Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Sept. 2004, p. 9016–9029 Vol. 78, No. 170022-538X/04/$08.00�0 DOI: 10.1128/JVI.78.17.9016–9029.2004
Palindrome Regeneration by Template Strand-Switching Mechanism atthe Origin of DNA Replication of Porcine Circovirus via the
Rolling-Circle Melting-Pot Replication ModelAndrew K. Cheung*
Virus and Prion Diseases of Livestock Research Unit, National Animal Disease Center,USDA Agricultural Research Service, Ames, Iowa
Received 8 March 2004/Accepted 30 April 2004
Palindromic sequences (inverted repeats) flanking the origin of DNA replication with the potential offorming single-stranded stem-loop cruciform structures have been reported to be essential for replication ofthe circular genomes of many prokaryotic and eukaryotic systems. In this study, mutant genomes of porcinecircovirus with deletions in the origin-flanking palindrome and incapable of forming any cruciform structuresinvariably yielded progeny viruses containing longer and more stable palindromes. These results suggest thatorigin-flanking palindromes are essential for termination but not for initiation of DNA replication. Detectionof template strand switching in the middle of an inverted repeat strand among the progeny viruses demon-strated that both the minus genome and a corresponding palindromic strand served as templates simulta-neously during DNA biosynthesis and supports the recently proposed rolling-circle “melting-pot” replicationmodel. The genome configuration presented by this model, a four-stranded tertiary structure, provides insightsinto the mechanisms of DNA replication, inverted repeat correction (or conversion), and illegitimate recom-bination of any circular DNA molecule with an origin-flanking palindrome.
The rolling-circle replication mechanism has been demon-strated in the replication of circular DNAs of phages (32),bacterial plasmids (7, 15), plant geminiviruses (11, 13, 34), andanimal DNA viruses (10, 28, 29). The rolling-circle cruciformreplication model postulates that a replicator initiation protein(Rep) binds its cognate sequence at the origin of DNA repli-cation, unwinds the double-stranded DNA, and causes extru-sion of two single-stranded stem-loops to generate a cruciformstructure. Rep then nicks the cognate sequence present in theplus strand to generate a free 3�-OH end for new leading-strand DNA synthesis. During geminivirus replication (11, 13,34), the closed circular single-stranded genome is first con-verted to a superhelical double-stranded DNA replication in-termediate. The virus-encoded Rep binds to and nicks (indi-cated by 2) between the seventh T and the eighth A of a“conserved” nonanucleotide (TAATATT2AC) at the originto initiate plus-strand DNA replication. This nonanucleotide ispresent among all members of the Geminiviridae family and isflanked by two inverted repeat (palindromic) sequences, whichcan potentially base pair together to form the stems of a cru-ciform structure during DNA replication.
Similar to geminiviruses, porcine circovirus (PCV) of theCircoviridae family has a closed circular single-stranded DNAgenome (21, 35, 40). Two genotypes of PCV have been iden-tified. PCV type 1 (PCV1) is nonpathogenic, while PCV2 hasbeen implicated as the etiological agent of a new disease,named postweaning multisystemic wasting syndrome (1). Thegenome nucleotide sequences of a number of PCV1 (1,759bases) and PCV2 (1,768 bases) isolates (2, 8, 12, 22, 23, 27)have been determined. It has been suggested that the PCV
genome is an intermediate between the genomes of geminivi-rus and plant circovirus (renamed nanovirus) (30, 36) and thatanimal circovirus was derived from a plant virus (probably aplant nanovirus) that switched hosts (via an insect vector) toinfect a vertebrate and then recombined with a vertebrate-infecting virus (probably a single-stranded RNA virus such asa calicivirus) (9).
The origin of PCV1 has been mapped to a 111-bp fragment(20) which contains a nonanucleotide (TAGTATT2AC) (Fig.1A) similar to that of geminiviruses. Recent work showed thatthe PCV2 nonanucleotide (AAGTATT2AC) can be furthercondensed to an octanucleotide motif sequence (AxTAxTAC)and is essential for PCV DNA replication (5). This octanucle-otide is flanked by a pair of 11-nucleotide palindromic se-quences. PCV DNA replication requires two Rep-associatedproteins, Rep and Rep� (REP complex) (2, 3, 18, 19). In vitroexperiments showed that PCV1 Rep binds to the right arm ofthe presumed stem-loop, while both Rep and Rep� bind to twoadjacent, almost perfect 6-nucleotide (CGGCAG or CGTCAG) tandem direct repeats located at nucleotide 13, 19, 30,and 36 (Fig. 1A) (39). However, in vivo analysis demonstratedthat Rep may interact with the palindromic stem only via the Cnucleotide at positions 3 and 10 (4). The presence of the“conserved” octanucleotide flanked by inverted repeats at theorigin and similarities among the Rep proteins essential forvirus replication suggest that PCV DNA may replicate via therolling-circle “cruciform” model proposed for the Mastrevirusgenus of the Geminiviridae family (18, 34) with modificationsspecified by the recently proposed rolling-circle “melting-pot”model (4).
The origin configuration of the melting-pot model differsfrom that of the rolling-circle cruciform model. In the melting-pot model, the REP complex binds its cognate sequence(AGTATT2AC), which is embedded in the 12-nucleotide
* Mailing address: National Animal Disease Center, P.O. Box 70,Ames, IA 50010. Phone: (515) 663-7497. Fax: (515) 663-7458. E-mail:[email protected].
9016
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
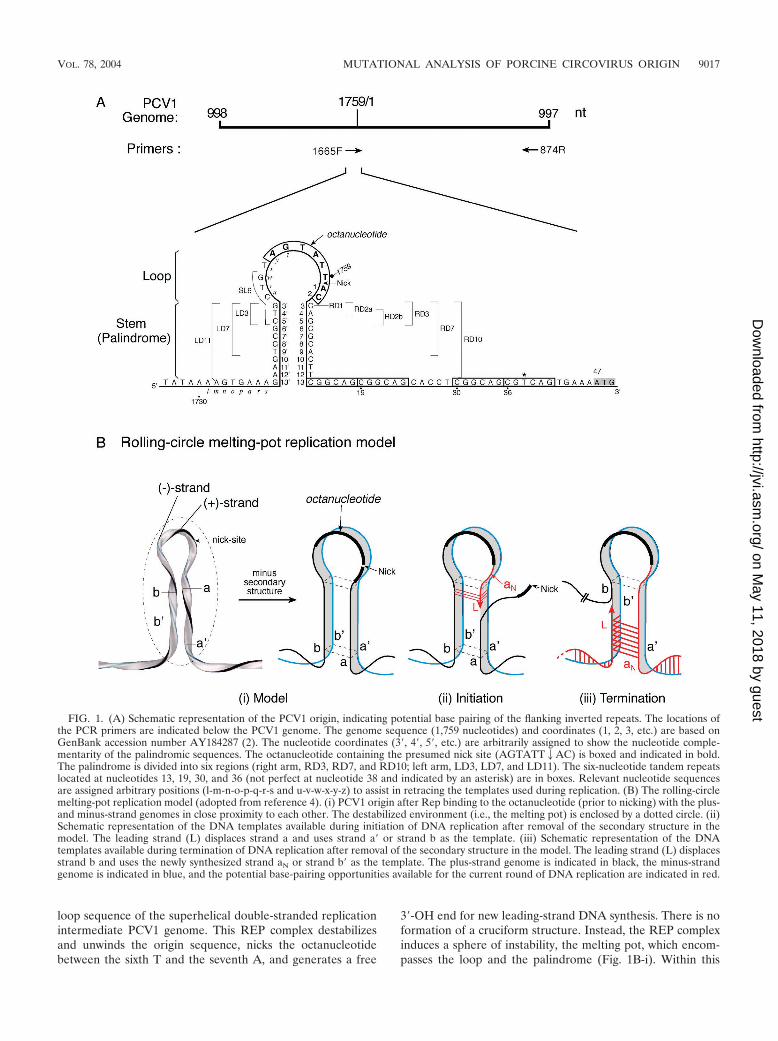
loop sequence of the superhelical double-stranded replicationintermediate PCV1 genome. This REP complex destabilizesand unwinds the origin sequence, nicks the octanucleotidebetween the sixth T and the seventh A, and generates a free
3�-OH end for new leading-strand DNA synthesis. There is noformation of a cruciform structure. Instead, the REP complexinduces a sphere of instability, the melting pot, which encom-passes the loop and the palindrome (Fig. 1B-i). Within this
FIG. 1. (A) Schematic representation of the PCV1 origin, indicating potential base pairing of the flanking inverted repeats. The locations ofthe PCR primers are indicated below the PCV1 genome. The genome sequence (1,759 nucleotides) and coordinates (1, 2, 3, etc.) are based onGenBank accession number AY184287 (2). The nucleotide coordinates (3�, 4�, 5�, etc.) are arbitrarily assigned to show the nucleotide comple-mentarity of the palindromic sequences. The octanucleotide containing the presumed nick site (AGTATT2AC) is boxed and indicated in bold.The palindrome is divided into six regions (right arm, RD3, RD7, and RD10; left arm, LD3, LD7, and LD11). The six-nucleotide tandem repeatslocated at nucleotides 13, 19, 30, and 36 (not perfect at nucleotide 38 and indicated by an asterisk) are in boxes. Relevant nucleotide sequencesare assigned arbitrary positions (l-m-n-o-p-q-r-s and u-v-w-x-y-z) to assist in retracing the templates used during replication. (B) The rolling-circlemelting-pot replication model (adopted from reference 4). (i) PCV1 origin after Rep binding to the octanucleotide (prior to nicking) with the plus-and minus-strand genomes in close proximity to each other. The destabilized environment (i.e., the melting pot) is enclosed by a dotted circle. (ii)Schematic representation of the DNA templates available during initiation of DNA replication after removal of the secondary structure in themodel. The leading strand (L) displaces strand a and uses strand a� or strand b as the template. (iii) Schematic representation of the DNAtemplates available during termination of DNA replication after removal of the secondary structure in the model. The leading strand (L) displacesstrand b and uses the newly synthesized strand aN or strand b� as the template. The plus-strand genome is indicated in black, the minus-strandgenome is indicated in blue, and the potential base-pairing opportunities available for the current round of DNA replication are indicated in red.
VOL. 78, 2004 MUTATIONAL ANALYSIS OF PORCINE CIRCOVIRUS ORIGIN 9017
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

destabilized environment, all four strands of the inverted re-peats (strands a, a�, b, and b�) are in a “melted” state and arejuxtaposed in a four-stranded tertiary structure.
After the REP complex nicks the octanucleotide, DNA rep-lication proceeds with the leading strand descending into thepalindromic stem portion of the melting pot through the rightarm and displacing the old strand a. Both minus-genomestrand a� and inverted repeat strand b are available as tem-plates (Fig. 1B-ii). For DNA termination, the leading strandascends into the melting pot through the left arm to displacethe old strand b. Both the minus-genome strand b� and thenewly synthesized palindromic strand aN are available as tem-plates (Fig. 1B-iii). Therefore, two DNA strands are availableas templates via template strand switching (i.e., from the minusgenome to the corresponding palindromic strand) during ini-tiation as well as termination of DNA replication. For thisreason, the rolling-circle melting-pot replication model permitsthe use of either the left-arm or right-arm inverted sequence toregenerate the wild-type palindrome (inverted repeat correc-tion, previously referred as gene correction) (38) or to adopt amutilated sequence to form a new palindrome (inverted repeatconversion) in the progeny virus (4).
In this study, the experiments were designed to examine themechanisms involved in palindrome regeneration by self-DNAreplication during production of infectious progeny viruses.This was done via deletion mutagenesis of either or both armsof the 11-nucleotide palindrome present at the origin of PCV1.
MATERIALS AND METHODS
Cell and virus propagation. Viruses were propagated in PCV1-free PK15 celllines maintained in minimal essential medium-Hank’s balanced salt solution(MEM-H) (Life Technologies, Grand Island, N.Y.) supplemented with 10%fetal calf serum.
DNA mutagenesis. Specific deletions were introduced into the cloned PCV1genome according to the manufacturer of the QuickChange site-directed mu-tagenesis kit (Stratagene, San Diego, Calif.). The primer sets for mutagenesiswere designed to contain 14 to 20 identical nucleotides on either side of thepredetermined deletion (indicated by lowercase letters), and only one strand ofeach mutagenic primer set is listed. The primers were as follows: LD3, AAAAGTGAAAGAAGTGCG(ctg)CTGTAGTATTACCAGCGCA; SL6, AAAAGTGAAAGAAGTGCG(ctgctg)TAGTATTACCAGCGCACT; LD7, CTATAAAAGTGAAAGAAG(tgcgctg)CTGTAGTATTACCAGCGCA; LD11, CATCCTATAAAAGTGAAA(gaagtgcgctg)CTGTAGTATTACCAGCGCA; RD1, TGCTGTAGTATTAC(c)AGCGCACTTCGGCA; RD2a, TGCTGTAGTATTAC(ca)GCGCACTTCGGCAG; RD2b, TGCTGTAGTATTACC(ag)CGCACTTCGGCAG; RD3, CGCTGCTGTAGTATTAC(agc)CGCACTTCGGCAGCGGCA; RD7, TGCGCTGCTGTAGTATTAC(cagcgca)CTTCGGCAGCGGCAGCACC; RD10, TGCGCTGCTGTAGTATTAC(cagcgcactt)CGGCAGCGGCAGCACCTC; Del3, TAAAAGTGAAAGAAGTGCG(ctg)CTGTAGTATTAC(agc)CGCACTTCGGCAGCGGCAG; Del7, TCCTATAAAAGTGAAAGAAG(tgcgctg)CTGTAGTATTAC(cagcgca)CTTCGGCAGCGGCAGCACCT; andDel10, TCATCCTATAAAAGTGAAAG(aagtgcgctg)CTGTAGTATTAC(cagcgcactt)CGGCAGCGGCAGCACCTCGG.
Transfection. After the viral genome was excised from the Bluescript plasmidand recircularized by T4 DNA ligase, the ligated DNA mixture was transfectedinto PK15 cells which had been seeded into 48-well tissue culture plates so thatapproximately 60 to 80% confluency was reached 24 h later. Transfection of theligated DNA mixture (0.5 �g) was carried out with a commercially availableLipofectamine reagent (30 �g/ml) in MEM-H. The DNA/Lipofectamine mixture(0.3 ml) was dispensed into each culture which had been freshly rinsed withMEM-H. After incubation for 5 h at 37 C, the DNA-Lipofectamine mixture wasreplaced with MEM-H with 10% fetal bovine serum.
Immunochemical staining. PK15 monolayer cells seeded in 48-well cultureplates were infected with virus or transfected with DNA. At 48 h, the cells wererinsed with water, fixed in a phosphate-buffered saline solution containing 40%acetone and 0.2% bovine serum albumin (�20°C) for 10 min, and dried for 30
min at 37°C. The cells were then incubated with a hyperimmune swine serum thatreacts with the Rep-associated proteins of PCV1 (2) diluted in binding buffer(0.01% Tween 20 and 0.5 M NaCl in phosphate-buffered saline) for 1 h at roomtemperature, washed twice with phosphate-buffered saline containing 0.05%Tween 20 (PBSW), incubated with protein G conjugated with horseradish per-oxidase (1:1,000) (Zymed Labs, Inc., San Francisco, Calif.) for 30 min, and rinsedwith PBSW twice. Color development was carried out with 3-amino-9-ethylcar-bazole and hydrogen peroxide in 0.05 M sodium acetate buffer (pH 5). Viralantigens were stained reddish brown in this assay.
PCR. Total cell DNAs were isolated with the STAT-60 DNA extraction kitpurchased from Tel-Text B, Inc. (Friendswood, Tex.). The cells were lysed andextracted with chloroform according to the manufacturer’s instructions. Theorigin sequence was amplified with oligonucleotide primers 1665F (CCAAGATGGCTGCGGGGG) and 874R (GTAATCCTCCGATAGAGAGC) (2). PCRwas carried out with 1 �g of DNA in the presence of 10 mM Tris-HCl (pH 8.3),0.2 mM each of the four deoxynucleoside triphosphates, 100 pM each of theprimers, and 2.5 U of Taq polymerase. The reaction mixture was amplified for 45cycles at 94°C for 10 s, 55°C for 30 s, and 70°C for 30 s and then kept at 70°C for10 min.
RESULTS
Construction of mutant genomes. A previously describedPCV1 genomic clone (J1) (2, 3) was employed to construct themutated genomes used in this study. A schematic representa-tion of the single-stranded plus-strand origin is denoted in Fig.1A. The octanucleotide sequence encompassing the presumednick site between the first nucleotide (position 1) and the lastnucleotide (position 1759) is enclosed in a box. A series ofmutations were engineered into the right arm (starting at nu-cleotide 3 of the genomic sequence) (2), the left arm, or botharms of the palindrome to destabilize the potential stem-loopstructure. A collection of 13 mutant genomes, organized intothree groups, were constructed. Those in the left arm arenamed LDx, those in the right arm are named RDx, and thosein both arms are named Delx. For the LD and RD genomes, xindicates the number of bases deleted; for the Del mutantgenomes, x indicates the number of palindromic base pairsdeleted. An SL6 mutant genome with a six-nucleotide (CTGCTG) deletion at the stem-loop junction of the left arm wasalso constructed.
To assist in tracing the template used during DNA synthesis,the relevant viral sequences were assigned arbitrary positions(Fig. 1A). After excision and recircularization, the viral ge-nomes were transfected into PK15 cells and then assayed forviral protein synthesis and progeny virus production.
Viral protein synthesis. Transfected cultures were assayed at48 h for Rep-associated protein by immunochemical stainingwith a hyperimmune swine serum (3, 6). In comparison withthe parent J1 genome, an equivalent number of Rep-associ-ated antigen-producing cells were detected with the left-armdeletion genomes (LD3, SL6, LD7, and LD11), while the num-ber of Rep-associated antigen-producing cells was greatly re-duced with the right-arm deletion genomes (RD1, RD2a,RD2b, RD3, RD7, and RD10) and the double-arm deletiongenomes (Del3, Del7, and Del10) (Fig. 2). The number ofRep-associated antigen-producing cells exhibited by RD1,RD2b, and the rest of the RD and Del mutant genomes was25%, �10%, and �1% of that with J1, respectively. It wasinteresting that the two-nucleotide deletion (CA) in RD2a (1%of J1) that involved the C nucleotide at position 3 had a moreinhibitory effect than the two-nucleotide deletion (AG) in
9018 CHEUNG J. VIROL.
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

RD2b (10% of J1) that did not involve the C nucleotide atposition 3.
Progeny virus production. At 7 days, a set of parallel trans-fected cultures were harvested, frozen and thawed three times,and then assayed for progeny virus by immunochemical stain-ing after passage on fresh PK15 cells for 48 h. Infectiousviruses were readily recovered in the cultures transfected withthe left-arm deletion genomes LD3, SL6, LD7, and LD11. Forthe right-arm deletion genomes, fewer viruses were recoveredfrom RD1, RD2a, and RD2b in comparison with parent J1,and none were detected with the RD3, RD7, or RD10 or withany of the double-arm mutant genomes immediately aftertransfection. However, a small number of progeny viruses ev-idently were produced with RD3, Del3, and RD10, becauseafter an additional two to three cell passages, infectious viruseswere recovered from RD3 and Del3 (two out of two experi-ments) and occasionally from RD10 (one of five experiments).
Genotype of recovered progeny viruses. After three cell pas-sages and confirmation by immunochemical staining, virus-infected cell DNAs were isolated and amplified by PCR withPCV1-specific primers 1665F and 874R (Fig. 1A). Each PCRproduct was subcloned into a TA cloning plasmid (Invitrogen,Carlsbad, Calif.) for nucleotide sequence determination. A
collection of 22 viruses with various palindromic sequenceswere obtained (Fig. 3 to 5). To retrace the DNA strand used asthe template during synthesis of the palindromic sequences ineach of the recovered viruses, the following criteria were used:(i) template strand switching was invoked as little as possibleand only when the minus genome could not possibly be thetemplate, (ii) template-dependent DNA replication along thesame template was favored as long as possible, and (iii) a largesingle deletion had priority over multiple small deletions. Se-quences derived from their respective palindromic templatesvia template strand switching are shaded but not labeled, ille-gitimate nucleotide are circled, and deleted nucleotide areenclosed in dotted ovals.
Left-arm deletions. (i) LD3. A copy of the tandem triplet(CTG/CTG) at positions 5�, 4�, and 3�/u, v, and w was deletedin LD3, and the recovered progeny viruses contained only onecopy of the triplet (Fig. 3). Since the remaining triplet canoccupy the left-arm positions 5�, 4�, and 3�, the recoveredviruses may exhibit the original 11-nucleotide palindrome buta shortened loop (9 instead of 12 nucleotides). The resultsfrom two independent experiments showed that the engi-neered mutation was retained in the progeny viruses.
FIG. 2. Immunochemical staining of Rep-associated antigens in PK15 cells transfected with deletion genomes. The input genome is indicatedin each panel.
VOL. 78, 2004 MUTATIONAL ANALYSIS OF PORCINE CIRCOVIRUS ORIGIN 9019
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

(ii) SL6. Both copies of the tandem triplet (CTG/CTG) atpositions 5�, 4�, and 3�/u, v, and w were deleted in SL6, and therecovered progeny viruses obtained in two separate experimentsdid not contain these six nucleotides. Thus, these viruses havea noncomplementary upper region (positions 3, 4, and 5) andmay exhibit a shortened palindromic stem of eight nucleotides.
(iii) LD7. Seven nucleotides (TGCGCTG) at positions 9�through 3� were deleted. Four independent experiments wereconducted, and each yielded a set of progeny viruses withslightly different palindromic sequences. Inspection of theprogeny virus genomes suggested that the input viral DNAsmay have assumed multiple configurations based on nucleotide
FIG. 3. Input genomes and recovered viruses from left-arm mutations. For the input genome, the location of the engineered deletion and thepotential configuration(s) of each genome are presented at the left of each panel. The octanucleotide sequences are enclosed in boxes, and theinverted repeat sequences regenerated via template strand switching are shaded but not labeled. “Illegitimate” nucleotides are circled. Deletionsare indicated by dotted ovals. The dotted line in the palindrome indicates possible duplication of the preceding sequence. The C3 and C10nucleotides of vRD3.5 are indicated by asterisks. The number of examples (specific subclones/total number recovered) of each recovered virus,determined by sequencing cloned PCR fragments, is indicated in parentheses.
9020 CHEUNG J. VIROL.
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

base-pairing availability. It is unlikely that any stable palin-drome could have been formed by the base-pairing nucleotidethat remained in the engineered genomes (between one andfour nucleotide) at the onset of DNA replication. As illus-
trated, virus genome A was likely derived from configuration A(with four base-pairing nucleotides), while virus genome B waslikely derived from configuration B (with three base-pairingnucleotides). Essentially, the right-arm sequences of the recov-
FIG. 3—Continued.
VOL. 78, 2004 MUTATIONAL ANALYSIS OF PORCINE CIRCOVIRUS ORIGIN 9021
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

ered viruses were identical to that of the input genomes, whichindicates that the minus genome was used as the templateduring initiation of DNA replication. During termination, aportion of the right arm served as the template for synthesis ofthe left arm (shaded nucleotides) to generate a longer palin-drome. A total of six genotypes were observed among theprogeny viruses, and each contained a longer and more stablepalindrome than that of the input DNA. Evidently, templatestrand switching from the minus genome template to the newlysynthesized palindromic strand aN template occurred duringtermination of DNA replication. In addition, nucleotide inser-tion or deletion was observed in each progeny virus.
For vLD7.1, leading-strand DNA synthesis proceeded throughpositions u, v, w, x, y, and z before template strand switching touse the right arm as the template. Upon reverse template strandswitching (from palindromic strand aN to genome strand b�) tosynthesize the loop sequence, the triplet sequence TAG at posi-tions x, y, and z was duplicated. For vLD7.2, leading-strand DNAsynthesis proceeded through positions l, m, and n and then the
nucleotides at positions o, p, q, r, s, 13�, 12�, 11�, 10�, u, v, and wwere deleted. Template strand switching occurred at the bottomof the left arm, and the leading strand used the right arm as thetemplate for synthesizing almost the entire new left arm. ForvLD7.3, leading-strand DNA synthesis proceeded through posi-tions u, v, and w, and then template strand switching used theright arm as the template before reverse template strand switch-ing to use the minus genome at positions x, y, and z. With vLD7.4,vLD7.5, and vLD7.6, leading-strand DNA synthesis proceededthrough positions l, m, n, o, p, q, and r before template strandswitching occurred. For vLD7.4 and vLD7.6, the nucleotides atpositions s, 13�, 12�, 11�, 10�, u, v, and w were deleted. For vLD7.5,a shorter sequence (s, 13�, 12�, 11�, 10�) was deleted, and reversetemplate strand switching occurred at positions u, v, and w. Thus,for each virus, a different length of the right arm was used as thetemplate to regenerate the left-arm sequence prior to reversetemplate strand switching to synthesize the loop nucleotide.
(iv) LD11. The entire left-arm sequence from nucleotides13� to 3� was deleted in LD11. Judging from the four recovered
FIG. 3—Continued.
9022 CHEUNG J. VIROL.
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

viruses, the input genomes would have existed in two config-urations (A and B) during replication. Configuration A ap-peared to have recruited the l, m, n, o, and p sequence into themelting pot to generate vLD11.1, vLD11.2, and vLD11.4. BothvLD11.1 and vLD11.2 had a deletion of six nucleotides atpositions q, r, s, u, v, and w, and vLD11.2 incorporated anillegitimate nucleotide at position o (circled). For vLD11.1 andvLD11.2, template strand switching occurred after positions l,m, n, o, and p and then reverse template strand switchingoccurred at positions x, y, and z. For vLD11.4, only two nucle-otides at positions 6 to 7 of the right leg were used as thetemplates, and the three nucleotides at positions q, r, and swere deleted. Since the nucleotides at positions l, m, n, o, andp were not recruited into a new palindrome, configuration Bwas void of any base-pairing nucleotide. However, it gave riseto vLD11.3, which exhibited the original 11-nucleotide palin-drome by regenerating the sequence at positions 13� through 6�with the right arm as the template and then reverse templatestrand switching at positions u, v, and w. Nucleotide deletionwas not observed with vLD11.3, but an illegitimate C nucleo-tide was incorporated into the right arm at position 5. Thiscollection of progeny viruses showed that a different length ofthe right arm was also used as the template to regenerate theleft-arm sequence.
Right-arm deletions. Previous work (4) showed that the Cnucleotides at positions 3 and 10 (designated C3 and C10) arecritical for PCV1 progeny virus regeneration. The followingexperiments were aimed at deleting C3 or disrupting the spatialdistance between C3 and C10.
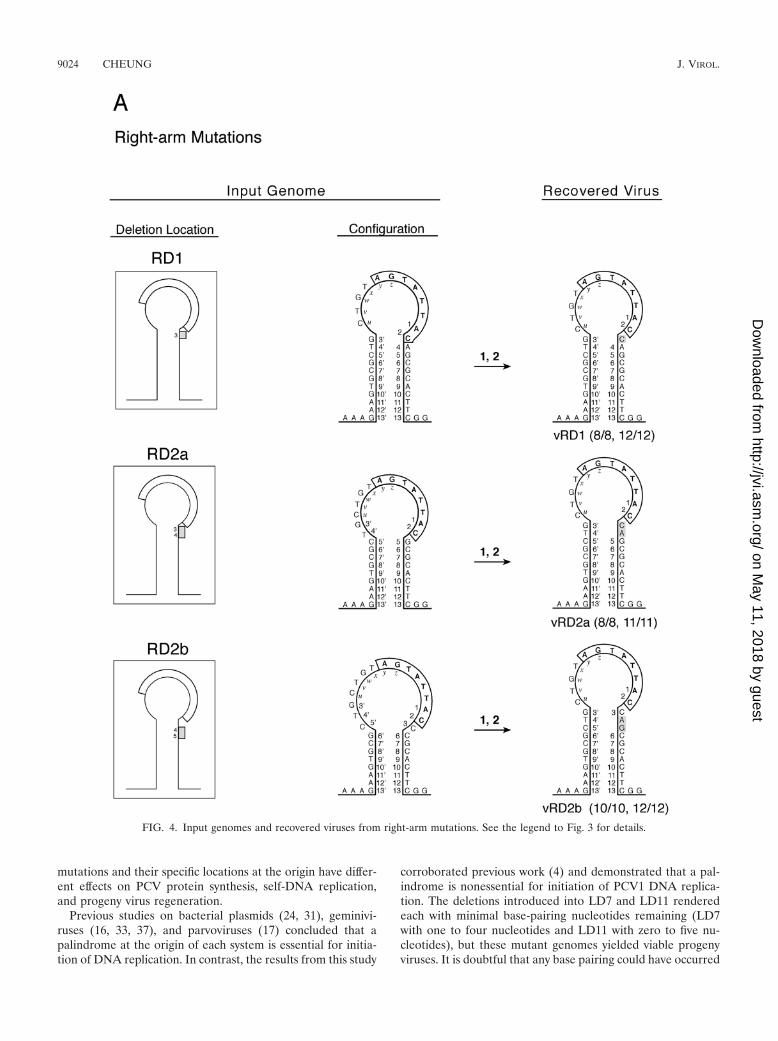
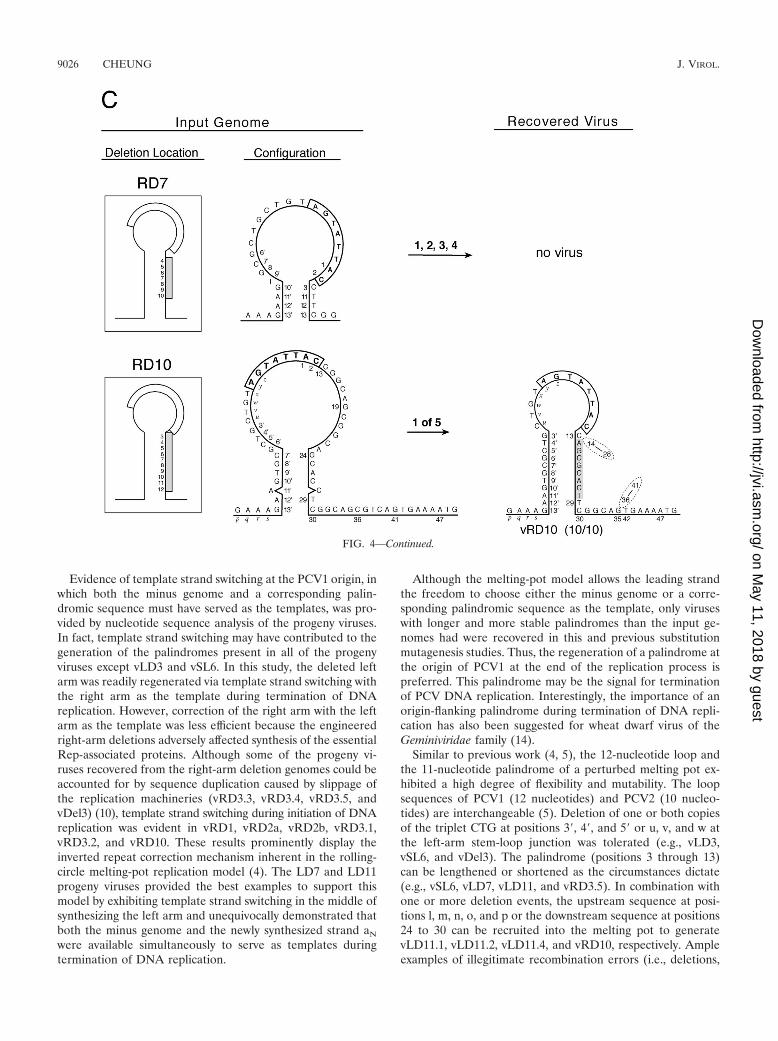
(i) RD1, RD2a, and RD2b. One or two nucleotides weredeleted from this group of mutant genomes. As shown above,the number of Rep-associated antigen-producing cells exhib-ited by RD1 (25%), RD2a (�1%), and RD2b (�10%) wasreduced, and fewer progeny viruses were recovered immedi-ately after transfection in comparison with the parent genomeJ1. However, all three input deletion genomes yielded wild-type viruses, and they could all be accounted for by templatestrand switching during initiation of DNA replication (Fig. 4).
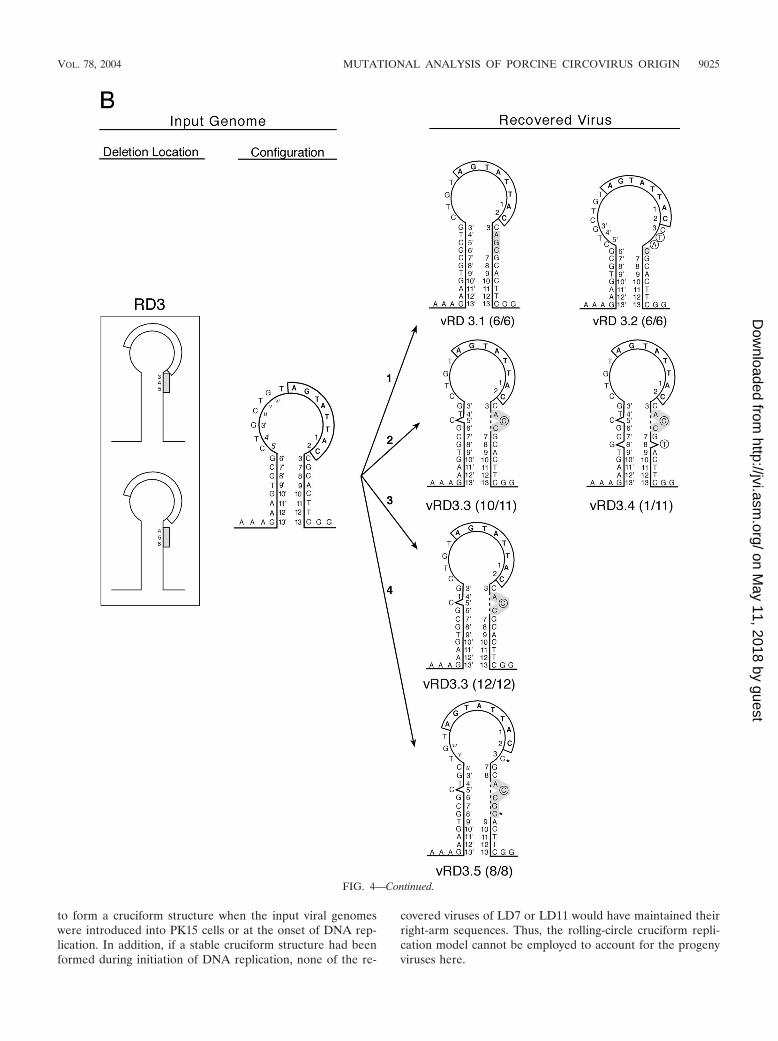
(ii) RD3. Three nucleotides were deleted from the right armat positions 3, 4, and 5 (CAG) or 4, 5, and 6 (AGC) in RD3,and five variant viruses were recovered. Presumably, this mu-tant genome would assume the 4-5-6 deletion configurationbecause C3 plays an important role in viral protein synthesis.For vRD3.1, template strand switching occurred after position3 at the onset of initiation of DNA replication, and the se-quence at 4�, 5�, and 6� was used as the template to regeneratethe wild-type palindrome. For vRD3.2, 2 illegitimate nucleo-tides (TA) were inserted prior to template strand switching touse position 6� as the template. This virus may exhibit anexpanded loop and a shortened palindrome. For vRD3.3 andvRD3.4, template strand switching occurred after position 3,and an illegitimate C nucleotide was incorporated in the re-generated sequence of each virus; in addition, vRD3.4 had asecond illegitimate T nucleotide at position 8. For vRD3.5,template strand switching occurred after position 8 and anillegitimate C nucleotide was incorporated in the regeneratedsequence; the actual C3 and C10 of the new genome are de-noted by asterisks. Alternatively, the regenerated sequences invRD3.3, vRD3.4, and vRD3.5 could also be the product ofduplication of the preceding sequence (ACC for vRD3.3 and
vRD3.4 and ACCGC for vRD3.5) (indicated by dotted lines)resulted from slippage of the replication machineries (10).
(iii) RD7 and RD10. Seven nucleotides from positions 3through 9 and 10 nucleotides from positions 3 through 12 weredeleted from RD7 and RD10, respectively. Progeny viruseswere not recovered from RD7, but on one occasion, viruseswere recovered from RD10. Apparently, the sequence at po-sitions 24 to 30 downstream from the original right arm wasrecruited into the stem portion of the melting pot but only thenucleotides at positions 29 and 30 were retained in the vRD10viruses. Template strand switching was evident at position 13to regenerate a wild-type sequence for the right arm, andconcurrently, two separate deletion events appeared to haveoccurred at positions 14 to 28 and 36 to 41. The data alsoshowed that only one copy of the hexanucleotide CGGCAGwas sufficient to sustain replication of the modified genome ofvRD10.
Double-arm deletions. (i) Del3. Three nucleotide at posi-tions 4, 5, and 6 of the right arm and a copy of the comple-mentary sequence at positions 4�, 5�, and 6� or u, v, and w ofthe left arm were deleted (Fig. 5). A variant virus with threenucleotides inserted into the right arm (positions 4, 5, and 6)but not the left arm was recovered. This new DNA sequencecould be accounted for by template strand switching at position3 with incorporation of an illegitimate C in the regeneratedsequence. Alternatively, the regenerated sequence (ACC)could also be the result of duplication of the preceding se-quence due to slippage of the replication machineries.
(ii) Del7 and Del10. Seven and 10 nucleotides on both sidesof the palindrome were deleted from Del7 and Del10, respec-tively. Progeny viruses were not recovered from either inputgenome in four independent experiments.
DISCUSSION
In this study, deletion mutagenesis was conducted to inves-tigate the importance of the origin-flanking palindromic se-quences of the PCV1 genome with respect to viral proteinsynthesis and progeny virus production via self-DNA replica-tion. A total of 13 mutant genomes were constructed. Three ofthe engineered genomes did not give any progeny virus, whilethe other 10 yielded 22 progeny virus variants. Deletions (from3 to 11 nucleotides) introduced into the left arm (LD3, SL6,LD7, and LD11) did not affect the number of viral antigen-producing cells, and progeny viruses were readily recoveredfrom cultures transfected with the excised and recircularizeddouble-stranded mutant genomes. In contrast, the right-armand double-arm deletion genomes exhibited greatly reducednumbers of Rep-associated protein-producing cells. Since theRep-associated antigens are essential for PCV DNA replica-tion, their reduction is expected to affect progeny virus pro-duction. Mutant genomes with deletions of three or fewernucleotides (RD1, RD2a, RD2b, RD3, and Del3) yieldedprogeny viruses consistently, while a seven-nucleotide-deletiongenome (RD7) did not give any progeny virus, and a 10-nucleotide-deletion genome (RD10) yielded infectious virusesonly occasionally. In comparison with a previous study (4),nucleotide substitutions engineered into the right arm, leftarm, or both arms of the palindrome did not affect proteinsynthesis or progeny virus production. Therefore, the types of
VOL. 78, 2004 MUTATIONAL ANALYSIS OF PORCINE CIRCOVIRUS ORIGIN 9023
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
mutations and their specific locations at the origin have differ-ent effects on PCV protein synthesis, self-DNA replication,and progeny virus regeneration.
Previous studies on bacterial plasmids (24, 31), geminivi-ruses (16, 33, 37), and parvoviruses (17) concluded that apalindrome at the origin of each system is essential for initia-tion of DNA replication. In contrast, the results from this study
corroborated previous work (4) and demonstrated that a pal-indrome is nonessential for initiation of PCV1 DNA replica-tion. The deletions introduced into LD7 and LD11 renderedeach with minimal base-pairing nucleotides remaining (LD7with one to four nucleotides and LD11 with zero to five nu-cleotides), but these mutant genomes yielded viable progenyviruses. It is doubtful that any base pairing could have occurred
FIG. 4. Input genomes and recovered viruses from right-arm mutations. See the legend to Fig. 3 for details.
9024 CHEUNG J. VIROL.
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
to form a cruciform structure when the input viral genomeswere introduced into PK15 cells or at the onset of DNA rep-lication. In addition, if a stable cruciform structure had beenformed during initiation of DNA replication, none of the re-
covered viruses of LD7 or LD11 would have maintained theirright-arm sequences. Thus, the rolling-circle cruciform repli-cation model cannot be employed to account for the progenyviruses here.
FIG. 4—Continued.
VOL. 78, 2004 MUTATIONAL ANALYSIS OF PORCINE CIRCOVIRUS ORIGIN 9025
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
Evidence of template strand switching at the PCV1 origin, inwhich both the minus genome and a corresponding palin-dromic sequence must have served as the templates, was pro-vided by nucleotide sequence analysis of the progeny viruses.In fact, template strand switching may have contributed to thegeneration of the palindromes present in all of the progenyviruses except vLD3 and vSL6. In this study, the deleted leftarm was readily regenerated via template strand switching withthe right arm as the template during termination of DNAreplication. However, correction of the right arm with the leftarm as the template was less efficient because the engineeredright-arm deletions adversely affected synthesis of the essentialRep-associated proteins. Although some of the progeny vi-ruses recovered from the right-arm deletion genomes could beaccounted for by sequence duplication caused by slippage ofthe replication machineries (vRD3.3, vRD3.4, vRD3.5, andvDel3) (10), template strand switching during initiation of DNAreplication was evident in vRD1, vRD2a, vRD2b, vRD3.1,vRD3.2, and vRD10. These results prominently display theinverted repeat correction mechanism inherent in the rolling-circle melting-pot replication model (4). The LD7 and LD11progeny viruses provided the best examples to support thismodel by exhibiting template strand switching in the middle ofsynthesizing the left arm and unequivocally demonstrated thatboth the minus genome and the newly synthesized strand aN
were available simultaneously to serve as templates duringtermination of DNA replication.
Although the melting-pot model allows the leading strandthe freedom to choose either the minus genome or a corre-sponding palindromic sequence as the template, only viruseswith longer and more stable palindromes than the input ge-nomes had were recovered in this and previous substitutionmutagenesis studies. Thus, the regeneration of a palindrome atthe origin of PCV1 at the end of the replication process ispreferred. This palindrome may be the signal for terminationof PCV DNA replication. Interestingly, the importance of anorigin-flanking palindrome during termination of DNA repli-cation has also been suggested for wheat dwarf virus of theGeminiviridae family (14).
Similar to previous work (4, 5), the 12-nucleotide loop andthe 11-nucleotide palindrome of a perturbed melting pot ex-hibited a high degree of flexibility and mutability. The loopsequences of PCV1 (12 nucleotides) and PCV2 (10 nucleo-tides) are interchangeable (5). Deletion of one or both copiesof the triplet CTG at positions 3�, 4�, and 5� or u, v, and w atthe left-arm stem-loop junction was tolerated (e.g., vLD3,vSL6, and vDel3). The palindrome (positions 3 through 13)can be lengthened or shortened as the circumstances dictate(e.g., vSL6, vLD7, vLD11, and vRD3.5). In combination withone or more deletion events, the upstream sequence at posi-tions l, m, n, o, and p or the downstream sequence at positions24 to 30 can be recruited into the melting pot to generatevLD11.1, vLD11.2, vLD11.4, and vRD10, respectively. Ampleexamples of illegitimate recombination errors (i.e., deletions,
FIG. 4—Continued.
9026 CHEUNG J. VIROL.
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

insertions, and illegitimate nucleotide incorporation) thatoccur frequently at the origin of phages and plasmids (25,26) were also detected here. Insertions or duplications (e.g.,vLD7.1, vRD3.2, and vRD3.5) and deletions (e.g., selectedvLD7, selected vLD11, and vRD10) were observed among theprogeny viruses. Incorporation of illegitimate nucleotide wasdetected in vLD11.2, vLD11.3, vRD3.2, vRD3.3, vRD3.4,
vRD3.5, and vDel3. However, the “illegitimate” nucleotides invRD3.3, vRD3.4, vRD3.5, and vDel3 could be attributed toduplication of the preceding sequence and not to illegitimatenucleotide incorporation. Finally, it is interesting that whenmultiple experiments were conducted with some of the mutantgenomes (four with LD7, three with LD11, four with RD3, andfive with RD10), multiple sets of progeny viruses were ob-
FIG. 5. Input genomes and recovered viruses from double-arm mutations. See the legend to Fig. 3 for details.
VOL. 78, 2004 MUTATIONAL ANALYSIS OF PORCINE CIRCOVIRUS ORIGIN 9027
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

tained. These observations suggest that the destabilized envi-ronment at the PCV1 origin, with all four strands of the in-verted repeats in the melted state, and the availability of twotemplates simultaneously during initiation and termination ofDNA replication may contribute to the flexibility as well as theincreased mutation frequency at the origin.
Integrated in this destabilized and very flexible melting-potenvironment is a certain degree of organization. Formation ofthe melting pot is dependent upon the interactions betweenthe REP complex and its cognate octanucleotide motif se-quence (5, 20). Mutant nucleotides engineered into positions 3and 10 of the right arm reverted back to wild-type C nucleo-tides in the progeny virus genomes irrespective of the minusgenome template or the corresponding palindromic strandtemplate (4). These two positions have been designated birth-right positions, and presumably their birthrights are “inher-ited” and dictated by the REP complex. All the progeny virusesrecovered from the right-arm deletion genomes contained a Cnucleotide at positions 3 and 10 through insertion or deletionof the appropriate number of nucleotides via template strandswitching or duplication. The most notable examples arevRD3.5 and vRD10, in which the genomes underwent multiplemodifications (a combination of insertion, duplication, dele-tion, and illegitimate nucleotide incorporation) to maintain theC3 and C10 configuration. These data indicate that the prede-termined status of positions 3 and 10 and the spatial distancebetween C3 and C10 are critical for PCV1 DNA replication.Furthermore, DNA sequence complementarity is not the onlydeciding factor for nucleotide incorporation during self-DNAreplication at the PCV1 origin.
Not unexpectedly, the results obtained in this investigationarrived at the melting-pot model independently of the substi-tution mutagenesis study (4). Thus, this work confirms andextends previous observations that the rolling-circle melting-pot replication model provides a viable mechanism for DNAreplication, inverted repeat correction (or conversion), andillegitimate recombination at the PCV1 origin and may beapplicable to the replication of other circular DNAs.
ACKNOWLEDGMENTS
I thank S. Pohl, D. Alt, K. Halloum, and O. Kohutyuk for technicalassistance and M. Marti and S. Ohlendorf for manuscript preparation.
Mention of trade names or commercial products in this article issolely for the purpose of providing specific information and does notimply recommendation or endorsement by the U.S. Department ofAgriculture.
REFERENCES
1. Allan, G. M., and J. A. Ellis. 2000. Porcine circoviruses: a review. J. Vet.Diagn. Investig. 12:3–14.
2. Cheung, A. K. 2003. Comparative analysis of the transcriptional patterns ofpathogenic and non-pathogenic circoviruses. Virology 301:41–49.
3. Cheung, A. K. 2004. Identification of the essential and non-essential tran-scriptional units for protein synthesis, DNA replication and infectious virusproduction of porcine circovirus type 1. Arch. Virol. 149:975–988.
4. Cheung, A. K. 2004. Detection of template strand-switching during initiationand termination of DNA replication of porcine circovirus. J. Virol. 78:4268–4277.
5. Cheung, A. K. Identification of an octanucleotide motif sequence essentialfor viral protein, DNA and progeny virus biosynthesis at the origin of DNAreplication of porcine circovirus type 2. Virology, 324:28–36.
6. Cheung, A. K., and S. R. Bolin. 2002. Kinetics of porcine circovirus type 2replication. Arch. Virol. 147:43–58.
7. del Solar, G., R. M. Giraldo, J. Ruiz-Echevarria, M. Espinosa, and R.Diaz-Orejas. 1998. Replication and control of circular bacterial plasmids.Microbiol. Mol. Biol. Rev. 62:434–464.
8. Fenaux, M., P. G. Halbur, M. Gill, T. E. Toth, and X. J. Meng. 2000. Geneticcharacterization of type 2 porcine circovirus (PCV-2) from pigs with post-weaning multisystemic wasting syndrome in different geographic regions ofNorth America and development of a differential PCR-restriction fragmentlength polymorphism assay to detect and differentiate between infectionswith PCV-1 and PCV-2. J. Clin. Microbiol. 38:2494–2503.
9. Gibbs, M. J., and G. F. Weiller. 1999. Evidence that a plant virus switchedhosts to infect a vertebrate and then recombined with a vertebrate-infectingvirus. Proc. Natl. Acad. Sci. USA 96:8022–8027.
10. Graham, F. L., J. Rudy, and P. Brinkley. 1989. Infectious circular DNA ofhuman adenovirus type 5: regeneration of viral DNA termini from moleculeslacking terminal sequences. EMBO J. 7:2077–2085.
11. Gutierrez, C. 1999. Geminivirus DNA replication. Cell. Mol. Life Sci. 56:313–329.
12. Hamel, A. L., L. L. Lin, and G. P. S. Nayar. 1998. Nucleotide sequence ofporcine circovirus associated with postweaning multisystemic wasting syn-drome in pigs. J. Virol. 72:5262–5267.
13. Hanley-Bowdoin, L., S. B. Settlage, B. M. Orozco, S. Nagar, and D. Robert-son. 2000. Geminiviruses: models for plant DNA replication, transcription,and cell cycle regulation Crit. Rev. Biochem. Mol. Biol. 35:105–140.
14. Kammann, M., H. J. Schalk, V. Matzeit, S. Schaefer, J. Schell, and B.Gronenborn. 1991. DNA replication of wheat dwarf virus, a geminivirus,requires two cis-acting signals. Virology 184:786–790.
15. Khan, S. A. 2000. Plasmid rolling-circle replication: recent developments.Mol. Microbiol. 37:477–484.
16. Lazarowitz, S. G., A. J. Pinder, V. D. Damsteegt, and J. S. Rogers. 1989.Maize streak virus genes essential for systemic and symptom development.EMBO J. 8:1023–1032.
17. Lefebvre, R. B., S. Riva, and K. I. Berns. 1984. Conformation takes prece-dence over sequence in adeno-associated virus DNA replication. Mol. Cell.Biol. 4:1416–1419.
18. Mankertz, A., and B. Hillenbrand. 2001. Replication of porcine circovirustype 1 requires two proteins encoded by the viral rep gene. Virology 279:429–438.
19. Mankertz, A., J. Mankertz, K. Wolf, and H.-J. Buhk. 1998. Identification ofa protein essential for replication of porcine circovirus. J. Gen. Virol. 79:381–384.
20. Mankertz, A., F. Persson, J. Mankertz, G. Blaess, and H.-J. Buhk. 1997.Mapping and characterization of the origin of DNA replication of porcinecircovirus. J. Virol. 71:2562–2566.
21. McNulty, M., J. Dale, P. Lukert, A. Mankertz, J. Randles, and D. Todd.2000. Circoviridae, p. 299–303. In M. H. V. van Regenmortel, C. M. Fauquet,D. H. L. Bishop, E. B. Carstens, M. K. Estes, S. M. Lemon, J. Maniloff, M. A.Mayo, D. J. McGeoch, C. R. Pringle, and R. B. Wickner (ed.), Seventhreport of the International Committee on Taxonomy of Viruses. AcademicPress, San Diego, Calif.
22. Meehan, B. M., J. L. Creelan, M. S. McNulty, and D. Todd. 1997. Sequenceof porcine circovirus DNA: affinities with plant circoviruses. J. Gen. Virol.78:221–227.
23. Meehan, B. M., F. McNeilly, D. Todd, S. Kennedy, A. Jewhurst, J. A. Ellis,L. E. Hassard, E. G. Clark, D. M. Haines, and G. M. Allan. 1998. Charac-terization of novel circovirus DNAs associated with wasting syndromes inpigs. J. Gen. Virol. 79:2171–2179.
24. Miao, D. M., Y. Honda, K. Tanaka, A. Higashi, T. Nakamura, Y. Taguchi,H. Sakai, T. Komano, and M. Bagdasarian. 1993. A base-paired hairpinstructure essential for the functional priming signal for DNA replicationof the broad host range plasmid RSF1010. Nucleic Acids Res. 21:4900–4903.
25. Michel, B., and S. D. Ehrlich. 1986. Illegitimate recombination at the rep-lication origin of bacteriophage M13. Proc. Natl. Acad. Sci. USA 83:3386–3390.
26. Michel, B., and S. D. Ehrlich. 1986. Illegitimate recombination occurs be-tween the replication origin of the plasmid pC194 and a progressing repli-cation fork. EMBO J. 5:3691–3696.
27. Morozov, I., T. Sirinarumitr, S. Sorden, P. G. Halbur, M. K. Morgan, K.-J.Yoon, and P. S. Paul. 1998. Detection of a novel strain of porcine circovirusin pigs with postweaning multisystemic wasting syndrome. J. Clin. Microbiol.36:2535–2541.
28. Musatov, S., J. Roberts, D. Pfaff, and M. Kaplitt. 2002. A cis-acting elementthat directs circular adeno-associated virus replication and packaging. J. Vi-rol. 76:12792–12802.
29. Musatov, S. A., T. A. Scully, L. Dudus, and K. J. Fisher. 2000. Induction ofcircular episomes during rescue and replication of adeno-associated virus inexperimental models of virus latency. Virology 275:411–432.
30. Niagro, F. D., A. N. Forsthoefel, R. P. Lawther, L. Kamalanathan, B. W.Ritchie, K. S. Latimer, and P. D. Lukert. 1998. Beak and feather diseasevirus and porcine circovirus genomes: intermediates between geminivirusesand plant circoviruses. Arch. Virol. 143:1723–1744.
31. Noirot, P., J. Bargonetti, and R. P. Novick. 1990. Initiation of rolling-circlereplication in pT181 plasmid: initiator protein enhances cruciform extrusionat the origin. Proc. Natl. Acad. Sci. USA 87:8560–8564.
9028 CHEUNG J. VIROL.
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

32. Novick, R. P. 1998. Contrasting lifestyles of rolling-circle phages and plas-mids. Trends Biochem. Sci. 23:434–438.
33. Orozco, B. M., and L. Hanley-Bowdoin. 1996. A DNA structure is requiredfor geminivirus replication origin function. J. Virol. 70:148–158.
34. Palmer, K. E., and E. O. Rybicki. 1998. The molecular biology of mastrevi-ruses. Adv. Virus Res. 50:183–234.
35. Pringle, C. R. 1999. Virus taxonomy at the XIth International Congress ofVirology, Sydney, Australia. Arch. Virol. 144:2065–2070.
36. Randles, J. W., P. W. G. Chu, J. L. Dale, R. M. Harding, J. Hu, M. Kojima,K. M. Makkouk, Y. Sana, J. E. Thomas, and H. J. Vetten. 2000. Nanovirus,p. 303–309. In M. H. V. van Regenmortel, C. M. Fauquet, D. H. L. Bishop,E. B. Carstens, M. K. Estes, S. M. Lemon, J. Maniloff, M. A. Mayo, D. J.McGeoch, C. R. Pringle, and R. B. Wickner (ed.), Seventh report of the
International Committee on Taxonomy of Viruses. Academic Press, SanDiego, Calif.
37. Revington, G. N., G. Sunter, and D. M. Bisaro. 1989. DNA sequencesessential for replication of the B genome component of tomato goldenmosaic virus. Plant Cell 1:985–992.
38. Samulski, R. J., A. Srivastava, K. I. Berns, and N. Muzyczka. 1983. Rescueof adeno-associated virus from recombinant plasmids: gene correction withinthe terminal repeats of AAV. Cell 33:135–143.
39. Steinfeldt, T., T. Finsterbusch, and A. Mankertz. 2001. Rep and Rep� pro-tein of porcine circovirus type 1 bind to the origin of replication in vitro.Virology 291:152–160.
40. Tischer, I., H. Gelderblom, W. Vettermann, and M. A. Koch. 1982. Avery small porcine virus with circular single-stranded DNA. Nature 295:64–66.
VOL. 78, 2004 MUTATIONAL ANALYSIS OF PORCINE CIRCOVIRUS ORIGIN 9029
on May 11, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from





























