Embed Size (px)
Citation preview

Page 1 of 5
Review
Licensee OA Publishing London 2013. Creative Commons Attribution License (CC-BY)
For citation purposes: Garcia MC, Lipke PN, Klotz SA. Pathogenic microbial amyloids: their function and the host response. OA Microbiology 2013 Dec 01;1(1):2. Co
mpe
ting
inte
rest
s: n
one
decl
ared
. Con
flict
of i
nter
ests
: non
e de
clar
ed.
All a
utho
rs c
ontr
ibut
ed to
con
cepti
on a
nd d
esig
n, m
anus
crip
t pre
para
tion,
read
and
app
rove
d th
e fin
al m
anus
crip
t.Al
l aut
hors
abi
de b
y th
e As
soci
ation
for M
edic
al E
thic
s (AM
E) e
thic
al ru
les o
f disc
losu
re.
Infe
ctio
n &
Imm
unity
Pathogenic microbial amyloids: their function and the host response
MC Garcia1, PN Lipke1, SA Klotz2*
AbstractIntroductionFunctional microbial amyloids are ubiquitous in nature and some contrib-ute to the pathogenesis of infectious diseases. Three pathogenic microbial amyloids are compared and their contribution to the disease process explained. The recent demonstration and visualisation of fungal amyloid in human invasive candidiasis are dis-cussed. Moreover, the binding of host serum amyloid P component to Can-dida functional amyloid in invasive human disease is presented in light of its possible role of masking fungi from the host defences. Conclusion Pathogenic fungi possess functional cell surface amyloids that are demon-strated by amyloidophilic dyes and these proteins bind serum amyloid P compound which may mask the fun-gal surface from host defences.
IntroductionAmyloid is a term ordinarily used to describe protein formed from spontaneously self-propagating, in-soluble, β-sheet rich fibrils. These fibrils are resistant to enzymatic di-gestion, have characteristic patterns observed through electron micros-copy and special tinctorial proper-ties such as staining with the dyes, Congo red and thioflavin-T. Although amyloid fibres have been associated with disease, it is now apparent that
the presence of amyloid is not al-ways pathological. Amyloidoses are diseases in which amyloid deposits accumulate extracellularly and dis-rupt the structure and function of tis-sues and organs. The most common is AL amyloidosis caused by monoclonal immunoglobulin light chain deposi-tion in the glomeruli of kidneys of patients with various plasma cell dyscrasias. One amyloidosis, AA, occurs in response to chronic inflam-matory disorders including infec-tious diseases such as tuberculosis and chronic bacterial osteomyelitis. Examples of pathological amyloid deposits, but not considered amyloi-doses, include Aβ-amyloid found in the plaques of Alzheimer’s disease and prions found in the spongiform encephalopathies. These disease states are often characterised by protein misfolding that exposes am-yloid-forming properties.
The structure of amyloid assem-blies can lead to functional proper-ties. Amyloid’s tensile strength and resistance to degradation are advan-tageous in nanotechnology where it is used to manufacture tissue scaf-folding, nanowires and nanotubes1. Natural amyloids include fibrils in skin cell melanosomes that impart a characteristic ultrastructure to the organelle, as seen on electron microscopy and are necessary for proper assembly and deposition of melanin2.
Microbes elaborate amyloids that are used to fasten the microorgan-isms to a substratum. They are ubiq-uitous in nature and are important components of microbial biofilms3,4. Since microbial amyloids perform a beneficial function for the microor-ganism they are referred to as ‘func-
tional amyloids’5. These fibrils serve to attach a microbe to a substratum and thus, secure a survival advantage for the microorganism. Some func-tional amyloids attach microbes to inanimate surfaces, others to host cells, still others attach microbes to one another and some amyloids serve to stabilise the biofilm during infection6. We will briefly discuss three microbial functional amyloid proteins presumed to be integral to the pathogenesis of disease in hu-mans: the curli protein of Escheri-chia coli, merozoite surface protein 2 (MSP2) of Plasmodium falciparum merozoites and the Als cell surface adhesins of Candida albicans.
DiscussionThe authors have referenced some of their own studies in this re-view. These referenced studies have been conducted in accordance with the Declaration of Helsinki (1964) and the protocols of these studies have been approved by the relevant ethics committees related to the insti-tution in which they were performed. All human subjects, in these refer-enced studies, gave informed consent to participate in these studies.
Three pathogenic amyloid adhesins of microbesCurli Pathogenic E. coli strains responsible for acute diarrhoeal diseases express multiple adhesins or attachment pro-teins including fimbriae which are long, hair-like appendages (micro-metres in length) and short, amyloid fibrils known as curli. Fimbriae are resilient fibrils composed of repeating units of amino acids and the length of the assembled fibre provides for
* Corresponding author Email: [email protected] Department of Biology, Brooklyn College of
City University of New York, Brooklyn, NY, USA
2 Department of Medicine, University of Ari-zona, Tucson, AZ, USA

Page 2 of 5
Review
Licensee OA Publishing London 2013. Creative Commons Attribution License (CC-BY)
For citation purposes: Garcia MC, Lipke PN, Klotz SA. Pathogenic microbial amyloids: their function and the host response. OA Microbiology 2013 Dec 01;1(1):2. Co
mpe
ting
inte
rest
s: n
one
decl
ared
. Con
flict
of i
nter
ests
: non
e de
clar
ed.
All a
utho
rs c
ontr
ibut
ed to
con
cepti
on a
nd d
esig
n, m
anus
crip
t pre
para
tion,
read
and
app
rove
d th
e fin
al m
anus
crip
t.Al
l aut
hors
abi
de b
y th
e As
soci
ation
for M
edic
al E
thic
s (AM
E) e
thic
al ru
les o
f disc
losu
re.
to the red blood cell (several of these proteins are being incorporated into vaccines). MSP2 is one of these sur-face proteins and is unique in that it contains a functional amyloid in the N-terminus and is likely important in attachment of the merozoite to the red blood cell9 (Table 1). The amyloid fibrils of MSP2 are resistant to pro-teinase, stain with Congo red and are formed under physiologic conditions, all characteristics of amyloid fibres10. After attachment of the merozoite to the red blood cell, the parasite reori-ents on the cell surface and, using dif-ferent adhesins, penetrates the cell. Once it is intracellular the merozoite is free to digest haemoglobin.
Als proteins of C. albicans There are eight different Als proteins in C. albicans located on the cell sur-face of yeasts and hyphae of this op-portunistic pathogen. Although curli and MSP2 proteins appear to be re-quired for pathogenesis of disease in the human, the intended purpose of Als proteins in C. albicans is likely to
length) that demonstrate character-istics of amyloid, that is, the ability to self-propagate7.
Merozoite surface protein 2 The most lethal form of malaria is caused by P. falciparum, which stands out from other less virulent species of malaria by reason of its ability to parasitise a high percent-age of red blood cells. The stage of the parasite that attaches and enters the red blood cell is the merozoite. It is released initially from the liver, enters red blood cells and ultimately ruptures the cell. The parasites at-tach and enter the red blood cell in order to obtain their food source, haemoglobin. The initial contact of a recently released merozoite and red blood cell is thought to occur by ran-dom collision. On the surface coat of P. falciparum merozoites are numer-ous proteins that extend out from the parasite surface like small knobs8. There are number of merozoite sur-face proteins believed to be impor-tant in the adherence of the parasite
long-range interactions with substra-ta. They can bend and resist torsion and stretch to five times their normal length. Fimbriae are important in colonisation of a surface, for exam-ple, the intestinal wall (Table 1). Curli proteins are highly hydrophobic, at-tached to the cell membrane and con-tribute not only to adherence to tissue but also attachment of bacterium to other bacteria, that is, cell-to-cell ag-gregation which is critical for biofilm formation. Curli are found throughout Enterobacteriaceae and in some spe-cies such as E. coli and Salmonella spp., which are important in the pathogen-esis of disease. In murine models, enterohaemorrhagic E. coli express curli in order to ensure cell-to-cell ag-gregation and adherence to intestinal cells. Curli are the product of several proteins. One subunit attaches to the outer cell membrane and then nucle-ates another protein on top of itself, a process that continues one after an-other to form a fibril3. It is interesting to note that some secreted proteins of E. coli form amyloid ropes (> cm in
Table 1 Characteristics of three microbial cell surface amyloids that mediate adherence of the microbe to human tissue and cells
Characteristic Escherichia coli curli Plasmodium falciparum MSP2 Candida albicans Als proteins
Molecular weightPolymers of ~15 kDa
protein30 kDa Heavily glycosylated, >200 kD
GPI cell surface anchor? No Yes Yes
Location of amyloidEntire structure is attached to the outer cell membrane
N-terminus of the proteinA threonine-rich repeat region
adjacent to N-terminal immunoglobulin region
Agglutination or aggregation of microbes?
Yes Not known Yes
Amyloid fibres demonstrated?
Yes Yes Yes
Attachment targetsHost proteins, other
Escherichia coli Possibly glycophorinPeptides/proteins of other candida
albicans or host tissue
Vaccine candidateAntibodies to curli can be detected in patients with disease (Bian et al. 2000)
Yes, one of several surface proteins being included in
potential vaccines Yes (Edwards 2012)
GPI, glycosylphosphatidyl inositol anchor. These amyloids are all expressed under physiological conditions.

Page 3 of 5
Review
Licensee OA Publishing London 2013. Creative Commons Attribution License (CC-BY)
For citation purposes: Garcia MC, Lipke PN, Klotz SA. Pathogenic microbial amyloids: their function and the host response. OA Microbiology 2013 Dec 01;1(1):2. Co
mpe
ting
inte
rest
s: n
one
decl
ared
. Con
flict
of i
nter
ests
: non
e de
clar
ed.
All a
utho
rs c
ontr
ibut
ed to
con
cepti
on a
nd d
esig
n, m
anus
crip
t pre
para
tion,
read
and
app
rove
d th
e fin
al m
anus
crip
t.Al
l aut
hors
abi
de b
y th
e As
soci
ation
for M
edic
al E
thic
s (AM
E) e
thic
al ru
les o
f disc
losu
re.
of ulceration of the bowel epithelium into the muscularis layer. The amy-loidophilic dyes Congo red and thio-flavin-T demonstrated the presence of amyloid on all morphologies of invading Candida (Figures 2 and 3). A monoclonal antibody directed against SAP bound to the same sur-faces as dyes (Figure 4). Accompa-nying in vitro work established the fact that SAP has a specific interac-tion with the fungi once functional amyloid is expressed on the fungal surface.
This interaction of SAP with Can-dida spp. was a surprising finding and not previously known to occur. The story is intriguing because the pres-ence of SAP is a characteristic find-ing of all the amyloidoses. SAP is a pentraxin, a pentameric protein that circulates at relatively stable concen-trations of 20–35 mg/L. (C-reactive protein, the acute phase reactant, has homologous structures to SAP and is also a pentraxin with a dynamic range of 0.05–500 mg/L.) Human SAP, how-ever, is not an acute phase reactant. It binds to amyloid whether it is in-tracellular or extracellular and is universally present where amyloid is deposited15. In amyloidoses, SAP contributes to the mass effect of the amyloid deposition that leads to tis-sue and organ dysfunction. Removal of SAP from amyloid deposits in pa-tients results in improvement16. It is easy to envision how pentameric SAP might cover up amyloid fibrils and mask recognition of amyloid. Perhaps this is one of the reasons
exposed on host cells and fungal cells (Figure 1). The threonine-rich domain is formed of β-sheets that form amyloid fibrils. In addition to amyloid formation this region con-tributes to the adherence process by stabilising adherence. It is charac-teristic of C. albicans adherence and aggregation that it is very resistant to physical and chemical force, for example, 8 M urea and formamide are required to undue adherence (as they are to break apart amyloid). We have termed the Als proteins the ‘perfect adhesive’: binding to a wide variety of protein targets with al-most unbreakable adhesion even in the face of shear forces11. These cell surface proteins likely contribute to the phenotypic characteristics of the fungus (Table 2).
Fungal amyloid in human infectionThe recent observation that Candida spp. elaborate amyloid in human dis-ease and that these fibrils are ‘recog-nised’ by the host resulting in the deposition of host serum amyloid P component (SAP) onto the fungi has opened a new chapter in can-didiasis12,13. The study involved 25 autopsy patients with invasive can-didiasis of the intestines. Many, but not all of the patients had neutrope-nia near the time of death and some had candidemia, detected either an-temortem or at autopsy14. Multiple specimens from each patient dem-onstrated yeast cells and hyphae en-tering the intestines through a point
serve a social function, that is, aggre-gating fungal cells to another as this is the preferred phenotype in na-ture. Candida albicans is considered a commensal of human mucous sur-faces. However, if host defences are compromised the Als proteins can mediate adherence of the fungus to peptides and proteins, the fun-gus does not ordinarily encounter, for example, human endothelium or urothelial cells. The best studied of the Als proteins, Als5p when at-tached to the surface of non-patho-genic Saccharomyces cerevisiae (bak-er’s yeast) has been used to model the functions of the different regions of the Als proteins. The protein is connected to the cell wall by a gly-cosylphosphatidyl inositol anchor where a C-terminal stalk region rais-es the protein above the cell surface (Figure 1). The stalk is followed by multiple 36-amino acid tandem re-peats (the number of repeats varies in each protein), which are instru-mental in cell-to-cell aggregation. A threonine-rich domain forms a con-nection between the tandem repeats, and the N-terminal immunoglobulin region binds to various peptides
Table 2 Cell surface functional amyloids of Candida albicans influence cellular phenotypic characteristics of the fungus
Characteristics of amyloids Does this apply to Candida albicans?
Hydrophobic Yes (Klotz et al. 1985)
Surface active Yes (Klotz 1989)
Avidity of Candida albicans amyloid for self
Yes (Garcia et al.20)
Aggregation of cells Yes (Klotz and Penn 1987)
Repetitive motif Yes (Lipke et al. 2011)
Figure 1: Schematic representation of Candida albicans Als cell surface protein.
Page 4 of 5
Review
Licensee OA Publishing London 2013. Creative Commons Attribution License (CC-BY)
For citation purposes: Garcia MC, Lipke PN, Klotz SA. Pathogenic microbial amyloids: their function and the host response. OA Microbiology 2013 Dec 01;1(1):2. Co
mpe
ting
inte
rest
s: n
one
decl
ared
. Con
flict
of i
nter
ests
: non
e de
clar
ed.
All a
utho
rs c
ontr
ibut
ed to
con
cepti
on a
nd d
esig
n, m
anus
crip
t pre
para
tion,
read
and
app
rove
d th
e fin
al m
anus
crip
t.Al
l aut
hors
abi
de b
y th
e As
soci
ation
for M
edic
al E
thic
s (AM
E) e
thic
al ru
les o
f disc
losu
re.
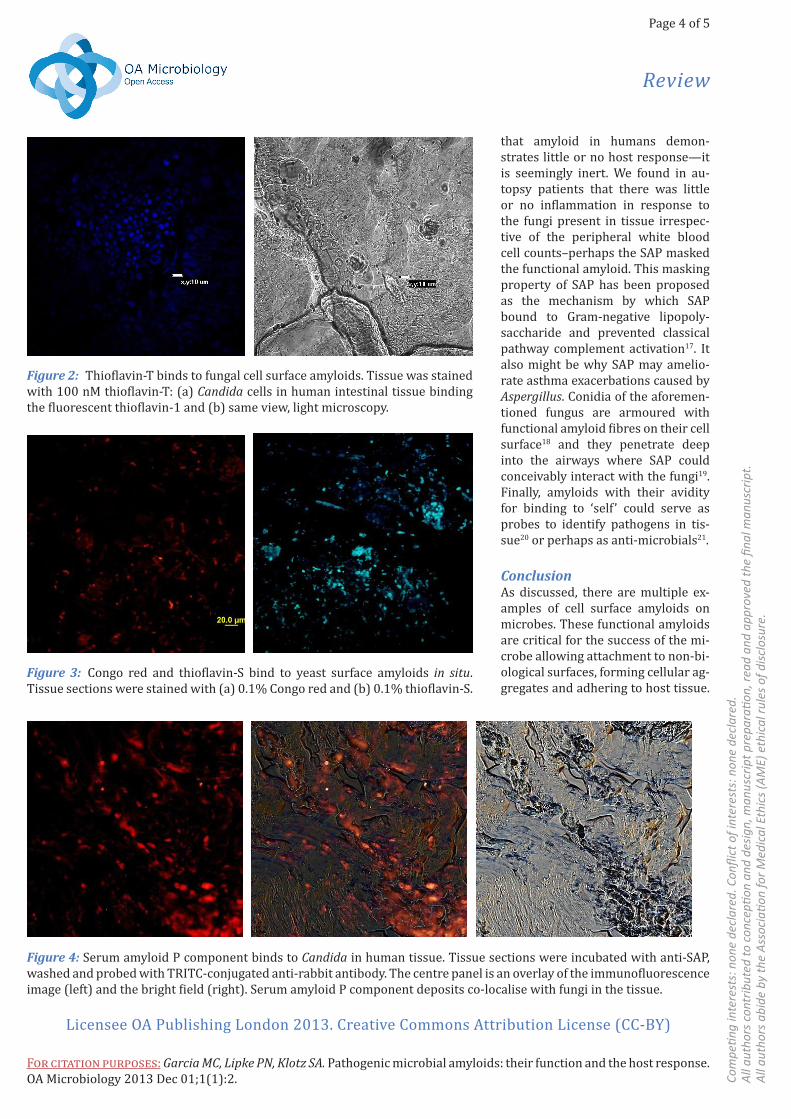
Figure 2: Thioflavin-T binds to fungal cell surface amyloids. Tissue was stained with 100 nM thioflavin-T: (a) Candida cells in human intestinal tissue binding the fluorescent thioflavin-1 and (b) same view, light microscopy.
Figure 3: Congo red and thioflavin-S bind to yeast surface amyloids in situ. Tissue sections were stained with (a) 0.1% Congo red and (b) 0.1% thioflavin-S.
Figure 4: Serum amyloid P component binds to Candida in human tissue. Tissue sections were incubated with anti-SAP, washed and probed with TRITC-conjugated anti-rabbit antibody. The centre panel is an overlay of the immunofluorescence image (left) and the bright field (right). Serum amyloid P component deposits co-localise with fungi in the tissue.
that amyloid in humans demon-strates little or no host response—it is seemingly inert. We found in au-topsy patients that there was little or no inflammation in response to the fungi present in tissue irrespec-tive of the peripheral white blood cell counts–perhaps the SAP masked the functional amyloid. This masking property of SAP has been proposed as the mechanism by which SAP bound to Gram-negative lipopoly-saccharide and prevented classical pathway complement activation17. It also might be why SAP may amelio-rate asthma exacerbations caused by Aspergillus. Conidia of the aforemen-tioned fungus are armoured with functional amyloid fibres on their cell surface18 and they penetrate deep into the airways where SAP could conceivably interact with the fungi19. Finally, amyloids with their avidity for binding to ‘self ’ could serve as probes to identify pathogens in tis-sue20 or perhaps as anti-microbials21.
ConclusionAs discussed, there are multiple ex-amples of cell surface amyloids on microbes. These functional amyloids are critical for the success of the mi-crobe allowing attachment to non-bi-ological surfaces, forming cellular ag-gregates and adhering to host tissue.

Page 5 of 5
Review
Licensee OA Publishing London 2013. Creative Commons Attribution License (CC-BY)
For citation purposes: Garcia MC, Lipke PN, Klotz SA. Pathogenic microbial amyloids: their function and the host response. OA Microbiology 2013 Dec 01;1(1):2. Co
mpe
ting
inte
rest
s: n
one
decl
ared
. Con
flict
of i
nter
ests
: non
e de
clar
ed.
All a
utho
rs c
ontr
ibut
ed to
con
cepti
on a
nd d
esig
n, m
anus
crip
t pre
para
tion,
read
and
app
rove
d th
e fin
al m
anus
crip
t.Al
l aut
hors
abi
de b
y th
e As
soci
ation
for M
edic
al E
thic
s (AM
E) e
thic
al ru
les o
f disc
losu
re.
15. Pepys MB, Rademacher TW, Amatay-akul-Chantler S, Williams P, Noble GE, Hutchinson WL, et al. Human serum amy-loid P component is an invariant constitu-ent of amyloid deposits and has a uniquely homogeneous glycostructure. Proc Natl Acad Sci USA. 1994 Jun;91(12):5602–6.16. Pepys MB. Pathogenesis, diagnosis and treatment of systemic amyloidosis. Philos Trans R Soc Lond B Biol Sci. 2001 Feb;356(1406):203–10.17. de Haas CJ, van Leeuwen EM, van Bommel T, Verhoef J, van Kessel KP, van Strijp JA. Serum amyloid P compo-nent bound to gram-negative bacteria prevents lipopolysaccharide-mediated classical pathway complement activa-tion. Infect Immun. 2000 Apr;68(4): 1753–9.18. Paris S, Debeaupuis JP, Crameri R, Car-ey M, Charlès F, Prévost MC, et al. Conid-ial hydrophobins of Aspergillus fumiga-tus. Appl Environ Microbiol. 2003 Mar; 69(3):1581–8.19. Moreira AP, Cavassani KA, Hullinger R, Rosada RS, Fong DJ, Murray L, et al. Serum amyloid P attenuates M2 mac-rophage activation and protects against fungal spore-induced allergic airway dis-ease. J Allergy Clin Immunol. 2010 Oct; 126(4):712–21.20. Garcia MC, Lysak N, Filonenko A, Rich-ards H, Sobonya RE, Klotz SA, Lipke PN. Peptide detection of fungal functional amyloids in infected tissue. PLoS One. 2014 Jan;9(1):e86067. 21. Scorciapino MA, Pirri G, Vargiu AV, Rug-gerone P, Giuliani A, Casu M, et al. A novel dendrimeric peptide with antimicrobial properties: structure-function analy-sis of SB056. Biophys J. 2012 Mar;102 (5):1039–48.
7. Xicohtencatl-Cortes J, Saldaña Z, Deng W, Castañeda E, Freer E, Tarr PI, et al. Bacterial macroscopic rope-like fibers with cytopathic and adhesive proper-ties. J Biol Chem. 2010 Oct;285(42): 32336–42.8. Bannister LH, Mitchell GH, Butch er GA, Dennis ED, Cohen S. Structure and development of the surface coat of erthrocytic merozoites of Plasmodium knowlesi. Cell Tissue Res. 1986;245(2): 281–90.9. Low A, Chandrashekaran IR, Adda CG, Yao S, Sabo JK, Zhang X, et al. Merozoite surface protein 2 of Plasmodium falci-parum: expression, structure, dynamics, and fibril formation of the conserved N-terminal domain. Biopolymers. 2007 Sep;87(1):12–22.10. Adda CG, Murphy VJ, Sunde M, Wad-dington LJ, Schloegel J, Talbo GH, et al. Plasmodium falciparum merozoite sur-face protein 2 is unstructured and forms amyloid like fibrils. Mol Biochem Parasi-tol. 2009 Aug;166(2):159–71.11. Klotz SA, Lipke PN, eds. The perfect adhesive. Badajoz, Spain: Formatex; 2010.12. Gilchrist KB, Garcia MC, Sobonya R, Lipke PN, Klotz SA. New features of in-vasive candidiasis in humans: amyloid formation by the fungi and deposition of serum amyloid P component by the host. J Infect Dis. 2012 Nov;206(9):1473–8.13. Pepys MB. Invasive candidiasis: new insights presaging new therapeu-tic approaches? J Infect Dis. 2012 Nov; 206(9):1339–41.14. Thorn JL, Gilchrist KB, Sobonya RE, Gaur NK, Lipke PN, Klotz SA. Postmor-tem candidaemia: marker of dissemi-nated disease. J Clin Pathol. 2010 Apr; 63(4):337–40.
They are also necessary for the suc-cess of biofilms and also in infection of the host. These unique structures are utilised for the benefit of the microbe but sometimes to the detriment of the host as seen in some infectious diseases.
Abbreviations listGPI, glycosylphosphatidyl inosi-tol anchor; SAP, serum amyloid P component.
References1. Marshall KE, Serpell LC. Structural in-tegrity of beta-sheet assembly. Biochem Soc Trans. 2009 Aug;37(4):671–6.2. Watt B, van Niel G, Raposo G, Marks MS. PMEL: a pigment cell-specific model for functional amyloid formation. Pig-ment Cell Melanoma Res. 2013 May; 26(3):300–15.3. Blanco LP, Evans ML, Smith DR, Badtke MP, Chapman MR. Diversity, biogen-esis and function of microbial amyloids. Trends Microbiol. 2012 Feb;20(2):66–73.4. Larsen P, Nielsen JL, Otzen D, Nielsen PH. Amyloid-like adhesins produced by floc-forming and filamentous bacteria in activated sludge. Appl Environ Microbiol. 2008 Mar;74(5):1517–265. Otzen D, Nielsen PH. We find them here, we find them there: functional bac-terial amyloid. Cell Mol Life Sci. 2008 Mar;65(6):910–276. Schwartz K, Syed AK, Stephenson RE, Rickard AH, Boles BR. Functional amyloi-ds composed of phenol soluble modulins stabilize Staphylococcus aureus biofilms. PLoS Pathog. 2012 Jun;8(6):e1002744.





![Colloid-amyloid Bodies in PUVA-treated Human Psoriatic ...Amyloid of primary cutaneous amyloidoses such as lichen amyloidosus [5, 17], macular amyloidosis [6] and amyloid dep- osition](https://img.dokumen.tips/doc/110x75/5e62f6a65098527daa05e73b/colloid-amyloid-bodies-in-puva-treated-human-psoriatic-amyloid-of-primary-cutaneous.jpg)


![Journal of BIOPHOTONICS - unipi.it · mer’s, systemic amyloidoses and Parkinson’s [1]. For this reason, the structure and the biology of the amy-loid fibrils are under extensive](https://img.dokumen.tips/doc/110x75/5b465ab97f8b9aa4148c3a91/journal-of-biophotonics-unipiit-mers-systemic-amyloidoses-and-parkinsons.jpg)












![Effects of Seeding on Lysozyme Amyloid Fibrillation in the ... · Alzheimer’s disease, type 2 diabetes, and several systemic amyloidoses [1-3]. These proteins, despite their unrelated](https://img.dokumen.tips/doc/110x75/5f66a2f7828269373b7d097b/effects-of-seeding-on-lysozyme-amyloid-fibrillation-in-the-alzheimeras-disease.jpg)







