germacrene-D 1482 0.1 69 cis-Muurola-4(14)-5-diene 1484 0.3 0.3 70 epi-Cubebol 1489 4.6 4.2 71 �-Muurolene 1494 0.6 1.2 72 �-Himachalene 1497 0.2 0.1 73 Z-�- Bisabolene 1504 0.3 0.1 74 �-Cadinene 1508 0.1 16.6 75 Cubebol 1516 5.6 4.8 76 �-Cadinene 1523 tr 0.1 77 cis-calamenene 1524 0.3 78 E-�-Bisabolene 1527 1.2 0.5 79 Cadina-1,4-diene 1531 tr 0.4 80 trans-Calamenene 1532 0.1 81 �-Cadinene 1537 tr 82 �-Calacorene 1540 tr 0.1 83 �-Gurjunene epoxide 1542 tr 84 Elemol 1547 tr 85 cis-Muurol-5-en-4-beta--ol 1549 tr 0.1 86 E-Nerolidol 1558 0.1 1.5 87 Germacrene B 1558 0.1 88 Ledol 1560 0.2 89 �-Calacorene 1563 0.1 0.4 90 Spathulenol 1574 0.1 0.1
91 1-Hydroxy-1,7-dimethyl-4-isopropyl-2,7 cyclodecadiene 1576 0.2 1.0
92 epi-Globulol 1580 0.5 0.4 93 Globulol 1583 0.1 94 Viridiflorol 1591 tr 0.1 95 Guaiol 1596 2.8 0.1
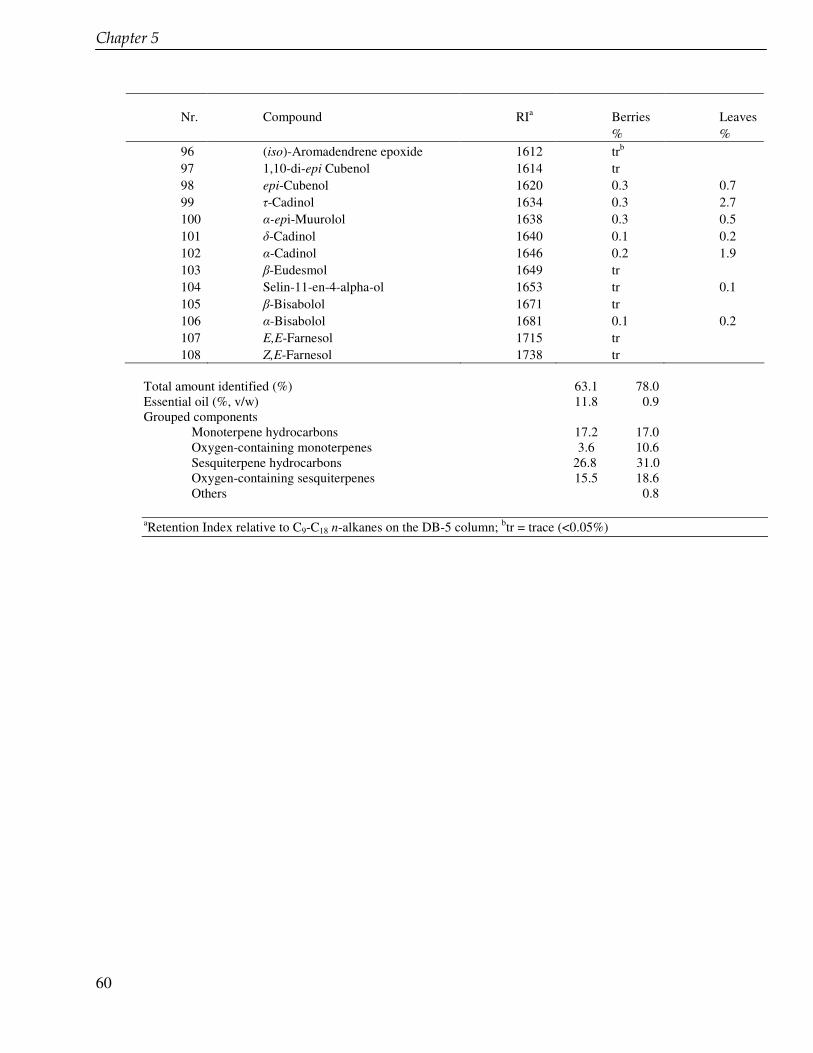
��������!
60
Nr. Compound RIa Berries Leaves % % 96 (iso)-Aromadendrene epoxide 1612 trb 97 1,10-di-epi Cubenol 1614 tr 98 epi-Cubenol 1620 0.3 0.7 99 �-Cadinol 1634 0.3 2.7 100 �-epi-Muurolol 1638 0.3 0.5 101 �-Cadinol 1640 0.1 0.2 102 �-Cadinol 1646 0.2 1.9 103 �-Eudesmol 1649 tr 104 Selin-11-en-4-alpha-ol 1653 tr 0.1 105 �-Bisabolol 1671 tr 106 �-Bisabolol 1681 0.1 0.2 107 E,E-Farnesol 1715 tr 108 Z,E-Farnesol 1738 tr
Total amount identified (%) 63.1 78.0 Essential oil (%, v/w) 11.8 0.9 Grouped components Monoterpene hydrocarbons 17.2 17.0 Oxygen-containing monoterpenes 3.6 10.6
Sesquiterpene hydrocarbons 26.8 31.0 Oxygen-containing sesquiterpenes 15.5 18.6 Others 0.8
aRetention Index relative to C9-C18 n-alkanes on the DB-5 column; btr = trace (<0.05%)

61
Chapter 6 Reduced coniferin and enhanced 6-methoxypodophyllotoxin production in Linum flavum cell suspension cultures after treatment with Na2EDTA Elfahmi, Sieb Batterman, Albert Koulman, Henricus L. Hoge, Rein Bos, Oliver Kayser, Herman J. Woerdenbag, Wim J. Quax Submitted

��������#
62
Abstract Treatment of cell suspension cultures of Linum flavum L. with Na2EDTA reduced the coniferin and enhanced the 6-methoxypodophyllotoxin (6-MPT) production in a concentration-dependent way, in a range of 0.1 - 5 mM. On day 14 after treatment with Na2EDTA, an inhibition of the coniferin production up to 88% was found (content in control cultures 120.7 mg g-1 DW). The maximum enhancement of the 6-MPT production was 320% on day 7 after treatment with 5 mM Na2EDTA (control value 0.16 mg g-1 DW).
The reduction in coniferin accumulation in the suspension cultures was correlated with an inhibition of coniferyl alcohol glucosyltransferase (CAGT) activity as determined in cell homogenates. On day 14 after treatment with 2 and 5 mM Na2EDTA the CAGT activity was inhibited up to > 89 % (control value 8.8 µkat g-1). The inhibitory effect of Na2EDTA on CAGT was also shown in a partially purified enzyme preparation. Several metal ions such as Fe3+, Cu2+, Zn2+, Li+, and the elicitors nigeran and salicylic acid had no significant effect on the production of coniferin and 6-MPT.

� "���������$��"%���������� � ��� ����������� �������������&���
63
Introduction Podophyllotoxin and podophyllotoxin-derived lignans possess cytotoxic and antiviral activities.
Teniposide and etoposide are semi-synthetic derivatives of podophyllotoxin that are clinically used as anticancer drugs (Moraes et al., 2002). Podophyllotoxin is also the starting compound for the rheumatoid arthritis drug CPH 82 (Reumacon) (Svensson and Pettersson, 2003). For the production of semi-synthetic podophyllotoxin derivatives on an industrial scale, podophyllotoxin is isolated from the rhizomes of Podophyllum plants from wild habitats, which are counted as endangered species (Smollny et al., 1998).
The use of biotechnological approaches to improve the production of podophyllotoxin or related lignans with plant cell and organ cultures, including the biotransformation of suitable precursors and the modification of biosynthetic pathways, is considered to be suitable and economically attractive (Giri and Narasu, 2000). Several investigations to enhance the production of podophyllotoxin-derived lignans by manipulation of cell and organ cultures have been carried out (Van Uden et al., 1990a, 1991a, Woerdenbag et al., 1990, Berim et al., 2005, Petersen and Alfermann, 2001). The production of podophyllotoxin, 6-methoxypodophyllotoxin (6-MPT) and its glucoside could be enhanced in cell cultures of Podophyllum hexandrum Royle (Woerdenbag et al., 1990), Linum flavum L. (Van Uden et al., 1990a, 1992, Molog et al., 2001) Callitris drummondii F. Mueller (Van Uden et al., 1990b), Linum album Kotschy (Seidel et al., 2002), and Linum nodiflorum (Berim et al., 2005).
Based on the close chemical resemblance with podophyllotoxin (see Fig. 1), 6-MPT is considered also as an interesting starting compound for the preparation of new semi-synthetic derivatives with antitumor properties. Cell cultures of L. flavum produce 6-MPT and its glucoside. The cytotoxicity of 6-MPT in vitro against tumor cell lines was comparable with that of podophyllotoxin (Middel et al., 1995).
Coniferyl alcohol is an early precursor of both lignins and lignans. The glucosylation of coniferyl alcohol yields coniferin that is accumulated endogenously in L. flavum cultures up to 12 % on a dry weight basis (Van Uden et al., 1991b). This reaction is catalysed by coniferyl alcohol glucosyltransferase (CAGT, EC number: 2.4.1.111). Lignans are formed through radical-mediated dimerisation of two coniferyl alcohol units. Blocking the branch leading to the formation of coniferin by inhibiting CAGT could result in an enhanced production of lignans, such as 6-MPT in the cell suspension cultures of L. flavum. High coniferin contents in the cell suspension culture correspond with low 6-MPT levels, as was demonstrated in feeding experiment with cell cultures of P. hexandrum (Van Uden et al., 1990a). It is known that Na2EDTA (Ibrahim and Grisebach, 1976) and metal ions such as Cu2+, Zn2+ , Li+ and Fe2+ (Schmid and Grisebach, 1982, Wunder and William, 1999, Reed et al., 1993) are able to inhibit the glucosyltransferase activity. Addition of one of these compounds may interfere with CAGT.
The aim of this paper is to explore the effect of Na2EDTA and several inorganic salts on the production of coniferin and 6-MPT in cell suspension cultures of L. flavum. This effect is compared to that of the elicitors salicylic acid and nigeran. These compounds enhance various biosynthetic pathways, but so far there is little evidence of their effect on the lignan biosynthesis (Guan and Scandalios, 1995, Chong et al., 1999).

��������#
64
Materials and methods Plant material and culture conditions
Cell suspensions of Linum flavum L. (Linaceae) leaves have been initiated and are maintained at the Department of Pharmaceutical Biology, University of Groningen. The cell suspensions are cultured routinely every two weeks by transferring 100 mL of fully grown suspension aseptically into 200 mL of fresh liquid medium. The medium is a MS (Murashige and Skoog 1962) containing 0.2 mg of indole-3-acetic acid (IAA) and 0.2 mg of 6-benzylaminopurine (BAP), purchased from Duchefa (Haarlem, the Netherlands). The cell suspensions were incubated on a rotary shaker (175 rpm) at 26°C under a day/night regime (16/8 h: 3,000 lux, day light L 36W/10, OSRAM, Germany).
Treatment of cell suspensions
Disodium ethylenediamine tetra acetate (Na2EDTA) was purchased from Duchefa, copper (II) sulphate pentahydrate, zinc chloride, and lithium chloride from Merck (Darmstadt, Germany), iron (II) sulphate pentahydrate, salicylic acid from Sigma-Aldrich (Zwijndrecht, the Netherlands), and nigeran from Sigma, St. Louis (USA).
These compounds were added to the culture media used for the cell suspension cultures of L. flavum yielding the following final concentrations: Na2EDTA : 0.01, 0.5, 1, 2 and 5 mM, Cu2+, Zn2+ and Li+: 0.1, 1 and 5 mM, Fe2+: 0.01, 0.1 and 1 mM, salicylic acid: 0.1, 0.5, 1 and 5 mM; nigeran 20 mg L-1. Suspension-grown cells were harvested each 2 days during the growth cycle of 14 days. Samples of about 10 mL were taken aseptically and transferred into a calibrated conical tube and centrifuged for 5 min at 1,500 g. In order to monitor the viability and growth of the cell cultures, the medium pH and the conductance were routinely measured in the supernatant. The cells were filtered using Buchner funnel. Fresh weight (FW) was determined and the cells put overnight in the freezer and subsequently freeze dried. Dry weight (DW) was also determined. Coniferin and 6-MPT contents were subsequently analyzed by HPLC.
Extraction
About 100 mg, accurately weighed of freeze dried and powdered cell material were extracted by ultrasonification in 2 mL methanol (80%; v/v) for 1 h. Dichloromethane (4.0 mL) and water (4.0 mL) were added. The mixture was vortexed and centrifuged (5 min; 1,500 g). For the determination of the 6-MPT concentration, 2.0 mL of the dichloromethane phase were taken and evaporated to dryness. The residue was redissolved in 1.0 mL methanol and centrifuged. For the determination of coniferin 50 µL water phase were diluted with water until 1.0 mL and centrifuged (2 min; 10,000 g).
Treatment of aqueous phase with ββββ-glucosidase
To confirm the coniferin production, the water phase was submitted to enzymatic hydrolysis. A 3.5% (w/v) solution of β-glucosidase (Sigma G-0395) was prepared in 0.1 M phosphate buffer, pH 5.0. To 2.0 mL samples of the water phase 0.5 mL was added, followed by incubation for 5 h at 37°C. The aglucone formed was extracted with 2.0 mL dichloromethane. Of the dichloromethane phase 1.5 mL were taken and evaporated to dryness. The residue was redissolved in 1.0 mL methanol and centrifuged (2 min; 10,000 g). Coniferyl alcohol was determined by HPLC.

� "���������$��"%���������� � ��� ����������� �������������&���
65
Protein purification
Cells were harvested on day 1 after subculturing and stored at -20°C overnight. Frozen cells were suspended in an equal volume of the homogenisation buffer that consisted of 0.2 M Tris-HCl, pH 7.5, 5% polyvinylpyrrolidone, 0.2 % DOWEX�- 1*2 – 100, 0.1 % DTT (w/w) and 10% ethylenglycol. The mixture was homogenised using an ultraturrax (Janke & Kunkel, IKA-WERK, Staufen, Germany). The homogenate was filtered through Miracloth and clarified by centrifugation for 20 min at 20,000 g. Proteins dissolved in the supernatant were then fractionated by (NH4)2SO4 precipitation. The fraction obtained between 40 and 80% saturation was desalted on a HiPrep 26/10 desalting column (Amersham Biosciences, Uppsala, Sweden) previously equilibrated with 0.02 M Tris-HCl buffer, pH 7.5. The protein eluting from the desalting column was applied to a HiTrap DEAE FF column (Amersham Biosciences, Uppsala, Sweden) which had been equilibrated with 0.02 M Tris-HCl buffer, pH 7.5. The protein was eluted first with 100 mL of 0.02 M Tris-HCl buffer, followed by a linear gradient from 0.02 to 0.2 M Tris-HCl buffer and finally with 100 mL of 0.4 M Tris-HCl buffer, all at pH 7.5. Fractions of 5 mL each were collected at a flow rate of 1 mL min-1 and assayed directly for CAGT activity. Fractions containing the highest activity were combined and concentrated by vivaspin 6 mL concentrator (Vivasciences, Hannover, Germany). The combined fractions with the highest CAGT activity resulting from the desalting column (the third step of the purification procedure, see Table 2) were exposed to Na2EDTA. The Na2EDTA was added to 5 mL of the partially purified CAGT yielding final concentrations of 0.1, 0.5, 1, 2 and 5 mM. The mixtures were incubated at 4°C for one day. 200 µL (3x) of the mixture were taken and submitted for CAGT assay.
CAGT assay
The enzyme assay for CAGT was developed from the methods used by Ibrahim et al. (1976) and Schmid et al. (1982). Cells were treated with Na2EDTA in the range of 0.1-5 mM and harvested at different time points during the growth cycle and stored at -20°C overnight. Frozen cells, 2-3 g, were suspended in homogenisation buffer and the mixture was homogenised using an ultraturrax. The homogenate was centrifuged (3,000 g; 25 min, 2°C). The supernatant was separated from the pellet. The assay buffer was prepared containing 0.2 M Tris-HCl pH 7.5, DTT 0.1 %. The standard assay mixture consisted of 0.32 µmol coniferyl alcohol in 40 µL ethyleneglycol monomethylether, 0.32 µmol uridine diphosphate (UDP)-glucose in 40 µL assay buffer, 200 µL protein homogenate or partially purified CAGT and assay buffer in a total volume of 320 µL. The reaction was started by the addition of protein and vortexed for 5 s immediately followed by incubation for 30 min at 30°C. The reaction was stopped by adding 2.0 mL dichloromethane followed by vortexing the mixture for 20 s and centrifugation (5 min; 1,500 g). The dichloromethane and water layers were used for HPLC analysis of coniferyl alcohol and coniferin, respectively. Protein was determinated using the Bradford assay (Bradford, 1976). Analysis of coniferin, coniferyl alcohol and 6-MPT
Coniferin, coniferyl alcohol and 6-MPT were analyzed by HPLC. The HPLC system consisted of an ISCO Model 2350 pump, a Shimadzu photodiode array detector (Shimadzu, ‘s-Hertogenbosch, the Netherlands), UV absorbance at 230 and 290 nm and LiChrocart RP-18 column (250 x 4.6 mm i.d.) (Merck, Darmstadt, Germany). The mobile phase for coniferyl alcohol and 6-MPT analysis was acetonitrile (LAB-SCAN Analytical-sciences, Dublin, Ireland) / water (40:60 v/v; 0.1% phosphoric acid) and for coniferin, methanol/water (30:70; 0.1 % phosphoric acid). Calibration curves were made using coniferyl alcohol (Sigma), coniferin and 6-MPT, which were isolated from L. flavum cell suspension cultures as published previously (van Uden et al., 1991b, 1992). For the statistical evaluation of the data the Sudent’s t-test was used. A P-value <0.05 was considered as significant.

��������#
66
O
O
OH
O
OMeOMe
MeO
R
CH2OH
OCH3
OR
1. R = H2. R = OMe
3. R = H4. R = Glucose
Fig. 1. Chemical structures of podophyllotoxin (1), 6-methoxypodophyllotoxin (2), coniferyl alcohol (3) and coniferin (4).
Results and discussion
The effect of Na2EDTA on the growthy of L. flavum cell suspension cultures was determined on the basis of dry weight accumulation as shown in Fig. 2. The growth period of the cultures was 14 days. From day 1 after inoculation the cells grew until day 8. The stationary phase was reached between day 8 and 12. At the end of the period (day 14) the cell suspension was refreshed. There was no significant effect of Na2EDTA on the growth of the cell suspensions or on the viability parameters at concentrations of 0.1, 0.5 and 1 mM (dry weight and conductivity). At a concentration of 2 and 5 mM, Na2EDTA inhibited cell growth up to 22% and 59% respectively (maximal effect) on day 8.
In Fig. 3 the effect of treatment with Na2EDTA on the coniferin production in L. flavum cells is shown. Untreated cells (control) contained up to 12.0 % coniferin on a dry weigh basis on day 14 of the growth cycle. After treatment with Na2EDTA, the coniferin production was reduced in a concentration dependent way by 18-88% (Fig. 5) on day 14, although it should be noted that the higher concentrations of Na2EDTA (2 and 5 mM) also inhibited the cell growth.
To confirm the coniferin production, the water phase that contained coniferin was submitted to enzymatic hydrolysis using β-glucosidase. This enzyme catalyses the hydrolysis of monolignol glucosidases, that lead to the release of the corresponding alcohols (Dharmawardhana et al., 1995). The coniferyl alcohol formed fully correlated with the coniferin content as found after hydrolysis.
The accumulation of 6-MPT was enhanced 1.2, 1.9 and 3.2 fold at a concentration of Na2EDTA of 1, 2 and 5 mM respectively on day 7 in the cell suspensions of L. flavum (Fig. 4, Fig. 5). Adding 0.1 mM and 0.5 mM Na2EDTA did not enhance the 6-MPT production.
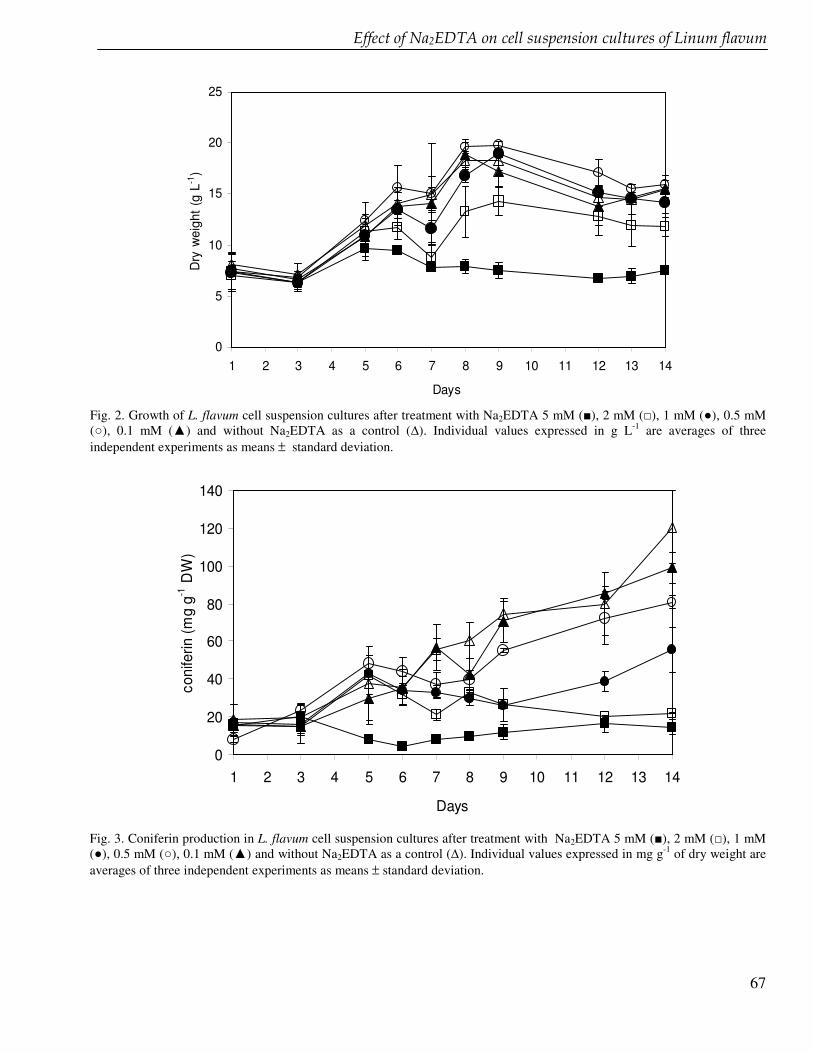
� "���������$��"%���������� � ��� ����������� �������������&���
67
0
5
10
15
20
25
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Days
Dry
wei
ght (
g L-1
)
Fig. 2. Growth of L. flavum cell suspension cultures after treatment with Na2EDTA 5 mM (), 2 mM (�), 1 mM (�), 0.5 mM (�), 0.1 mM (�) and without Na2EDTA as a control (�). Individual values expressed in g L-1 are averages of three independent experiments as means ± standard deviation.
0
20
40
60
80
100
120
140
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Days
coni
ferin
(mg
g-1 D
W)
Fig. 3. Coniferin production in L. flavum cell suspension cultures after treatment with Na2EDTA 5 mM (), 2 mM (�), 1 mM (�), 0.5 mM (�), 0.1 mM (�) and without Na2EDTA as a control (�). Individual values expressed in mg g-1 of dry weight are averages of three independent experiments as means ± standard deviation.
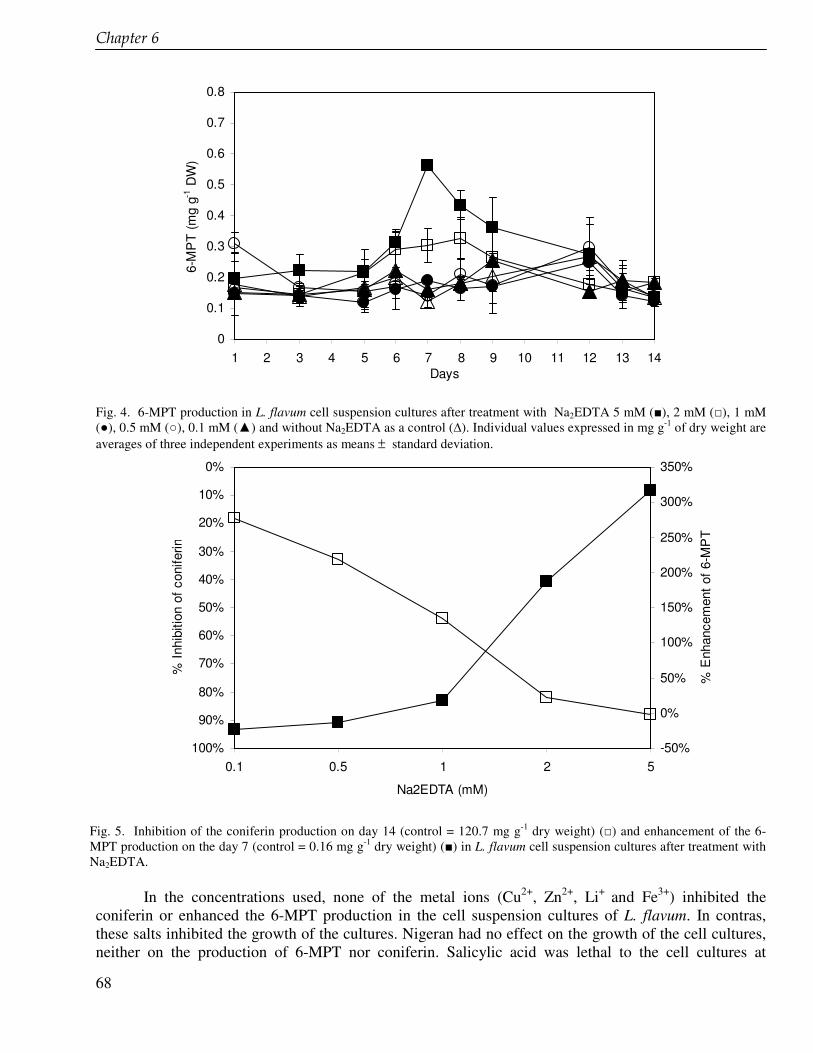
��������#
68
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
1 2 3 4 5 6 7 8 9 10 11 12 13 14Days
6-M
PT
(mg
g-1 D
W)
Fig. 4. 6-MPT production in L. flavum cell suspension cultures after treatment with Na2EDTA 5 mM (), 2 mM (�), 1 mM (�), 0.5 mM (�), 0.1 mM (�) and without Na2EDTA as a control (�). Individual values expressed in mg g-1 of dry weight are averages of three independent experiments as means ± standard deviation.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%0.1 0.5 1 2 5
Na2EDTA (mM)
% In
hibi
tion
of c
onife
rin
-50%
0%
50%
100%
150%
200%
250%
300%
350%
% E
nhan
cem
ent o
f 6-M
PT
Fig. 5. Inhibition of the coniferin production on day 14 (control = 120.7 mg g-1 dry weight) (�) and enhancement of the 6-MPT production on the day 7 (control = 0.16 mg g-1 dry weight) () in L. flavum cell suspension cultures after treatment with Na2EDTA.
In the concentrations used, none of the metal ions (Cu2+, Zn2+, Li+ and Fe3+) inhibited the coniferin or enhanced the 6-MPT production in the cell suspension cultures of L. flavum. In contras, these salts inhibited the growth of the cultures. Nigeran had no effect on the growth of the cell cultures, neither on the production of 6-MPT nor coniferin. Salicylic acid was lethal to the cell cultures at
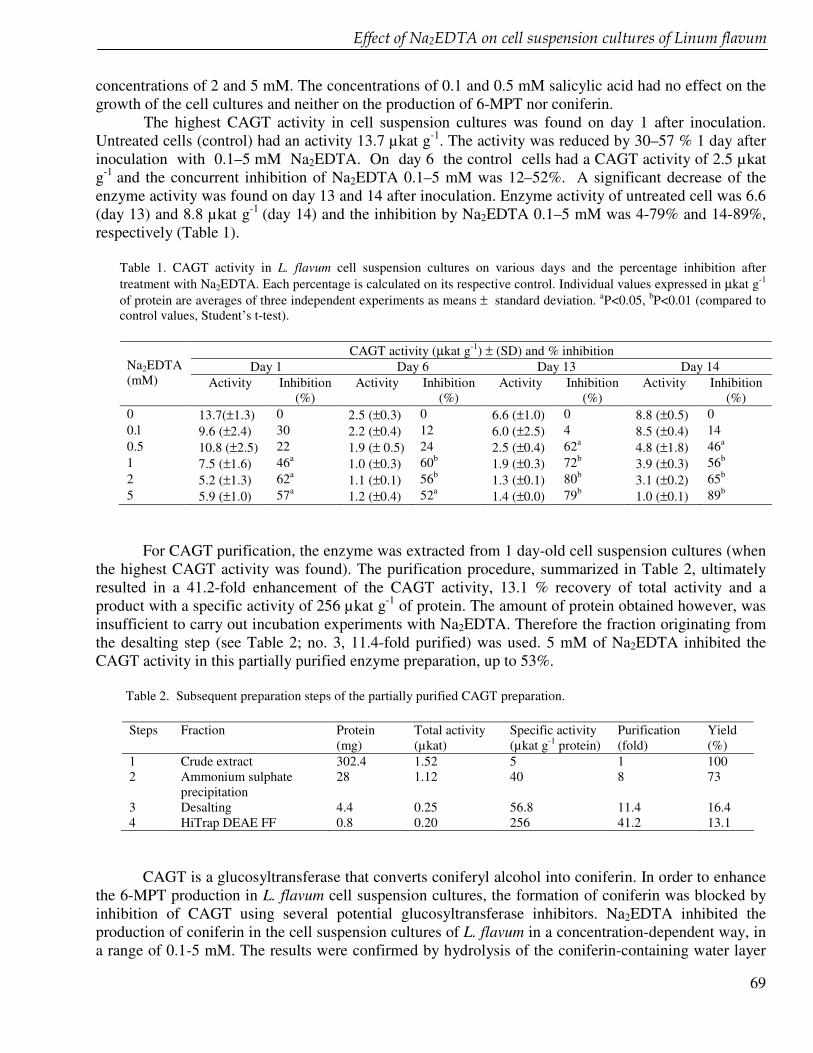
� "���������$��"%���������� � ��� ����������� �������������&���
69
concentrations of 2 and 5 mM. The concentrations of 0.1 and 0.5 mM salicylic acid had no effect on the growth of the cell cultures and neither on the production of 6-MPT nor coniferin. The highest CAGT activity in cell suspension cultures was found on day 1 after inoculation. Untreated cells (control) had an activity 13.7 µkat g-1. The activity was reduced by 30–57 % 1 day after inoculation with 0.1–5 mM Na2EDTA. On day 6 the control cells had a CAGT activity of 2.5 µkat g-1 and the concurrent inhibition of Na2EDTA 0.1–5 mM was 12–52%. A significant decrease of the enzyme activity was found on day 13 and 14 after inoculation. Enzyme activity of untreated cell was 6.6 (day 13) and 8.8 µkat g-1 (day 14) and the inhibition by Na2EDTA 0.1–5 mM was 4-79% and 14-89%, respectively (Table 1).
Table 1. CAGT activity in L. flavum cell suspension cultures on various days and the percentage inhibition after treatment with Na2EDTA. Each percentage is calculated on its respective control. Individual values expressed in µkat g-1 of protein are averages of three independent experiments as means ± standard deviation. aP<0.05, bP<0.01 (compared to control values, Student’s t-test).
CAGT activity (µkat g-1) ± (SD) and % inhibition
Day 1 Day 6 Day 13 Day 14 Na2EDTA (mM) Activity Inhibition
(%) Activity Inhibition
(%) Activity Inhibition
(%) Activity Inhibition
(%) 0 13.7(±1.3) 0 2.5 (±0.3) 0 6.6 (±1.0) 0 8.8 (±0.5) 0 0.l 9.6 (±2.4) 30 2.2 (±0.4) 12 6.0 (±2.5) 4 8.5 (±0.4) 14 0.5 10.8 (±2.5) 22 1.9 (± 0.5) 24 2.5 (±0.4) 62a 4.8 (±1.8) 46a 1 7.5 (±1.6) 46a 1.0 (±0.3) 60b 1.9 (±0.3) 72b 3.9 (±0.3) 56b 2 5.2 (±1.3) 62a 1.1 (±0.1) 56b 1.3 (±0.1) 80b 3.1 (±0.2) 65b 5 5.9 (±1.0) 57a 1.2 (±0.4) 52a 1.4 (±0.0) 79b 1.0 (±0.1) 89b
For CAGT purification, the enzyme was extracted from 1 day-old cell suspension cultures (when
the highest CAGT activity was found). The purification procedure, summarized in Table 2, ultimately resulted in a 41.2-fold enhancement of the CAGT activity, 13.1 % recovery of total activity and a product with a specific activity of 256 µkat g-1 of protein. The amount of protein obtained however, was insufficient to carry out incubation experiments with Na2EDTA. Therefore the fraction originating from the desalting step (see Table 2; no. 3, 11.4-fold purified) was used. 5 mM of Na2EDTA inhibited the CAGT activity in this partially purified enzyme preparation, up to 53%.
Table 2. Subsequent preparation steps of the partially purified CAGT preparation. Steps Fraction Protein
(mg) Total activity (µkat)
Specific activity (µkat g-1 protein)
Purification (fold)
Yield (%)
1 Crude extract 302.4 1.52 5 1 100 2 Ammonium sulphate
precipitation 28 1.12 40 8 73
3 Desalting 4.4 0.25 56.8 11.4 16.4 4 HiTrap DEAE FF 0.8 0.20 256 41.2 13.1
CAGT is a glucosyltransferase that converts coniferyl alcohol into coniferin. In order to enhance
the 6-MPT production in L. flavum cell suspension cultures, the formation of coniferin was blocked by inhibition of CAGT using several potential glucosyltransferase inhibitors. Na2EDTA inhibited the production of coniferin in the cell suspension cultures of L. flavum in a concentration-dependent way, in a range of 0.1-5 mM. The results were confirmed by hydrolysis of the coniferin-containing water layer

��������#
70
of the cell extract by enzymatic hydrolysis using �-glucosidase. Coniferin was completely converted into coniferyl alcohol and the concentration of the formed coniferyl alcohol related to the original coniferin concentration. There was a correlation between the coniferin and the 6-MPT production in L. flavum cell suspension cultures. Higher coniferin contents corresponded with lower 6-MPT levels. This supports our hypothesis that blocking the branch leading to the formation of lignins by inhibition of glucosyltransferase may result in an enhanced production of lignans. By inhibiting the coniferin production, coniferyl alcohol accumulates and is available as a substrate to produce of 6-MPT and other lignans. The high 6-MPT content on day 6-9 of the growth cycle (Fig. 4), correlates with the low CAGT content as measured on day 6 (Table 1). This is in agreement with earlier observations (Van Uden et al. 1991b), showing that a maximum coniferyl alcohol content in L. flavum cell suspension cultures was preceded by a maximal activity of the enzyme �-glucosidase. At the beginning of the growth cycle of the control cell suspension cultures no clear relationship existed between CAGT activity and coniferin content. However, after day 6 it appeared that a low activity of CAGT related to a low coniferin content. CAGT activity then increased until day 14, with a simultaneous increase of the coniferin accumulation. The highest CAGT activity in cell suspensions was found on day 1. Then it declined to a lower level on day 6 and re-increased on day 13 and 14. This is probably affected by �-glucosidase that converts coniferin into coniferyl alcohol. The reaction catalyzed by �-glucosidase is opposite to that of CAGT. In L. flavum cell suspension cultures, �-glucosidase activity increased to a maximal value on day 4 of the growth cycle and declined to lowest activity on day 14 (Van Uden et al., 1991b). A high CAGT activity apparently relates to a low �-glucosidase activity. Reduction of the CAGT activity correlated with a reduction of the coniferin production in the cell suspensions. The coniferin production and CAGT activity were reduced to >88% at the end of a growth cycle after treatment with 5 mM Na2EDTA, while control values of the coniferin production were at their maximum at this time point. These results strongly suggest that Na2EDTA inhibits CAGT activity thereby inhibiting the conversion of coniferyl alcohol into coniferin in L. flavum cell suspension cultures. Our hypothesis that Na2EDTA (0.5 mM) is an inhibitor of CAGT activity is further supported by the inhibitory effect on CAGT activity in an 11.4-fold purified enzyme preparation. The effect, in terms of percentage inhibition, however, is less pronounced than in the cell suspensions. This difference may be due to a toxic effect of Na2EDTA on the cell suspension cultures. In conclusion, Na2EDTA appears to be an inhibitor of CAGT activity both in vivo and in situ. Because of a lack of information about the structure and the function of the CAGT it is not yet clear which mechanism underlies the inhibition by EDTA. If CAGT needs a metal ion as a co-factor for its activity, it can be understood that the Na2EDTA complexes with the metal ion, thereby reducing the enzyme activity. Further studies directed to the purification, structure and function determination of the enzyme are in progress. Acknowledgement The research was supported by the QUE Project Batch II, Department of Biology Institut Teknologi Bandung ITB, Indonesia, under the contract No. 3028-IX/P3S-1/KON-QUE II/2000, IBRD LOAN No. 4193 – IND.

71
Chapter 7
Cloning of glucosyltransferase genes from cell suspension cultures of Linum flavum in E. coli

��������'�
72
Abstract Glucosyltransferases (GTs) from cell suspension cultures of Linum flavum were successfully
cloned in E. coli. Total RNA was isolated from the cell suspension cultures at day 1 of subculturing, when the highest activity of GTs was reached. With degenerate primers designed on a conserved region from plant glucosyltransferases, RT-PCR was used to amplify specific cDNA fragments. The sequences of these cDNA fragments showed strong similarity with those encoding for GTs in various other plant species, previously deposited in the databases. Using primer extension and inverse PCR the full length gene was amplified from genomic DNA extracted from Linum flavum cells. Based on phylogenetic analysis, one of the cloned GTs belongs to Group D, Family 1 of glucosyltransferases that are active for phenylpropanoid compounds. This is the first time that a gene involved in the lignan biosynthesis from L. flavum is cloned.

Cloning of glucosyltransferase genes from Linum flavum cell suspension cultures in E. coli
73
Introduction Podophyllotoxin, an aryltetraline lignan with antiviral and antineoplastic activities, is used as a
precursor for the semi-synthesis of established cancer therapeutics, i.e. etoposide, teniposide and etopophos. The use of plants as a source of podophyllotoxin is limited due to difficulties in the (large scale) cultivation of the endangered Podophyllum hexandrum (Gordaliza et al., 2004). There is a need to find alternative sources for podophyllotoxin. Biotechnological and enzymatic approaches have been considered as a potential solution for the problem. Petersen and Alfermann (2001) described the state of the art of the use of plant cells and organ cultures as a strategy in this respect. In vitro production of podophyllotoxin and 6-methoxypodophyllotoxin and related lignans has been shown in cell cultures of Podophyllum and Linum species (Woerdenbag et al., 1990, Empt et al., 2000, Chattopaday et al., 2002, Berim et al., 2005). An understanding of the metabolic pathway of lignans and the cloning of the corresponding genes may allow genetic modifications leading to a higher production of lignans. In this respect the branch of lignan and lignin biosynthesis after the common early pathway opens an interesting possibility. Reducing the lignin biosynthesis may lead to a channelling of precursors towards the synthesis of lignans (Gordaliza et al., 2004).
Cell suspension cultures of Linum flavum (family Linaceae) accumulate high amounts of coniferin that is synthesized from coniferyl alcohol by glucosyltransferases (GTs). Coniferyl alcohol is a precursor of coniferin and lignans through different biosynthetic branches (see Fig. 1) (Van Uden et al., 1991). Inhibition of the coniferin production by inhibition of GTs could lead to the accumulation of coniferyl alcohol that subsequently leads to an increase in the production of cytotoxic lignans. GTs are a large group of enzymes that catalyze the transfer of a sugar moiety from an activated donor onto saccharide or non-saccharide acceptors. These enzymes play a key role in the biosynthesis of natural products and in recent years the information on GTs of small molecules has increased enormously.
Many recombinants GTs have been characterized from different plant species (Vogt and Jones 2000, Bowles et al., 2005). From the genome sequence of Arabidopsis thaliana, more than 110 GTs have been identified and based on a phylogenetic tree, they were grouped into 12 distinct classes (Li et al., 2001, Bowles et al., 2005). Several important natural products have been used as the substrate for classification such as curcumin (Kaminaga et al. 2004), phenylpropanoids (Taguchi et al., 2001, Chong et al., 2002), dihydroxyphenylalanine (DOPA) (Sasaki et al., 2005), steroidal alkaloids (Kohara et al., 2005) anthocyanins (Imayama, et al., 2004, Lorenc-Kukula et al., 2004), coumarins and flavonoids (Taguchi et al., 2001). Glucosylation has biological significance influencing toxicity, bioavailability, solubility, adsorption, and metabolism of compounds. It also enables the storage of potent toxic and deterrent chemicals at high concentration in vacuoles (Lorenc-Kukula et al., 2004, Bowles et al., 2005).
The detection of coniferin in L. flavum cells cultures (Van Uden et al., 1991) indicates the presence of one or more GTs are involved in its synthesis from coniferyl alcohol. Based on conserved regions we have designed PCR primers for the amplification of GT genes of group D of family 1 from mRNA isolated from lignan synthesizing L. flavum cells. The conserved region, called the PSPG box, is highly characteristic and present in all GTs involved in natural product biosynthesis. This domain may also define the active site of GTs of animals and microorganisms. The PSPG box is considered to represent the nucleotide diphosphate binding site. In this study we report the isolation of glucosyltransferase cDNAs from cell suspension cultures of L. flavum, and the cloning of the corresponding genes in E. coli.
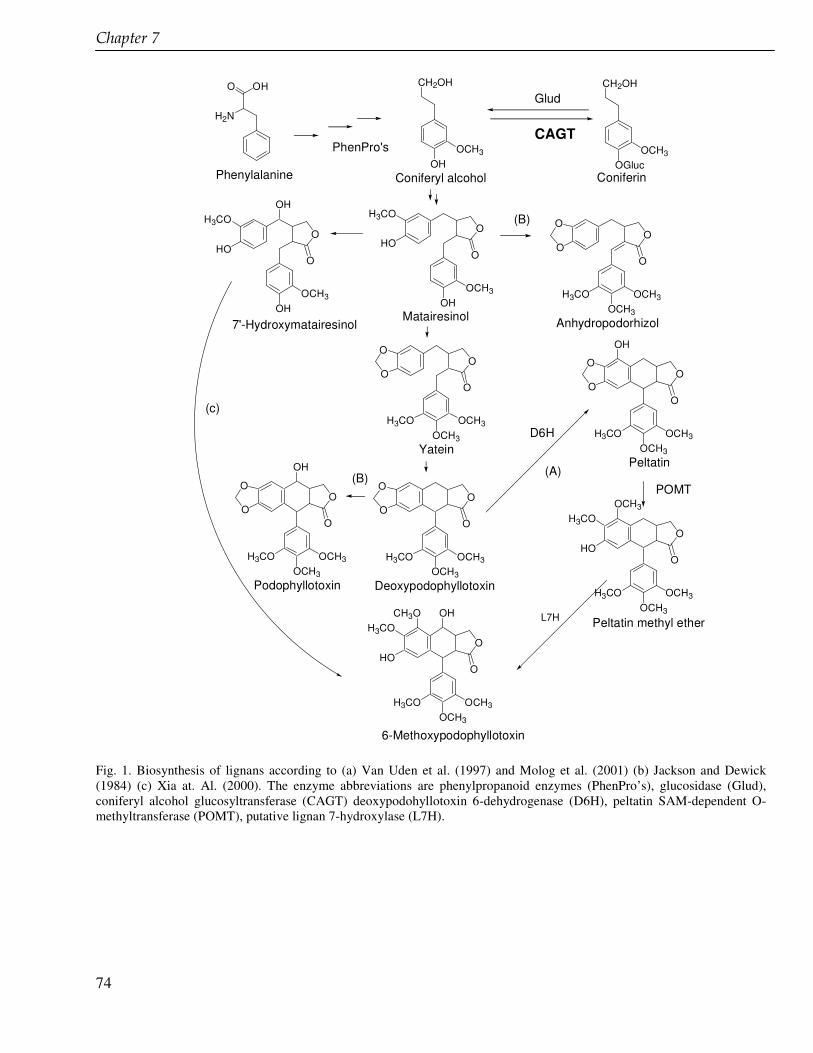
��������'�
74
OH3CO
HO
OCH3
OCH3
O
CH3O OH
H3CO
O
OCH3
OCH3
O
O
O
H3CO
OH(B)
O
OCH3
OCH3
O
O
O
H3CO
OH3CO
HO
OHOCH3
O
OH
O
OCH3
OCH3
O
O
O
H3CO
OH3CO
HO
OCH3
OCH3
O
OCH3
H3CO
(A)
D6H
OHO
H2N
OH3CO
HO
OHOCH3
O
POMT
O
OCH3
OCH3
O
O
O
H3CO
OH
(B)
CH2OH
OHOCH3
O
OCH3
OCH3
O
O
O
H3CO
CH2OH
OGlucOCH3
CAGT
Glud
PhenPro's
Phenylalanine Coniferyl alcohol Coniferin
Matairesinol
Yatein
DeoxypodophyllotoxinPodophyllotoxin
Peltatin
7'-Hydroxymatairesinol Anhydropodorhizol
Peltatin methyl etherL7H
6-Methoxypodophyllotoxin
(c)
Fig. 1. Biosynthesis of lignans according to (a) Van Uden et al. (1997) and Molog et al. (2001) (b) Jackson and Dewick (1984) (c) Xia at. Al. (2000). The enzyme abbreviations are phenylpropanoid enzymes (PhenPro’s), glucosidase (Glud), coniferyl alcohol glucosyltransferase (CAGT) deoxypodohyllotoxin 6-dehydrogenase (D6H), peltatin SAM-dependent O-methyltransferase (POMT), putative lignan 7-hydroxylase (L7H).

Cloning of glucosyltransferase genes from Linum flavum cell suspension cultures in E. coli
75
Materials and methods Plant material and culture conditions
Cell suspensions of Linum flavum L. (Linaceae) leaves have been initiated and are maintained at the Department of Pharmaceutical Biology, University of Groningen. The cell suspensions are cultured routinely every two weeks by transferring 100 mL of a fully grown suspension aseptically into 200 mL of fresh liquid medium. The medium contains MS medium (Murashige and Skoog,1962), 0.2 mg of indole-3-acetic acid (IAA) and 0.2 mg of 6-benzylaminopurine (BAP), purchased from Duchefa, Haarlem, the Netherlands. The cell suspensions were incubated on a rotary shaker (175 rpm) at 26°C under a day/night regime (16/8 h: 3,000 lux, day light L 36W/10, OSRAM, Germany). Isolation of plant RNA, DNA and cDNA construction
One day old cell suspension cultures were harvested, filtered and the remaining cell material was put directly into liquid nitrogen and grinded. 500 mg of powdered cell material were used for total RNA isolation using nucleospin RNA plant isolation kits (Macherey-Nagel, GmbH & Co., Düren, Germany). cDNA was constructed from 2 µg total RNA using Omniscript RT (Qiagen GmbH, Hilden, Germany). Genomic DNA was isolated from 30 g of powdered cell material using the cetyltrimethyl ammoniumbromide (CTAB) method. 30 mL of extraction buffer containing 350 mM sorbitol, 100 mM Tris HCl pH 7.5, 5 mM EDTA and 20 mM sodium bisulfide (added just before use) were added to the cell frozen material, mixed until thawn and resuspended. The mixture was centrifuged for 5 min at 3000 x g. The supernatant was separated and the pellet was resuspended using extraction buffer (3.5 mL) and nucleus lysis buffer (5 mL). The nucleus lysis buffer contained 200 mM Tris HCl pH 7.5, 50 mM EDTA, 2 M sodium chloride and 2% (w/v) CTAB. To the suspension, 5% sarkosyl was added and the mixture was incubated at 65oC for 1 h. The mixture was extracted using 8 mL chloroform : isoamylalcohol (24:1) and rotated for 30 min. The upper layer was separated, 5 mL of isopropanol were added and mixed thoroughly for 5 min at room temperature and subsequently centrifuged for 10 min (3000 x g). The pellet was washed using 70% ethanol and dissolved by 1 mL TE buffer with 1 µL RNAse, then incubated at 65oC for 10 min.
PCR amplification
Alignment of 106 sequences of GTs from various other plant species deposited in the NCBI database using MegAlign was performed to identify regions of conserved amino acid sequences. The region between position 254 and 298 and the region between 342 and 386 (44 amino acids) were found to be the most conserved. The following two degenerated primers were designed for amplification of GT fragment from L. flavum cDNA: Pgt-1f: 5′-GGNWTRMSRNVVDNNNTGGGCHCCGCA-3′ and Pgt-1r: 5′-TGCCGATGGTGNCNTGGCCHTT NTTRGCNGANCA-3′. PCR amplification of cDNA was performed using a Mastercycler gradient thermocycler (Eppendorf) in 50 µL containing 50 ng cDNA, 200 µM dNTP and 1.25 units of Taq polymerase (Fermentas GMBH, St. Leon-Rot, Germany). The PCR program was denaturation at 95oC for 30 s, annealing at 60oC for 40 s, and extension at 72oC for 2 min. The PCR product was cloned into the pGEM-T easy vector system I (Promega, Madison, USA) and the sequences were determined with a Megabase Sequencer (a Alf Express II using ThermoSequenase fluorescent primer cycle kit, Amersham Pharmacia, Sweden).

��������'�
76
Rapid amplification of the cDNA ends (RACE)
The amplification of the 3′- and the 5′-ends of two coniferyl alcohol glucosyltransferase (CAGT) candidates was performed using 3′ and the 5′ RACE kits (Invitrogen, Carlsbad, Canada). Two sets of primers were designed (Table 1) based on the sequences found in the first PCR round. Pgt-2r (1-3) and Pgt-3f (1-3) were used for the amplification of the 3′- and 5′ ends of CAGT-A, respectively. Pgt-4r (1-3) and Pgt-5f (1-3) were used for the amplification of the 3′- and 5′ ends of CAGT-B, respectively. The applied touchdown PCR program was initial denaturation at 95oC for 3 min, one cycle; denaturation at 95oC for 40 s, annealing at 68oC for 40 s, extension at 72oC for 3 min, 95oC for 40 s, 42oC for 2 min, and extension at 72oC for 3 min, 5 cycles; 94oC for 40 s, 72oC for 3 min, 5 cycles; 94oC for 40 s, 72oC for 3 min, 25 cycles; 95oC for 40 s, 68oC for 2 min, and extension at 72oC for 3 min and the last extension 72oC for 5 min. PCR products were cloned into pGEM-T easy. The sequences were determined.
Inverse PCR (iPCR)
10 µg of genomic DNA were digested using HaeIII in total volume of 100 µL, then concentrated using sodium acetate precipitation. HaeIII was inactivated by heating at 60oC for 10 min. 10 µL were ligated using T4 DNA ligase and purified using a QIA quick gel extraction kit (Qiagen GmbH, Hilden, Germany). Two sets of primers were designed based on the first PCR products. They were Pgt-3f3, Pgt-2r3 for CAGT-A and Pgt-5f3, Pgt-4r3 for CAGT B. The applied PCR program was initial denaturation at 94oC for 5 min, 10 cycles; denaturation at 94oC for 15 s, annealing at 60oC for 40 s, and extension at 72oC for 3 min, 25 cycles; at 94oC for 15 s, 60oC for 30 s, 72oC, plus 5 min at 72oC per cycle, the last extension at 72oC for 7 min. PCR products were cloned into pGEM-T easy. The sequences were determined. The second series of iPCR was done based on the obtained sequences. Genomic DNA was digested using DpnI, and ligated. The primers were Pgt-6f, Pgt-6r for CAGT-A and Pgt-5f2, Pgt-8r for CAGT B. The last iPCR series was done using the restriction enzyme MseI, and the primers Pgt-6f, Pgt-7r for CAGT-A and Pgt-9f, Pgt-8r for CAGT B. All PCR products were cloned into pGEM-Teasy and the sequences determined. (All restriction enzymes were purchased from New Englands Biolabs.)
Table 1. The nucleotide sequences of primers used to amplify the sequence of glucosyltransferases. Primer Nucleotide sequence Length (bp) 5′ 3′ Pgt-2r1 TAAAGGCCAAGTCACCATCGGCACACCCCGCCGT 35 Pgt-2r2 ACACGCTCTCCAGCGTCGAGTTCCACCCGCAAT 34 Pgt-2r3 ACTCCAACGCATCCATGGCGAAGAATCCTCA 30 Pgt-3f1 TGAGAGTTAAAGTGGGCACCGCAGCTGAGGA 32 Pgt-3f2 CGCCATGGATGCGTTGGAGGGTTCGTGACTCATTGC 36 Pgt-3f3 TGGAACTCGACGCTGGAAGGGGTGAC 27 Pgt-4r1 AGCAAACAATGGCCAGGGCACCATCGGCACCCCGT 35 Pgt-4r2 CAGCGTCGAGTTCCACCCGCAATGAGTCACGAACC 35 Pgt-4r3 TATTGGCGCACAAGGCAGTGGGAGGATTCGTGT 33 Pgt-5f1 ACACGAATCCTCCCACTGCCTTGTGCGCCAATA 34 Pgt-5f2 AGTCGAGGTATTGGCGCACAAGGCAGTGGGAGGA 34 Pgt-5f3 TGGAACTCGACGCTGGAGAGCGTGTGGAAC 30 Pgt-6r GTATTACCAACCCTCTCCCTTCCGTTCTCTCCTCAA 36 Pgt-6f ATGGTGACATGGCCGCTTCAGGCGGATCAA 30 Pgt-7r GATGGAGGAAGCAGAGGAGAGAAGCTA 28 Pgt-8r TGACATTCGCCGGACTTACAGAATTTCACT 30 Pgt-9f TTGTACGCGGAGCAGCAGTGCAATGCGTTTCA 32

Cloning of glucosyltransferase genes from Linum flavum cell suspension cultures in E. coli
77
Results and discussion The total RNA isolation of the cell suspension cultures of Linum flavum provided high amounts
of RNA. Checking the product by gel electroforesis yielded two intensive bands with the size of 1900 and 4700 bp respectively, probably representing ribosomal RNA. The PCR product of cDNA, using primers that were designed based on the conserved region of the amino acids, called PSPG box, was 130 bp in length. Sequences of 12 randomly picked clones of this products showed homology with GTs. On the basis of the sequence they were divided into 3 different candidate groups and called CAGT A, B and C. The use of the 3′ and the 5′ RACE methods to find the full lengths of CAGT A, B and C did not provide the expected fragments. Due to these problems, iPCR amplifications were done to find the full length. This method is a normal PCR that has the primers oriented in the reverse direction of the usual orientation. iPCR permits the amplification of the regions that flank any DNA segment of known sequence, either upstream or downstream or both (Ochman et al., 1988). Restriction enzymes that were used to digest genomic DNA should not cut the internal known sequence. The first iPCR products extended to 400 bp of CAGT A and 350 bp of CAGT B both at the 5′ end. No extension was found for CAGT C. These sequenced were extended using a second round of iPCR to 774 for CAGT A and CAGT B 563 of CAGT B, both the extensions are to the 5′. After the third iPCR the full length of both CAGT was found with additional upstream and downstream sequences. Start and stop codons were determined. From the alignment with known GT sequences it can be concluded that CAGT A and CAGT B do not harbour any intron sequences.
It has been suggested that GTs are highly regiospecific (or regioselective) rather than substrate specific, e.g. uridine 5′-diphosphoglucose 5-O- and 6-glucosyltransferases. Both GTs catalyze the transfer of glucose not only to the hydroxyl groups of betanidin but also to those of flavonoids, i.e. flavonols, anthocyanidins and flavones (Vogt et al., 1997). This was also shown by UGT72E2 from Arabidopsis thaliana that has broad substrate recognition. It showed activity towards coniferyl alcohol, sinapyl alcohol, coniferyl aldehyde and sinapyl aldehyde, in contrast to UGT72E1 that is highly specific to coniferyl aldehyde and sinapyl aldehyde (Lim et al., 2005). Phylogentic analysis (Fig. 4) using MegAlign software from DNASTAR showed that CAGT A belongs to group E of glucosyltransferase family 1. Most of GTs from this group have substrate specificity on phenylpropanoid compounds. It indicates that CAGT A may be active to phenylpropanoid compounds. Based on the enrichment of mRNA and the phylogenetic analysis, we conclude that CAGT A may be coniferyl alcohol glucosyltransferase. Expression experiments are needed to confirm this.

��������'�
78
1 atggagcact cacagcagct tcacgtcgtc tttctcccgt acatggcgca cggccacatg m e h s q q l h v v f l p y m a h g h m 20 61 acccccttcg ccgacatggc aagactcttc gcccgccacg gcgccaagtc gaccatcatc t p f a d m a r l f a r h g a k s t i i 40 121 accactcccc tcaacgcccc tttcttctcc ggcaagatcc tcgccgacgt tcaacaacta t t p l n a p f f s g k i l a d v q q l 60 181 ggccttcaaa tccgaatcca catcgtcgac ttcccttccg ccgccgccgg cctccccgaa g l q i r i h i v d f p s a a a g l p e 80 241 ggctgtgaga acgtccgctc cgctatccaa tcccctgacg ccggccacga catcttcttc g c e n v r s a i q s p d a g h d i f f 100 301 aaattcataa cctccatgga ccatttccag cgcccggtgg aggagctcct ccagcagtgg k f i t s m d h f q r p v e e l l q q w 120 361 cggcggactg catcgtcgcc gacttcgtct tccactgggc tgaccgaatc agcccgccgg r r t a s s p t s s s t g l t e s a r r 140 421 ctcggcatcc cgaggctcta cttcaacggg atggggacgc tcgccatgtg ttgtacaatt l g i p r l y f n g m g t l a m c c t i 160 481 gcctcaaacg ctaccagcct cacaaaaacc gtggactccg attccgagcc acgttctcgt a s n a t s l t k t v d s d s e p r s r 180 541 cccccggggc cggggctttg gcagacggaa agaatacccg ggtttcacgc cggcgaggct p p g p g l w q t e r i p g f h a g e a 200 601 gtcgagttca acaagcagct gcagcggatg gaggaagcag aggagagaag ctacggaact v e f n k q l q r m e e a e e r s y g t 220 661 ctggtgaaca gcttcctcga gctcgaacct gggtatccgg agtatttcag gaaagtgatg l v n s f l e l e p g y p e y f r k v m 240 721 gggagaaggg cgtggttcgt cggtcctctg ttgctctgca acaaggacct gcaacaagga g r r a w f v g p l l l c n k d l q q g 260 781 cggggaaggg gaatctgccg ccattgttgt acacgaatgc ctaaatggct cgactcgaag r g r g i c r h c c t r m p k w l d s k 280 841 aaactggcca attcagtcat ctacatatgc ttcggcagcg aggctgtctt ctctgctgcg k l a n s v i y i c f g s e a v f s a a 300 901 cagttaaacg agatcgctgc agctctggcg gaatcggagc agagcttcat ctgggtagtg q l n e i a a a l a e s e q s f i w v v 320 961 aaggaagaat cgctgccgga agggtttgag gagagaacgg aagggagagg gttggtaata k e e s l p e g f e e r t e g r g l v i 340 1021 cggggatggg caccccagct gaggattctt cgccatggat gcgttggagg gttcgtgact r g w a p q l r i l r h g c v g g f v t 360 1081 cattgcgggt ggaactcgac gctggaaggg gtgacggcgg gggtgccgat ggtgacatgg h c g w n s t l e g v t a g v p m v t w 380 1141 ccgcttcagg cggatcaatt cgcaaacgag aagttggtga cggaggttct gaagatcggg p l q a d q f a n e k l v t e v l k i g 400 1201 gtcggagtcg gagctgagga atggtcgacg ggcgagagga ggatcgtggt gggaagggaa v g v g a e e w s t g e r r i v v g r e 420 1261 ggtataaaga gtgcggtaag ccgggtgatg gttggtgcag agggtggaga gatgaggggc g i k s a v s r v m v g a e g g e m r g 440 1321 agagttaagg agcttgcaga aagggctgcc atggcgatgg attaa r v k e l a e r a a m a m d - 460 Fig. 2. Nucleotide and deduced amino acid sequences of CAGT A of Linum flavum cell suspension cultures. The nucleotide sequence is numbered on the left. The amino acid sequence is given in the single-letter code and is numbered on the right.

Cloning of glucosyltransferase genes from Linum flavum cell suspension cultures in E. coli
79
1 atgaaccaga cagttcatct agctttcgtc ccggctcccg gaatcggcca tctcgtctcc m n q t v h l a f v p a p g i g h l v s 20 61 acaatcgaat tcgcccgccg cttgctccgc cgcgacacca ccatctccgt cctcatcctc t i e f a r r l l r r d t t i s v l i l 40 121 ctcatcaagt tcccgccgcc gttcggcgac gacatcgaca gcttcgtcaa atcaatttcc l i k f p p p f g d d i d s f v k s i s 60 181 ggcgaggacg atgaagatca ccgccgcgtc gaattcgtca ctctccccca actactcccc g e d d e d h r r v e f v t l p q l l p 80 241 cctgcttcct cctcctccgc tggagattct tccaaatcgc cggaggcttt cgtcacacag p a s s s s a g d s s k s p e a f v t q 100 301 gccgctagtc aaagaagcaa ttgtaaaccc tgctctgccc cggtcaccgg tctggtcgtt a a s q r s n c k p c s a p v t g l v v 120 361 gacttgttct gtacttccat gatcgacgtc gccgacgagc taggaatccc ttcctacatc d l f c t s m i d v a d e l g i p s y i 140 421 ttcttcactt cctcaattgc gtttctaggg ttcatgctct acctcccgac ccgccacgac f f t s s i a f l g f m l y l p t r h d 160 481 cgggtcggat ctgtattcga gctgaccgac gacccggtgc cggtgcccag ctactccgac r v g s v f e l t d d p v p v p s y s d 180 541 ccgtttcctt ctcgggtcct gccgtccgtc ttcctcaaca agcacggcgg gtacacgacg p f p s r v l p s v f l n k h g g y t t 200 601 atgatgaacc acggccggag attctcggaa gctaagggca taatcgtcaa ttcatttgcc m m n h g r r f s e a k g i i v n s f a 220 661 gagctggagc cgtacgcttt gaaatcactg atctcctcct cgtttactct gccgccgccg e l e p y a l k s l i s s s f t l p p p 240 721 ccggtctacg ccccgattct ggatttgaag gctcaggggc aagtgaaatt ctgtaagtcc p v y a p i l d l k a q g q v k f c k s 260 781 ggcgaatgtc aggagatcgt gaggtggctg gacggtcagc cggaggagtc ggtgatcttc g e c q e i v r w l d g q p e e s v i f 280 841 ctctgcttcg ggagcatggg agctttcgag aaggatcaat tgagggaaat cgctacaggg l c f g s m g a f e k d q l r e i a t g 300 901 ctggagcgga gtggctgccg gttcctctgg tcgatccgga aacctcctcc ggcggaagag l e r s g c r f l w s i r k p p p a e e 320 961 ttctcgatgc cggcggatta tggagggagt tacggggaga tcttgccgga agggtttgag f s m p a d y g g s y g e i l p e g f e 340 1021 gacaggacga gagggttggg tttgatctgc aaatggcgcc gcaagtcgag agtcgaggta d r t r g l g l i c k w r r k s r v e v 360 1081 ttggcgcaca aggcagtggg aggattcgtg tcgcattgcg ggtggaactc gacgctggag l a h k a v g g f v s h c g w n s t l e 380 1141 agcgtgtgga acggggtgcc gatggtggcg tggcctgttg tacgctggat gcagcagtgc s v w n g v p m v a w p v v r w m q q c 400 1201 aatgcgtttc agctggctga gtggagctgg tgtgctggct gggtggatgc tgagtgctgg n a f q l a e w s w c a g w v d a e c w 420 1261 actaccggat taagatgggc ggagtatgaa gaagcaggag acggcgtgag tgaggagttg t t g l r w a e y e e a g d g v s e e l 440 1321 tttactggcg gagtgagatt gagaaggcct gtaaaatgtg tgatggatag gggaagcgcg f t g g v r l r r p v k c v m d r g s a 460 1381 gtgaggaaga gggcgaagga gatgggctga v r k r a k e m g - 469 Fig. 3. Nucleotide and deduced amino acid sequences of CAGT B of Linum flavum cell suspension cultures. The nucleotide sequence is numbered on the left. The amino acid sequence is given in the single-letter code and is numbered on the right.

��������'�
80
AB070753 Vigna angularis AB176523 Nicotiana tabacum AY345975 Stevia rebaudiana AC013430 Arabidopsis thaliana AC006551 Arabidopsis thaliana
AB191249 Dianthus caryophyllus (chalcon) AB017060 Arabidopsis thaliana
AC008153 Arabidopsis thaliana AB025604 Arabidopsis thaliana
AC006922 Arabidopsis thaliana AC005851 Arabidopsis thaliana AB047095 Vitis vinifera (flavonoid) AY695816. Fragaria x ananas (flavonoid)
AC009917 Arabidopsis thaliana NM_190981 Oryza sativa NM_196703 Oryza sativa
AF331854 Zea mays AF503433 Sorghum bicolor
AF303396 Phaseolus vulgaris AC011664 Arabidopsis thaliana
AC004165 Arabidopsis thaliana AC004165.2 Arabidopsis thaliana
AP003232 Oryza sativa AY663786. Fragaria x ananas DQ289587. Fragaria x ananas
AB072918 Nicotiana tabacum AY345976. Stevia rebaudiana AB025634 Arabidopsis thaliana
CAGT B Linum flavum AL049862 Arabidopsis thaliana
AY345983 Stevia rebaudiana AC016662 Arabidopsis thaliana
AB182387 Solanum aculeatissimum AP000373 Arabidopsis thaliana
AF190634 Nicotiana tabacum (salicylic acid) AJ889012 Lycopersicum esculentum (phenolic)
AY345982 Stevia rebaudiana AC002396 Arabidopsis thaliana AC002333 Arabidopsis thaliana AC006533 Arabidopsis thaliana
AC007153 Arabidopsis thaliana AF287143 Brassica napus (sinapate) AM231594 Brassica napus (hydroxycinnamate)
AC005106 Arabidopsis thaliana AF346431 Nicotiana tabacum (phenylpropanoid) U32644 Nicotiana tabacum (salicylate) AF346432 Nicotiana tabacum (phenylpropanoid)
U32643 Nicotiana tabacum (salicylate) AL021961 Arabisopsis thaliana CAGT A Linum flavum AC006282 Arabidopsis thaliana AC005167 Arabidopsis thaliana AC005489 Arabidopsis thaliana
XM_450574 Oryza sativa (betanidin) AB018115 Arabidopsis thaliana AL035602 Arabidopsis thaliana AB192315 Ipomoea purpurea (antochyanin)
AC006340 Arabidopsis thaliana Group A
Group B
Group C
Group D
Group E
Group F
Group G
Group H
Group I
Group J
Group L
Group K
Fig. 4. Phylogenetic analysis of CAGT A, B and other plant glucosyltransferases. Grouping is based on phylogenetic analysis of Arabidopsis thaliana which showed 12 distinct groups. Each glucosyltransferase contains accession number, plant name and substrate (in the bracket, if any).

Cloning of glucosyltransferase genes from Linum flavum cell suspension cultures in E. coli
81
Acknowledgement The research was supported by the QUE Project Batch II, Department of Biology Institut Teknologi Bandung ITB, Indonesia, under the contract No. 3028-IX/P3S-1/KON-QUE II/2000, IBRD LOAN No. 4193 – IND.

82

83
Chapter 8 Production of a cytotoxic arylnaphthalene lignan using genetically transformed root cultures of Linum leonii
Nikolay Vasilev, Elfahmi, Rein Bos, Oliver Kayser, Georgi Momekov, Spiro Konstantinov, Herman J. Woerdenbag, Wim J. Quax, Iliana Ionkova This chapter is based on Journal of Natural Products, in press (2006)

��������(�
84
Abstract Callus and hairy roots cultures of Linum leonii F.W. Schulz. were established. The genetic
transformation in hairy roots was proven by PCR analysis, which showed integration of rol A and rol C genes into the plant genome. Callus and hairy roots accumulated the arylnaphthalene lignan justicidin B as a major constituent. Hairy roots produced 5-fold higher yields of justicidin B (10.8 mg g-1 DW) compared to callus. Justicidin B demonstrated strong cytotoxicity to the chronic leukemic cell lines LAMA-8, K-562 and SKW-3 with IC50 values of 1.1, 6.1 and 1.6 �M, respectively. Apoptotic properties of justicidin B were reported for the first time.

��������������������������������������������������������������������� ������������������� �����������������
85
Introduction Justicidin B is an arylnaphthalene lignan which exerts cytotoxic (Joseph et al., 1988, MacRae et
al., 1989), antiviral (Gertsch et al., 2003), fungicidal, antiprotozoal (Chen et al. 1996) and antiplatelet properties (Baba et al., 1996). Several tumor types including sarcomas and breast, prostate, and lung carcinomas grow in or preferentially metastasize the skeleton where they proliferate, and induce significant bone remodelling, bone destruction, and cancer pain (Mohagheghzadeh et al., 2002). Thus, justicidin B may have significant clinical utility as a lead compound in the management of bone cancer and osteoclastogenesis, due to its cytotoxic and bone resorption inhibitory properties. The potent bone resorption inhibitor justicidin B was used as a lead compound for design of new antirheumatic drugs (Sabino et al., 2002).
Justicidin B was first isolated from Justicia spp. (Acanthaceae) and Haplophyllum spp. (Rutaceae) (Okigawa et al., 1970, Pettit and Schaufelberger, 1988). Justicidin B has further been isolated from different Phyllanthus species (Euphorbiaceae) (Bachmann et al., 1993). It was shown that cell cultures of Linum austriacum produce justicidin B, which is the first report on the existence of arylnaphthalene lignans in a species of the Linaceae (Mohagheghzadeh et al., 2002). Since there is a growing interest in justicidin B due to its various pharmacological effects, the sustainable biotechnological supply of this valuable lignan would be a feasible alternative.
Our preliminary experiments exhibited that justicidin B is the main cytotoxic principle in the methanolic extract of callus of Linum leonii F.W. Schulz. (Linaceae) (Vasilev and Ionkova, 2005). Therefore, we decided to establish hairy roots from this species in hope of produce justicidin B in high yields. This paper describes the isolation, structure elucidation and cytotoxic evaluation of the major lignan produced by conventional and genetically transformed cultures of L. leonii.
H3CO
H3COO
OO
O1'2 '
3 '
4'5 '
6 '
7'
71
22
3
45
8 '
8
Fig. 1. Chemical structure of justicidin B.
Material and methods General experimental procedures
NMR spectra measurements were carried out on a Bruker WM 400 (400 MHz) and a Bruker DRX 500 (500 MHz) spectrometer in CDCl3. TMS was used as an internal standard. GC analysis was performed on a Hewlett Packard 5890 series II gas chromatograph equipped with a 7673 autosampler, and a Hewlett Packard 3365 Chemstation software A10.02 under the following conditions: column: WCOT fused silica CPsil 5 CB lowbleed/MS, # CP7810; 15 m x 0.25 mm ID; film thickness 0.10 �m; (Varian Middelburg, The Netherlands); temperature program: 150°C – 320°C at 15°C min-1 injector temperature 250ºC; detector (FID) temperature 300ºC; carrier gas: helium; inlet pressure 125 kPa; linear gas velocity 40 cm s-1; split ratio 100:1; injected volume 1 �L. GC-MS analysis was performed on a

��������(�
86
Shimadzu QP5000 GC-MS system equipped with a 17A GC, an AOC-20i autoinjector, and the GC-MS solution software 1.10. GC conditions: WCOT fused silica CPsil 5 CB lowbleed/MS, # CP7810, film thickness 0.10 �m; 15 m x 0.25 mm ID (Varian Middelburg, The Netherlands). Temperature program: 150°C – 320°C at 15 °C min-1. Injector temp.: 275°C; inlet pressure: 75 kPa; column flow: 2.1 mL min-
1: linear velocity: 75.5 cm sec-1 ; split ratio: 20:1; total flow: 46.2 mL min-1; carrier flow (He): 46.2 mL min-1; injection volume: 2 µL. temperature program 35 min at 90°C, 90-170 °C at 4°C min-1. MS conditions: ionization energy, 70 eV; ion source temperature, 250°C; interface temperature, 250°C; scan speed, 2 scans s-1; mass range, 34-600 u
Plant material
The seeds of Linum leonii F.W. Schulz. (Linaceae) were a kind gift (No 1636; No com. 253/1999) from the botanical garden Nancy (France). Callus cultures were initiated and grown as described previously (Vasilev and Ionkova, 2005). Hairy roots were induced by direct incubation of segments from sterile grown plants with Agrobacterium rhizogenes strain ATCC 15834 cultured in YMB medium in the presence of 20 �M acetosyringone for 2 days in the dark, which increased susceptibility toward infection. The fast growing hairy roots were further maintained under permanent dark on a rotary shaker (80 rpm) and refreshed by a new medium every two weeks. Hairy root cultures were maintained at 25 ± 1 °C as described in Mohagheghzadeh et al. (2002). DNA analysis
DNA isolation was conducted from the dry plant material of the intake plant, calli and hairy roots according to a protocol for rapid isolation from dry and fresh samples (Khanuja et al., 1999). The isolation of DNA of A. rhizogenes ATCC 15834 was performed following the instructions of Qiaprep spin miniprep kit from Qiagen (Westburg, BV, Leusden, The Netherlands). The integration of rol A and rol C genes from A. rhizogenes into the plant genome, which is the genetic evidence for hairy roots transformation, was proven by PCR reaction. Therefore the following specific primers were designed (Nader et al., 2004): for rol A gene, nucleotide positions 21-42 (5’-CGTTGTCGGAAT-GGCCCAGACC-3’) and 268-246 (5’-CGTAGGTCTGAATAT-TCCGGTCC-3’), totally 248 bp; for rol C gene, positions 51-70 (5’-TGTGA-CAAGCAGCGATGAGC-3’) and 550-531 (5’-GATTGCAAACTTGCACTCGC-3’), a fragment of 490 bp totally. Vir D2 gene is not involved in the plant genome during the transformation. The specific primers for the detection of vir D2 are: primer A (5’-ATGCCCGATCGAGCTCAAGT-3’) and primer E (5’-CCTGACCCAAACATCTCG-GCTGCCCA-3’), ending in a fragment of 338 bp (Hass et al., 1995). All PCR reactions were performed in a Mastercycler gradient thermocycler (Eppendorf) with recombinant taq DNA polymerase (Fermentas GMBH, St. Leon-Rot, Germany). The PCR program was 5 min at 95°C, 35 cycles of 30 s at 95°C, 40 s at 50°C, 2 min at 72°C and a final step of 5 min at 72°C. Extraction, isolation and quantification
Air-dried plant material from hairy roots (10 g) was extracted with 80% MeOH (200 mL for 1 h sonification at 25ºC). The extract was separated with 3 x 200 mL dichloromethane. Dichloromethane layers were filtered (Na2SO4 was used as a drying agent), combined, concentrated under reduced pressure at 50ºC, dried and kept at -20ºC. The initial amount of hairy roots yielded 780 mg dry dichloromethane extract. The dichloromethane extract was subjected to preparative TLC separation using silica gel 60 F254 (Merck): 10 x 20 cm, 2 mm, toluene: acetone 10:1, development length: 9 cm and = 254 nm. The most abundant fraction (Rf = 0.45) was pooled and evaporated to dryness consequently. The residue was further purified by recrystallisation in cold MeOH to yield 3.1 mg justicidin B. Quantitative determination of justicidin B in callus and hairy roots was performed by GC

��������������������������������������������������������������������� ������������������� �����������������
87
analysis as described by Koulman et al. (2001). Cinchonidine was used as an internal standard. The response factor (RF) was calculated using 3 concentration ratios (3:1, 1:1, 1:3) between justicidin B and cinchonidine; RF=1.80 (CV=1.11%, n=5). The limit of detection (LOD) was established as the amount of analyte that provided a signal-to-noise ratio of 3. LOD was 0.1 µg mL-1. The limit of quantification (LLOQ) was defined as the lowest calibration standard that could be quantified with an accuracy of 90-110% and a precision of 15%. LLOQ of justicidin B was 1 µg mL-1. Intraday (n=6) and interday (n=5) coefficients of variations (CV) were determined. Intra-day CV from callus and hairy roots determinations are 7.4 and 4.9% respectively, and inter-day variations for callus and hairy roots analyses are 1.8 and 3.4% respectively.
Leukemic cell lines and culture conditions
The three leukemic cell lines LAMA-84, K-562 and SKW-3 were supplied from the German Collection of Microorganisms and Cell Cultures (DSMZ). The culture conditions are as previously described (Vasilev et al., 2005). Cytotoxicity assay
The MTT-dye reduction assay was carried out as described by Mossmann (1983) with some modifications (Konstantinov, 1999). The clinically applied epipodophyllotoxin derivative etoposide was used as reference cytotoxic drug. Briefly, 100 µL aliquots of cell suspension (1×105 cells mL-1) were seeded in 96-well microplates. Following 24 h incubation at 37°C the cells were exposed to the newly isolated lignan or to etoposide for 72 h. After the incubation period MTT solution (10 mg mL-1 in PBS) was added (10 �L/well) and the plates were further incubated for 4 h at 37°C. Thereafter the formazan crystals formed were dissolved through addition of 100 �L/well 5% formic acid in 2-propanol (Merck) and the absorption of the samples was measured with an ELISA reader (Uniscan Titertec) at 580 nm. 100 �L RPMI 1640 medium (Sigma), 10 �L MTT stock and 100 �L 5% formic acid in 2-propanol served as a blank solution. The results were expressed as survival fraction (% of untreated control). All values were expressed as the mean ± SD (n=8). The data processing included the Student s̀ t-test with P ≤ 0.05 taken as significance level, using Microsoft EXCEL and OriginPlot software for PC. Apoptosis assay
The DNA extraction and horizontal gel electrophoresis procedures were performed as previously described (Vasilev et al., 2005). About 5×106 SKW-3 cells – treated with justicidin B (at 0.25 or 0.5 �M) and untreated controls, were washed in PBS and spun at 1,800 x g for 5 min. The cell pellets were resuspended in 0.25 mL PBS and lysed through addition of 0.5 mL buffer containing 0.5% Triton X-100, 20 mM Tris-HCl and 1mM EDTA (pH = 7.4). Samples were incubated at 0°C (on ice) for 5 min and thereafter spun at 11,000 x g for 20 min. The supernatants were transferred into 2 mL ‘safe lock’ test tubes and then 0.937 mL 2-propanol as well as 0.187 mL 6 M solution of NaCl was added to each sample. The tubes were gently agitated and incubated at –20°C for 12 h in order to allow precipitation of the hydrophilic DNA. The samples were centrifuged for 20 min at 11,000 g, the supernatants were decanted and DNA was washed in 1 mL ice cold 70% ethanol and then air dried. The isolated DNA was redissolved in 20 �L distilled water and analyzed by gel electrophoresis in 0.8% agarose gel. Finally DNA was stained with ethidium bromide and visualized using an UV transilluminator and photographed with a fixed digital camera (Bio Doc ITTM system).

��������(�
88
Results and discussion Callus cultures were developed as previously described (Vasilev et al., 2005). The genetically
modified cultures demonstrated typical hairy roots phenotype: intensive branching, hormone autotrophy and lack of geotropism. The most vigorous growth was observed when the bacterial growth stopped and there was no further necessity of antibiotic.
To our knowledge, no reports on the induction of hairy roots from L. leonii have been published till now. We performed DNA analysis in order to confirm the hairy roots transformation. The TL region in plasmid T-DNA of the agropine-type strain A. rhizogenes 15834 contains 18 open reading frames including several loci called rol (root loci). The products encoded by rol A and rol C genes were found to have a synergistic effect on root induction and induce increased sensitivity to auxin (Slightom et al., 1996, Day et al., 1999). PCR analysis showed that hairy roots from L. leonii contain rol C and rol A genes (Fig. 2, lanes 3 and 4) corresponding to the positive controls obtained by DNA from A. rhizogenes ATCC 15834 (lanes 7 and 6). Untransformed callus served as a negative control (lane 2). Vir D2 was not detected in the hairy roots (lane 5), thus showing that T-DNA is incorporated in the plant genome and it is not a residual bacterial contamination.
Fig. 2. PCR analysis of L. leonii roots transformed by A. rhizogenes ATCC15834. Lane 1 – DNA marker; lane 2 – untransformed L. leonii plantlets; lanes 3 and 4 – DNA from L. leonii hairy roots in which rol C of 490 bp and rol A of 248 bp integration was positive; lane 5 – DNA from hairy roots not expressing vir D2; lanes 6, 7 and 8 – positives controls of A. rhizogenes DNA showing roal A, rol C, and vir D2 (338 bp) respectively.
We undertook isolation and identification of the main component in hairy roots of L. leonii by preparative TLC and subsequent recrystallisation in cold MeOH. This isolate was analyzed by means of GC-MS and NMR. The EI-MS of the isolated compound showed a m/e value of 364 and mass fragmentation, that is consistent with the data for an arylnaphthalene lignan (Okigawa et al., 1970). Further NMR experiments were performed in order to distinguish between justicidin B and isojusticidin B as these two isomers have no different MS fragmentation pattern. A closer look at the 1H NMR spectrum showed that the proton signals at � 7.12 ppm and � 7.05 ppm appeared as singlets, which is indicative for 4,5-substitution (Okigawa et al., 1970). Therefore the resonance signals at � 7.12 ppm and � 7.05 ppm were assigned to H-6 and H-3 respectively, due to the shielding effect of the piperonyl group on H-3. Thus the isolated compound was unambiguously identified as justicidin B. The 1H NMR data is in full agreement with a previous report of justicidin B (Pettit and Schaufelberger, 1988).
Callus and hairy roots of L. leonii retained the capability to produce justicidin B. This finding supports the hypothesis that arylnaphthalene lignans are characteristic of the section Linum. Hairy roots accumulated 5 times higher amounts of justicidin B (10.8 mg g-1 DW) than the conventional cultures of callus (Table 1). The content of justicidin B in L. leonii hairy roots after 14 days period is very close to
1 2 3 4 5 6 7 8
500 bp
100 bp
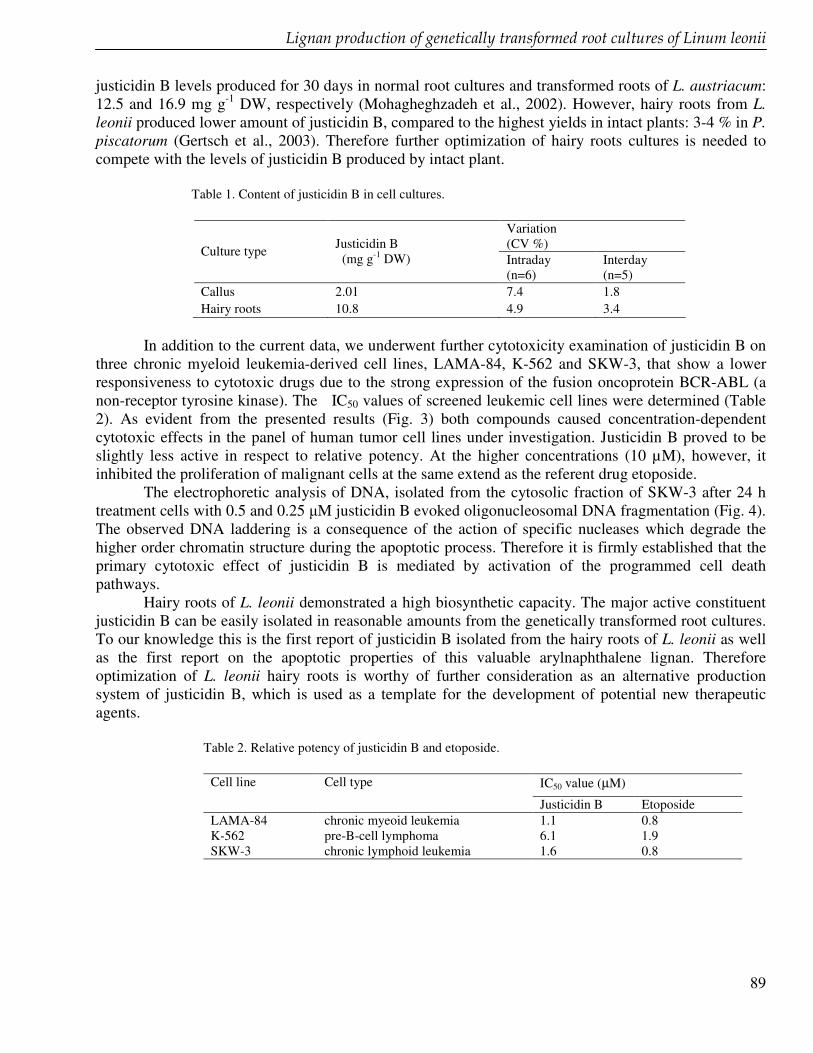
��������������������������������������������������������������������� ������������������� �����������������
89
justicidin B levels produced for 30 days in normal root cultures and transformed roots of L. austriacum: 12.5 and 16.9 mg g-1 DW, respectively (Mohagheghzadeh et al., 2002). However, hairy roots from L. leonii produced lower amount of justicidin B, compared to the highest yields in intact plants: 3-4 % in P. piscatorum (Gertsch et al., 2003). Therefore further optimization of hairy roots cultures is needed to compete with the levels of justicidin B produced by intact plant.
Table 1. Content of justicidin B in cell cultures.
Variation (CV %) Culture type Justicidin B
(mg g-1 DW) Intraday (n=6)
Interday (n=5)
Callus 2.01 7.4 1.8 Hairy roots 10.8 4.9 3.4
In addition to the current data, we underwent further cytotoxicity examination of justicidin B on
three chronic myeloid leukemia-derived cell lines, LAMA-84, K-562 and SKW-3, that show a lower responsiveness to cytotoxic drugs due to the strong expression of the fusion oncoprotein BCR-ABL (a non-receptor tyrosine kinase). The IC50 values of screened leukemic cell lines were determined (Table 2). As evident from the presented results (Fig. 3) both compounds caused concentration-dependent cytotoxic effects in the panel of human tumor cell lines under investigation. Justicidin B proved to be slightly less active in respect to relative potency. At the higher concentrations (10 µM), however, it inhibited the proliferation of malignant cells at the same extend as the referent drug etoposide.
The electrophoretic analysis of DNA, isolated from the cytosolic fraction of SKW-3 after 24 h treatment cells with 0.5 and 0.25 �M justicidin B evoked oligonucleosomal DNA fragmentation (Fig. 4). The observed DNA laddering is a consequence of the action of specific nucleases which degrade the higher order chromatin structure during the apoptotic process. Therefore it is firmly established that the primary cytotoxic effect of justicidin B is mediated by activation of the programmed cell death pathways.
Hairy roots of L. leonii demonstrated a high biosynthetic capacity. The major active constituent justicidin B can be easily isolated in reasonable amounts from the genetically transformed root cultures. To our knowledge this is the first report of justicidin B isolated from the hairy roots of L. leonii as well as the first report on the apoptotic properties of this valuable arylnaphthalene lignan. Therefore optimization of L. leonii hairy roots is worthy of further consideration as an alternative production system of justicidin B, which is used as a template for the development of potential new therapeutic agents.
Table 2. Relative potency of justicidin B and etoposide.
IC50 value (µM) Cell line
Cell type
Justicidin B Etoposide LAMA-84 chronic myeoid leukemia 1.1 0.8 K-562 pre-B-cell lymphoma 6.1 1.9 SKW-3 chronic lymphoid leukemia 1.6 0.8
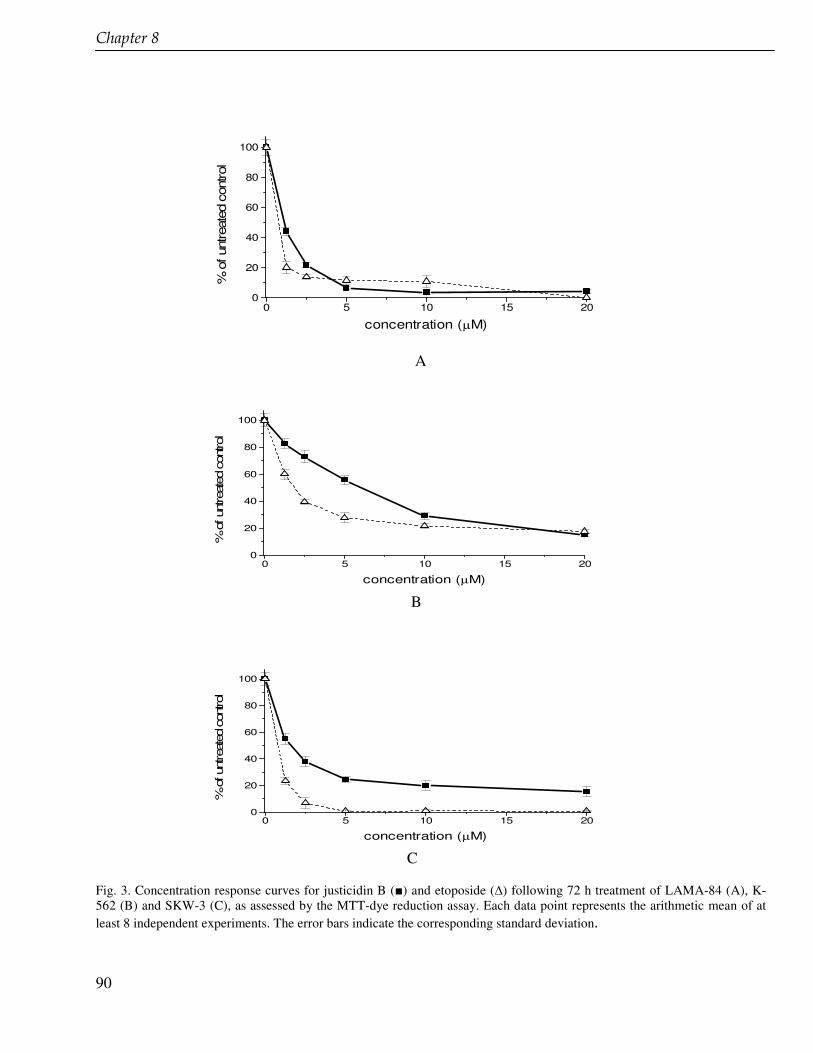
��������(�
90
A
0 5 10 15 200
20
40
60
80
100
% o
f unt
reat
ed c
ontro
l
concentration (µM) B
0 5 10 15 200
20
40
60
80
100
% o
f unt
reat
ed c
ontro
l
concentration (µM) C Fig. 3. Concentration response curves for justicidin B () and etoposide (�) following 72 h treatment of LAMA-84 (A), K-562 (B) and SKW-3 (C), as assessed by the MTT-dye reduction assay. Each data point represents the arithmetic mean of at least 8 independent experiments. The error bars indicate the corresponding standard deviation.
0 5 10 15 200
20
40
60
80
100
% o
f unt
reat
ed c
ontro
l
concentration (µM)

��������������������������������������������������������������������� ������������������� �����������������
91
1 2 3
Fig. 4. Imaging of DNA laddering induced by justicidin B treatment. DNA was extracted from the cytosolic fraction of untreated 5×106 SKW-3 cells (1) or following 24 h exposure to justicidin B at 0.25 µM (2) or 0.5 µM (3). Acknowledgements
The authors thank T. Hackl, Institut für Organische Chemie, Universität Hamburg, for recording NMR spectra. The financial support by the Huygens Program for N. Vasilev is gratefully acknowledged.

92

93
Summary and concluding remarks

94
Summary and concluding remarks In this thesis phytochemical and biosynthetic studies of lignans are described. The focus is on the
Indonesian medicinal plants Phyllanthus niruri and Piper cubeba and on two Linum species, Linum flavum and L. leonii, native to European countries.
Both Indonesian plants are used in jamu. Jamu is the Indonesian traditional herbal medicine, practised for many centuries in the Indonesian community to maintain good health and to treat diseases. The manufacturing of jamu is shifting more and more from household scale to the bigger industries. As the economical and clinical value of jamu nowadays increases in Indonesia, there is a need for further scientific proof and well conducted research. Jamu has to be developed in order to assure its efficacy and safety.
Chapter 2 reviews the research carried out on jamu and jamu plants, covering a broad range of aspects including phytochemistry, pharmacology, toxicologicy and clinical studies. In addition, ethical issues such intellectual property right (IPR), benefit sharing, and conservation are addressed.
Phyllanthus niruri is an important medicinal plant for jamu. All parts of the plant are used, among others to treat gonorrhea, syphilis, nephralgia, diarrhea, fever, and tetanus. The leaves serve to treat epilepsy, malaria, constipation, hypertension, and menstrual disorders. Lignans seem to be an important group of secondary metabolites, responsible for the biological activity of P. niruri. We initiated cell cultures of P. niruri that were able to produce lignans. The lignan profiles of cell suspensions, callus cultures, aerial plant parts, roots and seeds were compared and significant differences, both qualitatively and quantitatively, were found (chapter 3). Two compounds that were not found in P. niruri before were isolated from the cell suspension cultures: a new dihydrocubebin-dimethylether and urinatetralin, a new lignan from P. niruri, but reported earlier from P. urinaria. Feeding 0.5 mM of ferulic acid or 0.5 mM of caffeic acid, being early precursors of lignan biosynthesis, resulted in an increase up to 0.7 mg g-1 DW of the dihydrocubebin-dimethylether (control value 0.1 mg g-1 DW) and up to 0.3 mg g-1 DW of urinatetralin (control value 0.2 mg g-1 DW) in the suspension cultures. Lignans are also found in another important medicinal plant for jamu, Piper cubeba. The berries of this plant are used to treat gonorrhea, dysentery, syphilis, abdominal pain, diarrhea, enteritis and asthma. The lignan profile of berries (the particular plant part used in jamu), leaves and stalks were investigated and compared using gas chromatography (GC), gas chromatography coupled to mass spectrometry (GC-MS), and high pressure liquid chromatography (HPLC) (chapter 4). Thirteen lignans were identified in the berries, fifteen in the leaves and five in the stalks.
Our further phytochemical investigation of P. cubeba berries and leaves focused on the essential oil composition (chapter 5). The essential oil of the berries is commonly used as a constituent of cosmetics as well as for medicinal purposes. Antimicrobial, antiherpes simplex, antifungal, cardiovascular, and gastroprotective are mentioned in the latter context. Hydrodistillation of the berries of P. cubeba yielded 11.8% (w/w) and the leaves 0.9% (v/w) oil. In total 105 components could be identified (GC, GC-MS) in the berries, dealing with 63.1% of the oil. In the leaves, 63 components could be identified, corresponding with 78.0% of the oil. The total amount of monoterpenes was comparable in both oils (17.2% and 17.0%, for berries and leaves, respectively). The main monoterpenes in the berries and leaves oil were �-thujene, �-pinene, sabinene, and limonene. In the oxygenated monoterpene fractions trans-sabinene hydrate was the main component. �-Copaene, �-elemene, E-caryophyllene, and caryophyllene were the main sesquiterpenes in the berries oil. E-caryophyllene, and -cadinene were the main sesquiterpenes in the leaves oil.
Based on the similarity of the lignan and essential oil profiles we conclude that, in addition to the berries, also the leaves may be used for medicinal purposes. This knowledge can be used for the further development of (rationally designed) phytomedicines from P. cubeba.

)���������������������������* �
95
Studies on the lignan biosynthesis were performed with the European plant, Linum flavum. L. flavum cell cultures accumulate a high amount of coniferin (12% on a dry weight basis). Cell suspension cultures of leaves from this plant were treated with glucosyltransferase inhibitors in order to enhance the production of the lignan 6-methoxypodophyllotoxin (6-MPT) and to reduce the coniferin production (chapter 6). These two compounds originate from the common precursor coniferyl alcohol by different biosynthetic branches. Enzymatic transformation of this substrate by coniferyl alcohol glucosyltransferase (CAGT) yields coniferin. It was hypothized that by inhibiting this step more precursor would be available for the biosynthetic branch leading to the formation of lignans. Na2EDTA inhibited the production of coniferin up to 88% and on the other hand enhanced of the 6-MPT production up to 0.6 mg g-1 DW, 3.2-fold more than in untreated cultures. The inhibition of the coniferin production related to an inhibition of the CAGT activity, as shown in cell homogenates incubated with coniferyl alcohol. This indicates that Na2EDTA inhibits CAGT and therefore reduces the production of coniferin. The mechanism of inhibition is not clear as yet. To further study the role of CAGT in the biosynthesis of lignans, we partially purified this enzyme from cell suspension cultures of L. flavum. A complete purification of CAGT, however, appeared to be impossible because of its unstable nature. We tried to clone the CAGT from cell suspension cultures of L. flavum in Escherichia coli (chapter 7). The total RNA isolation revealed that the quality of RNA was sufficient to synthesize the cDNA. A forward and a reverse degenerated primer were designed based on the most conserved region of 100 glucosyltransferases from various species. These glucosyltransferases were aligned using MegAline software from DNASTAR Inc. The conserved region, called the PSPG box, is highly characteristic and present in all GTs involved in natural product biosynthesis. This domain may also define the active site of GTs of animals and microorganisms. The PSPG box is considered to represent the nucleotide diphosphate binding site. Several GT-encoding genes are suitable candidates to be inserted into a variety of plants with the aim of improving food, crop quality as well understanding the biosynthesis of valuable natural products for medicine.
Three different potential GTs were cloned from cell suspension cultures of L. flavum in E. coli. The gene sequence of three different products was elucidated. Two of them were complete sequences (ORF): CAGT A and CAGT B. We were able to find the conserved region (PSPG box) for the third one. Alignment of CAGT A and B with the sequences from the plant GTs obtained from databases showed 50% and 40% homology, repectively. The conserved region (PSPG box) of CAGT A and B showed 80% and 89% homology respectively. Phylogenetic analysis revealed that CAGT A and B belong to the same subfamily together with other phenylpropanoid glucosyltransferases. Although the expression of these GTs is not yet finished, it may be concluded that at least one of the two CAGT is involved in the biosynthetic step from coniferyl alcohol to coniferin in L. flavum cell suspension cultures.
In chapter 8, we describe the production of justicidin B, a cytotoxic aryltetraline lignan in cell and hairy root cultures of Linum leonii. The hairy root cultures were obatained by genetic transformation using the agropine-type strain Agrobacterium rhizogenes 15834. The products encoded by rol A and rol C genes were found to have a synergistic effect on root induction and to induce increased sensitivity to auxin. The transformation of these genes from A. rhizogenes into the hairy root was checked by PCR (polymerase chain reaction). Proof of transformation was given by the PCR products showing that rol A and rol C genes were present in the hairy roots of L. leonii. Genetically modified hairy roots produced 5-fold higher yields of justicidin B (10.8 mg g-1 DW) compared to untreated callus. This suggests that this technique may be used to enhance the accumulation of justicidin B. In addition to the production, we investigated the cytotoxic effect of justicidin B in three chronic human myeloid leukemia-derived cell lines (LAMA-84, K-562 and SKW-3), that show a lower responsiveness to cytotoxic drugs due to a strong expression of the fusion oncoprotein BCR-ABL (a non-receptor tyrosine kinase). IC50 values of justicidin B were 1.1, 6.1 and 1.5 µM for the chronic myeloid leukemia (LAMA-84), pre-B-cell

96
lymphoma (K-562) and chronic lymphoid leukemia (SKW) cell lines respectively. These IC50 were comparable to the anticancer drug etoposide (a semi-syntehetic lignan derivative). We conclude that the phytochemical studies of the two selected Indonesian medicinal plants as well as the production of lignans in cell suspension cultures provide further scientific approach for the development of jamu. Furthermore the genetic engineering studies of two Linum species contribute to medicinal plant research with respect to a better understanding of the lignan biosynthesis. The biotechnology approach may also be applied to use medicinal plants as a source for drugs discovery.

97
Samenvatting (Dutch) Aspecten betreffende fytochemie en biosynthese van lignanen, met de nadruk op Indonesische medicinale planten
Het gebruik van planten, plantenextracten en uit planten geïsoleerde verbindingen voor het behandelen van ziekten, als voedingssupplement en voor cosmetica is diep geworteld in het verleden en nog steeds in ontwikkeling. Veel geneesmiddelen die in de hedendaagse gezondheidszorg worden toegepast zijn gebaseerd op planten en ontdekt op basis van het traditionele (medicinale) gebruik door inheemse bevolkingsgroepen. Podofyllotoxine, vincristine, vinblastine, camptothecine, taxol, artemisinine, aspirine, atropine, efedrine, kinine, reserpine en digoxine zijn bekende voorbeelden van zulke geneesmiddelen. Ontwikkelingen in wetenschap en techniek geven een verdere impuls aan de exploratie van medicinale planten als bron voor nieuwe geneesmiddelen. Moderne analysetechnieken, biotechnologische benaderingen, genomics, proteomics en metabolomics worden toegepast in het onderzoek met medicinale planten en dragen bij aan de vooruitgang van dit vakgebied. Dikwijls echter vormen de relatief lage gehaltes van biologisch actieve verbindingen (zogenaamde ‘secundaire metabolieten’) en problemen bij het standaardiseren een knelpunt in de exploitatie van medicinale planten. Wereldwijd zijn pogingen ondernomen om de productie van biologisch actieve verbindingen te verhogen met behulp van biotechnologie. In dit proefschrift worden aspecten betreffende fytochemie en biosynthese van lignanen onderzocht die aanwezig zijn in twee medicinale planten uit Indonesië, Phyllanthus niruri L. (Euphorbiaceae) en Piper cubeba L. (Piperaceae), en in twee Linum-soorten die inheems zijn in Europa, Linum flavum L. en Linum leonii F.W. Schulz. (Linaceae). De beide Indonesische planten worden in jamu gebruikt. Jamu is de traditionele Indonesische geneeskunde en is gebaseerd op het volksgebruik van medicinale planten. Hoofdstuk 2 geeft een uitgebreid overzicht van jamu. De Indonesische bevolking gebruikt jamu al eeuwenlang om een goede gezondheid te behouden en om ziekten te behandelen. Wij bespreken de relatie van jamu met de biodiversiteit van Indonesië (die één der grootste ter wereld is), de positie van jamu in de huidige Indonesische samenleving, wet- en regelgeving, de bereiding van jamu (die meer en meer verschuift van kleinschalige huisvlijt naar groter industrieel), economische vooruitzichten, rationele farmacotherapie met deze middelen en ethische overwegingen die in beschouwing moeten worden genomen bij het ontwikkelen van geneesmiddelen uit traditionele bronnen. Wij geven uitgebreide informatie over de biologische activiteit, de therapeutische waarde van planten en bestanddelen uit planten die het meest frequent in jamu worden toegepast, bespreken klinisch onderzoek dat beschikbaar is en besteden aandacht aan toxicologische aspecten. Aangezien de economische en klinische betekenis van jamu in Indonesië stijgt, is een verdere wetenschappelijke onderbouwing nodig. Op basis van kennis verkregen uit goed opgezet onderzoek kan jamu in Indonesië verder worden ontwikkeld en kan de veiligheid en effectiviteit van deze middelen worden gewaarborgd. In Hoofdstuk 3 bestuderen wij lignanen in celcultures van Phyllanthus niruri, in Indonesië bekend als meniran. P. niruri is een belangrijke plant in jamu. Alle delen van de plant worden gebruikt, ondermeer om gonorroe, syfilis, nefralgie, diarree, koorts en tetanus te behandelen. De bladeren worden toegepast bij epilepsie, malaria, obstipatie, hypertensie en menstruatieproblemen. Lignanen vormen een belangrijke groep van secundaire metabolieten in deze plant en worden verantwoordelijk gehouden voor de biologische activiteit. Wij hebben celcultures opgezet van de bladeren van de plant. In zowel callusweefsel als in celsuspensies waren lignanen aanwezig. Er waren kwalitatieve en kwantitatieve verschillen tussen de lignanen in bovengrondse delen van de plant, wortels en zaden. Uit de celsuspensiecultures isoleerden wij twee lignanen die niet in de plant zelf voorkomen: een nieuwe dimethylether van cubebine en urinatetraline, dat eerder in P. urinaria werd gevonden. Koffiezuur en

98
ferulazuur, vroege voorlopers in de biosynthese van lignanen, werden aan het groeimedium van de celsuspensiecultures toegevoegd (‘precursor feeding’) met als doel de lignaanproductie in de cellen te verhogen. Normale celsuspensiecultures (controle) produceerden 0,1 mg g-1 drooggewicht cubebine dimethylether en 0,2 mg g-1 urinatetraline. In een concentratie van 0,5 mM gaven zowel koffiezuur als ferulazuur een verhoging van de productie van het cubebine dimethylether als van urinatetraline in de cellen (tot respectievelijk 0,7 mg g-1 en 0,3 mg g-1).
Lignanen zijn ook aanwezig in Piper cubeba, een andere belangrijke jamu-plant en bekend als kemukus. De bessen van P. cubeba worden gebruikt ter behandeling van gonorroe, dysenterie, syfilis, buikpijn, diarree, enteritis, en astma. Hoofdstuk 4 beschrijft de lignaanprofielen van verschillende delen van van de plant: bessen, bladeren en takjes. Als methoden werden dunnelaagchromatografie (TLC), hogedruk-vloeistofchromatografie (HPLC), gaschromatografie (GC) en gaschromatografie gekoppeld aan massaspectrometrie (GC-MS) gebruikt. In de bladeren vonden wij vijftien lignanen, in de bessen dertien en in de takjes vijf. Cubebine, hinokinine, yateine, en isoyateine zijn algemeen voorkomende lignanen in het geslacht Piper en bleken ook de belangrijkste bestandelen te zijn in de bovengrondse delen van P. cubeba die wij onderzochten.
Om een completer fytochemisch beeld van P. cubeba te krijgen bestudeerden wij de samenstelling van de vluchtige olie van bessen en bladeren van deze plant met behulp van GC en GC-MS (Hoofdstuk 5). Liganen en vluchtige olie (met mono- en sesquiterpenen en fenylpropaanderivaten) vormen de belangrijkste groepen secundaire metabolieten in P. cubeba. De vluchtige olie uit de bessen wordt gebruikt in cosmetica en voor medische doeleinden. De werking is antimicrobieel, antiviraal (bij herpes simplex), antischimmel, cardiovasculair en maagbeschermend. Stoomdestillatie van de bessen leverde 11,8% en van de bladeren 0,9% olie op. In totaal werden in de bessenolie 105 bestanddelen geïdentificeerd, overeenkomend met 63,1% van de olie. In de bladolie werden 63 bestanddelen geïdentificeerd, overeenkomend met 78,0% van de olie. De hoeveelheid monoterpenen was vergelijkbaar in beide oliën (17,2% en 17,0% voor respectievelijk bessen en bladeren). De belangrijkste monoterpenen waren �-thujeen, �-pineen, sabineen en limoneen. In de geoxygeneerde monoterpeenfractie was trans-sabineenhydraat het hoofdbestanddeel. �-Copaeen, �-elemeen, E-caryofylleen en caryofylleen waren de belangrijkste sesquiterpenen in de bessenolie; E-caryofylleen en -cadineen de belangrijkste in de bladolie. Op basis van de overeenkomstige profielen van lignanen en vluchtige olie van bessen en bladeren concluderen wij dat ook de bessen voor medicinale doeleinden bruikbaar zouden kunnen zijn. Deze kennis kan worden gebruikt voor de verdere ontwikkeling van fytotherapeutica op basis van P. cubeba. Biosynthesestudies van lignanen werden uitgevoerd met een Europese plant, Linum flavum (gele vlas). Het gebruik van podofyllotoxine als uitgangsstof voor de semi-synthetische cytostatica etoposide, teniposide en etophos, stimuleert het onderzoek naar lignanen, inclusief podofyllotoxinederivaten en tussenproducten uit de biosynthese. In Hoofdstuk 6 beschrijven wij de verhoogde productie van 6-methoxypodofyllotoxine (6-MPT) in celsuspensiecultures van blaadjes van L. flavum onder invloed van ethyleendiaminetetraacetaat (EDTA) en andere glucosyltransferaseremmers. Het enzym coniferylalcohol glycosyltransferase (CAGT) zet coniferylalcohol (een fenylpropaanderivaat en vroege voorloper in de biosynthese van lignanen) om in coniferine. Celcultures van de L. flavum accumuleren grote hoeveelheden coniferine (tot 12%, berekend op drooggewicht). De hypothese is dat door remming van de vorming van coniferine er meer coniferylalcohol beschikbaar komt, waardoor de productie van 6-MPT wordt verhoogd. Na toevoeging van EDTA werd de productie van coniferine tot 88% van de controlewaarde geremd, terwijl de 6-MPT productie met een factor 3,2 toenam, tot 0,6 mg g-1 drooggewicht. De remming van de coniferineproductie correleerde met remming van de CAGT-activiteit in celhomogenaten van L. flavum die werden geïncubeerd met coniferylalcohol. Het mechanisme achter de enzymremming is nog niet opgehelderd. Om de rol van CAGT in de lignaanbiosyntehse verder te bestuderen hebben we het enzym gedeeltelijk gezuiverd uit

)����&�������+%����,�
99
celsuspensiecultures van L. flavum. Een totale zuivering bleek, vanwege de instabiliteit van CAGT, niet mogelijk.
De biosynthese van lignanen werd ook op genetisch niveau onderzocht. Wij trachtten CAGT uit celsuspensiecultures van L. flavum te kloneren en tot expressie te brengen in de bacterie Escherichia coli (Hoofdstuk 7). De kwaliteit van het geïsoleerde RNA was voldoende om cDNA te maken. Op basis van de meest geconserveerde gebieden van het DNA dat codeert voor honderd glycosyltransferases in verschillende plantensoorten, werden primers ontworpen (‘forward en reverse degenerated primers’). Het geconserveerde gebied, ‘PSPG box’ genaamd, is zeer karakteristiek en aanwezig in alle glycosyltransferases uit biosyntheseroutes van natuurstoffen. De PSPG box is waarschijnlijk de nuceotidedifosfaat-bindingsplaats. Verschillende genen die coderen voor glycosyltransferases kunnen worden ingebouwd in planten, met als doel de kwaliteit ervan als voedingsmiddel of als landbouwgewas te verbeteren. Daarnaast kan op deze manier meer kennis worden vergaard over de biosynthese van medicinaal toegepaste natuurstoffen. Het uiteindelijke doel is het ontwikkelen van een efficiënt systeem voor de productie van cytotoxische lignanen.
Er werden drie verschillende glycosyltransferases uit celsuspensiecultures van L. flavum in E. coli gekloneerd en de sequentie werd opgehelderd. Twee producten waren complete sequenties: CAGT A en CAGT B. Van het derde product konden wij alleen het geconserveerde gebied (PSPG box) aantonen. Vergelijking (‘alignment’) van CAGT A en B met sequenties van glycosyltransferases uit planten die beschikbaar waren in databases, gaf respectievelijk 50% en 40% homologie. De geconserveerde gebieden van CATG A en B vertoonden 80% en 89% homologie. Fylogenetische analyse van de resultaten bracht aan het licht dat CAGT A en B tot dezelfde subfamilie behoren, samen met andere fenylpropanoïd glycosyltransferases. Hoewel de expressie van deze glycosyltransferases nog niet helemaal afgerond is, concluderen wij dat minstens één van de twee CAGT’s betrokken is bij de biosynthesestap van coniferylalcohol naar coniferine in L. flavum celcultures. In Hoofdstuk 8 beschrijven wij de productie van justicidine B in celcultures en in hairy-rootcultures van Linum leonii. Justicine is een arylnaftaleen-lignaan met cytotoxische, antivirale, fungicide, antiprotozoaire en bloedplaatjesaggregatieremmende eigenschappen. De hairy-rootcultures werden verkregen door genetische modificatie van L. leonii met behulp van Agrobacterium rhizogenes-stam 15834 (agropinetype). De producten waarvoor de rol A en rol C genen coderen hadden een synergistisch effect op de wortelinductie en verhoogden de gevoeligheid voor het plantenhormoon auxine. Met behulp van de PCR-techniek (polymerase chain reaction) werd bewezen dat de transformatie van de rol A en rol C genen uit A. rhizogenes in de hairy-rootcultures L. leonii geslaagd was. Genetisch gemodificeerde hairy-rootcultures produceerden vijfmaal hogere gehaltes aan justicidine B (10,8 mg g-1 drooggewicht) dan normaal callusweefsel. Daarnaast hebben wij de cyotoxische werking van justicidine B bestudeerd tegen drie humane chronische myeloïde leukemie cellijnen, LAMA-84, K-562 en SKW. Deze cellijnen zijn minder gevoelig voor cytostatica vanwege een hoge expressie van het eiwit BCR-ABL (‘fusion oncoprotein’; een non-receptor tyrosinekinase). De IC50-waarden (continue incubatie in de MTT-assay) van justicidine bedroegen 1,1 �M (LAMA-84), 6,1 �M (K-562) en 1,5 �M (SKW) en waren vergelijkbaar met die van het referentiecytostaticum etoposide. Wij concluderen dat het fytochemische onderzoek van de twee Indonesische planten en het onderzoek naar de productie van liganen in celsuspensiecultures de verdere ontwikkeling van jamu wetenschappelijke ondersteunen. Het moleculair-biologisch en genetisch onderzoek dat met twee Europese Linum-soorten is uitgevoerd draagt bij aan een beter begrip van de lignaanbiosynthese. De biotechnologische benadering kan ook worden toegepast om medicinale planten te gebruiken als bron voor geneesmiddelontwikkeling.

100

101
Ringkasan (Indonesian) Studi fitokimia dan biosintesis lignan, dengan fokus pada tumbuhan obat Indonesia
Penggunaan tumbuh-tumbuhan, ekstrak dan senyawa kimia dari tumbuhan serta turunannya, dalam pengobatan berbagai penyakit, makanan tabahan dan bahan baku kosmetik telah berlangsung sejak lama dan terus berkembang sampai sekarang. Banyak obat modern diturunkan dari tumbuhan yang pada awalnya ditemukan melalui penggunaannya secara tradisional. Beberapa contoh diantaranya: obat anti kanker (podophyllotoxin, vincristine, vinblastin, taxol), anti malaria (quinine dan artemisinin), obat penguat jantung (digoxin), dan obat demam (aspirin). Perkembangan ilmu pengetahuan dan teknologi menstimulasi pengembangan tumbuhan obat sebagai sumber yang berguna untuk penemuan obat. Metoda analitik modern, pendeketan bioteknologi, metabolomik, proteomik dan genomik sekarang sudah diaplikasikan dalam penelitian tumbuhan obat dan memberikan kontribusi yang besar dalam pengembangan tumbuhan obat. Pendekatan ini sudah digunakan diseluruh dunia untuk mengatasi permasaalahan yang sering muncul dalam pengembangan tumbuhan obat. Permasaalah tersebut diantaranya rendahnya kandungan senyawa berkhasiat dari tumbuhan obat dan standardisasinya.
Pada tesis ini dimuat studi tentang aspek fitokimia dan bioteknologi terhadap senyawa aktif lignan dari dua tumbuhan obat yang digunakan di Indonesia yaitu Phyllanthus niruri L (Euphorbiaceae) dan Piper cubeba L(Piperaceae) dan dua tumbuhan obat dari Eropa yaitu Linum flavum L dan Linum leoni F.W. Schulz (Linaceae). Kedua tumbuhan Indonesia tersebut digunakan sebagai komponen jamu. Jamu adalah obat tradisional di Indonesia yang dibuat dari tumbuhan dan digunakan oleh masyarakat Indonesia semenjak beberapa abad yang lalu untuk memelihara kesehatan dan mengobati penyakit. Produksi jamu telah berkembang dari skala industri rumah tangga ke skala industri yang lebih besar. Karena nilai klinis dan ekonomis dari jamu sekarang terus meningkat di Indonesia, maka dibutuhkan bukti-bukti ilmiah dan penelitian lanjut terhadap tumbuhan obat. Jamu harus dikembangkan untuk menjamin aktifitas farmakologi dan keamanannya. Tinjauan pustaka tentang penelitian-penelitian yang dilakukan terhadap tumbuhan yang digunakan untuk memproduksi jamu, secara komprehensif dimuat pada Bab 2. Tinjauan ini mencakup berbagai aspek termasuk fitokimia, farmakologi, toksikologi dan uji klinis serta berbagai isu etika seperti hak kekayaan intelektual, pembagian keuntungan dan konservasi.
Phyllanthus niruri (meniran) adalah salah satu tumbuhan obat yang sering digunakan untuk jamu. Seluruh bagian dari tumbuhan ini digunakan untuk mengobati gonore, sifilis, nefralgia, diare, demam dan tetanus. Daun meniran digunakan untuk mengobati epilepsi, malaria, konstipasi, hipertensi dan kelainan menstruasi. Senyawa lignan adalah metabolit sekunder yang penting pada tumbuhan ini, yang betanggung jawab terhadap aktivitas biologinya. Kultur sel dari P. niruri menghasilkan senyawa lignan. Terdapat perbedaan yang signifikan secara kuanlitatif dan kuantitatif antara profil senyawa lignan dari kultur sel, kultur kalus, tumbuhan asli, akar dan biji dari P. niruri. Dua senyawa lignan yang tidak ditemukan sebelumnya dari tumbuhan ini berhasil diisolasi dan dimurnikan. Satu diantaranya adalah senyawa baru yaitu cubebin dimetilleter dan senyawa baru untuk P. niruri tetapi telah dilaporkan sebelumnya dari tumbuhan lain, P. urinaria, yaitu urinatetralin (lihat Bab 3). Penambahan dua senyawa-antara untuk biosintesis lignan pada kultur sel P. niruri dapat menstimulasi peningkatan produksi cubebin dimetileter hingga 0,7 mg g-1 berat kering (kontrol sel hanya 0,1 mg g-1 berat kering) dan urinatetralin hingga 0,3 mg g-1 berat kering (control sel hanya 0,2 mg g-1 berat kering). Dua senyawa-antara tersebut adalah asam ferulat (0,5 mM) dan asam kafeat (0,5 mM).
Lignan juga ditemukan dari tumbuhan obat lainnya yang digunakan untuk jamu, yaitu Piper cubeba. Biji dari tumbuhan ini digunakan untuk mengobati gonore, disentri, sifilis, sakit pada perut, diare, enteritis dan asma. Profil senyawa lignan dari biji, daun dan batang P. cubeba dibuat dan dibandingkan dengan menggunakan khromatografi gas (GC), khromatografi gas-spektrometer masa (GC-MS) dan khromatografi cair tegangan tinggi (HPLC) (Bab 4). Tiga belas lignan ditemukan pada

102
biji, lima belas pada daun dan lima pada batang. Pada Bab 5 dimuat hasil penelitian terhadap komposisi minyak atsiri dari P. cubeba. Minyak atsiri biasanya digunakan sebagai bahan pembuat kosmetik serta untuk tujuan pengobatan. Beberapa aktifitas farmakologi dari minyak atsiri adalah sebagai antimikroba, anti herpes simplex, antijamur, obat jantung, proteksi lambung. Destilasi air dari biji P. cubeba menghasilkan minyak atsiri sebanyak 11,8% dan dari daun 0,9%. Dari biji dapat diidentifikasi 105 komponen, sebesar 63,1% dari minyak atsiri, sedangkan dari daun 63 komponen dengan jumlah 78,0% dari minyak atsiri. Jumlah total monoterpen dari minyak yang diperoleh dari biji dan daun hampir sama, yaitu 17,2% dan 17,%. Monoterpen utama dari biji dan daun adalah �-thujene, �-pinene, sabinene, and limonene. Trans-sabinenhidrat adalah komponen utama dari fraksi monoterpen teroksigenasi. �-Copaene, �-elemene, E-caryophyllene, and caryophyllene adalah sesquiterpen utama dari minyak atisiri biji, sedangkan E-caryophyllene, and -cadinene adalah sesquiterpen utama dari daun. Berdasarkan hasil penelitian terhadap lignan dan minyak atsiri dari P. cubeba dapat disimpulkan bahwa selain dari biji, daun dari tumbuhan ini juga dapat dimanfaatkan untuk tujuan pengobatan. Kesimpulan ini dapat dijadikan dasar untuk pengembangan lebih lanjut sehingga P. cubeba bisa dijadikan untuk pengobatan rasional (fitofarmaka).
Studi terhadap biosintesis lignan dilakukan dengan menggunakan tumbuhan Eropa, Linum flavum L. Penggunaan podofilotoksin sebagai bahan dasar untuk produksi obat antikanker etoposide, teniposide dan etopophos menstimulasi penelitian terhadap lignan, termasuk turunan podofilotoksin lainnya dan produk antara dari biosintesis lignan. Pada Bab 6 ditulis studi tentang peningkatan produksi 6-metoksipodofilotoksin (6-MPT) dan penurunan produksi coniferin pada kultur suspensi sel dari L. flavum dengan menggunakan natrium etilendiamina tetraasetat (Na2EDTA) dan senyawa penghambat enzim glucosylrasnferase lainnya. Enzim coniferil alkohol glucosyltransferase (CAGT) menjadi katalis pembentukan senyawa coniferin dengan menggunakan coniferil alkohol sebagai substrat. Produksi coniferin pada kultur suspensi sel L. flavum sangat besar yaitu 12% dari berat kering. Coniferil alkohol juga digunakan untuk pembentukan senyawa 6-MPT melalui jalur biosintesis yang berbeda dengan coniferin. Aktivitas CAGT yang dihambat akan mengurangi produksi coniferin, sehingga jumlah coniferil alkohol meningkat. Sebaliknya, akumulasi coniferil alkohol ini bisa digunakan sebagai bahan antara untuk produksi senyawa 6-MPT. Na2EDTA menghambat produksi coniferin sampai 88% dan di sisi lain meningkatkan produksi 6-MPT sampai 0,56 mg g-1 berat kering, lebih dari tiga kali lipat dibanding kultur control. Pengurangan produksi coniferin berkorelasi dengan berkurangnya aktivitas CAGT, seperti terlihat pada aktivitas ekstrak sel terhadap coniferil alkohol. Ini menunjukkan bahwa Na2EDTA menghambat CAGT dan oleh karena itu mengurangi produksi coniferin. Mekanisme penghambatan terhadap CAGT masih belum jelas. Untuk studi lebih lanjut terhadap peran CAGT dalam biosintesis lignan telah dilakukan pemurnian parsial enzim ini dari kultur suspensi sel L. flavum. Pemurnian sampai pada tingkat 100% sangat sulit karena ketidakstabilan enzim.
Penelitian lanjutan untuk memahami fungsi enzim CAGT adalah cloning CAGT dari kultur suspensi sel L. flavum pada Escherichia coli (Bab 7). Hasil isolasi total RNA menunjukkan kualitas yang cukup untuk sintesis cDNA. Satu forward dan satu reverse degenerated primer didisain berdasarkan daerah lestari seratus glucosyltransferase dari berbagai spesies tumbuhan. Glcosyltransferas ini di analisis menggunkan perangkat lunak MegAline dari DNASTAR Inc. Daerah lestari, yang dikenal dengan kotak PSPG, sangat khas dan ada di semua glucosyltransferase yang berperan dalam biosintesis bahan alam. Daerah ini juga didefenisikan sebagai sisi aktif enzim glucosyltransferase dari hewan dan mikroorganisme. Kotak PSPG dipercayai sebagai representasi dari sisi yang berikatan dengan nukleotida difosfat. Beberapa gen pengkode glucosyltransferase adalah kandidat yang cocok untuk disisipkan kedalam tanaman untuk tujuan peningkatan kualitas makanan, hasil pertanian dan untuk memahami biosintesis bahan alam yang berguna sebagai obat.
Tiga glucosyltransferase telah berhasil di kloning dari kultur suspensi sel L. flavum pada E. coli. Dua diantaranya telah memiliki urutan nukleotida lengkap (ORF) yaitu CAGT A dan CAGT B.

-���*� ���+������ ���,�
103
Sedangkan urutan lengkap dari CAGT C belum berhasil diidentifikasi. Penjajaran CAGT A dan CAGT B dengan beberapa glucosyltransferase dari berbagai tumbuhan berdasarkan basis data, menunjukkan 50% dan 40% homolog berturut-turut. Sedangkan kotak PSPG box dari CAGT A dan B menunjukkan 80% dan 89% homolog, berturut-turut. Analisis filogenetik menunjukkan bahwa CAGT A dan B tergolong ke dalam subfamili yang sama dengan fenilpropanoid glucosyltransferase yang lain. Walaupun ekspresi dari kedua gen ini belum berhasil dilakukan, dapat disimpulkan bahwa sekurangnya satu dari dua CAGT berperan dalam tahap biosintesis dari coniferil alkohol untuk pembentukan coniferin pada kultur suspensi sel L. flavum.
Pada Bab 8 dijelaskan produksi justicidin B, sebuah senyawa sitotoksik ariltetralin lignan pada kultur sel dan akar rambut dari Linum leonii. Kultur akar rambut dihasilkan dari transformasi genetik menggunakan Agrobacterium rhizogenes 15834 strain (tipe agropin). Produk yang dikode oleh gen rol A dan rol C mempunyai efek sinergis pada induksi akar dan meningkatkan sensitifitas auxin. Keberhasilan transformasi gen ini dari A. rhizogenes pada kultur akar rambut diperiksa dengan PCR. Pada produk PCR ditunjukkan bahwa gen rol A dan rol C ditemukan pada kultur akar rambut L. Leoni. Akar rambut yang dimodifikasi secara genetik menghasilkan produksi justicidin B (10 mg g-1 berat kering) lima kali lipat lebih tinggi bila dibandingkan dengan kalus kontrol. Hasil ini menunjukkan bahwa teknik ini dapat digunakan untuk meningkatkan akumulasi justicidin B. Di samping itu, penelitian terhadap efek sitotoksik justicidin B terhadap tiga sel LAMA-84, K-562 dan SKW yang diturunkan dari leukimia kronis dari manusia, yang menunjukkan respon yang lebih rendah terhadap obat sitotoksik karena ekspresi yang tinggi dari protein BCR-ABL (tirosin kinase non reseptor). IC50 dari justicidin B adalah 1,2, 6,1 dan 1,5 µM untuk sel LAMA-84, K-562 dan leukimia limfoid kronis (SKW), berturut-turut. Nilai IC50 sebanding dengan obat antikanker etoposide.
Dapat disimpulkan bahwa studi fitokimia dari dua tumbuhan obat Indonesia, juga produksi lignan pada kultur suspensi sel pada tesis ini memberikan pendekatan ilmiah lanjut untuk pengembangan jamu. Sedangkan studi rekayasa genetik pada dua spesies Linum memberikan kontribusi untuk penelitian tumbuhan obat, khususnya dalam memahami biosintesis lignan sebagai salah satu golongan metabolit sekunder penting untuk aktifitas farmakologi. Pendekatan bioteknologi dapat pula diaplikasikan dalam upaya menggunakan tumbuhan obat sebagai sumber penemuan obat baru.

104

105
References
Adams, R.P., Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, Allured Publishing Corporation, Carol Stream, Illinois, 2001
Aggarwal, B.B., Kumar, A., Bharti, A.C., 2003. Anticancer potential of curcumin: preclinical and clinical studies. Anticancer Research, 23, 363-398
Ahmed, S., Anuntiyo, J., Malemud, C.J., Haqqi, T.M., 2005. Biological basis for the use of botanicals in osteoarthritis and rheumatoid arthritis: A review. Evidence-Based Complementary and Alternative Medicine, 2, 301-308
Akhtar, M.S., Khan, M.A., Malik, M.T., 2002. Hypoglycaemic activity of Alpinia galanga rhizoma and its extracts in rabbits. Fitoterapia, 73, 623-628
Alarcon-Aguilara, F.J., Roman-Ramos, R., Perez-Gutierrez, S., Aguilar-Contreras, A., Contreras-Weber, C.C., Flores-Saenz, J.L., 1998. Study of the anti-hyperglycemic effect of plants used as antidiabetics. Journal of Ethnopharmacology, 61, 101-110
Alen, Y., Nakajima, S., Nitoda, T., Baba, N., Kanzaki, H., Kawazu, K., 2000. Antinematodal activity of some tropical rainforest plants against the pinewood nematode, Bursaphelenchus xylophilus. Zeitschrift für Naturforschung C (A Journal of Biosciences), 55, 295-299
Alexander-Lindo, R.L., Morrison, E.Y.S., Nair, M.G., 2004. Hypoglycaemic effect of stigmast-4-en-3-one and its corresponding alcohol from the bark of Anacardium occidentale (Cashew). Phytotherapy Research, 18, 403-407
Ali, N., Hashim, N.H., Saad, B., Safan, K., Nakajima, M., Yoshizawa, T., 2005. Evaluation of a method to determine the natural occurrence of aflatoxins in commercial traditional herbal medicines from Malaysia and Indonesia. Food and Chemical Toxicology, 43, 1763-1772
Altman, R.D., Marcussen, K.C., 2001. Effects of a ginger extract on knee pain in patients with osteoarthritis. Arthritis and Rheumatism, 44, 2531-2538
Anjaneyulu, A.S.R., Ramaiah, P.A., Ramachandra, L., Venkateswarlu, R., 1981. New Lignans from the heartwood of Cleistanthus collinus. Tetrahedron, 37, 3641-3652
Anjaneyulu, A.S.R., Rao, K.J., Row, L.R., Subrahmanyam, 1973. Crystalline constituents of Euphorbiaceae, 12. Isolation and structural elucidation of 3 new lignans from leaves of Phyllanthus niruri Linn. Tetrahedron, 29, 1291-1298
Anonymous, Nederlandse Farmacopee, 6th ed., 2nd printing, 1966 Staatsuitgeverij, ‘s-Gravenhage, p. 72
Antony, S., Kuttan, R., Kuttan, G., 1999. Immunomodulatory activity of curcumin. Immunological Investigations, 28, 291-303
Arambewela, L.S.R., Arawwawala, L.D.A.M., Ratnasooriya, W.D., 2005. Antidiabetic activities of aqueous and ethanolic extracts of Piper betle leaves in rats. Journal of Ethnopharmacology, 102, 239-245
Aruna, K., Sivaramakrishnan, V.M., 1998. Anticarcinogenic effects of some Indian plant-products. Food and Chemical Toxicology, 953-956

106
Baba, A., Kawamura, N., Makino, H., Ohta, Y., Taketomi, S., Sohda, T., 1996. Studies on disease-modifying antirheumatic drugs: synthesis of novel quinoline and quinazoline derivatives and their anti-inflammatory effect. Journal of Medicinal Chemistry, 39, 5176-5182
Bachmann, T.L., Ghlia, F., Thorssell, K.G.B., 1993. Lignans and lactones from Phyllanthus anisolobus. Phytochemistry, 32, 189-191
Backer, C.A., Van den Brink, R.C.B., 1968. Flora of Java, NVP Noordhoff, Groningen, p.168-170
Backer, C.A., Van den Brink, R.C.B., 1968. Flora of Java. NVP Noordhoff, Groningen, p. 466-469
Badheka, L.P., Prabhu, B.R., Mulchandani, N.B., 1986. Dibenzylbutyrolactone lignans from Piper cubeba. Phytochemistry, 25, 487-489
Badheka, L.P., Prabhu, B.R., Mulchandani, N.B., 1987. Lignans of Piper cubeba. Phytochemistry, 26, 2033-2036
Bastos, J.K., Carvalho, J.C.T., de Souza, G.H.B., Pedrazzi, A.H.P., Sarti, S.J., 2001. Anti-inflammatory activity of cubebin, a lignan from the leaves of Zanthoxyllum naranjillo Griseb. Journal of Ethnopharmacology, 75, 279-282
Beers, S.J., 2001. Jamu, the ancient Indonesian art of herbal healing. Periplus Editions (HK) Ltd.
Bendjeddou, D., Lalaoui, K., Satta, D., 2003. Immuno stimulating activity of the hot water-soluble polysaccharide extracts of Anacyclus pyrethrum, Alpinia galanga and Citrullus colocynthis. Journal of Ethnopharmacology, 88, 155-160
Berim, A., Spring, O., Conrad, J., Maitrejean, M., Boland, W., Petersen, M., 2005. Enhancement of lignan biosynthesis in suspension cultures of Linum nodiflorum by coronalon, indanoyl-isoleucine and methyl jasmonate. Planta, 222, 769-776
Bermawie, N., Hernani, Suwijo, P., Kardono, L.B.S., 2005. Approaches for sustainable utilization of biodiversity of medicinal and aromatic plants in Indonesia. http://www.ics.trieste.it//Documents/Downloads/df3204.pdf
Bhat, G.P., Surolia, N., 2001. In vitro antimalarial activity of extracts of three plants used in the traditional medicine of India. American Journal of Tropical Medicine and Hygiene, 65, 304-308
Borsato, M.L.C., Grael, C.F.F., Souza, G.E.P., Lopes, N.P., 2000. Analgesic activity of the lignans from Lychnophora ericoides. Phytochemistry, 55, 809-813
Bouttier, S., Fourniat, J., Garofalo, C., Gleye, C., Laurens, A., Hocquemiller, R., 2002. β-lactamase inhibitors from Anacardium occidentale. Pharmaceutical Biology, 40, 231-234
Bowles, D., Isayenkova, J., Lim, E.K., Poppenberger, B., 2005. Glycosyltransferases: managers of small molecules. Current Opinion in Plant Biology, 2005, 8, 254-263
Bradford, M.M., 1976. A rapid and sensitive methode for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72, 248-254
Bunpo, P., Kataoka, K., Arimochi, H., Nakayama, H., Kuwahara, T., Bando, Y., Izumi, K., Vinitketkumnuen, U., Ohnishi, Y., 2004. Inhibitory effects of Centella asiatica on azoxymethane-induced aberrant crypt focus formation and carcinogenesis in the intestines of F344 rats. Food and Chemical Toxicology, 42, 1987-1997
Caksen, H., Odabas, D., Akbayram, S., Cesur, Y., Arslan, S., Uner, A., Oner, A.F., 2003. Deadly nightshade (Atropa belladonna) intoxication: an analysis of 49 children. Human & Experimental Toxicology, 22, 665-668

-�������� �
107
Calabrese, C., Berman, S.H., Babish, J.G., Ma, X.F., Shinto, L., Dorr, M., Wells, K., Wenner, C. A., Standish, L.J., 2000. A phase I trial of andrographolide in HIV positive patients and normal volunteers. Phytotherapy Research, 14, 333-338
Calixto, J.B., Santos, A.R.S., Filho, V.C., Yunes, R.A., 1998. A review of the plants of the genus Phyllanthus: Their chemistry, pharmacology and their therapeutic potential. Medical Research Review, 18, 225-258
Campos, A.H., Schor, N., 1999. Phyllanthus niruri inhibits calcium oxalate endocytosis by renal tubular cells: its role in urolithiasis. Nephron, 81, 393-397
Cesarone, M.R., Incandela, L., De Sanctis, M.T., Belcaro, G., Bavera, P., Bucci, M., Ippolito, E., 2001. Evaluation of treatment of diabetic microangiopathy with total triterpenic fraction of Centella asiatica: A clinical prospective randomized trial with a microcirculatory model. Angiology, 62, S49-S54
Chander, R., Srivastava,V., Tandon, J.S., Kapoor, N.K., 1995. Antihepatotoxic activity of diterpenes of Andrographis paniculata (Kal-Megh) against Plasmodium berghei-induced hepatic damage in Mastomys natalensis. International Journal of Pharmacognosy, 33, 135-138
Chang, C.C., Lien, Y.C., Liu, K.C.S., Lee, S.S., 2003. Lignans from Phyllanthus urinaria. Phytochemistry, 63, 825-833
Channa, S., Dar, A., Ahmed, S., Atta’ur Rahman, 2005. Evaluation of Alstonia scholaris leaves for broncho-vasodilatory activity. Journal of Ethnopharmacology, 97, 469-476
Chattopaday, S., Srivastava, A.K., Bhojwani, S.S., Bisaria, V.S., 2002. Production of podophyllotoxin by plant cell cultures of Podophyllum hexandrum in bioreactor. Journal of Bioscience & Bioengineering, 93, 215-220
Chattopadhyay, D., Arunachalam, G., Ghosh, L., Mandal, A.B., 2004. CNS activity of Alstonia macrophylla leaves extracts: an ethnomedicine of Onge of Bay Islands. Fitoterapia, 75, 673-682
Chattopadhyay, I., Biswas, K., Bandyopadhyay, U., Banerje, R.K., 2004. Turmeric and curcumin: biological actions and medicinal applications. Current Science, 87, 44-53
Cheenpracha, S., Karalai, C., Ponglimanont, C., Subhadhirasakul, S., Tewtrakul, S., 2006. Anti-HIV-1 protease activity of compounds from Boesenbergia pandurata. Bioorganic & Medicinal Chemistry, 14, 1710-1714
Chen, C.C., Hsin, W.C., Ko, F.N., Huang, Y.L., Ou, J.C., Teng, C.M., 1996. Antiplatelet arylnaphthalide lignans from Justicia procumbens. Jornal of Natural Products, 59,1149-1150
Chen, J.Y., Liu, P.Y., Chen, J.H., Lin, L.J., 2004. Safety of transvenous temporary cardiac pacing in patients with accidential digoxin overdose and symptomatic bradycardia. Cardiology, 102, 152-155
Cheng, C.L., Guo, J.S., Luk, J., Koo, M.W.L., 2004. The healing effects of Centella extract and asiaticoside on acetic acid induced gastric ulcers in rats. Life sciences, 74, 2237-2249
Chi, C.W., Chang, Y.F., Chao, T.W., Chiang, S.H., Peng, F.K., Lui, W.Y., Liu, T.Y., 1994. Flow-cytometric analysis of the effect of berberine on the expression of glucocorticoid receptors in human hepatoma hepg2 cells. Life Sciences, 26, 2099-2107
Chithra, V., Leelamma, S., 2000. Coriandrum sativum - effect on lipid metabolism in 1,2-dimethyl hydrazine induced colon cancer. Journal of Ethnopharmacology, 71, 457-463

108
Choi, E.M., Hwang, J.K., 2003. Investigations of anti-inflammatory and antinociceptive activities of Piper cubeba, Physalis angulata and Rosa hybrida. Journal of Ethnopharmacology, 89, 171-175
Choi, E.M., Hwang, J.K., 2005. Effect of some medicinal plants on plasma antioxidant system and lipid levels in rats. Phytotherapy Research, 19, 382-386
Choi, E.M., Hwang, J.K., 2005. Screening of Indonesian medicinal plants for inhibitor activity on nitric oxide production of RAW264.7 cells and antioxidant activity. Fitoterapia, 76, 194-203
Chong, J., Baltz, R., Fritig, B., Saindrenan, P., 1999. An early salicylic acid-, pathogen- and elicitor-inducible tobacco glucosyltransferase: role in compartmentalization of phenolics and H2O2 metabolism. FEBS Letter, 458, 204-208
Chong, J., Baltz, R., Schmitt, C., Beffa, R., Fritig, B., Saindrenan, P., 2002. Downregulation of a pathogen-responsive tobacco UDP-Glc: phenypropanoid glucosyltransferase reduces scopoletin glucoside accumulation, enhance oxidative stress, and weakens virus resistance. Plant Cell, 14, 1093-1107
Chrubasik, S., Pittler, M.H., Roufogalis, B.D., 2005. Zingiberis rhizoma: A comprehensive review on the ginger effect and efficacy profiles. Phytomedicine, 12, 684-701
Chularojmontri, L., Wattanapitayakul, S.K., Herunsalee, A., Charuchongkolwongse, S., Niumsakul, S., Srichairat, S., 2005. Antioxidative and cardioprotective effects of Phyllanthus urinaria L. on doxorubicin-induced cardiotoxicity. Biological & Pharmaceutical Bulletin, 28, 1165-1171
Coon, J.T., Ernst, E., 2002. Panax ginseng: A systematic review of adverse effect and drug interactions. Drug Safety, 25, 323-344
Coutinho, P.M., Deleury, E., Davies, G.J., Henrissat, B., 2003. An evolving hierarchical family classification for glycosyltransferases. Journal of Molecular Biology, 328, 307-317
Cragg, G.M., Newman, D.J., 2005. Plants as a source of anti-cancer agents. Journal of Ethnopharmacology, 100, 72-79
Da Silva, R., De Souza, G.H.B., Da Silva, A.A., De Souza, V.A., Pereira, A.C., Royo, V.D., Silva, M.L.A.E., Donate, P.M., Araujo, A.L.S.D., Carvalho, J.C.T., Bastos, J.K., 2005. Synthesis and biological activity evaluation of lignan lactones derived from (-)-cubebin. Bioorganic & Medicinal Chemistry Letters, 15, 1033-1037
Dawkins, G., Hewitt, H., Wint, Y., Obiefuna, P.C.M., Wint, B., 2003. Antibacterial effects of Carica papaya fruit on common wound organisms. West India Medical Journal, 52, 290-292
Day, S.H., Chiu, N.Y., Won, S.J., Lin, C.N., 1999. Cytotoxic lignans of Justicia ciliate. Journal of Natural Products, 62, 1056-1058
De Clercq, E., 2000. Current lead natural products for the chemotherapy of human immunodefiency virus (HIV) infection. Medicinal Research Review, 20, 323-349
De Mejia, E.G., Ramirez-Mares, M.V., Arce-Popoca, E., Wallig, M., Villa-Trevino, S. 2004. Inhibition of liver carcinogenesis in Wistar rats by consumption of an aqueous extract from leaves of Ardisia compressa. Food and Chemical Toxicology, 42, 509-516
De Sanctis, M.T., Belcaro, G., Incandela, L., Cesarone, M.R., Griffin, M., Ippolito, E., Cacchio, M., 2001. Treatment of edema and increased capillary filtration in venous hypertension with total triterpenic fraction of Centella asiatica: A clinical, prospective, placebo-controlled, randomized, dose-ranging trial. Angiology, 52, S55-S59

-�������� �
109
De Souza, V.A., Da Silva, R., Pereira, A.C., Royo, V.D., Saraiva, J., Montanheiro, M., De Souza, G.H.B., Da Silva, A.A., Grando, M.D., Donate, P.M., Bastos, J.K., Albuquerque, S., Silva, M.L.A.E., 2005. Trypanocidal activity of (-)-cubebin derivatives against free amastigote forms of Trypanosoma cruzi. Bioorganic & Medicinal Chemistry Letters. 15, 303-307
Dhandapani, S., Subramanian, R., Rajagopal, S., Namasivayam, N., 2002. Hypolipidemic effect of Cuminum cyminum L. on alloxan-induced diabetic rats. Pharmacological Research, 46, 251-255
Dharmawardhana, D.P., Ellis, B.E., Carlson, J.E., 1995. A β-glucosidase from lodgepole pine xylem specific for the lignin precursor coniferin. Plant Physiology, 107, 331-339
Dobhal, M.P., Li, G.L., Gryshuk, A., Graham, A., Bhatanager, A.K., Khaja, S.D., Joshi, Y.C., Sharma, M.C., Oseroff, A., Pandey, R.K., 2004. Structural modifications of plumieride isolated from Plumeria bicolor and the effect of these modifications on in vitro anticancer activity. Journal of Organic Chemistry, 69, 6165-6172
Dua, V.K., Qjha, V.P., Roy, R., Joshi, B.C., Valecha, N., Devi, C.U., Bhatnagar, M.C., Sharma, V.P., Subbatao, S.K., 2004. Anti-malarial activity of some xanthones isolated from the roots of Andrographis paniculata. Journal of Ethnopharmacology, 95, 247-251
Eisei Indonesia PT., Medicinal Herb Index in Indonesia, 1995. 2nd ed.; PT Eisei Indonesia, Jakarta
Empt, U., Alferman, A.W., Pras, N., Petersen, M., 2000. The use of plant cell cultures for the production of podophyllotoxin and related lignans. Journal of Applied Botany, 74, 145-150
Eno, A.E., Owo, O.I., Itam, E.H., Konya, R.S., 2000. Blood pressure depression by the fruit juice of Carica papaya (L.) in renal and DOCA-induced hypertension in the rat. Phytotherapy Research, 14, 235-239
Ernst, E., Pittler, M.H., 2002. Risks associated with herbal medicinal products. Wiener Medizinische Wochenschrift 152, 183-189
Ficker, C.E., Smith, M.L., Susiarti, S., Leaman, D.J., Irawati, C., Arnason, J.T., 2003. Inhibition of human pathogenic fungi by members of Zingiberaceae used by the Kenyah (Indonesian Borneo). Journal of Ethnopharmacology, 85, 289-293
Flavor & Fragrance Library Shimadzu Benelux, ‘s-Hertogenbosch.The Netherlands.
Freitas, A.M., Schor, N., Boim, M.A., 2002. The effect of Phyllanthus niruri on urinary inhibitors of calcium oxalate crystallization and other factors associated with renal stone formation. B. J. U International, 89, 829-834
Gamborg, O.L., Miller, R.A., Oijama, V., 1968. Nutrient requirements of suspension cultures of soybean root cells. Experimental Cell Research, 50, 151-158
Gao, Y.H., Gao, H., Chan, E., Tang, W.B., Xu, A.L., Yang, H.Y., Huang, M., Lan, J., Li, X.T., Duan, W., Xu, C.J., Zhou, S.F., 2005. Antitumor activity and underlying mechanisms of ganopoly, the refined polysaccharides extracted from Ganoderma lucidum, in mice. Immunology Investigations, 34, 171-198
Gertsch, J., Tobler, R.T., Brun, R., Sticher, O., Heilmann, J., 2003. Antifungal, antiprotozoal, cytotoxic and piscicidal properties of Justicidin B and a new arylnaphthalide lignan from Phyllanthus piscatorum. Planta Medica, 69, 420-424
Giri, A., Narasu, M.L., 2000. Production of podophyllotoxin from Podophyllum hexandrum : a potential natural product for clinically useful anticancer drug. Cytotechnology, 34, 17-26

110
Gnanapragasam, A., Ebenezar, K.K., Sathish, V., Govindaraju, P., Devaki, T., 2004. Protective effect of Centella asiatica on antioxidant tissue defense system against adriamycin induced cardiomyopathy in rats. Life Sciences, 76, 585-597
Gordaliza, M., Garcia, P.A., Miguel del Corral, J.M., Castro, M.A., Gomez-Zurita, M.A., 2004. Podophyllotoxin: distribution, sources, applications and new cytotoxic derivatives. Toxicon, 44, 441-459
Goto, H., Sasaki, Y., Fushimi, H., Shibahara, N., Shimada, Y., Komatsu, K., 2005. Effect of Curcuma herbs on vasomotion and hemorheology in spontaneously hypertensive rat. American Journal of Chinese Medicine, 33, 449-457
Goun, E., Cunningham, G., Chu, D., Nguyen, C., Miles, D., 2003. Antibacterial and antifungal activity of Indonesian ethnomedical plants. Fitoterapia, 74, 592-596
Grosvenor, P.W., Gothard, P.K., McWilliam, N.C., Supriono, A., Gray, D.O., 1995. Medicinal-plants from Riau province, Sumatra, Indonesia .1. Uses. Journal of Ethnopharmacology, 45, 75-95
Guan, L., Scandalios, J.G., 1995. Developmentally related responses of maize catalase genes to salicylic acid. Proceeding of National Academic Sciences, 92, 5930-5934
Gupta, P.P., Tandon, J.S., Patnaik, G.K., 1998. Anti-allergic activity of andrographolides isolated from Andrographis paniculata (Burm F) Wall. Pharmaceutical Biology, 36, 72-74
Harada, T., Ohtaki, E., Misu, K., Sumiyoshi, T., Hososda, S., 2002. Congestive heart failure caused by digitalis toxicity in an elderly man taking a licorice-containing Chinese herbal laxative. Cardiology , 98, 218
Harish, R., Shivanandappa, T., 2006. Antioxidant activity and hepatoprotective potential of Phyllanthus niruri. Food Chemistry, 95, 180-185
Harmatha, J., Nawrot, J., 2002. Insect feeding deterrent activity of lignans and related phenylpropanoids with a methylenedioxyphenyl (Piperonyl) structure moiety. Entomologia Experimentalis et Applicata, 104, 51-60
Hartati, M.S.W., Wahyuono, S., Khasanah, N., 1999. Identification of antimicrobial compound in volatile oil of the tagetes leaves (Tagetes erecta L. fam. Compositae) (in Indonesian). Indonesian Journal of Pharmacy, 10, 40-47
Hass, J.H., Moore, L.W., Ream, W., Manulis, S., 1995. Universal PCR primers for detection of phytophatogenic Agrobacterium strains. Applied Environmental Microbiology. 61, 2879-2884
Huang, R.L., Huang, Y.L., Ou, J.C., Chen, C., Hsu, F.L., Chang, C.M., 2003. Screening of 25 compounds isolated from Phyllanthus species for anti-human hepatitis B virus in vitro. Phytotherapy Research, 17, 449-453
Huang, Y.L., Chen, C.C., Ou, J.C., 1992. Isolintetralin : a new lignan from Phyllanthus niruri. Planta Medica, 58, 473-474
Huntley, A. L., Coon, J. T., Ernst, E., 2005. The safety of herbal medicinal products derived from Echinacea species: A systematic review. Drug Safety, 28, 387-400
Hussein, G., Miyashiro, H., Nakamura, N., Hattori, M., Kakiuchi, N., Shimotohno, K., 2000. Inhibitory effects of Sudanese medicinal plant extracts on hepatitis C virus (HCV) protease. Phytotherapy research, 14, 510-516

-�������� �
111
Iacobellis, N.S., Lo Cantore, P., Capasso, F., Senatore, F., 2005. Antibacterial activity of Cuminum cyminum L. and Carum carvi L. essential oils. Journal of Agricultural and Food Chemistry, 53, 57-61
Ibrahim, R.K., Grisebach, H., 1976. Purification and properties of UDP-glucose: coniferyl alcohol glucosyltransferase from suspension cultures of Paul’s Scarlet Rose. Archives of Biochemistry and Biophysics, 176, 700-708
Ichikawa, H., Takada, Y., Murakami, A., Aggarwal, B.B., 2005. Identification of a novel blocker of I kappa B alpha kinase that enhances cellular apoptosis and inhibits cellular invasion through suppression of NF-kappa B-regulated gene products. Journal of Immunology, 174, 7383-7392
Ikawati, Z., Wahyuono, S., Maeyama, K., 2001. Screening of several Indonesian medicinal plants for their inhibitory effect on histamine release from RBL-2H3 cells. Journal of Ethnopharmacology. 75, 249-256
Imayama, T., Yoshihara, N., Fukuchi-Mizutani, M., Tanaka, Y., Ino, I., Yabuya, T., 2004. Isolation and characterization of a cDNA clone of UDP-glucose: anthocyanin 5-O-glucosyltransferase in Iris hollandica. Plant Cell, 167, 1243-1248
Ishikawa, T., Funahashi, T., Kudo,J., 2000. Effectiveness of the Kampo kami-shoyo-san (TJ-24) for tremor of antipsychotic-induced parkinsonism. Psychiatry and Clinical Neurosciences, 54, 579-582
Ishimaru, K., Yoshimatsu, K., Yamakawa, T., Kamada, H., Shimomura, K., 1992. Phenolic Constituents in Tissue-Cultures of Phyllanthus niruri. Phytochemistry 1992, 31, 2015-2018
Ismail, N., Pihie, A.H.L., Nallapan M., 2005. Xanthorrhizol induces apoptosis via the up-regulation of Bax and p53 in Hela cells. Anticancer Reasearch, 25, 2221-2227
Iwo, M.I., Soemardji, A.A., Retnoningrum, D.S., Sukrasno, Mar’u, U.U., 2000. Immunostimulating effect of Pule (Alstonia scholaris L. R.Br., Apocynaceae) bark extracts. Clinical Hemorheology and Microcirculation, 23, 177-183
Jackson, D.E., Dewick, P.M., 1984. Biosynthesis of Podophyllum Lignans .1. Cinnamic acid precursors of podophyllotoxin in Podophyllum hexandrum. Phytochemistry, 23, 1029-1035
Jagetia, G.C., Baliga, M.S. 2005. The effect of seasonal variation on the antineoplastic activity of Alstonia scholaris R. Br. in HeLa cells. Journal of Ethnopharmacology, 96, 37-42
Jagetia, G.C., Baliga, M.S., 2004. Effect of Alstonia scholaris in enhancing the anticancer activity of berberine in the Ehrlich ascites carcinoma-bearing mice. Journal of Medical Food, 7, 235-244
Janssen, A.M., Scheffer, J.J.C., 1985. Acetoxychavicol acetate, an antifungal component of Alpinia galanga. Planta Medica, 6, 507-511
Ji, L.L., Wang, Z., Dong, F., Zhang, W.B., Wang, Z.T., 2005. Andrograpanin, a compound isolated from anti-inflammatory traditional Chinese medicine Andrographis paniculata, enhances chemokine SDF-1 alpha-induced leukocytes chemotaxis. Journal of Cellular Biochemistry, 95, 970-978
Jirovetz, L., Buchbauer, G., Ngassoum, M.B., Geissler, M., 2002. Aroma compound analysis of Piper nigrum and Piper guineense essential oils from Cameroon using solid-phase microextraction-gas chromatography, solid-phase microextraction-gas chromatography-mass spectrometry and olfactometry. Journal of Chromatography A, 976, 265-275
Jitoe, A., Masuda, T., Tengah, I.G.P., Suprapta, D.N., Gara, I.W., Nakatani, N., 1992. Antioxidant activity of tropical ginger extracts and analysis of the contained curcuminoids. Journal of Agricultural and Food Chemistry, 40, 1337-1340

112
Joseph, H., Gleye, J., Moulis, C., Mensah, L.J., Roussakis, C., Gratas, C., 1988. Justicidin B, a cytotoxic principle from Justicia pectoralis. Journal of Natural Products, 51, 599-600.
Joulain, D., König, W.A., The Atlas of Spectral Data of Sesquiterpene Hydrocarbons. E.B-Verlag, Hamburg, 1998.
Kamei, J., Saitoh, A., Asano, T., Nakamura, R., Ichiki, H., Iiduka, A., Kubo, M., 2005. Pharmacokinetic and pharmacodynamic profiles of the antitussive principles of Glycyrrhizae radix (licorice), a main component of the Kampo preparation Bakumondo-to (Mai-men-dong-tang). European Journal of Pharmacology, 507, 163-168
Kamil, W.D., Dewick, P.M., 1986. Biosynthesis of Podophyllum lignans .4. Biosynthetic relationship of aryltetralin lactone lignans to dibenzylbutyrolactone lignans. Phytochemistry, 25, 2093-2102
Kaminaga, Y., Sahin, F.P., Mizukami, H., 2004. Molecular cloning and characterization of a glucosyltransferase catalyzing glucosylation of curcumin in cultured Catharanthus roseus cells. FEBS Letters, 567, 197-202
Kardono, L.B.S., Angerhofer, C.K., Tsauri, S., Padmawinata, K., Pezzuto, J.M., Kinghorn, A.D., 1991. Cytotoxic and antimalarial constituents of the roots of Eurycoma longifolia. Journal of Natural Products, 54, 1360-1367
Kardono, L.B.S., Tsauri, S., Padmawinata, K., Pezzuto, J.M., Kinghorn, A.D., 1990. Studies on Indonesian Medicinal-Plants 2. Cytotoxic constituents of the bark of Plumeria rubra collected in Indonesia. Journal of Natural Products, 53, 1447-1455
Karunagaran, D., Rashmi, R., Kumar, T.R.S., 2005. Induction of apoptosis by curcumin and its implications for cancer therapy. Current Cancer Drug Target, 5, 117-129
Kassuya, U.A.L., Leite, D.F.P., De Melo, L.V., Rehder, V.L.G., Calixto, J.B., 2005. Anti-inflammatory properties of extracts, fractions and lignans isolated from Phyllanthus amarus. Planta Medica, 71,721-726
Keawpradub, N., Kirby, G.C., Steele, J.C.P., Houghton, P.J., 1999. Antiplasmodial activity of extracts and alkaloids of three Alstonia species from Thailand. Planta Medica, 65, 690-694
Khan, M.R., Omoloso, A.D., Kihara, M., 2003. Antibacterial activity of Alstonia scholaris and Leea tetramera. Fitoterapia, 74, 736-740
Khanna, A.K., Rizvi, R., Chander, R.J., 2002. Lipid lowering activity of Phyllanthus niruri in hyperlipernic rats. Journal of Ethnopharmacology, 82, 19-22
Khanuja, S.P.S., Shasany, A.K., Darokar, M.P., Kumar, S.J., 1999. Rapid isolation of DNA from dry and fresh samples of plants producing large amounts of secondary metabolites and essential oils. Plant Molecular Biology Reporter, 17, 1-7
Kim, I.T., Park, H.J., Nam, J.H., Park, Y.M., Won, J.H., Choi, J., Choe, B.K., Lee, K.T., 2005. In vitro and in vivo anti-inflammatory and antinociceptive effects of the methanol extract of the roots of Morinda officinalis. Journal of Pharmacy and Pharmacology, 57, 607-615
Kim, K.J., Yu, H.H., Cha, J.D., Seo, S.J., Choi, N.Y., You, Y.O., 2005. Antibacterial activity of Curcuma longa L. against methicillin-resistant Staphylococcus aureus. Phytotherapy Research, 19, 599-604
King, S.R., Carlson, T.J., Moran, K., 1996. Biological diversity, indigenous knowledge, drug discovery and intellectual property rights: creating reciprocity and maintaining relationships. Journal of Ethnopharmacology, 51, 45-57

-�������� �
113
Kirana C., Record, I.R., McIntosh, G.H., Jones, G.P., 2003. Screening for antitumor activity of 11 species oh Indonesian Zingiberaceae using human MCF-7 and HT-29 cancer cells. Pharmaceutical Biology, 41, 271-276
Koba, K., Sanda, K., Raynaud, C., Mandin, D., Millet, J., Chaumont, J.P., 2003. Antimicrobial activity of essential oils of Cymbopogon citratus L. (DC) Stapf., C. nardus L. rendle and C. schoenanthus L. Spreng. Journal de Mycologie Medicale, 13, 175-180
Kohara, A., Nakajima, C., Hashimoto, K., Ikenaga,T., Tanaka, H., Shoyama, Y., Yoshida, S., Muranaka, T., 2005. A novel glucosyltransferase involved in steroid saponin biosynthesis in Solanum aculeatissimum. Plant Molecular Biology, 57, 2, 225-239
Konstantinov, S.M., Eibl, H., Berger, M.R., 1999. BCR-ABL influences the antileukaemic efficacy of alkylphosphocholines. British Journal of Haematology, 107, 365-374
Konturek, J.W., Thor, P., Maczka, M., Stoll, R., Domschke, W., Konturek, S.J., 1994. Role of cholecystokinin in the control of gastric emptying and secretory response to a fatty meal in normal subjects and duodenal ulcer patients. Scandinavian Journal of Gastroenterology, 29, 583-590
Koul, J.L., Koul, S.K., Taneja, S.C., Dhar, K.L., 1996. Oxygenated cyclohexanes from Piper cubeba. Phytochemistry, 41, 1097-1099
Koulman, A., Bos, R., Medarde, M., Pras, N., Quax, W.J., 2001. A fast and simple GC-MS method for lignan profiling in Anthriscus sylvestris and biosynthetically related plant species. Planta Medica, 67, 858-862
Kubo, I., Murai, Y., Soediro, I., Soetarno, S., Sastrodihardjo, S., 1992. Cytotoxic Anthraquinones from Rheum palmatum. Phytochemistry, 31, 1063-1065
Kubo, I., Murai, Y., Soediro, I., Soetarno, S., Sastrodihardjo, S., 1991. Efficient isolation of glycosidase inhibitory stilbene glycosides from Rheum palmatum. Journal of Natural Products, 54, 1115-1118
Kubo, M., Ma, S.P., Wu, J.X., Matsuda, H., 1996. Anti-inflammatory activities of 70% methanolic extract from Cinnamomi cortex. Biological & Pharmaceutical Bulletin, 19, 1041-1045
Kumar, R.A., Sridevi, K., Kumar, N.V., Nanduri, S., Rajagopal, S., 2004. Anticancer and immunostimulatory compounds from Andrographis paniculata. Journal of Ethnopharmacology, 92, 291-295
Langmead, L., Makins, R.J., Rampton, D.S., 2004. Anti-inflammatory effects of Aloe vera gel in human colorectal mucosa in vitro. Alimentary Pharmacology & Therapeutics, 19, 521-527
Lawrence, B.M., 1980. New trends in essential oils. Perfumer & Flavorist, 5, 27-32
Leaman, D.J., Arnason, J.T., Yusuf, R., Roemantyo, H.S., Soedjito, H., Angerhofer, C.K., Pezzuto, J.M., 1995. Malaria remedies of the Kenyah of the Apo Kayan, East Kalimantan, Indonesian Borneo: A quantitative assessment of local consensus as an indicator of biological efficacy. Journal of Ethnopharmacology, 49, 1-16
Lee, C.W., Hong, D.H., Han, S.B., Park, S.H., Kim, H.K., Kwon, B.M., Kim, H.M., 1999. Inhibition of human tumor growth by 2 '-hydroxy- and 2 '-benzoyloxycinnamaldehydes. Planta Medica, 65, 263-266
Lee, S.A., Hong, S.S., Han, X.H., Hwang, J.S., Oh, G.J., Lee, K.S., Lee, M.K., Hwang, B.Y., Ro, J.S., 2005. Piperine from the fruits of Piper longum with inhibitory effect on monoamine oxidase and antidepressant-like activity. Chemical & Phamaceutical Bulletin, 53, 832-835

114
Lee, H.S., Ahn, Y.J., 1998. Growth-inhibiting effects of Cinnamomum cassia bark-derived materials on human intestinal bacteria. Journal of Agricultural and Food Chemistry, 46, 8-12
Li, Y., Baldauf, S., Lim, E.K., Bowles, D.J., 2001. Phylogenetic analysis of the UDP-glycosyltransferase multigene family of Arabidopsis thaliana. Journal of Biological Chemistry, 276, 4338-4343
Li, Y., Xu, C., Zhang, Q.A., Liu, J.Y., Tan, R.X., 2005. In vitro anti-Helicobacter pylori action of 30 Chinese herbal medicines used to treat ulcer diseases. Journal of Ethnopharmacology, 98, 329-333
Lim, E.K., Jackson, R.G., Bowles, D.J., 2005. Identification and characterisation of Arabidopsis glycosyltransferases capable of glucosylating coniferyl aldehyde and sinapyl aldehyde. FEBS Letters, 579, 2802-2806
Limyati, D.A., Juniar, B.L.L., 1998. Jamu gendong, a kind of traditional medicine in Indonesia: the microbial contamination of its material and end products. Journal of Ethnopharmacology, 63, 201-208
Lin, S.C., Lin, C.C., Lin, Y.H., Supriyatna, S., Pan, S.L., 1996. The protective effect of Alstonia scholaris R Br on hepatotoxin-induced acute liver damage. American Journal of Chinese Medicine, 24, 153-164
Liu, I.M., Liou, S.S., Lan, T.W., Hsu, F.L., Cheng, J.T., 2005. Myricetin as the active principle of Abelmoschus moschatus to lower plasma glucose in streptozotocin-induced diabetic rats. Planta Medica, 71, 617-621
Liu, J., Lin, H., McIntosh, H., 2001. Genus Phyllanthus for chronic hepatitis B virus infection: a systematic review. Journal of Viral Hepatitis, 8, 358-366
Lo Cantore, P., Iacobellis, N.S., De Marco, A., Capasso, F., Senatore, F., 2004. Antibacterial activity of Coriandrum sativum L. and Foeniculum vulgare Miller var. vulgare (Miller) essential oils. Journal of Agricultural and Food Chemistry, 52, 7862-7866
Lohezic-Le Devehat, F., Bakhtiar, A., Bezivin, C., Amoros, M., Boustie, J., 2002. Antiviral and cytotoxic activities of some Indonesian plant. Fitoterapia, 73, 400-405
Lorenc-Kukula, K., Korobczak, A., Aksamit-Stachurska, A., Kostyn, K., Lukaszewicz, M., Szopa, J., 2004. Glucosyltransferase: The gene arrangement and enzyme function. Cellular & Molecular Biology Letters, 9, 935-946
Ma’at, S. 2002. Protective and immunomodulating effect of phyllanthus niruri's extract in the treatment of cancer. International Journal of Cancer, 100 (S13), 72
MacRae, W.D., Hudson, J.B., Towers, G.H., 1989. The antiviral action of lignans. Planta Medica, 55, 531-535
Manju, V., Nalini, N., 2005. Chemopreventive efficacy of ginger, a naturally occurring anticarcinogen during the initiation, post-initiation stages of 1,2 dimethylhydrazine-induced colon cancer. Clinica Chimica Acta, 358, 60-67
Martins, A.P., Salgueiro, L., Vila, R., Tomi, F., Canigueral, S., Casanova, J., Proença da Cuhna, A., Adzett, T., 1998. Essential oils from four Piper species. Phytochemistry, 49, 2019-2023
Matsubara, T., Bohgaki, T., Watarai, M., Suzuki, H., Ohashi, K., Shibuya, H., 1999. Antihypertensive actions of methylripariochromene A from Orthosiphon aristatus, an Indonesian traditional medicinal plant. Biological & Pharmaceutical Bulletin, 22, 1083-1088

-�������� �
115
Matsuda, H., Morikawa, T., Managi, H., Yoshikawa, M., 2003. Antiallergic principles from Alpinia galanga: Structural requirements of phenylpropanoids for inhibition of degranulation and release of TNF-alpha and IL-4 in RBL-2H3 cells. Bioorganic & Medicinal Chemistry Letters, 13, 3197-3202
Matsuda, H., Pongpiriyadacha, Y., Morikawa, T., Ochi, M., Yoshikawa, M., 2003. Gastroprotective effects of phenylpropanoids from the rhizomas of Alpinia galanga in rats: structural requirements and mode of action. European Journal of Pharmacology, 471, 59-67
Mehrotra, R., Rawat, S., Kulshreshtha, D.K., Patnaik, G.K., Dhawan, B.N., 1990. In vitro studies on the effect of certain natural products against hepatitis B virus. Indian Journal of Medicinal Research, 92, 133-138
Middel, O., Woerdenbag, H.J., Van Uden, W., Van Oeveren, A., Jansen, J.F.G.A., Feringa, B.L., Konings, A.W.T., Pras, N., Kellogg, R.M., 1995. Synthesis and cytotoxicity of novel lignans. Jornal of Medicinal Chemistry, 38, 2112-2117
Millonig, G., Stadlmann, S., Vogel, W., 2005. Herbal hepatotoxicity: Acute hepatitis caused by a Noni preparation (Morinda citrifolia). European Journal of Gastroenterology and Hepatology, 17, 445-447
Miyakoshi, M., Yamaguchi, Y., Takagaki, R., Mizutani, K., Kambara, T., Ikeda, T., Zaman, M. S., Kakihara, H., Takenaka, A., Igarashi, K., 2004. Hepatoprotective effect of sesquiterpenes in turmeric. Biofactors, 21, 167-170
Mohagheghzadeh, A., Schmidt, T.J., Alfermann, A.W., 2002. Arylnaphthalene lignans from in vitro cultures of Linum austriacum. Journal of Natural Products, 65, 69-71
Mojica-Henshaw, M.P., Francisco, A.D., De Guzman, F., Tigno, X.T., 2003. Possible immunomodulatory actions of Carica papaya seed extract. Clinical Hemorheology and Microcirculation, 29, 219-229
Molog, G., Empt, U., Kuhlman, S., Van Uden, W., Pras, N., Alferman, A.W., Petersen, M., 2001. Deoxypodophyllotoxin 6-hydroxylase, a cytochrome P450 monooxygenase from cell cultures of Linum flavum involved in the biosynthesis of cytotoxic lignans. Planta, 214, 288-294
Moraes, R.M., Bedir, E., Barret, H., Burandt Jr., C., Canel, C., Khan, I.A., 2002. Evaluation of Podophyllum peltatum; accessions for podophyllotoxin production. Planta Medica, 68, 341-344
Morikawa, T., Matsuda, H., Yamaguchi, I., Pongpiriyadacha, Y., Yoshikawa, M., 2004. New amides and gastroprotective constituents from the fruit of Piper chaba. Planta Medica, 70, 152-159
Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. Journal of Immunology Methods, 65, 55-63.
MS Library NIST107, National Institute of Standard and Technology.
Murashige, T., Skoog, F., 1962. A Revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum, 15, 473-497
Mutalik, S., Chetana, M., Sulochana, B., Devi, P.U., Udupa, N., 2005. Effect of Dianex, a herbal formulation on experimentally induced diabetes mellitus. Phytotherapy Research, 19, 409-415
Nader, B.L., Cardoso, Taketa, A.T., Iturriaga, G., Pereda-Miranda, R., Villarreal, M.L., 2004. Genetic transformation of Glaphimia glauca by Agrobacterium rhizogenes and the production of norfriedelanes. Planta Medica, 70, 1174-1179

116
Naik, A.D., Juvekar, A.R., 2003. Effects of alkaloidal extract of Phyllanthus niruri on HIV replication. Indian Journal of Medical Sciences, 57, 387-393
Nawawi, A., Nakamura, N., Hattori, M., Kurokawa, M., Shiraki, K., 1999. Inhibitory effects of Indonesian medicinal plants on the infection of herpes simplex virus type 1. Phytotherapy Research, 13, 37-41
Notka, F., Meier, G., Wagner, R., 2004. Concerted inhibitory activities of Phyllanthus amarus on HIV replication in vitro and ex vivo. Antiviral Research, 64, 93-102
Ochman, H., Gerber, A.S., Hartl, D.L., 1988. Genetic applications of an inverse polymerase chain reaction. Genetics, 120, 621-623
Ogata, T., Higuchi, H., Mochida, S., Matsumoto, H., Kato, A., Endo, T., Kaji, A., Kaji, H., 1992. HIV-1 reverse transcriptase inhibitor from Phyllanthus niruri. AIDS Research and Human Retroviruses, 11, 1937-1944
Ohashi, K., Bohgaki, T., Matsubara, T., Shibuya, H., 2000. Indonesian medicinal plants. XXIII. Chemical structures of two new migrated pimarane-type diterpenes, neoorthosiphols A and B, and suppressive effects on rat thoracic aorta of chemical constituents isolated from the leaves of Orthosiphon aristatus (Lamiaceae). Chemical & Pharmaceutical Bulletin, 48, 433-435
Ojewole, J.A.O., 2004. Potentiation of the anti-inflammatory effect of Anacardium occidentale (Linn.) stem-bark aqueous extract by grapefruit juice. Methods and Findings in Experimental and Clinical Phamacology, 26, 183-188
Okazaki, Y., Iqbal, M. Okada, S., 2005. Suppressive effects of dietary curcumin on the increased activity of renal ornithine decarboxylase in mice treated with a renal carcinogen, ferric nitrilotriacetate. Biochimica et Biophysica Acta, 1740, 357-366
Okigawa, M., Maeda, T., Kawano, N., 1970. The isolation and structure of three new lignans from Justicia procumbens Linn. Var. Leucantha Honda. Tetrahedron, 26, 4301–4305
Omar, S., Zhang, J., MacKinnon, S., Leaman, D., Durst, T., Philogene, B.J.R., Arnason, J.T., Sanchez-Vindas, P.E., Poveda, L., Tamez, P.A., Pezzuto, J.M., 2003. Traditionally-used antimalarials from the Meliaceae. Current Topics in Medicinal Chemistry, 3, 137-139
Orav, A., Stylova, I., Kailas, T., Müürisepp, M., 2004. Effect of storage on the essential oil composition of Piper nigrum L. fruits of different ripening states. Journal of Agricultural and Food Chemistry. 52, 2582-2586
Otaka, T., Mori, H., Morimoto, M., Ueba, N., Sutardjo, S., Kusumoto, I.T., Hattori, M., Namba, T., 1995. Screening of Indonesian plant-extracts for anti-human-immunodeficiency-virus type-1 (HIV-1) activity. Phytotherapy research, 9, 6-10
Oyedeji, A.O., Adeniyi, B.A., Ajayi, O., König, W.A., 2005. Essential oil composition of Piper guineense and its antimicrobial activity. Another chemotype from Nigeria. Phytotherapy Research, 19, 362-364
Padma, P., Pramod, N.P., Thyagarajan, S.P., Khosa, R.L., 1998. Effect of the extract of Annona muricata and Petunia nyctaginiflora on herpes simplex virus. Journal of Ethnopharmacology, 61, 81-83
Park, K.M., Cho, J.H., Sohn, J.H., Lee, S.H., Hwang, J.K., 2005. Antibacterial activity of panduratin A isolated from Kaempferia pandurata against Porphyromonas gingivalis. Food Science and Biotechnology, 14, 286-289

-�������� �
117
Parmar, V.S., Jain, S.C., Bisht, K.S., Jain, R., Taneja, P., Jha, A., Tyagi, O.D., Prasad, A.K., Wengel, J., Olsen, C.E., Boll, P.M., 1997. Phytochemistry of the genus Piper. Phytochemistry, 46, 597-673
Paul, J., Duncan, J.R., Sharp, P., Norris, A., Siddiq, M.A., Bacon, C., Weighill, J., 2005. Agranulocytosis and citrobacter infection associated with jamu, a herbal remedy containing phenylbutazone. Clinical Infectious Diseases. 40, 1859-1860
Petersen, M., Alfermann, A.W., 2001. The production of cytotoxic lignans by plant cell cultures. Applied Microbiology & Biotechnology, 55, 135-142
Pettit, G.R., Schaufelberger, D.E., 1988. Isolation and structure of the cytostatic lignan glycoside phyllanthostatin A. Journal of Natural Products, 51, 1104-1112
Poolsup, N., Suthisisang, C., Prathanturarug, S., Asawamekin, A., Chanchareon, U., 2004. Andrographis paniculata in the symptomatic treatment of uncomplicated upper respiratory tract infection: systematic review of randomized controlled trials. Journal of Clinical Pharmacy and Therapeutics, 29, 37-45
Prabhu, B.R., Mulchandani, N.B., 1985. Lignans from Piper cubeba. Phytochemistry, 24, 329-331
Pramono, E., 2002. The traditional use of traditional knowledge and medicinal plants in Indonesia. Multi-Stakeholder dialoque on trade, intellectual property and biological resources in Asia, BRAC Centre for Development Management, Rajendrapur, Bangladesh
Pu, H.F., Huang, W.J., Tseng, W.M., Wang, S.W., Liu, Y.W., Doong, M.L., Wang, P.S., 2004. Effects of juice from Morinda citrifolia (Noni) on gastric emptying in male rats. Chinese Journal of Physiolgy, 47, 169-174
Rajasekaran, S., Sivagnanam, K., Ravi, K., Subramanian, S., 2004. Hypoglycemic effect of Aloe vera gel on streptozotocin-induced diabetes in experimental rats. Journal of Medicinal Food, 7, 61-66
Ram, V.J., 2001. Herbal preparations as a source of hepatoprotective agents. Drug News & Perspective, 14, 353-363
Reddy, S.V., Srinivas, P.V., Praveen, B., Kishore, K.H., Raju, B.C., Murthy, U.S., Rao, J.M., 2004. Antibacterial constituents from the berries of Piper nigrum. Phytomedicine, 11, 697-700
Reed, D.W., Davin, L., Jain, J.C., Deluca, V., Nelson, L., Underhill, E.W., 1993. Purification and properties of UDP-glucose thiohydroxymate glucosyltransferase from Brasica napus L. seedling. Archives Biochemistry and Biophysics, 305, 526-532
Rilantono, L.I., Yuwono, H.S., Hugrahadi, T., 2000. Dietary antioxidative potential in arteries. Clinical Hemorheology and Microcirculation, 23, 113-117
Riswan, S., Roemantyo, H.S., 2002. Jamu as traditional medicine in Java, Indonesia. South Pacific Study, 23, 2-10
Row, L.R., Srinivasulu, C., 1964. New Lignans from Phyllanthus niruri Linn. Tetrahedron Letters, 24, 1557-1567
Rukachaisirikul, T., Siriwattanakit, P., Sukcharoenphol, K., Wongvein, C., Ruttanaweang, P., Wongwattanavuch, P., Suksamrarn, A., 2004. Chemical constituents and bioactivity of Piper sarmentosum. Journal of Ethnopharmacology, 93, 173-176
Runnie, I., Salleh, M.N., Mohamed, S., Head, R.J., Abeywardena, M.Y., 2004. Vasorelaxation induced by common edible tropical plant extracts in isolated rat aorta and mesenteric vascular bed. Journal of Ethnopharmacology, 92, 311-316

118
Sabino, M.A.C., Ghilardi, J.R., Jongen, J.L.M., Keyser, C.P., Luger, N.M.. Mach, D.B., Peters C.M., Rogers, S.D., Schwei, M.J., De Felipe, C., Mantyh, P.W., 2002. Studies on disease-modifying antirheumatic drugs: synthesis of novel quinoline and quinazoline derivatives and their anti-inflammatory effect. Cancer Research, 62, 7343–7349
Santos, A.R., Filho, V.C., Niero, R., Vianna, A.M., Moreno, F.N., Campos, M.M., Yunes, R.A., Calixto, J.B., 1994. Analgesic effects of callus culture extracts from selected species of Phyllanthus in Mice. Journal of Pharmacy and Pharmacology, 46, 755-759
Sasaki, N., Wada, K., Koda, T., Kasahara, K., Adachi, T., Ozeki, Y., 2005. Isolation and characterization of cDNAs encoding an enzyme with glucosyltransferase activity for cyclo-DOPA from four o’clocks and feather cockscombs. Plant and Cell Physiology, 46, 666-670
Satroamidjojo, 2001. Obat Asli Indonesia, Dian Rakyat, Jakarta
Satyanarayana, P., Venkateswarlu, 1991. Isolation, structure and synthesis of new diarylbutane lignans from Phyllanthus niruri - synthesis of 5'-desmethoxy niranthin and an antitumor extractive. Tetrahedron, 47, 8931-8940
Satyanarayana, P., Subrahmanyam, P., Viswanatham, K.N., 1988. New seco- and hydroxyl-lignans from Phyllanthus niruri. Journal of Natural Products, 51, 44-49
Sayyah, M., Mahboubi, A., Kamalinejad, M., 2002. Anticonvulsant effect of the fruit essential oil of Cuminum cyminum in mice. Pharmaceutical Biology, 40, 478-4809
Schmid, G., Grisebach, H., 1982. Enzymatic synthesis of lignin precursor: purification and properties of UDP-glucose: coniferyl alcohol glucosyltransferase from cambial sap of spruce (Picea abies L.). European Journal of Biochemistry, 123, 362-370
Seidel, V., Windhövel, J., Eaton, G., Alfermann, A.W., Arroo, R.R.J., Medarde, M., Petersen, M., Woolley, J.G. 2002. Biosynthesis of podophyllotoxin in Linum album cell cultures. Planta, 215, 1031-1039
Sharma, R.A., Gescher, A.J., Steward, W.P., 2005. Curcumin: The story so far. European Journal of Cancer, 41, 1955-1968
Sheena, N., Ajith, T.A., Janardhanan, K.K., 2003. Anti-inflammatory and anti-nociceptive activities of Ganoderma lucidum occurring in South India. Pharmaceutical Biology, 41, 301-304
Shibuya, H., Kitagawa, I., 1996. Chemical study of Indonesian medicinal plants. Yakugaku Zasshi-Journal of the Pharmaceutical Society of Japan, 116, 911-927
Shimizu, M., Horie, S., Terashima, S., Ueno, H., Hayashi, T., Arisawa, M., Suzuki, S., Yshizaki, M., Morita, N., 1989. Studies on aldose reductase inhibitors from natural-products. 2. Active components of a Paraguayan crude drug para-parai mi, Phyllanthus-niruri. Chemical & Pharmaceutical Bulletin, 37, 2531-2532
Singha, P.K., Roy, S., Dey, S., 2003. Antimicrobial activity of Andrographis paniculata. Fitoterapia, 74, 692-694
Singh, B., Agrawal, P.K., Thakur, R.S., 1989. A new lignan and a new neolignan from Phyllanthus niruri. Journal of Natural Products, 52, 48-51
Slightom, J.L., Durand-Tardif, M., Jouanin, L., Tepfer, D., 1986. Nucleotide sequence analysis of TL-DNA of Agrobacterium rhizogenes agropine type plasmid. Identification of open reading frames. Journal of Biology and Chemistry, 261, 108–121

-�������� �
119
Smollny, T., Wichers, H. J., Kalenberg, S., Shahsavari, A., Petersen, M., Alfermann, A.W., 1998. Accumulation of podophyllotoxin and related lignans in cell suspension cultures of Linum album. Phytochemistry, 48, 975-979
Soedjarto, 1996. Biodiversity prospecting and benefit-sharing: perspectives from the field. Journal of Ethnopharmacology, 51, 1-15
Sripanidkulchai, B., Wongpanich, V., Laupattarakasem, P., Suwansaksri, J., Jirakulsomchok, D., 2001. Diuretic effects of selected Thai indigenous medicinal plants in rats. Journal of Ethnopharmacology, 75, 185-190
Sridhar, S.B., Sheetal, U.D., Pai, M.R.S.M., Shastri, M.S., 2005. Preclinical evaluation of the antidiabetic effect of Eugenia jambolana seed powder in streptozotocin-diabetic rats. Brazilian Journal of Medical an Biological Research, 38, 463-468
Steffan, R., Watjen, W., Michels, G., Niering, P., Wray, V., Ebel, R., Edrada, R., Kahl, R., Proksch, P., 2005. Polyphenols from plants used in traditional Indonesian medicine (Jamu): uptake and antioxidative effects in rat H4IIE hepatoma cells. Journal of Pharmacy and Pharmacology, 57, 233-240
Stevinson, C., Huntley, A., Ernst, E., 2002. A systematic review of the safety of kava extract in the treatment of anxiety. Drug Safety, 25: 251-261
Subeki, Matsura, H., Takahashi, K., Yamasaki, M., Yamato, O., Maede, Y., Katakura, K., Suzuki, M., Trimurningsih, Chairul, Yoshihara,T., 2005. Antibabesial activity of protoberberine alkaloids and 20-hydroxyecdysone from Arcangelisia flava against Babesia gibsoni in culture. Journal of Veterinary Medical Science, 66, 871-874
Subeki, S., Matsuura, H., Takahashi, K., Yamasaki, M., Yamato, O., Maede, Y., Katakura, K., Kobayashi, S., Trimurningsih,, Chairul, Yoshihara T., 2005. Anti-babesial and anti-plasmodial compounds from Phyllanthus niruri. Journal of Natural Products, 68, 537-539
Suharmiati, 2003. Menguak tabir dan potensi jamu gendong. Agromedia Pustaka, Jakarta
Sukandar, E.Y., Suganda, A.G., Muslikhati., 1999. The effect of the volatile oil of bark and leaves of Cinnamomum burmanni against bacteria and fungi (in Indonesian). Indonesian Journal of Pharmacy, 10, 31-39
Sukardiman, Darwanto, A., Tanjung, M., Darmadi, M.A., 2000. Cytotoxic mechanism of flavonoid from temu kunci (Kaempferia pandurata) in cell culture of human mammary carcinoma. Clinical Hemorheology and Microcirculation, 23, 185-190
Sumathykutty, M.A., Rao, J.M., Padmakumari, K.P., Narayanan, C.S., 1999. Essential oil constituents of some Piper species. Flavour and Fragrance Journal, 14, 279-282
Svensson, B., Pettersson, H., 2003. Reumacon (CPH82) showed similar x-ray progression and clinical effects as methotrexate in a two year comparative study on patients with early rheumatoid arthritis. Scandinavian Journal of Rheumatology, 32, 83-88
Syamasundar, K.V., Singh, B., Thakur, R.S., Husain, A., Kiso, Y., Hikino, H., 1985. Series on liver-protective drugs .19. Antihepatotoxic principles of Phyllanthus niruri herbs. Journal of Ethnopharmacology, 14, 41-44

120
Taguchi, G., Yazawa, T., Hayashida, N., Okazaki, M., 2001. Molecular cloning and heterologous expression of novel glucosyltransferase fro tobacco cultured cells that have broad substrate specificity and are induced by salicylic acid and auxin. European Journal of Biochemsitry, 268, 4086-4094
Takara, K., Horibe, S., Obata, Y., Yoshikawa, E., Ohnishi, N., Yokoyama, T., 2005. Effects of 19 herbal extracts on the sensitivity to paclitaxel or 5-fluorouracil in HeLa cells. Biological & Pharmaceutical Bulletin, 28, 138-142
Tan, M.L., Kuroyanagi, M., Sulaiman, S.F., Najimudin, N., Muhammad, T.S.T., 2005. Cytotoxic activities of major diterpenoid constituents of Andrographis paniculata in a panel of human tumor cell lines. Pharmaceutical Biology, 43, 501-508
Taneja, S.C., Koul, S.K., Pushpangadan, P., Dhar, K.L., Daniewski, W.M., Schilf, W., 1991. Oxygenated cyclohexanes from Piper species. Phytochemistry, 30, 871-874
Tawata, M., Aida, K., Noguchi, T., Ozaki, Y., Kume, S., Sasaki, H., Chin, M., Onaya, T., 1992. Antiplatetlet action of isoliquiritigenin, An aldose reductase inhibitor in licorice. European Journal of Pharmacology, 212, 87-92
Tchoumbougnang, F., Zollo, P.H.A., Dagne, E., Mekonnen, Y., 2005. In vivo antimalarial activity of essential oils from Cymbopogon citratus and Ocimum gratissimum on mice infected with Plasmodium berghei. Planta Medica, 71, 20-23
Tezuka, Y., Terazono, M., Kusumoto, T.I., Hatanaka, Y., Kadota, S., Hattori, M., Namba, T., Kikuchi, T., Tanaka, K., Supriyatna, S., 2000. Helicterins A-F, six new dimeric (7.5 ',8.2 ')-neolignans from the Indonesian medicinal plant Helicteres isora. Helvetica Chimica Acta, 2908-2919
Tinker, A.C., Wallace, A.V., 2006. Selective inhibitors of inducible nitric oxide synthase: potential agents for the treatment of inflammatory diseases. Current Topic of Medicinal chemistry, 6, 77-92
Tona, L., Cimanga, R.K., Mesia, K., Musuamba, C.T., De Bruyne, T., Apers, S., Hernans, N., Van Miert, S., Pieters, L., Totte, J., Vlietinck, A.J., 2004. In vitro antiplasmodial activity of extracts and fractions from seven medicinal plants used in the Democratic Republic of Congo. Journal of Ethnopharmacology, 93, 27-32
Tona, L., Mesia, K., Ngimbi, N.P., Chrimwami, B., Okond’ahoka, Cimanga, K., de Bruyne, T., Apers, S., Hermans, N., Totte, J., Pieters, L., Vlietinck, A.J., 2001. In-vivo antimalarial activity of Cassia occidentalis, Morinda morindoides and Phyllanthus niruri. Annals of Tropical Medicine and Parasitology, 95, 47-57
Trimurningsih, Subeki, Matsuura, H., Takahashi, K., Yamasaki, M., Yamato, O., Maede, Y., Katakura, K., Suzuki, M., Kobayashi, S., Chainul, Yoshihara, T., 2005. Evaluation of the inhibitory activities of the extracts of Indonesian traditional medicinal plants against Plasmodium falciparum and Babesia gibsoni. Journal of Veterinary Medical Sciences, 67, 829-831
Tuchinda, P., Reutrakul, V., Claeson, P., Pongprayoon, U., Sematong, T., Santisuk, T., Taylor, W.C., 2002. Anti-inflammatory cyclohexenyl chalcone derivatives in Boesenbergia pandurata. Phytochemistry, 59, 169-173
Udoh, F.V., Udoh, P.B., 2005. Hepatotoxicity of the methanol extract of Carica papaya (paw-paw) seeds in Wistar rats. Pharmaceutical Biology, 43, 349-352
United Nations, Convention on Biological Diversity, Rio de Janeiro, 1992

-�������� �
121
Unny, R., Chauhan, A.K., Joshi, Y.C., Dobhal, M.P., Gupta, R.S., 2003. A review on potentiality of medicinal plants as the source of new contraceptive principles. Phytomedicine, 10, 233-260
Usia, T., Watabe, T., Kadota, S., Tezuka, Y., 2005. Potent cyp3A4 inhibitory constituents of Piper cubeba. Journal of Natural Products, 68, 64-68
Usia, T., Watabe, T., Kadota, S., Tezuka, Y., 2005. Metabolite-cytochrome P450 complex formation by methylenedioxyphenyl lignans of Piper cubeba: mechanism-based inhibition. Life Sciences, 76, 2381-2391
Van Uden, W., Pras, N., Malingré, T.M., 1990a On the improvement of the podophyllotoxin production by phenylpropanoid precursor feeding to cell cultures of Podophyllum hexandrum Royle. Plant Cell Tissue and Organ Culture, 23, 217-224
Van Uden, W., Pras, N., Malingré, T.M., 1990b. The accumulation of podophyllotoxin-β-D-glucoside by cell suspension cultures derived from the conifer Callitris drummondii. Plant Cell Reports, 9, 257-269
Van Uden, W., Homan, B., Woerdenbag, H.J., Pras, N., Malingré, T.M., Wichers, H.J., Harkes, M., 1992. Isolation, purification, and cytotoxicity of 6-methoxypodophyllotoxin, a lignan from root cultures of Linum flavum L. Journal of Natural Products, 55, 102-110
Van Uden, W., Pras, N., Homan, B., Malingré, T.M., 1991a. Improvement of 5-methoxypodophyllotoxin using a new selected root culture of Linum flavum L. Plant Cell Tissue and Organ Culture, 27, 115-121
Van Uden, W., Pras, N., Batterman, S., Visser, J.F., Malingré, T.M., 1991b. The accumulation and isolation of coniferin from a high-producing cell suspension of Linum flavum L. Planta 183: 25-30
Van Uden, W., Bos, J.A., Boeke, G.M., Woerdenbag, H.J., Pras, N., 1997. The large scale isolation of deoxypodophyllotoxin from rhizomas of Antriscus sylvestris followed by its bioconversion into 5-methoxypodophyllotoxin β-D-glucoside by cell cultures of Linum flavum. Journal of Natural Products, 60, 401-403
Vasilev, N.P., Ionkova, I., 2005. Cytotoxic activity of extracts from Linum cell cultures. Fitoterapia, 76, 50-53
Vasilev, N., Momekov, G., Zaharieva, M., Konstantinov, S., Bremner, P., Heinrich, M., Ionkova, I., 2005. Cytotoxic activity of a podophyllotoxin-like lignan from Linum tauricum Willd. Neoplasma, 52, 425-429
Violon, C., Simeray, J., Leger, D., Chaumont, J. P., 1991 Plantes Med. Phytother. 1991, 25, 118-122
Vogt, T., Jones, P., 2000. Glycosyltransferases in plant natural products synthesis: characterization of a supergene family. Trends in Plant Science, 5, 380-386
Vogt, T., Zimmermann, E., Grimm, R., Meyer, M., Strack, D., 1997. Are the caracteristics of betanidin glucosyltransferases from cell suspension cultures of Dorotheanthus bellidiformis indicative of their phylogenetic relationship with flavonoid glucosyltransferases?. Planta, 2003, 349-361
Walsky, R.L., Obach, R.S., 2004. Validated assays for human cytochrome P450 activities. Drug Metabolism and Disposition, 32, 647-660
Wang, X.S., Liu, L., Fang, J.N., 2005. Immunological activities and structure of pectin from Centella asiatica. Carbohydrate Polymers, 60, 95-101
WHO strategy for traditional medicine 2002-2005, Provisional Agenda, Regional Committee, p. 1-8

122
Woerdenbag, H.J., Van Uden, W., Frijlink, H.W., Lerk, C.F., Pras, N., Malingré, T.M., 1990. Increased podophyllotoxin production in Podophyllum hexandrum cell suspension cultures after feeding coniferyl alcohol as a �-cyclodextrin complex. Plant Cell Reports, 9, 97-100
Wunder, D., William, H.B., 1999. Action of agent on glucosyltransferase from Streptococcus mutans in solution and adsorbed to experimental pellicle. Archives Oral Biology, 44, 203-214
Xu, Y., Ku, B.S., Yao, H.Y., Lin, Y.H., Ma, X., Zhang, Y.H., Li, X.J., 2005. Antidepressant effects of curcumin in the forced swim test and olfactory bulbectomy models of depression in rats. Pharmacology Biochemistry and Behavior, 82, 200-206
Yang, B.K., Wilson, M.A., Cho, K.Y., Song, C.H., 2004. Hypoglycemic effect of exo- and endo-biopolymers produced by submerged mycelial culture of Ganoderma lucidum in streptozotocin-induced diabetic rats. Journal of Microbiology and Biotechnology, 14, 972-977
Yu, B.C., Hung, C.R., Chen, W.C., Cheng, J.T., 2003. Antihyperglycemic effect of andrographolide in streptozotocin-induced diabetic rats. Planta Medica, 69, 1075-1079
Yun, J.M., Kwon, H.J., Mukhtar, H., Hwang, J.K., 2005. Induction of apoptosis by panduratin a isolated from Kaempferia pandurata in human colon cancer HT-29 cells. Planta Medica, 701, 501-507
Yusuf, S., Agunu, A., Diana, M., 2004. The effect of Aloe vera A. Berger (Liliaceae) on gastric acid secretion and acute gastric mucosal injury in rats. Journal of Ethnopharmacology, 93, 33-37
Zhang, C.Y., Kuroyangi, M., Tan, B.K., 1998. Cardiovascular activity of 14-deoxy-11,12-didehydroandrographolide in the anaesthetised rat and isolated right atria. Pharmacological Research, 38, 413-417
Zhang, C.Y., Tan, B.K.H., 1996. Hypotensive activity of aqueous extract of Andrographis paniculata in rats. Clinical and Experimental Pharmacology and Physiology, 23, 675-678
Zheng, G.Q., Kenney, P.M., Lam, L.K.T., 1993. Potential anticarcinogenic natural products isolated from lemongrass oil and galanga root oil. Journal of Agricultural and Food Chemistry, 41, 153-156
Zhou, Y., Hu, C.P., Deng, P.Y., Deng, H.W., Yuan-Jian, L., 2004. The protective effects of ligustrazine on ischemia reperfusion and DPPH free radical induced myocardial injury in isolated rat hearts. Planta Medica, 70, 818-822
Zuhud, E.A.M., Aziz, S., Ghulamahdi, M., Andarwulan, N., Darusman, L.K., 2001. Dukungan teknologi pengembangan obat asli Indonesia dari segi budaya, pelestarian dan pasca panen. Paper presented at the workshop on agribusiness development based on biopharmaca, 13-15 November 2001, Departemen Pertanian, Jakarta, Indonesia

123
Curriculum Vitae Elfahmi was born on 25 April 1969 in Ampang Pulai, a village 56 km South of Padang, West
Sumatera, Indonesia. He finished the primary and elementary school in the village where he was born. In 1989, he moved from Ampang Pulai to Padang when he passed the university entrance examination, for the Department of Pharmacy, Andalas University. He obtained his doctorandus degree in 1993 and became a pharmacist (apoteker) in 1995. Immediately after completing his study in Padang, he received a grant from the Indonesian government through the University Research and Undergraduate Education (URGE) project for his master degree. He obtained his master degree from the Department of Pharmacy (since 2006 named School of Pharmacy), Institut Teknologi Bandung (ITB) in 1997. He started his PhD study in June 2001 at the Department of Pharmaceutical Biology, University of Groningen with the research topic plant biotechnology. His PhD study was financially supported by the Quality for Undergraduate Education (QUE) project of the Department of Biology, ITB, and partly by the University of Groningen. In July 2006, he will continue his work at the School of Pharmacy, ITB, as a lecturer and researcher. During his study in Groningen he got a chance to attend scientific meetings in several countries to present his results and to publish his research output as listed below.
List of publications
1. Elfahmi, Batterman, S., Koulman, A., Hackl, T., Bos, R., Kayser, O., Woerdenbag, H.J., Quax, W.J., 2006. Lignans from cell suspension cultures of Phyllanthus niruri, an Indonesian medicinal plants. Journal of Natural Products, 2006, 55-58
2. Vasilev, N., Elfahmi, Bos, R., Kayser, O., Momekov, G., Konstantinov, S., Ionkova, I., 2006. Production of cytotoxic arylnaphthalene lignan from genetically transformed root cultures of Linum leonii. Journal of Natural Products (in press)
3. Elfahmi, Ruslan, K., Bos, R., Kayser, O., Woerdenbag, H.J., Quax, W.J. Jamu: The Indonesian traditional herbal medicine (submitted)
4. Elfahmi, Ruslan, K., Batterman, S., Bos, R., Kayser, O., Woerdenbag, H.J., Quax, W.J. Lignan profile of Piper cubeba, an Indonesian medicinal plant (submitted)
5. Elfami, Rein Bos, Komar Ruslan, Herman J. Woerdenbag, Oliver Kayser, and Wim J. Quax. Essential oil constituents of Piper cubeba from Indonesia (submitted)
6. Elfahmi, Sieb Batterman, Albert Koulman, Henricus L. Hoge, Rein Bos, Oliver Kayser, Herman J. Woerdenbag, Wim J. Quax. Reduced coniferin and enhanced 6-methoxypodophyllotoxin production in Linum flavum cell suspension cultures after treatment with Na2EDTA (submitted)

124

125
Acknowledgements Alhamdulillah, finally this thesis is finished. This could not happen without the support from
many people, especially from the members of the Pharmaceutical Biology group, University of Groningen. Their support does not only include the scientific and academic aspects, but also the social life during the years of my stay in the nice small city (fietsstad), Groningen. I would like to express my gratitude to all of you.
I would like to thank the QUE Project of the Department of Biology, Institut Teknology Bandung (ITB) for financial support during my PhD study, especially to Dr. Agus Dana Perdana and Dr. Taufikurrahan, the directors of the project. My research was also partially supported by University of Groningen through a scholarship.
I wish to thank my promotor Prof. Dr. Wim J. Quax who gave me a chance to do research for my doctoral degree in his department, for his support and approval of this thesis. Together with Dr. Niesko Pras, you introduced me to the scientific atmosphere in your country. Although it was only for short time being supervised by Niesko, I appreciate it so much.
I would like to express my sincere appreciation to my copromotors, Dr. Herman Woerdenbag and Dr. Oliver Kayser, for patience and guidance during my PhD period. Herman, you have always tried to help me to find the solution for a problem I had. I know that you left the lab long time ago, but your knowledge in practical manners still helps me a lot. Fast in answers, immediately in responding emails, excellent in English, fast in correcting manuscript are things that I will never forget. It was nice when I tried to send you an email in Dutch, you replied in Indonesian. Good luck with your Indonesian course, and then we can continue talking in such way.
Working together in the lab with colleagues has stimulated me to finish my study. Therefore, many thanks I want to address to my colleagues especially to my roommate Mattijs Julsing for friendship, help and nice discussions. Sometime we had a quite hard discussion, even people thought that we were fighting, but actually we were sharing ideas. I really enjoyed it, my best friend, Mattijs. Albert Koulman, I remember that you sent me much literature before I started my study here, followed by a lot of help and discussion concerning the research project. It is pity that you will not be here when I defend my thesis. My thanks also go to other colleagues, Monique (ex-roommate), Almer, Charles, Lidia, Johanna, Michiel, Asia, Agata, Ana, Ykelin, Henco, Ronald D., Ronald M., Robert, Linda, Helga, Geeske, Nickolay, Carlos, Mariana, Melloney, Mags, and Peter. I want to thank my paranimfen Mattijs and M. Chalid for helping me to arrange my deffence and subsequent party.
I greatly appreciate the important assistance from Rein Bos, Sieb Battermann, Janita Zwinderman and Freeke van Putten. Rein’s expertise in analytical instruments (GC, GC-MS, HPLC) helped me a lot. You are hand van de meester and more than that, you also supervised me in many things. Sieb, you have prepared and maintained the plant cell culture that I have used for my research project. You also introduced me to some nice Gronings words. Janita, I thank you for helping me to arrange administration stuffs. Freeke, you helped me to prepare samples to be analysed. I appreciate it.
I would also like to thank Prof. Jacques Hille, Dr. Paul Dijkwel and Drs. Marcel G. Sturee who gave me an opportunity to do work at the Department of Plant Molecular Biology in Haren. Paul and Marcel your expertise in plant genetics improved my knowledge in this field that I just entered. I am glad that I have good results on this topic although the work has to be continued to yield a nice publication.
I am grateful to the members of the reading committee, Prof. Dr. F.A.J. Muskiet, Prof. Dr. T.J. Schmidt and Prof. Dr. R. Verpoorte, for the evaluation of the manuscript and for their valuable comments.

126
I want to thank my colleagues as well as my neighbours in Groningen, Heni Rachmawati, Ikhlasul Amal and their family for many things. My thanks also go to all members of deGromiest (the Indonesian Moslim Society in Groningen) for friendship. My family felt at home being together with you all. It was nice to have routine gathering, discussion, tadarus, sharing ideas. I hope it can be kept and developed further. To all members of PPI (Persatuan Pelajar Indonesia, Indonesian student society in Groningen), I thanks you for your moral support.
My warmest gratitude goes to my colleagues in several cities in the Netherlands, Deden, Iskandar, Zul, Zayd, Gun, Indra, Hasyim, Didin, Lukman, Satria, Iman, Billi and Oka for many helps and kind friendship. Your houses were “my hotels” when I travelled to your city.
I would like to thank Martin and Sanneke Broekhuijsen for hospitality and friendship. My wife learned Dutch from you and you learned Indonesian from her. It was nice time to have dinners at your house. Martin, you are a good cooker of lekkere dingen. Also to Henk and Aaf Euverman, I enjoyed to stay in your house before my family came. Unfortunately I miss the contact with you. Tante Ice, Tante Caroline, Mbak Nanie, Pak Ahmad, Mbak Ellis, Wak Asyiah, I thank you for your moral support and we keep in contact.
My best gratitude goes to my parents and my brothers Da Manto, Da Piyan, Ef and my sister Uni Etimurni for support in many ways, praying and a lot of other things you spent to help me. You have stimulated me to develop my career.
Finally I come to address my appreciation to my wife, Yulia Helmi. It is difficult to find words to express my gratitude for you. I was happy to live in Groningen with you and our children, although we were far from our home country. My children, Fathiya Mufidah, Nurul Rahimah and Ahmad Muzakki Imanullah have made me enjoy the life. Unfortunately you will not be here when I defend my thesis, a moment when family and colleagues come together. You have to go back to work and that was our best decision. I am sure that you will enjoy it as well
I am very sorry that I am not able to mention all people who have given a contribution to this thesis. I am sure that some people will be missing to be mentioned. I would like to thank all of you.
Thank you very much Dank u wel Terima kasih





























