Embed Size (px)
Citation preview

Peroxidative Metabolism of �2-Agonists Salbutamol and Fenoteroland Their Analogues
Krzysztof J. Reszka,* Dennis W. McGraw, and Bradley E. Britigan
Research SerVices and Department of Internal Medicine, Department of Veterans Affairs Medical Center,Cincinnati, Ohio 45220, and Department of Internal Medicine, UniVersity of Cincinnati, Cincinnati, Ohio 45267
ReceiVed February 18, 2009
Phenolic �2-adrenoreceptor agonists salbutamol, fenoterol, and terbutaline relax smooth muscle cellsthat relieve acute airway bronchospasm associated with asthma. Why their use sometimes fails to relievebronchospasm and why the drugs appear to be less effective in patients with severe asthma exacerbationsremains unclear. We show that in the presence of hydrogen peroxide, both myeloperoxidase, secreted byactivated neutrophils present in inflamed airways, and lactoperoxidase, which is naturally present in therespiratory system, catalyze oxidation of these �2-agonists. Azide, cyanide, thiocyanate, ascorbate,glutathione, and methimazole inhibited this process, while methionine was without effect. Inhibition byascorbate and glutathione was associated with their oxidation to corresponding radical species by theagonists’ derived phenoxyl radicals. Using electron paramagnetic resonance (EPR), we detected freeradical metabolites from �2-agonists by spin trapping with 2-methyl-2-nitrosopropane (MNP). Formationof these radicals was inhibited by pharmacologically relevant concentrations of methimazole and dapsone.In alkaline buffers, radicals from fenoterol and its structural analogue, metaproteronol, were detected bydirect EPR. Analysis of these spectra suggests that oxidation of fenoterol and metaproterenol, but notterbutaline, causes their transformation through intramolecular cyclization by addition of their aminonitrogen to the aromatic ring. Together, these results indicate that phenolic �2-agonists function as substratesfor airway peroxidases and that the resulting products differ in their structural and functional propertiesfrom their parent compounds. They also suggest that these transformations can be modulated bypharmacological approaches using appropriate peroxidase inhibitors or alternative substrates. Theseprocesses may affect therapeutic efficacy and also play a role in adverse reactions of the �2-agonists.
Introduction
Asthma is a chronic inflammatory disease characterized bybronchial smooth muscle contraction and episodic narrowingof the airway. This, along with edema of the bronchial walland accumulation of airway mucus, limits airflow and gasexchange. Standard treatment of acute asthma exacerbationsincludes inhalation of �2-adrenergic agonists, which activate �2-adrenergic receptors (�2AR)1 on bronchial smooth muscle cells,triggering an increase in cyclic AMP that leads to smooth musclerelaxation. Short-acting �2-adrenergic agonists used to relieveacute airway bronchospasm have included salbutamol, fenoterol,and terbutaline. Why their use sometimes fails to relievebronchospasm and why the drugs appear to be much lesseffective in relieving bronchoconstriction in patients with severeasthma exacerbations (status asthmaticus), or increases risk ofdeath (1-3), is largely unknown.
It is now recognized that inflammation is an importantcomponent of asthma (4-9). Elevated levels of inflammatory
cells, particularly neutrophils (PMN) and eosinophils (EOS) andtheir secretory products, are present in asthmatic airways andincrease during clinical exacerbations of the disease. Uponactivation, EOS and PMN generate superoxide (O2
•-), hydrogenperoxide (H2O2), and secrete unique peroxidases: eosinophilperoxidase (EPO) by EOS and myeloperoxidase (MPO) byPMN. These heme enzymes functionally resemble lactoperoxi-dase (LPO) that is normally present in lung lining fluid, whereit plays a protective role against pathogens. It is now believedthat oxidative processes supported by these enzymes contributeto tissue injury (4-9). MPO, EPO, and LPO commonly utilizeendogenously generated H2O2 to convert substrates such astyrosine (TyrOH), SCN-, NO2
-, Br-, and Cl- (only MPO) to
* To whom correspondence should be addressed. Department of InternalMedicine, University of Cincinnati College of Medicine, P.O. Box 670557,231 Albert Sabin Way, Cincinnati, OH 45267-0557. Tel: (513) 558-6608.Fax: (513) 558-0852. E-mail: [email protected].
1 Abbreviations: �2AR, �2-adrenergic receptors; ABTS, 2,2′-azino-di-(3-ethyl-benzthiazoline-6-sulfonic acid); AscH-, ascorbate; Asc•-, ascorbateradical; BSA, bovine serum albumin; DMPO, 5,5′-dimethyl pyrroline-N-oxide, spin trap; DTPA, diethylenetriamine pentaacetic acid; EPO, eosinophilperoxidase; EPR, electron paramagnetic resonance; GlOx, glucose oxidase;hfsc’s, hyperfine splitting constants; LPO, lactoperoxidase; MNP, 2-methyl-2-nitrosopropane, spin trap; MPO, myeloperoxidase; PMN, neutrophils;TyrOH, tyrosine; TyrO•, tyrosyl radical.
Figure 1. Structures of �2-agonists (salbutamol, fenoterol, and ter-butaline) and the related compounds investigated.
Chem. Res. Toxicol. 2009, 22, 1137–1150 1137
10.1021/tx900071f CCC: $40.75 This article not subject to U.S. Copyright. Published 2009 by American Chemical Society.Published on Web 05/22/2009

reactive metabolites that interact with cell components causingtheir modification and resulting in the loss of normal physi-ological functions. Studies in Vitro showed that �2-agonists affectthe function of granulocytes. Treatment of PMN and EOS withsalbutamol and fenoterol inhibited superoxide production anddegranulation (10, 11). Antioxidant activity with respect tosuperoxide, hydrogen peroxide, hypochlorous acid, and hydroxylradicals was reported for a number of �2-agonists (12). It wasspeculated that the antioxidant properties of the agonists aredue to their scavenging of oxidants (13).
Phenols are typical peroxidase substrates, and their oxidationcan be described by reactions given by eqs 1-3 with MPO asa representative peroxidase and TyrOH as a substrate. Theimmediate metabolite of TyrOH is the tyrosyl radical (TyrO•).
In this mechanism, H2O2 first oxidizes the enzyme in theresting (ferric) state to a highly reactive compound I (MPO-I)(eq 1), with two oxidizing equivalents above ground level.Compound I is then reduced back to the ferric state throughinteraction with 2 molecules of TyrOH. This occurs throughthe intermediacy of compound II (MPO-II), which is the productof one-electron reduction of MPO-I by the substrate (eq 2).During the reaction, two TyrO• radicals are formed. In theabsence of other substrates, phenoxyl radicals typically formdimers through o,o′-biphenyl or p-phenoxyphenyl ether linkages(eq 4) (14, 15). Specifically, tyrosyl radicals recombine to formo,o′-dityrosine as the major product (16-18). The TyrO• radicalsalso react with other targets such as tyrosine residues in proteins(19) or react with reduced glutathione, causing its oxidation (20).Oxidation of phenolics by LPO and EPO systems occurs, inprinciple, according to the same mechanism (eqs 1-4) (21-23).
Given that �2-agonists (salbutamol, fenoterol, and terbutaline)also possess phenolic character (Figure 1) and that, by necessity,they must function in the oxidizing environment of inflamedairways, we hypothesized that they too can be metabolized byairway peroxidases. This metabolic pathway of �2-agonists hasnever been explored. Such a peroxidase-mediated oxidationwould be expected to cause structural modifications of the drugssimilar to those reported for tyrosine, rendering them less active.Consequently, their therapeutic activity might decrease duringtimes of increased airway inflammation that characterize acuteasthma exacerbations.
In this article, we show that salbutamol, fenoterol, andterbutaline (Figure 1) are oxidatively modified in Vitro byperoxidases likely to be present in asthmatic airways, MPO andLPO. It is also shown that these drugs differ markedly in theircapacity to undergo oxidation and that their oxidation productsare highly reactive. Our in Vitro data also suggest that it maybe possible to minimize the oxidative transformation of �2-agonists by peroxidase inhibitors and antioxidants, thus preserv-ing their bronchodilation capacity. Therefore, these observationsmay be pertinent to therapeutic and toxicological functions of�2-agonists.
Experimental Procedures
Materials. Lactoperoxidase (LPO) from bovine milk (EC1.11.1.7), catalase from bovine liver (EC 1.11.1.6; 2,350 U/mg),
horseradish peroxidase (HRP), terbutaline hemisulfate, metaprot-erenol hemisulfate, L-tyrosine, and all other chemicals (hydrogenperoxide (30%), L-GSH, ascorbic acid, methimazole, dapsone,L-methionine, NaSCN, NaCN, NaN3, diethylenetriamine pentaaceticacid (DTPA), 2,2′-azino-di-(3-ethyl-benzthiazoline-6-sulfonic acid)(ABTS), 5,5-dimethyl pyrroline N-oxide (DMPO), 2-methyl-2-nitrosopropane (MNP), and albumin (bovine serum, BSA) wereobtained from Sigma-Aldrich Co. (St. Louis, MO). LPO concentra-tion was determined using ε412 of 1.12 × 105 M-1 cm-1 (24).Myeloperoxidase (MPO) from human leucocytes (lyophilizedpowder, 25 U, RZ (A429/A280) of 0.61) and SOD from bovine liver(5000 U/mg) were obtained from Axxora, LLC (San Diego, CA).MPO was reconstituted with 0.25 mL of distilled water before use.Salbutamol hemisulfate, fenoterol hydrobromide, and glucoseoxidase type X were from MP Biochemicals, Inc. (Solon, OH).R-D(+)-Glucose was from Across Organic (Belgium). All chemicalswere used as received. H2O2 concentration was determined usingε240 of 39.4 M-1 cm-1 (25) and that of DMPO using ε227 of 8 ×103 M-1 cm-1 in water (26). Stock solution of MNP (10 mM indimers) was prepared in 0.1 M phosphate buffers (pH 7.0 and 8.0)containing DTPA (0.1 or 0.2 mM) by stirring overnight in a vesselprotected from light. This procedure generates a significant amountof MNP monomers capable of trapping radicals. Stock solutionsof other reagents were prepared in glass-distilled water.
Spectrophotometric Measurements. Spectra were measuredusing an Agilent diode array spectrophotometer model 8453(Agilent Technologies, Inc., Santa Clara, CA). Oxidation of �2-agonists was studied by measuring absorption spectra at designatedtime points following the start of the reaction. Samples wereprepared in 50 mM acetate buffer (pH 5.0), 50 and 100 mMphosphate buffers (pH 7.0 and 8.0), and 100 mM Tris/HCl (pH9.19). All buffers contained DTPA (100 and 200 µM), andmeasurements were performed at ambient temperature of 20 °C.Typically the reaction was started by the addition of a small aliquotof H2O2 (2, 5, or 10 µL) or glucose oxidase (1 µL), if glucose/glucose oxidase was used to generate H2O2, to a sample containinga studied compound, peroxidase, and, if required, an inhibitingcofactor. Time course measurements were carried out followingchanges in absorbance at 315 nm in 15 s intervals versus absorbanceat 800 nm, where none of the compounds absorb. The 315 nmwavelength was chosen because the �2-agonists’ oxidation productsabsorb intensely near 315 nm and because it is close to theabsorption maximum of tyrosine dimers.
In certain experiments, oxidation of �2-agonists by peroxidaseswas carried out using H2O2 generated by the reaction of glucose (1mM) with glucose oxidase (0.2 µg/mL). The rate of H2O2 generationin these systems was estimated based on the rate of oxidation ofABTS (1 mM) to the green ABTS radical cation (ABTS•+) by HRP,at increasing concentrations of the enzyme. Concentrations ofglucose and glucose oxidase were the same as those used inexperiments with �2-agonists. The plot of the rate of ABTS•+
oxidation at 420 nm (determined from the linear portion of kineticruns) versus [HRP] is a curve, which plateaus above a certainthreshold value [HRP]. The mean value of the rate from the plateauregion (dA420/dt ) ε420 × d[ABTS•+]/dt) was taken as the rate atwhich all H2O2 produced by glucose/glucose was immediately usedup by the enzyme to oxidize ABTS. Calculations were performedusing ε420 (ABTS•+ of 3.6 × 104 M-1 cm-1) (27) and assumingthat stoichiometry for the reaction is 1 mol of H2O2 to 2 mols ofABTS. The rate of H2O2 generation determined in this way was3.33 µM/min, based on two separate determinations.
Because the commercially available fenoterol exists in the formof hydrobromide, and because the bromide anion (Br-) is convertedby peroxidases to brominating hypobromous acid (HOBr), therewas the possibility that Br- might interfere with enzymatic oxidationof the drug. However, experiments performed in the presence oftaurine and L-methionine (traps for HOBr), as well as additionaldoses of bromide (as NaBr) added to the sample, did not revealany meaningful changes in the oxidation kinetics of fenoterol. Incontrast to taurine, L-methionine is considered to afford a lessreactive product upon reaction with HOBr. However, 2 mM
MPO + H2O2 f MPO-I + H2O (1)
MPO-I + TyrOH f MPO-II + TyrO• (2)
MPO-II + TyrOH f MPO + TyrO• (3)
TyrO•+TyrO• f f (TyrOH)2 dimer (4)
1138 Chem. Res. Toxicol., Vol. 22, No. 6, 2009 Reszka et al.

L-methionine only minimally inhibited the oxidation of fenoterol(95% of the control (see Table 2)). Therefore, it was concludedthat Br- that is naturally present in the sample does not significantlyinfluence the metabolism of fenoterol. To evaluate the role ofoxygen in oxidative processes, spectrophotometric experiments wereperformed after bubbling N2 gas through the sample (1 mL volume)for 5 min before the start of the reaction (H2O2 addition) and thenbetween readouts, which were collected every 1 min.
EPR Measurements. EPR spectra were recorded using a BrukerEMX EPR spectrometer (Bruker Biospin Co., Billerica, MA),operating in X band and equipped with a high sensitivity resonatorER 4119HS. Formation of free radicals from �2-agonists was studiedin samples prepared in 100 mM phosphate buffer (pH 7.0 and 8.0)/DTPA (0.2 mM) (total volume 250 µL) containing MNP, MPO(or LPO), and the agonists. The reaction was initiated by theaddition of H2O2 as the last component. In experiments, in whichH2O2 was generated using glucose (1 mM) and glucose oxidase(3.9 µg/mL), glucose oxidase was added as a last component. Thesample was transferred to a flat aqueous EPR cell, and recordingwas started 1 min after initiation of the reaction. Typically, spectraof MNP adducts were recorded using microwave power of 20 mW,modulation amplitude of 0.1 mT, receiver gain of 2 × 105,conversion time of 40.96 ms, time constant of 81.92 ms, and scanrate of 10 mT/41.92 s. EPR spectra are the average of 5 scans andrepresent results of typical experiments. Unless stated otherwise,direct EPR measurements (spin traps omitted) of free radicalsderived from �2-agonists were performed using conditions similarto those for the detection of MNP adducts. EPR spectra weresimulated using WINSIM software developed at NIEHS/NIH (RTP,NC).
The effect of methimazole and dapsone on the formation of freeradicals from drugs was studied in phosphate buffers (pH 7.0 and8.0) containing DTPA (0.2 mM) and MNP (10 mM in dimers).Oxidation was carried out by MPO (0.43 units/mL)/H2O2 (37 µM)and LPO (0.39 µM)/glucose (1 mM)/glucose oxidase (0.8 µg/mL).In experiments involving methimazole, the concentrations offenoterol and terbutaline were 0.47 mM, while in those involvingdapsone, their concentrations were 0.047 mM.
The effects of AscH- and GSH on the metabolism of drugs wereinvestigated using samples in 100 mM phosphate buffer (pH 7.0)/DTPA (0.1 mM) (total volume 250 µL) containing salbutamol orfenoterol and MPO, and the reaction was initiated by the addition
of H2O2 as the last component. When the effect of GSH was studied,spin trap DMPO (18 mM) was also present. The sample wastransferred to a flat aqueous EPR cell, and recording was started 1min after initiation of the reaction. The spectra of DMPO adductswere recorded using the same parameters as those described above,but the sweeping rate was 8 mT/41.92 s. The EPR spectra ofascorbate radicals were obtained using microwave power of 5 mW,modulation amplitude of 0.05 mT, and scan rate of 4 mT/41.92 s.The EPR spectra shown are the average of 5 scans and representresults of a typical experiment.
Results
Interaction of �2-Agonists with the MPO System. Tyrosineand other phenolics are oxidized by peroxidases to free radicalsthat can recombine to give rise to corresponding dimericproducts (15-18, 23). Because the �2-adrenergic agonistssalbutamol, fenoterol, and terbutaline also contain phenolicmoieties (Figure 1), we hypothesized that they too can bemetabolized by peroxidases. Spectrophotometric measurementsshowed that these compounds have no significant absorptionaround 300 nm and none above this wavelength (Figure 2A,B,D,traces a). However, when H2O2 was added to salbutamol inbuffer (pH 7.0) containing MPO, a new absorption band centeredaround 315 nm appeared (Figure 2A), suggesting the formationof a new metabolite. The new absorption is in the rangecharacteristic of tyrosine dimers (16-18, 23). Measurementsof the absorbance at 315 nm (A315) following additions of smallaliquots of H2O2 (9.85 µM per dose) to salbutamol (1 mM) inbuffer (pH 7.0) containing MPO (200 mU/mL) showed that inthe range 0-100 µM, the plot of ∆A315 versus [H2O2] is linear(Figure 2A, inset). On the basis of this relationship, a molecularabsorptivity, ε315, for the generated mixture of products wasdetermined to be 1210 ( 19 M-1 cm-1 (N ) 3). This value isin the range of molar absorptivities at 300 nm determined fora mixture of products derived from phenolics oxidized enzy-matically at pH 5.0 (23). The time course of the reactionfollowing a single bolus addition of H2O2 shows that A315
plateaus after ∼10 min of reaction (Figure 3, trace a). The lackof further increase in A315 is chiefly due to the consumption of
Figure 2. Absorption spectra observed during the oxidation of salbutamol (A), tyrosine (B), fenoterol (C), and terbutaline (D) by MPO (200mU/mL) and H2O2 in 50 mM phosphate buffer (pH 7.0) containing 100 µM DTPA. The reaction was initiated by a bolus addition of H2O2 to a finalconcentration of 50 µM (in A, B, and D) and 25 µM (in C). Spectra were recorded every 30 s. Initial concentrations of �2-agonists were 1 mM.Inset in A and C: ∆A315 versus H2O2 consumed during oxidation by MPO of salbutamol and fenoterol, respectively. H2O2 (0.98 mM) was beingadded in small portions (5 µL to the salbutamol sample and 2 µL to the fenoterol sample), and when A315 stabilized, ∆A315 was determined aftereach single dose of H2O2. Other conditions were as described for the main panel.
Oxidation of �2-Agonists Chem. Res. Toxicol., Vol. 22, No. 6, 2009 1139

H2O2. For comparison, absorption spectra observed during theoxidation of tyrosine by MPO/H2O2 are shown in Figure 2B.Because both the spectral changes and the kinetics of oxidationof salbutamol and tyrosine (Figure 3, traces a and b, respec-tively) are similar, we propose that oxidation of this �2-agonistgenerates the corresponding dimer, o,o′-disalbutamol, amongother possible products. The efficacy of salbutamol oxidationby MPO/H2O2 is pH-dependent. After a 30 min reaction ofsalbutamol (1 mM) with MPO (2 mU/mL) and H2O2 (119 µM),∆A315 increased from near zero at pH 5.0, to 0.12 at pH 7.0and 0.178 at pH 8.0. This dependence is similar to that reportedfor oxidation of tyrosine (17, 18).
Under the conditions used, we observed that the efficacy ofsalbutamol oxidation was dependent upon the concentration ofthe agonist. At constant concentration of H2O2 of 52 µM, therate of oxidation and the amount of the metabolite formed(measured as ∆A315 over 25 min) increased as the concentrationof the drug increased from 100 µM to 1 mM (Figure 4A, tracesa-d). This type of response is suggestive of a slow reaction ofsalbutamol with peroxidase compound II. The yield of salbuta-mol oxidation was also measured at constant concentration ofthe agonist of 1 mM and changing [H2O2] from 25 to 200 µM.The maximum yield was found at around 100 µM of theperoxide. The decrease in yield at higher concentrations of H2O2
may be due to the inhibition of the peroxidase. Cyanide andazide (1 mM each) substantially inhibited the oxidation ofsalbutamol by MPO and H2O2, while catalase (235 units/mL)completely blocked the reaction (Table 1), confirming that theoxidation of salbutamol is a peroxidase-dependent process.
Oxidation of salbutamol was compared with that of fenoterol.In contrast to salbutamol, which contains only one monophenolicmoiety, fenoterol possesses both a monophenolic and a diphe-nolic (1,3-dihydroxybenzene or resorcinol) moiety. It wastherefore anticipated that fenoterol might behave differently fromsalbutamol when exposed to peroxidases, especially sinceresorcinols are oxidized by peroxidases (28). Figure 2C showsspectra observed during the interaction of fenoterol (1 mM) withMPO (200 mU/mL) and H2O2 (25 µM) at pH 7.0. Under theconditions used, and in contrast to salbutamol and tyrosine, thenew species formed absorbs both in the UV and visible regionsof the spectrum. The spectral lines intersect at 322, 505, and735 nm and show new absorption maxima at 360 and 550 nm.At higher concentrations of H2O2, the absorption maxima areless pronounced, and the intersection points are absent (notshown), presumably due to the formation of a mixture of variousdimeric and/or polymeric products. We confirmed that oxidation
of the resorcinol moiety of the drug is responsible for theseeffects by showing that oxidation of a related �2-agonist,terbutaline (Figure 1), which contains only the resorcinol group,yields spectral features (Figure 2D) similar to those of oxidizedfenoterol. However, for oxidized terbutaline, the absorptionmaxima (340 and 500 nm) are blue-shifted compared to thosefound for fenoterol. Measurements of A315 following additionsof small aliquots of H2O2 (3.9 µM per dose) to fenoterol (1mM) in buffer (pH 7.0) containing MPO (200 mU/mL) showedthat the plot of ∆A315 versus [H2O2] is nonlinear (Figure 2C,inset), presumably due to inactivation of the enzyme. Thus,under similar conditions, fenoterol behaves differently fromsalbutamol, for which the relationship between ∆A315 and [H2O2]was linear (Figure 2A, inset).
The time course of absorption changes at 315 nm recordedduring the oxidation of fenoterol following single bolus addition
Figure 3. Time course of absorption changes measured at 315 nmduring the oxidation of salbutamol (a), tyrosine (b), and fenoterol (c).The reaction was initiated by the addition of a small aliquot of H2O2
(final concentration 60 µM) to pH 7.0 buffer (50 mM phosphate)containing a drug (1 mM each), MPO (0.2 U/mL), and DTPA (100µM). Absorbance was read every 30 s. N ) 2 (salbutamol and fenoterol)and 3 (tyrosine).
Figure 4. Dependence of the oxidation of salbutamol (A) and fenoterol(B) on drug concentration. The rate of oxidation by MPO (200 mU/mL) and H2O2 (50 µM) was measured in 50 mM phosphate buffer(pH 7.0) containing 100 µM DTPA, by following changes in absorbanceat 315 nm.
Table 1. Efficacy of the Oxidation of Fenoterol (1 mM) andSalbutamol (1 mM) by MPO (200 mU/mL) and H2O2 (50µM) in 50 mM Phosphate Buffer (pH 7.0) Containing 0.1
mM DTPA in the Presence of Modulating Co-Factorsa
∆A315 (%)
fenoterol salbutamol
control 100 100catalase (235 U/mL) 0.0 0.0NaCN (1 mM) 0.24 ( 0.32 12.5 ( 9.1NaN3 (1 mM) 17.9 ( 4.2 6.6 ( 1.3GSH (0.1 mM)b 43.3 ( 14.5 62.0 ( 17.3BSA (0.5 mg/mL) 110 ( 2.0 99.0 ( 2.0
a The extent of oxidation is expressed as ∆A315 ( SE versus control(in %) during 22 min of reaction at 20° C. Values are the mean of atleast duplicate determinations. b In experiments with GSH, con-centrations of salbutamol and fenoterol were 100 µM each, and thereaction was continued for 30 min.
1140 Chem. Res. Toxicol., Vol. 22, No. 6, 2009 Reszka et al.

of H2O2 is shown in Figure 3 (trace c). At this wavelength, thefenoterol-derived metabolites absorb approximately twicemore intensely than metabolites from salbutamol or tyrosine(Figure 3, traces a and b). This may be due to the presence ofthe resorcinol moiety, oxidation of which may give rise to morecomplex polyphenolic products with higher molar absorptivity.Similar to salbutamol, oxidation of fenoterol by MPO/H2O2
depends on pH. Measurements of ∆A315 during 30 min ofreaction of fenoterol (1 mM) with MPO (200 mU/mL) and H2O2
(119 µM) showed that ∆A315 increased from 0.0165 at pH 5.0to 0.097 at pH 7.0, and 0.129 at pH 8.0 (corrected forautoxidation). However, in contrast to salbutamol, at pH 8 (andhigher) fenoterol undergoes autoxidation, as evidenced by a slowincrease in A315 without externally added H2O2 and/or MPO.When MPO was added, A315 increased faster (not shown),suggesting the accumulation of H2O2 during the autoxidationphase. This was verified by the addition of catalase, whichabolished the effect of MPO (not shown).
The rate of fenoterol oxidation by MPO, following a singlebolus addition of H2O2 (52 µM), changed minimally when theconcentration of fenoterol was increased from 100 to 1000 µM(Figure 4B). This behavior is different from that observed duringthe oxidation of salbutamol, for which the rate of oxidation wasstrongly dependent on the concentration of the drug (Figure 4A).The observation that ∆A315 reached almost the same final levelsat each concentration of fenoterol tested (Figure 4B), suggeststhat the extent of fenoterol oxidation depends on the concentra-tion of the peroxide. Measurements of ∆A315 versus [H2O2] atconstant fenoterol concentration (1 mM) and MPO of 0.1 U/mLshowed that the maximum yield was at [H2O2] near 50 µM.The decrease at higher concentrations of the peroxide is probablydue to inactivation of the enzyme as was already suggested bydata in Figure 2C (inset). Addition of SOD (100 units) waswithout any apparent effect on the oxidation of fenoterol,suggesting that the free superoxide radical was not involved inthe reaction. Similarly to salbutamol, the oxidation of fenoterolwas substantially inhibited by the peroxidase poisons cyanideand azide and completely prevented by catalase (Table 1),further supporting the peroxidative metabolism of the drug’soxidation. Together, these observations confirm that salbutamoland fenoterol undergo oxidation by MPO in the presenceof H2O2. They also demonstrate that the MPO metabolism ofsalbutamol differs from that of fenoterol both in the nature ofthe products formed and in responses to changes in H2O2 andsubstrate concentrations.
The role of oxygen was examined using fenoterol as asubstrate as this compound produced more profound changesin absorption spectra. When the sample of fenoterol wasdeaerated by bubbling N2 gas through the solution, the rate ofthe reaction and the extent of oxidation both decreased byapproximately 50% (not shown), suggesting that oxygen isinvolved, at least partially, in the oxidative transformation ofthe agonist. One possible explanation of this effect may be thatoxygen adds to phenoxyl radicals resulting in hydroxylation ofthe aromatic ring, a process known to occur during aerobic(photo)oxidation of resorcinols (29).
Effect of Ascorbate and Glutathione on the Oxidationof Salbutamol and Fenoterol. Given that the respiratory tractlining fluid contains antioxidants such as ascorbate (AscH-) andglutathione (GSH) (30), it was expected that they could affectthe oxidation of �2-agonists. The concentration of AscH- inairway fluid was estimated to be near 100 µM (30), and in ourexperiments, we used concentrations within this physiologicalrange. Figure 5 shows changes in A315 versus time for salbutamol
(A) and fenoterol (B) reacting with MPO/H2O2 in the absenceand presence of 10, 20, 40, and 100 µM AscH- (traces a-e,respectively). It is found that AscH- affects the oxidation ofsalbutamol and fenoterol in a similar manner, causing delay inthe net oxidation of the drugs. An increase in A315 is observedafter a lag period, the duration of which depends on AscH-
concentration. It is concluded that only when AscH- isconsumed the net oxidation of the drugs is observed. At 100µM AscH-, there is a complete inhibition of oxidation of bothsalbutamol and fenoterol samples. This is understandable giventhat the concentration of H2O2 was only 50 µM and that theperoxide was used to oxidize both the drugs and AscH-. Theobservation that oxidation of �2-agonists is resumed after a lagperiod suggests that this delay is not due to inhibition/inactivation of the enzyme but rather due to the interaction ofAscH- with the drugs’ derived phenoxyl radicals. The proposedmechanism of the inhibition is described by the reaction givenin Scheme 1A,B, which shows that recovery of the drug occursat the expense of AscH-, which is oxidized to the ascorbateradical (Asc•-).
We investigated the formation of Asc•- during the oxidationof �2-agonists by MPO/H2O2 in the presence of AscH- (100µM) by EPR. Ascorbate radicals are relatively stable andproduce a distinct EPR spectrum, a doublet with a hyperfinesplitting constant of 0.18 mT. Oxidation of AscH- by MPO/H2O2 alone is relatively inefficient, as evidenced by the weakEPR signal of Asc•- generated by the system (Figure 6, spectrumB). In contrast, EPR spectra generated by oxidation of AscH-
in the presence of salbutamol (40 and 100 µM) are more intense(Figure 6C and D, respectively), approximately by 76% and117%, and in the presence of fenoterol (20 µM) by 270% (Figure6, spectrum E), indicating that both of these agonists stimulatethe oxidation of AscH-, with fenoterol being substantially more
Figure 5. Oxidation of salbutamol (A) and fenoterol (B) by MPO/H2O2; effect of ascorbic acid. Salbutamol (1 mM) and fenoterol (200µM) were exposed to MPO (200 mU/mL) and H2O2 (50 µM) in theabsence and presence of 10, 20, 40, and 100 µM ascorbic acid (tracesa-e, respectively) in pH 7.0 buffer (50 mM phosphate containing 100µM DTPA), and A315 was measured in 30 s intervals following H2O2
addition.
Oxidation of �2-Agonists Chem. Res. Toxicol., Vol. 22, No. 6, 2009 1141

effective. The spectrum in Figure 6A shows that when MPOand H2O2 are absent, the level of Asc•- is below the detectionlimit.
Our spectrophotometric measurements showed that GSH alsoinhibits the oxidation of �2-agonists by MPO and H2O2 (Table1), consistent with prior observations that oxidation of tyrosineto dityrosine by MPO/H2O2 can be inhibited by GSH (20).Because GSH is a poor peroxidase substrate, one potentialmechanism of inhibition is the interaction of the generatedphenoxyl radical metabolites with the thiol as depicted inScheme 1A,B. This reaction is accompanied by the formationof GS• radicals as described for tyrosine and other phenolics(31). To verify that this mechanism operates also for �2-agonists,we performed EPR experiments combined with spin trapping,
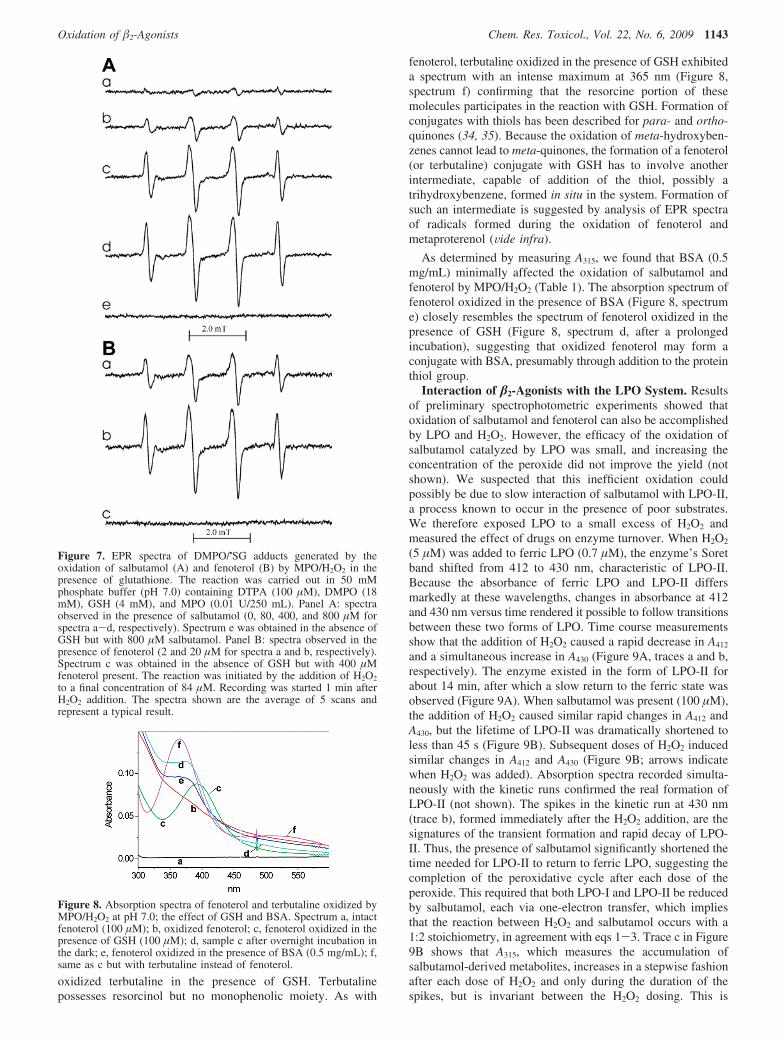
in order to detect GS• radicals. When GSH was exposed toMPO/H2O2 in the presence of the spin trap DMPO andsalbutamol, EPR spectra of the DMPO/•SG adduct weredetected. The hfsc’s aN ) 1.51 mT and a�
H ) 1.61 mT are inagreement with those determined in earlier reports for the sameDMPO adduct (31-33). In Figure 7, panel A, are shown spectrarecorded in the absence (a) and presence of 80, 400, and 800µM salbutamol (spectra b-d, respectively). They show thatsalbutamol in a concentration-dependent manner enhances theoxidation of GSH to GS• evidenced by more intense EPR spectraof DMPO/•SG adducts. No radicals were detected when salb-utamol alone (GSH omitted) was incubated with MPO and H2O2
(Figure 7A, spectrum e), perhaps due to a low efficacy of theaddition of the drug-derived phenoxyl radicals to DMPO and/or to the poor stability of the resulting adduct.
When similar experiments were conducted using fenoterol(2 and 20 µM) instead of salbutamol, the EPR spectra shownin Figure 7, panel B (spectra a and b, respectively) weredetected. Although the general pattern is similar to that observedin the presence of salbutamol, namely, that the intensity of theEPR spectra increases as the concentration of fenoterol increasesfrom (spectra a and b), fenoterol appears to be markedly moreefficient in stimulating the oxidation of GSH. The EPR spectrumgenerated in the presence of 2 µM fenoterol (Figure 7B,spectrum a) is approximately 2-fold more intense than thatobserved in the presence of 80 µM salbutamol (Figure 7A,spectrum b). This further confirms the higher reactivity of ametabolite derived from fenoterol. When GSH was omitted, noradicals from fenoterol were detected by spin trapping withDMPO (Figure 7B, spectrum c). The higher stimulatory actionof fenoterol, when compared to that of salbutamol, implies thatthe compound’s resorcinol moiety may play a dominating rolein the interaction with GSH. The proposed cycle of redoxreactions involving fenoterol, AscH-, and GSH is depicted inScheme 1B.
We found that oxidation of fenoterol in the presence of GSHgenerates a new species with an absorption maximum at 395nm (Figure 8, spectrum c), which upon longer incubation shiftedto 375 nm (spectrum d). The new species was tentativelyascribed to a fenoterol-SG conjugate. Because the oxidationof salbutamol in the presence of GSH did not produce thisspectral feature, it suggested that the fenoterol resorcine moietymight be involved (Scheme 1B). To test this possibility, we
Scheme 1. Proposed Reactions of Salbutamol (A) and Fenoterol (B) Initiated by MPO (or LPO) and H2O2a
a Pathway B is also pertinent for reactions involving terbutaline and metaproterenol.
Figure 6. EPR spectra of the ascorbate radical in pH 7.0 buffer (50mM) containing DTPA (0.2 mM). (A) Ascorbate (100 µM) in buffer,no additives. (B) Same as in A in the presence of MPO (0.01 U/250µL) and H2O2 (39 µM). (C and D) Same as B but in the presence ofsalbutamol (40 and 100 µM, respectively). (E) Same as B but in thepresence of fenoterol (20 µM). The reaction was initiated by the additionof H2O2. Recording was started 1 min after H2O2 addition. Instrumentalsettings: modulation amplitude, 0.05 mT; receiver gain, 2 × 105; timeconstant, 40.96 ms; conversion time, 40.96 ms; seep time, 41.94 s.Sample volume was 250 µL. The spectra shown are the average of 5scans and represent typical results.
1142 Chem. Res. Toxicol., Vol. 22, No. 6, 2009 Reszka et al.
oxidized terbutaline in the presence of GSH. Terbutalinepossesses resorcinol but no monophenolic moiety. As with
fenoterol, terbutaline oxidized in the presence of GSH exhibiteda spectrum with an intense maximum at 365 nm (Figure 8,spectrum f) confirming that the resorcine portion of thesemolecules participates in the reaction with GSH. Formation ofconjugates with thiols has been described for para- and ortho-quinones (34, 35). Because the oxidation of meta-hydroxyben-zenes cannot lead to meta-quinones, the formation of a fenoterol(or terbutaline) conjugate with GSH has to involve anotherintermediate, capable of addition of the thiol, possibly atrihydroxybenzene, formed in situ in the system. Formation ofsuch an intermediate is suggested by analysis of EPR spectraof radicals formed during the oxidation of fenoterol andmetaproterenol (Vide infra).
As determined by measuring A315, we found that BSA (0.5mg/mL) minimally affected the oxidation of salbutamol andfenoterol by MPO/H2O2 (Table 1). The absorption spectrum offenoterol oxidized in the presence of BSA (Figure 8, spectrume) closely resembles the spectrum of fenoterol oxidized in thepresence of GSH (Figure 8, spectrum d, after a prolongedincubation), suggesting that oxidized fenoterol may form aconjugate with BSA, presumably through addition to the proteinthiol group.
Interaction of �2-Agonists with the LPO System. Resultsof preliminary spectrophotometric experiments showed thatoxidation of salbutamol and fenoterol can also be accomplishedby LPO and H2O2. However, the efficacy of the oxidation ofsalbutamol catalyzed by LPO was small, and increasing theconcentration of the peroxide did not improve the yield (notshown). We suspected that this inefficient oxidation couldpossibly be due to slow interaction of salbutamol with LPO-II,a process known to occur in the presence of poor substrates.We therefore exposed LPO to a small excess of H2O2 andmeasured the effect of drugs on enzyme turnover. When H2O2
(5 µM) was added to ferric LPO (0.7 µM), the enzyme’s Soretband shifted from 412 to 430 nm, characteristic of LPO-II.Because the absorbance of ferric LPO and LPO-II differsmarkedly at these wavelengths, changes in absorbance at 412and 430 nm versus time rendered it possible to follow transitionsbetween these two forms of LPO. Time course measurementsshow that the addition of H2O2 caused a rapid decrease in A412
and a simultaneous increase in A430 (Figure 9A, traces a and b,respectively). The enzyme existed in the form of LPO-II forabout 14 min, after which a slow return to the ferric state wasobserved (Figure 9A). When salbutamol was present (100 µM),the addition of H2O2 caused similar rapid changes in A412 andA430, but the lifetime of LPO-II was dramatically shortened toless than 45 s (Figure 9B). Subsequent doses of H2O2 inducedsimilar changes in A412 and A430 (Figure 9B; arrows indicatewhen H2O2 was added). Absorption spectra recorded simulta-neously with the kinetic runs confirmed the real formation ofLPO-II (not shown). The spikes in the kinetic run at 430 nm(trace b), formed immediately after the H2O2 addition, are thesignatures of the transient formation and rapid decay of LPO-II. Thus, the presence of salbutamol significantly shortened thetime needed for LPO-II to return to ferric LPO, suggesting thecompletion of the peroxidative cycle after each dose of theperoxide. This required that both LPO-I and LPO-II be reducedby salbutamol, each via one-electron transfer, which impliesthat the reaction between H2O2 and salbutamol occurs with a1:2 stoichiometry, in agreement with eqs 1-3. Trace c in Figure9B shows that A315, which measures the accumulation ofsalbutamol-derived metabolites, increases in a stepwise fashionafter each dose of H2O2 and only during the duration of thespikes, but is invariant between the H2O2 dosing. This is
Figure 7. EPR spectra of DMPO/•SG adducts generated by theoxidation of salbutamol (A) and fenoterol (B) by MPO/H2O2 in thepresence of glutathione. The reaction was carried out in 50 mMphosphate buffer (pH 7.0) containing DTPA (100 µM), DMPO (18mM), GSH (4 mM), and MPO (0.01 U/250 mL). Panel A: spectraobserved in the presence of salbutamol (0, 80, 400, and 800 µM forspectra a-d, respectively). Spectrum e was obtained in the absence ofGSH but with 800 µM salbutamol. Panel B: spectra observed in thepresence of fenoterol (2 and 20 µM for spectra a and b, respectively).Spectrum c was obtained in the absence of GSH but with 400 µMfenoterol present. The reaction was initiated by the addition of H2O2
to a final concentration of 84 µM. Recording was started 1 min afterH2O2 addition. The spectra shown are the average of 5 scans andrepresent a typical result.
Figure 8. Absorption spectra of fenoterol and terbutaline oxidized byMPO/H2O2 at pH 7.0; the effect of GSH and BSA. Spectrum a, intactfenoterol (100 µM); b, oxidized fenoterol; c, fenoterol oxidized in thepresence of GSH (100 µM); d, sample c after overnight incubation inthe dark; e, fenoterol oxidized in the presence of BSA (0.5 mg/mL); f,same as c but with terbutaline instead of fenoterol.
Oxidation of �2-Agonists Chem. Res. Toxicol., Vol. 22, No. 6, 2009 1143

consistent with oxidation of the drug only during enzymeturnover. Trace c in Figure 9A shows that when salbutamolwas omitted, changes in A315 versus time were minimal.
Responses of LPO to H2O2 depended on the concentrationof salbutamol. In the presence of 10 µM salbutamol, the lifetimeof LPO-II was approximately 420 s after the first dose of H2O2,markedly longer than that at 100 µM salbutamol. Duringsubsequent doses of H2O2, as the drug was consumed, thelifetime of LPO-II increased (not shown). However, also at 1mM salbutamol, the formation of LPO-II was detected, sug-gesting that the interaction of the drug with LPO-II is the rate-limiting step. This is consistent with the view that peroxidativeactivity depends on the rate with which a substrate reducescompound II to the native enzyme (36). Together, theseobservations suggest that salbutamol undergoes one-electronoxidation both by LPO-I and LPO-II, in agreement withreactions described by eqs 2 and 3, with the reaction involvingLPO-II being the slowest step.
Under conditions similar to those in Figure 9B, kinetic tracesrecorded in the presence of fenoterol (100 µM) did not showthe formation of LPO-II (Figure 9C; traces a and b show thatA412 and A430 are nearly constant). Thus, in contrast to salbuta-
mol, fenoterol appears to readily react with both LPO-I andLPO-II, providing rapid enzyme turnover.2 The stepwiseincrease in A315 following every dose of the peroxide (Figure9C, trace c) is indicative of the formation of a fenoterol-derivedmetabolite. This occurs only during the brief period duringwhich H2O2 is available. The small monotonous increase inabsorbance at 412 and 430 nm over the time of observation(Figure 9C) is due to the accumulation of fenoterol metabolites,which absorb in this region (see spectra in Figure 2C).
We next conducted experiments to find a relationship between∆A315 and H2O2 consumed for salbutamol and fenoterol oxidizedby a LPO/H2O2 system. In the H2O2 concentration range 0-50µM, the plot of ∆A315 versus [H2O2] consumed is linear(correlation coefficient 0.99-0.999) for both salbutamol andfenoterol (not shown). On the basis of this relationship, the molarabsorptivity for products derived from salbutamol was deter-mined to be ε315 ) 1149 ( 66 M-1 cm-1. This value is close tothe 1210 M-1 cm-1 determined in this study using the MPO/H2O2 system. For products derived from the oxidation offenoterol, ε315 was determined to be 1688 ( 37 M-1 cm-1. Bothof these values are in the range of molar absorptivitiesdetermined at 300 nm for a number of phenolics oxidized byLPO/H2O2 at pH 5 (23).
For LPO, low concentrations of H2O2 generated smallamounts of metabolites, and the use of higher concentrationsof H2O2 did not improve the yield. We expected that moreefficacious oxidation of salbutamol by LPO could be achievedby continuous generation of low H2O2 fluxes using the glucose/glucose oxidase system, where inactivation of LPO could beminimized by choosing appropriately low concentrations ofglucose oxidase. An additional advantage of this system is thatit better mimics the generation of the peroxide in ViVo. Whensalbutamol (100 µM) was exposed to LPO (0.7 µM), glucose(1 mM), and glucose oxidase (0.1 µg/mL), there was acontinuous increase in A315 (Figure 10, trace c), indicating theoxidation of the drug. During the entire time of observation,LPO remained in its ferric form (A412 and A430 approximatelyconstant) (Figure 10, traces a and b, respectively), suggestingthat under the conditions of slow generation of H2O2, LPO-IIdoes not accumulate in quantities detectable by our recordingsystem, being rapidly reduced to ferric LPO. Oxidation ofsalbutamol was still observed when the concentration of glucoseoxidase was increased 2-fold or more, to increase the rate of
2 We note that Br-, copresent with fenoterol, may participate in thisreaction and contribute to the rapid enzyme turnover.
Figure 9. Oxidation of salbutamol and fenoterol by a LPO/H2O2 system.(A) Time course of absorption changes at 412 nm (ferric LPO, trace a)and 430 nm (LPO compound II, trace b) following the addition of H2O2
(5 µM) to LPO (0.7 µM) in 0.1 M phosphate buffer (pH 7.0) in theabsence of �2-agonists. (B) The same as in A but in the presence ofsalbutamol (100 µM). Trace c shows changes in absorbance at 315 nmdue to the accumulation of salbutamol oxidation products followingthe addition of H2O2 (indicated by arrows). (C) The same as in A butin the presence of fenoterol (100 µM). Data points were collected in15 s intervals.
Figure 10. Oxidation of salbutamol by LPO and H2O2 (generated byglucose and glucose oxidase) at pH 7.0. The sample containedsalbutamol (100 µM), LPO (0.7 µM), and glucose (1 mM), and thereaction was initiated by the addition of glucose oxidase (0.1 µg/mL)as the last component (arrow). Traces a, b, and c correspond toabsorption changes at 412 nm, 430 nm, and 315 nm, respectively. GlOxdesignates glucose oxidase. Data points were collected in 15 s intervals.
1144 Chem. Res. Toxicol., Vol. 22, No. 6, 2009 Reszka et al.

H2O2 generation, but the process was less efficient, presumablydue to the formation of the less reactive intermediate LPOcompound III (not shown). LPO-III is formed in the presenceof a large excess of H2O2 over LPO and is characterized by itsSoret band at 424 nm (24, 37). Oxidation of salbutamol by LPO-III is in agreement with prior observations that phenols reactwith LPO-III converting it to the native enzyme, which thenre-enters the peroxidative cycle (23, 38).
Effect of Inhibitors on the Oxidation of �2-Agonists byLPO/H2O2. Oxidation of salbutamol and fenoterol by LPO/glucose/glucose oxidase was strongly inhibited by azide butweakly by cyanide (Table 2). Thiocyanate, the natural substratefor LPO, and GSH markedly inhibited the oxidation of fenoteroland salbutamol (Table 2). In contrast, L-methionine, in whichthe thiol group is methylated, was inactive, emphasizing theimportance of the free -SH group for effective antioxidant action.Because LPO-catalyzed oxidation could be a mechanism thatinactivates �2-agonists in the airways, we sought to determinewhether pharmacological inhibitors of peroxidase could affectoxidation of these drugs. Methimazole, an antithyroid drug, isknown to inhibit thyroid peroxidase and LPO (39, 40). Weobserved that methimazole markedly inhibited the oxidation ofboth agonists by the LPO system (Table 2). In the presence ofmethimazole, the intensity of the LPO Soret band markedlydecreased during the reaction indicating that inhibition of�-agonist oxidation was primarily due to inactivation of theenzyme, in agreement with previous reports (40-42). We alsosought to examine the effect of dapsone, another inhibitor ofLPO/H2O2 and MPO/H2O2/Cl- systems (43-46), but its inhibi-tory capacity could not be precisely established because of itsown absorption in the analytical region of the spectrum.
EPR Measurements of Radicals from �2-Agonists. Phe-noxyl radicals are thought to be the primary metabolites of theenzymatic oxidation of phenols. Formation of these radicals hasbeen confirmed by EPR using tyrosine as a substrate (47, 48)and is consistent with the formation of tyrosine dimers throughradical-radical coupling. We thus sought to verify whetherperoxidative metabolism of phenolic �2-agonists also generatescorresponding radicals. When fenoterol was exposed to MPOand H2O2 in pH 7.0 buffer containing the spin trap MNP, anintense EPR spectrum attributed to a MNP adduct with thefenoterol-derived radical was detected (aN ) 1.49 mT) (Figure11A). In contrast, with salbutamol only a weak EPR signal ofa MNP adduct was observed after 15 min of reaction (Figure11B). No radicals were detected at pH 7.0 when MNP wasomitted in any sample.
The formation of radicals from agonists shows pH depen-dence. When fenoterol was exposed to MPO and H2O2 in pH 8buffer containing the spin trap MNP, the EPR spectrum shownin Figure 11C was observed. The spectrum contains contributionfrom two species: one is an MNP adduct, a triplet with aN )1.500 mT (labeled as o), and the other species characterized bya multiline EPR spectrum (labeled as *) has been attributed toa fenoterol-derived radical. The latter assignment was confirmedby an experiment conducted in the absence of MNP, in whichan EPR spectrum of the fenoterol radical alone was observed(Figure 11D). Initial simulations of this spectrum were per-formed assuming that the species is a meta-semiquinone formedvia the one-electron oxidation of the resorcinol moiety of thedrug. The characteristic feature of such a radical would bethe presence in its EPR spectrum of a triplet of doublets due to
Table 2. Oxidation of Fenoterol and Salbutamol by LPO/H2O2: Effect of Inhibitorsa
Amount of Metabolite Formed (∆A315 (%))
fenoterol salbutamol
control 100 100NaN3 (1 mM) 45.5 ( 3.9 17.4 ( 7.1NaCN (1 mM) 95.2 ( 5.3 80.0 ( 3.2NaSCN (0.1 mM) 22.8 ( 3.8 5.5 ( 7.3GSH (0.1 mM) 38.1 ( 3.2 42.7 ( 3.4methionine (0.1 mM) 103.7 ( 2.0 105.5 ( 16.2methionine (2 mM) 95.0 ( 3.5methimazole (20 µM) 30.0 ( 2.9 16.7 ( 4.6
a H2O2 was generated by glucose (1 mM) glucose oxidase (0.2 µg/mL). Oxidation of fenoterol (50 µM) and salbutamol (100 µM) wascarried out in 0.1 M potassium phosphate buffer (pH 7.0) containing 0.1mM DTPA in the presence of LPO (158 nM LPO for fenoterol and 216nM for salbutamol). The extent of inhibition was determined bymeasuring ∆A315 during 30 min of reaction and is expressed as % ofcontrol (mean ( SE from at least two determinations).
Figure 11. EPR spectra of radicals generated by the oxidation of �2-agonists by MPO and LPO systems. (A) Fenoterol (1.44 mM) oxidizedby MPO (0.19 U/mL) and H2O2 (113 µM) in pH 7.0 buffer containingMNP. The structure shown is that of a MNP/fenoterol radical adduct.(B) Salbutamol (2.28 mM) oxidized by MPO (0.38 U/mL) and H2O2
(113 µM) in pH 7.0 buffer containing MNP. (C) Fenoterol (2 mM)oxidized by MPO (0.4 U/mL) and H2O2 (99 µM) at pH 8.0 in thepresence of MNP. The spectrum is a composite of two spectra: of aMNP adduct (labeled o) and a fenoterol-derived semiquinone radical(labeled *). The dotted line is a simulated spectrum. (D) Observedspectrum of a radical derived from fenoterol in the absence of MNP.The dotted line is a simulated spectrum. (E) Oxidation of fenoterol (2mM) by LPO (0.425 µM) and H2O2 generated by glucose (1 mM) andglucose oxidase (4 µg/mL) at pH 8.0. (F) Oxidation of salbutamol (4.7mM) by MPO (0.94 U/mL) and H2O2 (0.43 mM) at pH 8.0. (G)Oxidation of salbutamol (2 mM) by MPO (0.4 U/mL) and H2O2 (99µM) at pH 8.0 in the presence of MNP. (H) Spectrum recorded whenMNP alone was exposed to MPO and H2O2 at pH 8.0. Buffers containedDTPA (0.2 mM) and, where indicated, MNP (10 mM in dimers).Instrumental settings: microwave power, 20 mW (41 mW in B);modulation amplitude, 0.1 mT (0.2 mT in B and F); receiver gain, 2 ×104 (A) and 2 × 105 (2.5 × 105 for E); time constant, 40.96 ms;conversion time, 40.96 ms; sweep time, 10 mT/41.94 s. Sample volumewas 250 µL. The intensity of original spectra A and C was decreasedby a factor 0.2 and spectrum E by 0.5. The spectra shown are theaverage of 5 scans and represent typical results.
Oxidation of �2-Agonists Chem. Res. Toxicol., Vol. 22, No. 6, 2009 1145
the interaction of the unpaired electron with two equivalentprotons (at C4,6) and one nonequivalent proton at C2 in thering (49). Although the spectrum calculated using this assump-tion reproduced well the positions of hyperfine components,amplitudes of the outermost lines did not match those of theexperimental spectrum (not shown). Also, the splitting on thetwo equivalent protons (0.67 mT) is smaller than that determinedfor meta-semiquinones derived from substituted resorcinols, 1.1mT and 0.99 mT (in alkaline and acidic solutions, respectively)(49). Finally, because of their reactivity, meta-semiquinonescannot be detected using static EPR, a technique that was usedin this study. These collective findings thus suggested that thespectrum detected is not that of the primary metabolite, (i.e., afenoterol meta-semiquinone) but rather of a secondary radicalproduct. A markedly better fit was obtained when the tripletcomponents were calculated assuming splitting on the nitrogenatom instead of on two equivalent protons. The dotted line inFigure 11D is a simulated spectrum calculated using thefollowing hfsc’s: 0.678 mT (1N), 0.190 mT (1H), 0.700 mT(1H), and 0.889 mT (1H) (correlation coefficient 0.974). Asimulated composite spectrum in Figure 11C (dotted line) fittingwell the experimental one (solid line) was obtained using hfsc’sfor the MNP adduct (1.495 mT (1N)) and for the fenoterolradical (0.698 mT (1N), 0.180 mT (1H), 0.6565 mT (1H), and0.903 mT (1H) (correlation coefficient 0.996)). Because thereis only one nitrogen atom in the molecule, in the side chain atC5 (Figure 1), the splitting on nitrogen is suggestive ofintramolecular cyclization through the amino nitrogen addingto the resorcine moiety to form a fused 5-membered ring. Noradicals were detected when MNP alone was exposed to MPOand H2O2 (Figure 11H).
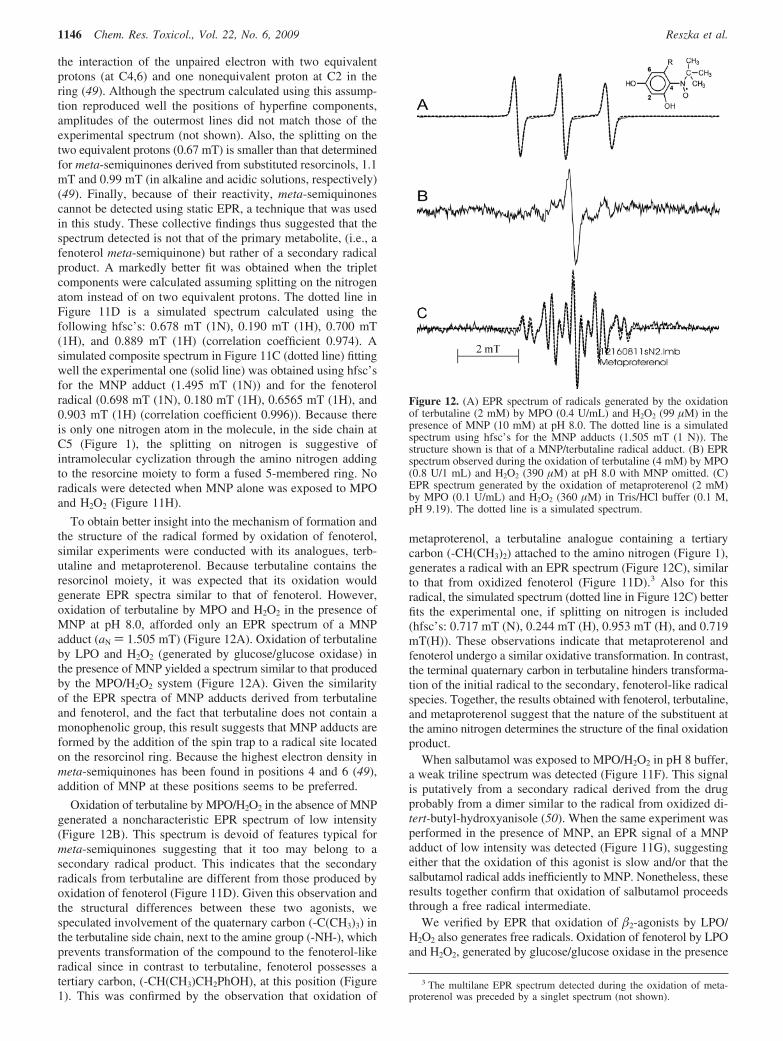
To obtain better insight into the mechanism of formation andthe structure of the radical formed by oxidation of fenoterol,similar experiments were conducted with its analogues, terb-utaline and metaproterenol. Because terbutaline contains theresorcinol moiety, it was expected that its oxidation wouldgenerate EPR spectra similar to that of fenoterol. However,oxidation of terbutaline by MPO and H2O2 in the presence ofMNP at pH 8.0, afforded only an EPR spectrum of a MNPadduct (aN ) 1.505 mT) (Figure 12A). Oxidation of terbutalineby LPO and H2O2 (generated by glucose/glucose oxidase) inthe presence of MNP yielded a spectrum similar to that producedby the MPO/H2O2 system (Figure 12A). Given the similarityof the EPR spectra of MNP adducts derived from terbutalineand fenoterol, and the fact that terbutaline does not contain amonophenolic group, this result suggests that MNP adducts areformed by the addition of the spin trap to a radical site locatedon the resorcinol ring. Because the highest electron density inmeta-semiquinones has been found in positions 4 and 6 (49),addition of MNP at these positions seems to be preferred.
Oxidation of terbutaline by MPO/H2O2 in the absence of MNPgenerated a noncharacteristic EPR spectrum of low intensity(Figure 12B). This spectrum is devoid of features typical formeta-semiquinones suggesting that it too may belong to asecondary radical product. This indicates that the secondaryradicals from terbutaline are different from those produced byoxidation of fenoterol (Figure 11D). Given this observation andthe structural differences between these two agonists, wespeculated involvement of the quaternary carbon (-C(CH3)3) inthe terbutaline side chain, next to the amine group (-NH-), whichprevents transformation of the compound to the fenoterol-likeradical since in contrast to terbutaline, fenoterol possesses atertiary carbon, (-CH(CH3)CH2PhOH), at this position (Figure1). This was confirmed by the observation that oxidation of
metaproterenol, a terbutaline analogue containing a tertiarycarbon (-CH(CH3)2) attached to the amino nitrogen (Figure 1),generates a radical with an EPR spectrum (Figure 12C), similarto that from oxidized fenoterol (Figure 11D).3 Also for thisradical, the simulated spectrum (dotted line in Figure 12C) betterfits the experimental one, if splitting on nitrogen is included(hfsc’s: 0.717 mT (N), 0.244 mT (H), 0.953 mT (H), and 0.719mT(H)). These observations indicate that metaproterenol andfenoterol undergo a similar oxidative transformation. In contrast,the terminal quaternary carbon in terbutaline hinders transforma-tion of the initial radical to the secondary, fenoterol-like radicalspecies. Together, the results obtained with fenoterol, terbutaline,and metaproterenol suggest that the nature of the substituent atthe amino nitrogen determines the structure of the final oxidationproduct.
When salbutamol was exposed to MPO/H2O2 in pH 8 buffer,a weak triline spectrum was detected (Figure 11F). This signalis putatively from a secondary radical derived from the drugprobably from a dimer similar to the radical from oxidized di-tert-butyl-hydroxyanisole (50). When the same experiment wasperformed in the presence of MNP, an EPR signal of a MNPadduct of low intensity was detected (Figure 11G), suggestingeither that the oxidation of this agonist is slow and/or that thesalbutamol radical adds inefficiently to MNP. Nonetheless, theseresults together confirm that oxidation of salbutamol proceedsthrough a free radical intermediate.
We verified by EPR that oxidation of �2-agonists by LPO/H2O2 also generates free radicals. Oxidation of fenoterol by LPOand H2O2, generated by glucose/glucose oxidase in the presence
3 The multilane EPR spectrum detected during the oxidation of meta-proterenol was preceded by a singlet spectrum (not shown).
Figure 12. (A) EPR spectrum of radicals generated by the oxidationof terbutaline (2 mM) by MPO (0.4 U/mL) and H2O2 (99 µM) in thepresence of MNP (10 mM) at pH 8.0. The dotted line is a simulatedspectrum using hfsc’s for the MNP adducts (1.505 mT (1 N)). Thestructure shown is that of a MNP/terbutaline radical adduct. (B) EPRspectrum observed during the oxidation of terbutaline (4 mM) by MPO(0.8 U/1 mL) and H2O2 (390 µM) at pH 8.0 with MNP omitted. (C)EPR spectrum generated by the oxidation of metaproterenol (2 mM)by MPO (0.1 U/mL) and H2O2 (360 µM) in Tris/HCl buffer (0.1 M,pH 9.19). The dotted line is a simulated spectrum.
1146 Chem. Res. Toxicol., Vol. 22, No. 6, 2009 Reszka et al.

of MNP, gives rise to both a MNP adduct (o) and the fenoterolradical (*) (Figure 11E). A well fitted simulated spectrum (dottedline) was obtained using hfsc’s for the fenoterol-derived radicalof 0.6836 mT (1N), 0.187 mT (1H), 0.9054 mT (1H), and0.6927 mT (1H), and for the triplet from the MNP adduct of1.495 mT (1N) (correlation coefficient 0.992), confirming thatLPO and MPO oxidize fenoterol to the same type of free radical.Also, oxidation of terbutaline by LPO/glucose/glucose oxidasein the presence of MNP afforded a MNP adduct whose EPRspectrum is similar to that generated by MPO/H2O2 (Figure12A).
Effect of Peroxidase Inhibitors on Free Radical Forma-tion: EPR Study. We investigated the effect of methimazoleand dapsone on radical formation from fenoterol and terbutalineby measuring EPR spectra of their MNP adducts. Table 3summarizes the results obtained at pH 8.0. Methimazole (0.86mM) and dapsone (1 mM) markedly inhibited the formation ofradicals from fenoterol and terbutaline oxidized by LPO/glucose/glucose oxidase (or H2O2 reagent). When the MPO system wasused, only methimazole was inhibitory. This is consistent withthe known inhibitory action of both methimazole and dapsoneon LPO activity and methimazole on MPO activity only (46).In contrast, dapsone has only a minimal effect on MPO activity.Results obtained at pH 7.0 qualitatively agree with those at pH8.0 (not shown).
Discussion
This study shows that �2-agonists salbutamol, fenoterol, andterbutaline undergo oxidation by MPO and LPO systems, whichmay be pertinent to their fate in asthmatic airways, for whichsignificant peroxidase activities have been reported (4-9). Thereaction was totally dependent on the simultaneous presenceof peroxidases, and H2O2 was abolished by catalase and wasinhibited by heme poisons (azide and cyanide) indicating thatoxidation of these �2-agonists requires an active peroxidase.Because these agonists contain phenolic groups, their metabo-lism seems to be best described by the process that is typicalfor peroxidative oxidation of phenols with phenoxyl radicalsas primary metabolites (eqs 1-3). Our observations are alsoconsistent with results of studies on oxidation of tyrosine andother phenolics by peroxidase systems (15, 17, 18, 23, 28).
For LPO, the effect of �2-agonists on the peroxidative cyclewas readily observed. Thus, while in the absence of the agonistsa small dose of H2O2 induced the formation of LPO-II lastingfor 14 min, in the presence of salbutamol (100 µM) the lifetimeof LPO-II was dramatically shortened, to less than 45 s (Figure9A,B). Only during the brief period during the formation anddecay of LPO-II was salbutamol oxidized, as evidenced by themomentary increase in A315 (Figure 9B). The fact that LPO-IIwas detected even in the presence of 1 mM salbutamol (i.e.,when [salbutamol] . [H2O2]) indicates that the interaction ofthe drug with LPO-II is the slowest step. In contrast tosalbutamol, fenoterol appeared to be a markedly more effectivesubstrate since in its presence, LPO-II did not accumulate indetectable quantities, suggesting that it was rapidly reduced toferric LPO (Figure 9C).4
Oxidation of salbutamol by LPO was also observed whenH2O2 was generated by glucose/glucose oxidase. In this system,the rate of H2O2 generation could be regulated by the amountof glucose oxidase applied and could be adjusted so that allH2O2 produced, was immediately used up for oxidation of thesubstrate, without apparent accumulation of LPO-II.
The formation of the agonist-derived free radicals during theiroxidation by peroxidases has been verified by EPR and spintrapping with MNP, and by their stimulatory action on peroxi-dative oxidation of glutathione and ascorbate, a process knownto be catalyzed by phenolics. The putative stable products ofthe oxidation of �2-agonists are, among other possible products,dimers formed by recombination of phenoxyl radicals. Thistransformation may be especially pertinent to salbutamol, asthis agent possesses one phenolic moiety, and its metabolismseems to resemble that of tyrosine, known to produce o,o′-dityrosine dimers upon enzymatic oxidation. It has to beemphasized, however, that in contrast to tyrosine, the formationof salbutamol dimers may be restricted by the presence of asubstituent (-CH2OH) in one ortho-position in the drug’saromatic ring, which decreases the number of possible radicalsites that can participate in recombination.
Fenoterol is unique among the �-agonists investigated in thatit possesses both the mono- and 1,3-dihydroxybenzene moietiesand may thus be subjected to peroxidative metabolism charac-teristic of a simple phenolic and that of meta-hydroquinones.Using terbutaline as a model compound, we found that whenoxidized it forms adducts with MNP whose EPR spectra arevery similar to those generated by the oxidation of fenoterol.On the basis of this result, we conclude that MNP traps theradical located on resorcinol ring, most likely by adding at C4or C6, which are the sites of the highest spin density. Formationof MNP adducts with the radical from the fenoterol monophe-nolic group was not detected, but if formed, their contributionmight be small, judging by the low intensity of EPR spectragenerated by oxidation of salbutamol. It has been reported thatalkyl resorcinols undergo oxidation by peroxidases and formdimeric products through the addition of the correspondingphenoxyl radicals (28). Thus, theoretically, peroxidative me-tabolism of fenoterol may lead to dimers through recombinationof two types of phenoxyl radicals: (1) those derived from themonophenolic moiety (to form dityrosine type dimers) and (2)those derived from meta-semiquinone-type radicals (to formdiresorcinols). Formation of mixed type dimers may also bepossible.
We found that AscH- and GSH at concentrations approx-imating those in airway fluids (∼100 µM) (30) inhibit oxidation
4 See footnote 2.
Table 3. Effect of Dapsone and Methimazole on EPRSpectra of MNP Adducts with Radicals from Fenoterol and
Terbutaline Generated by MPO/H2O2 and LPO/H2O2
Systems at pH 8.0a
+ MPO + H2O2 + LPO + H2O2
Fenoterol-no additives 100b 100c
+ dapsone (1 mM) d 13.9 ( 0.6b
+ methimazole (0.86 mM) 22.4 ( 0.6b 26.9 ( 8.2c
Terbutaline
- no additives 100b 100c
+ dapsone (1 mM) d 54.6 ( 20.0b
+ methimazole (0.86 mM) 33.5 ( 3.5b 33.9c
a Concentrations of MNP spin adducts were calculated as secondintegrals of the low-field component of the respective spectra and areexpressed as % of control samples (no additives). Mean ( SE of at leasttwo measurements. In experiments with methimazole, concentrations offenoterol and terbutaline were 0.47 mM and in experiments withdapsone 0.047 mM. The activity of MPO was 500 mU/1 mL. Theconcentration of LPO was 72 nM. b The oxidant was the reagent H2O2
(37 µM). c H2O2 was generated using glucose (1 mM) and glucoseoxidase (0.8 µg/mL). d Not determined as dapsone does not inhibitMPO.
Oxidation of �2-Agonists Chem. Res. Toxicol., Vol. 22, No. 6, 2009 1147

of �2-agonists. During this reaction, AscH- and GSH areoxidized to their respective radicals as documented by our EPRdata. Absorption spectra measured during the oxidation offenoterol in the presence of albumin indicated the formation ofan albumin-agonist conjugate. A similar conjugate was prob-ably formed when fenoterol and terbutaline were oxidized inthe presence of GSH as judged by the appearance of charac-teristic absorption bands at 395 and 360 nm, respectively, thatwere not observed when GSH was omitted. Formation of suchconjugates with thiols has been described for other quinonoidcompounds (34, 35). Thus, GSH plays a dual role during theoxidation of polyphenols. First, it functions as a reducing agent,converting semiquinones back to hydroquinones, and second,it may act as an alkylating agent (Scheme 1B). It is thereforepossible that if the oxidative metabolism of �2-agonists occursin ViVo it may contribute to the depletion of cellular antioxidantsand promote oxidative injury in tissues.
We found that oxidation of fenoterol and metaproterenol, butnot terbutaline, afforded radicals that could be detected by directEPR. Spectral analysis of these radicals suggested that theseare not primary semiquinones formed by one-electron oxidationof resorcinol moieties in fenoterol and metaproterenol but rathersecondary radicals formed by oxidation of as yet not identifiedtransient products derived from these drugs. Because the bestfitted simulated spectra were obtained including splitting on thenitrogen atom, it was tentatively assumed that the new radicalwas formed by the amino nitrogen in the side chain at C5 addingto the resorcinol ring in oxidized fenoterol and metaproterenol,forming a fused 5-member ring, followed by further oxidation.Such a transformation is known to occur for catecholamines,in which the two hydroxy groups are in ortho-position and canform electrophilic ortho-quinone. Meta-hydroquinones cannotform quinones, although the formation of biradicals having ameta-quinone character has been considered (51). However, ifformed, such a molecule would be highly electrophilic and afterhydrolysis might produce a 2-hydroxy-1,4-benzoquinone-typechromophore, for which intramolecular cyclization would bepossible. Formation of 2-hydroxy-1,4-benzoquinone from re-sorcinol has been reported to occur through enzymatic conver-sion (52, 53) and reaction of the resorcinol semiquinone withoxygen (29). The observation that the rate and the extent ofoxidation of fenoterol depend on oxygen (they both decreasein deaerated solutions) partially supports the latter mechanism.Hydroxylation of phenolics by MPO compound III has also beenreported (54). Thus, hydroxylation of the resorcinol ring infenoterol, terbutaline, and metaproterenol through an enzymaticor chemical process seems feasible. Oxidation of terbutalineprobably did not lead to the corresponding fenoterol-type radicalor the reaction was inefficient, possibly because the bulky natureof the substituent on the amino nitrogen (quaternary carbon)prevented the cyclization.
It can be hypothesized that structural modifications of�-agonists catalyzed by peroxidases, such as the formation ofdimers, cyclization, or the formation of conjugates with GSHand albumin may impact their therapeutic activity, which isdependent on their interaction with �2-adrenergic receptors onsmooth muscle cells, because their affinity to these receptorswill be altered, presumably diminished. Because of this, it wasimportant to find out whether oxidation of �2-agonists byperoxidases can be prevented or at least inhibited. For thispurpose, we investigate two compounds, methimazole anddapsone. Methimazole acts as a suicidal inhibitor of thyroidperoxidase and LPO (40, 55) and is used in the treatment ofhyperthyroid conditions. Methimazole is oxidized by MPO to
a free radical product without inactivation of the enzyme (56).Dapsone, an anti-inflammatory and an antileprotic drug, wasused as a corticosteroid sparing agent in the treatment of asthma(57). Dapsone inhibits LPO activity (46) but is neutral withrespect to MPO unless chloride is also a substrate (43). Ourspectrophotometric measurements revealed that methimazoleinhibits oxidation of salbutamol and fenoterol by the LPOsystem. This was further supported by EPR studies, whichshowed that methimazole and dapsone inhibit the generationof free radicals from fenoterol and terbutaline by LPO/H2O2,thus preventing their transformation to products with alteredreactivity. Methimazole also effectively inhibited oxidation byMPO, whereas dapsone did not. However, dapsone may havethe potential to inhibit modifications such as chlorination andnitration to which �-agonists may also be subject. Moreover,these results suggest the intriguing possibility that peroxidaseinhibitors may have the potential to prevent or minimize thedegradation of �-agonists in asthmatic airways.
In summary, we have found that salbutamol and fenoterolare metabolized by endogenous airway peroxidases, (MPO andLPO), to free radicals, which potentially give rise to structurallymodified products. Salbutamol and fenoterol differ markedlyin their capacity to undergo oxidation by the peroxidases aswell as in the reactivity of their resulting metabolites, withfenoterol being more susceptible to oxidation and affording morereactive intermediates. Because the therapeutic activity of thesedrugs depends on their binding to �2-adrenergic receptors onsmooth muscle cells, structural modifications resulting fromoxidation are likely to inhibit the interaction with their cognatereceptor, resulting in diminished therapeutic efficacy. Peroxidasemediated oxidation and consequent drug inactivation, particularwhen airway inflammation is severe, may thus be a mechanismthat renders patients refractory or resistant to the bronchodilatoryeffects of �2-agonists.This oxidative transformation, however,can be inhibited by physiological antioxidants/substrates (ascor-bate, glutathione, and thiocyanate) and pharmacological inhibi-tors of peroxidase (methimazole and dapsone), suggesting thatsuch agents may offer the means to prevent and/or enhance thetherapeutic activity or other functions of the agonists in clinicalsettings.
Acknowledgment. This study was supported by a grant fromNIH (HL072068 to D.W.M.).
Note Added after Print Publication. The print publicationof June 15, 2009 (Vol. 22, Issue 6), contained an incompleteversion of Figure 1. Figure 1 was corrected in the web versionon July 7, 2009. An Addition and Correction also appears inthe August 17, 2009 (Vol. 22, Issue 8), print issue.
References
(1) Spitzer, W. O., Suissa, S., Ernst, P., Horwitz, R. I., Habbick, B.,Cockcroft, D., Boivin, J. F., McNutt, M., Buist, A. S., and Rebuck,A. S. (1992) The use of beta-agonists and the risk of death and neardeath from asthma. N. Engl. J. Med. 326, 501–506.
(2) Suissa, S., Ernst, P., Boivin, J. F., Horwitz, R. I., Habbick, B., Cockroft,D., Blais, L., McNutt, M., Buist, A. S., and Spitzer, W. O. (1994) Acohort analysis of excess mortality in asthma and the use of inhaledbeta-agonists. Am. J. Respir. Crit. Care Med. 149 (3 Pt 1), 604–610.
(3) Abramson, M. J., Walters, J., and Walters, E. H. (2003) Adverse effectsof beta-agonists: are they clinically relevant? Am. J. Respir. Med. 2(4), 287–297.
(4) Aldridge, R. E., Chan, T., van Dalen, C. J., Senthilmohan, R., Winn,M., Venge, P., Town, G. I., and Kettle, A. J. (2002) Eosinophilperoxidase produces hypobromous acid in the airways of stableasthmatics. Free Radical Biol. Med. 33, 847–856.
1148 Chem. Res. Toxicol., Vol. 22, No. 6, 2009 Reszka et al.

(5) Duguet, A., Iijima, H., Eum, S. Y., Hamid, Q., and Eidelman, D. H.(2001) Eosinophil peroxidase mediates protein nitration in allergicairway inflammation in mice. Am. J. Respir. Crit. Care Med. 164,1119–1126.
(6) Wu, W. J., Samoszuk, M. K., Comhair, S. A. A., Thomassen, M. J.,Farver, C. F., Dweik, R. A., Kavuru, M. S., Erzurum, S. C., and Hazen,S. L. (2000) Eosinophils generate brominating oxidants in allergen-induced asthma. J. Clin. InVest. 105, 1455–1463.
(7) Andreadis, A. A., Hazen, S. L., Comhair, S. A. A., and Erzurum, S. C.(2003) Oxidative and nitrosative events in asthma. Free Radical Biol.Med. 35 (3), 213–225.
(8) Wardlaw, A. J., Dunnette, S., Gleich, G. J., Collins, J. V., and Kay,A. B. (1988) Eosinophils and mast cells in bronchoalveolar lavagefluid in subjects with mild asthma. Am. ReV. Respir. Dis. 137, 62–69.
(9) Bousquet, J., Jeffery, P. K., Busse, W., Johnson, M., and Vinola, A. M.(2000) Asthma: from bronchoconstriction to airway inflammation andremodeling. Am. J. Respir. Crit. Care Med. 161, 1720–1745.
(10) Yasui, K., Kobayashi, N., Yamazaki, T., Agematsu, K., Matsuzaki,S., Nakata, S., and Baba, A. (2006) Differential effects of short-actingbeta2-agonists on human granulocyte functions. Int. Arch. AllergyImmunol. 139 (1), 1–8.
(11) Tachibana, A., Kato, M., Kimura, H., Fujiu, T., Suzuki, M., andMorikawa, A. (2002) Inhibition by fenoterol of human eosinophilfunctions including beta2-adrenoceptor-independent actions. Clin. Exp.Immunol. 130 (3), 415–23.
(12) Gillissen, A., Jaworska, M., Scharling, B., van Zwoll, D., and Schultze-Werninghaus, G. (1997) Beta-2 agonists have antioxidant function invitro. 1. Inhibition of superoxide anion, hydrogen peroxide, hypochlo-rous acid and hydroxyl radical. Respiration 64, 16–22.
(13) Gillissen, A., Wickenburg, D., van Zwoll, D., and Schultze-Werning-haus, G. (1997) Beta-2-agonists have antioxidant function in vitro. 2.The effect of beta-2-agonists on oxidant mediated cytotoxicity andon superoxide anion generated by human polymorphonuclear leuko-cytes. Respiration 64, 23–28.
(14) Sawahata, T., and Neal, R. A. (1982) Horseradish peroxidase-mediatedoxidation of phenol. Biochem. Biophys. Res. Commun. 109 (3), 988–994.
(15) Yu, J., Taylor, K. E., Zou, H., Biswas, N., and Bewtra, J. K. (1994)Phenol conversion and dimeric intermediates in horseradish peroxidase-catalyzed phenol removal from water. EnViron. Sci. Technol. 28, 2154–2160.
(16) Bayse, G. S., Michaelis, A. W., and Morrison, M. (1972) Theperoxidase catalyzed oxidation of tyrosine. Biochim. Biophys. Acta284, 34–42.
(17) Marquez, L. A., and Dunford, H. B. (1995) Kinetics of oxidation oftyrosine and dityrosine by myeloperoxidase compounds I and II.J. Biol. Chem. 270 (51), 30434–30440.
(18) Heinecke, J. W., Li, W., Daehnke, H. L., III, and Goldstein, J. A.(1993) Dityrosine, a specific marker of oxidation, is synthesized bythe myeloproxidase-hydrogen peroxide system of human neutrophilsand macrophages. J. Biol. Chem. 268 (6), 4069–4077.
(19) Heinecke, J. W., Li, W., Francis, G. A., and Goldstein, J. A. (1993)Tyrosyl radical generated by myeloperoxidase catalyzes the oxidativecross-linking of proteins. J. Clin. InVest. 91, 2866–2872.
(20) Tien, M. (1999) Myeloperoxidase-catalyzed oxidation of tyrosine.Arch. Biochem. Biophys. 367 (1), 61–66.
(21) Metodiewa, D., Reszka, K., and Dunford, H. B. (1989) Oxidation ofthe substituted catechols dihydroxyphenylalanine methyl ester andtrihydroxyphenylalanine by lactoperoxidase and its compounds. Arch.Biochem. Biophys. 274 (2), 601–608.
(22) Metodiewa, D., Reszka, K., and Dunford, H. B. (1989) Evidence fora peroxidatic oxidation of norepinephrine, a catecholamine, bylactoperoxidase. Biochem. Biophys. Res. Commun. 160 (3), 1183–1188.
(23) Monzani, E., Gatti, A. L., Profumo, A., Casella, L., and Gullotti, M.(1918) (1997) Oxidation of phenolic compounds by lactoperoxidase.Evidence for the presence of a low-potential compound II duringcatalytic turnover. Biochemistry 36, 1926.
(24) Jenzer, H., Jones, W., and Kohler, H. (1986) On the molecularmechanism of lactoperoxidase-catalyzed H2O2 metabolism and ir-reversible enzyme inactivation. J. Biol. Chem. 261 (33), 15550–15556.
(25) Nelson, D. P., and Kiesow, L. A. (1972) Enthalpy of decomposition ofhydrogen peroxide by catalase at 25°C (with molar extinction coefficientsof H2O2 solution in the UV). Anal. Biochem. 49, 474–478.
(26) Kalyanaraman, B., Felix, C. C., and Sealy, R. C. (1982) Photoionizationof melanin precursors: an electron spin resonance investigation usingthe spin trap 5,5-dimethyl-1-pyrroline-1-oxide (DMPO). Photochem.Photobiol. 36 (1), 5–12.
(27) Childs, R. E., and Bardsley, W. G. (1975) The steady-state kineticsof peroxidase with 2,2′-azino-di-(3-ethyl-benzthiazoline-6-sulphonicacid) as chromogen. Biochem. J. 145, 93–103.
(28) Divi, R. L., and Doerge, D. R. (1994) Mechanism-based inactivationof lactoperoxidase and thyroid peroxidase by resorcinol derivatives.Biochemistry 33, 9668–9674.
(29) Perbet, G., Filiol, C., Boule, P., and Lemaire, J. (1979) Photolyse etphoto-oxydation des diphenols en solution aqueous diluee. J. Chim.Phys. 76 (1), 89–96.
(30) Cross, C. E., van der Vliet, A., O’Neill, C. A., Louie, S., and Halliwell,B. (1994) Oxidants, antioxidants, and respiratory tract lining fluids.EnViron. Health Perspect. 102 (Suppl 10), 185–191.
(31) Sturgeon, B. E., Jr., Barr, D. P., Corbett, J. T., Martinez, J. G., andMason, R. P. (1998) The fate of the oxidizing tyrosyl radical in thepresence of glutathione and ascorbate. Implications for the radical sinkhypothesis. J. Biol. Chem. 273 (46), 30116–30121.
(32) Harman, L. S., Carver, D. K., Schreiber, J., and Mason, R. P. (1986)One- and two-electron oxidation of reduced glutathione by peroxidases.J. Biol. Chem. 261 (4), 1642–1648.
(33) Reszka, K. J., Matuszak, Z., Chignell, C. F., and Dillon, J. (1999)Oxidation of biological electron donors and antioxidants by a reactivelactoperoxidase metabolite from nitrite (NO2
-): an EPR and spintrapping study. Free Radical Biol. Med. 26 (5-6), 669–678.
(34) Takahashi, N., Schreiber, J., Fischer, V., and Mason, R. P. (1987)Formation of glutathione-conjugated semiquinones by the reaction ofquinines with glutathione: an ESR study. Arch. Biochem. Biophys.252 (1), 41–48.
(35) Rao, D. N. R., Takahashi, N., and Mason, R. P. (1988) Characterizationof a glutathione conjugate of 1,4benzosemiquinone-free radical formedin rat hepatocytes. J. Biol. Chem. 263 (34), 17981–17986.
(36) Yamazaki, I., Tamura, M., Nakajima, R., and Nakamura, M. (1985)Physiological aspects of free-radical reactions. EnViron. HealthPerspect. 64, 331–342.
(37) Kohler, H., Taurog, A., and Dunford, H. B. (1988) Spectral studieswith lactoperoxidase and thyroid peroxidase: interconversions betweennative enzyme, compound II, and compound III. Arch. Biochem.Biophys. 264 (2), 438–449.
(38) Metodiewa, D., Marquez, L. A., and Dunford, H. B. (1991) The activityof mammalian peroxidases (lactoperoxidase and myeloperoxidase) andtheir compounds III toward 2-t-butyl-4-methoxyphenol (butylatedhydroxyanisole) and its dimer (2,2′-dihydroxy-3,3′-di-t-butyl-5,5′-dimethoxydiphenyl). Biochem. Int. 23 (2), 281–290.
(39) Davidson, B., Soodak, M., Neary, J. T., Strout, H. V., Kieffer, J. D.,Mover, H., and Maloof, F. (1978) The irreversible inactivation ofthyroid peroxidase by methylmercaptoimidazole, thiouracil, and pro-pylthiouracil in vitro and its relationship to in vivo findings. Endo-crinology 103 (3), 871–882.
(40) Ohtaki, S., Nakagawa, H., Nakamura, M., and Yamazaki, I. (1982)Reactions of purified hog thyroid peroxidase with H2O2, tyrosine, andmethylmercaptoimidazole (goitrogen) in comparison with bovinelactoperoxidase. J. Biol. Chem. 257 (2), 761–766.
(41) Michot, J. L., Nunez, J., Johnson, M. L., Irace, G., and Edelhoch, H.(1979) Iodide binding and regulation of lactoperoxidase activity towardgoitrogens. J. Biol. Chem. 254, 2205–2209.
(42) Edelhoch, H., Irace, G., Johnson, M. L., Michot, J. L., and Nunez, J.(1979) The effects of thioureylene compounds (goitrogens) onlactoperoxidase activity. J. Biol. Chem. 254, 11822–11830.
(43) van Zyl, J. M., Basson, K., Kriegler, A., and van der Walt, B. J. (1991)Mechanisms by which clofazimine and dapsone inhibit the myelop-eroxidase system. A possible correlation with their anti-inflammatoryproperties. Biochem. Pharmacol. 42 (3), 599–608.
(44) Kettle, A. J., and Winterbourn, C. C. (1991) Mechanism of inhibitionof myeloperoxidase by anti-inflammatory drugs. Biochem. Pharmacol.41 (10), 1485–1492.
(45) Bozeman, P. M., Learn, D. B., and Thomas, E. L. (1992) Inhibitionof the human leukocyte enzymes myeloperoxidase and eosinophilperoxidase by dapsone. Biochem. Pharmacol. 44 (2), 553–563.
(46) Thomas, E. L., Jefferson, M. M., Joyner, R. E., Cook, G. S., and King,C. C. (1994) Leukocyte myeloperoxidase and salivary lactoperoxidase:identification and quantitation in human mixed saliva. J. Dent. Res.73 (2), 544–55.
(47) McCormick, M. L., Gaut, J. P., Lin, T. S., Britigan, B. E., Buettner,G. R., and Heinecke, J. W. (1998) Electron paramagnetic resonancedetection of free tyrosyl radical generated by myeloperoxidase,lactoperoxidase, and horseradish peroxidase. J. Biol. Chem. 273 (48),32030–32037.
(48) Gunther, M. R., Tschirret-Guth, R. A., Witkowska, H. E., Fann, Y. C.,Barr, D. P., Ortiz De Montellano, P. R., and Mason, R. P. (1998)Site-specific spin trapping of tyrosine radicals in the oxidation ofmetmyoglobin by hydrogen peroxide. Biochem. J. 330 (Pt. 3), 1293–1299.
(49) Stone, T. J., and Waters, W. A. (1964) Aryloxy-radicals. Part III.Electron spin resonance spectra of radicals from some substitutedresorcinols. J. Chem. Soc. 4302–4307.
(50) Valoti, M., Jr., Sgaragli, G., and Mason, R. P. (1989) Free radicalintermediates during peroxidase oxidation of 2-t-butyl-4-methoxyphe-nol, 2,6-di-t-butyl-4-methylphenol, and related phenol compounds.Arch. Biochem. Biophys. 269 (2), 423–432.
Oxidation of �2-Agonists Chem. Res. Toxicol., Vol. 22, No. 6, 2009 1149

(51) Roithova, J., Schroder, D., and Schwartz, H. (2005) Generation ofthe elusive meta-benzoquinone in gas phase. Angew. Chem., Int. Ed.117, 3152–3156.
(52) Philipp, B., and Schink, B. (1998) Evidence of two oxidative reactionsteps initiating anaerobic degradation of resorcinol (1,3-dihydroxy-benzene) by the denitrifying bacterium Azoarcus anaerobius. J.Bacteriol. 180 (14), 3644–3649.
(53) Gallus, C., and Schink, B. (1998) Anaerobic degradation of a-resorcylate by Thaurea aromatica strain AR-1 prceeds via oxidationand decarboxylation to hydroxyhydroquinone. Arch. Microbiol. 169,333–338.
(54) Kettle, A. J., and Winterbourn, C. C. (1994) Superoxide-dependenthydroxylation by myeloperoxidase. J. Biol. Chem. 269 (25), 17146–17151.
(55) Bandyopadhyay, U., Bhattacharyya, D. Kr., Chatterjee, R., andBanerjee, R. K. (1995) Irreversible inactivation of lactoperoxidase bymercaptomethylimidazole through generation of a thiyl radical: its useas a probe to study the active site. Biochem. J. 306, 751–757.
(56) McGirr, L. G., Jatoe, S. D., and O’Brien, P. J. (1990) Myeloperoxidasecatalysed cooxidative metabolism of methimazole: oxidation ofglutathione and NADH by free radical intermediates. Chem.-Biol.Interact. 73 (2-3), 279–295.
(57) Dewey, A., Bara, A., Dean, T., and Walters, H. (2002) Dapsone asan oral corticosteroid sparing agent for asthma. Cochrane DatabaseSyst. ReV. 4, CD003268, 1-8.
TX900071F
1150 Chem. Res. Toxicol., Vol. 22, No. 6, 2009 Reszka et al.





























