Embed Size (px)
Citation preview

Parathyroid Gland Structure of Some Tropical Lizards WILLIAM SENIOR Depar tmen t of Biological Sciences, Ahmadzc Bello Universi ty , Zaria, Nigeria
ABSTRACT The number, location, size, shape and microstructure of the parathyroid glands of Agarna agarna agama, Hemidactylus broohi angulatus and Pytodactylus hasselquisti hasselquisti was investigated using approximately 250 specimens of each species from the Zaria area of Nigeria.
Only parathyroid 111 was found. Additional patches of tissue in Hemidactylus, though possibly parathyroid IV, are considered to be derived from parathy- roid 111. It was found that the amount of parathyroid tissue per gram of body weight was similar in the three species used, and that females had more parathyroid tissue than males. The same situation seems possible in other species.
The structure of the parathyroid glands could not be related to taxonomic grouping within the Sauria, but the general picture was found to be more similar to that of birds and mammals than to that of amphibians.
The anatomy, histology and cytology of mammalian parathyroid glands have been extensively studied in connection with medical and veterinary research. Other tetrapod groups have not been the sub- ject of comparative detailed investigations. Amphibian parathyroid glands have been studied by Waggener (’29, ’30), Cortelyou, Hibner-Owerko and Mulroy (‘60), Bosch- witz (‘61), and Cortelyou (‘62). Avian stud- ies have been carried out by Polin, Sturkie and Hunsaker (’57), Urist, Deutsch, Pom- erantz and McLean (‘60), and Simkiss (‘61). Veterinary studies on the domestic fowl have also been carried out by Urist (‘59), Taylor and Hertelendy (‘61), Taylor, Morris and Hertelendy (‘62) and Itakura and Yamagiwa (‘70).
The structure of reptile parathyroid glands is less well known. There are many reports of the position of endocrine struc- tures in investigations concerned with gen- eral anatomy, but on only a few occasions have histological methods been used to show that these structures in lizards are parathyroid in type (Adams, ’39; Peters, ’41; Underwood, ’57; Sidky, ’65; Clark, ’6 7, ’68).
There is also some doubt as to the num- ber of parathyroid glands occurring in reptiles. Greep (‘63) states, “The lizards are usually categorically stated to possess only one pair of parathyroid glands- parathyroid 111. . . . Exceptions have been
J. MORPH., 142: 91-108.
noted however.” Gorbman and Bern (’62) state, “In reptiles there are most often two pairs of parathyroids.” No reference denies the presence of parathyroid I11 in lizards, but the regular occurrence of para- thyroid IV has been reported in only a few instances (table 1).
Table 1 does not indicate the rarity of parathyroid IV in lizards so much as the rarity of studies involving lizard parathy- roid glands.
Goin and Goin (‘62) dismiss the subject by saying, “There is little to be said about the structure of the endocrine glands (in herptiles). Most of them are found in all vertebrates and the morpho- logical variations from one group to an- other seem in general to have little evolu- tionary significance.” The extensive work on the parathyroid glands edited by von Euler and Heller (‘63) contains only one photograph of the internal structure of a reptilian gland, taken from Peters (‘41). Later papers by Rogers (‘63), Sidky (‘65), and Clark (‘65, ’67, ’68) have been con- cerned at least in part with the histology of reptilian parathyroid glands. Also, the recent work edited by Gans (‘70) contains an article (Clark, ’70) devoted to reptile parathyroid glands. However, no tropical lizard or gecko has been investigated since a study by Weber (‘09).
This study was carried out on three common lizard species in the Zaria area
91

92 WILLIAM SENIOR
TABLE 1
Reports of presence ( +) or absence ( - ) of parathyroid IV i n lizards
- Genus P IV Reference Group
Iguania Iguanidae
Agamidae Chamaeleontidae
Nyctisauria Gekkonidae
P ygopodidae
Leptoglossa Xantusidae Teiidae Scincidae
Lacertidae
Cordylidae Dibanidae Feyliniidae
Diploglossa Anguidae Anniellidae Xenosauridae Helodermatidae Varanidae Lanthanotidae
Annulata Amphisbaenidae
Anolis Cyclura Iguuna Phrynosomu Xiphocercus Agama
Anc ylodactylus Hemidactylus Lygodactylus Ptyodactylus Tarentola Aprasia Delma Lialis
_-_ Chalcides Gong ylus Mnbuya Scincus Tiliqua Trachysaurus Lacerta Latastiu
---
Ang u i s
Underwood ('63), Clark ('68) Underwood ('63) Underwood ('63) Adams ('53) Underwood ('63) This Study
This Study This Study This Study This Study This Study Underwood ('57) Underwood ('57) Underwood ('57)
Sidky ('65) Saint Remy and Prenant ('04) This Study Sidky ('65) Rogers ('63) Rogers ('63) Adams ('39). Peters ('41) This Study
- Saint Remy and Prenant ('04)
- Adams ('52), Sidky ('65), This Study
of Nigeria to determine the number and location of the parathyroid glands of each species and to ascertain the normal histo- logical structure of these endocrine or- gans. Occasional comparative observations were made on the occurrence and struc- ture of the parathyroid glands of other lizards in the area as they became avail- able.
Seasonal variation of the glands, and relationship between gland structure and type of calcium deposits are the subjects of further papers.
MATERIALS AND METHODS
Three species were used to determine
the structure of the parathyroid glands in detail. Agama agama agama (L)
This lizard was easily obtained around buildings in the Zaria area. It is large (50-70 gm) and does not have otic calcium deposits. In view of the presence of other species of Agama in the area (Grandison, '68), the specimens used were examined critically, and any individuals were dis- carded unless clearly Agama agama agama.
Hemidactylus broohi angulatus (Hallowell)
A small (3-6 gm) gecko which was eas-

LIZARD PARATHYROID GLANDS
ily caught on the walls of buildings at night. It has large amounts of otic cal- cium.
Ptyodactylus hasselquisti hasselquisti (Donndorff)
A rock living gecko which was relatively easily obtained. It is larger than Hemidac- ty lus (9-13 gm) and also has otic calcium deposits.
Other species processed occasionally were Latastia longicaudata, Ancylodnctylus spi- nicollis, Lygodactylus picturatus , Taren- tola ephippiata, Mabuya perrotetii and Varanus exanthemat icus .
Regular collection of the three species Agama, Hemidactylus and Ptyodactylus was carried out twice each month between March 1967 and March 1968. Five males and five females were used when available, and the largest specimens were chosen. All were dissected within 24 hours of cap- ture to prevent any changes due to shock or keeping under laboratory conditions. Specimens were weighed, measured, and then dissected to locate and remove any parathyroid tissue. No other structure was deliberately preserved.
The dissections were flooded with for- mol saline, and the arteries of the carotid fork region were removed. In some of the smaller specimens, and especially in Hemi- dactylus, the anterior part of the systemic arch was included in the tissue removed. Parathyroid I11 is located very close to the junction of the carotid vessels in all the lizards studied, and, being firmly attached, was removed with them to formol saline for fixation. Figures 1, 2 and 3 show the relationship of parathyroid I11 and the carotid arteries .
After fixation and dehydration, speci- mens were cleared in benzene-methylene green (Senior, '69) and examined in ter- pineol. At this stage, connective tissue was dissected away to reveal the parathyroid glands. When the parathyroid tissue was clearly visible the three principal dimen- sions were measured. The tissue speci- mens were then stored in fresh terpineol. Selected material was stained with May- er's carmalum (Humason, '62), and mount- ed in polyester resin (Senior, '70a).
Ehrlich's haematoxylin and eosin were used on some sections from all specimens examined histologically, making it possi- ble to compare the results obtained with
Abbwuia tioizs
93
CA, Carotid artery IC, Internal Csl, Capsule carotid artery DC, Ductus caroticus P 111, Parathyroid I11
P IV, Parathyroid IV EC, External carotid artery SA, Systemic arch
Ves, Vesicle
1 I . O h n
Fig. 1 Agamtr. Ventral view of left carotid fork region.
those of previous workers. The microana- tomical picture of the parathyroid I11 re- gion required a connective tissue stain. Greenstein's five dye stain (Greenstein, '61) was adopted as a second staining method for use on some sections of all glands used for comparative purposes. Serial sections intended for cine reconstruction (Senior, '70b) were stained only with Greenstein's method.
Parathyroid 111 glands from the left side of one male and one female Agama, Hemi- dactylus and Ptyodactylus were selected from each month of the year. Sections pro- duced from these glands were examined to determine what should be considered the normal or natural histological picture at all times of the year.
RESULTS
1. N u m b e r and location of lizard
Two hundred and fifty-two specimens of parathyroid glands

94 WILLIAM SENIOR
Fig. 2 Hemidac ty lus . Ventral view of left ca- rotid fork region.
1.0 nm.
Fig. 3 Ptyodactylus. Ventral view ,of left ca- rotid fork region.
Agama were dissected. In two males the left gland was not found, and in one male the right gland was missing. No specimen was found to have any other patches of parathyroid tissue, and in no specimen was the parathyroid I11 gland of either side in an unusual position. Parathyroid I11 in Agama was invariably found on the dorsal surface of the carotid arch, with the long axis lying along a line joining the root of the ductus caroticus to the root of the external carotid artery. In life the gland is thus very close to the thymus glands but separated from them and at the same time closely bound to the blood vessels by tough connective tissue which contains many melanocytes. The whole area was frequently a site of fat deposi- tion in well fed individuals.
Of 251 Hemidactylus dissected the right gland could not be found in one female. However, 148 patches of parathyroid tis- sue additional to parathyroid 111 were found. Parathyroid I11 was again found invariably on the dorsal surface of the carotid arch at a point between the roots
of the ductus caroticus and the external carotid artery. The other patches of para- thyroid tissue were found in the connec- tive tissue between the systemic and caro- tid arches. These were not found in all specimens, and often were not found on both sides of the same animal. In no case was more than one patch found on one side of an animal. These patches of tis- sue, although not investigated embryo- logically, are referred to as “parathyroid IV.” They possibly occur more regularly than these results show, as they lie loosely in the connective tissue and could easily be left behind in the specimen during dis- section. When parathyroid IV has been found it has been close to parathyroid I11 and has not been visible until stained.
Of the 256 Pytodactylus dissected three females and one male had no right para- thyroid I11 gland. In one male the left gland was missing. No trace of a n y other parathyroid tissue was found. Parathyroid I11 in Ptyodactylus was very frequently found on the dorsal surface of the carotid

LIZARD PARATHYROID GLANDS 95
arch between the roots of the ductus caro- ticus and the external carotid artery. How- ever, in a few specimens one gland was found more loosely attached to the carotid arch, lying in fact between the internal carotid artery and the ductus caroticus. This occurred on the right side in 11 speci- mens and on the left in four cases. On no occasion were both glands ectopic, but on two occasions one gland was ectopic and the other missing.
In the above three species it has not been possible to establish a connection be- tween ectopic or missing glands and age, sex, season, state of cdcium deposits or degree of parasitisation.
Eight specimens of Mabuya were dis- sected. In every case there were two para- thyroid I11 glands and no others. Parathy- roid 111 was found on the dorsal surface of the carotid arch near the roots of the ductus caroticus and the external carotid artery. There was a considerable amount of thick connective tissue present but it was not pigmented.
Of the one or two specimens of Ancylo- dactylus, Latastia, Lygodactylus, Taren- tola and Vararzus examined, all have had two parathyroid 111 glands. These were found on the dorsal surface of the carotid arch near the junction of the internal ca- rotid artery and the ductus caroticus. In the case of Varanus the ductus caroticus is atrophied but the position of the root on the carotid arch is indicated by a small raised pigmented region. A similar struc- ture remains on the systemic arch which was at first thought to be parathyroid IV. Although no thorough search has been made it is considered that Varanus and the other four species examined briefly do not have more than two parathyroid glands.
2. Size and shape of lizard parathyroid glands
In Agama, parathyroid 111 is rather vari- able in shape, but is most frequently a flattened ovoid applied to the blood vessel. This basic shape was measured in three dimensions which were called antero-pos- terior (AP), left-right (LR) and dorso-ven- tral (DV). The longest axis was always designated (AP), the shortest (DV) and the other (LR) regardless of the actual anatomical axis. Gland volumes were cal-
culated by multiplying the three param- eters together and dividing by two. This mathematical approximation was con- firmed by making models of typical gland shapes from a known volume of modelling clay, taking the three measurements and calculating the volume. Using 100 ml of clay and measuring to the nearest 0.5 cm with a desk ruler gave calculated volumes within 15%.
It was not possible to obtain figures for unfixed material as the thick pigmented connective tissue rendered the gland in- visible. Attempts to remove the connective tissue before fixation resulted in distortion or destruction of the gland.
The average gland shape in A g a m a is a slightly flattened ovoid with the principal axes in the proportions 1.6:l.l:l.O. Fe- males had 13% more parathyroid tissue per gram of body weight than males.
Hemidactylus females also had 13% more parathyroid tissue per gram of body weight than males. The actual amount of parathyroid I11 tissue per gram of body weight in Agama was found to be 10% more than in Hemidactylus . Those Hemi- dactylus specimens with parathyroid IV however, had a total amount almost iden- tical to that in Agama.
Parathyroid I11 in Hemidactylus was found to be less variable in shape than in Agama. The principal axes had the pro- portions 1.5: 1 .O: 1 .O.
Ptyodactylus females also had 13 % more parathyroid tissue per gram of body weight than the males, and the amount of para- thyroid tissue in this species was the same as that in Agama when related to weight. The gland shape was the most regular of the species studied and had the propor- tions 1.7: 1.15: 1 .O.
Table 2 gives the actual measurements obtained.
3. The structure of lizard parathyroid glands
Agama parathyroid glands are surround- ed by a connective tissue capsule which is continuous with the outer fibrous layer of the major blood vessels to which they are attached. The capsule was found to be generally 15 p thick, becoming much thicker near the attachment to the blood vessel and thinner at the opposite end. Melanocytes were always present. Fat is

96 WILLIAM SENIOR
TABLE 2
Size und shape of parathyroid I l l in liznrds
Agama Hemidactylus Ptyodactylus
Male Female Male Female Male Female __
Number of specimens 124 122 115 135 106 144
Average weight (gm) 53.8 25.5 4.3 3.8 10.6 7.8
Dimensions of 0.79 0.63 0.32 0.32 0.47 0.44 parathyroid 111 (mm) 0.53 0.43 0.23 0.23 0.32 0.30
0.50 0.40 0.21 0.21 0.28 0.26
Total volume of parathyroid 111 tissue (cu. mm) 0.224 0.128 0.01 7 0.016 0.045 0.038
Volume of parathyroid tissue per gram of body weight 0.0042 0.0047 0.0038 0.0043 0.0042 0.0049
always excluded from the gland by the capsule.
The connective tissue often extends into the glandular parenchyma, and in extreme cases divides the gland into several sheets or lobes. Immediately under the capsule there is a blood capillary network which extends into and through the gland in such a way that it has exactly the same appearance regardless of the plane of sec- tion. The network was seen to be supplied with blood directly through a simple aper- ture in the blood vessel wall. The channel so formed was sometimes short and straight (fig. 5) but on occasions was seen to bend and pass for a short distance within the wall of the artery and the gland capsule before turning again to pass into the glan- dular parenchyma (figs. &8).
A small vein was seen to leave the gland and pass to the internal jugular vein. The gland capsule merges with the connective tissue sheath of this vein.
Within the gland the capillaries vary in size from 15 to 50 p , though the larger sizes appeared to be spherical rather than tubular and were connected to several smaller capillary vessels. Many of the cap- illaries were not circular in cross section. The shape varied from oval to rectangular and was sometimes almost linear. This configuration is similar to that frequently found in glandular tissue with a rich blood supply. Fingerlike cords of cells are found, separated from the capillary cavities by such a thin lining of squamous epithelium that at first sight it appears as if the glan-
dular cells are in direct contact with the blood.
In most glands there were found some much smaller vessels which did not have an epithelial lining. These vessels did not contain red blood cells, but occasional white cells were seen, they ran inside the strands of glandular parenchyma rather than between them. These vessels and their contents are in some ways similar in appearance to parts of the lymphatic sys- tem, but it is not possible to state that they form part of such a system.
The glandular parenchyma in Agama consists of irregular strands or rods of cells which follow random paths amongst one another. The strands were seen to branch or to join one another forming a three di- mensional network. There has been no indication that fibres surround the paren- chyma cells or join them to the connective tissue, but rather it seems that the paren- chyma is in simple contact with the cap- sule. This view is strengthened by the fact that the parenchyma sometimes separated from the connective tissue during section cutting.
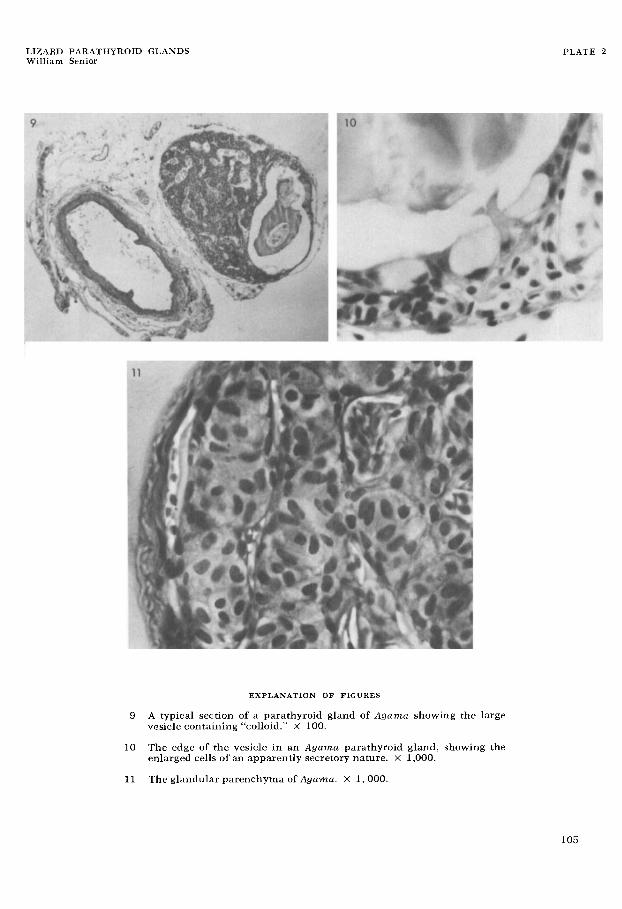
At the distal end of Agama glands there was found in about 40% of cases a rela- tively large vesicle (fig. 9) which at times occupied more than 25% of the area of a section of the gland. This has a definite structure with a distinct lining epithelium (fig. lo). The contents of the vesicle stained in a similar but not identical way to blood plasma. Some of the lining cells seem to be glandular and to be discharg-

LIZARD PARATHYROID GLANDS 97
ing their secretion into the lumen of the vesicle. Lymphocytes but not red cells have been found in the lumen, which connects by minute pores to nearby capillaries.
No evidence has been found to suggest that the vesicles were formed in reaction to parasites. Also there is no indication that the animals with glands containing vesicles were diseased or healthy, or at a particular stage of the life cycle. Males and females have shown this structure with equal frequency.
Fixed and stained parenchyma cells are generally ovoid in shape, being about 8 p across and 12 p long. The nuclei have a similar shape to the cell outline with di- mensions one-third to one-half smaller (fig. 11). Most of the cells are oriented so that the long axis runs from the centre to the surface of a strand. When a strand is seen in cross section the cells appear as spokes of a wheel. If the inner extremities of the cells do not meet, the strand has a tubular appearance. The central lumen may be an artifact, or may in life be filled with tissue fluid.
In general, differently shaped cells are scattered throughout the tissue, but in about 25% of the glands examined, patch- es of smaller rounded cells in a section were present in numbers indicating a den-
sity as high as 7.5 million per cubic milli- meter. This density is no doubt greater than in the living gland due to shrinkage during processing.
The cells of the glandular parenchyma showed considerable variations in staining properties (table 3). The cytoplasm was never as basophil as normal nuclei and was never as acidophil as the cytoplasm of the red blood cells present in the capil- laries. The presence of connective tissue and red blood cells under parenchyma cells has at times confused the picture in sec- tions 6 p thick. The red blood cells are flat oval plates with strongly acidophilic cytoplasm. They can be located in such a way that the strongly stained cytoplasm filters the light passing through another cell, thus changing the overall colour.
The nuclei of Agama parenchyma cells also showed variation in shape and stain- ing properties. Mitosis was found to be very rare. More than 1000 sections were examined but less than ten mitotic figures were seen. Occasionally some nuclei in a gland appeared shrunken with slightly ir- regular outlines. In extreme cases, spindle shaped nuclei were seen which stained brown rather than blue with Ehrlich’s haematoxylin.
When sections were stained with Green-
TABLE 3
Summary of parathyroid gland structure in lizards
Agama Hemidactylus Pytodactylus
Capsule thickness (p) 15
Capillary diameter (p) 15-50
Millions of cells per cu. mm 3.5-7.5
Normal cell size (p) 1 2 x 8
Cytoplasmic staining Basophil 70 7% Amphoteric 20 c/o Weak acidophil 2% Neutrophil 7%
Normal nuclear size (p)
Nuclear colours with Blue, Purple Greenstein’s stain Pink, Brown
7 X 5
Nuclear shapes Ovoid Prismatic Elongate
Common Present Rare
Vesicles Present
6
8-12
“4-1
Syncytium
Nil Common Present Nil
7.5 x 5
Blue-Grey, Reddish-Brown
Common Present Present
Absent
10
5-10
5.2.56.75
12 x 7.5
Common Rare Nil Present
7.5 x 5
Blue-Grey Reddish-Brown
Present Common Rare
Absent

98 WILLIAM
stein’s method the spindle shaped nuclei were again stained brown. The differen- tial uptake of acid fuchsin and haema- toxylin has not been correlated with any physical properties of the nuclei. All shapes and sizes have at times been stained blue, purple or pink in the same section.
The capsule of Hemidactylus glands was found to be identical to that of Agama ex- cept that melanocytes were not present and the average thickness was only 6 p. The blood supply to the gland is again di- rect from the carotid artery. In some cases several very narrow channels were found, of small capillary size, but usually one vessel about 10 p across pierced the ar- tery wall. Occasionally a single vessel was seen to leave the gland and pass to the internal jugular vein.
Hemidactylus glands are much less vas- cular than those of Agama, the paren- chyma is not divided to the same extent into strands of cells and there is very little division of the parenchyma by connective tissue. No channels other than capillaries have been found. The capillaries contain only a few red blood cells, and these seem almost too large to pass through the small- er vessels. It is possible that in life the blood pressure is high enough within the gland to engorge the capillaries and so in- crease their cross sectional area.
The parenchyma strands are not clearly defined, but seem to be shorter separate lengths than in Agama (fig. 13). Cell mem- branes have not been revealed by the methods used in this study even when sec- tions have been examined under phase contrast. It is possible that in Hemidac- ty lus the glandular parenchyma is largely if not entirely syncytial in nature. This would explain the frequent occurrence of nuclei with irregular outlines of a shape indicating that they had been in contact. In some areas the nuclei were tightly packed whilst in others a more normal open pattern was seen. Also, the cytoplasm stained uniformly in these glands.
The most common shape of nucleus in Hemidactylus is an ovoid 7.5 by 5 p. Greenstein’s method sometimes stained the tightly packed nuclei a darker colour leading to reddish brown. No mitotic fig- ures have been seen.
The structure of parathyroid IV in Hem-
SENIOR
idactylus is not markedly different from that of parathyroid 111. All the morpho- logical differences observed can be re- lated to the difference in size of the glands. During dissection of a few very large specimens, a minute blood vessel was seen to leave parathyroid IV and pass to the internal jugular vein. The blood supply to the gland may possibly be from the vessel leaving parathyroid 111. Very few capillaries have been found within the glandular parenchyme.
The connective tissue capsule of Ptyo- dactylus glands is integral with the fibrous coat of the carotid artery wall to which it is attached (fig. 14). The fibrous layer of the artery wall is only half its normal thickness alongside the parathyroid gland. Removal of the gland by physical means has resulted in the rupture of the carotid artery due to this structure. Parathyroid- ectomy has only been possible by electro- cautery.
The capsule is generally 10 p thick, and is often integral with the walls of the in- ternal carotid artery and the ductus ca- roticus as seen in a transverse section of the region of attachment. However, the attachment to the ductus caroticus is never so complete, as that to the internal carotid artery.
The blood supply to the parathyroid glands of Pytodactylus is by means of a series of perforations of the carotid artery direct to the capillary network. No connec- tion with the ductus caroticus has been observed. The capillaries close to the sup- plying perforations are strengthened so that in thin sections they appear to be run- ning in sheets of connective tissue. Also these vessels are seen to be at first lined with a continuation of the inner layers of the nearby artery and should therefore be classed as arterioles rather than capil- laries. The capillaries join together in the distal region to form a single thin walled vessel leaving the gland. It is probable but not certain that this vessel eventually con- nects with the internal jugular vein.
A clear channel was frequently found in the centre of parenchyma strands in Ptyodactylus. No evidence was obtained of the relationship of these structures with the body lymphatic system.
The glandular parenchyma strands were found to be shorter and less intertwined

LIZARD PARATHYROID GLANDS 99
than in Agama. Some of the cells were in spherical groups. In these cell groups the long axis of any nucleus is generally found to be oriented along a radius. Not all the cell groups are clearly delineated but the general configuration of a lizard parathy- roid gland is more easily understood in this species (fig. 15).
The cells of the glandular parenchyma in Ptyodactylus have well marked bound- aries. The most frequent cell shape is an ovoid narrower at one end. The nuclei are very variable in shape, but when ovoid are 7.5 by 5 ,.A. Some very large cells were observed in most of the glands examined. These measured up to 15 by 10 p with nu- clei of a normal type and size.
The cytoplasm of the parenchyma cells was always basophil and, apart from the largest cells, appeared very granular in nature. The amount of stain taken up var- ied from gland to gland and there was some variation within glands. The very large cells were sometimes almost un- stained but were never acidophilic. In Ptyodactylus the parathyroid cell cyto- plasm stained in an almost identical way to that of the blood plasma, though the plasma was not granular.
The nuclei in Ptyodactylus glands are more frequently angular than in the other two species, but the staining properties are much less variable. It is of interest that with Greenstein’s method the nuclei of the red blood cells always stained pink, whilst the white blood cell nuclei stained blue-grey llke the parathyroid cell nuclei. No mitotic figures were observed in Ptyo- dactylus glands.
Glands from other lizards processed for histological study have not shown any structure or property not present in the three species studied in detail. It is not possible to comment on the shape or size of these glands except to state that the few examples seem to conform to the same general size in comparison with body size, and no extremely different shapes have been seen.
The geckos Lygodactylus and Ancylo- dactylus showed a structure similar to that of Hemidactylus which is of similar size, and Tarentola, another gecko, was more similar to Ptyodactylys which is a similar size. Latastza, a lacertid from the infraorder Scincomorpha had a structure
very similar to that of Agarna from the in- fraorder Iguania. These two species were the only ones in which large vesicles were found within the parathyroid glands. Ma- buya , another member of the Scincomor- pha was different in structure from La- tastia and was in fact similar to Varanus from the infraorder Platynota.
Mabuya and Varanus both had extreme- ly uniform glandular parenchyma which did not show any marked division into groups of cells. In both cases the glands were ramified by prominent blood capil- laries supplied with blood through pores in the wall of the carotid artery and drain- ing to the internal jugular vein. The cyto- plasm and the nuclei both stained uni- formly in the few specimens processed.
DISCUSSION
According to Gabe and Saint-Girons (‘70) all studies of lizards have indicated that the normal number of parathyroid glands is one pair, and that these are derived from the third branchial pouch. Agama and Ptyodactylus can therefore be consid- ered to conform to the general lizard pat- tern. Microscopic sections of A g a m a glands sometimes gave the impression that two separate bodies were present. However, reconstruction of these bodies revealed a single structure with a forked end (fig. 4).
Hemidactylus frequently showed sepa- rate patches of tissue additional to the main pair of glands. An embryological study is needed to determine whether or not these structures are derived from the fourth branchial pouches, but the evidence obtained in this study indicates rather that they are probably isolated patches derived from parathyroid 111. The patches occurred more frequently at certain times of the year, they occurred in young and old males and females, and they were not closely applied to the aortic arch. A true parathyroid IV would be expected to occur more frequently in young animals and to be closely connected with the aortic arch.
The normal location of parathyroid 111 in all the lizards studied at Zaria was found to be on the dorsal surface of the carotid arteries, close to the carotid fork. It is possible that the few cases of para- thyroid I11 found loose in the connective tissue were due to damage during removal in the first dissection. This is again in

100 WILLIAM SENIOR
D C D C
Dorsal View of Right Carotid Fork
0.5 -. Fig. 4 Agctma. Various shapes ofgland and appearance in section.
strict accord with the collected informa- tion of Gabe and Saint-Girons ('70).
It is of interest that two species as dif- ferent as Agama and Ptyodactylus should have practically the same amount of para- thyroid tissue when expressed as volume per unit weight. Hemidactylus has less when only parathyroid I11 is considered, but when a correction is made for the presence of the additional patches of tis- sue, the final figure is very similar to that found in the other two species. Again, Mabuya seems to have a similar amount of parathyroid tissue considering that the figures were obtained from only a few specimens.
It is possible that the amount of para- thyroid tissue per gram of body weight normally present in lizards is fairly con- stant, and it is also possible that if a spe- cies is found to have very much less than this amount, a search for additional tissue would be indicated.
The volume of parathyroid tissue and
body weight are not clearly stated in the literature on other tetrapod groups or other reptiles. Most references give sizes of between 0.5 and 1.0 mm diameter for reptilian and amphibian parathyroid glands, but the weight of the animals con- cerned is not stated, and emphasis tends to be on a comparative change in size of the glands. An investigation is needed to determine whether or not the average amount of parathyroid tissue normally present in tetrapods as a whole is reason- ably constant. The evidence so far ob- tained does not preclude such a possibility.
The shape of lizard parathyroid glands seems to be less spherical than that men- tioned for amphibia (Gorbman and Bern, '62) and other reptile groups (Clark, '70). The shape can vary considerably when there is a large amount of fat in the area, but spherical glands have not been found apart from some of the additional patches of tissue in Hemidactylus . The slight flat- tening of the ovoid shape is probably con-

LIZARD PARATHYROID GLANDS 101
nected with the close binding of the gland to the carotid artery with the surrounding connective tissue. Gabe and Saint-Girons ('70) consider the normal shape to be a disc or a flattened ovoid.
The three major components of a lizard parathyroid gland are the connective tis- sue sheath, the glandular parenchyma and the blood capillary network. Nervous or muscular components have not been found.
Studies of the blood supply of parathy- roid glands in lizards and of the capil- laries within the glands do not appear to have been carried out in detail. Gabe and Saint-Girons ('70) mention that the blood supply is by means of arterioles coming from the carotid arch. It was found in this study that the usual blood supply was by simple apertures in the internal carotid artery leading direct to the capillary net- work within the gland. A strong capsule is therefore necessary to withstand the re- sulting high blood pressure.
Observation of the gland was sometimes possible before removal from a specimen whilst the heart was still beating. In these cases the gland was seen to pulsate in rhythm with the heart beat. The elas- ticity of the capsule thus permitted the capillaries to enlarge under high blood pressure, but expelled blood from the gland as the pressure fell. The capillaries of the parathyroid gland can be consid- ered to open directly to the carotid artery.
The single small blood vessel which drains a lizard parathyroid gland seems too small to carry away blood at low pres- sure as fast as it is delivered to the gland under high pressure. It is therefore likely that some of the blood, having entered the gland, is returned to the arterial trunk. Whether or not red blood cells are re- turned to the arterial trunk is not known, but the capillaries in the processed glands were frequently packed tight with red cells as if plasma had been lost. It may be that this packing between pulsations, if it occurs in life, gives an increased oxy- gen supply to the parenchyma.
The strong capsule of reptile parathy- roid glands in general and of lizards in particular is mentioned frequently in the literature. In general terms, the findings of this study agree with previous reports that the capsule thickness is largely de-
pendent upon the size of the individual (Peters, '41 ; Adams, '52).
Previous descriptions of the parenchyma of lizard parathyroid glands indicate that there are overall similarities but species differences. This study shows the same to be true. The parenchyma of the para- thyroid glands of Agamu, Hemidactylus and Ptyodactylus was found to be suffi- ciently different to allow identification of the species from an examination of a stained microscope slide. Other species which have been examined also show pat- terns of internal structure which seem to be characteristic for each species whilst showing similarities with one or other of the three species studied in detail.
The vesicle found in many Agama glands has the appearance of an enlarged lumen of a cell strand. This is especially so with the smaller sized vesicles. However, when large vesicles are examined, the lining parenchyma is different from that of the rest of the gland. A similar situation is re- ported in three species of Lacerta by Pe- ters ('41). Some glands have been found with a very large vesicle having only a single layer of cells between the lumen and the capsule. The impression given is that the interior of the gland is breaking down and that the vesicle has become a receptacle for cellular debris. If this is so, it could be related to the seasonal varia- tion in parathyroid gland structure re- ported in some anurans (Waggener, '29), and some lizards (Peters, '41; Sidky, '65).
The occurrence of the vesicle in Agama is more frequent between January and April than during the rest of the year, but has been recorded for every month. This lizard has no inactive period in tropical regions comparable with that of lacertids of temperate regions. Such a period of in- activity would, however, be most advan- tageous to Agama if it did in fact occur between January and April when food is very scarce.
The general appearance of the glandu- lar parenchyma of Hemidactylus is similar to that reported by von Brehm ('64) and earlier workers for anurans except that in Hemidactylus there is no clear indication of the presence of cell membranes, and the cells do not easily separate during processing. A peripheral zone of more transparent cells is limited to a single

102 WILLIAM SENIOR
layer under the capsule, and sometimes lining the larger capillaries. This type of configuration is not reported in mamma- lian glands.
The parenchyma of Agama parathyroid glands is, in contrast, closely comparable with that of some mammals. Lever ('57, '62) noted a grouping of cells of different appearances in the rat parathyroid. Such cell groups are often seen in Agama. The differences in staining of Agama paren- chyma cells are often seen also in mam- malian glands as differences in basophilia (Davis and Enders, '61). It seems that in Agama there is only one major type of cell present. This is in accord with the obser- vations of Clark ('65) on turtles, Hohn ('61) on birds, and Lever ('57) on rodents, but contrasts with the structure found in most other mammals, including man, where at least two kinds of cell are recog- nised.
Ptyodactylus glands are essentially sim- ilar to those of Agama in that the cell boundaries are easily seen, and a lobular appearance is presented. This is again very similar to the turtle, bird and rodent pattern but different from that of anurans and Hemidactylus . Stained sections of Ptyodactylus glands show intercellular material present especially at the centre of the lobules. It is possible that the spaces containing such material are smaller in the living animal due to the high blood pressure acting on the outside of the lob- ules and pressing the cells towards the central lumen.
In brief, it may be said that lizard para- thyroid glands are more similar to those of mammals and birds than they are to those of anurans. The general appear- ance of bird parathyroid glands is the most similar to lizard glands, but is especially similar to Agama. One major difference is that at no time has any trace of fat been found inside the capsule of a lizard parathyroid gland, whereas fat is com- monly reported in mammalian and bird glands.
ACKNOWLEDGMENTS
The author is greatly indebted to Pro- fessor A. P. Mead, Principal of the Col- lege of Science and Technology, Port Har- court, Nigeria, for suggesting the topic of study and for guidance throughout the
period of investigation. Also the design and running of the computer program used to process the results was by Mr. A. H. Hartley, Department of Mathemat- ics, Ahmadu Bello University, Zaria, Ni- geria.
LITERATURE CITED
Adams, W. E. 1939 The cervical region of the Lacertilia. - A critical review of certain aspects of its anatomy. J. Anat., 74: 57-71.
1952 The carotid sinus complex and epithelial body of Vnrnnus van'us. Anat. Rec., 113: 1-27.
1953 The carotid arch in lizards with particular reference to the origin of the internal carotid artery. J. Morph., 92. 115-155.
Baker, J. R. 1966 Cytblogical Technique. Meth- uen and Co., London.
Boschwitz, D. 1961 The parathyroids of Bufo viridis Laurenti. Herpetologica, 17: 192-199.
Clark, N. B. 1965 Experimental and histological studies of the parathyroid glands of fresh water turtles. Gen. comp. Endocr., 5: 297-312.
1967 Parathyroid glands in reptiles. Am. Zool., 7: 869-881.
1968 Effect of parathyroidectomy in the lizard, Anolzs carolinensis. Gen. comp. Endocr.,
The parathyroid. In: Biology of the Reptilia. C. Gans and T. S. Parsons, eds. Mor- phology C. Volume 3. Academic Press, London, pp. 235-262.
Cortelyou, J. R. 1962 Phosphorus changes in to- tally parathyroidectomised Rnnn pipiens. Endo- crinology, 70. 618-621.
Cortelyou, J. R., A. Hibner-Owerko and J. Mulroy 1960 Blood and urine calcium changes in to- tally parathyroidectomised Rana p ip iens . Endo- crinology, 66: 441-450.
Davis, R., and A. C. Enders 1961 Light and elec- tron microscope studies on the parathyroid gland. In: The Parathyroids. R. 0. Greep and R. V. Talmage, eds. Charles C Thomas, Spring- field, Illinois, pp. 76-92.
Gabe, M., and H. Saint-Girons 1970 In: Trait6 de Zoologie. P.-P. Grass&, ed. XIV (111). Masson et Cie, Paris, pp. 759-770.
Goin, C. J., and 0. B. Goin 1962 Introduction to Herpetology. W. H. Freeman and Co., San Francisco.
Gorbman, A., and H. J. Bern 1962 A Textbook of Comparative Endocrinology. John Wiley and Sons, New York.
Grandison, A. G. C. 1968 Nigerian lizards of the genus Agamu (Sauria: Agamidae). Bull. Br. Mus. nat. Hist. (Zool.), 17: 3-90.
Greenstein, J. S. 1961 A simplified five-dye stain for sections and smears. Stain Technol., 36: 87- 88.
Greep, R. 0. 1963 Comparative Endocrinology. Vol. 1. U. S. von Euler and H. Heller, eds. Aca- demic Press, New York, pp. 325370.
Hohn, E. 0. 1961 Endocrine glands, thymus and pineal body. In: The Biology and Com- parative Physiology of Birds. Vol. 2. A. J. Mar- shall, ed. Academic Press, New York, pp. 87- 114.
10: 99-102. 1970

LIZARD PARATHYROID GLANDS 103
Humason, G. L. 1962 Animal Tissue Techniques. W. H. Freeman and Co., San Francisco.
Itakura, C., and S. Yamagiwa 1970 Histopatho- logical studies on bone dysplasia of chickens. 11. Histopathology of the parathyroid gland. Jap. J. vet. Sci., 32: 243-249.
Lever, J. D. 1957 Fine structural appearances i n the rat parathyroid. J. Anat., 91 : 73-81.
Fine structural organisation in en- docrine tissues. Br. med. Bull., 18: 229-233.
Peters, H. 1941 Morphologische und experi- mentelle Untersuchungen iiber die Epithelkorper bei Eidechsen. Z. mikrosk.-anat. Forsch., 49: 1-40,
Polin, D., P. D. Sturkie and W. Hunsaker 1957 The blood calcium response of the chicken to parathyroid extracts. Endocrinology, 60: 1-5.
Rogers, D. C. 1963 A cytological and cytochem- ical study of the “epithelial body” on the carotid artery of the lizards Trachysaurus rugosus and Tiliqua occipitalis. Q. J1. microsc. Sci., 104: 197-205.
Saint Remy, G., and A. Prenant 1904 Re- cherches sur la d6veloppement des dkriv6s bran- chiaux chez les Sauriens et les Ophidiens. Archs. Biol., 20: 145-216.
Senior, W. 1969 Staining of animal tissues with the dye base of methylene green in benzene to facilitate identification and selection of material. Stain Technol., 44: 26S271.
1970a Polyester resin as a mounting medium for light microscopy. J. Microsc., 91 207-210.
1970b Reconstruction of microscopic objects by photographing serial sections onto cine film for projection. J. Microsc., 92: 223- 228.
Sidky, Y . A. 1965 Histological studies on the parathyroid glands of lizards. Z. Zellforsch., 65: 760-769.
Simkiss, K. 1961 Calcium metabolism during avian reproduction. Biol. Rev., 36: 321-367.
1962
Taylor, T. G., and F. Hertelendy 1961 Changes in the blood calcium associated with eggshell calcification in the domestic fowl. 2. Changes in the diffusible level. Poult. Sci., 40: 115-123.
Taylor, T. G., T. R. Morris and F. Hertelendy 1962 The effect of pituitary hormones on ovu- lation in calcium deficient pullets. Vet. Rec., 74: 123-125.
Underwood, G. 1957 On lizards of the family Pygopodidae. (A contribution to the morphol- ogy and phylogeny of the Squamata.) J. Morph., 100: 207-268.
1963 In: Comparative Endocrinology. Vol. 1. U. S. von Euler and H. Heller, eds. Aca- demic Press, New York, p. 345.
Urist, M. R. 1959 The effects of calcium de- privation upon the blood, adrenal cortex, ovary and skeleton in domestic fowl. Recent Prog. Horm. Res., 15: 455-477.
Urist, M. R., N. M. Deutsch, G. Pomerantz and F. C. McLean 1960 Interrelations between actions of parathyroid hormone and estrogens on bone and blood in avian species. Am. J. Physiol., 199: 851-855.
von Brehm, H. 1964 Experimentelle Studie zur Frage der Jahreszyklischen Veranderungen. Mo- phologische Untersuchungen an Epithelkor- perchen (Glandulae parathyreoideae) von An- uren. 11. Z. Zellforsch. mikrosk. Anat., 61: 725- 741.
von Euler, U. S., and H. Heller, eds. 1963 Com- parative Endocrinology. Acadcmic Press, New York.
Waggener, R. A. 1929 A histological study of the parathyroids of the Anura. J. Morph. Phys- iol., 48: 1-43.
1930 An experimental study of the para- thyroids in the Anura. J. Exp. Zool., 57: 13-55.
Weher, A. 1909 Recherches cytologiques sur la secrktion des glandes parathyroides du gecko. C. r. SBanc. Soc. Biol., 67: 17-18.

LIZARD PARATHYROID GLANDS William Senior
EXPLANATION O F FIGURES
5 Agnmcc parathyroid gland blood supply, a single almost straight channel leads from the carotid artery to the glandular tissue. x 250.
The blood supply is less direct than in figure 5. Opening from the carotid artery (fig. 6) a blood vessel passes for a short distance in the connective tissue (fig. 7) before opening into the parathyroid gland parenchyma (fig. 8). x 250.
G 8
PLATE 1
104
LIZARD PARATHYROID GLANDS William Senior
EXPLANATION O F FIGURES
9 A typical section of a parathyroid gland of Agama showing the large vesicle containing “colloid.” X 100.
The edge of the vesicle in an Agama parathyroid gland, showing the enlarged cells of an apparently secretory nature. X 1,000.
11 The glandular parenchyma of Agumu. X 1,000.
10
PLATE 2
105

LIZARD PARATHYROID GLANDS William Senior
EXPLANATION OF FIGURES
12 A typical section of the parathyroid glands of Hemiductglus , showing the similarity of structure of the two bodies. X 250.
13 The glandular parenchyma of Hemiductylus showing closely packed nuclei. X 1,000.
PLATE 3
106

LIZARD PARATHYROID GLANDS William Senior
EXPLANATION O F FIGURES
14 A typical section of a parathyroid gland of Ptyoductylus which also shows the entry and exit points of the blood supply. x 250.
15 The glandular parenchyma of Ptyodactylus showing the “wheel” pattern. X 1,000.
PLATE 4
107