Embed Size (px)
Citation preview

Determinants in V2C2 region of HIV-1 clade C primary envelopesconferred altered neutralization susceptibilities to IgG1b12 andPG9 monoclonal antibodies in a context-dependent manner
Shilpa Patil a,1, Ipsita Choudhary b,1, Nakul K. Chaudhary a, Rajesh Ringe b,2, Manish Bansal a,Brihaspati Narayan Shukla a, Saikat Boliar a, Bimal K. Chakrabarti a, Jayanta Bhattacharya a,n
a HIV Vaccine Translational Research Laboratory, THSTI-IAVI HIV Vaccine Design Program, Translational Health Science and Technology Institute,450, Udyog Vihar, Phase-III, Gurgaon 122016, Haryana, Indiab National AIDS Research Institute, Pune, Maharashtra, India
a r t i c l e i n f o
Article history:Received 31 March 2014Returned to author for revisions14 April 2014Accepted 16 June 2014
Keywords:HIV-1Clade CEnvelopeNeutralizing antibodyCD4bsIgG1b12VRC01PG9PG16
a b s t r a c t
In the present study by examining pseudoviruses expressing patient chimeric envelopes (Envs) madebetween an IgG1b12 (b12)-sensitive (2-5.J3) and a b12-resistant (4.J22) HIV-1 clade C envelope, weidentified determinants in the V2C2 region that governed susceptibility to b12 monoclonal antibody, butnot to other CD4 binding site antibodies. Interestingly, when the V2C2 sequence of the 2-5.J3 Env wastransferred to other b12-resistant primary clade C Envs, their susceptibility to b12 varied, indicating thatthis effect was context dependent. In addition, we identified determinants within the V2 region in theb12-resistant envelope that significantly modulated the neutralization of Env-pseudotyped virusesto PG9/PG16 MAbs. The enhanced neutralization susceptibilities of Envs to b12 and PG9 MAbs werecorrelated with increased exposure of their corresponding epitopes highlighting vulnerabilities in theV2C2 region that altered Env conformation necessary for the efficient accessibility of b12 and PG9antibodies.
& 2014 Elsevier Inc. All rights reserved.
Introduction
The entry of the Human Immunodeficiency Virus Type 1 (HIV-1)into the target cells depends primarily on the interaction of gp120 oftrimeric envelope (Env) glycoprotein (gp160) with CD4 and subse-quently with a coreceptor, usually CCR5 or CXCR4, which facilitatesfusion of viral membrane to cell membrane. In recent years, anumber of broad and potent neutralizing human monoclonalantibodies were isolated from slow progressing individuals(Huang et al., 2012; Walker et al., 2011; Walker et al., 2009; Wuet al., 2010) with unique specificities which has provided clues ofseveral vulnerabilities associated with immune evasion by HIV-1.The CD4 binding sites (CD4bs) and quaternary epitopes (QNE) onthe HIV-1 Env trimers are important targets for vaccine develop-ment. b12 is the first identified neutralizing monoclonal antibody(MAb) (Burton et al., 1994) whose epitopes in HIV-1 Env overlap
the CD4 binding domain of gp120 (Burton et al., 1994; Mo et al.,1997). The b12-mediated neutralization has been shown to beinfluenced by the mutations in V2 and C3 of gp120 (Mo et al.,1997). Additionally, altered conformation of Env due to subtlemutation in variable loops changed the sensitivity of neutraliza-tion of virus to b12 (Duenas-Decamp et al., 2008; Ly andStamatatos, 2000; Utachee et al., 2010). By alanine scanningmutagenesis, Pantophlet et al. (2003) demonstrated several resi-dues in gp120 that are important for gp120 recognition byneutralizing and non-neutralizing MAbs to CD4bs. Otherstructure-based studies also demonstrated distinct vulnerablesites in gp120 conferring b12 susceptibility and/or resistance(Chen et al., 2009; Liu et al., 2008; Wu et al., 2009; Zhou et al.,2007). In addition, in monkey models, using SHIV-HXBc2 andSHIV-89.6P viruses, specific amino acid changes in V2 region(SHIV-89.6P) and V1, V2, V3, and gp41 regions (SHIV-HXBc2) thatwere associated with b12 resistance were documented (Etemad-Moghadam et al., 1999; Si et al., 2001). Recently, we have alsodemonstrated a single amino acid substitution at the 681 positionin the gp41 membrane proximal external region (MPER) enhancedbinding and neutralization of HIV-1 clade A, B and CRF02-AG-235envelopes to CD4-Ig and b12 MAb, indicating gp41 as referred
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/yviro
Virology
http://dx.doi.org/10.1016/j.virol.2014.06.0180042-6822/& 2014 Elsevier Inc. All rights reserved.
n Corresponding author.E-mail addresses: [email protected],
[email protected] (J. Bhattacharya).1 Equal contribution.2 Current address: Weill Cornell Medical College, New York, USA.
Virology 462-463 (2014) 266–272
above can also modulate HIV-1 to b12 susceptibility (Ringe andBhattacharya, 2012). Very recently Utachee et al. (2014) demon-strated that the presence of aspartic acid at the position 185 in V2loop confers HIV-1A/E subtype envelopes with enhanced suscept-ibility to b12 monoclonal antibodies and that this effect was foundto be not dependent on glycan residues present in 186 and 197positions previously described to be regulating envelope suscept-ibility to b12 (Utachee et al., 2010). On the other hand, the variableloops particularly V1, V2 and V3 regions have also been shown tobe important for the recognition of recently discovered broad andpotent MAbs, such as PG9, PG16, PGT121 and PGT145 (Doores andBurton, 2010; Moore et al., 2011; Ringe et al., 2012; Thenin et al.,2012; Walker et al., 2011; Walker et al., 2009). Interestingly, HIV-1clade C has been widely shown to be refractory to neutralizationby b12 MAb in different geographic settings (Gray et al., 2006;Kulkarni et al., 2009; Ringe et al., 2010; Zhang et al., 2010).
In natural infection, the antibodies usually develop within the firstfew weeks of infection (Moore et al., 1994; Tomaras et al., 2008).However, the antibodies which neutralize autologous viruses werefound to be elicited after several weeks of initial infection (Montefioriet al., 2007) and have been demonstrated to exert immune selectionpressure on autologous viral variants (Bunnik et al., 2008; Frost et al.,2005; Gray et al., 2007; Li et al., 2006; Richman et al., 2003; Wei et al.,2003). It has been demonstrated earlier that HIV-1 variants thatbecome resistant to autologous neutralizing antibodies in the courseof infection by virtue of their escape strategies developed differentsusceptibilities to monoclonal antibodies and/or heterologous serumor plasma antibodies (Mascola, 2009; Mascola and Montefiori, 2010).We previously reported that env clones obtained at different timepoints from an Indian patient (NARI-IVC2) with early infection wasunusually sensitive to b12, an MAb that targets CD4 binding site(CD4bs) on HIV-1 Env (Ringe et al., 2010). This observation promptedus to investigate the determinants in Env responsible for enhancedsensitivity of these Envs to b12. Since, pseudoviruses expressing all theenvs obtained from the NARI-IVC2 patient at different time points
showed comparable sensitivity to b12, we examined the b12-sensitiveEnv, 2-5.J3 with that of 4.J22, also a clade C but b12-resistant Envobtained from a different patient (NARI-IVC4) towards mappingdeterminants responsible for altered sensitivity to b12. Constructingthe chimeric Envs taking both sensitive (2-5.J3) and resistant (4.J22)isolates and subsequently by fine mapping, we identified amino acidresidues in V2C2 regions that conferred reciprocal susceptibilities tob12 and PG9/16 MAbs.
Results and discussion
V2C2 region modulated susceptibility of primary patient-derived Envsto b12 MAb
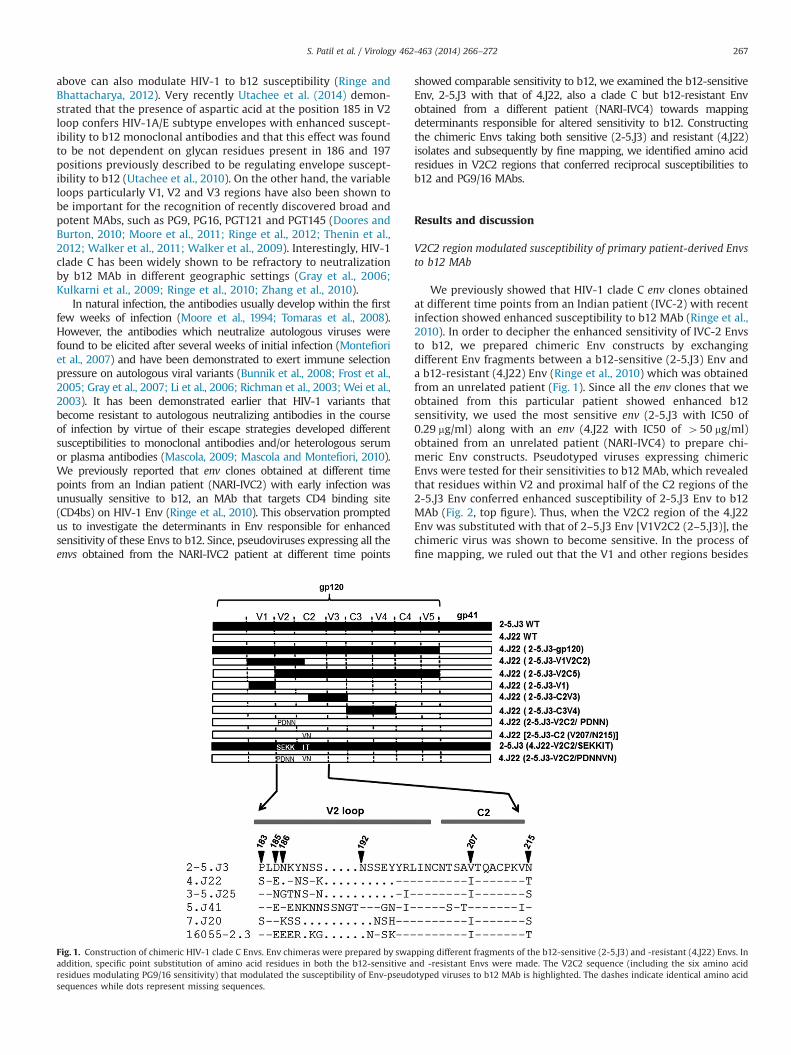
We previously showed that HIV-1 clade C env clones obtainedat different time points from an Indian patient (IVC-2) with recentinfection showed enhanced susceptibility to b12 MAb (Ringe et al.,2010). In order to decipher the enhanced sensitivity of IVC-2 Envsto b12, we prepared chimeric Env constructs by exchangingdifferent Env fragments between a b12-sensitive (2-5.J3) Env anda b12-resistant (4.J22) Env (Ringe et al., 2010) which was obtainedfrom an unrelated patient (Fig. 1). Since all the env clones that weobtained from this particular patient showed enhanced b12sensitivity, we used the most sensitive env (2-5.J3 with IC50 of0.29 mg/ml) along with an env (4.J22 with IC50 of 450 mg/ml)obtained from an unrelated patient (NARI-IVC4) to prepare chi-meric Env constructs. Pseudotyped viruses expressing chimericEnvs were tested for their sensitivities to b12 MAb, which revealedthat residues within V2 and proximal half of the C2 regions of the2-5.J3 Env conferred enhanced susceptibility of 2-5.J3 Env to b12MAb (Fig. 2, top figure). Thus, when the V2C2 region of the 4.J22Env was substituted with that of 2–5.J3 Env [V1V2C2 (2–5.J3)], thechimeric virus was shown to become sensitive. In the process offine mapping, we ruled out that the V1 and other regions besides
Fig. 1. Construction of chimeric HIV-1 clade C Envs. Env chimeras were prepared by swapping different fragments of the b12-sensitive (2-5.J3) and -resistant (4.J22) Envs. Inaddition, specific point substitution of amino acid residues in both the b12-sensitive and -resistant Envs were made. The V2C2 sequence (including the six amino acidresidues modulating PG9/16 sensitivity) that modulated the susceptibility of Env-pseudotyped viruses to b12 MAb is highlighted. The dashes indicate identical amino acidsequences while dots represent missing sequences.
S. Patil et al. / Virology 462-463 (2014) 266–272 267

V2C2 region (Fig. 1) played any role in altering b12 sensitivity.Interestingly, apart from having difference in V2 loop lengths,between 2-5.J3 and 4.J22 Envs, only six amino acid differenceswere observed in the V2C2 region. To understand whether thesesix residues (P183/D185/N186/N192 in V2 and V207/N215 in C2regions) were responsible for enhanced susceptibility of 2-5.J3 Envto b12 MAb, we substituted these residues in 2-5.J3 Env with theircorresponding ones from b12-resistant 4.J22 Env to give rise toP183S/D185E/N186K/N192K/V207I/N215T [referred to as 2–5.J3 (4.J22-V2C2/SEKKIT)] and examined their sensitivity to b12 MAb. Asshown in Fig. 2 (middle figure) 2-5.J3 (4.J22-V2C2/SEKKIT) dis-played comparable b12 sensitivity as 2-5.J3 wild type Env, sug-gesting that these residues did not play any role in b12susceptibility and indicated that enhanced susceptibility of 2-5.J3Env was due to the coordinated effects of residues in the V2region.
We next examined whether increased sensitivity of 2-5.J3 Envto b12 was due to increased shedding of gp120. Thus, pseudotypedviruses expressing 2-5.J3 and V1V2C2 (2-5.J3) Envs were incu-bated with b12 at a concentration that provided 50% neutraliza-tion (0.3 mg/ml) and those expressing 4.J22 Env with 10 mg/ml at37 1C in a CO2 incubator for different times (Fig. S1). The relativegp120 shedding was assessed by measuring the infectivity ofpseudotyped virus-b12 complex at indicated times in TZM-bl cells.
We found that the reduction in infectivity was comparablebetween the b12-sensitive and -resistant variants. Our dataindicated that enhanced sensitivity of 2-5.J3 Env was not due tob12-induced gp120 shedding.
Substitution of 2-5.J3 V2C2 sequence in unrelated b12-resistantprimary clade C Envs displayed varied b12 and VRC01 susceptibilities
We next examined the ability of the 2-5.J3 V2C2 sequence toalter b12 susceptibility of few unrelated primary clade C Envs. Forthis, we selected four b12-resistant clade C Envs: 3-5.J25, 5.J41, 7.J20 and 16055-2.3, and substituted their V2C2 with that of 2-5.J3Env. As shown in Fig. 2 (bottom figure), all the four Envs contain-ing 2–5.J3 V2C2 sequence differed in their degree of susceptibilityto b12 MAb. While 3-5.J25 and 16055-2.3 Envs containing 2-5.J3V2C2 sequence displayed substantial and modest increase in b12susceptibilities respectively compared to their corresponding wildtypes, 7.J20 (2-5.J3 V2C2) showed evidence of marginal increase inb12 susceptibility. Conversely, 2-5.J3 V2C2 sequence was not foundto influence 5.J41 Env in modulating its susceptibility to b12 MAb.Interestingly, presence of aspartic acid at the position of 185(D185) (Fig. 1) was not found to affect the neutralization of these
Fig. 2. Neutralization susceptibilities of wild type, chimeric and mutant pseudotyped viruses expressing 2-5.J3 and 4.J22 Envs (top and middle panel) and other unrelatedprimary Envs expressing 2-5.J3 V2C2 sequence (bottom panel) to b12 MAb. The neutralization assays were carried out in duplicates in at least three independent time points.
S. Patil et al. / Virology 462-463 (2014) 266–272268
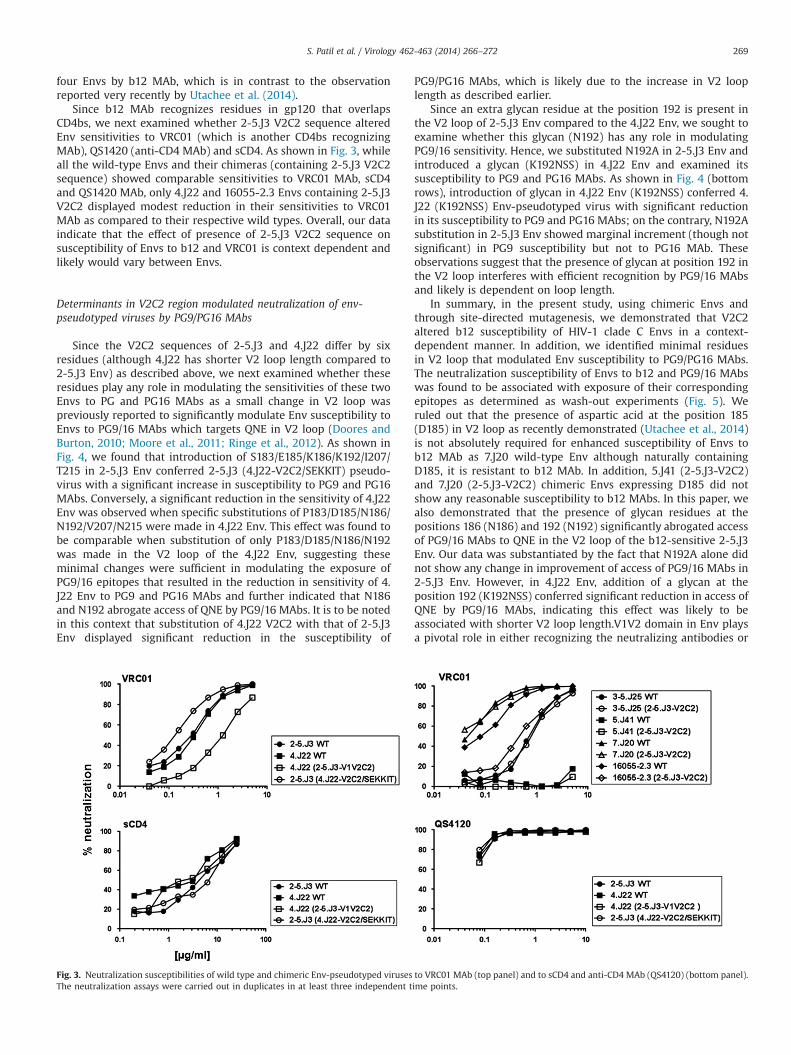
four Envs by b12 MAb, which is in contrast to the observationreported very recently by Utachee et al. (2014).
Since b12 MAb recognizes residues in gp120 that overlapsCD4bs, we next examined whether 2-5.J3 V2C2 sequence alteredEnv sensitivities to VRC01 (which is another CD4bs recognizingMAb), QS1420 (anti-CD4 MAb) and sCD4. As shown in Fig. 3, whileall the wild-type Envs and their chimeras (containing 2-5.J3 V2C2sequence) showed comparable sensitivities to VRC01 MAb, sCD4and QS1420 MAb, only 4.J22 and 16055-2.3 Envs containing 2-5.J3V2C2 displayed modest reduction in their sensitivities to VRC01MAb as compared to their respective wild types. Overall, our dataindicate that the effect of presence of 2-5.J3 V2C2 sequence onsusceptibility of Envs to b12 and VRC01 is context dependent andlikely would vary between Envs.
Determinants in V2C2 region modulated neutralization of env-pseudotyped viruses by PG9/PG16 MAbs
Since the V2C2 sequences of 2-5.J3 and 4.J22 differ by sixresidues (although 4.J22 has shorter V2 loop length compared to2-5.J3 Env) as described above, we next examined whether theseresidues play any role in modulating the sensitivities of these twoEnvs to PG and PG16 MAbs as a small change in V2 loop waspreviously reported to significantly modulate Env susceptibility toEnvs to PG9/16 MAbs which targets QNE in V2 loop (Doores andBurton, 2010; Moore et al., 2011; Ringe et al., 2012). As shown inFig. 4, we found that introduction of S183/E185/K186/K192/I207/T215 in 2-5.J3 Env conferred 2-5.J3 (4.J22-V2C2/SEKKIT) pseudo-virus with a significant increase in susceptibility to PG9 and PG16MAbs. Conversely, a significant reduction in the sensitivity of 4.J22Env was observed when specific substitutions of P183/D185/N186/N192/V207/N215 were made in 4.J22 Env. This effect was found tobe comparable when substitution of only P183/D185/N186/N192was made in the V2 loop of the 4.J22 Env, suggesting theseminimal changes were sufficient in modulating the exposure ofPG9/16 epitopes that resulted in the reduction in sensitivity of 4.J22 Env to PG9 and PG16 MAbs and further indicated that N186and N192 abrogate access of QNE by PG9/16 MAbs. It is to be notedin this context that substitution of 4.J22 V2C2 with that of 2-5.J3Env displayed significant reduction in the susceptibility of
PG9/PG16 MAbs, which is likely due to the increase in V2 looplength as described earlier.
Since an extra glycan residue at the position 192 is present inthe V2 loop of 2-5.J3 Env compared to the 4.J22 Env, we sought toexamine whether this glycan (N192) has any role in modulatingPG9/16 sensitivity. Hence, we substituted N192A in 2-5.J3 Env andintroduced a glycan (K192NSS) in 4.J22 Env and examined itssusceptibility to PG9 and PG16 MAbs. As shown in Fig. 4 (bottomrows), introduction of glycan in 4.J22 Env (K192NSS) conferred 4.J22 (K192NSS) Env-pseudotyped virus with significant reductionin its susceptibility to PG9 and PG16 MAbs; on the contrary, N192Asubstitution in 2-5.J3 Env showed marginal increment (though notsignificant) in PG9 susceptibility but not to PG16 MAb. Theseobservations suggest that the presence of glycan at position 192 inthe V2 loop interferes with efficient recognition by PG9/16 MAbsand likely is dependent on loop length.
In summary, in the present study, using chimeric Envs andthrough site-directed mutagenesis, we demonstrated that V2C2altered b12 susceptibility of HIV-1 clade C Envs in a context-dependent manner. In addition, we identified minimal residuesin V2 loop that modulated Env susceptibility to PG9/PG16 MAbs.The neutralization susceptibility of Envs to b12 and PG9/16 MAbswas found to be associated with exposure of their correspondingepitopes as determined as wash-out experiments (Fig. 5). Weruled out that the presence of aspartic acid at the position 185(D185) in V2 loop as recently demonstrated (Utachee et al., 2014)is not absolutely required for enhanced susceptibility of Envs tob12 MAb as 7.J20 wild-type Env although naturally containingD185, it is resistant to b12 MAb. In addition, 5.J41 (2-5.J3-V2C2)and 7.J20 (2-5.J3-V2C2) chimeric Envs expressing D185 did notshow any reasonable susceptibility to b12 MAbs. In this paper, wealso demonstrated that the presence of glycan residues at thepositions 186 (N186) and 192 (N192) significantly abrogated accessof PG9/16 MAbs to QNE in the V2 loop of the b12-sensitive 2-5.J3Env. Our data was substantiated by the fact that N192A alone didnot show any change in improvement of access of PG9/16 MAbs in2-5.J3 Env. However, in 4.J22 Env, addition of a glycan at theposition 192 (K192NSS) conferred significant reduction in access ofQNE by PG9/16 MAbs, indicating this effect was likely to beassociated with shorter V2 loop length.V1V2 domain in Env playsa pivotal role in either recognizing the neutralizing antibodies or
Fig. 3. Neutralization susceptibilities of wild type and chimeric Env-pseudotyped viruses to VRC01 MAb (top panel) and to sCD4 and anti-CD4 MAb (QS4120) (bottom panel).The neutralization assays were carried out in duplicates in at least three independent time points.
S. Patil et al. / Virology 462-463 (2014) 266–272 269

facilitating the escape of HIV-1 variants to the neutralizing anti-bodies including autologous antibodies, possibly due to its con-formation as well as glycosylation pattern (Overbaugh and Morris,2012; Pinter et al., 2004; Rong et al., 2007). Historically, the V1V2sequences have been shown to undergo changes earlier than thatof the other regions of the Env (Burns and Desrosiers, 1991;Johnson et al., 1991; Laird et al., 2008; Moore et al., 2011; Mooreet al., 2009; Overbaugh and Rudensey, 1992; Overbaugh et al.,1991). Our data suggest that the V2C2 region of the 2-5.J3 Envpossibly affected the spike apex region, which in turn influencedEnv to undergo conformational changes resulting in differentialmodulation of exposure of epitopes targeted by b12 MAb. Becauseconformational change is a complex phenomenon and is oftencontext dependent, this may possibly explain why despite contain-ing the same 2-5.J3 V2C2 sequence, other Envs displayed distinctsusceptibilities to b12 MAb.
Materials and methods
HIV-1 Envs and antibodies
The HIV-1 clade C envelopes 2-5.J3, 3-5.J25 and 4.J22 obtainedfrom Indian patients with recent infection were described pre-viously (Ringe et al., 2010). 7.J20 is also an HIV-1 clade C Envcloned in pcDNA3.1D/V5-His-TOPO (Invitrogen Inc.) obtained froman Indian patient with recent infection, while 16055-2.3 waskindly provided by Dr. David Montefiori. b12 monoclonal antibodywas kindly provided by Dr. Dennis Burton, and was also obtainedfrom the NIH AIDS Research Reagent and Reference Program.VRC01 and sCD4 were obtained from NIH AIDS Research Reagentand Reference Program. PG9 and PG16 MAbs were kindly providedby Dr. Wayne Koff, International AIDS Vaccine Initiative (IAVI),New York. QS4120 MAb was purchased from Calbiochem Inc.
Construction of Env chimera and point mutations
Chimeric Env constructs were made (Fig. 1) by swapping differentvariable and constant domains of b12-sensitive Env (2-5.J3) intoresistant Env (4.J22) by blunt ligation of PCR products using eitherIn-Fusion Dry Down PCR kit (Clontech Inc.) as described before
(Ringe and Bhattacharya, 2012) following the manufacturer's protocolor by two rounds of overlapping PCR in which first-round ampliconswere used as primers in the second round. Site-directed mutagenesisto substitute specific residues in V2 and C2 regions were carried outusing tailor-made primers by PCR using high fidelity polymerase,Platinum Taq (Invitrogen Inc.) or Phusion high fidelity polymerase(New England Biolabs Inc.). The purified PCR products were thendigested with DpnI joined with backbone plasmid by annealing in thepresence of pox DNA polymerase as described before (Ringe et al.,2011).
Preparation of Env-pseudotyped viruses and infectivity assay
Env-pseudotyped viruses were prepared by co-transfection ofpSVIIIenv containing patient Envs and pSG3Δenv in 293T cellsusing calcium phosphate (Promega, Inc.) as described before(Ringe et al., 2010). The infectivity assay was carried out in TZM-bl cells expressing CD4, CCR5 and CXCR4.
Neutralization assay
Neutralization assays were carried out using TZM-bl cells asdescribed before (Ringe et al., 2010). Env-pseudotyped viruses wereincubated with varying dilutions of plasma (heat-inactivated) andvarying concentrations of b12 and PG9/16 MAbs for 1 h at 37 1C, andsubsequently 1�104 TZM-bl cells were added into the mixture in thepresence of 25 μg/ml DEAE dextran (Sigma Inc.). The plates werefurther incubated in a CO2 incubator at 37 1C for two days and thedegree of virus neutralization was assessed by measuring relativeluminescence units (RLU) in a Luminometer (Victor 4, PerkinElmerInc.).
Virus-antibody washout assay
In order to assess the degree of exposure of neutralizingepitopes recognized by b12 and PG9 MAbs, virus antibody wash-out assay was carried out as essentially described by Chakrabartiet al. (2011). Briefly, MAbs (b12 and PG9) were serially 5-folddiluted in 250 ml of 10% DMEM media to which 250 ml of Env-pseudoviruses were added such that the final concentrations of
Fig. 4. Effect of glycan modification and specific amino acid substitutions in 2–5.J3 and 4.J22 Envs towards modulation of their susceptibilities to PG9 and PG16 MAbs.The point substitutions were made by site-directed mutagenesis. The data are representation of three independent experiments done in duplicates.
S. Patil et al. / Virology 462-463 (2014) 266–272270

antibodies became 50�0.08 mg/ml in a total volume of 500 ml,while in the “no inhibitor” control, the same volume of media wasadded instead of antibody. The reaction mixture was incubated for30 min at 37 1C following which 250 ml of reaction mixture wasadded in ultra-clear centrifuge tubes (Beckman Coulter, Inc.),which was diluted to 4 ml by complete DMEM and centrifugedat 35,000 rpm in an SW60 Ti swinging rotor (Beckman Optima-XE-100, Inc.) for 2 h at 4 1C. The virus pellet was washed twice with4 ml of media by centrifuging at 40,000 rpm for 30 min at 4 1C.After the final wash, 250 ml of DMEM was added to the virus pelletand suspended by gentle shaking at 4 1C for 30 min. 100 ml of thesuspended virus was used to infect 100 ml of TZM-b cells(0.2�106/ml) in duplicate. From the remaining 250 ml of reactionmixture, an equal volume of the antibody virus mixture was usedas a “no washout” control. Plates were incubated at 37 1C in a CO2
incubator for 2 days, following which the degree of virus neu-tralization was assessed by measuring the reduction of infection inTZM-bl cells.
GenBank accession number
The GenBank accession number of HIV-1 clade C Env, 7.J20 isEU908223.1.
Acknowledgments
This work was supported in part by a grant from the Depart-ment of Biotechnology, Government of India, to JB (BT/PR12853/MED/29/141/2009) and by the intramural support from the IAVI-THSTI HIV Vaccine Design Program. IAVI's work was made possibleby generous support from many donors including: the Bill &Melinda Gates Foundation; the Ministry of Foreign Affairs ofDenmark; Irish Aid; the Ministry of Finance of Japan; the Ministryof Foreign Affairs of the Netherlands; the Norwegian Agency forDevelopment Cooperation (NORAD); the United Kingdom
Department for International Development (DFID); and the UnitedStates Agency for International Development (USAID). The full listof IAVI donors is available at www.iavi.org. This study was madepossible by the generous support of the American people throughUSAID. The contents are the responsibility of the authors and donot necessarily reflect the views of USAID or the United StatesGovernment. We thank Dr. Dennis Burton, Scripps ResearchInstitute, La Jolla, California, and Dr. Wayne Koff, InternationalAIDS Vaccine Initiative, New York, for help with obtaining b12, PG9and PG16 monoclonal antibodies, respectively. We sincerelyacknowledge the support of the members of the HVTR laboratory,Prof. G Balakrish Nair, Prof. Sudhanshu Vrati, Dr. Rick King and Dr.Rajat Goyal. We thank the Director, National AIDS ResearchInstitute, Pune, for making available the HIV-1 clade C envelopeclones.
Appendix A. Supporting information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.virol.2014.06.018.
References
Bunnik, E.M., Pisas, L., van Nuenen, A.C., Schuitemaker, H., 2008. Autologousneutralizing humoral immunity and evolution of the viral envelope in thecourse of subtype B human immunodeficiency virus type 1 infection. J. Virol.82, 7932–7941.
Burns, D.P., Desrosiers, R.C., 1991. Selection of genetic variants of simian immuno-deficiency virus in persistently infected rhesus monkeys. J. Virol. 65,1843–1854.
Burton, D.R., Pyati, J., Koduri, R., Sharp, S.J., Thornton, G.B., Parren, P.W., Sawyer, L.S.,Hendry, R.M., Dunlop, N., Nara., P.L., 1994. Efficient neutralization of primaryisolates of HIV- 1 by a recombinant human monoclonal antibody. Science 266,1024–1027.
Chakrabarti, B.K., Walker, L.M., Guenaga, J.F., Ghobbeh, A., Poignard, P., Burton, D.R.,Wyatt, R.T., 2011. Direct antibody access to the HIV-1 membrane-proximalexternal region positively correlates with neutralization sensitivity. J. Virol. 85,8217–8226.
Fig. 5. The effect of presence of the 2-5.J3 V2C2 sequence in 4.J22 Env (top panel) and substitution of P183S/D185E/N186K/N192K/V207I/N215T in 2-5.J3 Env (bottom panel)on exposure of b12 and PG9 epitopes, respectively. Virus-MAb washout assay was carried out using an ultracentrifuge to remove unbound b12 and PG9 MAbs from theEnv-pseudotyped viruses. MAb bound Env-pseudotyped viruses were mixed with TZM-bl cells and reduction in virus infection was assessed in a dose-dependent manner.
S. Patil et al. / Virology 462-463 (2014) 266–272 271

Chen, L., Kwon, Y.D., Zhou, T., Wu, X., O'Dell, S., Cavacini, L., Hessell, A.J., Pancera, M.,Tang, M., Xu, L., Yang, Z.Y., Zhang, M.Y., Arthos, J., Burton, D.R., Dimitrov, D.S.,Nabel, G.J., Posner, M.R., Sodroski, J., Wyatt, R., Mascola, J.R., Kwong, P.D., 2009.Structural basis of immune evasion at the site of CD4 attachment on HIV-1gp120. Science 326, 1123–1127.
Doores, K.J., Burton, D.R., 2010. Variable loop glycan dependency of the broad andpotent HIV-1-neutralizing antibodies PG9 and PG16. J. Virol. 84, 10510–10521.
Duenas-Decamp, M.J., Peters, P., Burton, D., Clapham, P.R., 2008. Natural resistance ofhuman immunodeficiency virus type 1 to the CD4bs antibody b12 conferred by aglycan and an arginine residue close to the CD4 binding loop. J. Virol. 82, 5807–5814.
Etemad-Moghadam, B., Sun, Y., Nicholson, E.K., Karlsson, G.B., Schenten, D.,Sodroski, J., 1999. Determinants of neutralization resistance in the envelopeglycoproteins of a simian-human immunodeficiency virus passaged in vivo.J. Virol. 73, 8873–8879.
Frost, S.D., Wrin, T., Smith, D.M., Kosakovsky Pond, S.L., Liu, Y., Paxinos, E., Chappey,C., Galovich, J., Beauchaine, J., Petropoulos, C.J., Little, S.J., Richman, D.D., 2005.Neutralizing antibody responses drive the evolution of human immunodefi-ciency virus type 1 envelope during recent HIV infection. Proc. Natl. Acad. Sci.U. S. A. 102, 18514–18519.
Gray, E.S., Meyers, T., Gray, G., Montefiori, D.C., Morris, L., 2006. Insensitivity ofpaediatric HIV-1 subtype C viruses to broadly neutralising monoclonal anti-bodies raised against subtype B. PLoS Med. 3, e255.
Gray, E.S., Moore, P.L., Choge, I.A., Decker, J.M., Bibollet-Ruche, F., Li, H., Leseka, N.,Treurnicht, F., Mlisana, K., Shaw, G.M., Karim, S.S., Williamson, C., Morris, L.,2007. Neutralizing antibody responses in acute human immunodeficiency virustype 1 subtype C infection. J. Virol. 81, 6187–6196.
Huang, J., Ofek, G., Laub, L., Louder, M.K., Doria-Rose, N.A., Longo, N.S., Imamichi, H.,Bailer, R.T., Chakrabarti, B., Sharma, S.K., Alam, S.M., Wang, T., Yang, Y., Zhang, B.,Migueles, S.A., Wyatt, R., Haynes, B.F., Kwong, P.D., Mascola, J.R., Connors, M.,2012. Broad and potent neutralization of HIV-1 by a gp41-specific humanantibody. Nature 491, 406–412.
Johnson, P.R., Hamm, T.E., Goldstein, S., Kitov, S., Hirsch, V.M., 1991. The genetic fateof molecularly cloned simian immunodeficiency virus in experimentallyinfected macaques. Virology 185, 217–228.
Kulkarni, S.S., Lapedes, A., Tang, H., Gnanakaran, S., Daniels, M.G., Zhang, M.,Bhattacharya, T., Li, M., Polonis, V.R., McCutchan, F.E., Morris, L., Ellenberger,D., Butera, S.T., Bollinger, R.C., Korber, B.T., Paranjape, R.S., Montefiori, D.C.,2009. Highly complex neutralization determinants on a monophyletic lineageof newly transmitted subtype C HIV-1 Env clones from India. Virology 385,505–520.
Laird, M.E., Igarashi, T., Martin, M.A., Desrosiers, R.C., 2008. Importance of the V1/V2loop region of simian-human immunodeficiency virus envelope glycoproteingp120 in determining the strain specificity of the neutralizing antibodyresponse. J. Virol. 82, 11054–11065.
Li, B., Decker, J.M., Johnson, R.W., Bibollet-Ruche, F., Wei, X., Mulenga, J., Allen, S.,Hunter, E., Hahn, B.H., Shaw, G.M., Blackwell, J.L., Derdeyn, C.A., 2006. Evidencefor potent autologous neutralizing antibody titers and compact envelopes inearly infection with subtype C human immunodeficiency virus type 1. J. Virol.80, 5211–5218.
Liu, J., Bartesaghi, A., Borgnia, M.J., Sapiro, G., Subramaniam, S., 2008. Moleculararchitecture of native HIV-1 gp120 trimers. Nature 455, 109–113.
Ly, A., Stamatatos, L., 2000. V2 loop glycosylation of the human immunodeficiencyvirus type 1 SF162 envelope facilitates interaction of this protein with CD4 andCCR5 receptors and protects the virus from neutralization by anti-V3 loop andanti-CD4 binding site antibodies. J. Virol. 74, 6769–6776.
Mascola, J.R., 2009. The cat and mouse of HIV-1 antibody escape. PLoS Pathog. 5,e1000592.
Mascola, J.R., Montefiori, D.C., 2010. The role of antibodies in HIV vaccines. Annu.Rev. Immunol. 28, 413–444.
Mo, H., Stamatatos, L., Ip, J.E., Barbas, C.F., Parren, P.W., Burton, D.R., Moore, J.P., Ho,D.D., 1997. Human immunodeficiency virus type 1 mutants that escapeneutralization by human monoclonal antibody IgG1b12. off. J. Virol. 71,6869–6874.
Montefiori, D.C., Morris, L., Ferrari, G., Mascola, J.R., 2007. Neutralizing and otherantiviral antibodies in HIV-1 infection and vaccination. Curr. Opin. HIV AIDS 2,169–176.
Moore, J.P., Cao, Y., Ho, D.D., Koup, R.A., 1994. Development of the anti-gp120antibody response during seroconversion to human immunodeficiency virustype 1. J. Virol. 68, 5142–5155.
Moore, P.L., Gray, E.S., Sheward, D., Madiga, M., Ranchobe, N., Lai, Z., Honnen, W.J.,Nonyane, M., Tumba, N., Hermanus, T., Sibeko, S., Mlisana, K., Abdool Karim, S.S., Williamson, C., Pinter, A., Morris, L., 2011. Potent and broad neutralization ofHIV-1 subtype C by plasma antibodies targeting a quaternary epitope includingresidues in the V2 loop. J. Virol. 85, 3128–3141.
Moore, P.L., Ranchobe, N., Lambson, B.E., Gray, E.S., Cave, E., Abrahams, M.R.,Bandawe, G., Mlisana, K., Abdool Karim, S.S., Williamson, C., Morris, L., 2009.Limited neutralizing antibody specificities drive neutralization escape in earlyHIV-1 subtype C infection. PLoS Pathog. 5, e1000598.
Overbaugh, J., Morris, L., 2012. The Antibody Response against HIV-1. Cold SpringHarb. Perspect. Med. 2, a007039.
Overbaugh, J., Rudensey, L.M., 1992. Alterations in potential sites for glycosylationpredominate during evolution of the simian immunodeficiency virus envelopegene in macaques. J. Virol. 66, 5937–5948.
Overbaugh, J., Rudensey, L.M., Papenhausen, M.D., Benveniste, R.E., Morton, W.R.,1991. Variation in simian immunodeficiency virus env is confined to V1 and V4during progression to simian AIDS. J. Virol. 65, 7025–7031.
Pantophlet, R., Ollmann Saphire, E., Poignard, P., Parren, P.W., Wilson, I.A., Burton, D.R., 2003. Fine mapping of the interaction of neutralizing and nonneutralizingmonoclonal antibodies with the CD4 binding site of human immunodeficiencyvirus type 1 gp120. J. Virol. 77, 642–658.
Pinter, A., Honnen, W.J., He, Y., Gorny, M.K., Zolla-Pazner, S., Kayman, S.C., 2004. TheV1/V2 domain of gp120 is a global regulator of the sensitivity of primaryhuman immunodeficiency virus type 1 isolates to neutralization by antibodiescommonly induced upon infection. J. Virol. 78, 5205–5215.
Richman, D.D., Wrin, T., Little, S.J., Petropoulos, C.J., 2003. Rapid evolution of theneutralizing antibody response to HIV type 1 infection. Proc. Natl. Acad. Sci. U.S. A. 100, 4144–4149.
Ringe, R., Bhattacharya, J., 2012. Association of enhanced HIV-1 neutralization by asingle Y681H substitution in gp41 with increased gp120-CD4 interaction andmacrophage Infectivity. PLoS One 7, e37157.
Ringe, R., Phogat, S., Bhattacharya, J., 2012. Subtle alteration of residues includingN-linked glycans in V2 loop modulates HIV-1 neutralization by PG9 and PG16monoclonal antibodies. Virology 426, 34–41.
Ringe, R., Sharma, D., Zolla-Pazner, S., Phogat, S., Risbud, A., Thakar, M., Paranjape,R., Bhattacharya, J., 2011. A single amino acid substitution in the C4 region ingp120 confers enhanced neutralization of HIV-1 by modulating CD4 bindingsites and V3 loop. Virology 418, 123–132.
Ringe, R., Thakar, M., Bhattacharya, J., 2010. Variations in autologous neutralizationand CD4 dependence of b12 resistant HIV-1 clade C env clones obtained atdifferent time points from antiretroviral naive Indian patients with recentinfection. Retrovirology 7, 76.
Rong, R., Bibollet-Ruche, F., Mulenga, J., Allen, S., Blackwell, J.L., Derdeyn, C.A., 2007.Role of V1V2 and other human immunodeficiency virus type 1 envelopedomains in resistance to autologous neutralization during clade C infection.J. Virol. 81, 1350–1359.
Si, Z., Cayabyab, M., Sodroski, J., 2001. Envelope glycoprotein determinants ofneutralization resistance in a simian-human immunodeficiency virus (SHIV-HXBc2P 3.2) derived by passage in monkeys. J. Virol. 75, 4208–4218.
Thenin, S., Roch, E., Samleerat, T., Moreau, T., Chaillon, A., Moreau, A., Barin, F.,Braibant, M., 2012. Naturally occurring substitutions of conserved residues inHIV-1 variants of different clades are involved in PG9 and PG16 resistance toneutralization. J. Gen. Virol. 93, 1495–1505.
Tomaras, G.D., Yates, N.L., Liu, P., Qin, L., Fouda, G.G., Chavez, L.L., Decamp, A.C.,Parks, R.J., Ashley, V.C., Lucas, J.T., Cohen, M., Eron, J., Hicks, C.B., Liao, H.X., Self,S.G., Landucci, G., Forthal, D.N., Weinhold, K.J., Keele, B.F., Hahn, B.H., Greenberg,M.L., Morris, L., Karim, S.S., Blattner, W.A., Montefiori, D.C., Shaw, G.M.,Perelson, A.S., Haynes, B.F., 2008. Initial B-cell responses to transmitted humanimmunodeficiency virus type 1: virion-binding immunoglobulin M (IgM) andIgG antibodies followed by plasma anti-gp41 antibodies with ineffectivecontrol of initial viremia. J. Virol. 82, 12449–12463.
Utachee, P., Isarangkura-na-ayuthaya, P., Tokunaga, K., Ikuta, K., Takeda, N.,Kameoka, M., 2014. Impact of amino acid substitutions in the V2 and C2regions of human immunodeficiency virus type 1 CRF01_AE envelope glyco-protein gp120 on viral neutralization susceptibility to broadly neutralizingantibodies specific for the CD4 binding site. Retrovirology 11, 32.
Utachee, P., Nakamura, S., Isarangkura-Na-Ayuthaya, P., Tokunaga, K., Sawanpanya-lert, P., Ikuta, K., Auwanit, W., Kameoka, M., 2010. Two N-linked glycosylationsites in the V2 and C2 regions of human immunodeficiency virus type1 CRF01_AE envelope glycoprotein gp120 regulate viral neutralization suscept-ibility to the human monoclonal antibody specific for the CD4 binding domain.J. Virol. 84, 4311–4320.
Walker, L.M., Huber, M., Doores, K.J., Falkowska, E., Pejchal, R., Julien, J.P., Wang, S.K.,Ramos, A., Chan-Hui, P.Y., Moyle, M., Mitcham, J.L., Hammond, P.W., Olsen, O.A.,Phung, P., Fling, S., Wong, C.H., Phogat, S., Wrin, T., Simek, M.D., 2011. Broadneutralization coverage of HIV by multiple highly potent antibodies (PrincipalInvestigators, P.G., Koff, W.C., Wilson, I.A., Burton, D.R., Poignard, P.). Nature 477,466–470.
Walker, L.M., Phogat, S.K., Chan-Hui, P.Y., Wagner, D., Phung, P., Goss, J.L., Wrin, T.,Simek, M.D., Fling, S., Mitcham, J.L., Lehrman, J.K., Priddy, F.H., Olsen, O.A., Frey,S.M., Hammond, P.W., Kaminsky, S., Zamb, T., Moyle, M., Koff, W.C., Poignard, P.,Burton, D.R., 2009. Broad and potent neutralizing antibodies from an Africandonor reveal a new HIV-1 vaccine target. Science 326, 285–289.
Wei, X., Decker, J.M., Wang, S., Hui, H., Kappes, J.C., Wu, X., Salazar-Gonzalez, J.F.,Salazar, M.G., Kilby, J.M., Saag, M.S., Komarova, N.L., Nowak, M.A., Hahn, B.H.,Kwong, P.D., Shaw, G.M., 2003. Antibody neutralization and escape by HIV-1.Nature 422, 307–312.
Wu, X., Yang, Z.Y., Li, Y., Hogerkorp, C.M., Schief, W.R., Seaman, M.S., Zhou, T., Schmidt,S.D., Wu, L., Xu, L., Longo, N.S., McKee, K., O'Dell, S., Louder, M.K., Wycuff, D.L., Feng,Y., Nason, M., Doria-Rose, N., Connors, M., Kwong, P.D., Roederer, M., Wyatt, R.T.,Nabel, G.J., Mascola, J.R., 2010. Rational design of envelope identifies broadlyneutralizing human monoclonal antibodies to HIV-1. Science 329, 856–861.
Wu, X., Zhou, T., O'Dell, S., Wyatt, R.T., Kwong, P.D., Mascola, J.R., 2009. Mechanismof human immunodeficiency virus type 1 resistance to monoclonal antibodyB12 that effectively targets the site of CD4 attachment. J. Virol. 83,10892–10907.
Zhang, H., Rola, M., West, J.T., Tully, D.C., Kubis, P., He, J., Kankasa, C., Wood, C., 2010.Functional properties of the HIV-1 subtype C envelope glycoprotein associatedwith mother-to-child transmission. Virology 400, 164–174.
Zhou, T., Xu, L., Dey, B., Hessell, A.J., Van Ryk, D., Xiang, S.H., Yang, X., Zhang, M.Y.,Zwick, M.B., Arthos, J., Burton, D.R., Dimitrov, D.S., Sodroski, J., Wyatt, R., Nabel,G.J., Kwong, P.D., 2007. Structural definition of a conserved neutralizationepitope on HIV-1 gp120. Nature 445, 732–737.
S. Patil et al. / Virology 462-463 (2014) 266–272272





























