Embed Size (px)
Citation preview

Phylogeny and species delimitation in the genus Coprinelluswith special emphasis on the haired species
Laszlo G. Nagy1
Judit HaziCsaba VagvolgyiTamas Papp
Department of Microbiology, Faculty of Science andInformatics, University of Szeged, Kozep fasor 52,H-6726 Szeged, Hungary
Abstract: We inferred the phylogenetic structureand species limits within Coprinellus by using a newlygenerated multigene alignment of LSU, ITS and b-tubulin sequences. We sampled 154 specimens of 71species out of ca. 80 known taxa in Coprinellus andinferred phylogenetic relationships by BayesianMCMC and ML bootstrapping. Morphological andphylogenetic analyses revealed 17 new species, five ofwhich are described herein. Coprinellus was split intothree large clades, one comprising species with aninitially continuous sheath of veil (Domestici/Micaceiclade) and two consisting of primarily setulose(haired) species (eurysporoid and Core Setulosiclades). The separation of veiled species and theclade structure therein corresponds well to themorphology-based sectional classification, althoughthe inclusion of certain setulose taxa (e.g. C.disseminatus, C. verrucispermus, C. curtus etc.) inthese clades necessitates a new morphological defini-tion for the sections. Morphological traits arediscussed for all clades of Coprinellus. Species limitsin the eurysporoid and Core Setulosi clades arescrutinized in detail. Future directions of species-rankresearch and an artificial key to the recovered setulosespecies of Coprinellus is presented.
Key words: Coprinus sensu lato, Domestici, Micacei,new species, pilocystidia, Setulosi
INTRODUCTION
Molecular phylogenies have significantly rearrangedour views on the taxonomy of coprinoid mushroomsin the past few years (Hopple and Vilgalys 1994, 1999;Walther et al. 2005; Larsson and Orstadius 2008;Vasutova et al. 2008; Padamsee et al. 2008; Nagy et al.2010, 2011). The splitting of species of the collectivegenus Coprinus into Coprinopsis, Parasola, Coprinellusand some residual species not placed in any of the
former genera (Redhead et al. 2001, Redhead 2001,Simpson and Grgurinovic 2001) has received muchattention and general acceptance among taxono-mists. Yet the infrageneric relationships and theaffinities of the residual species have remained largelyuntested on a molecular phylogenetic basis. Despitegreat morphological diversity, Coprinopsis and Copri-nellus has not been investigated or split further whilethe paraphyly of Psathyrella has received moreattention (Larsson and Orstadius 2008, Vasutova etal. 2008, Padamsee et al. 2008). Molecular studiesrevealed a noteworthy infrageneric phylogeneticstructure both in Coprinellus and Coprinopsis (Nagyet al. 2010, 2011), whereas Parasola appeared to beuniform in accordance with its low morphologicalvariation (Nagy et al. 2009). This was particularlystriking in Coprinellus, where multigene phylogeniescalled the monophyly of the genus into question(Nagy et al. 2011). Subsequently the monophyly ofthe two large clades (Coprinellus p.p.1 and 2.) hasbeen established and the conflicting results attributedto MCMC convergence or taxon sampling issues(Nagy pers comm). The previously inferred infra-generic structure is particularly noteworthy withinCoprinellus. Studies outlined two large clades ofCoprinellus (Walther et al. 2005; Vasutova et al.2008; Nagy et al. 2010, 2011), one in a significantoverlap with subsection Setulosi, the other withsubsections Domestici and Micacei of a morphology-based subdivision of Coprinus s.l. (Partick 1977, Ulje2005, Orton and Watling 1979). Despite the overlapthese clades are in conflict with the sectionalsubdivisions proposed by Ulje (1991), Singer (1986),Kuhner and Romagnesi (1953) and Orton andWatling (1979), which merits further study.
At the time of splitting Coprinus s.l. into Coprinop-sis, Coprinellus and Parasola, in the absence ofsequence information, several species with ambiguousaffinities have not been placed in any of the newlyerected genera (Redhead et al. 2001). These includeseveral taxa of the cortinati group (sensu Ulje 2005),that is C. cortinatus, C. bellulus etc., certain membersof section Narcotici, some lesser known taxa and C.patouillardii (5 C. cordisporus). Subsequent to therearrangement of Coprinus s.l. into segregate genera(Redhead et al. 2001), several species, including thosedescribed by Ulje and Keizer (2003), Ulje andNoordeloos (2003), Ulje and Verkeben (2002), Nagy(2006), have been recombined in Coprinellus based
Submitted 12 May 2011; accepted for publication 13 Jul 2011.1 Corresponding author. E-mail: [email protected]
Mycologia, 104(1), 2012, pp. 254–275. DOI: 10.3852/11-149# 2012 by The Mycological Society of America, Lawrence, KS 66044-8897
254

on phylogenetic and/or morphological evidence(Simpson and Grgurinovic 2001). Of these taxaDoveri et al. (2011) recombined C. allovelus, C.canistri, C. callinus var. limicola, C. minutisporus andC. pseudoamphithallus while Hazi et al. (2010) andRoux and Garcia (2006) placed C. doverii, C.parvulus, C. pusillulus, C. pallidissimus and C. bipellisin Coprinellus respectively. However a classification inthe new system of coprionoid fungi has not yet beenproposed for several well known or recently describedspecies.
Setulose species of Coprinellus have been subject toa commonly recognized, narrow morphological spe-cies concept that has been uniform among authors(Lange 1952, Lange and Smith 1953, Ulje and Bas1991, Doveri et al. 2011, Nagy 2006, Hazi et al. 2010).This is based on (conjectured) low morphologicalplasticity within species and was partially supported bythe results of mating experiments (Kemp 1975, Lange1952). Indeed most species have low intraspecificvariability and are easily identified on the basis ofmicromorphology. However a comprehensive exam-ination of herbarium materials revealed a largenumber of deviating specimens with considerablevariation, primarily in spore shape, size and cheilo-cystidia. This variation would qualify the respectivespecimens as new species based on the currentlyaccepted narrow morphological species concept.Molecular phylogenetic approaches permit a criticaltest of this morphological species concept by com-paring the phylogenetic placement and branchlengths of the sequenced specimens to that of theirmorphological traits, which we undertake in thispaper.
In view of the above-mentioned points in this studywe assemble a new multigene dataset of Coprinellusand use the resulting phylogeny to analyze (i) thephylogenetic structure within Coprinellus, (ii) thelarge-scale polyphyly of species with hairs (pileo- andcaulocystidia), that is those of subsection Setulosi, (iii)species limits within setulose species and (iv) whethera narrow species concept is applicable across setulosetaxa as suggested previously.
MATERIALS AND METHODS
Taxon sampling.—An attempt was made to sequence alldescribed species of Coprinellus. Herbarium specimens werecollected to achieve a uniform coverage across all taxa byselecting 2–6 specimens per species. Material was obtainedfrom the herbaria WU, M, WTU, L, K, MICH as well as ourown collection of coprinoid species (SZMC, Szeged Micro-biological Collections). Specimens originating from twomonographic treatments of coprinoid fungi of C.B. Ulje(Ulje and Bas 1991, Ulje and Keizer 2003, Ulje andNoordeloos 2003, Ulje and Verkeben 2002) and F. van de
Bogart (1975) also were revised. Type collections wereincluded whenever the condition of the material alloweddestructive sampling. Previous work indicated a closerelationship between Ps. candolleana and Coprinellus (e.g.Hopple and Vilgalys 1999, Padamsee et al. 2008, Nagy et al.2011), so we used this group as outgroup in our analyses.
DNA extraction and sequencing.—Genomic DNA wasextracted with the DNEasy Plant Mini Kit (QIAGEN),following the manufacturer’s protocol with minor adjust-ments to the fruiting bodies of certain species. Primer setsand PCR amplification of the ITS, LSU and b-tubulin geneswere performed as described by Nagy et al. (2011). Weamplified the ITS and b-tubulin fragments for all speci-mens, while the LSU region was targeted only for one or fewrepresentative collection(s) per species. In some cases onlypoor quality DNA extracts could be produced because ofthe diminutive fruiting bodies. In these cases PCR amplifi-cation of all loci employed the JumpStart Taq DNApolymerase (Sigma-Aldrich). Amplicons were sequencedcommercially (by LGC Genomics Ltc, Berlin, Germany)from both directions and readings were assembled intocontigs with the Pregap4 and Gap programs of the Stadenpackage (Staden et al. 2000). All sequences were depositedin GenBank (SUPPLEMENTARY TABLE I.)
Alignments and phylogenetic analyses.—LSU and b-tubulinsequences were aligned with Clustal W (Thompson et al.2002), ITS was aligned by PRANK (Probabilistic alignmentkit, Loytyona et al. 2008). The latter is a model-basedalignment approach that distinguishes insertions fromdeletions and reconstructs the multiple alignment with atwo-step progressive algorithm that uses an accurate MLphylogeny for the assembly of the second multiplealignment. For the distinction between insertions anddeletions we used the + F option, which producesalignments with more gaps but has been shown to be moreaccurate in finding positional homologies than all previ-ously available alignment approaches (Loytyona et al.2008). Overly divergent intron regions of the b-tubulingene and leading and trailing gaps of all alignments weretrimmed before analysis. Indels in the ITS region wererecoded by means of the simple indel-coding algorithm(Simmons and Ochoterena 2001) as implemented inFastGap 1.2.1 (Borchsenius 2007). The simple indel codingalgorithm treats multiresidue indels as a single evolutionaryevent (Simmons and Ochoterena 2001) and has beenshown to compete well with the performance of thecomplex indel-coding algorithm (Simmons et al. 2007).Combinability of the genes was assessed by inspecting MLbootstrap values obtained for single-gene alignments.Mutually exclusive, strongly supported (. 70%) relation-ships were interpreted as significant conflict between thealignments. Models of nucleotide substitution were selectedfor each alignment with AICc in jModelTest (Posada 2008).During model selection we disregarded the invariant sites(I) option because this accounts for the same phenomenonas the gamma-distributed rate heterogeneity model andsimulation studies revealed a risk of parameter nonidenti-fiablility when the two are applied simultaneously (Rannala
NAGY ET AL.: COPRINELLUS PHYLOGENY 255

2002, Stamatakis 2006). Alignments and trees were depos-ited in TreeBASE (study 11515).
Maximum likelihood bootstrapping was performed in theMPI-enabled parallel version of RaxML 7.5.1 (Stamatakis2006). The dataset was partitioned as above, but the indelmatrix was omitted and the GTR + G model was invoked foreach partition separately. The gamma distribution wasseparated into four rate categories. One thousand nonpara-metric bootstrap replicates were run. Each bootstrap replicatewas started from a maximum parsimony tree built in RaxML.
We inferred phylogenies under maximum likelihood andtwo Bayesian methods. For Bayesian estimation we usedMrBayes 3.1.2 (Altekar et al. 2004) and BayesPhylogenies 2.0(Pagel and Meade 2007). MrBayes was launched with defaultpriors and these settings: chain length 30 000 000 genera-tions, sampling frequency 100, two replicates, with threeheated and one cold chains per replicate; while in Baye-sPhylogenies no metropolis coupling was applied, thesampling frequency was set to 1000, replicates to three andchain length was 15 000 000. The concatenated alignmentwas partitioned into ITS1, 5.8S, ITS2, b-tubulin, LSU andindel matrices, and the parameters of the evolutionary modelwere estimated separately for each partition. In BayesPhylo-genies we invoked a mixture model with three patterns foreach alignment site in each partition (Pagel et al. 2004). Theburn-in was established by inspecting the convergence oflikelihood values in Tracer (Rambaut 2004) and tree-splitposterior probabilities in AWTY (Wilgenbusch et al. 2004).Post burn-in trees were used to compute a 50% majority ruleconsensus tree in the SUMTREES script of the Dendropypackage (Sukumaran and Holder 2010). To examine theoptimal phylogenetic placement of C. verrucispermus and C.deliquescens we performed a maximum likelihood constraintanalysis as described in Nagy et al. (2010).
Morphological studies.—Characters suitable for differentia-tion among species were collected from literature searchesand microscopic examination of dried herbarium speci-mens. Microscopic examination was performed on a ZeissAxiolab light microscope after mounting the dried materi-als in 10% NH4OH and Congo red. Dimensions and shapeof caulo-, cheilo-, pleuro- and pilocystidia, veil remnants,basidia, basidiospores, the angle and presence of a germpore were noted. Cheilo- and pleurocystidia were observedseparately by cutting the gill edge from the rest of the gill toavoid blending of the two types of cystidia. Measurements ofbasidia and basidiospores included sterigmata and thehilum respectively. Q values refer to the length divided bythe width of the spores in frontal view. Macromorphologicalcharacters were described from fresh collections or weretaken from herbarium notes. Line drawings and measure-ments were carried out at 4003 or 10003 magnification.Line drawings were converted to vector graphics format inCorelDraw (Corel Co., Canada) after optimizing filter andsmoothing values while tracing bitmaps.
RESULTS
Altogether 160 new sequences were produced for thisstudy; accession numbers are provided (SUPPLEMEN-
TARY TABLE I). The concatenated dataset contained154 specimens of 71 species. We did not obtainspecimens for Coprinellus dilectus, C. minutisporusand C. allovelus, while PCR amplification or sequenc-ing failed for Coprinellus singularis, C. pyrrhanthes, C.heptemerus and C. ephemerus. The latter turned out tobe much rarer than previously assumed, with mostspecimens labeled C. ephemerus actually representingC. congregatus. Out of 154 specimens ITS, b-tubulinand LSU were available respectively for 153, 134 and79. Sequencing of ITS fragments failed for C.simulans, so this species was represented only by theb-tubulin and LSU genes. Both the ITS and the b-tubulin sequences of C. parvulus had an excessivenumber of ambiguities and potentially misidentifiedbases, so we excluded this taxon from the analyses.
After the exclusion of nonoverlapping leading andtrailing regions as well as introns, the alignmentsspanned 1022 characters for ITS, 369 for b-tubulinand 1296 for LSU regions, producing a concatenatedalignment of 2689 nucleic acid sites. This wassupplemented with 238 binary characters obtainedby recoding the indels in the ITS alignment. Based onthe AICc distances, the best fit model was found to bethe GTR + G for all partitions except the 5.8S region,for which the JC + G was proposed. The indelpartition was modeled by a two-state Markov modelimplemented for restriction sites in MrBayes. In bothBayesian and ML analyses the ITS region waspartitioned further into ITS1 (1–450), 5.8S (451–631) and ITS2 (632–1022), following the results ofNagy et al. (2011). The BayesPhylogenies analysis wasstopped after 15 000 000 generations because AWTYanalyses indicated sufficient topological convergence.The burn-in was established respectively as 25 000 000and 10 000 000 generations for the MrBayes andBayesPhylogenies runs. Thus posterior probabilitieswere calculated for 2 3 50 000 and 3 3 5000 treesfrom the output of the two programs. Unexpectedplacements of C. verrucispermus and C. silvaticus(basally in the Domestici clade) were examined byconstraint analyses. When these taxa were constrainedin the Micacei clade, the likelihood values did notdrop significantly, yielding the following P valueintervals for the approximately unbiased Kishino-Hasegawa and Shimodaira-Hasegawa tests: 0.18–0.29,0.13–0.25 and 0.65–0.67. This suggests a plausiblephylogenetic position for C. verrucispermus and C.silvaticus in the Micacei clade.
The trees inferred in the Bayesian MCMC and MLanalyses were largely congruent with regard to deepdivergences; the species were split into three largeclades hereafter referred to as the Core Setulosi clade(BPP: 0.56 MLBS: 90% MBPP: 0.65), eurysporoidclade (BPP: 1.00 MLBS: 100% MBPP: 1.00) and the
256 MYCOLOGIA

Micacei/Domestici (BPP: 0.50 MLBS: 56% MBPP:0.86) clade (FIG. 1). The latter was split into twosubclades, one containing species of the morpholog-ically discerned section Domestici plus C. pusillulus(5 C. heptemerus f. parvisporus), C. curtus and Copri-nellus sp. 7 (SZMC-NL-0150), hereafter referred to asDomestici clade (BPP: 0.50 MLBS: 2MBPP: 0.76) andthe other corresponding to section Micacei (Micaceiclade, BPP: 1.00 MLBS: 89% MBPP: 0.94), includingthe setulose taxa C. disseminatus, C. aureogranulatus,C. silvaticus, C. verrucispermus and C. xylophilus. Theposition of Coprinellus sp. 8 (SZMC-NL-2933) hasproved difficult to resolve due to a highly divergentITS sequence. Based on morphological features (rich,thick-walled, colored veil), it would fit well in sectionDomestici but is not conspecific with any of thedescribed species therein. All analyses place it in atransitional position between the Micacei/Domesticiand the setulose clades. Similarly C. simulans couldnot be assigned a certain position in any of theanalyses.
TAXONOMY
Coprinellus cinereopallidus L. Nagy, Hazi, Papp &Vagvolgyi, sp. nov. FIGS. 2, 3
MycoBank MB561851Pileus 3–6 (7) 3 3–4 mm in iuventute atque clausus,
usque ad 10–20 mm latus si plane expansus, ellipsoideus velsubglobosus, deinde convexus, applanatus in maturitate,leviter deliquescens, pellucide striatus usque ad centrum siudus, radialiter canaliculatus, plicatus in expansis specimi-nibus, omnino tenuissimis micaceis pilis obtectus, sine velo,pallidissime ochraceus vel pallide griseolus, in maturitatepaene albidus vel pallide griseolo-ochraceus. Lamellaeliberae, confertae, ventricosae, usque ad 1–1.5 mm latae,in iuventute albidae, deinde griseolae, demum nigrescentes,acie minute fimbriata albida. Stipes 0.5–1 mm crassus, 30–80 mm longus, elatus, fistulosus, fragilis, cylindricus velinconspicue inflatus basim versus, minutissime omninoalbis pilis pruinosus, albidus vel pallide ochraceus inmaturitate. Caro inconspicua, fragilis, sine odore vel saporeconspicuo.
Sporae 8.9–14.2 3 5.8–7 mm, plerumque 12.6 3 6.6 mm, Q5 1.76–2.18, in fronte ellipsoideae apice fusiformi velconico vel cylindrico atque basi obtusa, a latere fusiformesvel cylindricae, haud lentiformes, mediocriter rubrobrun-neae, poro germinativo valde excentrico, 2–2.3 mm latoatque parvo hilo praeditae. Basidia tetrasporigera, biformia,clavata vel in medio constricta, 23–30 3 8–10 mm. Pleur-ocystidia absunt. Cheilocystidia lageniformia, 35–40 3 10–12 mm. Pileocystidia numerosissima, delicata, lagoeniformiacollo cylindrico vel saepe sinuoso atque basi parva, elata,apice paene eadem latitudine, rotundato-inflato, interdumcapitato, 62–93 3 9–18 mm, saepe luteola refractivasubstantia praedito; intermixtis nonnullis robustis firmior-ibus cystidiis (110 3 15 mm), plerumque versus pilei
centrum. Caulocystidia numerosa, lagoeniformia, collocylindrico, plerumque sinuoso atque apice inflato-clavato,cuspide delicate rotundata, 55–88 3 17–29 mm. Sclerocys-tidia absunt. Velum abest. Fibulae absunt.
Pileus 4–7 3 3–5 mm when closed, ellipsoid tosubcylindrical, expanding to convex-plano-convexthen convex, 10–30 mm wide when fully expanded,surface translucently striate up to center when young,later radially sulcate-grooved, densely covered withshort, hardly noticeable hairs, pale grayish brown tobeige, darker in the center, grayish toward marginwhen moist, pale ochraceous brown to almost whitishwhen dry, weakly deliquescent; lamellae distant, freeto narrowly adnate, verntricose, up to 1–1.5 mmbroad, white when young becoming grayish thenblackish upon spore maturation, with whitish, fim-briate edge; stipe 0.8–1.5 3 20–50 mm, fragile,fistulose, equal or slightly broadening downward,pure white, densely covered with glimmering, shorthairs; context insignificant, brittle without peculiarodor or flavor.
Basidiospores 9.2–13.3 3 (5)5.5–7 mm, on average12.1 3 6.5 mm, Q 5 1.76–2.18, in frontal view ellipsoidto subcylindrical with obtuse base and often acute orsubpapillate apex, in lateral view ellipsoid to sub-amygdaloid, not lentiform, with small hilum and a(strongly) eccentric, ca. 1.8–2.1 mm wide germ pore;Basidia four-spored, clavate or elongate with a medianconstriction, bimorphic, 21–33 3 8–10 mm; Cheilo-cystidia lageniform with a tapering neck and capitateapex, sparse, 35–40 3 10–12 mm; pleurocystidiaabsent; pilocystidia sparse, lageniform with a tapering,rarely cylindrical neck and obtuse, rounded orsubcapitate, often definitely capitate apex, 67–88 3
12–22 mm; caulocystidia seldom similar to pilocystidiabut generally with a larger rounded base, lageniformwith cylindrical or tapering neck and rounded apex57–113 3 15–27.5 mm; cylindrical to diverticulate veilelements present on pileus surface. Clamp connec-tions present.
HOLOTYPE. HUNGARY, Szeged, Szeged BotanicalGarden, in mown lawn on clay-like soil, 25 May 2007,L. Nagy, SZMC-NL-0177, (HOLOTYPE, BP).
Additional specimens examined. HUNGARY, Lakitelek,Toserdo, Bromo sterili-Robinietum, 14 Oct 2006., L. Nagy,SZMC-NL-1145; THE NETHERLANDS, Alphen an derRijn, Rioolwaterzwivering, in leaf litter, C.B. Ulje, Ulje 1163.
Habitat and distribution. Growing solitary on mossy,clay-like soil or leaf litter. Known from Hungary andthe Netherlands.
Remarks. On account of the diverticulate veilelements on the pileus, this species belongs to thegroup around C. hiascens, which also is supported bythe phylogeny. Within this group it is closest to C.velatopruinatus both morphologically and phylogenet-ically. The molecular phylogeny however clearly
NAGY ET AL.: COPRINELLUS PHYLOGENY 257

FIG. 1. Bayesian 50% majority rule consensus phylogram for 154 specimens of Coprinellus with the Psathyrella candolleanaclade as outgroup. The partitioned BMCMC analysis was performed on the combined ITS + b-tubulin + nrLSU alignment andthe tree and posterior probabilities were computed from 100 000 and 15 000 trees for the MrBayes MCMC andBayesPhylogenies mixture-model MCMC analyses respectively. Bootstrap values were estimated in 1000 replicates.
258 MYCOLOGIA

separates it from the three sequenced collections of C.velatopruinatus (BPP: 1.00 MLBS: 100% MBPP: 1.00).Although C. cinereopallidus resembles C. velatopruina-tus in many respects, it differs in having distinctlycapitate pileocystidia and lageniform cheilocystidia.The latter character is shared with C. heterothrix, inwhich the apices of the cheilocystidia are not capitateand both ends of the spores are obtuse. The grayishbrown cap of C. cinereopallidus, which has no or faintochraceous hues, also distinguishes it from C.velatopruinatus and C. heterothrix, which has a mainly
ochraceous cinnamon-brown cap, similarly to mostother species of the C. hiascens clade. Another speciesin this group is C. minutisporus, distinguished by itssmall, 7–8.5 mm long, basidiospores, a central germpore and diminutive basidiocarps.
Coprinellus deminutus (Enderle) L. Nagy, Hazi,Vagvolgyi and Papp, comb nov.
MycoBank MB561846Basionym. Coprinus deminutus Enderle Zeitschrift fur
Mykologie 70:158. 2004.
FIG. 2. Fruiting bodies of the species described in this paper. A. C. cinereopallidus dry specimens (holotype). B. C.cinereopallidus, fresh, wet specimen. C. C. uljei (holotype). D. C. sabulicola (holotype), inset picture shows young fruiting body.E. C. fuscocystidiatus (holotype).
NAGY ET AL.: COPRINELLUS PHYLOGENY 259

FIG. 3. Coprinellus cinereopallidus. A. Spores. B. Basidia. C. Pileocystidia. D. Caulocystidia. E. Veil elements. F.Cheilocystidia. Bars: A 5 5 mm; B, E, F 5 10 mm; C, D 5 20 mm.
260 MYCOLOGIA

Coprinellus fuscocystidiatus L. Nagy, Hazi, Papp &Vagvolgyi, sp. nov. FIGS. 2, 4
MycoBank MB561848Pileus 5–12 3 4–7 mm in iuventute, ellipsoideus vel plus
minusve cylindraceus, deinde convexus vel plano-convexusin maturitate, usque ad 25 mm latus, valde deliquescens; iniuventute pellucide striatus et radialiter canaliculatususque ad centrum, deinde radiatim sulcatus, sine manifes-tis veli reliquis, minutissime, vix manifeste, omninopruinosus; ochraceus vel pallide brunneorubeolus si udus,griseobrunneus vel pallide ochraceus si siccus, centroobscuriore. Lamellae liberae, confertae, ventricosae, usquead 2 mm latae, albae in iuventute, deinde griseolae velnigrescentes, acie minute fimbriata pallidiore. Stipes 1–2 mm crassus, 40–90 mm longus, filiformis, cylindricus,fragilis, fistulosus, tenuibus hyalinis pilis dense obtectusper cunctam longitudinem, albidus, vitreus vel pallideochraceus. Caro tenuis, albida, sine odore vel saporeconspicuo.
Sporae 11.7–15 3 6.4–7.5 3 5,8–6.5 mm, plerumque 13.83 7.1 3 6.3 mm, Q 5 1.82–2.02, ellipsoideae in fronte apiceconico et basi obtusa praeditae, a latere cylindricae velnaviculares, mediocriter rubrobrunneae, poro germinativovalde excentrico, circiter 2–2.2 mm lato, et parvo hiloinstructae. Basidia tetrasporigera, biformia, clavata vel inmedio constricta, 22–30 3 8–10 mm. Pleurocystidia absunt.Cheilocystidia (i) globosa, vesiculosa, dense collecta, 15–25 mm diam, et (ii) satis numerosa, lageniformia, collo valdeattenuato et apice obtuso, 20–43 3 10–15 mm. Pileocystidiamoderate numerosa, lagoeniformia, collo attenuato et apiceobtuso, basi pallide castanea sub lente, laevia, haudincrustata, 80–145 3 16–22 mm. Sclerocystidia absentia.Caulocystidia numerosa, lagoeniformia, basi ampla velcylindrica, collo attenuato, hyalina, basi haud brunnea,57–109 3 17–28 mm. Velum abest. Fibulae absunt.
Pileus 5–12 3 4–7 mm when young, ellipsoid to+/2 cylindrical, expanding to convex/plano-convexwhen old, up to 25 mm across, strongly deliquescent;surface when young translucently striate and radiallygrooved up to center, later radially sulcate, withoutvisible veil remnants and with fine, faintly pruinose;ochraceous pale reddish brown when moist, beige topale ochraceous when dry, darker in the center.Lamellae free, crowded, ventricose up to 2 mm broad,white when young, then grayish blackish, with finelyfrimbriate, paler edge. Stipe 1–2 3 40–90 mm,filiform, cylindrical, fragile, fistulose, densely coveredwith fine hyaline hairs over its length, whitish, vitreousto pale ochraceous. Context without characteristicodor or flavor, thin, whitish.
Basidiospores 11.7–15 3 6.4–7.5 3 5.8–6.5 mm, onaverage 13.8 3 7.1 3 6.3 mm, Q 5 1.82–2.02, ellipsoidin frontal view with +/2 conical apex and obtusebase, in lateral view cylindrical or navicular, mediumred-brown, with strongly eccentric, ca. 2–2.2 mm widegerm pore and small hilum. Basidia bimorphic, four-spored, clavate or with median constriction, 22–30 3
8–10 mm; pleurocystidia absent; cheilocystidia of twotypes: (i) globose, vesiculose densely packed cells, 15–25 mm diam and (ii) abundant lageniform cheilocys-tidia with a strongly tapering neck and obtuse apex,20–43 3 10–15 mm; pileocystidia moderately numer-ous, lageniform with tapering neck and obtuse apex,base pale brown chestnut under the microscope,color is similar to that of pileipellis cells, smooth, notincrusted, 80–145 3 16–22 mm; sclerocystidia absent;caulocystidia numerous, lageniform with a broad baseor cylindrical with tapering neck, hyaline, base notbrown, 57–109 3 17–28 mm; veil absent; clampconnections absent.
HOLOTYPE. NORWAY, Steinkjer, Skrattasen, ondung mixed with straw, 5 Sep 2009, L. Nagy, NL-2720(HOLOTYPE, BP);
Additional specimens examined. SWEDEN, Oland, NedVasterstad, on cow dung. 10 Sep 2009, L. Nagy & T.Knutsson, NL-3622, SZMC.
Habitat and distribution. Known from Sweden andNorway. Both collections were growing on dung,although the holotype was found on a mixture ofhorse dung and straw.
Remarks. This species can be recognized by thestriking brown base of pileocystidia in combinationwith both globose and lageniform cheilocystidia. Noother species in Coprinellus is known to have coloredpileocystidia. Both of the collections examined by uspossess both types of cheilocystidia, so we included itas a diagnostic character, even though it is not alwaystaxonomically reliable. Macroscopically this species issimilar to C. callinus, although the pileus is somewhatdarker and the neck of its pileocystidia is tapering.Phylogenetically its closest relative is C. amphithallus, atwo-spored species with similar colors but lackingglobose cheilocystidia.
C. pallidus L. Nagy, Hazi, Papp & Vagvolgyi, sp. nov.FIG. 5
MycoBank MB561849Pileus 3–6 (7) 3 3–4 mm in iuventute atque clausus,
usque ad 10–20 mm latus si plane expansus, ellipsoideus velsubglobosus, deinde convexus, applanatus in maturitate,leviter deliquescens, pellucide striatus usque ad centrum siudus, radialiter canaliculatus, plicatus in expansis specimi-nibus, omnino tenuissimis micaceis pilis obtectus, sine velo,pallidissime ochraceus vel pallide griseolus, in maturitatepaene albidus vel pallide griseolo-ochraceus. Lamellaeliberae, confertae, ventricosae, usque ad 1–1.5 mm latae,in iuventute albidae, deinde griseolae, demum nigrescentes,acie minute fimbriata albida. Stipes 0.5–1 mm crassus, 30–80 mm longus, elatus, fistulosus, fragilis, cylindricus velinconspicue inflatus basim versus, minutissime omninoalbis pilis pruinosus, albidus vel pallide ochraceus inmaturitate. Caro inconspicua, fragilis, sine odore vel saporeconspicuo.
NAGY ET AL.: COPRINELLUS PHYLOGENY 261

FIG. 4. Coprinellus fuscocystidiatus. A. Spores. B. Basidia. C. Pileocystidia. D. Caulocystidia. E. Lageniform cheilocystidia. F.Globose cheilocystidia. Bars: A 5 5 mm; B, F 5 10 mm; C–E 5 20 mm.
262 MYCOLOGIA

FIG. 5. Coprinellus pallidus. A. Spores. B. Basidia. C. Pileocystidia. D. Caulocystidia. Bars: A 5 5 mm, B 5 10 mm, C, D 5
20 mm.
NAGY ET AL.: COPRINELLUS PHYLOGENY 263

Sporae 8.9–14.2 3 5.8–7 mm, plerumque 12.6 3 6.6 mm, Q5 1.75–2.03, in fronte ellipsoideae apice fusiformi velconico vel cylindrico atque basi obtusa, a latere fusiformesvel cylindricae, haud lentiformes, mediocriter rubrobrun-neae, poro germinativo valde excentrico, 2–2.3 mm latoatque parvo hilo praeditae. Basidia tetrasporigera, biformia,clavata vel in medio constricta, 23–30 3 8–10 mm. Pleur-ocystidia absunt. Cheilocystidia haud notata, verisimiliterglobosa-clavata. Pileocystidia numerosissima, delicata, la-goeniformia collo cylindrico vel saepe sinuoso atque basiparva, elata, apice paene eadem latitudine, rotundato-inflato, interdum capitato, 62–93 3 9–18 mm, saepe luteolarefractiva substantia praedito; intermixtis nonnullis robustisfirmioribus cystidiis (110 3 15 mm), plerumque versus pileicentrum. Caulocystidia numerosa, lagoeniformia, collocylindrico, plerumque sinuoso atque apice inflato-clavato,cuspide delicate rotundata, 55–88 3 17–29 mm. Sclerocys-tidia absunt. Velum abest. Fibulae absunt.
Pileus 3–6(7) 3 3–4 mm when young and closed,up to 10–20 mm broad when fully expanded,ellipsoid-subglobose, then convex, flattened whenmature, weakly deliquescent, translucently striate upto center when moist, radially grooved, plicate whenexpanded, the surface is covered with fine glimmer-ing hairs, veil not seen, pale ochraceous to palegrayish, fading to almost whitish, pale grayishochraceous when mature; lamellae free, crowded,ventricose, up to 1–1.5 mm broad, whitish whenyoung, then grayish, finally blackish, with minutelyfimbriate whitish edge; stipe 0.5–1 3 30–80 mm,slender, fistulose, fragile, cylindrical or insignificantlybroadening toward base, surface covered with finewhitish pruinose hairs, whitish to pale ochraceouswhen mature. Context insignificant, brittle, with nonoticeable odor or flavor.
Basidiospores 8.9–14.2 3 5.8–7 3 6.2–6.7 mm, onaverage 12.6 3 6.6 3 6.5 mm, Q 5 1.75–2.03, in frontalview ellipsoid with fusiform, conical or cylindricalapex and obtuse base, in lateral view fusiform tocylindrical, slightly lentiform, medium red-brown,with 2–2.3 mm wide, strongly eccentric germ poreand small a hilum; basidia four-spored, bimorphic,clavate or with median constriction, 23–30 3 8–10 mm;pleurocystidia absent; cheilocystidia not seen, proba-bly globose-clavate; pileocystidia abundant, delicate,lageniform with cylindrical, or often sinuous, neckand a small slender base and almost equally broadrounded-enlarged, sometimes capitate apex, 62–93 3
9–18 mm, apex often filled with yellowish refractivematerial; interspersed with the before-mentionedtype, a small number of robust, firmer cystidia (1103 15 mm) mainly toward the center of the pileus;caulocystidia abundant, lageniform, with cylindrical,mostly sinuous neck and enlarged-clavate apex withdelicate rounded tip, 55–88 3 17–29 mm; sclerocysti-dia absent; veil absent; clamp connections absent.
HOLOTYPE. HUNGARY, Kecskemet, Kistemeto:on wood chips along a path, 5 Aug 2006, L. Nagy &Zs. Gorliczai, NL-1556 (HOLOTYPE, BP);
Additional specimens examined. HUNGARY, Szeged, Ne-pliget, in mown lawn, 14 May 2008, L. Nagy, NL-4218,SZMC; Szeged Botanical Garden, among grass on clay-likesoil, 9 May 2008, L. Nagy, NL-0625, SZMC.
Habitat and distribution. So far known from humidplaces with mossy, clay-like soil from three localities inHungary.
Remarks. The pale colors and the slender, capitatepileocystidia with frequently sinuous neck are distinc-tive for this species. Capitate or subcapitate pileocys-tidia also are found in other species of Coprinellus,such as C. curtus, C. plagioporus, C. marculentus, C.heterothrix, C. velatopruinatus, C. subdisseminatus andC. cinereopallidus. Of these C. curtus differs in havingstrongly flattened spores, more vivid colors and a richcoverage of velar spherocysts on the pileus; the fruitingbodies of C. plagioporus and C. marculentus are darkbrown and pileocystidia are broader. C. heterothrixpossesses diverticulate veil elements on the pileussurface unlike C. pallidus, which can vanish quicklyfrom the maturing fruiting bodies. In such cases thefusiform spores and the narrow, slender pileocystidiaare indicative of C. pallidus.
Coprinellus ramosocystidiatus (Bender) L. Nagy,Hazi, Vagvolgyi and Papp, comb nov.
MycoBank MB561847Basionym. Coprinus ramosocystidiatus Bender, Zeitschrift
fur Mykoogie 56:35. 1990.
Coprinellus sabulicola L. Nagy, Hazi, Papp & Vagvol-gyi, sp. nov. FIGS. 2, 6
MycoBank MB561850Pileus primo ellipsoideus vel etiam conicus, usque ad 3 3
2 mm in inexpanso tempore, deinde campanulatus cumobtuso centro, ad postremum applanatus vel proprierevolutus, usque ad 15 mm latus, etiam in iuvenili temporeusque ad centrum radialiter sulcatus sed perluciditate nonstriatus, in senectute fissuratus, non deliquescens. Cuticulaopaca, primo levis atque pallide ochraceofusca griseis cumvarietatibus, sed etiam fuscorutilans, deinde pallidior floslactis-ochracea griseis cum varietatibus atque austerioripapilla. Lamellae remotae, ventricosae, usque ad 1 mmlatae, ex albo nigricantes, cum albidiore margine. Stipes 20–45 3 0.5–0.9 mm, filiformis, haud bulbosus, fragilis, densepruinosus, albovitreus, ad basim pallide ochraceus. Odoratque sapor nulli.
Sporae 15–21.8 3 10–13 mm, fere 17.3 3 10.9 mm,ellipsoideae-late ellipsoideae, etiam subovoideae, a laterelineamentis angulatae, ad apicem obtusissimae, hilo latopraeditae, haud vel vix lentiformes, nigriculo- vel purpureo-fuscae, cum externo, 2.8–3 mm lato, germinabili poro.Basidia clavata, unimorpha, bispora, fere 32.5 3 11.3 mm.
264 MYCOLOGIA

Cheilocystidia globosa usque ad vesiculosa vel late ellipsoi-dea, brevistipitata vel sessilia, 17–32 3 12.5–27 mm. Pleur-ocystidia absentia. Pileocystidia biformia: (i) lageniformiausque ad fusiformia, valde contracta atque capitata,plerumque crustata, 22–63 3 8–19 mm; (ii) mucronata, adaliquando crustatam basim ellipsoidea, ad apicem obtusaatque non capitata, 30–33 3 12–14 mm. Velum haudagnitum sed nonnullae crustatae hyphae in pileipelleiacentes. Caulocystidia lageniformia usque ad fusiformia,contracta atque late capitata, 41–60 3 10–20 mm. Pileipellisanguste clavatis, plerumque aliquantum angulatis cellulisinstructa. Subpellis diverticulatis atque fibulatis hyphispraedita.
Pileus ellipsoid to conical when young, up to 3 3
2 mm when closed, later campanulate with more orless obtuse center, becoming applanate or, more
typically concave when mature, not or weaklydeliquescent. Margin strongly radially sulcate up tocenter even when young, becoming strongly fissuredwhen old, in young stages not or slightly translu-cently striate. Surface mat, smooth when young, veilor setules not visible to the naked eye but somewhatpruinose when very young. In young stages paleocher-brown with grayish hue to warm reddishbrown (like typical young Parasola misera), becom-ing paler ochraceous-beige with grayish hue due tomaturing spores, typically with a darker button incenter when mature. Rarely the mature pileus is justpale gray to almost whitish (mostly in dry environ-ment). Lamellae free, distant (almost like P. misera),ventricose up to 1 mm broad, white when young,
FIG. 6. Coprinellus sabulicola. A. Spores. B. Basidia. C. Pileocystidia. D. Caulocystidia. E. Cheilocystidia. Bars: A, B, E 5
10 mm; C, D 5 20 mm.
NAGY ET AL.: COPRINELLUS PHYLOGENY 265

becoming grayish-blackish with age, but the gill edgeremains white for a long time, strongly contrastingwith the sides of the gills. Stipe 0.5–0.9 3 20–45 mmlong, slender, fistulose, fragile, equal without basalbulb, surface densely pruinose from short caulocys-tidia, whitish, vitreous to pale ochraceous towardbase. Context thin, insignifcant, odor and flavorindistinct.
Basidiospores 15–21.8 3 10–13 mm, on average 17.33 10.9 mm, Q 5 1.43–1.74, ellipsoid-broadly ellipsoidto almost ovoid, with strongly obtuse apex and largehilum, not or slightly lentiform, dark blackish reddishbrown, opaque, germ pore large, strongly eccentric,ca. 2.8–3 mm wide in lateral view often somewhatmore ellipsoid with an angular outline. Basidiaclavate, not bimorphic, two-spored, ca. 32.5–11.3 mm.Cheilocystidia abundant, globose to vesiculose orbroadly ellipsoid, typically with short but broadpedicel or no pedicel at all, and then the cystidiumends in a broad flat base, 17–32 3 12.5–27 mm.Pleurocystida absent. Pseudoparaphyses present. Pi-leocystidia of two types; (i) normal lageniformcystidia with tapering neck and capitate apex, thesecystidia are small compared to other species ofSetulosi, they have an inconspicuous neck and apex(somewhat similar to that of C. heptemerus), basalpart sometimes inconspicuous, making the cystidiumjust fusiform, sometimes a more pronounced basalpart is observable, incrusted with crystal not dissolv-ing in NH4OH in most cases, 22–63 3 8–19 mm; (ii)typically mucronate with an ellipsoid basal partending in a short neck with obtuse, noncapitateapex, basal part sometimes incrusted, 30–33 3 12–14 mm. Veil not seen but several hyphae lying on thepileipellis are incrusted. Caulocystidia abundant,lageniform to fusiform with a tapering neck andgenerally large capitulum, incrustation scarce, 41–603 10–20 mm. Pileipellis cells unusually narrowlyclavate but mostly somewhat angular. Hyphae under-neath pileipellis layer rather narrow (2.5–4 mm wide),richly diverticulate; veil absent; clamp connectionspresent.
HOLOTYPE. HUNGARY, Orgovanyi Osborokas: onsand, attached to bark of Populus nigra 24 Oct 2009,L. Nagy and M. Jeppson, NL-0872 (HOLOTYPE, BP).
Additional specimens examined. HUNGARY, Alfold, Asot-thalom: Festucetum vaginatae stipaetosum, on herbs 2 Nov2008, L. Nagy, NL-4140, SZMC; ibid: Festucetum vaginataestipaetosum, on dead stems of 9 May 2007, L. Nagy & Zs.Gorliczai, NL-1147, SZMC; ibid: roadside, on sedges, 20 Jun2008 L. Nagy, NL-2907, SZMC; ibid: Bromo sterili-Robinie-tum, on sandy roadside, 20 Jun 2008, L. Nagy, NL-2906,SZMC; ibid: on vegetable debris, buried in sand, 10 Jun2008, L. Nagy, NL-1763, SZMC; ibid: Festucetum vaginatae,on dead grasses and stems of Euphorbia cyparassias, 20 Aug2005, L. Nagy, NL-1027, SZMC; Nyomasi – forest:
Cynodonti-Festucetum vaginatae, 8 Sep 2007, L. Nagy, NL-057, SZMC 9; ibid: 16 May 2005, L. Nagy, NL-3063, SZMC;Nyarjas: Artemisio-Festucetum, on herbaceous debris, 1 Nov2004, L. Nagy & Zs. Gorliczai, NL-1031, SZMC; Torokfai:among grass on bare sandy soil, 31 Aug 2004, L. Nagy, NL-1560, SZMC; Balazspuszta, Kunsag-major: Cynodonti-Festu-cetum pseudovinae, 6 Jun 2010, L. Nagy, NL-0825, SZMC;Fulophaza: on rabbit dung, 6 Jun 2006, L. Nagy, NL-3536;Nemetker, Lato-hegy: Festucetum vaginatae, 6 Nov 2009, P.Finy, FP/2009.11.06/1.
Habitat and distribution. Grows in sand dunesalways in the vicinity of smaller open, sandy areas,often found on naked sand but is always emergingfrom dead sedges, grasses or dung buried in sand(Euphorbia cyparassias, Artemisia absynthium, Stipaarenaria); the type material was growing on bark ofPopulus nigra exposed on bare sand. Rarely found onpure dung (deer, horse). So far known only fromHungary.
Remarks. This species bears some spectacularfeatures compared with other Coprinellus species;among these are relatively large basidiospores, thelack of a pedicel on the cystidia, a habitat in dry,sandy sites and short, capitate pileocystidia withincrusted base. These characters readily distinguishit from any other member of Coprinellus. Macroscop-ically C. sabulicola has superficial resemblance to C.curtus or C. deminutus, both of which differ inhaving much smaller basidiospores, globose veil andfour-spored basidia. Furthermore, unlike in othercoprinoid species, its basidia are not bi- or trimor-phic. Bi- and trimorphic basidia have been found tobe linked to the presence of pseudoparaphyses,plicate pileus, increased size of cystidia and autodi-gestion across the entire Psathyrellaceae, which hasbeen referred to as coprinoidization (Nagy et al.2011). Being an exception to this rule, three of thesetraits can be found in C. sabulicola, that is plicatepileus, increased cell size and pseudoparaphyses,whereas the basidia are monomorphic. Thus wehypothesize that C. sabulicola represents a transition-al state between deliquescent and nondeliquescentspecies, reflecting the characteristics of ancestralmembers of the genus Coprinellus. In agreementwith this assumption it has a basal, albeit somewhatuncertain phylogenetic position, either as a sistergroup to the eurysporoid clade, or between theDomestici + Micacei clades and the Core Setulosi +eurysporoid clade. Both alternatives received lowsupport in the Bayesian and ML analyses. A thirdline of evidence for the ancestral nature of C.sabulicola may be the short branch it occupies,although this conclusion may be affected by potentialrate variation among lineages and decoupling of therate of morphological and molecular evolution(Pagel 1999, Rabosky et al. 2007).
266 MYCOLOGIA

Coprinellus uljei L. Nagy, Hazi, Papp & Vagvolgyi, sp.nov. FIGS. 2, 7
MycoBank MB561852Pileus 4–10 3 3–7 mm in iuventute, ellipsoideus vel
cylindricus, deinde convexus vel plano-convexus obtusoumbone centrali praeditus, demum applanatus marginerevoluto, valde pellucide striatus usque ad centrum iniuventute, radialiter sulcatus in maturitate, tenuibus mica-ceis pilis obtectus, brunneo-ochraceus in iuventute, brun-neogriseolus in maturitate, deliquescens. Lamellae liberae,confertae, ventricosae, usque ad 1 mm latae, albae iniuventute, deinde griseolae, demum nigrescentes, aciealbida fimbriata. Stipes 30–90 longus, 0.8–1.5 mm crassus,elatus, fragilis, fistulosus, cylindricus vel leviter inflatusbasim versus, dense pruinosus per cunctam longitudinem,albidus vel pallide ochraceus. Caro inconspicua, odore etsapore inconspicuo.
Sporae 11.3–14 3 6.2–8 mm, plerumque 13.1 3 7.1 mm, Q5 1.75–1.98 mm, ellipsoideae, latiores basim versus, haudlentiformes, poro germinativo valde excentrico, 1.7–1.9 mmlato, praeditae. Basidia tetrasporigera, biformia, clavata velin medio constricta, 18–27 3 8–10 mm. Pleurocystidiaabsunt. Cheilocystidia plerumque globosa, vesiculosa, 20–30 mm diam, intermixta pluribus lagoeniformibus cystidiis,45–65 3 17–20 mm. Pileocystidia abundantissima, lagoeni-formia, collo attenuato atque apice obtuso, inflato velclavato, sed haud manifeste capitato, 65–115 3 12–25 mm.Sclerocystidia sparsa vel sparsissima, moderate crassituni-cata, lagoeniformia-fusiformia, circiter 62 3 10 mm. Caulo-cystidia numerosa, lageniformia, collo attenuato, lato, atqueapice obtuso, inflato. Velum abest. Fibulae haud notatae.
Pileus 4–10 3 3–7 mm when young, ellipsoid tosubcylindrical, later convex-plano convex with obtuseumbo in the center, finally applanate with up-rolledmargin, surface strongly translucently striate up tocenter when young, radially sulcate upon expansionof pilei, covered with fine, glimmering hairs, ochra-ceous brown when young, grayish brown when older,deliquescent. Lamellae free, crowded, ventricose, upto 1 mm broad, white when young, later grayish, thenblackish, edge whitish fimbriate. Stipe 30–90 3 0.8–1.5 mm, slender, fragile, fistulose, equal or somewhatbroadening toward base, densely pruinose its entirelength, whitish to pale ochraceous. Context insignif-icant, without particular odor or flavor.
Basidiospores 11.3–14 3 6.2–8 mm, on average 13.13 7.1 mm, Q 5 1.75–1.98, ellipsoid, somewhat broadertoward base, not lentiform, with a strongly eccentric,1.7–1.9 mm wide germ pore; Basidia four-spored,bimorphic, clavate or with median constriction, 18–273 8–10 mm; pleurocystidia absent; cheilocystidiapredominantly globose, vesiculose, 20–30 mm diam,interspersed with several lageniform cystidia, 45–65 3
17–20 mm; Pileocystidia abundant, lageniform with atapering neck and obtuse, enlarged or clavate, but notdefinitely capitate apex, 65–115 3 12–25 mm, Scler-ocystidia sparse to very sparse, moderately thick-
walled, lageniform-fusiform, ca. 62 3 10 mm; caulo-cystidia abundant, lageniform with a tapering, broadneck and obtuse, enlarged apex; veil absent; clampconnections not seen.
HOLOTYPE. SLOVAKIA, Muranska Planina Na-tional Park, Studna, on wood chips in Piceetum, 10Oct 2008, L. Nagy, SZMC-NL-3985 (HOLOTYPE, BP).
Additional specimens examined. SWEDEN, Oland, Tor-slunda Parish, Skogsby/Tveta-hagnet, Corylus-Betula-Quer-cus-Juniperus mosaic vegetation, on soil, 26 Sep 2007, L.Nagy and T. Knutsson, SZMC-NL-0157; Honstorps utmar-ker, Corylus-Betula-Quercus-Juniperus mosaic vegetation, onsoil, 27 Sep 2007, L. Nagy and T. Knutsson, SZMC-NL-2492.
Habitat and distribution. Growing on soil and woodchips in broadleaf and coniferous forests. Solitary or inlarge troops. So far known only from warm, dryhabitats of Oland (Sweden) and Slovakia.
Remarks. The presence of lageniform cheilocystidiain addition to globose ones is somewhat uncertain fortwo out of the three examined collections. Morpho-logically this species is close to C. sclerocystidiosus withwhich it shares the obtuse-enlarged apex of thepileocystidia, eccentric germ-pore and spore size butdiffers in having globose and (optionally) lageniformcheilocystidia, whereas C. sclerocystidiosus has globose-ellipsoid cheilocystidia, more abundant sclerocystidiaand the apices of its pileocystidia are never enlarged.In addition C. sclerocystidiosus is phylogenetically wellseparated from C. uljei. Another similar species is C.plagioporus, which can have pileocystidia with enlargedapex and also possesses similar basidiospores, but has adifferent cap color (dark brown) and exclusivelyglobose cheilocystidia and lacks sclerocystidia.
ARTIFICIAL KEY TO THE SETULOSE (HAIRED) SPECIES OF COPRINELLUS
1 Pileus with a macroscopically visible, either con-tinuous or abundant granulose veil coverage whenyoung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
1 Veil on pileus absent or observable only under themicroscope in the form of discontinuous sphaer-ocysts or hyphae . . . . . . . . . . . . . . . . . . . . . . . . 112 Basidiospores ovoid with conical base, mitri-
form in lateral view often amygdaliform . . . 32 Basidiospores ovoid with obtuse base or
ellipsoid . . . . . . . . . . . . . . . . . . . . . . . . . . . 63 Basidiospores ornamented . . . . . . . . . . . . . . . . . 43 Basidiospores smooth . . . . . . . . . . . . . . . . . . . . . 5
4 Basidia four-spored . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . C. deliquescens (5 C. silvaticus)
4 Basidia two-spored . . . . . . . C. verrucispermus5 Pileus up to 20 mm wide when expanded, fruiting
bodies small, fragile . . . . . . . . . . . C. disseminatus5 Pileus 20–50 mm broad when expanded, fruiting
bodies robust, habit like C. impatiens or C.micaceus . . . . . . . . . . . . . . . . . . . . . . C. xylophilus
NAGY ET AL.: COPRINELLUS PHYLOGENY 267
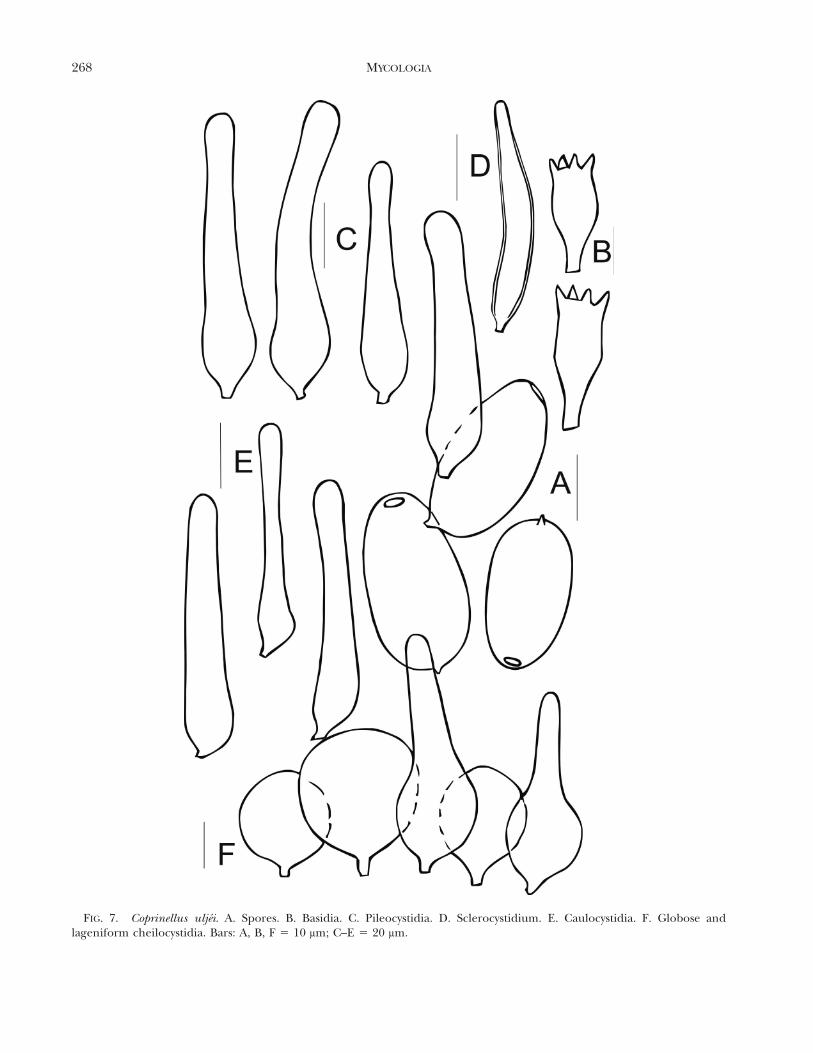
FIG. 7. Coprinellus uljei. A. Spores. B. Basidia. C. Pileocystidia. D. Sclerocystidium. E. Caulocystidia. F. Globose andlageniform cheilocystidia. Bars: A, B, F 5 10 mm; C–E 5 20 mm.
268 MYCOLOGIA

6 Pileocystidia capitate and veil cells with up to5 mm thick, brown walls . . . . . . . . . . . . . . . . 7
6 Pileocystidia not capitate or veil cell walls, 2 mm . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
7 Pileus 4–13 mm diam, veil patches pale ocher, ondung . . . . . . . . . . . . . . . . . . . . . . . . . . . C. curtus
7 Pileus up to 25 mm diam, veil patches blackish, onsoil . . . . . . . . . . . . . . . . . C. sp. 7 (SZMC-NL-0159)8 Apex of pileocystidia obtuse to capitate . . . . 98 Apex of pileocystidia acute . . . . . . . . . . . . 10
9 Pileus bright golden yellow-orange . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . C. aureogranulatus
9 Pileus dull . . . . . . . . . . . . . . . . . . . C. pyrrhanthes10 Basidiospores 12–16 mm long . . . C. heptemerus10 Basidiospores up to 7–11 mm long . . . . . . . .
. . . C. pusillulus (5 C. heptemerus f. parvisporus)11 Basidia two-spored . . . . . . . . . . . . . . . . . . . . . . 1211 Basidia four-spored . . . . . . . . . . . . . . . . . . . . . . 19
12 Basidia monomorphic, basidiospores 15–21 mm long . . . . . . . . . . . . . . . . . C. sabulicola
12 Basidia bi-(tri)morphic, basidiospores smaller. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
13 Germ pore central, pileocystidia with a taperingneck and acute apex, habitat on sandy places . . .
. . . . . . . . . . . . Coprinellus sp. 2 (SZMC-NL-1356)13 Not with this combination of characters . . . . . . 14
14 Cheilocystidia globose to ellipsoid . . . . . . . 1514 Cheilocystidia lageniform . . . . . . . . . . . . . 17
15 Sclerocystidia present . . . . . . . . . . . . . . . . C. sassi15 Sclerocystidia lacking . . . . . . . . . . . . . . . . . . . . 16
16 Velar spherocysts present on pileus . . . . . . .. . . Coprinellus sp. 4 (SZMC-NL-3843 and 2644)
16 Velar spherocysts lacking . . . . . . . . . . . . . .. . . . . . . . . . . . . . C. bisporus and C. bisporiger
17 Sclerocystidia present, av. Q of basidiospores . 2. . . . . . . . . . . . . . . . . . . . . C. pseudoamphithallus
17 Sclerocystidia present, spore length/width ratiosmaller . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1818 Germ pore eccentric, av. Q of basidiospores
1.7–1.8 . . . . . . . . . . . . . . . . . . C. amphithallus18 Germ pore central, av. Q of basidiospores 1.3–
1.5 . . . . . . . . . . . . . . . . . . . . . . . C. singularis19 Diverticulate or hyphal veil elements present on
pileus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2019 Veil, if present composed of sphaerocysts . . . . . 25
20 Base of basidiospores obconical, germ porecentral, cheilocystidia lageniform . . . C. hiascens
20 Base of basidiospores obtuse . . . . . . . . . . . 2121 Basidiospores 7–8.5 mm long . . . . . C. minutisporus21 Basidiospores longer . . . . . . . . . . . . . . . . . . . . . 22
22 Veil on pileus unbranched, hyphal (likesection Lanatuli) C. allovelus (doubtful spe-cies, known only from one collection and onefruiting body)
22 Veil diverticulate . . . . . . . . . . . . . . . . . . . . 2323 Cheilocystidia globose-ellipsoid . . . C. velatopruinatus23 Cheilocystidia lageniform . . . . . . . . . . . . . . . . . 24
24 Cheilocystidia with capitate apex . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . C. cinereopallidus
24 Cheilocystidia with equal or acute apex . . . .. . . . . . . . . . . . . . . . . . . . . . . . . C. heterothrix
25 Basidiospores hexagonal, nodulose or mitriform 2625 Basidiospores ellipsoid, ovoid, but not angular . . . 29
26 Basidiospores mitriform, fruiting body robust,pileus 30–70 mm when expanded . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . C. angulatus26 Basidiospores hexagonal or nodulose . . . . 27
27 Basidiospores hexagonal . . . . . . . . . C. marculentus27 Basidiospores nodulose . . . . . . . . . . . . . . . . . . . 28
28 Basidiospores 6–8 mm long, sclerocystidialacking . . . . . . . . . . . . . . . . . . . . . . C. doverii
28 Basidiospores 9–11 mm long, sclerocystidiapresent . . . . . . . . . . . . . . C. mitrinodulisporus
29 Apex of pileocystidia obtuse or capitate, neckcylindrical or tapering . . . . . . . . . . . . . . . . . . . . 30
29 Apex of pileocystidia +/2 acute, neck tapering . . . 3730 Sclerocystidia present . . . . . . . . . . . . . . . . 3130 Sclerocystidia absent . . . . . . . . . . . . . . . . . 33
31 Cheilocystidia globose-ellipsoid . C. sclerocystidiosus31 Cheilocystidia globose-ellipsoid plus lageniform . . . 32
32 Pileocystidia with enlarged to subcapitateapex . . . . . . . . . . . . . . . . . . . . . . . . . . C. uljei
32 Pileocystidia with acute or equal apex . . . . .. . . . . . . . . . . . . . . . . . . . . . . C. subimpatiens
33 Color dark brown, with or without purplish tints orgrayish . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
33 Color of young fruiting bodies ocher melleous,yellow-brown or cinnamon brown . . . . . . . . . . . 3634 Color of fruiting bodies pale gray, pileocysti-
dia slender, delicate, with capitate apex . . .. . . . . . . . . . . . . . . . . . . . . . . . . . C. pallidus
34 Fruiting bodies dark brown, pileocystidiastout . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
35 Color purplish, apex of pileocystidia equal, orenlarged, not capitate . . . . . . . . . . . . . . . . . . . . . . .. . . . C. subpurpureus (C. fallax, a doubtful species
comes close, should differ in a reddish spore print)35 Color without purplish hues, pileocystidia (sub)-
capitate . . . . . . . . . . . . . . . . . . . . . C. plagioporus36 Cheilocystidia lageniform, basidiospores sub-
cylindrical (Q 5 1.8–1.9) . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . C. subdisseminatus
36 Cheilocystidia globose, basidiospores elliptical(Q , 1.8) . . . . . . . . . . . . . . . . . . . C. simulans
37 Cheilocystidia lageniform or globose plus lageni-form . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
37 Cheilocystidia globose-ellipsoid . . . . . . . . . . . . . 4438 Cheilocystidia of two types, globose and
lageniform . . . . . . . . . . . . . . . . . . . . . . . . 3938 Cheilocystidia lageniform only . . . . . . . . . . 41
39 Base of pileocystidia brown but not incrusted . . .. . . . . . . . . . . . . . . . . . . . . . . . C. fuscocystidiatus
39 Base of pileocystidia not brown . . . . . . . . . . . . . 4040 Pileus bright orange-brown, sclerocystidia ab-
sent . . . . . . . C. dilectus (based on the carefulredescriptionof Schafer)(see alsoC. pellucidus, whichoccasionally can have lageniform cheilocystidia too)
40 Pileus dull, sclerocystidia present . . . . . . . . . . .. . . . . . . . . . Coprinellus sp. 9 (SZMC-NL-1070)
NAGY ET AL.: COPRINELLUS PHYLOGENY 269

41 Pileus up to 2–3 mm when expanded, habitat ondung . . . . . . . . . . . . . . . . . . . . . . . . . C. parvulus
41 Pileus larger, noncoprophilous taxa . . . . . . . . . . 4242 Basidiospores 11–13 mm long, sclerocystidia
absent . . . . Coprinellus sp. 3 (SZMC-NL-3854)42 Basidiospores . 10 mm, sclerocystidia present
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4343 Fruiting body nondeliquescent, germ pore central
. . . . . . . . . . . . . . . . . . . . . . . . . . . . C. impatiens43 Fruiting body deliquescent, germ pore eccen-
tric . . . . . . . . . . Coprinellus sp. 9 (SZMC-NL-1070)44 Pleurocystidia present . . . . . . . . . . . . . . . . 4544 Pleurocystidia absent . . . . . . . . . . . . . . . . . 49
45 Noncoprophilous species . . . . . . . . . . . C. canistri45 Coprophilous taxa . . . . . . . . . . . . . . . . . . . . . . 46
46 Basidiospores 8–11.5 mm long; pileocystidia upto 70 mm long . . . . . . . . . . . . . . . . . . . . . . 47
46 Basidiospores larger (. 12 mm) ; pileocystidialonger . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
47 Caulocystidia unbranched . . . . . . . C. brevisetulosus47 Caulocystidia branched . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . Coprinellus sp. 5 (SZMC-NL-1752)48 Growing on soil in large groups, basidiospores
+/2 amygdaloid in lateral view . . . . . . . . . .. . . . . . . . . Coprinellus sp. 11 (Daams72–68a)
48 Growing on dung, basidiospores sllipsoid inside view . . . . . . . . . . . . . . . . . . . . . . . . . . 49
49 Clamp connections present . . . . . . . . C. ephemerus49 Clamp connections absent . . . . . . . C. congregatus
50 Basidiospores fusiform, with obconical base . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
50 Basidiospores ellipsoid, ovoid to somewhatmitriform . . . . . . . . . . . . . . . . . . . . . . . . . 52
51 Sclerocystidia absent, basidiospores flattened, 11–13 mm long . . . . Coprinellus sp. 3 (SZMC-NL-3854)
51 Base of sclerocystidia encrusted, basidiospores 10–11 mm long . . . Coprinellus sp. 10 (SZMC-NL-3101)52 Sclerocystidia present . . . . . . . . . . . . . . . . 5352 Sclerocystidia absent . . . . . . . . . . . . . . . . . 54
53 Germ pore eccentric, on dung . . . C. heterosetulosus53 Germ pore central, on wood chips . . . . . . . . . . .
. . . . . . . . . . . . Coprinellus sp. 6 (SZMC-NL-2617)54 Basidiospores ovoid to subglobose, Q 5 1.1–
1.3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5554 Basidiospores ellipsoid to somewhat mitri-
form . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5655 Cheilocystidia lageniform . . . . . . . . . C. eurysporus55 Cheilocystidia globose . . . C. aff. eurysporus (SZMC-
NL-1761, 3418, Arnolds 4845, Ulje 1191, Hoijer 95067)56 Average spore length , 10 mm . . . . . . . . . 5756 Average spore length . 10 mm . . . . . . . . . 59
57 Basidiospores submitriform, strongly lentiform 7–9 mm long, on dead grasses . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . Coprinellus sp. 9 (SZMC-NL-1070)
57 Basidiospores ellipsoid, not lentiform, on dung . . . 5858 Basidiospores 8–11 mm long, stipe often
radicate in the substrate . . . . . . . C. radicellus58 Basidiospores 6–9 mm long, stipe not radicate
. . . . . . . . . . . . . . . . . . . . . . . . . . C. pellucidus59 Sclerocystidia present . . . . . . . . . . . . . . C. callinus
59 Sclerocystidia absent . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . C. limicola + Coprinellus sp. 1 (SZMC-
NL-0195 and 1751) (the two species seem morpho-logically identical but are phylogenetically separated)
DISCUSSION
In this study we analyzed the phylogeny and diversityof the genus Coprinellus, one of the most species-richgroups in the Psathyrellaceae. The phylogeny in-ferred here, with an almost complete species-leveldataset (71 out of ca. 80 taxa sampled), does notprovide evidence for the monophyly of morphologi-cally based sections of previous classifications (Ulje2005, Orton and Watling 1979, Kuhner and Romag-nesi 1953, Patrick 1977, Schafer 2010, Keirle et al.2005, Singer 1986) or for the monophyly of specieswith pileo- and caulocystidia. Below we attempt tosynthesize morphological synapomorphies to explainthe phylogenetic distribution of the species.
Higher level relationships within Coprinellus.—Phy-logenetic analyses do not support the monophyly ofCoprinellus species with pileo- and caulocystidia.These, formerly classified in subsection Setulosi ofsection Pseudocoprinus (Ulje and Bas 1991, Ulje 2005,Orton and Watling 1979, Kuhner 1928), are distrib-uted throughout Coprinellus and are found in all fourlarger clades of the genus. The relationship of certainsetulose species to veiled ones accords with formerlypublished phylogenies; the affinities of C. dissemina-tus to veiled Coprinellus species, in particular to C.micaceus, has been noted (Vasutova et al. 2008;Larsson and Orstadius 2008; Walther et al. 2005;Nagy et al. 2011, 2010) and the position of C. curtusand C. heptemerus in the vicinity of C. domesticus andC. radians also has been inferred (Larsson andOrstadius 2008; Nagy et al. 2010, 2011), althoughsatisfying morphological synapomorphies could notbe defined in either case. In fact, although thegeneral appearance of C. disseminatus and C.micaceus is different, the presence of a rich, granuloseveil and the shape of the basidiospores are indicativeof their relationship. In addition to C. disseminatustwo other setulose species, C. aureogranulatus and C.pyrrhanthes, belong to this clade, although the latterconclusion is based only on morphological similarityin the shape of basidiospores and pileocystidia as wellas veil characters. The mitriform spores furthersupport the placement of C. silvaticus and C.verrucispermus in the Micacei clade, which has beenjudged plausible by the constraint analyses as well.Thus we propose that mitriform spores can beregarded as a synapomorphy for this clade and canbe used in the taxonomic circumscription of section
270 MYCOLOGIA

Micacei. Although the basidiospores of C. truncorum,C. aureogranulatus and two incompletely known taxa,C. pallidissimus and C. saccharinus, are not explicitlymitriform, they do have a tendency to produce(sub)mitriform spores, which is evident when sporepopulations of mature specimens are inspected.
How morphological traits support the inclusion ofC. curtus and C. pusillulus (and C. heptemerus) in theDomestici clade is more difficult to establish. Thesetwo species share a rich, granulose veil that forms analmost continuous sheath over the pileus on youngfruiting bodies and strongly flattened basidiosporeswith an eccentric germ pore, the latter being unusualfor the rest of the clade. The only character thatshows an overlap between the setulose and veiledspecies of this clade is the tendency of terminal veilcells to have thick, often brownish walls withcrystalline incrustation. To our best knowledge noother Coprinellus species possess thick-walled, coloredand/or incrusted veil cells and thus an almostcontinuous brownish velar sheath on the young pilei,so these can be considered unique synapomorphiccharacters for this group. Our results further supportthe affinity of C. deminutus to the Domestici clade,which is well supported by morphological features.Another tiny species with morphological features ofthe Domestici clade is C. ramosocystidiatus, a rare oroverlooked taxon with granular veil and ellipsoid-subglobose pileipellis cells, which bear finger-likeprojections. Because both C. deminutus and C.ramosocystidiatus fit well into the Domestici clade wepropose new combinations for them in Coprinellus(see below).
Two clades consist entirely of setulose species. Ofthe four major clades of Coprinellus the Micacei andDomestici clades are easy to separate from the CoreSetulosi and eurysporoid clades on the basis of a richveil coverage on the pileus. The Core Setulosi plus theeurysporoid clades together compose about 90% ofthe species classified in subsection Setulosi (Ulje andBas 2003, Ulje 2005, Schafer 2010, Keirle et al. 2005,Orton and Watling 1979). To our best knowledgesubsection Setulosi has not been subdivided further,which reflects the morphological uniformity of thisgroup. The key characters that have been used forguidance in identification keys are among others thenumber of sterigmata on the basidia, the shape of thepileocystidia (tapering/cylindrical), the shape of thebasidiospores, the presence or absence and shape ofpleuro- and cheilocystidia, however none of thecharacters show a correlation with the phylogeneticstructure of these clades. Thus a morphology-baseddistinction between the Core Setulosi and theeurysporoid clades is not straightforward. The lattercontains robust, soil-inhabiting species without veil,
although these characters do not separate it unambi-guosly from the Core Setulosi clade. Further study isnecessary to establish reliable characters that can beused in identification keys.
The core setulosi clade.—The backbone of the CoreSetulosi clade has proved difficult to resolve, probablybecause of an episode of accelerated speciation in theevolution of this group (Nagy et al. unpubl).Consistent with this the most significant incongru-ence between the ML and the two Bayesian analysesinvolved the branching order at the backbone of theCore Setulosi clade. Although the majority of nodescould not be resolved, several strongly supportedand/or morphologically homogeneous clades wererecovered in the phylogenetic analyses. These, here-after named C. bisporus, C. callinus, C. hiascens andC. pellucidus clades, can be unambiguously circum-scribed morphologically, as presented below.
C. bisporus clade. Although this clade received lowsupport values (BPP: 0.51, MLBS: 2, MBPP: 0.53), it isworth mentioning because all taxa are characterized bytwo-spored basidia except for C. canistri, for which theinclusion of more materials might reveal a moreplausible position in other parts of the tree. Fourmorphologically established species, C. canistri, C. sp.11, C. bisporus as well as C. bisporiger.
C. callinus clade. On the basis of morphology fourspecies can be discerned in this group. However onlythree of these could be inferred as monophyletic. Twocollections (SZMC-NL-0195 and 1751) differ from C.callinus in having somewhat smaller basidiospores(perhaps only statistically), central or hardly eccentricgerm pore and the lack of sclerocystidia. HoweverSZMC-NL-3101 is completely different, growing ondung, up to 2–3 mm diam, with sclerocystidia andnavicular spores. Coprinellus sp. 2 is characterized bytwo-spored basidia, Parasola-like appearance and 8–10 mm long ellipsoid spores, which are obtuse at bothends. Formal description of the mentioned taxa willrequire more specimens.
C. hiascens clade. C. hiascens is delimited here in abroad sense, including C. heterothrix, which implies aconsiderable morphological plasticity for C. hiascens.Traditional characters used to delimit these twospecies (shape of the spore base, angle of germ poreand the shape of the pileocystidia; Orton and Watling1979, Ulje 2005) have considerable overlap betweencollections and no correlation with clade structure canbe recognized. In addition to C. hiascens the phylog-eny provides evidence for two species, C. velatoprui-natus and C. cinereopallidus, with diverticulate veilelements on the pileus. Although the phylogenyclearly separates the latter two taxa (BPP: 1.0, MLBS:100%, MBPP: 1.0), morphologically they are difficult
NAGY ET AL.: COPRINELLUS PHYLOGENY 271

to separate. Both species differ from C. hiascens inhaving globose-ellipsoid cheilocystidia, whereas thecheilocystidia of C. hiascens are lageniform.
‘‘Coprinus’’ crassitunicatus, a species informallydescribed by van de Bogart (1975) also is nested inthis clade. For this species a scattered universal veil ofglobose, thick-walled elements has been reported.Despite this type of veil, the phylogeny suggests aclose relationship to or conspecifity with C. hiascens.
C. pellucidus clade. C. brevisetulosus, C. radicellusand C. pellucidus form a well supported monophyleticgroup (BPP: 1.0, MLBS: 68%, MBPP: 0.97), character-ized by 7–11 mm long ellipsoid basidiospores with acentral germ pore, four-spored basidia and short, upto ca. 90 mm long pileocystidia, and a habitat on dung.The cheilocystidia are globose, except for C. pellucidus,in which lageniform cheilocystidia intermixed withglobose ones have been encountered frequently. Theidentity of such collections has been questioned andthe possibility of being separate taxa have been raised,however our results confirm that these collectionsbelong to C. pellucidus.
Eurysporoid clade.—Species limits within this claderequire more investigation. Several hitherto poorlydefined species belong to this clade coupled with thelowest mean path length between specimens acrossthe entire phylogeny, resulting in poor resolution ofspecies limits and polyphyly of even well establishedspecies such as C. angulatus even when the clade isanalyzed separately (results not shown). The highnumber of incongruent nodes inferred in themixture-model and traditional Bayesian as well asthe ML analyses also can be considered a symptom ofthe weak phylogenetic signal within this clade. Out ofthe species included C. sclerocystidiosus and C.subimpatiens were inferred as monophyletic withstrong support (BPP/MLBS/MBPP 1.00/76/0.67and 1.00/100/1.00 respectively). A weakly supportedclade of specimens labeled C. aff. eurysporus could bediscerned (BPP: 0.87 MLBS: 2MBPP: 0.88), which onthe other hand does not include the holotypespecimen of C. eurysporus. However for the timebeing we refrain from splitting these specimens intotwo species because it would trigger a cascade oftaxon splits, given the present phylogeny.
Velar spherocysts as a phylogenetically informativecharacter.—The structure, color and existence ofvarious veil structures have long been consideredamong the most important characters in coprinoidfungi for delimiting species or higher taxonomicgroups. For instance taxa with globose veil elementshave been classified in section Veliformes (Orton andWatling 1979, Ulje 2005, Keirle et al. 2005, Schafer2010, Horak 2005), which proved polyphyletic (Hop-
ple and Vilgalys 1999; Nagy et al. 2010, 2011; Keirle etal. 2005; Walther et al. 2005), included many speciesnow placed in Coprinopsis (subsections Nivei andNarcotici p.p.). Of section Veliformes subsectionsMicacei and Domestici belong to Coprinellus andcomprise most species in this genus with a richgranulose veil coverage on the pileus. However anumber of representatives of the Core Setulosi clade(e.g. C. doverii, C. mitrinodulisporus, C. plagioporus, C.marculentus Coprinellus sp. 4.) possess globose veilelements, which renders this type of veil homoplastic.In other words the phylogenetic explanatory power ofgranulose veil is low not only in the Psathyrellaeae butalso in Coprinellus. Although literature descriptionstreat all granular-globose veil cells as one and thesame character, their manifestation in the CoreSetulosi and Micacei/Domestici clades representstwo morphological types. Those found in the CoreSetulosi clade are mostly smooth, hyaline, thin-walledand scarce, often difficult to notice under themicroscope and never visible macroscopically or withhand lenses, whereas those in the Micacei/Domesticiclades are more rich and macroscopically observable,usually incrusted or have thickened wall. Thereforewe propose that these two types of veil can be used asphylogenetically informative characters for definingspecies groups when distinguished this way. On theother hand understanding the evolution of velarstructures is an interesting future prospect.
Shall we apply a narrow species concept in Coprinel-lus?—The phylogeny inferred here provides evidencefor the monophyly of most morphologically estab-lished species. During the years of research onCoprinus s.l. taxa a practice of using a narrowmorphological species concept has become fixedamong mycologists. For instance mating experimentsconfirmed the existence of both C. congregatus and C.ephemerus, two extremely closely related speciesdiffering only in the absence or presence of clampconnections, which are difficult to observe, especiallywhen clamp connections are rare and cannot befound on each of the septa, and a hardly detectabledifference in spore sizes (Lange 1952, Ulje 2005).Multispecimen sampling in our dataset is suitable fortesting this narrow species concept in a phylogeneticframework. We intentionally sampled many speci-mens that morphologically fall close to certain taxa inthe Core Setulosi clade but showed one or moredeviating characters (TABLE I). Of these specimens asmaller portion proved to be enigmatic lineages whilethose clustered in already described taxa contributevaluable information on the taxonomic value ofmorphological characters. The species affiliations(TABLE I) imply that the least reliable character is
272 MYCOLOGIA

the occurrence of lageniform cheilocystidia in specieswith predominantly globose-ellipsoid cystidia. Al-though the occurrence of both types of cystidia isknown to be diagnostic for species such as C.subimpatiens, C. dilectus, C. fuscocystidiatus as well asCoprinellus sp. 9 (SZMC-NL-1070) (Ortega andEsteve-Raventos 2003, Ulje 2005, Ulje and Bas 1991,Schafer 2011, Lange and Smith 1953), our resultssuggest that they can occur occasionally in C.pellucidus, C. congregatus and C. uljei as well. Similarlyspore size and shape are usually considered taxonom-ically reliable characters with little amplitude; ourphylogeny suggests that spore size and shape can behighly variable in C. congregatus and the C. hiascensgroup respectively. Taken together molecular data donot support the taxonomic recognition of specimensthat were easily distinguished morphologically, sug-gesting that the narrow morphological species con-cept proposed for setulose Coprinellus taxa is notuniversal, with at least the above-mentioned charac-ters forming exceptions. Whether other charactersshow the same plasticity is an open question, howevera phylogenetic approach for proving the autonomy ofproposed new species is advisable, even in case thecandidate morphological difference seems constantacross collections.
ACKNOWLEDGMENTS
The authors thank the curators of herbaria WU, MICH,WTU, L, M as well as Tommy Knuttson, Francesco Doveri,Jan Cervenka and Peter Finy for loaning and contributingspecimens, and Joe Ammirati for his generous help withliterature. The first author has been supported by theFungal Research Trust. The SYNTHESYS grant of theEuropean Union enabled the first author to visit and revisematerials collected by Kees Ulje in the Nationaal HerbariumNederland, Leiden, in 2006 and 2010. Giovanni Consiglio isthanks for preparing the Latin diagnoses.
LITERATURE CITED
Altekar G, Dwarkadas S, Huelsenbeck JP, Ronquist F. 2004.Parallel Metropolis coupled Markov chain Monte Carlofor Bayesian phylogenetic inference. Bioinformatics 20:407–415, doi:10.1093/bioinformatics/btg427
Borchsenius F. 2007. FastGap 1.0.8. Software distributed bythe authors at (http://192.38.46.42/aubot/fb/FastGap_home.htm).
Doveri F, Sarrocco S, Pecchia S, Forti M, Vannacci G. 2010.Coprinellus mitrinodulisporus, a new species fromchamois dung. Mycotaxon (In press).
Hazi J, Nagy GL, Vagvolgyi C, Papp T. 2010. Coprinellusradicellus, a new species northern distribution. MycolProgress, doi:10.1007/s11557-010-0709-y
TABLE I. Morphological amplitude of certain species of the Core Setulosi clade of Coprinellus
Voucher numbera Species affiliation Morphological difference to typical specimens of that species
SZMC-NL-0764 C. congregatus Spore up to 16 3 8.5 mmSZMC-NL-1221 C. congregatus Spore up to 15 3 5 mm, Pileus consistently acute, like C. sassiSZMC-NL-1443 C. congregatus Lageniform cheilocystidia between globose-ellipsoid onesSZMC-NL-1449 C. congregatus Lageniform cheilocystidia between globose-ellipsoid ones, spore up to 17 3
9.2 mmSZMC-NL-3064 C. congregatus Base of pileocystidia with thickened walls, cheilocystidia globose (not ellipsoid)
with some lageniform interspersed, pileus vivid orange-yellowSZMC-NL-1128 C. congregatus Base of pileocystidia with slightly thickened walls, cheilocystidia globose (not
ellipsoid) with some lageniform interspersed, pileus vivid orange-yellowSZMC-NL-0636 C. congregatus? Pileus up to 4–8 mm wide when fully expanded, cheilocystidia predominantly
utriform, basidiospores 9.5–12(12.4) 3 5.4–6.6 mmSZMC-NL-0628 C. hiascens Pileocystidia with cylindrical neck and +/2 obtuse apex, basidiospores with a
central germ pore and obtuse baseSZMC-NL-1329 C. hiascens Pileocystidia with cylindrical neck and +/2 obtuse apex, basidiospores with a
central germ pore and obtuse baseSZMC-NL-1349 C. hiascens Pileocystidia with cylindrical neck and +/2 obtuse apex, basidiospores with a
central germ pore and obtuse baseSZMC-NL-3856 C. bisporus Cheilocystidia globose (not ellipsoid), basidiospores fusiform, with
pronouncedly conical baseSZMC-NL-1233 C. heterosetulosus Sclerocystidia absent, basidiospores not lentiformSZMC-NL-2928 C. pellucidus Scarce velar sphaerocysts present on pileusSZMC-NL-1446 C. pellucidus Basidiospores dark brown, with distinctly thickened wall, lageniform
cheilocystidia present but scarce
a Deviating specimens qualifying to species or varieties under the narrow morphological species concept applied insubsection Setulosi, their species affiliation based on the phylogenetic analyses and the morphological differences as comparedto typical appearance of the species they belong to. For the phylogenetic placement of these specimens, see FIG. 1.
NAGY ET AL.: COPRINELLUS PHYLOGENY 273

Hopple JS, Vilgalys R. 1994. Phylogenetic relationshipsamong coprinoid taxa and allies based on data fromrestriction site mapping of nuclear rDNA. Mycologia86:96–107, doi:10.2307/3760723
———, ———. 1999. Phylogenetic relationships in themushroom genus Coprinus and dark-spored alliesbased on sequence data from the nuclear gene codingfor the large ribosomal subunit RNA: divergentdomains, outgroups, and monophyly. Mol PhylogenetEvol 13:1–19, doi:10.1006/mpev.1999.0634
Horak E. 2005. Die Rohrlinge und Blatterpilze in Europa.Amsterdam: Elsevier. 555 p.
Keirle MR, Hemmes DE, Desjardin DE. 2004. Agaricales ofthe Hawaiian Islands 8: Agaricaceae. Coprinus andPodaxis, Psathyrellaceae: Coprinellus, Coprinopsis andParasola. Fungal Divers 15:33–124.
Kemp RFO. 1975. Breeding biology of Coprinus species inthe section Lanatuli. Trans Br Mycol Soc 65:375–388,doi:10.1016/S0007-1536(75)80034-9
Kuhner R. 1928. Le developpement et al. potision taxono-mique de l’Agaricus disseminatus Pers. Le Botaniste 20:147–195.
———, Romagnesi H. 1953. Flore Analytique des Champi-gnons Superieures. Paris.
Lange M. 1952. The species concept in the genus Coprinus.Dansk Bot Arkiv 14.
———, Smith AH. 1953. The Coprinus ephemerus group.Mycologia 45:747–780.
Larsson E, Orstadius L. 2008. Fourteen coprophilousspecies of Psathyrella identified in the Nordic countriesusing morphology and nuclear rDNA sequence data.Mycol Res 112:1165–1185, doi:10.1016/j.mycres.2008.04.003
Loytynoja A, Goldman N. 2008. An algorithm for progres-sive multiple alignment of sequences with insertions.Proc Natl Acad Sci USA 102:10557–10562, doi:10.1073/pnas.0409137102
Nagy GL. 2006. Coprinus doverii sp. nov., a unique newspecies of subsection Setulosi from central and south-ern Europe. Mycotaxon 98:147–151.
———, Kocsube S, Papp T, Vagvolgyi C. 2009. Phylogenyand character evolution of the coprinoid mushroomgenus Parasola as inferred from LSU and ITS nrDNAsequence data. Persoonia 22:28–37, doi:10.3767/003158509X422434
———, Urban A, Orstadius L, Papp T, Larsson E, VagvolgyiC. 2010. The evolution of autodigestion in themushroom family Psathyrellaceae (Agaricales) inferredfrom maximum likelihood and Bayesian methods.Mol Phylogenet Evol 57:1037–1048, doi:10.1016/j.ympev.2010.08.022
———, Walther G, Hazi J, Vagvolgyi C, Papp T. 2011.Understanding the evolutionary processes of fungalfruiting bodies: correlated evolution and divergencetimes in the Psathyrellaceae. Syst Biol 60:303–317,doi:10.1093/sysbio/syr005
Ortega A, Esteve-Raventos F. 2003. New and interestingspecies of Coprinus (Coprinaceae, Agaricales) fromAndalusia (southern Spain). Nova Hedwig 76:465–475,doi:10.1127/0029-5035/2003/0076-0465
Orton PD, Watling R. 1979. Coprinaceae 1: Coprinus. BritishFungus Flora. Agarics and Boleti 2. Edinburgh, UK:Royal Botanic Garden. 148 p.
Padamsee M, Matheny PB, Dentinger BT, McLaughlin DJ.2008. The mushroom family Psathyrellaceae: evidencefor large-scale polyphyly of the genus Psathyrella. MolPhylogenet Evol 46:415–429, doi:10.1016/j.ympev.2007.11.004
Pagel M. 1999. Inferring the historical patterns of biologicalevolution. Nature 401:877–884, doi:10.1038/44766
———, Meade A. 2004. A phylogenetic mixture model fordetecting pattern-heterogeneity in gene sequence orcharacter-state data. Syst Biol 53:571–581, doi:10.1080/10635150490522232
———, ———. 2007. BayesPhylogenies 1.0. Softwaredistributed by the authors.
Patrick WWJ. 1977. Sectional nomenclature in the genusCoprinus. Mycotaxon 7:341–355.
Posada D. 2008. jModelTest: phylogenetic model averaging.Mol Biol Evol 25:1253–1256, doi:10.1093/molbev/msn083
Rabosky DL, Dollenan SC, Talaba AL, Lovette IJ. 2007.Exceptional among-lineage variation in diversificationrates during the radiation of Australia’s most diversevertebrate clade. Proc Biol Sci 274:2915–2923,doi:10.1098/rspb.2007.0924
Rambaut A, Drummond A. 2008. Tracer 1.4.1. Softwaredistributed by the authors (http://beast.bio.ed.ac.uk/).
Rannala B. 2002. Identifiability of parameters in MCMCBayesian inference of phylogeny. Syst Biol 51:754–760,doi:10.1080/10635150290102429
Redhead S. 2001. A bully for Coprinus—a story of manure,minutiae and molecules. Field Mycol 2:118–126,doi:10.1016/S1468-1641(10)60532-4
———, Vilgalys R, Moncalvo JM, Johnson J, HoppleJS. 2001. Coprinus Persoon and the disposition ofCoprinus species sensu lato. Taxon 50:203–241,doi:10.2307/1224525
Reijnders AFM. 1979. Developmental anatomy of Coprinus.Persoonia 10:383–424.
Roux P. 2006. Mille et un Champignons. 1223 p.Schafer D. 2010. Keys to sections of Parasola, Coprinellus,
Coprinopsis and Coprinus in Britain. Field Mycol 11:44–51, doi:10.1016/j.fldmyc.2010.04.006
———. 2011. Coprinus dilectus Fr. sensu Josserand. FieldMycol 10:15–18, doi:10.1016/S1468-1641(10)60491-4
Simmons MP, Muller K, Norton AP. 2007. The relativeperformance of indel-coding methods in simulations.Mol Phylogenet Evol 44:724–740, doi:10.1016/j.ympev.2007.04.001
———, Ochoterena H. 2000. Gaps as characters insequence-based phylogenetic analyses. Syst Biol 49:369–381, doi:10.1093/sysbio/49.2.369
Simpson JA, Grgurinovic CA. 2001. The nomenclature ofspecies of Coprinus recorded from south Australia.Australas Mycol 20:55.
Singer R. 1969. Mycoflora australis. Nova Hedwigia Beiheft 29.———. 1986. The Agaricales in modern taxonomy. 4th ed.
Koenigstein, Germany: Koeltz Scientific Books. 441 p.
274 MYCOLOGIA

Staden R, Beal KF, Bonfield JK. 2000. The Staden Package.Methods Mol Biol 132:115–130.
Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa andmixed models. Bioinformatics 22:2688–2690, doi:10.1093/bioinformatics/btl446
Sukumaran J, Holder MT. 2010. Dendropy: a phythonlibrary for phylogenetic computing. Bioinformatics 26:1569–1571, doi:10.1093/bioinformatics/btq228
Thompson JD, Gibson TJ, Higgins DG. 2002. Multiplesequence alignment using Clustal W and Clustal X.Curr Protoc Bioinformatics. Chapter 2, Unit 23.
Ulje CB. 1988. Four new species of Coprinus from theNetherlands. Persoonia 13:479–488.
———. 2005. Coprinus. In: Noordeloos ME, Kuyper TW,Vellinga EC, eds. Flora Agaricina Neerlandica 6:22–109.
———, Bas C. 1991. Studies in Coprinus II. SubsectionSetulosi of section Pseudocoprinus. Persoonia 14:275–339.
———, Keizer J-P. 2003. Coprinus parvulus, a new Coprinusfrom the Netherlands. Persoonia 18:281–283.
———, Noordeloos ME. 2003. Notulae ad Floram agarici-nam neerlandicam XLII. Additions to Coprinus subsec-tion Setulosi. Persoonia 18:259–264.
———, Verkeben A. 2002. A new species of Coprinussubsection Setulosi. Persoonia 18:143–145.
van de Bogart F. 1975. The genus Coprinus in Washingtonand adjacent western states [doctoral dissertation].Seattle: Univ Washington Press. 366 p.
Vasutova M, Antonin V, Urban A. 2008. Phylogenetic studies inPsathyrella focusing on sections Pennatae and Spadiceae—new evidence for the paraphyly of the genus. Mycol Res112:1153–1164, doi:10.1016/j.mycres.2008.04.005
Walther G, Garnica S, Weiss M. 2005. The systematicrelevance of conidiogenesis modes in the gilledAgaricales. Mycol Res 109:525–544, doi:10.1017/S0953756205002868
Wilgenbusch JC, Warren DL, Swofford DL. 2004. AWTY: asystem for graphical exploration of MCMC conver-gence in Bayesian phylogenetic inference. http://ceb.csit.fsu.edu/awty
NAGY ET AL.: COPRINELLUS PHYLOGENY 275





























